Back to Journals » Cancer Management and Research » Volume 18

The Interaction of PI3K/AKT/mTOR Pathway with the Tumor Microenvironment: A Dual Engine Driving Breast Cancer Progression and Reshaping Treatment Strategies
Authors Huang X, Xia C, Jin X, Wang Y
Received 13 April 2026
Accepted for publication 24 June 2026
Published 30 June 2026 Volume 2026:18 616696
DOI https://doi.org/10.2147/CMAR.S616696
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yong Teng
Xi Huang, Chunjun Xia, Xiangyue Jin, Yu Wang
Department of Oncology, Jianhu Clinical Medical College of Yangzhou University, Yancheng, 224700, People’s Republic of China
Correspondence: Xi Huang, Email [email protected]
Abstract: The PI3K/AKT/mTOR signaling pathway, as a core regulatory network for cell growth, proliferation and metabolism, frequently undergoes abnormal activation in breast cancer, becoming a key driver of tumor occurrence and development. In recent years, research perspectives have shifted from solely focusing on intrinsic signals within tumor cells to the complex interactions between these cells and the tumor microenvironment (TME). This review systematically explores how the PI3K/AKT/mTOR pathway shapes an immunosuppressive, pro-angiogenic, and pro-metastatic microenvironment through interactions with immune cells, fibroblasts, vascular endothelial cells, and the extracellular matrix in the TME, thereby promoting the malignant progression, drug resistance, and recurrence of breast cancer. Additionally, it analyzes novel therapeutic strategies targeting this pathway and its interactions with the microenvironment, including combination immunotherapy, anti-angiogenic therapy, and metabolic modulation therapy, providing new ideas and potential directions to overcome the current limitations in breast cancer treatment.
Keywords: PI3K/AKT/mTOR pathway, tumor microenvironment, breast cancer, therapy
Introduction
Breast cancer (BC), as the most common malignant tumor among women globally, presents significant clinical challenges due to its high heterogeneity, treatment resistance, and recurrence.1 In recent years, research perspectives have shifted from tumor cells themselves to the tumor microenvironment (TME), a dynamic ecosystem that includes immune cells, cancer-associated fibroblasts (CAFs), endothelial cells, and extracellular matrix components, closely related to tumor progression.2 The PI3K/AKT/mTOR pathway, as a key intracellular pro-survival signaling axis, has transcended its regulatory role over the autonomous behavior of tumor cells, becoming a core hub for “cross-border communication” between tumor cells and TME.1
In this review, a deep analysis of the bidirectional interaction network between the PI3K/AKT/mTOR pathway and the TME not only reveals the mechanisms of breast cancer progression but also provides a theoretical basis for developing combination therapies and overcoming resistance.1 Future research needs to further explore the synergistic effects of pathway inhibitors and microenvironment-targeted drugs to optimize clinical efficacy and improve patient prognosis.3
The Core Role and Mechanisms of Abnormal Activation of the PI3K/AKT/mTOR Pathway in Breast Cancer
Basic Structure and Physiological Function of PI3K/AKT/mTOR Pathway
The PI3K/AKT/mTOR signaling pathway is a core signal transduction network regulating growth, proliferation, survival, and metabolism within cells, playing a key role in maintaining cellular homeostasis by integrating various extracellular and intracellular signals.4 This pathway is primarily composed of key molecules such as phosphoinositide 3-kinase (PI3K), protein kinase B (AKT), and mammalian target of rapamycin (mTOR). Under physiological conditions, growth factors and cytokines bind to receptors (such as receptor tyrosine kinases) on the cell membrane, activating PI3K, which catalyzes the production of the second messenger PIP3. PIP3 recruits and activates downstream AKT, and activated AKT phosphorylates various substrates, ultimately activating mTOR complexes (mTORC1 and mTORC2), thereby regulating a series of cellular processes such as protein synthesis, glycolysis, and autophagy.1,4 This pathway is not only crucial for the physiological functions of normal cells, but its signal output also influences the differentiation and function of immune cells in the tumor microenvironment, serving as an important bridge connecting the intrinsic characteristics of tumor cells with the external microenvironment (Figure 1).4
 |
Figure 1 Physiological function of the PI3K/AKT/mTOR pathway. The activation and downstream effects of the PI3K/AKT/mTOR pathway are involved in various biological processes. Created with BioRender.com. Abbreviations: PI3K, phosphatidylinositol 3-kinase; PIP3, phosphatidylinositol (3,4,5)-trisphosphate; AKT, protein kinase B; mTORC1/2, mechanistic target of rapamycin complex 1/2. |
Genetic and Epigenetic Changes of the Pathway in Breast Cancer
In breast cancer, the PI3K/AKT/mTOR pathway is one of the most frequently activated pathways, with its dysregulation primarily driven by genetic and epigenetic changes. The most common genetic alteration is somatic mutations in the PIK3CA gene, which encodes the catalytic subunit p110α of PI3K, found in approximately 40% of hormone receptor-positive (HR+) breast cancers, leading to constitutive activation of PI3K.5,6 Additionally, the loss or functional impairment of the tumor suppressor gene PTEN, as well as mutations in genes such as AKT1 and PIK3R1, are also important causes of abnormal activation of this pathway.6,7 Besides genetic mutations, epigenetic regulation is also involved. For example, the overexpression of long non-coding RNA (lncRNA) such as MALAT1 can promote the proliferation and metastasis of breast cancer cells by upregulating the expression of key genes in the PI3K/AKT/mTOR pathway.8 Dysregulation of circular RNA (circRNA) has also been shown to influence the progression of breast cancer by regulating this pathway.9 These multi-layered changes collectively lead to the sustained activation of the PI3K/AKT/mTOR signaling pathway, becoming a core driving force in the occurrence and development of breast cancer (Figure 2).
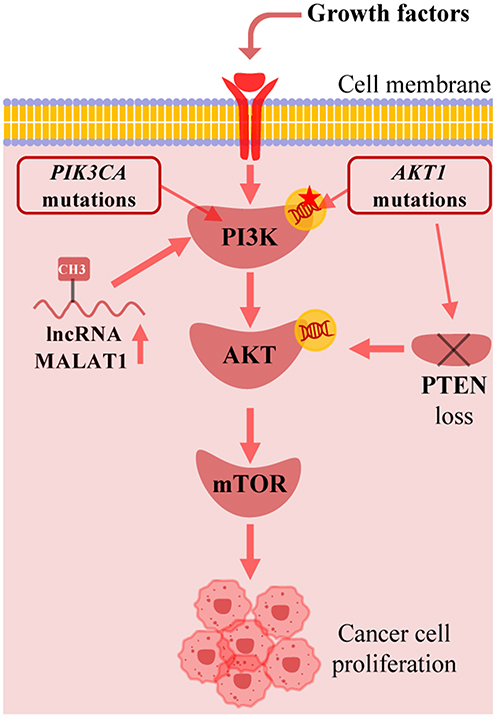 |
Figure 2 Genetic and epigenetic dysregulation in breast cancer. PIK3CA mutation, AKT1 mutations, lncRNA MALAT1 and PTEN loss are able to cause constitutive activation of PI3K/AKT/mTOR pathway to promote cancer cell growth. Created with BioRender.com. Abbreviations: lncRNA, long non-coding RNA; PTEN, phosphatase and tensin homolog. |
Association of Pathway Activation with Molecular Subtypes and Prognosis of Breast Cancer
Abnormal activation of the PI3K/AKT/mTOR pathway is closely related to different molecular subtypes, clinical pathological features, and patient prognosis in breast cancer:
(1) HR+/HER2− subtype: Roughly 40% patients carry activating PIK3CA hotspot mutations, persistent pathway activation is the core driver of endocrine therapy resistance; Alpelisib combined with fulvestrant is the standard targeted regimen for mutated patients.4
(2) HER2+ subtype: HER2 gene amplification induces continuous upstream pathway activation; dual anti-HER2 combined with PI3K inhibitor achieves better disease control compared with single-agent treatment.
(3) TNBC subtype: PTEN gene deletion is the predominant driver of pathway overactivation instead of PIK3CA mutation, and pathway upregulation largely fuels immunosuppressive TME formation and early distant metastasis.5
Studies have shown that the number of PIK3CA mutations and specific loci (such as the C2 domain) are associated with shortened overall survival in patients.6 Additionally, the overexpression of pathway-related genes such as Exportin-T (XPOT) is associated with adverse clinical features such as negative estrogen receptor, negative progesterone receptor, large tumor size, and lymph node metastasis, indicating poorer prognosis.10 The excessive activation of this pathway not only drives malignant behaviors of tumor cells, such as proliferation, survival, migration, and metabolic reprogramming, but is also related to the maintenance of tumor stem cell characteristics, epithelial-mesenchymal transition, and treatment resistance, collectively leading to a more aggressive tumor phenotype and worse clinical outcomes.7 Therefore, assessing the status of this pathway is of great value for molecular typing, prognosis assessment, and treatment strategy selection in breast cancer.
The Role of the PI3K/AKT/mTOR Pathway in Shaping the Tumor Immune Microenvironment and Immune Evasion
Regulation of Polarization and Function of Tumor-Associated Macrophages
The PI3K/AKT/mTOR pathway significantly influences the immunosuppressive characteristics of the tumor microenvironment by regulating the polarization state of tumor-associated macrophages (TAMs). Studies have shown that activation of this pathway can promote macrophage polarization towards the M2 type, characterized by high expression of immunosuppressive markers such as CD206 and IL-10 (Figure 3).4 In breast cancer models, mTOR signaling induces the conversion of macrophages to the M2 phenotype through upregulation of transcription factors ATF3 and the CXCL8/IL-8 axis, thereby promoting tumor angiogenesis and metastasis.11 PI3K/AKT/mTOR signaling shifts macrophage functional phenotype toward an immune-suppressive M2-like status rather than complete subtype conversion. Of note, Torin1-mediated TAM repolarization data was originally observed in hepatocellular carcinoma, further in vivo verification within breast cancer models is required to confirm translational feasibility.12 Notably, the metabolic reprogramming of TAMs (such as enhanced glycolysis) is also regulated by this pathway, further strengthening their immunosuppressive function.4
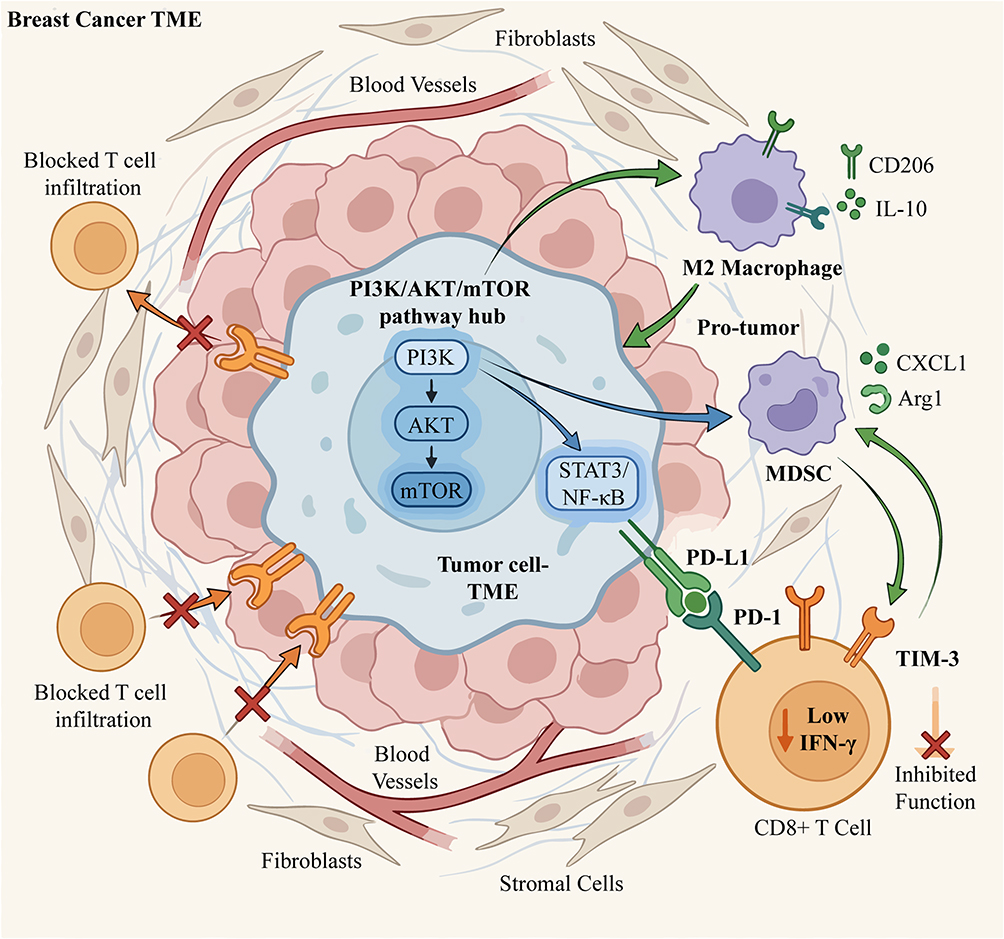 |
Figure 3 The PI3K/AKT/mTOR pathway mediates immunosuppression in the breast cancer tumor microenvironment (TME). The PI3K/AKT/mTOR signaling in breast cancer cells upregulates downstream transcription factors (eg, STAT3, NF-κB), which orchestrate immunosuppressive processes, including inducing M2-like macrophage differentiation and promoting MDSC accumulation to suppress anti-tumor immunity. Moreover, the PI3K/AKT/mTOR signaling prevents T cell infiltration and upregulates PD-L1, which binds PD-1 on CD8+ T cells, to reduce IFN-γ secretion and impair T cell function. Created with BioRender.com. Abbreviations: TME, tumor microenvironment; MDSC, myeloid-derived suppressor cell; PD-L1, programmed death-ligand 1; PD-1, programmed death-1; TIM-3, T cell immunoglobulin and mucin-domain containing-3; IFN-γ, interferon-gamma; Arg1, arginase 1. |
Impact on the Recruitment and Activity of Myeloid-Derived Suppressor Cells
The expansion and function of myeloid-derived suppressor cells (MDSCs) are highly dependent on the PI3K/AKT/mTOR pathway. In non-small cell lung cancer, RNF7 upregulates CXCL1 expression by activating NF-κB signaling, promoting the recruitment of MDSCs to the tumor site.13 In pancreatic ductal adenocarcinoma (PDAC) models, the PI3K inhibitor taselisib reduces MDSC infiltration while increasing the proportion of CD8+ T cells, reshaping the immune microenvironment.14 Mechanistically, mTOR signaling regulates the phosphorylation of GM-CSF receptors, affecting the activity of transcription factors such as HOXA10 and IRF8, thereby controlling the differentiation fate of MDSCs.15 Additionally, lactic acid secreted by tumor cells can activate the PI3K/AKT/mTOR pathway, inducing MDSCs to highly express arginase 1 (Arg1), thereby inhibiting T cell function (Figure 3).16
Inhibition of Cytotoxic T Lymphocyte Infiltration and Function
Abnormal activation of the PI3K/AKT/mTOR pathway weakens the anti-tumor effects of CD8+ T cells through multiple mechanisms (Figure 3). In the breast cancer microenvironment, this pathway inhibits T cell receptor signaling, leading to reduced secretion of IFN-γ and granzyme B.17 Spatial transcriptomic analysis shows that activation of the PI3K/AKT signaling is significantly associated with the absence of CD8+ T cell infiltration in cervical cancer.18 Furthermore, tumor cells induce T cell exhaustion phenotypes (such as PD-1+TIM-3+) through mTOR-dependent upregulation of PD-L1. Notably, the loss of ULK1 can promote CD8+ T cell infiltration and enhance their cytotoxicity by activating the PI3K/AKT/mTOR pathway.19 In colorectal cancer models, miR-26a-5p significantly reduces the survival and killing ability of CD8+ tumor-infiltrating lymphocytes by inhibiting the EP300/PI3K/AKT/mTOR axis.20
Regulation of Immune Checkpoint Molecules Such as PD-L1 in Tumor Cells
The PI3K/AKT/mTOR pathway can directly regulate the transcription and translation of PD-L1 (Figure 3). In non-small cell lung cancer, activation of the EGFR/PI3K/AKT/mTOR cascade upregulates PD-L1 protein expression through downstream effector molecules STAT3, NF-κB, and c-MYC.21 Experimental evidence confirms that the nuclear form of PD-L1 maintains sister chromatid cohesion by binding to PDS5B, suggesting its pro-cancer function independent of immune checkpoints.22 In head and neck squamous cell carcinoma, disruption of the HER3-PI3K-mTOR axis simultaneously downregulates PD-L1 expression and reverses the immunosuppressive microenvironment.23 Additionally, SMARCAL1 stabilizes the chromatin accessibility of the PD-L1 transcription regulatory region, cooperating with JUN to maintain PD-L1 expression and promote immune evasion.24 These findings provide a theoretical basis for treatment strategies targeting both the PI3K/mTOR and PD-1/PD-L1 pathways.
Interaction Between the Pathway and Cancer-Associated Fibroblasts and Extracellular Matrix
Promotion of CAF Activation and Pro-Tumor Factor Secretion
The PI3K/AKT/mTOR pathway plays a central role in the activation of CAFs. Studies indicate that this pathway promotes the secretion of CXCL10 through upregulation of the Toll-like receptor 3-interferon regulatory factor-interferon β (TLR3-IRF-IFNβ) signaling axis, thereby activating CAFs and enhancing their pro-tumor functions (Figure 4).25 In colon cancer, FGFR4 drives CAF differentiation through the CXCL10-CXCR3 axis, forming an immunosuppressive TME that further promotes tumor progression.25 Moreover, activation of the PI3K/AKT/mTOR pathway can induce CAFs to secrete pro-inflammatory factors such as IL-6 and IL-8, promoting tumor cell proliferation and metastasis through paracrine mechanisms.26 In pancreatic ductal adenocarcinoma (PDAC), CAF activation also relies on the synergistic action of TGF-β signaling and the PI3K/AKT/mTOR pathway, leading to matrix stiffening and limiting chemotherapy drug penetration.14 These findings reveal how the PI3K/AKT/mTOR pathway regulates CAF activation and their pro-tumor functions through multiple pathways.
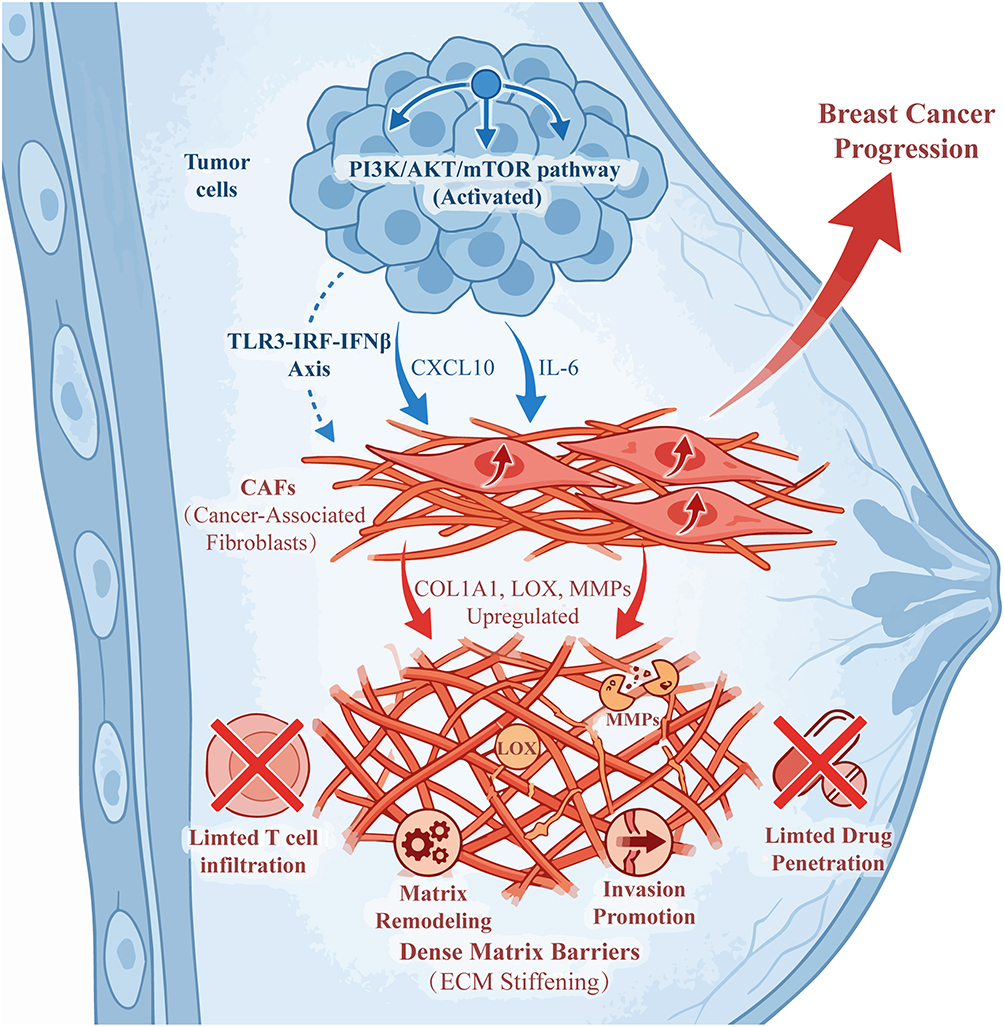 |
Figure 4 Activated PI3K/AKT/mTOR pathway in breast cancer cells drives tumor progression via cancer-associated fibroblast (CAF)-mediated extracellular matrix (ECM) remodeling. Activated PI3K/AKT/mTOR pathway in breast cancer cells upregulates secretion of cytokines (eg, CXCL10, IL-6) and activates the TLR3-IRF-IFNβ axis, which together induce CAF activation. Activated CAFs upregulate expression of ECM regulators (COL1A1, LOX, MMPs), leading to ECM stiffening (dense matrix barrier formation) and matrix remodeling. Dense ECM barriers block anti-tumor T cell infiltration and ECM stiffening impairs therapeutic drug delivery to tumor cells. MMP-mediated matrix degradation facilitates tumor cell invasion and dissemination. Created with BioRender.com. Abbreviations: CAF, cancer-associated fibroblast; ECM, extracellular matrix; TLR3, toll-like receptor 3; IRF, interferon regulatory factor; IFNβ, interferon beta; COL1A1, collagen type I alpha 1 chain; LOX, lysyl oxidase; MMP, matrix metalloproteinase. |
Driving Extracellular Matrix Remodeling and Stiffening
The PI3K/AKT/mTOR pathway directly participates in the dynamic remodeling of the ECM by regulating the synthesis of matrix metalloproteinases (MMPs) and collagen (Figure 4). In hepatocellular carcinoma, YAP1-positive CAFs activate the PI3K/AKT/mTOR pathway, upregulating the expression of COL1A1, COL3A1, and LOX, leading to increased ECM cross-linking and matrix stiffening.27 This stiffening not only enhances tissue mechanical strength but also promotes tumor cell invasion through integrin signaling.28 Furthermore, activation of mTOR can inhibit the degradation activity of MMP-14, reducing collagen cleavage and thus maintaining the dense structure of the ECM.29 In glioblastoma, CAFs induce the activation of astrocytes via the AKT/mTOR-SERPINH1/COL5A1 axis, further promoting ECM deposition and tumor progression.30 Abnormal remodeling of the ECM also limits T cell infiltration by forming physical barriers, exacerbating immune evasion.31
Mediating the Formation of Physical Barriers for Treatment Resistance
Excessive deposition and cross-linking of the ECM create physical barriers to treatment resistance through the PI3K/AKT/mTOR pathway (Figure 4). In non-small cell lung cancer (NSCLC), COL3A1 and COL6A1 expressed by tumor cells construct “castle-like” and “armor-like” structures through TGF-β-dependent mechanisms, hindering T cell penetration and weakening the efficacy of immune checkpoint inhibitors.32 Similarly, the dense stroma in pancreatic cancer increases tissue hydraulic pressure and limits drug diffusion, leading to chemotherapy resistance.14 Inhibitors targeting this pathway (such as taselisib) can reverse ECM stiffening, improve drug delivery, and enhance CD8+ T cell infiltration.14 Additionally, SMYD3 activates Src/PKCδ signaling through epigenetic regulation of CDCP1 expression, further promoting CAF-mediated ECM remodeling and oxaliplatin resistance.31 These findings emphasize the potential value of targeting the PI3K/AKT/mTOR pathway to overcome ECM-related treatment barriers.
The Role of PI3K/AKT/mTOR Signaling in Tumor Angiogenesis and Establishment of Pre-Metastatic Microenvironment
Regulation of Vascular Endothelial Growth Factor Expression and Angiogenesis
The PI3K/AKT/mTOR signaling pathway is a key molecular hub regulating tumor angiogenesis, one of its core mechanisms being the upregulation of vascular endothelial growth factor (VEGF) expression (Figure 5). This pathway integrates various extracellular and intracellular signals from the TME to directly or indirectly promote the transcription and translation of VEGF, thereby driving the proliferation, migration, and formation of new blood vessels in endothelial cells.4 For example, in PDAC, sustained activation of the PI3K/AKT/mTOR pathway is a key driver of tumor progression, with downstream effector molecules such as HIF-1α directly inducing VEGF expression and promoting abnormal tumor vascularization.33 In lung cancer models, targeting the PI3K/AKT/mTOR/HIF-1α signaling pathway effectively inhibits CD31-positive microvessel density (MVD) and levels of pro-angiogenic factors such as VEGF, bFGF, and TGF-β1 in serum, thereby improving the hypoxic tumor microenvironment and inhibiting tumor growth and metastasis.34 Additionally, in hepatocellular carcinoma, the chemokine CXCL3 activates the PI3K/AKT/mTOR pathway, upregulating various pro-angiogenic factors including VEGF, thereby reshaping the tumor microenvironment and promoting tumor progression.12 Collectively, these studies indicate that the PI3K/AKT/mTOR pathway is a core bridge connecting intrinsic signals in tumor cells with the angiogenic microenvironment.
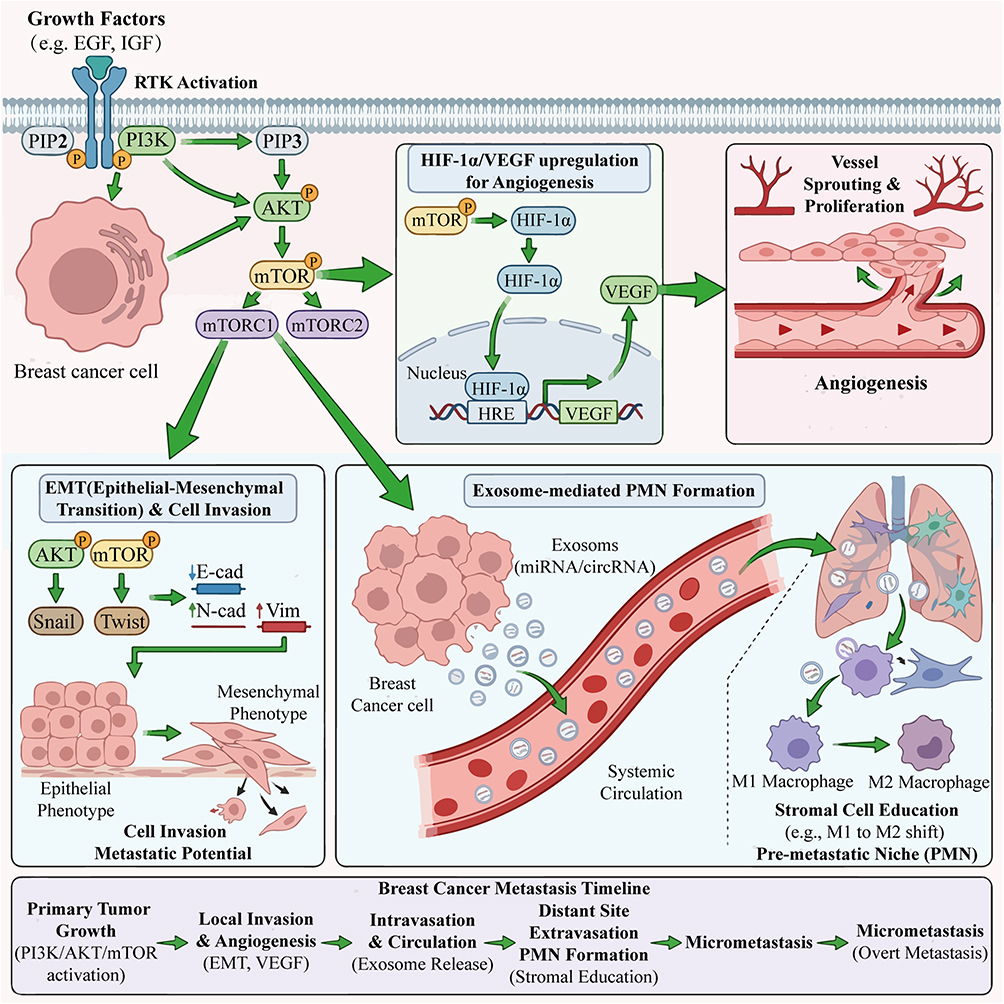 |
Figure 5 The PI3K/AKT/mTOR pathway drives breast cancer progression and metastasis via multiple mechanisms. Growth factors (eg, EGF, IGF) bind to receptor tyrosine kinases (RTKs) on breast cancer cells, activating the PI3K/AKT/mTOR pathway. Activated mTOR upregulates HIF-1α, which transcriptionally induces VEGF (via HRE binding). VEGF promotes vessel sprouting/proliferation, supporting tumor growth and nutrient supply. AKT/mTOR signaling upregulates Snail/Twist, suppressing E-cadherin and upregulating N-cadherin/vimentin. This induces EMT, enhancing cancer cell invasion and metastatic potential. Breast cancer cells secrete exosomes (carrying miRNA/circRNA) into systemic circulation. These exosomes educate stromal cells at distant sites, forming a PMN that supports subsequent metastasis. Created with BioRender.com. Abbreviations: RTK, receptor tyrosine kinase; EMT, epithelial-mesenchymal transition; HIF-1α, hypoxia-inducible factor 1α; VEGF, vascular endothelial growth factor; HRE, hypoxia response element; E-cad, E-cadherin; N-cad, N-cadherin; Vim, vimentin; PMN, pre-metastatic niche. |
Promotion of Epithelial-Mesenchymal Transition and Cell Invasion
The PI3K/AKT/mTOR signaling pathway plays a crucial role in driving epithelial-mesenchymal transition (EMT) and enhancing the invasive capabilities of tumor cells (Figure 5). EMT is a key step for tumor cells to acquire migratory and invasive characteristics, enabling distant metastasis. This pathway promotes the expression of a series of downstream transcription factors, such as Snail, Twist, and ZEB1, to suppress the expression of epithelial markers (such as E-cadherin) while upregulating mesenchymal markers (such as N-cadherin and Vimentin), thereby facilitating changes in cell morphology and function.35 In cervical cancer, ERBB3 significantly alters the expression profile of EMT-related genes and upregulates matrix metalloproteinase MMP9 through activation of the PI3K/AKT/mTOR pathway, thereby altering macrophage infiltration in the tumor microenvironment and ultimately affecting patient prognosis.35 Similarly, in Ewing sarcoma, the downstream mediator of the PI3K/AKT/mTOR pathway, ATF3, regulates the expression of chemokines such as CXCL8/IL-8, influencing not only the invasiveness of tumor cells but also reshaping the immune cell phenotypes in the tumor microenvironment.11 These findings reveal that the PI3K/AKT/mTOR pathway, by regulating EMT programs, lays the cellular biological foundation for tumor cells to break through the basement membrane restrictions, enter the circulatory system, and undergo distant metastasis.
Remote Microenvironment Regulation Mediated by Exosomes
Tumor-derived exosomes are important carriers for the PI3K/AKT/mTOR signaling pathway to conduct remote communication and shape the pre-metastatic microenvironment (PMN) (Figure 5). These nano-sized vesicles carry proteins, lipids, and nucleic acids (such as miRNA, circRNA) from the parent tumor cells, capable of circulating through body fluids to reach distant organs and create suitable “soil” for tumor metastasis by “educating” local stromal cells.36 For instance, in NSCLC, tumor-derived exosomes can be engineered into targeted delivery systems, loaded with drugs such as ginsenoside Rb1, which inhibit the PI3K/AKT/mTOR signaling pathway while reshaping the immune microenvironment (such as promoting M1 macrophage polarization), thereby overcoming immune therapy resistance and inhibiting tumor metastasis.37 In glioblastoma, ginsenoside-derived C3DM inhibits tumor growth by downregulating the EGFR/PI3K/AKT/mTOR signaling pathway and modulating the acidification state of the tumor microenvironment, enhancing T cell function.38 Additionally, exosome-mediated circRNA delivery also plays an important role in regulating the tumor microenvironment, influencing various processes such as tumor immunity, metabolism, angiogenesis, and EMT.36 Therefore, targeting tumor exosomes and their associated PI3K/AKT/mTOR signaling molecules provides new strategies for intervening in tumor metastasis.
Treatment Strategies Targeting the PI3K/AKT/mTOR Pathway and Its Microenvironment Interaction Interface
Efficacy and Limitations of Existing Pathway Inhibitors (Such as Alpelisib, Everolimus)
Existing PI3K/AKT/mTOR pathway inhibitors such as Alpelisib (a PI3Kα-specific inhibitor) and Everolimus (an mTOR inhibitor) have demonstrated clear clinical value in breast cancer treatment, but they have significant limitations (Figure 6 and Table 1). Alpelisib combined with fulvestrant can extend progression-free survival (PFS) in patients with PIK3CA mutation-positive HR+/HER2- advanced breast cancer, but toxicities such as hyperglycemia and rash limit its application.39 Everolimus inhibits tumor growth by blocking the mTORC1 complex, but feedback activation of AKT and MAPK pathways leads to resistance.40 In renal cell carcinoma studies, mTOR inhibitors can upregulate TFE3 expression, indirectly inhibiting p-Akt, but monotherapy efficacy is affected by the infiltration of immunosuppressive macrophages in the microenvironment.26 Additionally, PI3K/AKT/mTOR inhibitors generally have significant tissue distribution differences and insufficient targeting, with limited regulatory effects on metabolic reprogramming in the microenvironment (such as lactic acid accumulation).4
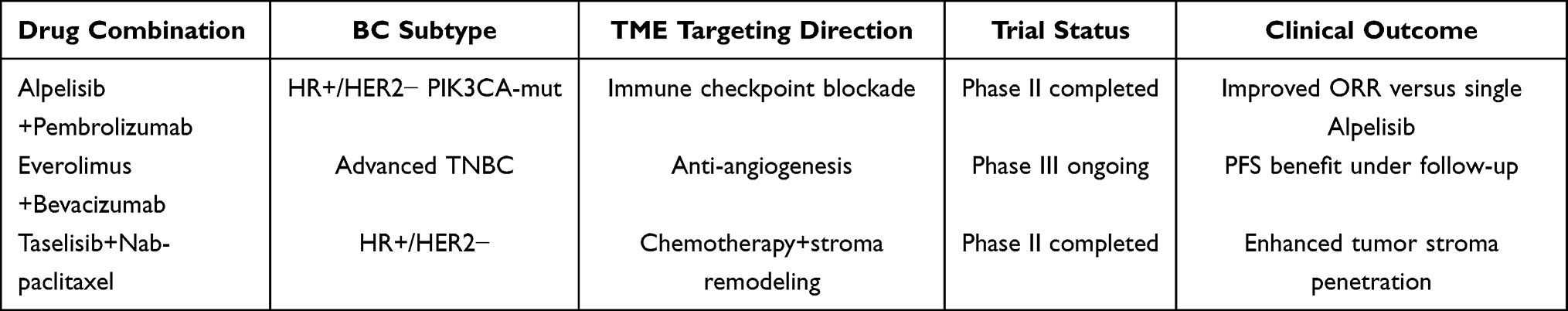 |
Table 1 Summary of BC Clinical Trials Combining PI3K/AKT/mTOR Inhibitors and TME-Targeted Therapies |
 |
Figure 6 Therapeutic strategies targeting the PI3K/AKT/mTOR pathway in PIK3CA+ HR+ breast cancer: single agents (efficacy and challenges) and synergistic combinations. 1. Single agent inhibitors: PI3Kα inhibitors (eg, Alpelisib) target PI3K to block pathway signaling, extending progression-free survival, which induces hyperglycemia and insulin feedback. And mTORC1 inhibitors (eg, Everolimus) suppress mTORC1 activity, which triggers a feedback activation loop of AKT, leading to treatment resistance. 2. Synergistic combinations to overcome resistance and enhance efficacy: A Immunotherapy combinations: PI3K/AKT/mTOR inhibitors downregulate PD-L1 expression on tumor cells, combining with anti-PD-1 (eg, Pembrolizumab) enhances anti-tumor T cell activity. B. Anti-angiogenic combination: PI3K/AKT/mTOR inhibitors reduce vascular hyperpermeability, combining with anti-VEGF (eg, Bevacizumab) improves drug delivery to tumor cells. C. Chemotherapy and metabolic targeting: PI3K/AKT/mTOR inhibitors block autophagy and synergize with chemotherapy (e.g, Paclitaxel). Concurrent targeting of metabolic regulators (HK2, GLUT1) further enhances apoptosis induction. Created with BioRender.com. Abbreviations: PFS, progression-free survival; HR+, hormone receptor-positive; PD-L1, programmed death-ligand 1; PD-1, programmed death-1; VEGF, vascular endothelial growth factor; HK2, hexokinase 2; GLUT1, glucose transporter 1. |
Synergistic Anti-Tumor Effects of Combination with Immune Checkpoint Inhibitors
The combination of PI3K/AKT/mTOR pathway inhibition with immune checkpoint blockade (such as anti-PD-1/PD-L1) shows synergistic mechanisms: pathway inhibitors can downregulate PD-L1 expression in tumor cells, reducing immune evasion (Figure 6).21 In head and neck squamous cell carcinoma models, HER3 antibody blockade of PI3K/AKT/mTOR signaling simultaneously reverses the immunosuppressive microenvironment, significantly extending survival when combined with PD-1 inhibitors.23 In hepatocellular carcinoma, CXCL3 recruits M2-type macrophages through activation of the PI3K/AKT/mTOR pathway, while the mTOR inhibitor Torin1 can block this process and enhance CD8+ T cell infiltration.12 However, it should be noted that PI3Kδ inhibitors may deplete regulatory T cells, leading to increased autoimmune toxicity, necessitating optimization of dosing schedules.11 Preclinical data suggest that monitoring the degree of PI3K/AKT/mTOR pathway inhibition during combination therapy is necessary, as excessive inhibition may impair T cell function.5
Combination Strategies with Anti-Angiogenic Drugs or Chemotherapy
The combination of PI3K/AKT/mTOR inhibitors with anti-angiogenic drugs (such as bevacizumab) can synergistically improve tumor perfusion and drug delivery (Figure 6). In pancreatic cancer models, the PI3K inhibitor taselisib reduces microvessel density while increasing CD8+ T cell infiltration, significantly inhibiting tumor growth when combined with gemcitabine.14 In breast cancer, Alpelisib enhances the effect of anti-angiogenic drugs on vascular normalization by reducing VEGF secretion through inhibition of HIF-1α.7 Chemotherapeutic drugs such as paclitaxel can induce immunogenic cell death, while the mTOR inhibitor Everolimus enhances chemotherapy sensitivity by blocking the protective autophagy induced by chemotherapy in tumor cells.41 However, caution is warranted regarding the cumulative toxicity of triple combinations (chemotherapy + anti-angiogenic + pathway inhibitors), and a metronomic chemotherapy regimen is recommended to reduce adverse reactions.42
New Approaches Targeting Tumor Metabolism and Microenvironment Metabolic Reprogramming
Emerging strategies targeting metabolic reprogramming mediated by the PI3K/AKT/mTOR pathway include (Figure 6): ① inhibiting key glycolytic enzymes HK2 or GLUT1 to reverse microenvironment acidification (for example, in follicular lymphoma with high GLUT1 expression, lactic acid promotes M2 macrophage polarization, which is associated with POD24 progression43); ② regulating glutamine metabolism, where DDIT4 induces autophagy by inhibiting mTOR, reducing glutamine-dependent tumor growth;44 ③ targeting lipid metabolism, where SCD1 inhibitors block PI3K-AKT-mTOR signaling to inhibit lipid synthesis in head and neck squamous cell carcinoma, synergizing with chemotherapy.45 Ginsenoside C3DM reduces glioma microenvironment lactic acid levels and enhances T cell function by inhibiting the EGFR/PI3K/AKT/mTOR pathway.38 Additionally, TLR7 agonists promote M1 macrophage polarization through the PI3K/AKT/mTOR pathway, improving the immunosuppressive microenvironment.46
Patient Stratification and Efficacy Prediction Based on Microenvironment Biomarkers
Microenvironment characteristics can predict responses to PI3K/AKT/mTOR targeted therapy (Figure 6): ① immune cell characteristics: those with a high CD8+ T cell/Treg ratio are more likely to benefit from Alpelisib + PD-1 inhibitors;14 ② metabolic biomarkers: high GLUT1 expression is associated with resistance to mTOR inhibitors, while high HK2 expression suggests the need for combination with glycolysis inhibitors;43 ③ spatial multi-omics analysis: tumor budding regions in cervical cancer show excessive activation of the PI3K/AKT/mTOR pathway accompanied by immune suppression, indicating the need for early intervention.18 In pancreatic ductal adenocarcinoma, clustering analysis based on PI3K/AKT/mTOR pathway genes can classify patients into immune hot (Cluster B) and cold (Cluster A) types, guiding precision treatment selection.14 Dynamic changes in circulating tumor DNA detection of PIK3CA mutations can monitor Alpelisib resistance in real-time.47
Conclusion
The interaction of the PI3K/AKT/mTOR pathway with the tumor microenvironment plays a core role in the occurrence, development, and treatment resistance of breast cancer. Existing breast cancer-specific evidence confirms that this pathway not only regulates intrinsic proliferation and invasion of BC cells but remodels immune suppression, angiogenesis and stromal architecture; multiple mechanistic conclusions cited from other solid tumors remain preliminary and need further BC-specific preclinical validation. Distinct regulatory patterns across HR+/HER2-, HER2+, TNBC subtypes lead to divergent therapeutic responses to PI3K/mTOR inhibitors and combination regimens. Single-agent PI3K/mTOR blockade fails largely due to adaptive TME compensation and immune escape.
Future priority research directions are refined as follows: (1) Preclinical validation of cross-cancer derived TME regulatory mechanisms using BC organoid and patient-derived xenograft models; (2) Optimize subtype-oriented combination schemes of PI3K/mTOR inhibitors plus TME-targeted drugs according to Table 1 trial data; (3) Develop metabolism or immune-related TME biomarkers for dynamic resistance monitoring. Remaining clinical obstacles include combination drug overlapping toxicity and intratumoral TME heterogeneity. Collectively, subtype-focused pathway-TME crosstalk research facilitates individualized combinatorial therapy development for breast cancer.
AI Tools
In this manuscript, ChatGPT and DeepL were used for paraphrasing and grammatical corrections.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by Jiangsu Provincial Health Commission Geriatric Health Research Project (Grant No.: LKZ2025030 to W.Y.), Yancheng Health Commission Project (Grant No.: YK2023040 to W.X.).
Disclosure
The authors declare no competing interests.
References
1. Li H, Prever L, Hirsch E, Gulluni F. Targeting PI3K/AKT/mTOR signaling pathway in breast cancer. Cancers. 2021;13. doi:10.3390/cancers13143517
2. Mehraj U, Dar AH, Wani NA, Mir MA. Tumor microenvironment promotes breast cancer chemoresistance. Cancer Chemother Pharmacol. 2021;87:147–13. doi:10.1007/s00280-020-04222-w
3. Jiang M, Zhang K, Zhang Z, et al. PI3K/AKT/mTOR axis in cancer: from pathogenesis to treatment. MedComm. 2025;6:e70295. doi:10.1002/mco2.70295
4. Mafi S, Mansoori B, Taeb S, et al. mTOR-mediated regulation of immune responses in cancer and tumor microenvironment. Front Immunol. 2021;12:774103. doi:10.3389/fimmu.2021.774103
5. Alves CL, Ditzel HJ. Drugging the PI3K/AKT/mTOR pathway in ER+ breast cancer. Int J Mol Sci. 2023;24:4522. doi:10.3390/ijms24054522
6. Xiao W, Zhang G, Chen B, et al. Mutational landscape of PI3K-AKT-mTOR pathway in breast cancer: implications for targeted therapeutics. J Cancer. 2021;12:4408–4417. doi:10.7150/jca.52993
7. Garg P, Ramisetty S, Nair M, et al. Strategic advancements in targeting the PI3K/AKT/mTOR pathway for Breast cancer therapy. Biochem Pharmacol. 2025;236:116850. doi:10.1016/j.bcp.2025.116850
8. Naveed M, Malik A, Anjum H, Ijaz B. LncRNA MALAT1 expression regulates breast cancer progression via PI3K/AKT/mTOR pathway modulation. Biochem Genet. 2024;62:3421–3438. doi:10.1007/s10528-023-10592-6
9. Karami Fath M, Akhavan Masouleh R, Afifi N, et al. PI3K/AKT/mTOR signaling pathway modulation by circular RNAs in breast cancer progression. Pathol Res Pract. 2023;241:154279. doi:10.1016/j.prp.2022.154279
10. Hou Z, Ma W, Ren D, et al. Exportin-T promotes breast cancer progression via PI3K/AKT/mTOR signaling pathway. J Inflamm Res. 2025;18:6467–6481. doi:10.2147/JIR.S512905
11. Palombo R, Passacantilli I, Terracciano F, et al. Inhibition of the PI3K/AKT/mTOR signaling promotes an M1 macrophage switch by repressing the ATF3-CXCL8 axis in Ewing sarcoma. Cancer Lett. 2023;555:216042. doi:10.1016/j.canlet.2022.216042
12. Li Y, Liu T, Cai Z, et al. CXCL3 promotes liver cancer progression by modulating the tumor microenvironment via the PI3K/AKT/mTOR pathway. PLoS One. 2025;20:e0334639. doi:10.1371/journal.pone.0334639
13. Prabhu KS, Kuttikrishnan S, Mariyam Z, et al. PI3 K/AKT/mTOR pathway and its role in breast cancer stem cells. Naunyn Schmiedebergs Arch Pharmacol. 2025;398:16779–16795. doi:10.1007/s00210-025-04297-3
14. Xie P, Tan SY, Li HF, Tang HD, Zhou JH. Transcriptome data-based status of PI3K/AKT/mTOR pathway indicates heterogeneity and immune modulation in patients with pancreatic ductal adenocarcinoma. J Gene Med. 2024;26:e3570. doi:10.1002/jgm.3570
15. Christofides A, Katopodi X-L, Cao C, et al. SHP-2 and PD-1-SHP-2 signaling regulate myeloid cell differentiation and antitumor responses. Nat Immunol. 2023;24(1):55–68. doi:10.1038/s41590-022-01385-x
16. Oh MH, Sun IH, Zhao L, et al. Targeting glutamine metabolism enhances tumor-specific immunity by modulating suppressive myeloid cells. J Clin Invest. 2020;130:3865–3884. doi:10.1172/JCI131859
17. Guo Y, Xie YQ, Gao M, et al. Metabolic reprogramming of terminally exhausted CD8(+) T cells by IL-10 enhances anti-tumor immunity. Nat Immunol. 2021;22:746–756. doi:10.1038/s41590-021-00940-2
18. Choi Y, Park NJ, Le TM, et al. Immune pathway and gene database (IMPAGT) revealed the immune dysregulation dynamics and overactivation of the PI3K/Akt pathway in tumor buddings of cervical cancer. Curr Issues Mol Biol. 2022;44:5139–5152. doi:10.3390/cimb44110350
19. Jeong H, Lee J, Son JY, et al. ULK1 knockout suppresses pancreatic cancer progression by inhibiting autophagy and enhancing antitumor immunity. Exp Mol Med. 2025;57:2816–2832. doi:10.1038/s12276-025-01590-2
20. Wang C, Lin H, Zhao W, Liang Y, Chen Y, Wang C. MiR-26a-5p exerts its influence by targeting EP300, a molecule known for its role in activating the PI3K/AKT/mTOR signaling pathway in CD8+tumor-infiltrating lymphocytes of colorectal cancer. Cell Mol Biol. 2023;69:232–241. doi:10.14715/cmb/2023.69.12.37
21. Quan Z, Yang Y, Zheng H, et al. Clinical implications of the interaction between PD-1/PD-L1 and PI3K/AKT/mTOR pathway in progression and treatment of non-small cell lung cancer. J Cancer. 2022;13:3434–3443. doi:10.7150/jca.77619
22. Yu J, Qin B, Moyer AM, et al. Regulation of sister chromatid cohesion by nuclear PD-L1. Cell Res. 2020;30:590–601. doi:10.1038/s41422-020-0315-8
23. Wang Z, Goto Y, Allevato MM, et al. Disruption of the HER3-PI3K-mTOR oncogenic signaling axis and PD-1 blockade as a multimodal precision immunotherapy in head and neck cancer. Nat Commun. 2021;12:2383. doi:10.1038/s41467-021-22619-w
24. Leuzzi G, Vasciaveo A, Taglialatela A, et al. SMARCAL1 is a dual regulator of innate immune signaling and PD-L1 expression that promotes tumor immune evasion. Cell. 2024;187:861–881e832. doi:10.1016/j.cell.2024.01.008
25. Sun EG, Choi JN, Park MR, et al. FGFR4 promotes CAF activation through the CXCL10-CXCR3 axis in colon cancer. Cell Death Dis. 2025;16:424. doi:10.1038/s41419-025-07588-y
26. Hwang C, Kang YK, Kim JY, et al. TFE3/PI3K/Akt/mTOR axis in renal cell carcinoma affects tumor microenvironment. Am J Pathol. 2024;194:1306–1316. doi:10.1016/j.ajpath.2024.02.022
27. Yan W, Xiao GH, Wang LJ, Zhou Y, Yang F, Mou KH. CAFs activated by YAP1 upregulate cancer matrix stiffness to mediate hepatocellular carcinoma progression. J Transl Med. 2025;23:450. doi:10.1186/s12967-025-06325-5
28. DeLeon-Pennell KY, Barker TH, Lindsey ML. Fibroblasts: the arbiters of extracellular matrix remodeling. Matrix Biol. 2020;91-92:1–7. doi:10.1016/j.matbio.2020.05.006
29. Theofilis P, Sagris M, Oikonomou E, et al. Extracellular matrix remodeling biomarkers in coronary artery disease. Curr Top Med Chem. 2022;22:2355–2367. doi:10.2174/1568026623666221024091758
30. Zhang J, Chen Y, Xu H. Glioblastoma induces CAF-like astrocyte activation via the AKT/mTOR-SERPINH1/COL5A1 axis. Biomol Biomed. 2025;25:2737–2754. doi:10.17305/bb.2025.11898
31. Zhao L, Wang Z, Cheng P, Sheng G, Han M, Zheng Z. SMYD3-CDCP1 axis drives EMT and CAF activation in colorectal cancer and is targetable for oxaliplatin sensitization. Biomedicines. 2025;13:2737. doi:10.3390/biomedicines13112737
32. Wang M, Wang Y, Pan X, et al. Acquired resistance to immunotherapy by physical barriers with cancer cell-expressing collagens in non-small cell lung cancer. Proc Natl Acad Sci U S A. 2025;122:e2500019122. doi:10.1073/pnas.2500019122
33. Stanciu S, Ionita-Radu F, Stefani C, et al. Targeting PI3K/AKT/mTOR signaling pathway in pancreatic cancer: from molecular to clinical aspects. Int J Mol Sci. 2022;23:10132. doi:10.3390/ijms231710132
34. Mao QY, Wang XQ, Lin F, et al. Scorpiones, scolopendra and gekko inhibit lung cancer growth and metastasis by ameliorating hypoxic tumor microenvironment via PI3K/AKT/mTOR/HIF-1alpha signaling pathway. Chin J Integr Med. 2024;30:799–808. doi:10.1007/s11655-024-3803-8
35. Yang X, Zhu W. ERBB3 mediates the PI3K/AKT/mTOR pathway to alter the epithelial‑mesenchymal transition in cervical cancer and predict immunity filtration outcome. Exp Ther Med. 2023;25:146. doi:10.3892/etm.2023.11845
36. Li J, Zhang G, Liu CG, et al. The potential role of exosomal circRNAs in the tumor microenvironment: insights into cancer diagnosis and therapy. Theranostics. 2022;12:87–104. doi:10.7150/thno.64096
37. Jin X, Wuyun T, Zhang Y, Wang X, Zhao L. Overcoming acquired immunotherapy resistance in non-small cell lung cancer using ginsenoside Rb1-loaded, peptide-enhanced exosome delivery systems. J Nanobiotechnology. 2025;23:443. doi:10.1186/s12951-025-03456-1
38. Tang M, Deng H, Zheng K, He J, Yang J, Li Y. Ginsenoside 3beta-O-Glc-DM (C3DM) suppressed glioma tumor growth by downregulating the EGFR/PI3K/AKT/mTOR signaling pathway and modulating the tumor microenvironment. Toxicol Appl Pharmacol. 2023;460:116378. doi:10.1016/j.taap.2023.116378
39. Peng R, Zhan Y, Li A, Lv Q, Xu S. Research progress and development strategy of PI3K inhibitors for breast cancer treatment: a review (2016-present). Bioorg Chem. 2024;153:107934. doi:10.1016/j.bioorg.2024.107934
40. Dong C, Wu J, Chen Y, Nie J, Chen C. Activation of PI3K/AKT/mTOR pathway causes drug resistance in breast cancer. Front Pharmacol. 2021;12:628690. doi:10.3389/fphar.2021.628690
41. Liu QQ, Li YQ, Cheng WM. Research progress on the mechanism of PI3K/AKT/mTOR signaling pathway in multiple myeloma --review. Zhongguo Shi Yan Xue Ye Xue Za Zhi. 2024;32:292–296. doi:10.19746/j.cnki.issn.1009-2137.2024.01.047
42. Wang X, Chen X, Zhu J, Li S. Quantifying the impact of metronomic chemotherapy chemo-switch regimen and the sequencing of chemotherapy and radiotherapy on pancreatic ductal adenocarcinoma treatment. J Theor Biol. 2025;599:112033. doi:10.1016/j.jtbi.2024.112033
43. Deng Y, Ma J, Zhao S, Yang M, Sun Y, Zhang Q. Expression of glucose transporter-1 in follicular lymphoma affected tumor-infiltrating immunocytes and was related to progression of disease within 24 months. Transl Oncol. 2023;28:101614. doi:10.1016/j.tranon.2022.101614
44. Jiao Y, Xiang Y. A review of the participation of DDIT4 in the tumor immune microenvironment through inhibiting PI3K-Akt/mTOR pathway. Front Oncol. 2025;15:1595463. doi:10.3389/fonc.2025.1595463
45. Hsu CM, Yang MY, Chang SF, Su HC. Targeting Stearoyl-CoA Desaturase 1 through PI3K-AKT-mTOR signaling in head and neck squamous cell carcinoma. OTO Open. 2025;9:e70143. doi:10.1002/oto2.70143
46. Guo L, Zhou X, Zhang Z, Zhang X. TLR7 modulates glioblastoma progression through PI3K/AKT/mTOR pathway and immune microenvironment remodeling. Ir J Med Sci. 2025;195:597–607. doi:10.1007/s11845-025-04208-8
47. Hao C, Wei Y, Meng W, Zhang J, Yang X. PI3K/AKT/mTOR inhibitors for hormone receptor-positive advanced breast cancer. Cancer Treat Rev. 2025;132:102861. doi:10.1016/j.ctrv.2024.102861
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.