")
Back to Archived Journals » Research and Reports in Biodiversity Studies » Volume 4
The impact of CO2 emissions on 'nuisance' marine species
Authors Hall-Spencer J, Allen R
Received 29 May 2015
Accepted for publication 1 September 2015
Published 5 November 2015 Volume 2015:4 Pages 33—46
DOI https://doi.org/10.2147/RRBS.S70357
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr David Lane
Video abstract presented by Professor Hall-Spencer and Ro Allen.
Views: 1013
Jason M Hall-Spencer, Ro Allen
Marine Biology and Ecology Research Centre, School of Marine Science and Engineering, Plymouth University, Plymouth, Devon, UK
Abstract: Anthropogenic CO2 emissions are being taken up from the atmosphere by the oceans, increasing the availability of dissolved inorganic carbon but reducing both the carbonate saturation and pH of seawater. This ocean acidification affects biological processes in a wide range of marine taxa. Here, we assess the likely responses of 'nuisance' species to ocean acidification, meaning those organisms that have undesirable effects from a human perspective. Based on a synthesis of evidence available to date, we predict increased growth and toxicity in harmful algal bloom species, and a significant increase in invasive algae in response to increased CO2 availability. Blooms of stinging jellyfish are also expected to increase since they are highly resilient to acidification. The effects of ocean acidification on invasive molluscs (eg, oyster drills), damaging echinoderms (eg, crown-of-thorns starfish), and a wide range of nuisance taxa will vary depending on species and location. In the USA, for example, the invasive crab Carcinus maenas is resilient to projected increases in CO2 and its impact on marine communities is expected to increase since it feeds on organisms that respond to ocean acidification with weaker defensive traits and lower recruitment. Conversely, the Red King Crab, Paralithodes camtschaticus, is adversely affected by acidification and so is expected to die back in the Barents Sea which it has invaded. Overall, we suspect that there will be an increase in nuisance species, as many have traits that are resilient to the combined warming and acidification caused by rising CO2 levels; region-specific assessments are needed to understand responses of nuisance species in local habitats. Finally, we highlight the need for targeted studies of the effects of global change on particularly harmful marine taxa such as the seaweed Caulerpa taxifolia, the starfish Asterias amurensis, several invasive ascidians, and the lionfish Pterois volitans.
Keywords: ocean acidification, anthropocene, multiple impacts, harmful algal blooms, invasive species
Introduction
In 2013, atmospheric carbon dioxide reached 400 ppm at the long-term monitoring station of Mauna Loa, higher than at any time in the past 800,000 years and up from 280 ppm in the pre-industrial period of the 1700s.1 This rapid increase in atmospheric CO2 levels is causing ocean acidification, since 30% of human CO2 emissions have been absorbed by surface waters, driving down seawater pH and making it increasingly corrosive to calcium carbonate.2
A rapidly growing body of research indicates that ocean acidification will severely disrupt marine ecosystems, since it alters the balance of success between competing organisms.3 Ocean acidification can impact a wide range of processes across marine taxa, including photosynthesis, acid-base homeostasis, calcification, and behavior.4–7 Dissolved inorganic carbon is a resource for photosynthesis, and so increases in CO2 can benefit groups such as diatoms, brown algae, and seagrasses.8,9 Falling carbonate saturation is expected to degrade biogenic reef habitats and negatively impact calcified organisms.10–13 Organisms incapable of acid-base homeostasis may suffer metabolic depression, while those that are capable of regulation incur associated energetic costs.5 Observations show that there will be winners as well as losers as the oceans acidify, just as there were in previous high-CO2 mass extinction events.6,14 There is also a growing realization that secondary effects, mediated through changes in habitat and species’ interactions, will drive ecosystem change.15
Syntheses of the effects of ocean acidification have focused on key groups, such as photosynthetic organisms or on commercially important species, and show that even closely related species can have very different responses to ocean acidification.9–16 Over-arching meta-analyses conclude that ocean acidification will drive substantial changes in marine ecosystems and cause long-term biogeographic shifts because some organisms gain a competitive advantage over others.5,13 We were prompted to undertake this review since we have noticed that invasive algae and jellyfish thrive as CO2 levels ramp up around volcanic seeps in the Mediterranean (Figures 1 and 2). Here, we consider the possibility that ocean acidification may preferentially benefit ‘nuisance’ organisms, ie, those organisms that have undesirable effects from a human perspective. Such organisms include invasive species, which are ecologically disruptive non-native organisms, as well as native organisms such as harmful algal blooms, jellyfish swarms, and crown-of-thorn starfish outbreaks. Given that ocean acidification is occurring alongside other stressors, such as global warming and eutrophication,17 we include those investigations that consider these combined drivers of global change.
Algae
Algae can benefit from ocean acidification since increased availability of CO2 and HCO3- can stimulate photosynthesis.18 As some primary producers are better able to capitalize on increasing carbon availability than others, this is expected to alter marine communities.15,19 Invasive algae are expected to benefit in competitive interactions under acidified conditions since they tend to be non-calcareous, have wide thermal and salinity tolerances, are highly fecund, grow rapidly, and are often parthenogenic.9 As an example, elevated CO2 is expected to extend the range of the invasive warm water alga Neosiphonia harveyi since experiments have shown that the increased availability of dissolved inorganic carbon allows this seaweed to cope with colder temperatures, so it is expected to extend into higher latitude regions where native species die back due to warming.9 This carbon-boost to invasive algae may be coupled with temperature-driven range extensions of warm water herbivorous fish that can remove temperate macroalgae, facilitating the spread of warm water invasive species, such as N. harveyi.20
Surveys of Mediterranean CO2 seeps have repeatedly shown that coralline algae become less common as CO32− levels fall, whereas brown seaweeds and invasive Asparagopsis taxiformis (Figure 1) and Caulerpa spp. proliferate as CO2 and HCO3- levels rise.21,22 A strength of using these natural analogs for the effects of ocean acidification is that they reveal the long-term response of marine communities in areas where CO2 levels have been elevated for 100s of years. Shorter term mesocosm experiments (<1 year) allow tighter control of CO2 doses and have shown that Caulerpa taxifolia are resilient to simulated acidification. This is a concern as C. taxifolia has spread rapidly causing significant alterations to native algal communities worldwide.23 Mesocosm work on the ocean acidification response of invasive canopy-forming algae such as Sargassum muticum and Undaria pinnatifida (Table 1) also raises concerns since they may benefit from increased CO2 levels thereby decreasing native algal species’ richness and altering communities.24 S. muticum settlement increases when algal diversity is low, suggesting that less diverse communities will be more vulnerable to species’ invasions.25 Field sites with the elevated CO2 levels predicted this century show approximately 5% reduction in algal community diversity accompanied by wholesale shifts in community structure due to losses of calcified algal cover, with invasive algae look set to fill niches left by intolerant native species.21,26 Mesocosm work shows that the physiological performance of macroalgal assemblages invaded by S. muticum under near-future conditions may be superior to non-invaded assemblages, potentially accelerating the proliferation of the invasive species.27
 | Figure 1 Invasive Asparagopsis taxiformis thrive in endemic Posidonia oceanica seagrass beds at CO2 seeps in the Mediterranean (A). Fish farmers are facing increased losses due to blooms of spiky phytoplankton, such as this Chaetoceros sp., which are resilient to rising CO2 levels but damage gills and can kill fish (B). |
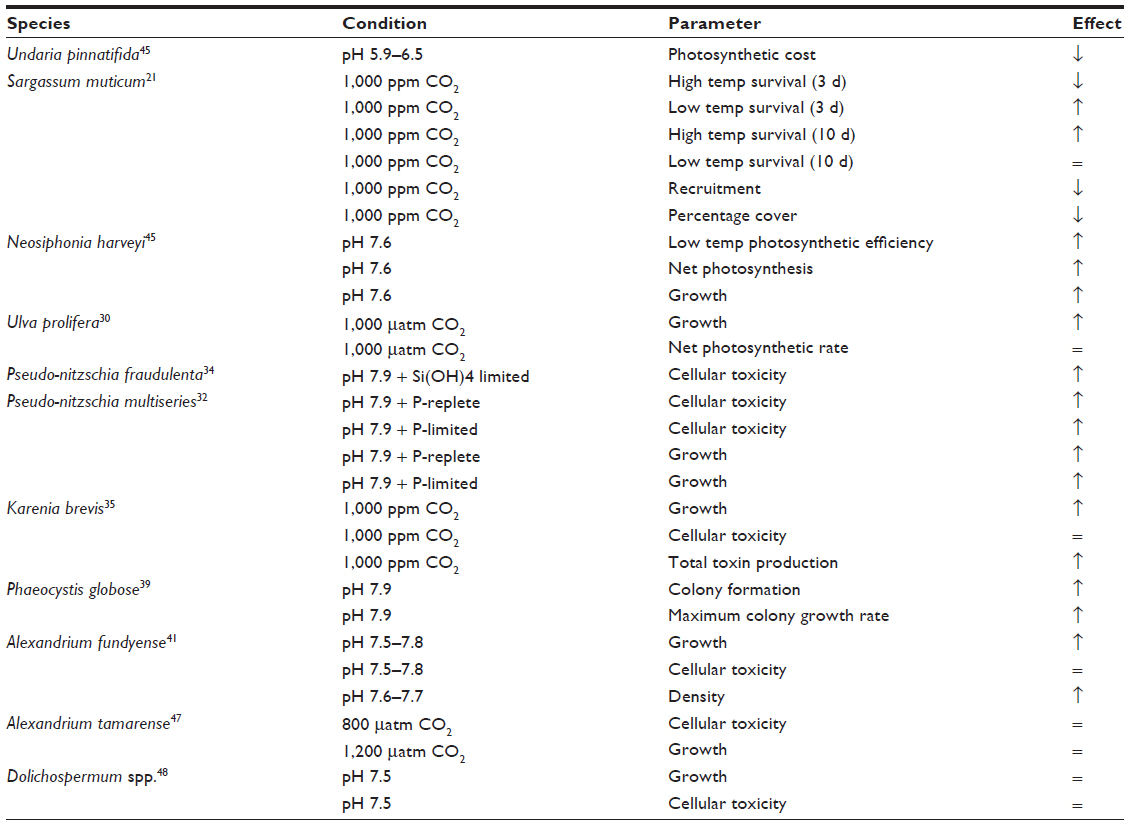 | Table 1 Algal responses to ocean acidification |
Ulva spp. have become notorious in recent years as they have started to regularly form green tides that rot and kill marine life, reducing the amenity value of beaches.28 In the Yellow Sea, for example, blooms of Ulva prolifera are causing large-scale disruption;29 experiments show that ocean acidification is likely to increase U. prolifera growth, with rising atmospheric CO2 levels expected to increase the intensity of green tide events.30
Harmful algal blooms cause significant problems through mechanical damage (such as to fish gills), toxic effects, and competition for resources. Increased availability of dissolved inorganic carbon risks increasing the toxicity of harmful algal blooms.31 Tests show that the Pseudo-nitzschia diatoms produce more of the toxins that cause amnesic shellfish poisoning with increases in pCO2 in nutrient limited conditions.32–34 Similarly, Karenia brevis, which causes neurotoxic shellfish poisoning, grew significantly faster and maintained the same toxin production per cell in laboratory simulations of ocean acidification.35 As increased growth rates increase the likelihood of blooms becoming nutrient limited, ocean acidification is expected to increase K. brevis bloom toxicity and the severity of its mechanical effects.36
Simulated acidification benefits Alexandrium minutum which can form harmful blooms,37 and genetic adaptation accentuates their positive responses to reduced pH.38 In the harmful algal bloom forming alga Phaeocystis globosa, elevated CO2 stimulates colony formation and increases maximal colonial growth rates although this may render them more susceptible to viral infection.39,40 Despite widespread inter- and intra-specific variability in responses (Table 1),41 trends of increased growth, toxicity, and synergistic effects are likely to increase the severity of future harmful algal bloom events. Furthermore, temperature stratification induced nutrient scarcity and elevated CO2 may interact to increase the likelihood of blooms becoming nutrient limited, subsequently leading to increased toxin production.36 This has implications for aquaculture and fisheries; the synergistic effects of simulated harmful algal bloom exposure and elevated CO2 resulted in increased mortality and decreased development and growth in the scallop Argopecten irradians, versus exposure to these stressors independently.37 The relationship between harmful algal blooms and climate change remains a research priority.42,43 Overall, ocean acidification is expected to profoundly alter benthic and pelagic algal communities,21,44 and is expected to benefit invasive species, increasing their range, diversity, and abundance.9,45–48
Jellyfish
‘Jellyfish’ refers to gelatinous zooplankton, including scyphozoans, cubozoans, and ctenophores. Many scientists are convinced that they are on the increase due to marine ecosystem degradation, for example due to eutrophication and overfishing.49 Jellyfish stings can be painful and even fatal to humans, impacting tourism and fish farm industries.50,51 Blooms of the invasive ctenophore Mnemiopsis coincided with the collapse of Black Sea and Caspian Sea pelagic fisheries,52–54 and may play a role in suppressing the population recovery of fish planktivores.52 Although anemones and soft corals are thought to be resilient to ocean acidification, to date little research effort has been committed to investigating the responses of jellyfish,55–57 which is surprising considering their economic impact.
Aurelia labiata is highly resilient to acidification in culture conditions, although the acidification slows statolith development.58 A similar study also found smaller statoliths and retarded asexual polyp reproduction at pH 7.6 in Alatina nr mordens.59 As statoliths are not exposed to seawater, it is likely that reduced statolith size results from the stress of living in high CO2 waters which alters energy budgets in many marine taxa.3,60 Attrill et al linked ocean acidification to increased jellyfish nematocyst abundance in the North Sea in plankton surveys, suggesting negative impacts on calcareous plankton as the indirect cause of the increase.61 A follow-up study analyzed coelenterate tissue occurrence instead of nematocyst presence and did not find a significant relationship between pH and jellyfish abundance.62 The contribution of ocean acidification to jellyfish blooms is clearly a matter of debate,63 yet as a group they seem resilient to the levels of acidification expected this century (Table 2). Our own observations show jellyfish can survive well in high abundance at CO2 seeps in the Mediterranean (Figure 2).
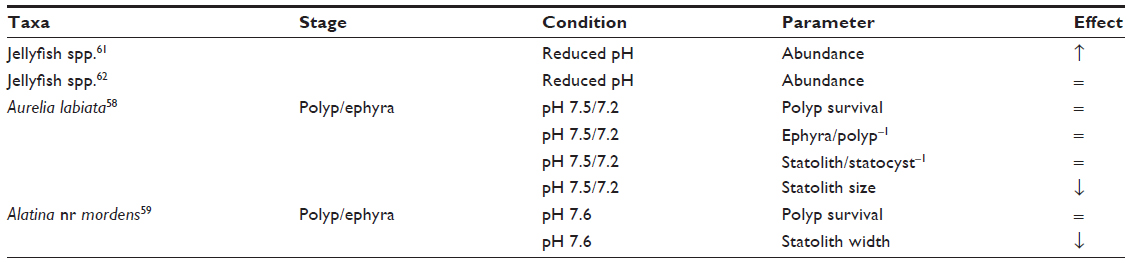 | Table 2 Jellyfish responses to ocean acidification parameters |
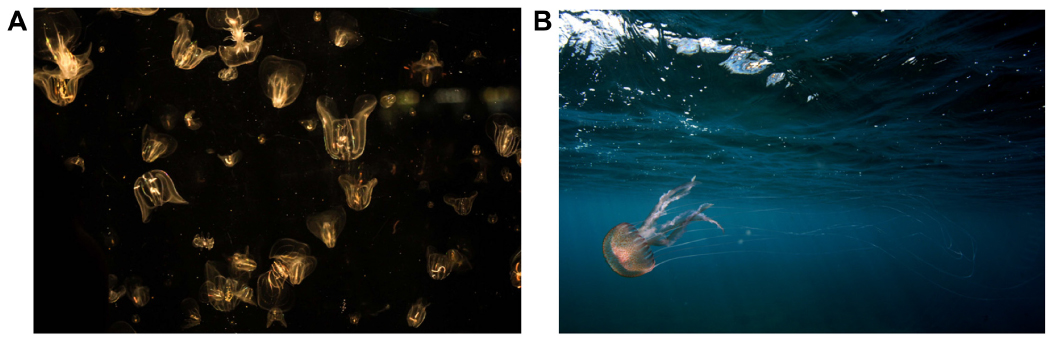 | Figure 2 Invasive Mnemiopsis contributed to the collapse of Black Sea and Caspian Sea pelagic fisheries (A). Native Pelagia noctiluca at CO2 seeps off Vulcano; jellyfish and anemones thrive where the water is too corrosive for hard corals (B). |
Invasive molluscs
Laboratory studies show that calcifying marine molluscs are predominantly negatively affected by ocean acidification, with adverse effects spanning calcification, growth, respiration, and survival (Table 3).5,64,65 Field observations in the Mediterranean show that mollusc recruitment and shell thickness are significantly reduced across a range of taxa under acidified conditions.12,66–68 In 2006, the oyster farming industry in the Northeast Pacific struggled to cope with the adverse effects of CO2-rich upwelling waters and now have to manipulate the carbonate chemistry of their hatcheries to help the spat survive.16 So what evidence is there for the effects of ocean acidification on highly invasive marine molluscs?
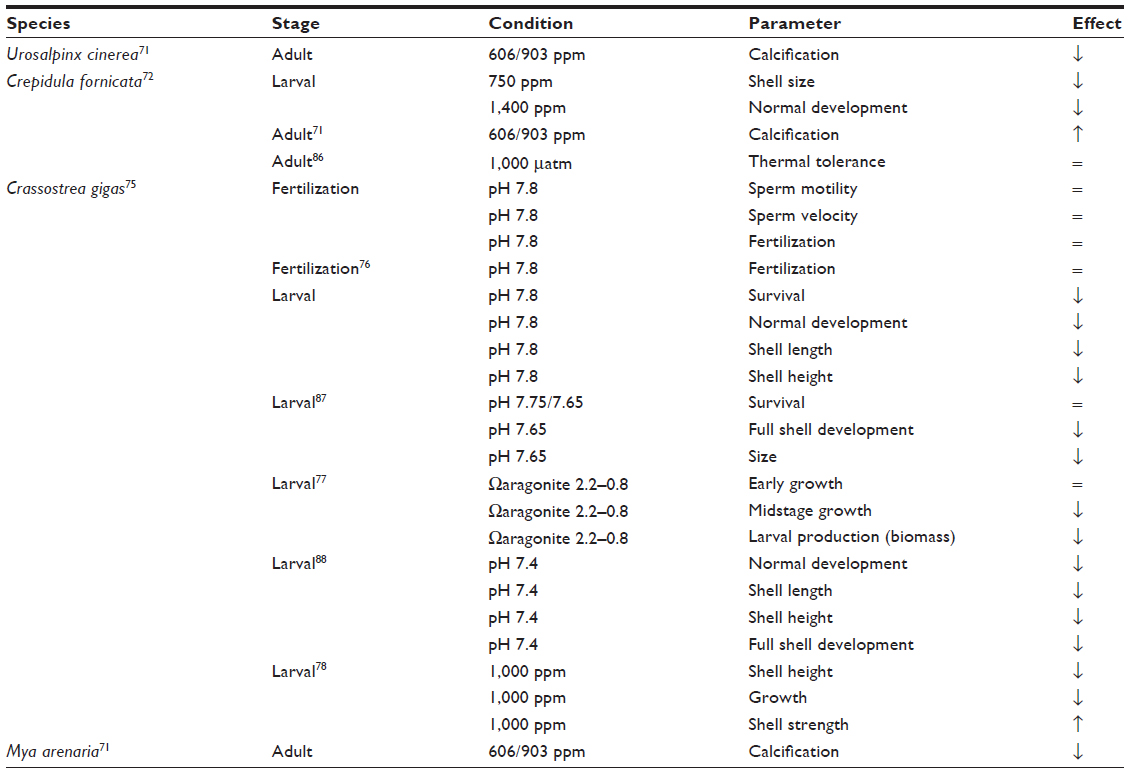 | Table 3 Responses of invasive molluscs to ocean acidification |
The American slipper limpet, Crepidula fornicata, is now widespread in Europe and is considered one of the 100 worst invasive species.69 In the Atlantic it can reduce growth and survival in Mytilus edulis.70 Ocean acidification may benefit adult C. fornicata since calcification increased at 606 ppm and 903 ppm CO2, with negative responses only observed at 2,856 ppm.71 However, larval shell growth was significantly reduced at 750 ppm CO2.72 As delayed larval development increases vulnerability to predation this may reduce recruitment. So it is hard to predict how ocean acidification will affect the spread of C. fornicata, it is a very robust organism as an adult yet the costs of coping with hypercapnia at the larval stage may impair recruitment.
The invasive NE Pacific oyster Crassostrea gigas has been introduced for aquaculture worldwide and has subsequently spread, becoming a threat to native bivalves such as the Sydney Rock oyster Saccostrea glomerata in Southern Australia.73,74 The fertilization of an invasive strain of C. gigas was not affected at pH 7.8,75,76 but larval mortality increased dramatically in acidified treatments with larval developmental abnormalities increasing from <20% in normal conditions to 54% at pH 7.8 and significantly reduced larval shell growth after 6 days.76 Natural reductions in carbonate saturation correlate with decreases in larval production, suggesting that ocean acidification may reduce C. gigas recruitment.77 In aquaria, juvenile and adult growth rates of the invasive oyster also decrease.78 However, the direct responses of C. gigas to ocean acidification vary as resilient genotypes are present in current populations which may facilitate adaptation to future ocean acidification.78 S. glomerata fertilization and early life history are more severely impacted than in C. gigas so the impact of invasive species on native oysters may increase due to ocean acidification.79
An invasive predatory snail from the Northwest Atlantic (Urosalpinx cinerea) is spreading in the Northeast Atlantic and the Northeast Pacific where it is impacting oyster and scallop aquaculture.80,81 Ries et al found a linear reduction in calcification in U. cinerea in response to rising pCO2 with significantly reduced adult growth at 606 ppm.71 Yet although it appears to respond negatively to ocean acidification in aquaria, its harmful effects in the wild may increase. Sanford et al found that ocean acidification caused a 29%–40% reduction in growth of the native oyster Ostrea lurida and that U. cinerea ate significantly more oysters in acidified treatments.82 This effect may be the result of reduced energetic value in prey species, increased energetic requirement in the predator, reduced prey handling time, or any combination of these points.83
The role of invasive predatory gastropods may increase in response to ocean acidification if recruitment rates are maintained as per capita consumption rates can increase.78,82,83 This may enhance disruption to bivalve fisheries and the destruction of native biogenic habitats by invasive snails.84 Indeed, bivalve aquaculture may increase in importance as global fish landings are expected to decrease significantly in response to climate stressors.85 Literature is lacking regarding the effect of ocean acidification in predatory gastropod recruitment, and this gap should be filled.65
Invasive crustaceans
Laboratory experiments indicate that although crustaceans may be one of the most resilient animal taxa to ocean acidification, ~30% of species studied to date are negatively impacted,5,13 which helps explain reduced crustacean diversity in acidified conditions at CO2 seeps.10,66,89 The resilience of certain crustaceans to near-future ocean acidification has been attributed to their high capacity for extracellular pH regulation.13,90
The barnacle Amphibalanus improvisus is native to the Atlantic Ocean, but has spread through fouling of artificial structures,91 vessels,92 and larval transport in ballast water, leading to a global distribution. It competes with native organisms for space, and may alter habitats since it promotes macroalgal colonization and creates refugia in empty shells.93,94 Furthermore, the species can foul native or co-invasive bivalves, altering predator-prey interactions.95 This barnacle is particularly resilient to the effects of ocean acidification, with no significant effects observed on larval growth and development, or juvenile growth and shell strength, even at pH conditions far below 2100 projections.96–98 Furthermore, intra-specific variability in response exists, suggesting populations are genotypically diverse, which may increase adaptive capacity in response to ocean acidification.97 The studies demonstrate the exceptional capacity of A. improvisus to persist under near-future acidified conditions. Contrastingly, Semibalanus balanoides show retarded embryonic development and reduced adult survival at pH 7.7.99 Thus, in certain situations, invasive barnacles may gain a competitive advantage over native barnacles due to ocean acidification.
The crab Carcinus maenas is native to the Northeast Atlantic but has established populations in Australia, Southern Africa, and North America where it has disrupted native communities and impacted mollusc fisheries.100–103 These crabs cope well with elevated CO2 by regulating extracellular pH, although the associated energetic costs,90,104,105 may have knock-on effects.106 Chemoreception may be impaired and limit predation in decapods as a result of ocean acidification,107 and mollusc prey may be less well defended due to ocean acidification.71,108,109 Despite this, no response in prey handling time or per capita consumption rate has been observed in C. maenas in response to ocean acidification,105,106 so we expect the impact of this invasive crab will increase as they are remarkably resilient.
The Red King Crab, Paralithodes camtschaticus, has invaded the Barents Sea, where it has disrupted native benthic communities.110–112 It is not as resilient as C. maenas to ocean acidification since larval survival and growth are reduced at pH 7.7 and juvenile survival and growth are dramatically reduced at pH 7.8.113,114 Thus, ocean acidification may have severe effects on P. camtschaticus populations, which would alleviate its effects in its invasive range but could also negatively impact crab fisheries in its native range, although the role of adaptation remains to be considered.114
Despite crustaceans being one of the most resilient taxonomic groups to near-future ocean acidification, there is such a high level of inter-specific variance in responses that the effects of ocean acidification on invasive crustaceans should be considered on a case-by-case basis (Table 4).
 | Table 4 Responses of invasive crustaceans to ocean acidification |
Echinoderms
Sea urchins appear to be especially vulnerable to ocean acidification, as demonstrated in laboratory and field studies.115–117 Centrostephanus rodgersii (Figure 3) recently spread to Tasmania due to a temperature-driven range extension. It is a prolific algal grazer, creating and maintaining coralline algal dominated ‘barrens’ with reduced biodiversity such that it now threatens commercially important abalone.118,119 Although acidification impedes embryo development, temperature appears to have a stronger effect on the early life history of C. rodgersii so the benefits of rising ocean temperature may outweigh the negative effects of acidification,120 as shown in other sea urchins.121 Thus, the impact of C. rodgersii in Tasmania is likely to be maintained, despite ocean acidification, since genotypes resilient to both acidification and warming are present in C. rodgersii populations of Southeast Australia.122 Sticking with Tasmania for a moment, the invasive cushion star Patiriella regularis may recede from this island as the oceans acidify, since its growth and survival are impaired in mesocosm experiments with elevated CO2 (Table 5).
 | Figure 3 The spread of Centrostephanus rodgersii sea urchins due to sea surface warming has created ‘Barrens’ in Tasmania (A).119 Acanthaster planci starfish seem resilient to rising CO2 levels but their outbreaks result in mass coral mortality on the Australian Great Barrier Reef (B).123 |
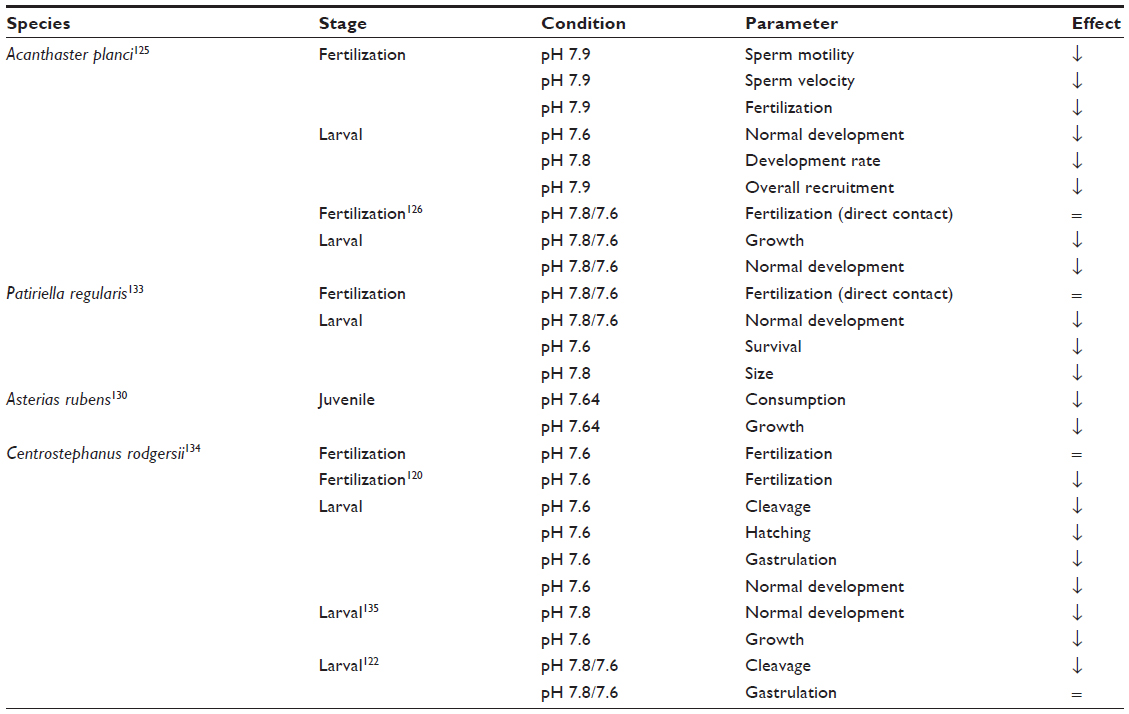 | Table 5 Examples of responses of echinoderms to ocean acidification |
Crown-of-thorns starfish, Acanthaster planci, eat coral and their outbreaks can cause >50% coral mortality at affected sites;123 they also increase coral vulnerability to disease (Katz et al).124 Uthicke et al found that elevated CO2 reduced A. planci sperm motility and estimated that fertilization success reduced by 29% at pH 7.8 and by 75% at pH 7.6.125 The rates of larval development and growth are also reduced under near-future acidified conditions.125,126 Retarded development led to an extended duration in the plankton, where larvae were vulnerable to predation, likely reducing recruitment.125 Compounding the effect of ocean acidification on A. planci early life history stages, near-future acidified conditions affect crustose coralline algae and biofilms, reducing the successful settlement of A. planci on these substrata by an average of 36%.125 Based on the effects of fertilization and settlement alone, A. planci recruitment is projected to reduce by approximately 50% under near-future acidified conditions.125
Thus, ocean acidification may reduce the impact of echinoderm outbreaks (Table 5); however, responses must be considered in conjunction with other factors such as food availability and temperature, which may be more influential than pCO2 levels in seawater in determining the response of A. planci outbreaks in future.127,128 There is clearly room for further investigation – for example, the starfish Asterias rubens is a significant pest to Irish bivalve fisheries,129 and responds to ocean acidification with reduced consumption, growth,130 and increased vulnerability to pathogens.131 But, little work has been conducted on the congeneric North Pacific seastar, Asterias amurensis, despite the fact that it is considered to be one of the most disruptive invasive species in the world.132
Ascidians
Invasive ascidians are having significant impacts worldwide, outcompeting native species, smothering benthic habitats, and biofouling maritime infrastructure such as vessels, marinas, and aquaculture facilities.136–138 Few studies have investigated the impact of ocean acidification on ascidians. Ciona intestinalis and Ascidiella aspersa appear to benefit from increased larval development rates and survival under near-future acidified conditions,139 which could enhance their recruitment. Field data from CO2 seep studies describe variable responses. Fabricius et al found that ascidian abundance decreased at elevated CO2 conditions in Papua New Guinea.10 At comparable CO2 seeps in Italy, Donnarumma et al found that ascidians dominated faunal assemblages at extremely low pH sites (6.57) on seagrass beds.140 Peck et al investigated biofouling communities and found ascidian abundance dramatically increased in response to a 100-day exposure to pH 7.7.141 Thus, certain ascidians can be expected to benefit from ocean acidification,139 although competitive interactions will determine their site-specific success. As with other invasive groups of organisms, there is significant scope for further investigation of ascidian responses to ocean acidification, due to their environmental impacts and threats to aquaculture.
Fish
Fish have high metabolic rates and a well-developed capacity for extracellular pH regulation.5 Their early life stages are often resilient to elevated CO2 and adult fish survive well in tests.13,142 However, ocean acidification has significant behavioral and sensory impacts attributed to impacts on the nervous system of several species of fish.143,144 Dixson et al (2010) found that settlement-stage larvae could not discriminate between predator and non-predator chemical cues at pH 7.8,145 and studies at CO2 seeps demonstrate altered predator cue responses in juvenile fish,146 increasing vulnerability to predation. Visual and auditory predator cue responses can also be compromised by near-future ocean acidification.144,147,148
Although no work on ocean acidification has been carried out on invasive fish, we can consider what current knowledge might mean for Pterois volitans, a lionfish native to the Indo-Pacific that has invaded the Atlantic coast of North America.149 P. volitans has had profound impacts on food webs and on prey fish populations in the Caribbean.150,151 Lesser and Slattery consider the invasive lionfish has played a key role in shifting benthic communities from coral to algal dominance by reducing herbivorous fish populations (Figure 4).152 On balance, negative effects on tropical coral reef prey species are expected to outweigh effects on predatory species.153
 | Figure 4 The invasive lionfish Pterois volitans eats native fish in the Caribbean (A). Caribbean reefs are shifting toward algal dominance, partially through the suppression of herbivorous fish (B). |
We fear that predation pressure from P. volitans could maintain low populations of herbivorous fish.151,154 This could combine with the adverse effects of ocean acidification on grazing sea urchins and the benefits of elevated CO2 on fleshy seaweeds and contribute to an ongoing decline in coral reefs due to algal overgrowth, especially in areas affected by eutrophication.155 Studies of the effects of ocean acidification on P. volitans and its ecological interactions would be worthwhile considering the rapid spread of this species.
Conclusion
There is plenty of evidence that ocean acidification can be expected to exacerbate the problems associated with ‘nuisance’ species thereby degrading ecosystem services. The traits that help many invasive organisms succeed, such as wider physiological tolerance and a reproductive advantage in the face of multiple stressors,156 are traits that look set to help them in an acidifying ocean.
We expect that the impacts of harmful bloom-forming algae and invasive seaweeds will increase as oceans acidify, although responses in other groups are less predictable. Inter-specific variance in responses to near-future ocean acidification is prevalent, with even closely related species showing very different responses.115,157 This highlights the importance of species-specific investigations, as extrapolations from similar organisms could be inaccurate. It will be important to consider the limitations in applying organism responses from laboratory experiments to predictive modeling of natural habitats, as laboratory responses will not necessarily translate to the wild. The effect of ocean acidification on nuisance species must be considered in the context of their community, as their impact will largely depend on the relative responses of surrounding organisms.15,158
Invasive species are, by nature, adaptable and this will play an important role in determining their success as the chemistry of the oceans continues to change.159 The role of adaptation has not been explored extensively with regard to ecologically harmful marine species, however, the increasing rate of ocean acidification is narrowing the time window available for marine organisms to adapt,17 thus organisms with resilient genotypes present in current populations have an advantage.97,122 Some invasive species have strains that are particularly resilient to ocean acidification, for example the invasive C. gigas may adapt more readily to ocean acidification than its native competitor S. glomerata in Southern Australia.78
Although this review focuses on ocean acidification, multiple drivers of global change such as warming, eutrophication, overfishing, and oxygen depletion will determine the prevalence of outbreaks of nuisance marine organisms. For example, ocean acidification can enhance the success of invasive marine organisms near their thermal limits at the same time as causing a decline in native marine organisms.9,120,160 Indeed, strong variability in responses to multiple climatic stressors have been identified in marine taxa, and moving forward, these stressors should be considered synergistically wherever possible.161 In carrying out this review we found a few instances where invasive or harmful marine organisms might be negatively affected by ocean acidification, but far more instances in which ocean acidification can be reasonably expected to make matters worse. This highlights the necessity of regional studies of the responses of harmful marine taxa (e.g. C. taxifolia, A. amurensis, invasive ascidians, P. volitans) to rising CO2 levels. This review also reinforces the overall imperative to reduce CO2 emissions, since they may be opening the door to the spread of harmful marine organisms.
Acknowledgment
This review contributes to the EU FP7 project ‘Mediterranean Sea Acidification under a changing climate’ (grant agreement no. 265103) and to the UK Ocean Acidification research programme, co-funded by NERC, Defra and DECC (Grant: NE/H016996/1 “Ocean acidification impacts on sea-surface biogeochemistry and climate”).
Disclosure
There are no conflicts of interest to disclose in this work.
References
Hoegh-Guldberg OR, Cai R, Poloczanska ES, et al. The Ocean. In: Turley C, Omar L, editors. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part B: Regional Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York, NY, USA: Cambridge University Press; 2014:1655–1731. | |
Feely RA, Sabine C, Lee K, et al. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science. 2004;305(5682):362–366. | |
Barry JP, Widdicombe S, Hall-Spencer JM. Effects of ocean acidification on marine biodiversity and ecosystem function. In: Gattuso JP, Lansson L, editors. Ocean Acidification. Oxford: Oxford University Press; 2011:192–209. | |
Harley CDG, Anderson K, Demes K, et al. Effects of climate change on global seaweed communities. J Phycol. 2012;48(5):1064–1078. | |
Wittmann AC, Pörtner HO. Sensitivities of extant animal taxa to ocean acidification. Nature Climate Change. 2013;3(11):995–1001. | |
Garilli V, Rodolfo-Metalpa R, Scuderi D, et al. Physiological advantages of dwarfing in surviving extinctions in high-CO2 oceans. Nature Climate Change. 2015;5(7):678–682. | |
Welch MJ, Watson SA, Welsh JQ, McCormick MI, Munday PL. Effects of elevated CO2 on fish behaviour undiminished by transgenerational acclimation. Nature Climate Change. 2014;4(12):1086–1089. | |
Connell SD, Kroeker KJ, Fabricius KE, Kline DI, Russell BD. The other ocean acidification problem: CO2 as a resource among competitors for ecosystem dominance. Philos Trans R Soc Lond B Biol Sci. 2013; 368(1627):20120442. | |
Brodie J, Williamson C, Smale D, et al. The future of the northeast Atlantic benthic flora in a high CO2 world. Ecol Evol. 2014;4(13):2787–2798. | |
Fabricius KE, De’ath G, Noonan S, Uthicke S. Ecological effects of ocean acidification and habitat complexity on reef-associated macroinvertebrate communities. Proc Biol Sci. 2014;281(1775):20132479. | |
Jackson EL, Davies AJ, Howell KL, Kershaw PJ, Hall-Spencer JM. Future-proofing marine protected area networks for cold water coral reefs. ICES J Mar Sci. 2014;71(9):2621–2629. | |
Milazzo M, Rodolfo-Metalpa R, Chan V, et al. Ocean acidification impairs vermetid reef recruitment. Sci Rep. 2014;4:4189. | |
Kroeker KJ, Kordas R, Crim R, et al. Impacts of ocean acidification on marine organisms: quantifying sensitivities and interaction with warming. Glob Chang Biol. 2013;19(6):1884–1896. | |
Fabricius KE, Langdon C, Uthicke S, et al. Losers and winners in coral reefs acclimatized to elevated carbon dioxide concentrations. Nature Climate Change. 2011;1(3):165–169. | |
Gaylord B, Kroeker K, Sunday J. Ocean acidification through the lens of ecological theory. Ecology. 2015;96(1):3–15. | |
Branch TA, Dejoseph BM, Ray LJ, Wagner CA. Impacts of ocean acidification on marine seafood. Trends Ecol Evol. 2013;28(3):178–186. | |
Gattuso JP, Magnan A, Billé R, et al. Contrasting futures for ocean and society from different anthropogenic CO2 emissions scenarios. Science. 2015;349(6243):aac4722. | |
Cornwall CE, Revill AT, Hurd CL. High prevalence of diffusive uptake of CO2 by macroalgae in a temperate subtidal ecosystem. Photosynth Res. 2015;124(2):181–190. | |
Koch M, Bowes G, Ross C, Zhang XH. Climate change and ocean acidification effects on seagrasses and marine macroalgae. Glob Chang Biol. 2013;19(1):103–132. | |
Vergés A, Steinberg PD, Hay ME, et al. The tropicalization of temperate marine ecosystems: climate-mediated changes in herbivory and community phase shifts. Proc Biol Sci. 2014;281(1789):20140846. | |
Porzio L, Buia MC, Hall-Spencer JM. Effects of ocean acidification on macroalgal communities. Journal of Experimental Marine Biology and Ecology. 2011;400(1–2):278–287. | |
Baggini C, Salomidi M, Voutsinas E, Bray L, Krasakopoulou E, Hall-Spencer JM. Seasonality affects macroalgal community response to increases in pCO2. PLoS One. 2014;9(9):e106520. | |
Grant WM. Molecular Phylogeography and Climate Change Biology of the Invasive Green Marine Macroalgae Caulerpa Taxifolia and Caulerpa Cylindracea in Australia [PhD thesis]. Australia: The University of Adelaide; 2015. | |
Britton-Simmons KH. Direct and indirect effects of the introduced alga Sargassum muticum on benthic, subtidal communities of Washington State, USA. Marine Ecology Progress Series. 2004;277:61–78. | |
Vaz-Pinto F, Olabarria C, Gestoso I, et al. Functional diversity and climate change: Effects on the invasibility of macroalgal assemblages. Biological Invasions. 2013;15(8):1833–1846. | |
Porzio L, Garrard SL, Buia MC. The effect of ocean acidification on early algal colonization stages at natural CO2 vents. Marine Biology. 2013;160(8):2247–2259. | |
Olabarria C, Arenas F, Viejo R, et al. Response of macroalgal assemblages from rockpools to climate change: Effects of persistent increase in temperature and CO2. Oikos. 2013;122(7):1065–1079. | |
Scherner F, Horta PA, Oliveira EC, et al. Coastal urbanization leads to remarkable seaweed species loss and community shifts along the SW Atlantic. Mar Pollut Bull. 2013;76(1–2):106–115. | |
YeXiao-wen NH, Zhang XW, Mao YZ, et al. ‘Green tides’ are overwhelming the coastline of our blue planet: Taking the world’s largest example. Ecological Research. 2011;26(3):477–485. | |
Xu J, Gao K. Future CO2 Induced ocean acidification mediates the physiological performance of a green tide alga. Plant Physiol. 2012;160(4):1762–1769. | |
Fu FX, Tatters AO, Hutchins DA. Global change and the future of harmful algal blooms in the ocean. Mar Ecol Prog Ser. 2012;470:207–233. | |
Sun J, Hutchins D, Feng Y, et al. Effects of changing pCO2 and phosphate availability on domoic acid production and physiology of the marine harmful bloom diatom Pseudo-nitzschia multiseries. Limnol Oceanogr. 2011;56(3):829–840. | |
Lelong A, Hégaret H, Soudant P, Bates SS. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: revisiting previous paradigms. Phycologia. 2012;51(2):168–216. | |
Tatters AO, Fu FX, Hutchins D. High CO2 and silicate limitation synergistically increase the toxicity of Pseudo-nitzschia fraudulenta. PLoS One. 2012;7(2):e32116. | |
Errera RM, Yvon-Lewis S, Kessler JD, Campbell L. Reponses of the dinoflagellate Karenia brevis to climate change: PCO2 and sea surface temperatures. Harmful Algae. 2014;37:110–116. | |
Hardison DR, Sunda WG, Tester P, Shea D, Litaker RW. Increased cellular brevetoxins in the red tide dinoflagellate Karenia brevis under CO2 limitation of growth rate: Evolutionary implications and potential effects on bloom toxicity. Limnol Oceanogr. 2014;59(2):560–577. | |
Hwang DF, Lu YH. Influence of environmental and nutritional factors on growth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum. Toxicon. 2000;38(11):1491–1503. | |
Flores-Moya A, Rouco M, García-Sánchez M, et al. Effects of adaptation, chance, and history on the evolution of the toxic dinoflagellate Alexandrium minutum under selection of increased temperature and acidification. Ecol Evol. 2012;2(6):1251–1259. | |
Wang Y, Smith WO, Wang X, Li S. Subtle biological responses to increased CO2 concentrations by Phaeocystis globosa Scherffel, a harmful algal bloom species. Geophys Res Lett. 2010;37(9):1–5. | |
Chen S, Gao K, Beardall J. Viral attack exacerbates the susceptibility of a bloom-forming alga to ocean acidification. Glob Chang Biol. 2014;21(2):629–636. | |
Hattenrath-Lehmann TK, Smith J, Wallace R, et al. The effects of elevated CO2 on the growth and toxicity of field populations and cultures of the saxitoxin – producing dinoflagellate, Alexandrium fundyense. Limnol Oceanogr. 2014;60(1):198–214. | |
Talmage SC, Gobler CJ. Effects of CO2 and the harmful alga Aureococcus anophagefferens on growth and survival of oyster and scallop larvae. Mar Ecol Prog Ser. 2012;464:121–134. | |
Fleming L, McDonough N, Austen M, et al. Oceans and human health: a rising tide of challenges and opportunities for Europe. Mar Env Res. 2014;99:16–19. | |
Hall-Spencer JM, Rodolfo Metalpa R, Martin S, et al. Volcanic carbon dioxide vents show ecosystem effects of ocean acidification. Nature. 2008;454:96–99. | |
Olischläger M, Wiencke C. Ocean acidification alleviates low-temperature effects on growth and photosynthesis of the red alga Neosiphonia harveyi (Rhodophyta). J Exp Bot. 2013;64(18):5587–5597. | |
Carvalho M, Hayashizaki K, Ogawa, H. Effect of pH on the carbon stable isotope fractionation in photosynthesis by the kelp Undaria pinnatifida. Coastal Marine Science. 2010;34(1):135–139. | |
Van de Waal DB, Eberlein T, John U, Wohlrab S, Rost B. Impact of elevated pCO2 on paralytic shellfish poisoning toxin content and composition in Alexandrium tamarense. Toxicon. 2014;78:58–67. | |
Brutemark A, Engström-öst J, Vehmaa A, Gorokhova E. Growth, toxicity and oxidative stress of a cultured cyanobacterium (Dolichospermum sp.) under different CO2/pH and temperature conditions. Phycol Res. 2015;63:56–63. | |
Brotz L, Cheung WWL, Kleisner K, Pakhomov E, Pauly D. Increasing jellyfish populations: trends in Large Marine Ecosystems. Hydrobiologia. 2012;690(1):3–20. | |
Purcell JE, Uye SI, Lo WT. Anthropogenic causes of jellyfish blooms and their direct consequences for humans: A review. Mar Ecol Prog Ser. 2007;350:153–174. | |
Richardson AJ, Bakun A, Hays GC, Gibbons MJ. The jellyfish joyride: causes, consequences and management responses to a more gelatinous future. Trends Ecol Evol. 2009;24(6):312–322. | |
Roohi A, Kideys A, Sajjadi A, et al. Changes in biodiversity of phytoplankton, zooplankton, fishes and macrobenthos in the Southern Caspian Sea after the invasion of the ctenophore Mnemiopsis Leidyi. Biol Invasions. 2010;12(7):2343–2361. | |
Condon RH, Steinberg D, del Giorgio PA, et al. Jellyfish blooms result in a major microbial respiratory sink of carbon in marine systems. Proc Natl Acad Sci U S A. 2011;108(25):10225–10230. | |
Daskalov G, Grishin A, Rodionov S, Mihneva V. Trophic cascades triggered by overfishing reveal possible mechanisms of ecosystem regime shifts. Proc Natl Acad Sci U S A. 2007;104(25):10518–10523. | |
Olariaga A, Guallart E, Fuentes V, et al. Polyp flats, a new system for experimenting with jellyfish polyps, with insights into the effects of ocean acidification. Limnol Oceanogr Methods. 2014;12(4):212–222. | |
Lacoue-Labarthe T, Nunes PALD, Cinar M, et al. Mediterranean and the Black Seas region. In: Hilmi N, Allemand D, Kavanagh C, Laffoley D, Metian M, Osborn D, Reynaud S, editors. Bridging the gap between ocean acidification impacts and economic valuation: regional impacts of ocean acidification on fisheries and aquaculture. IUCN 2015 DOI: 10.2305/IUCN.CH.2015.03.en. | |
Suggett DJ, Hall-Spencer JM, Rodolfo-Metalpa R, et al. Sea anemones may thrive in a high CO2 world. Glob Chang Biol. 2012;18(10):3015–3025. | |
Winans AK, Purcell JE. Effects of pH on asexual reproduction and statolith formation of the scyphozoan, Aurelia labiata. Hydrobiologia. 2010;645(1):39–52. | |
Klein SG, Pitt K, Rathjen K, Seymour JE. Irukandji jellyfish polyps exhibit tolerance to interacting climate change stressors. Glob Chang Biol. 2014;20(1):29–37. | |
Melzner F, Stange P, Trübenbach K, et al. Food supply and seawater pCO2 impact calcification and internal shell dissolution in the blue mussel Mytilus edulis. PLoS One. 2011;6(9):e24223. | |
Attrill MJ, Wright J, Edwards M. Climate-related increases in jellyfish frequency suggest a more gelatinous future for the North Sea. Limnol Oceanogr. 2007;52:480–485. | |
Richardson AJ, Gibbons MJ. Are jellyfish increasing in response to ocean acidification? Limnol Oceanogr. 2008;53(5):2040–2045. | |
Duarte CM, Fulweiler R, Lovelock C, et al. Reconsidering Ocean Calamities. Bioscience. 2014;65(2):130–139. | |
Gazeau F, Parker L, Comeau S, et al. Impacts of ocean acidification on marine shelled molluscs. Mar Biol. 2013;160:2207–2245. | |
Parker LM, Ross P, O’Connor W, et al. Predicting the response of molluscs to the impact of ocean acidification. Biology. 2013;2(2):651–692. | |
Cigliano M, Gambi MC, Rodolfo-Metalpa R, Patti FP, Hall-Spencer JM. Effects of ocean acidification on invertebrate settlement at volcanic CO2 vents. Mar Biol. 2010;157(11):2489–2502. | |
Rodolfo-Metalpa R, Houlbreque F, Tambutte E, et al. Coral and mollusc resistance to ocean acidification adversely affected by warming. Nat Clim Chang. 2011;1(6):308–312. | |
Basso L, Hendriks IE, Rodríguez-Navarro AB, Gambi C, Duarte CM. Extreme pH Conditions at a Natural CO2 Vent System (Italy) Affect Growth, and Survival of Juvenile Pen Shells. Estuaries Coasts. 2015:1. | |
Streftaris N, Zenetos A. Alien marine species in the Mediterranean – the 100 ‘worst invasives’ and their impact. Mediterr Mar Sci. 2006;7(1):87–118. | |
Thieltges DW. Impact of an invader: Epizootic American slipper limpet Crepidula fornicata reduces survival and growth in European mussels. Mar Ecol Prog Ser. 2005;286:13–19. | |
Ries JB, Cohen AL, McCorkle DC. Marine calcifiers exhibit mixed responses to CO2-induced ocean acidification. Geology. 2009;37(12):1131–1134. | |
Noisette F, Comtet T, Legrand E, et al. Does encapsulation protect embryos from the effects of ocean acidification? The example of Crepidula fornicata. PLoS One. 2014;9(3):e93021. | |
Nell J. The history of oyster farming in Australia. Mar Fish Rev. 2001; 63(3):14–25. | |
Diederich S. Differential recruitment of introduced Pacific oysters and native mussels at the North Sea coast: coexistence possible? J Sea Res. 2005;53(4):269–281. | |
Havenhand JN, Schlegel P. Near-future levels of ocean acidification do not affect sperm motility and fertilization kinetics in the oyster Crassostrea gigas. Biogeosciences Discuss. 2009;6(12):3009–3015. | |
Barros P, Sobral P, Range P, Chícharo L, Matias D. Effects of sea-water acidification on fertilization and larval development of the oyster Crassostrea gigas. J Exp Mar Bio Ecol. 2013;440:200–206. | |
Barton A, Hales B, Waldbusser GG, Langdon C, Feely R. The Pacific oyster, Crassostrea gigas, shows negative correlation to naturally elevated carbon dioxide levels: Implications for near-term ocean acidification effects. Limnol Oceanogr. 2012;57(3):698–710. | |
Wright JM, Parker L, O’Connor W, et al. Populations of pacific oysters Crassostrea gigas respond variably to elevated CO2 and predation by Morula marginalba. Biol Bull. 2014;226(3):269–281. | |
Parker LM, Ross PM, O’Connor WA. Comparing the effect of elevated pCO2 and temperature on the fertilization and early development of two species of oysters. Mar Biol. 2010;157(11):2435–2452. | |
Ordzie CJ, Garofalo GC. Predation, attack success, and attraction to the bay scallop, Argopecten irradians (Lamarck) by the oyster drill, Urosalpinx cinerea (Say). J Exp Mar Bio Ecol. 1980;47(1):95–100. | |
Buhle ER, Ruesink JL. Impacts of invasive oyster drills on olympia oyster (Ostrea lurida Carpenter 1864) Recovery in Willapa Bay, Washington, United States. J Shellfish Res. 2009;28(1):87–96. | |
Sanford E, Gaylord B, Hettinger A, et al. Ocean acidification increases the vulnerability of native oysters to predation by invasive snails. Proc Biol Sci. 2014;281(1778):20132681. | |
Kroeker K, Sanford E, Jellison B, Gaylord B. Predicting the effects of ocean acidification on predator-prey interactions: a conceptual framework based on coastal molluscs. Biol Bull. 2014;226(3):211–222. | |
Kimbro DL, Grosholz E, Baukus A, et al. Invasive species cause large-scale loss of native California oyster habitat by disrupting trophic cascades. Oecologia. 2009;160(3):563–575. | |
iddri.org [homepage on the Internet]. Weatherdon L, Rogers A, Sumaila R, Magnan A, Cheung W. The Oceans 2015 Initiative, Part II: An updated understanding of the observed and projected impacts of ocean warming and acidification on marine and coastal socioeconomic activities/sectors. Available from: http://www.iddri.org/Publications/The-Oceans-2015-Initiative,Part-II-An-updated-understanding-of-the-observed-and-projected-impacts-of-ocean-warming-and-acidific. Accessed September 7, 2015. | |
Noisette F, Richard J, Le Fur I, et al. Metabolic responses to temperature stress under elevated pCO2 in Crepidula fornicata. Journal of Molluscan Studies. 2014;81(2). | |
Timmins-Schiffman E, O’Donnell MJ, Friedman CS, Roberts SB. Elevated pCO2 causes developmental delay in early larval Pacific oysters, Crassostrea gigas. Mar Biol. 2013;160(8):1973–1982. | |
Kurihara H, Kato S, Ishimatsu A. Effects of increased seawater pCO2 on early development of the oyster Crassostrea gigas. Aquat Biol. 2007;1(1):91–98. | |
Garrard SL, Gambi M, Scipione M, et al. Indirect effects may buffer negative responses of seagrass invertebrate communities to ocean acidification. J Exp Mar Bio Ecol. 2014;461:31–38. | |
Whiteley NM. Physiological and ecological responses of crustaceans to ocean acidification. Mar Ecol Prog Ser. 2011;430:257–271. | |
Foster BA, Willan RC. Foreign barnacles transported to New Zealand on an oil platform. NZ J Mar Freshw Res. 1979;13:143–149. | |
Zvyagintsev AY. Fouling of ocean-going shipping and its role in the spread of exotic species in the seas of the Far East. Sessile Org. 2000;17:31–43. | |
Kotta J, Kotta I, Simm M, et al. Ecological consequences of biological invasions: Three invertebrate case studies in the north-eastern Baltic Sea. Helgol Mar Res. 2006;60(2):106–112. | |
Olenin S, Leppäkoski E. Non-native animals in the Baltic Sea: Alteration of benthic habitats in coastal inlets and lagoons. Hydrobiologia. 1999;393:233–243. | |
Laudien J, Wahl M. Indirect effects of epibiosis on host mortality: seastar predation on differently fouled mussels. Mar Ecol. 1999;20(1):35–47. | |
Pansch C, Nasrolahi A, Appelhans YS, Wahl M. Impacts of ocean warming and acidification on the larval development of the barnacle Amphibalanus improvisus. J Exp Mar Bio Ecol. 2012;420:48–55. | |
Pansch C, Schlegel P, Havenhand J. Larval development of the barnacle Amphibalanus improvisus responds variably but robustly to near-future ocean acidification. ICES J Mar Sci. 2013;70(4):805–811. | |
Pansch C, Nasrolahi A, Appelhans YS, Wahl M. Tolerance of juvenile barnacles (Amphibalanus improvisus) to warming and elevated pCO2. Mar Biol. 2013;160(8):2023–2035. | |
Findlay HS, Kendall M, Spicer JI, Widdicombe S. Future high CO2 in the intertidal may compromise adult barnacle Semibalanus balanoides survival and embryonic development rate. Mar Ecol Prog Ser. 2009;389:193–202. | |
Grosholz ED, Ruiz GM. Predicting the impact of introduced marine species: lessons from the multiple invasions of the European Green Crab Carinus maenas. Biol Conserv. 1996;78:59–66. | |
Thresher R, Proctor C, Ruiz G, et al. Invasion dynamics of the European shore crab, Carcinus maenas, in Australia. Mar Biol. 2003; 142(5):867–876. | |
Walton WC, MacKinnon C, Rodriguez LF, Proctor C, Ruiz GM. Effect of an invasive crab upon a marine fishery: green crab, Carcinus maenas, predation upon a venerid clam, Katelysia scalarina, in Tasmania (Australia). J Exp Mar Bio Ecol. 2002;272(2):171–189. | |
Murray LG, Seed R, Jones T. Predicting the impacts of Carinus maenas predation on cultivated Mytilus edulis beds. J Shellfish Res. 2007;26(4):1089–1098. | |
Fehsenfeld S, Kiko R, Appelhans Y, et al. Effects of elevated seawater pCO2 on gene expression patterns in the gills of the green crab, Carcinus maenas. BMC Genomics. 2011;12:488. | |
Appelhans YS, Thomsen J, Pansch C, Melzner F, Wahl M. Sour times: Seawater acidification effects on growth, feeding behaviour and acid-base status of Asterias rubens and Carcinus maenas. Mar Ecol Prog Ser. 2012;459:85–97. | |
Landes A, Zimmer M. Acidification and warming affect both a calcifying predator and prey, but not their interaction. Mar Ecol Prog Ser. 2012;450:1–10. | |
De la Haye KL, Spicer JI, Widdicombe S, Briffa M. Reduced pH sea water disrupts chemo-responsive behaviour in an intertidal crustacean. J Exp Mar Bio Ecol. 2012;412:134–140. | |
Bibby R, Cleall-Harding P, Rundle S, Widdicombe S, Spicer J. Ocean acidification disrupts induced defences in the intertidal gastropod Littorina littorea. Biol Lett. 2007;3(6):699–701. | |
Melatunan S, Calosi P, Rundle SD, Widdicombe S, Moody J. Effects of ocean acidification and elevated temperature on shell plasticity and its energetic basis in an intertidal gastropod. Mar Ecol Prog Ser. 2013;472:155–168. | |
Britayev T, Rzhavsky V, Pavlova LV, Dvoretskij G. Studies on impact of the alien Red King Crab (Paralithodes camtschaticus) on the shallow water benthic communities of the Barents Sea. J Appl Ichthyol. 2010;26(Suppl 2):66–73. | |
Falk-Petersen J, Renaud P, Anisimova N. Establishment and ecosystem effects of the alien invasive red king crab (Paralithodes camtschaticus) in the Barents Sea – A review. ICES J Mar Sci. 2011;68(3):479–488. | |
Oug E, Cochrane SKJ, Sundet JH, Norling K, Nilsson HC. Effects of the invasive red king crab (Paralithodes camtschaticus) on soft-bottom fauna in Varangerfjorden, northern Norway. Mar Biodivers. 2011;41(3):467–479. | |
Long WC, Swiney KM, Foy RJ. Effects of ocean acidification on the embryos and larvae of red king crab, Paralithodes camtschaticus. Mar Pollut Bull. 2013;69(1–2):38–47. | |
Long WC, Swiney KM, Harris C, Page HN, Foy RJ. Effects of ocean acidification on juvenile Red King Crab (Paralithodes camtschaticus) and Tanner Crab (Chionoecetes bairdi) growth, condition, calcification, and survival. PLoS One. 2013;8(4):e60959. | |
Byrne M, Lamare M, Winter D, Dworjanyn SA, Uthicke S. The stunting effect of a high CO2 ocean on calcification and development in sea urchin larvae, a synthesis from the tropics to the poles. Philos Trans R Soc Lond B Biol Sci. 2013;368(1627):20120439. | |
Johnson VR, Russell BD, Fabricius KE, Brownlee C, Hall-Spencer JM. Temperate and tropical brown macroalgae thrive, despite decalcification, along natural CO2 gradients. Glob Chang Biol. 2012;18(9):2792–2803. | |
Calosi P, Rastrick SPS, Graziano M, et al. Distribution of sea urchins living near shallow CO2 vents is dependent on species acid-base and ion-regulatory abilities. Mar Pollut Bull. 2013;73(2):470–484. | |
Strain EM, Johnson CR. Competition between an invasive urchin and commercially fished abalone: effect on body condition, reproduction and survivorship. Mar Ecol Prog Ser. 2009;377:169–182. | |
Byrne M, Andrew N. Centrostephanus rodgersii. In: Lawrence JM, editor. Sea Urchins: Biology and Ecology. San Diego: Academic Press; 2013:243–254. | |
Pecorino D, Barker MF, Dworjanyn S, Byrne M, Lamare MD. Impacts of near future sea surface pH and temperature conditions on fertilisation and embryonic development in Centrostephanus rodgersii from northern New Zealand and northern New South Wales, Australia. Mar Biol. 2014;161:101–110. | |
Byrne M, Foo S, Soars N, et al. Ocean warming will mitigate the effects of acidification on calcifying sea urchin larvae (Heliocidaris tuberculata) from the Australian global warming hot spot. J Exp Mar Bio Ecol. 2013;448:250–257. | |
Foo SA, Dworjanyn SA, Poore AG, Byrne M. Adaptive capacity of the habitat modifying sea urchin Centrostephanus rodgersii to ocean warming and ocean acidification: Performance of early embryos. PLoS One. 2012;7(8):e42497. | |
Baird AH, Pratchett MS, Hoey AS, Herdiana Y, Campbell SJ. Acanthaster planci is a major cause of coral mortality in Indonesia. Coral Reefs. 2013;32(3):803–812. | |
Katz SM, Pollock FJ, Bourne DG, Willis BL. Crown-of-thorns starfish predation and physical injuries promote brown band disease on corals. Coral Reefs. 2014;33(3):705–716. | |
Uthicke S, Pecorino D, Albright R, et al. Impacts of ocean acidification on early life-history stages and settlement of the coral-eating sea star Acanthaster planci. PLoS One. 2013;8(12):e82938. | |
Kamya PZ, Dworjanyn SA, Hardy N, et al. Larvae of the coral eating crown-of-thorns starfish, Acanthaster planci in a warmer-high CO2 ocean. Glob Chang Biol. 2014;20(11):3365–3376. | |
Fabricius KE, Okaji K, De’ath G. Three lines of evidence to link outbreaks of the crown-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral Reefs. 2010;29(3):593–605. | |
Uthicke S, Logan M, Liddy M, et al. Climate change as an unexpected co-factor promoting coral eating seastar (Acanthaster planci) outbreaks. Sci Rep. 2015;5:8402. | |
Calderwood J, O’Connor N, Roberts D. The effects of transportation stress and barnacle fouling on predation rates of starfish (Asterias rubens) on mussels (Mytilus edulis). Aquaculture. 2015;444:108–113. | |
Appelhans Y, Thomsen J, Opitz S, et al. Juvenile sea stars exposed to acidification decrease feeding and growth with no acclimation potential. Mar Ecol Prog Ser. 2014;509:227–239. | |
Hernroth B, Baden S, Thorndyke M, Dupont S. Immune suppression of the echinoderm Asterias rubens (L.) following long-term ocean acidification. Aquat Toxicol. 2011;103(3–4):222–224. | |
Lowe S, Browne M, Boudjelas S, De Poorter M. 100 of the world’s worst invasive alien species. Aliens. 2000;12:12. | |
Byrne M, Gonzalez-Bernat M, Doo S, Foo S, Soars N, Lamare M. Effects of ocean warming and acidification on embryos and non-calcifying larvae of the invasive sea star Patiriella regularis. Mar Ecol Prog Ser. 2013;473:235–246. | |
Byrne M, Soars N, Ho M, et al. Fertilization in a suite of coastal marine invertebrates from SE Australia is robust to near-future ocean warming and acidification. Mar Biol. 2010;157:2061–2069. | |
Doo SS, Dworjanyn S, Foo S, Soars N, Byrne M. Impacts of ocean acidification on development of the meroplanktonic larval stage of the sea urchin Centrostephanus rodgersii. ICES J Mar Sci. 2011;69:460–464. | |
Lengyel NL, Collie JS, Valentine PC. The invasive colonial ascidian Didemnum vexillum on Georges Bank – Ecological effects and genetic identification. Aquat Invasions. 2009;4(1):143–152. | |
Bock DG, Zhan A, Lejeusne C, MacIsaac HJ, Cristescu ME. Looking at both sides of the invasion: patterns of colonization in the violet tunicate Botrylloides violaceus. Mol Ecol. 2011;20(3):503–516. | |
Bullard SG, Davis CV, Shumway SE. Seasonal patterns of ascidian settlement at an aquaculture facility in the Damariscotta River, Maine. J Shellfish Res. 2013;32(2):255–264. | |
Dupont S, Thorndyke MC. Impact of CO2-driven ocean acidification on invertebrates early life-history – What we know, what we need to know and what we can do. Biogeosciences Discuss. 2009;6(2):3109–3131. | |
Donnarumma L, Lombardi C, Cocito S, Gambi MC. Settlement pattern of Posidonia oceanica epibionts along a gradient of ocean acidification: an approach with mimics. Mediterr Mar Sci. 2014;15(3):498–509. | |
Peck LS, Clark M, Power D, et al. Acidification effects on biofouling communities: winners and losers. Glob Chang Biol. 2015;21(5):1907–1913. | |
Munday PL, Donelson JM, Dixson DL, Endo GG. Effects of ocean acidification on the early life history of a tropical marine fish. Proc Biol Sci. 2009;276(1671):3275–3283. | |
Nilsson GE, Dixon D, Domenici P, et al. Near-future carbon dioxide levels alter fish behaviour by interfering with neurotransmitter function. Nat Clim Chang. 2012;2(3):201–204. | |
Chung WS, Marshall JN, Watson S, Munday PL, Nilsson G. Ocean acidification slows retinal function in damselfish through interference with GABAA receptors. J Exp Biol. 2014;217:323–326. | |
Dixson DL, Munday PL, Jones GP. Ocean acidification disrupts the innate ability of fish to detect predator olfactory cues. Ecol Lett. 2010;13(1):68–75. | |
Munday PL, Cheal AJ, Dixson DL, Rummer JL, Fabricius KE. Behavioural impairment in reef fishes caused by ocean acidification at CO2 seeps. Nat Clim Chang. 2014;4(6):487–492. | |
Simpson SD, Munday PL, Wittenrich ML, et al. Ocean acidification erodes crucial auditory behaviour in a marine fish. Biol Lett. 2011;7(6):917–920. | |
Ferrari MC, Mccormick M, Munday P, et al. Effects of ocean acidification on visual risk assessment in coral reef fishes. Funct Ecol. 2012;26(3):553–558. | |
Whitfield PE, Gardner T, Vives S, et al. Biological invasion of the Indo-Pacific lionfish Pterois volitans along the Atlantic coast of North America. Mar Ecol Prog Ser. 2002;235:289–297. | |
Arias-González JE, González-Gándara C, Luis Cabrera J, Christensen V. Predicted impact of the invasive lionfish Pterois volitans on the food web of a Caribbean coral reef. Environ Res. 2011;111(7):917–925. | |
Green SJ, Akins JL, Maljkovic A, Côté IM. Invasive lionfish drive Atlantic coral reef fish declines. PLoS One. 2012;7(3):e32596. | |
Lesser MP, Slattery M. Phase shift to algal dominated communities at mesophotic depths associated with lionfish (Pterois volitans) invasion on a Bahamian coral reef. Biol Invasions. 2011;13(8):1955–1868. | |
Cripps IL, Munday PL, McCormick MI. Ocean acidification affects prey detection by a predatory reef fish. PLoS One. 2011;6(7):e22736. | |
Albins M, Hixon M. Invasive Indo-Pacific lionfish Pterois volitans reduce recruitment of Atlantic coral-reef fishes. Mar Ecol Prog Ser. 2008;367:233–238. | |
Celis-Plá PS, Hall-Spencer JM, Horta P, et al. Macroalgal responses to ocean acidification depend on nutrient and light levels. Front Mar Sci. 2015;2:26. | |
Lenz M, daGama BAP, Gerner NV, et al. Non-native marine invertebrates are more tolerant towards environmental stress than taxonomically related native species: Results from a globally replicated study. Environ Res. 2011;111(7):943–952. | |
Freider CA. Present-day nearshore pH differentially depresses fertilization in congeneric sea urchins. Biol Bull. 2014;226(1):1–7. | |
Kroeker KJ, Micheli F, Gambi MC. Ocean acidification causes ecosystem shifts via altered competitive interactions. Nat Clim Change. 2012;3:156–159. | |
Sunday JM, Calosi P, Dupont S, et al. Evolution in an acidifying ocean. Trends Ecol Evol. 2014;29(2):117–125. | |
Poloczanska ES, Brown CJ, Sydeman WJ, et al. Global imprint of climate change on marine life. Nat Clim Chang. 2013;3(10):919–925. | |
iddri.org [homepage on the Internet]. Howes EL, Joos F, Eakin M, Gattuso J. The Oceans 2015 Initiative, Part I: An updated synthesis of the observed and projected impacts of climate change on physical and biological processes in the oceans. Available from: http://www.iddri.org/Publications/The-Oceans-2015-Initiative,Part-I-An-updated-synthesis-of-the-observed-and-projected-impacts-of-climate-change-on-physical-and. Accessed September 7, 2015. |
 © 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2015 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.