Back to Journals » Cancer Management and Research » Volume 12
The Hsa_circ_0091579/miR-940/TACR1 Axis Regulates the Development of Hepatocellular Carcinoma
Authors Jiang P, Han W, Fu Y, Chen Q
Received 21 April 2020
Accepted for publication 2 August 2020
Published 28 September 2020 Volume 2020:12 Pages 9087—9096
DOI https://doi.org/10.2147/CMAR.S259243
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ahmet Emre Eşkazan
This paper has been retracted.
Peiqiang Jiang, Wei Han, Yu Fu, Qingmin Chen
Department of Hepatobiliary and Pancreatic Surgery, The First Hospital of Jilin University, Changchun, Jilin, People’s Republic of China
Correspondence: Qingmin Chen
Department of Hepatobiliary and Pancreatic Surgery, The First Hospital of Jilin University, No. 71 Xinmin Street, Changchun, Jilin 130021, People’s Republic of China
Tel +86-431-81875133
Email [email protected]
Purpose: Circular RNAs (circRNAs) play important roles in hepatocellular carcinoma (HCC) development. The circRNA hsa_circ_0091579 (circ_0091579) is dysregulated in HCC, while the mechanism of circ_0091579 in HCC development is largely unknown.
Patients and Methods: Thirty paired cancer and adjacent normal tissues were harvested from HCC patients. SNU-387 and Huh7 cells were cultured in this study. circ_0091579, microRNA-940 (miR-940) and tachykinin-1 receptor (TACR1) abundances were measured via quantitative reverse transcription-polymerase chain reaction or Western blot. Cell viability, migration, invasion, colony ability, cell cycle distribution and apoptosis were assessed via 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide, transwell assay, colony formation assay and flow cytometry. The interaction among circ_0091579, miR-940 and TACR1 was tested via dual-luciferase reporter analysis. The anti-HCC role of circ_0091579 knockdown in vivo was investigated using xenograft model.
Results: circ_0091579 expression was enhanced in HCC tissue samples and cells. circ_0091579 silence inhibited cell viability, migration, invasion and colony formation, induced cell cycle arrest at G0/G1 phase, and promoted apoptosis in HCC cells. miR-940 was targeted via circ_0091579 and miR-940 knockdown reversed the suppressive effect of circ_0091579 silence on HCC development. miR-940 targeted TACR1 to repress HCC development. circ_0091579 could regulate TACR1 expression by mediating miR-940. Down-regulation of circ_0091579 decreased xenograft tumor growth.
Conclusion: Knockdown of circ_0091579 repressed HCC development by mediating miR-940/TACR1 axis, indicating a new pathogenesis of HCC.
Keywords: hepatocellular carcinoma, hsa_circ_0091579, miR-940, TACR1
Introduction
Hepatocellular carcinoma (HCC) accounts for up to 90% of liver malignancy with high incidence and mortality in the world.1 With significant insight into the pathogenesis of HCC, the diagnosis and treatment of HCC have gained great advance.2,3 Nevertheless, the outcome and effective therapy strategies are poor in HCC at advanced stage. Therefore, it is urgent to explore new target for HCC treatment.
Circular RNAs (circRNAs) are a type of highly expressed RNAs formed by a closed-loop structure without the 5ʹ caps and 3ʹ tail, which play important roles in human cancers.4 CircRNAs have multi-functions in the pathogenesis, development and treatment of HCC.5 Moreover, circRNAs usually take part in the development and treatment of HCC by mediating the competing endogenous RNA (ceRNA) network (circRNA-microRNA (miRNA)-mRNA).6 Previous studies have provided insight into multiple circRNAs in HCC. For example, hsa_circ_0001955, hsa_circ_0056836 and hsa_circ_0000092 have been reported to facilitate HCC development;7–9 while some circRNAs play the tumor-suppressive role in HCC, such as hsa_circ_5692, hsa_circ_0070269 and hsa_circ_0003418.10−12 A previous study reports that hsa_circ_0091579 (circ_0091579), a dysregulated circRNA derived from glypican 3 (GPC3) gene, is implicated in the detection, prognosis and treatment of HCC.13 However, the mechanism that circ_0091579 participates in the regulation of HCC development remains largely unknown.
miRNAs are single-stranded RNAs (~20 nucleotides) which exhibit important clinical values in liver disorders, including HCC.14,15 The former evidences indicate that miR-940 could play an anti-cancer role in HCC via inhibiting cell growth, migration and invasion.16,17 The tachykinin-1 receptor (TACR1) has been suggested to be an oncogene in many malignancies, including HCC.18–20 The bioinformatics analysis using CircInteractome21 and TargatScan22 predicts that miR-940 could bind with circ_0091579 and TACR1. However, it is not clear whether miR-940 and TACR1 are associated with circ_0091579-mediated HCC development.
In this research, we detected the expression of circ_0091579 and investigated the function of circ_0091579 on HCC development in vitro and in vivo. Additionally, we explored the ceRNA crosstalk of circ_0091579/miR-940/TACR1 in HCC cells.
Patients and Methods
Patients and Tissues
HCC patients (n=30) were recruited from the First Hospital of Jilin University. The HCC tissues and adjacent normal tissues were harvested and maintained at −80°C. Patients did not receive the other therapy before the tissue collection. All patients signed the written informed consents, and they provided the approval that the tissues could be stored and used for research. This work was in accordance with the Declaration of Helsinki. This research was approved via the ethics committee of the First Hospital of Jilin University.
Cell Culture and Transfection
HCC cell lines SNU-387 and Huh7 cells, and liver epithelial cell line THLE-2 cells were provided via Procell (Wuhan, China) and grown in RPMI-1640 medium (Procell) plus 10% fetal bovine serum (Zhejiang Tianhang Biotechnology, Huzhou, China) and 1% penicillin/streptomycin (Thermo Fisher, Waltham, MA, USA) in 5% CO2 at 37°C.
TACR1 overexpression vector (pc-TACR1) was generated by cloning TACR1 sequence into pcDNA3.1 vector in our laboratory, and the pcDNA3.1 vector (Thermo Fisher) acted as negative control (pc-NC). siRNA for circ_0091579 (si-circ_0091579-1, 5ʹ-GCACAUUAACCAGAGGCCUUU-3ʹ; si-circ_0091579-2, 5ʹ-CAUUAACCAGAGGCCUUUGAA-3ʹ), negative control of siRNA (si-NC, 5ʹ-AAGACAUUGUGUGUCCGCCTT-3ʹ), miR-940 mimic (5ʹ-AAGGCAGGGCCCCGCUCCCC-3ʹ), negative control of mimic (miRNA NC, 5ʹ-ACGUGACACGUUCGGAGAATT-3ʹ), miR-940 inhibitor (5ʹ-GGGGAGCGGGGCCCUGCCUU-3ʹ), and negative control of inhibitor (inhibitor NC, 5ʹ-CAGUACUUUUGUGUAGUACAA-3ʹ) were generated via Ribobio (Guangzhou, China). The vectors or these oligonucleotides (30 nM) were transfected into SNU-387 and Huh7 cells via Lipofectamine 2000 (Thermo Fisher) for 24 h.
Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
Tissues or cells were incubated in Trizol reagent (Thermo Fisher) and then total RNA was extracted using the acid guanidinium thiocyanate–phenol–chloroform extraction method.23 The reverse transcription was conducted using the specific reverse transcription kit (Thermo Fisher), and the generated cDNA was mixed with SYBR (Solarbio, Beijing, China) and specific primers (Genscript, Nanjing, China), followed via qRT-PCR. The primers were shown as: circ_0091579 (sense, 5ʹ-TGAGCCAGTGGTCAGTCAAA-3ʹ; antisense, 5ʹ-GTGGAGTCAGGCTTGGGTAG-3ʹ), GPC3 (sense, 5ʹ-CCATGCCAAGAACTACACCA-3ʹ; antisense, 5ʹ-GCCCTTCATTTTCAGCTCAT-3ʹ), miR-940 (sense, 5ʹ-GTATAAAGGGCCCCCGCT-3ʹ; antisense, 5ʹ-AGGGTCCGAGGTATTCGCACT-3ʹ), U6 (sense, 5ʹ-CTCGCTTCGGCAGCACA-3ʹ; antisense, AACGCTTCACGAATTTGCGT), and GAPDH (sense, 5ʹ-TGAATGGGCAGCCGTTAGG-3ʹ; antisense, 5ʹ-TGGACTCCACGACGTACTCA-3ʹ). U6 or GAPDH acted as reference control. Relative expression of circ_0091579, GPC3 or miR-940 was calculated via the 2−ΔΔCt method.24
RNase R Treatment and Structure of circ_0091579
RNase R could digest the linear RNAs but not the circRNAs. To test the stability of circRNAs, the isolated RNA was incubated with 3 U/μg RNase R (Geneseed, Guangzhou, China) for 30 min. Then, the levels of circ_0091579 and GPC3 were detected via qRT-PCR.
Furthermore, the structure of circ_0091579 was explored via the cancer-specific circRNA database (http://gb.whu.edu.cn/CSCD).25
Cell Viability
Cell viability was examined via 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H -tetrazolium bromide (MTT) analysis. 1 × 104 SNU-387 and Huh7 cells were added into 96-well plates and incubated for 48 h. Next, the MTT solution (Solarbio) was placed into each well with a final concentration of 0.5 mg/mL, and cells were cultured for 4 h. Then, the medium was removed and 100 μL of dimethyl sulfoxide (DMSO; Beyotime, Shanghai, China) was added. The absorbance at 570 nm was determined with a microplate reader (Molecular Devices, Sunnyvale, CA, USA). Cell viability was normalized to the control group.
Transwell Analysis
Transwell analysis was carried out to analyze the abilities of cell migration and invasion. For invasion assay, the transwell chamber (BD, Franklin Lakes, NJ, USA) was coated with Matrigel (BD). With regard to migration analysis, the chamber was not coated with Matrigel. 1 × 105 SNU-387 and Huh7 cells in non-serum medium were placed in the top chambers. The lower chambers were added with 600 μL of medium plus 10% serum. The cells were cultured for 20 h, and then cells passed the membranes were stained with 0.5% crystal violet (Beyotime), and counted under a microscope (Nikon, Tokyo, Japan) with 3 random fields.
Colony Formation Assay
For colony formation assay, 500 SNU-387 and Huh7 cells were added into the 6-well plates. After culture for 10 days, the cells were fixed with methanol (Aladdin, Shanghai, China) and stained with 0.5% crystal violet. The colonies were observed and counted.
Flow Cytometry
Cell cycle distribution and apoptosis were measured via flow cytometry. For cell cycle detection, 2 × 105 SNU-387 and Huh7 cells were maintained in 6-well plates for 48 h, and then fixed with 75% ethanol (Aladdin), followed by incubating with 50 μg/mL propidium iodide (PI; Solarbio) and RNase (Thermo Fisher). The cell cycle distribution was tested using a flow cytometer (Agilent, Hangzhou, China).
For cell apoptosis assay, 2 × 105 SNU-387 and Huh7 cells were placed into 6-well plates and incubated for 48 h. Then, the cells were harvested and incubated with Annexin V-FITC binding buffer (Sigma, St. Louis, MO, USA), followed via dying with Annexin V-FITC (Sigma) and propidium iodide (PI). Next, cells were detected with a flow cytometer. The apoptotic rate was presented as the percentage of cells with Annexin V-FITC+ and PI±.
Western Blot
Protein samples were isolated using RIPA buffer (Solarbio) and quantified using a BCA kit (Abcam, Cambridge, MA, USA). Twenty μg protein samples were separated via sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes (Solarbio). The transferred membranes were blocked in 5% fat-free milk, and then interacted with primary antibody anti-TACR1 (ab131091, 1:1000 dilution) or anti-GAPDH (ab9485, 1:5000 dilution), which were provided via Abcam. Then, the membranes were interacted with horseradish peroxidase-conjugated IgG (ab205718, 1:20000 dilution, Abcam). GAPDH served as a loading reference. Next, the membranes were incubated with ECL reagent (Beyotime). The protein blots were tested via Image Pro Plus software (Media Cybernetics, Rockville, MD, USA).
Dual-Luciferase Reporter Analysis
The complementary sequence of circ_0091579 and miR-940 was searched by CircInteractome,21 and the sequence of miR-940 and TACR1 was analyzed by TargatScan.22 The wild-type luciferase reporter plasmids (WT-circ_0091579 and WT-TACR1-3ʹUTR) were constructed by cloning the wild-type sequence of circ_0091579 or TACR1 3ʹUTR into pGL3-control vectors (YouBio, Changsha, China). The mutant-type luciferase reporter plasmids (MUT-circ_0091579 and MUT-TACR1-3ʹUTR) were generated via mutating the corresponding binding sites of miR-940. SNU-387 and Huh7 cells were co-transfected with these constructed vectors, Renilla luciferase vector, and miR-940 mimic or miRNA NC for 24 h. Next, the luciferase activity was examined via a dual-luciferase analysis kit (Genomeditech, Shanghai, China).
Xenograft Model
The lentiviral vector carrying shRNA for circ_0091579 (sh-circ_0091579) and its negative control (sh-NC) were produced via GenePharma (Shanghai, China), and transfected into Huh7 cells. The stably transfected cells were selected. Twelve BALB/c nude mice (male, 5-week-old) were purchased from Shanghai SLAC Laboratory Animal Co., Ltd. (Shanghai, China) and arbitrarily grouped into sh-circ_0091579 or sh-NC group (n=6/group) after the corresponding subcutaneous inoculation of the transfected Huh7 cells (4 × 106 cells/mouse). The tumor size was examined once a week and calculated with the formula: 0.5 × length × width.2 Twenty-eight days after inoculation, mice were killed, and all xenograft tumors were collected for weight and detection of circ_0091579, miR-940 and TACR1 expression. The animal experiments were performed in line with guidelines of the National Institutes of Health guide for the Care and Use of Laboratory animals (NIH Publications No. 8023, revised 1978), and had procured the permission of the Animal Ethical Committee of the First Hospital of Jilin University.
Statistical Analysis
The experiments were conducted 3 times. The data were shown as mean ± SD. The difference was tested via Student’s t-test or ANOVA with Dunnett’s test using SPSS 19.0 (SPSS, Chicago, IL, USA). It was statistically significant when P<0.05.
Results
circ_0091579 Level is Increased in HCC
We first measured the abnormally expressed circ_0091579 in HCC tissues and cells. First, we collected 30 paired HCC and adjacent normal tissues, and detected circ_0091579 expression. As shown in Figure 1A, a higher level of circ_0091579 was exhibited in HCC tissues than that in normal group. Furthermore, circ_0091579 level was also detected in HCC cell lines. Results showed that circ_0091579 level was evidently up-regulated in SNU-387 and Huh7 cells compared with THLE-2 cells (Figure 1B). In addition, after the treatment of RNase R, circ_0091579 was more resistant to RNase R than the linear form (GPC3) (Figure 1C and D). Besides, the cancer-specific circRNA database analyzed and described that circ_0091579 was located in the GPC3 gene and formed via head-to-tail splicing of GPC3 exons 5–9 (Figure 1E). These results indicated that the aberrant circ_0091579 might be associated with HCC development.
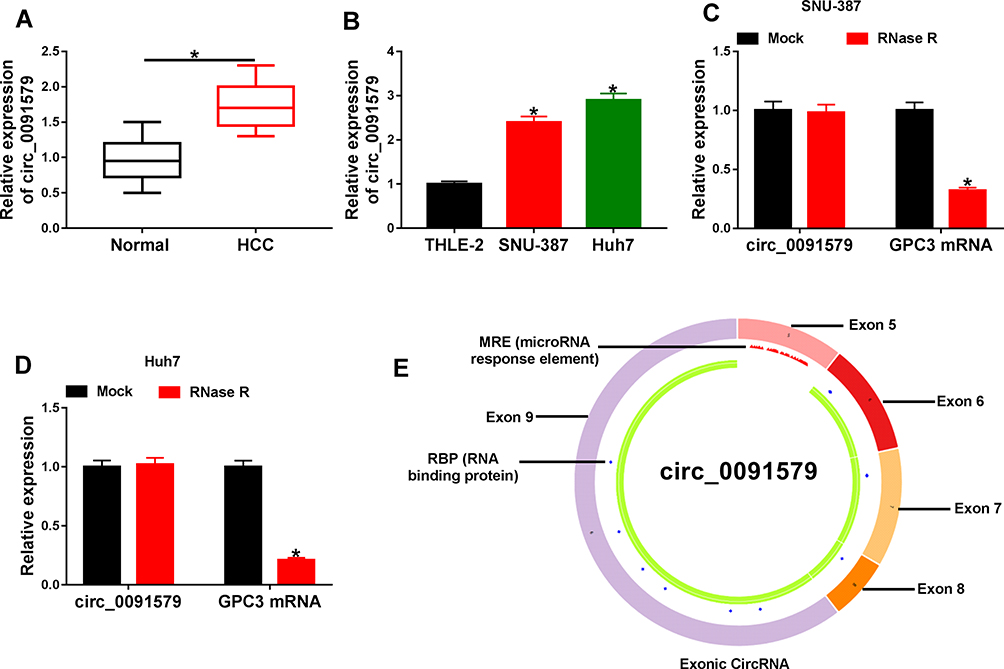 |
Figure 1 The expression of circ_0091579 in HCC. (A) circ_0091579 level was detected in HCC and adjacent normal tissues (n=30) via qRT-PCR. (B) circ_0091579 expression was measured in HCC cell lines (SNU-387 and Huh7) and control cells (THLE-2) by qRT-PCR. (C and D) circ_0091579 and GPC3 mRNA levels were examined after treatment of RNase R in SNU-387 and Huh7 cells. (E) The structure of circ_0091579 was analyzed via cancer-specific circRNA database. *P<0.05. |
Knockdown of circ_0091579 Inhibits HCC Development in vitro
To study the role of circ_0091579 in HCC development in vitro, circ_0091579 abundance was knocked down in SNU-387 and Huh7 cells via transfection of si-circ_0091579-1 or si-circ_0091579-2 (Figure 2A). Meanwhile, GPC3 expression was not changed (Figure 2B). The si-circ_0091579 (si-circ_0091579-1) with the relative higher inhibitive efficacy on circ_0091579 expression, and it was used for further experiments. The MTT assay showed that circ_0091579 knockdown evidently suppressed the viability of SNU-387 and Huh7 cells (Figure 2C). Moreover, circ_0091579 silence markedly reduced the abilities of migration and invasion in the two cell lines (Figure 2D). Additionally, the data of flow cytometry described that interference of circ_0091579 evidently increased the cells at G0/G1 phase and decreased the cells at S phase (Figure 2E). Furthermore, the down-regulation of circ_0091579 significantly repressed the colony formation ability (Figure 2F). Besides, inhibition of circ_0091579 obviously induced SNU-387 and Huh7 cell apoptosis (Figure 2G). These data indicated that circ_0091579 knockdown suppressed HCC development in SNU-387 and Huh7 cells.
 |
Figure 2 The influence of circ_0091579 on HCC development in vitro. (A and B) The levels of circ_0091579 and GPC3 were measured in SNU-387 and Huh7 cells transfected with si-circ_0091579-1, si-circ_0091579-2 or si-NC. Cell viability (C), migration and invasion (D), cycle distribution (E), colony formation (F) and apoptosis (G) were detected in SNU-387 and Huh7 cells transfected with si-circ_0091579-1 (si-circ_0091579) or si-NC. *P<0.05. |
miR-940 Knockdown Reverses the Function of circ_0091579 Silence in HCC Development
By detecting the level of circ_0091579 in nuclear and cytoplasmic fractions, we found that circ_0091579 was mainly expressed in cytoplasm (Figure 3A), suggesting circ_0091579 could serve as a ceRNA. The targets of circ_0091579 were explored using CircInteractome, and the binding sequences of circ_0091579 and miR-940 are displayed in Figure 3B. To validate the interaction between circ_0091579 and miR-940, we constructed the WT-circ_0091579 and MUT-circ_0091579 and transfected them into SNU-387 and Huh7 cells. The data of dual-luciferase reporter analysis displayed that miR-940 overexpression decreased more than 70% luciferase activity of WT-circ_0091579 in the two cell lines, while it did not alter the luciferase activity of MUT-circ_0091579 (Figure 3C). Moreover, miR-940 abundance was evidently reduced in HCC tissues and cell lines (SNU-387 and Huh7 cells) (Figure 3D and E). To analyze whether miR-940 was associated with circ_0091579-mediated HCC development, SNU-387 and Huh7 cells were transfected with si-NC, si-circ_0091579, si-circ_0091579 + miR-940 inhibitor or inhibitor NC. The transfection efficacy of miR-940 inhibitor is confirmed in Figure 3F. Furthermore, miR-940 expression was increased via circ_0091579 knockdown, which was weakened via transfection of miR-940 inhibitor (Figure 3G). Besides, down-regulation of miR-940 attenuated silence of circ_0091579-mediated suppression of cell viability, migration, invasion and colony formation, cell cycle arrest at G0/G1 phase, and promotion of apoptosis in SNU-387 and Huh7 cells (Figure 3H–L). These results indicated that circ_0091579 regulated HCC development by mediating miR-940.
 |
Figure 3 The effect of miR-940 on circ_0091579-mediated HCC development in vitro. (A) circ_0091579 expression in nuclear and cytoplasmic fractions. (B) The binding sites of circ_0091579 and miR-940, and the construction of WT-circ_0091579 and MUT-circ_0091579. (C) Luciferase activity was detected in SNU-387 and Huh7 cells transfected with WT-circ_0091579 or MUT-circ_0091579 and miR-940 mimic or miRNA NC. (D) miR-940 expression was measured in HCC and normal tissues (n=30). (E) miR-940 level was detected in SNU-387, Huh7 and THLE-2 cells. (F) miR-940 expression was examined in SNU-387 and Huh7 cells with transfection of miR-940 inhibitor or inhibitor NC. miR-940 level (G), cell viability (H), migration and invasion (I), cycle distribution (J), colony formation (K) and apoptosis (L) were examined in SNU-387 and Huh7 cells transfected with si-NC, si-circ_0091579, si-circ_0091579 + miR-940 inhibitor or inhibitor NC. *P<0.05. |
miR-940 Targets TACR1 to Suppress HCC Development in vitro
Next, the targets of miR-940 were searched using TargetScan, and the binding sequences of miR-940 and TACR1 are displayed in Figure 4A. To identify the relationship of miR-940 and TACR1, we constructed the WT-TACR1-3ʹUTR and MUT-TACR1-3ʹUTR and transfected them into SNU-387 and Huh7 cells. The results showed that miR-940 overexpression led to more than 67% reduction in luciferase activity of WT-TACR1-3ʹUTR, but it did not change the luciferase activity of MUT-TACR1-3ʹUTR (Figure 4B). Additionally, TACR1 protein expression was significantly elevated in HCC tissues and cells (Figure 4C and D). To explore the function of miR-940 and whether it required TACR1, SNU-387 and Huh7 cells were transfected with miRNA NC, miR-940 mimic, miR-940 mimic + pc-TACR1 or pc-NC. The transfection efficacy of miR-940 mimic and pc-TACR1 was validated in Figure 4E and F. Moreover, TACR1 protein expression was evidently declined via miR-940 overexpression, which was restored by the introduction of pc-TACR1 (Figure 4G). Besides, overexpression of miR-940 evidently suppressed cell viability, migration, invasion and colony formation, induced cell cycle arrest at G0/G1 phase, and triggered cell apoptosis in SNU-387 and Huh7 cells (Figure 4H–L). However, these events were mitigated by the restoration of TACR1. These data suggested that miR-940 regulated HCC development via targeting TACR1.
 |
Figure 4 The effect of miR-940 and TACR1 on HCC development in vitro. (A) The binding sequence of miR-940 and TACR1, and the construction of WT-TACR1-3ʹUTR and MUT-TACR1-3ʹUTR. (B) Luciferase activity was examined in SNU-387 and Huh7 cells transfected with WT-TACR1-3ʹUTR or MUT-TACR1-3ʹUTR and miR-940 mimic or miRNA NC. (C) TACR1 protein level was measured in HCC and normal tissues. (D) TACR1 protein level was examined in SNU-387, Huh7 and THLE-2 cells. (E) miR-940 abundance was examined in SNU-387 and Huh7 cells with transfection of miR-940 mimic or miRNA NC. (F) TACR1 protein expression was measured in SNU-387 and Huh7 cells with transfection of pc-TACR1 or pc-NC. TACR1 protein level (G), cell viability (H), migration and invasion (I), cycle distribution (J), colony formation (K) and apoptosis (L) were determined in SNU-387 and Huh7 cells with transfection of miRNA NC, miR-940 mimic, miR-940 mimic + pc-TACR1 or pc-NC. *P<0.05. |
circ_0091579 Regulates TACR1 Expression by miR-940
To test whether and how circ_0091579 could mediate TACR1, the influence of circ_0091579 on TACR1 expression was investigated. As shown in Figure 5A and B, TACR1 protein expression was evidently reduced via circ_0091579 knockdown in SNU-387 and Huh7 cells, which was reversed via TACR1 overexpression or miR-940 knockdown. These results indicated that circ_0091579 could mediate TACR1 by regulating miR-940.
 |
Figure 5 The effect of circ_0091579 on TACR1 expression. (A) TACR1 protein expression was measured in SNU-387 and Huh7 cells transfected with si-NC, si-circ_0091579, si-circ_0091579 + pc-TACR1 or pc-NC. (B) TACR1 protein level was detected in SNU-387 and Huh7 cells transfected with si-NC, si-circ_0091579, si-circ_0091579 + miR-940 inhibitor or inhibitor NC. *P<0.05. |
circ_0091579 Knockdown Decreases Xenograft Tumor Growth
To explore the function of circ_0091579 in HCC development in vivo, Huh7 cells with stable transfection of sh-circ_0091579 or sh-NC were applied to the establishment of xenograft model, and classified as sh-circ_0091579 or sh-NC group. After cell injection for 28 days, the volume and weight of the formed tumor were evidently reduced in sh-circ_0091579 group compared with sh-NC group (Figure 6A and B). Furthermore, the abundances of circ_0091579, miR-940 and TACR1 were detected in the formed tumor tissues. As displayed in Figure 6C–E, circ_0091579 and TACR1 levels were obviously declined in sh-circ_0091579 group, but miR-940 expression was enhanced. These data indicated that circ_0091579 knockdown reduced HCC cell xenograft tumor growth.
 |
Figure 6 The effect of circ_0091579 on xenograft tumor growth. (A and B) Tumor volume and weight were examined in xenograft tumor that was formed by Huh7 cells with transfection of sh-circ_0091579 or sh-NC. (C–E) circ_0091579, miR-940 and TACR1 levels were detected in xenograft tumor that was formed by Huh7 cells with transfection of sh-circ_0091579 or sh-NC. *P<0.05. |
Discussion
HCC is a major type of liver cancer worldwide.26 The biological function of circRNA in HCC development is being a cutting edge.27 Furthermore, the circRNA/miRNA/mRNA regulatory networks have key roles in the progression of HCC.28 In this research, we tested the function of circ_0091579 on HCC development and found the tumor-suppressive role of circ_0091579 knockdown in HCC. Moreover, this study aimed to explore a novel ceRNA mechanism addressed via circ_0091579. Here we were the first to identify the ceRNA crosstalk of circ_0091579/miR-940/TACR1 in HCC cells.
Zhang et al analyzed 20 dysregulated circRNAs using a circRNA microarray, and detected their expression in HCC via qRT-PCR.13 They found that circ_0091579 was a highly expressed circRNA in HCC. However, the role and mechanism of circ_0091579 in HCC are largely unclear. Niu et al suggested that circ_0091579 could promote HCC development via increasing cell viability, colony formation and migration.29 Similarly, we also confirmed these events. Moreover, we further validated that circ_0091579 knockdown could regulate HCC cell invasion and induce cell cycle arrest at G0/G1 phase and cell apoptosis. Collectively, our study indicated the oncogenic role of circ_0091579 in HCC development in vitro.
The circRNA-mediated ceRNA network is the main mechanism for circRNA in HCC development.30 A previous study indicated that circ_0091579 could sponge miR-490-3p in liver cancer.29 This study confirmed that miR-940 was targeted via circ_0091579. Here we found that miR-940 level was reduced in HCC tissues and cells, which was also in agreement with the previous study.31 The former evidences suggested that miR-940 could inhibit HCC cell growth, migration and invasion.16,17 Similarly, we also found that miR-940 overexpression repressed HCC development by decreasing cell viability, colony formation, migration and invasion, inducing cycle arrest at G0/G1 phase, and promoting apoptosis, which was also consistent with that in many other cancers, such as tongue squamous cell carcinoma, glioma and esophageal squamous cell carcinoma.32–34 However, it was opposite to that in endometrial carcinoma.35 We hypothesized it might be caused by the alteration of tumor microenvironment. Our study indicated the anti-cancer role of miR-940 in HCC development. Furthermore, we found that circ_0091579 could mediate HCC development via regulating miR-940.
Next, we validated the interaction between miR-940 and TACR1. A previous study suggested that TACR1 functioned as an oncogene in HCC development.20 In our study, TACR1 expression was enhanced in HCC, indicating the potential carcinogenic role of TACR1 in HCC. Moreover, we identified the oncogenic role of TACR1 in HCC by reversing the anti-cancer function of miR-940, which was also similar to that in neuroblastoma.19 Besides, our results validated that circ_0091579 could regulate TACR1 expression via competitively binding with miR-940, implying that circ_0091579 might target TACR1 by mediating miR-940 to be involved in HCC development in vitro. In xenograft model with nude mice as hosts, the tumors are formed via the injection of cancer cells, which could be used to assess the pathogenesis of HCC in vivo.36 To further explore the anti-cancer role of circ_0091579 in HCC in vivo, we established the murine xenograft model by injecting Huh7 cells, and confirmed that knockdown of circ_0091579 could decrease the tumor growth, which was associated with miR-940/TACR1 axis.
Conclusion
In conclusion, our study validated that circ_0091579 knockdown repressed HCC development in vitro and in vivo, possibly via mediating miR-940/TACR1 axis in a ceRNA-based mechanism. This research indicates a new mechanism for understanding the pathogenesis of HCC.
Acknowledgments
The authors would like to thank the participants in this study.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no conflicts of interest in this work.
References
1. Forner A, Reig M, Bruix J. Hepatocellular carcinoma. Lancet. 2018;391(10127):1301–1314. doi:10.1016/S0140-6736(18)30010-2
2. Grandhi MS, Kim AK, Ronnekleiv-Kelly SM, et al. Hepatocellular carcinoma: from diagnosis to treatment. Surg Oncol. 2016;25:74–85. doi:10.1016/j.suronc.2016.03.002
3. Dutta R, Mahato RI. Recent advances in hepatocellular carcinoma therapy. Pharmacol Ther. 2017;173:106–117. doi:10.1016/j.pharmthera.2017.02.010
4. Yin Y, Long J, He Q, et al. Emerging roles of circRNA in formation and progression of cancer. J Cancer. 2019;10:5015–5021. doi:10.7150/jca.30828
5. Wang M, Yu F, Li P. Circular RNAs: characteristics, function and clinical significance in hepatocellular carcinoma. Cancers (Basel). 2018;10:258. doi:10.3390/cancers10080258
6. Xiong DD, Dang YW, Lin P, et al. A circRNA-miRNA-mRNA network identification for exploring underlying pathogenesis and therapy strategy of hepatocellular carcinoma. J Transl Med. 2018;16:220. doi:10.1186/s12967-018-1593-5
7. Yao Z, Xu R, Yuan L, et al. Circ_0001955 facilitates hepatocellular carcinoma (HCC) tumorigenesis by sponging miR-516a-5p to release TRAF6 and MAPK11. Cell Death Dis. 2019;10:945. doi:10.1038/s41419-019-2176-y
8. Li Z, Liu Y, Yan J, et al. Circular RNA hsa_circ_0056836 functions an oncogenic gene in hepatocellular carcinoma through modulating miR-766-3p/FOSL2 axis. Aging (Albany NY). 2020;12:2485–2497. doi:10.18632/aging.102756
9. Pu J, Wang J, Li W, et al. hsa_circ_0000092 promotes hepatocellular carcinoma progression through up-regulating HN1 expression by binding to microRNA-338-3p. J Cell Mol Med. 2020. doi:10.1111/jcmm.15010
10. Liu Z, Yu Y, Huang Z, et al. CircRNA-5692 inhibits the progression of hepatocellular carcinoma by sponging miR-328-5p to enhance DAB2IP expression. Cell Death Dis. 2019;10(12):900. doi:10.1038/s41419-019-2089-9
11. Su X, Su J, He H, et al. Hsa_circ_0070269 inhibits hepatocellular carcinoma progression through modulating miR-182/NPTX1 axis. Biomed Pharmacother. 2019;120:109497. doi:10.1016/j.biopha.2019.109497
12. Chen H, Liu S, Li M, et al. circ_0003418 inhibits tumorigenesis and cisplatin chemoresistance through Wnt/beta-catenin pathway in hepatocellular carcinoma. Onco Targets Ther. 2019;12:9539–9549. doi:10.2147/OTT.S229507
13. Zhang C, Zhang C, Lin J, et al. Circular RNA Hsa_Circ_0091579 serves as a diagnostic and prognostic marker for hepatocellular carcinoma. Cell Physiol Biochem. 2018;51:290–300. doi:10.1159/000495230
14. Mahgoub A, Steer CJ. MicroRNAs in the evaluation and potential treatment of liver diseases. J Clin Med. 2016;5:52. doi:10.3390/jcm5050052
15. Anwar SL, Lehmann U. MicroRNAs: emerging novel clinical biomarkers for hepatocellular carcinomas. J Clin Med. 2015;4:1631–1650. doi:10.3390/jcm4081631
16. Yuan B, Liang Y, Wang D, et al. MiR-940 inhibits hepatocellular carcinoma growth and correlates with prognosis of hepatocellular carcinoma patients. Cancer Sci. 2015;106:819–824. doi:10.1111/cas.12688
17. Ding D, Zhang Y, Yang R, et al. miR-940 suppresses tumor cell invasion and migration via regulation of CXCR2 in hepatocellular carcinoma. Biomed Res Int. 2016;2016:7618342. doi:10.1155/2016/7618342
18. Xu H, Sun Y, Ma Z, et al. LOC134466 methylation promotes oncogenesis of endometrial carcinoma through LOC134466/hsa-miR-196a-5p/TAC1 axis. Aging (Albany NY). 2018;10:3353–3370. doi:10.18632/aging.101644
19. Pan J, Zhang D, Zhang J, et al. LncRNA RMRP silence curbs neonatal neuroblastoma progression by regulating microRNA-206/tachykinin-1 receptor axis via inactivating extracellular signal-regulated kinases. Cancer Biol Ther. 2019;20:653–665. doi:10.1080/15384047.2018.1550568
20. Hongfeng Z, Andong J, Liwen S, et al. lncRNA RMRP knockdown suppress hepatocellular carcinoma biological activities via regulation miRNA-206/TACR1. J Cell Biochem. 2020;121:1690–1702. doi:10.1002/jcb.29404
21. Dudekula DB, Panda AC, Grammatikakis I, et al. CircInteractome: a web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016;13:34–42. doi:10.1080/15476286.2015.1128065
22. Riffo-Campos AL, Riquelme I, Brebi-Mieville P. Tools for sequence-based miRNA target prediction: what to choose? Int J Mol Sci. 2016;17:1987. doi:10.3390/ijms17121987
23. Chomczynski P, Sacchi N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: twenty-something years on. Nat Protoc. 2006;1:581–585. doi:10.1038/nprot.2006.83
24. Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods. 2001;25:402–408. doi:10.1006/meth.2001.1262
25. Xia S, Feng J, Chen K, et al. CSCD: a database for cancer-specific circular RNAs. Nucleic Acids Res. 2018;46:D925–D929. doi:10.1093/nar/gkx863
26. Villanueva A. Hepatocellular Carcinoma. N Engl J Med. 2019;380:1450–1462. doi:10.1056/NEJMra1713263
27. Qiu L, Xu H, Ji M, et al. Circular RNAs in hepatocellular carcinoma: biomarkers, functions and mechanisms. Life Sci. 2019;231:116660. doi:10.1016/j.lfs.2019.116660
28. Sun X, Ge X, Xu Z, et al. Identification of circular RNA-microRNA-messenger RNA regulatory network in hepatocellular carcinoma by integrated analysis. J Gastroenterol Hepatol. 2020;35:157–164. doi:10.1111/jgh.14762
29. Niu WY, Chen L, Zhang P, et al. Circ_0091579 promotes proliferative ability and metastasis of liver cancer cells by regulating microRNA-490-3p. Eur Rev Med Pharmacol Sci. 2019;23:10264–10273. doi:10.26355/eurrev_201912_19664
30. Wu J, Liu S, Xiang Y, et al. Bioinformatic analysis of circular RNA-associated ceRNA network associated with hepatocellular carcinoma. Biomed Res Int. 2019;2019:8308694. doi:10.1155/2019/8308694
31. Li P, Xiao Z, Luo J, et al. MiR-139-5p, miR-940 and miR-193a-5p inhibit the growth of hepatocellular carcinoma by targeting SPOCK1. J Cell Mol Med. 2019;23:2475–2488. doi:10.1111/jcmm.14121
32. Ma T, Zhao Z, Wang Z, et al. MiR-940 inhibits migration and invasion of tongue squamous cell carcinoma via regulatingCXCR2/NF-kappaB system-mediated epithelial-mesenchymal transition. Naunyn Schmiedebergs Arch Pharmacol. 2019;392:1359–1369. doi:10.1007/s00210-019-01671-w
33. Luo H, Xu R, Chen B, et al. MicroRNA-940 inhibits glioma cells proliferation and cell cycle progression by targeting CKS1. Am J Transl Res. 2019;11:4851–4865.
34. Wang H, Song T, Qiao Y, et al. miR-940 inhibits cell proliferation and promotes apoptosis in esophageal squamous cell carcinoma cells and is associated with post-operative prognosis. Exp Ther Med. 2020;19:833–840. doi:10.3892/etm.2019.8279
35. Zhou Z, Xu Y, Wang L, et al. miR-940 potentially promotes proliferation and metastasis of endometrial carcinoma through regulation of MRVI1. Biosci Rep. 2019;39:BSR20190077. doi:10.1042/bsr20190077
36. Heindryckx F, Colle I, Van Vlierberghe H. Experimental mouse models for hepatocellular carcinoma research. Int J Exp Pathol. 2009;90:367–386. doi:10.1111/j.1365-2613.2009.00656.x
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.