Back to Journals » OncoTargets and Therapy » Volume 12
The functions of microRNA-124 on bladder cancer
Authors Cao Z, Xu L, Zhao S, Zhu X
Received 6 November 2018
Accepted for publication 2 March 2019
Published 7 May 2019 Volume 2019:12 Pages 3429—3439
DOI https://doi.org/10.2147/OTT.S193661
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr XuYu Yang
Zhigang Cao,1* Li Xu,1* Shuli Zhao,2 Xu Zhu1
1Department of Urology, Nanjing First Hospital, Nanjing Medical University, Nanjing, China; 2Central Laboratory, Nanjing First Hospital, Nanjing Medical University, Nanjing, China
*These authors contributed equally to this work
Background: To detect the expression of miR-124 in bladder cancer (BC) cell lines and tissue specimens and to analyze its association with the growth of the BC cells.
Methods: Quantitative real-time polymerase chain reaction (qPCR) was applied to examine the expression of miR-124 in BC cell lines and tissues. The function of miR-124 in modulating cell proliferation was assessed in BC cells with miRNA-124 overexpression; the cell viability was identified by Cell Count Kit-8; flow cytometry was employed to detect the cell cycle; the expressions of E2F3, cyclin-dependent kinase 4 (CDK4), Ki-67 and vascular endothelial growth factor (VEGF) were tested by qPCR and Western blot; angiogenesis experiment was performed to analysis changes in angiogenesis rate; and bioinformatics prediction and dual luciferase reporter system were employed to identify the target of miR-124.
Results: Survival curve data showed that the expression of MicroRNA-124 was positively correlated with survival. MicroRNA-124 expression was significantly decreased in BC cell lines and tissues. Bioinformatics prediction and dual luciferase reporter system verified CDK4 as a direct target of miR-124, which regulated the proliferation of BC cells by directly inhibiting CDK4. BC cells over-expressing miR-124 showed significantly inhibited cell viability, decreased angiogenesis rate, prevented cell proliferation and diminished the expression of E2F3, CDK4, Ki-67 and VEGF. All of these changes were reversed by over-expressing CDK4.
Conclusion: MicroRNA-124 suppressed the proliferation of CRC cells by directly targeting CDK4, which provides a target for improving the therapeutic effect of BC.
Keywords: miR-124, bladder cancer, CDK4, proliferation, VEGF
Introduction
One of the most common malignant tumors of the urinary system is bladder cancer (BC), and it is also one of the top ten common tumors in the body, accounting for the first incidence of urogenital tumors in China.1 Clinically, BC is often treated with tumor resection and postoperative adjuvant chemotherapy. However, due to the high metastatic nature of BC, the recurrence rate and mortality rate of BC remain high. Therefore, it is necessary to search for effective therapeutic targets and design effective treatment programs to improve clinical treatment efficiency and improve patient survival.
MiRNAs are a highly conserved class of non-coding small RNA molecules involved in many life and disease processes, not only in cell proliferation, apoptosis, differentiation, signal transduction and metabolism, but also in tumor cell transformation.2,3 And it also plays a vital role in the occurrence and development of various diseases.4,5 There is increasing evidence that the disordered expression of miRNAs contributes to the progression of human cancers.6–9 For example, abnormal expression of miRNA in BC is thought to benefit for the development of BC.10,11 Studies find that miRNA can be used as a marker for diagnosing BC and prognosis.12–14 MicroRNA-124 is considered to be brain-enriched and is an extremely conserved gene.15 The similarity of the mature sequence of the gene in different five is 89.54%, and most of the target sites are also very conservative.16 Therefore, MicroRNA-124 has extremely important biological functions, and miR-124 is one of the most well-studied microRNAs currently studied.17 MicroRNA-124 has been shown to have a potential antitumor function in glioma, lung cancer, hepatocellular carcinoma and BC.18–20 Studies have shown that miR-124 regulates cell cycle and movement by targeting CDK4.21–23 The recovery of miR-124 may be an effective anticancer therapy.24 The specific function of miR-124 in the progression of BC, particularly its molecular mechanism by which miR-124 exerts its function and regulates the malignant phenotype of BC cells, is poorly understood.
In this study, we aimed to conduct an in-depth study of miR-124 to elucidate its molecules in the progression and metastasis of malignant tumors, especially BC, and to explore a diagnostic method based on miR-124 and treatment with miR-124 for the theoretical value and clinical guiding significance of treating cancer.
Materials and methods
Clinical samples and cells
The current study was approved by the Ethics Committee of the Shaoyifu Hospital affiliated to Zhejiang University (Hangzhou, China). From March 2016 to April 2017, 83 bladder tissues and their adjacent non-tumor tissues were collected in the urology department of Shaoyifu Shaw Hospital, and informed consent was obtained from the patients. This research protocol has been approved by the ethics committee. Hek 293, human normal cell SV-HUC-1 and bladder cancer cell line T24, 5637, J82 and UM-UC-3 were all obtained from American Type Culture Collection (Manassas, VA) and cultured until they reached the log phase in Dulbecco’s Modified Eagle Medium (DMEM) containing 10% fetal bovine serum at 37°C in a 5% CO2 humidified incubator.
Reagent
Transfection reagent LipofectamineTM 3000 was purchased from Invitrogen (Thermo Fisher Scientific, Inc.). Cell Count Kit-8 (CCK-8) assay kit was obtained from Nanjing Jiancheng Bioengineering Institute (Nanjing, China). Transfected plasmids and miRNAmimics were bought from GenePharma (Shanghai, China),
Transfection
LipofectamineTM 3000 (Thermo Fisher Scientific, Inc.) was performed according to the instructions. In brief, the lipid complex was prepared by combining the reagent of 4 µL LFN Plus with 2 μg of plasmid DNA suspended in 1 mL serum-free medium and incubated for 15 mins at room temperature. The solution was then mixed with 40 µL of LFN in 1 mL serum-free medium and incubated further at room temperature for 15 mins. And lipid compounds will be diluted in serum-free medium to produce a 5 mL volume required concentration and with the cells at 37°C, 5% CO2 incubation under 24 hrs for subsequent experiments.
Bioinformatics prediction
In order to predict the potential target genes of miR-124, Targetscan and miRTarBase online software were performed, according to the manufacturer’s instruction. “miR-124” was inserted and “human” was selected. The putative target genes of miR-124 were scanned.
Dual-luciferase reporter gene assay
Hek 293 cells were transfected with 100 nM E2F3-3ʹ-UTR or CDK4-3ʹ-UTR plasmid, as well as with or without 100 nM miR-124 mimic. Then, luciferase activity was measured by luciferase reporter assay system (Promega Corporation) in Lmax II luminescence meter (Molecular Devices, LLC, Sunnyvale, CA, USA) for 48 hrs after transfection. Renin luciferase activity was defined as the standardization of firefly luciferase activity.
Cell viability assay
After the cells were transfected at 24 and 48 hrs post-culture, 10 μL of CCK-8 solution was added to the wells and the cells were incubated for 2 h at 37°C in an incubator with 5% CO2 in the dark. Subsequently, microplate reader (Bio-Rad Laboratories, Inc., Hercules, CA, USA) was employed to determine the OD of each well in different cell groups at an absorbance of 450 nm. Cell viability was detected by CCK-8 assay kit according to the manufacturer’s instructions.
Western blot
The phosphate buffer saline (PBS) was employed to wash cells twice, and the cells were added to protein lysis buffer (RIPA; Cell Signaling Technology, Inc., Danvers, MA, USA) for 2 hrs on ice, centrifuged at 12,000 x g for 30 mins at 4°C, and then the supernatant was collected. The protein concentration was tested by the BCA protein kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA) and adjusted to a concentration of 5 μg/μl using 1× loading and DEPC water. Six microliters (at least 30 μg) of the samples were electrophoresed (80 V for 30 min and then transferred to 120 V for 1.5 h) on a 10% running gels. The gels were transferred to polyvinylidene fluoride membrane (Bio-Rad Laboratories, Inc., Hercules, CA, USA) on ice for 110 mins at 110 V. A 5% nonfit milk was employed to block the membrane and eluted three times with PBS for 5 mins each time. The bands were then incubated overnight with the corresponding primary antibody, washed with PBS three times for 15 mins, incubated with secondary antibody: horseradish peroxidase–conjugated goat anti-mouse/rabbit IgG (1:2000; sc-516102/sc-2357; Santa Cruz Biotechnology, Inc. Dallas, TX, USA) for 2 hr at room temperature, washed with PBS 3 times for 15 mins each time, and furthermore washed once with PBS/0.1% Tween-20 (PBST) for 15 mins. Developments were carried out with a developer (EZ-ECL kit; Biological Industries BI), and the gray values of the strips were analyzed and counted by imageJ (version 5.0; Bio-Rad, Hercules, CA, USA). The antibodies used in the present study were anti-β-actin (rabbit; 1:1000; LS-B1625; LifeSpan BioSciences, Inc.), anti- E2F3 (rabbit; 1:1000; ab50917; Abcam), anti-CDK4 (rabbit; 1:1000; ab137675; Abcam), anti- VEGF ((rabbit; 1:1000; #2463; CST) and anti- Ki-67 (rabbit; 1:1000; ab16667; Abcam).
RNA isolation and real-time PCR
The cell culture medium in each well was aspirated as much as possible, and 1 mL of Trizol (Invitrogen, Carlsbad, California) was added to the MC3T3-E1 cells. The cells were placed horizontally for a while and blow evenly. The cells containing the lysate were transferred to a 1.5 mL EP tube and allowed to keep at room temperature for 5 mins. Two hundred microlieters of chloroform were added to each tube and inverted for 15 s. After emulsification, let stand for 5 mins. After centrifugation at 12,000 x g for 15 mins at 4°C, the upper aqueous phase was pipetted into a new 1.5 mL of EP and an equal volume of isopropanol (about 400 μL) was added to each tube and allowed to keep for 10 min at room temperature. The supernatant was discarded and 1 mL of pre-cooled 75% ice ethanol was added after centrifugation at 12,000 x g for 15 mins at 4°C. After centrifugation at 7500 x g for 10 mins at 4°C, the supernatant was discarded. An appropriate amount of DEPC (20 μL) was added to dissolve the RNA. The purity and concentration of RNA were tested by the NanoDrop nd-1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). According to the program provided by the manufacturer (Thermo Fisher Scientific, Waltham, USA), reverse transcription cDNA kit was used to reverse transcribe 1 µg total RNA for the synthesis of cDNA (42°C for 60 mins, 70°C for 5 mins, 4°C preservation). SYBR Green PCR Master Mix (Roche, Basle, Switzerland) was used to perform quantitative real-time polymerase chain reaction (qPCR) experiment using Opticon real-time PCR Detection System (ABI 7500, Life technology, USA), The PCR cycle was as follows: pretreatment at 95°C for 10 mins; followed by 40 cycles of 94°C for 15 s, 60°C for 1 min, finally at 60°C for 1 min and at 4°C for preservation. The relative mRNA quantity was determined using the comparative cycle threshold (ΔΔCt) method.25 GAPDH expression was used for normalization. The primer sequences are used for qRT-PCR analysis as shown in Table 1.
 | Table 1 Primers for RT-qPCR |
In vitro angiogenesis experiment
Matrigel plug angiogenesis assay (Corning, NY, USA) was used to evaluate the in vivo anti-angiogenic effect of miRNA-124. The matrigel stock solution was thawed overnight at 4°C. A gel solution was prepared using a Matrigel stock solution and serum-free DMEM, and the solution was placed in a 96-well plate and then allowed to incubate for 2 hrs to cure. The cultured cells were collected and digested to prepare a single cell suspension under aseptic conditions, and the cell suspension was adjusted to a density of 1×105/mL. The cells were seeded in 96-well plates at 100 μL per well. The plates were incubated for 6–8 hrs in an incubator (5% CO2, 37°C), and the cells were visualized using an inverted microscope (Thermo Fisher Scientific) and photographed.
Flow cytometry cell cycle analysis
According to the standard method, cell cycle analysis was performed. In short, 5×105 cells with 70% cold ethanol under –20°C overnight. The next day, the fixed cells were centrifuged at 1200 x g for 1 min and washed twice with PBS. After the cells with 200 µL RNase A (1 mg/ml) for 10 min at 37°C in suspension, then add 300 µl propidium iodine (PI, 100 mu l/mL, BioVision, Milpitas, CA, USA) to dyeing the DNA in cells in the dark. After being incubated at room temperature for 20 mins, cells in FAC Scan flow cytometer (Becton Dickinson, Franklin ‘lakes, NJ, USA) in the analysis of cellular DNA content, and through the Mod Fit LT software V2.0 (Becton Dickinson) analyzed the data.
Statistical analysis
Data appear as the means ± SD. Differences between the experimental groups were assessed by ANOVA or a two-tailed Student’s t-test. Differences were analyzed by GraphPad Prism 6, and all comparisons with a p<0.05 were considered statistically significant.
Results
MicroRNA-124 was obviously reduced in BC and correlated with malignant progression and poor prognosis
In the present study, qPCR data demonstrated that the expression of miR-124 was significantly lower in BC tissues than that in the adjacent non-tumor tissues (Figure 1A). Further exploration revealed that the BC patients with low miR-124 expression had a shorter survival time when compared with those presenting a high miR-124 expression (Figure 1B). Accordingly, miR-124 downregulation may contribute to the BC aggressiveness as well as the poor prognosis of patients.
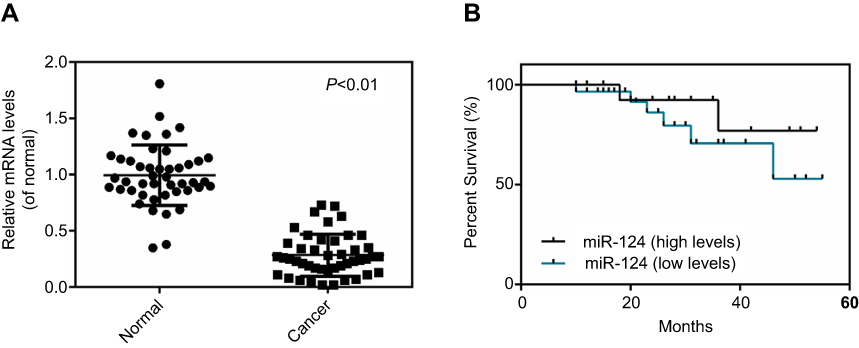 | Figure 1 MicroRNA-124 was downregulated inbladder cancer ( BC) tissues. (A) Reverse transcription-quantitative polymerase chain reaction was conducted to test the miR-124 expression in BC tissues and matched adjacent non-tumor tissues. (B) BC patients with low miR-124 expression presented a shorter survival time when compared with patients with a high expression of miR-124. P<0.01 vs normal. |
MicroRNA-124 was significantly decreased in BC cell lines
The miR-124 expression levels were then examined in several common BC cell lines (T24, 5637, J82 and UM-UC-3), using normal bladder epithelial SV-HUC-1 cells as the control. In accordance with the findings in the BC tissues, qPCR data revealed that miR-124 was also significantly downregulated in the four BC cell lines compared with the normal SV-HUC-1 cells (Figure 2). Since miR-124 expression was downregulated to a greater extent in T24 cells and 5637 cells, the two cell lines were used in subsequent experiments.
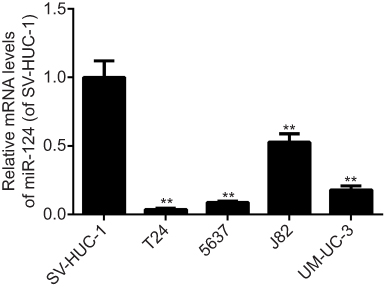 | Figure 2 MicroRNA-124 was obviously decreased in bladder cancer ( BC) cell lines. Reverse transcription-quantitative polymerase chain reaction was conducted to examine the miR-124 expression in four BC cell lines and normal bladder mucosa epithelial cells. **P<0.01 vs SV-HUC-1. |
Overexpression of miR-124 inhibited cells viability, E2F3 and CDK4 were predicted to be target genes of miR-124
T24 cells and 5637 cells were transfected with miR-124 mimic or miR-NC. As shown in (Figure 3A and B), the miR-124 levels were significantly higher in the mimic group compared with the miR-NC group and control. The study further demonstrated that the overexpression of miR-124 significantly decreased the cells viability when compared with the miR-NC group and control after transfection 24 and 48 hrs (Figure 3C and D). The potential target genes of miR-124 were further predicted using bioinformatics analysis (Figure 3E) and RT-PCR (Figure 3F and G), and the results showed that E2F3 and CDK4 were predicted to be putative target genes of miR-124.
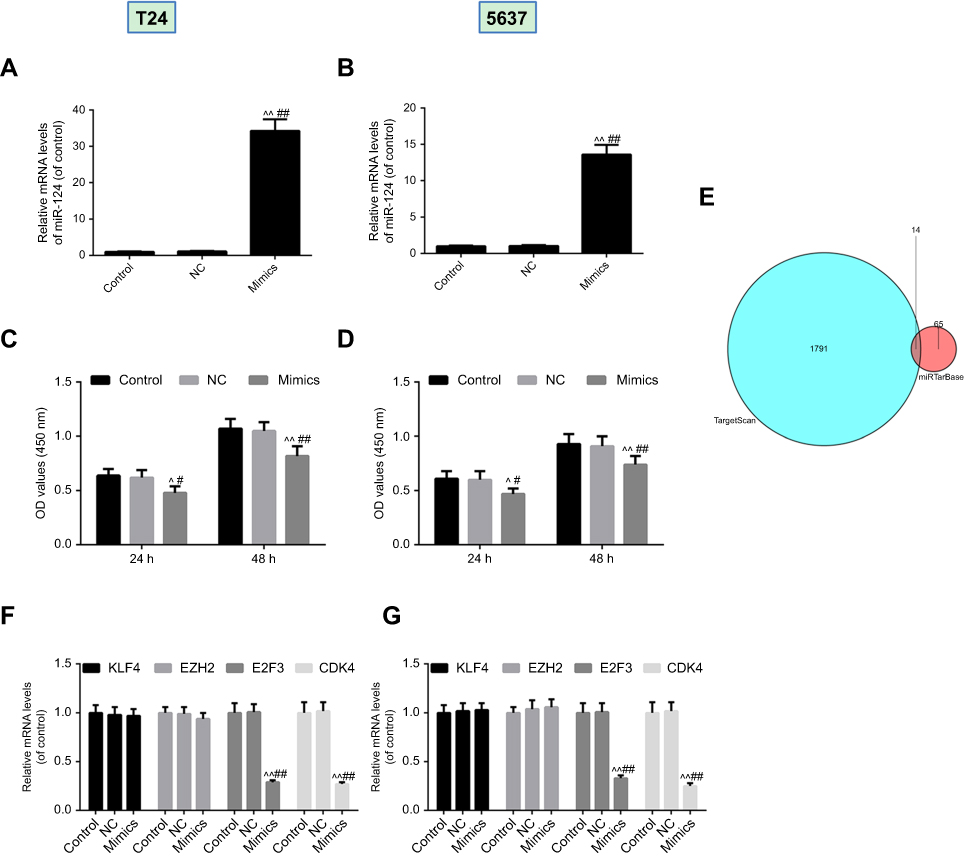 | Figure 3 Overexpression of miR-124 suppressed cells viability and E2F3 and CDK4 was predicted to be target genes of miR-124. T24 (A) cells and 5637 (B) cells transfected with miR-124 mimic or miR-NC. Cell Count Kit-8 (CCK-8) was used to detect the viability of T24 (C) cells and 5637 (D) cells. (E) The potential targets of miR-124 were analyzed by bioinformatics prediction. Reverse transcription-quantitative polymerase chain reaction was conducted to confirm the potential target of miR-124 in T24 (F) cells and 5637 (G) cells. ^P<0.05, ^^P<0.01 vs control, #P<0.05, ##P<0.01 vs NC. |
MicroRNA-124 has stronger binding ability to CDK4
To determine the binding capacity of miR-124 to CDK4 and E2F3, Targetscan website was used to find target sites (Figure 4A) The. dual-luciferase reporter gene assay data indicated that the luciferase activity was lower in CDK4- 3′-UTR than that in E2F3- 3′-UTR (Figure 4B).
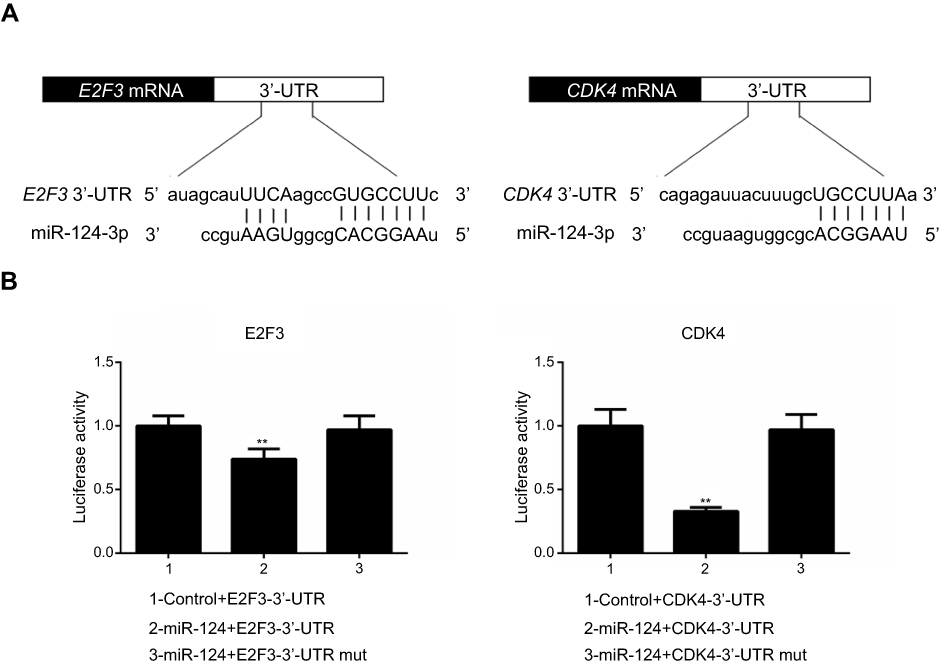 | Figure 4 (A) TargetScan website was used to find target sites. (B) The luciferase activity was detected by dual-luciferase reporter gene assay. **P<0.01 vs 1. |
Overexpression of CDK4 inhibited miRNA-124 inhibition of cell viability, angiogenesis and cell cycle on T24 cells
The effect of CDK4 was further examined in T24 cells. As shown in (Figure 5A), CDK4 was successfully transfected with T24 cells. Furthermore, we discovered that the inhibition of miRNA-124 on cell viability (Figure 5B), angiogenesis (Figure 5C) and cell cycle (Figure 5D) was reversed by CDK4. As shown in (Figure 5E and F) were angiogenesis and cell cycle images, respectively.
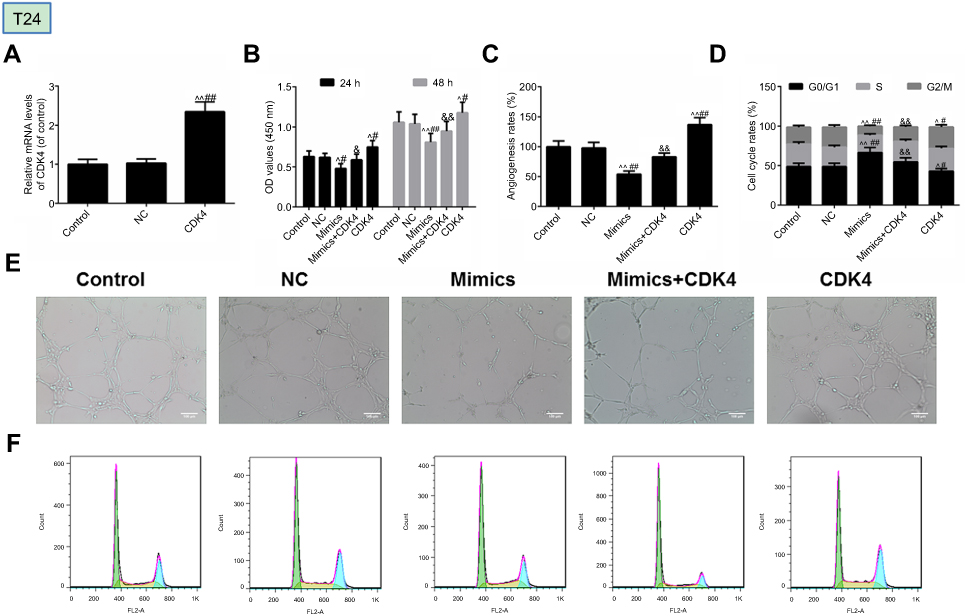 | Figure 5 MicroRNA-124 inhibits the malignant phenotypes of T24 cells. (A) Transfection efficiency of CDK4 was tested by reverse transcription-quantitative polymerase chain reaction. (B) Cell Count Kit-8 (CCK-8) was used to detect the viability of cells. (C) Angiogenesis rate was identified by in vitro angiogenesis experiment. (D) Cell cycle was described by flow cytometry. The images of angiogenesis rate and cell cycle were presented in (E) and (F), respectively. ^P<0.05, ^^P<0.01 vs control, #P<0.05, ##P<0.01 vs NC. &P<0.05, &&P<0.01 vs mimics. |
Overexpression of CDK4 inhibited miRNA-124 inhibition of cell viability, angiogenesis, and cell cycle on 5637 cells
The effect of CDK4 was further examined in 5637 cells. As shown in Figure 6A, CDK4 was successfully transfected with T24 cells. Furthermore, we found that the inhibition of miRNA-124 on cell viability (Figure 6B), angiogenesis (Figure 6C) and cell cycle (Figure 6D) was reversed by CDK4. As shown in Figure 6E and F were angiogenesis and cell cycle images, respectively.
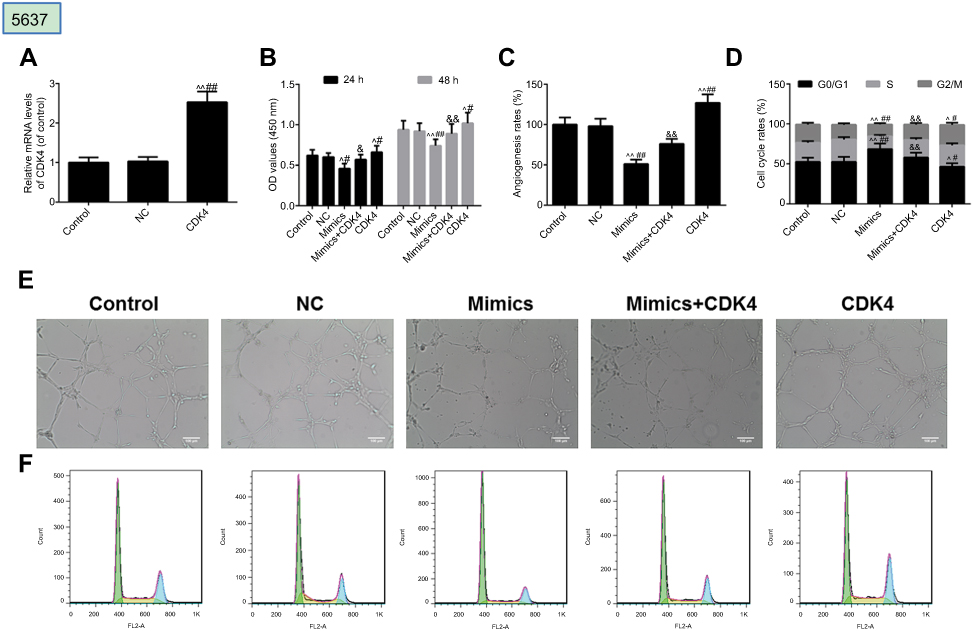 | Figure 6 MicroRNA-124 inhibits the malignant phenotypes of 5637 cells. (A) Transfection efficiency of CDK4 was tested by reverse transcription-quantitative polymerase chain reaction. (B) Cell Count Kit-8 (CCK-8) was used to detect the viability of cells. (C) Angiogenesis rate was identified by in vitro angiogenesis experiment. (D) Cell cycle was described by flow cytometry. The images of angiogenesis rate and cell cycle were presented in (E) and (F), respectively. ^P<0.05, ^^P<0.01 vs control, #P<0.05, ##P<0.01 vs NC. &P<0.05, &&P<0.01 vs mimics. |
Overexpression of miRNA-124 inhibited the expression of E2F3, CDK4, Ki-67 and VEGF, which reversed by CDK4
In this study, qPCR (Figure 7A and B) and Western blot (Figure 7C, D, E, F) were used to analyze the expression of E2F3, CDK4, Ki-67 and VEGF, and the findings demonstrated that miRNA-124 inhibited the expression of E2F3, CDK4, Ki-67 and VEGF, which were reversed by CDK4 in T24 cells and 5637 cells.
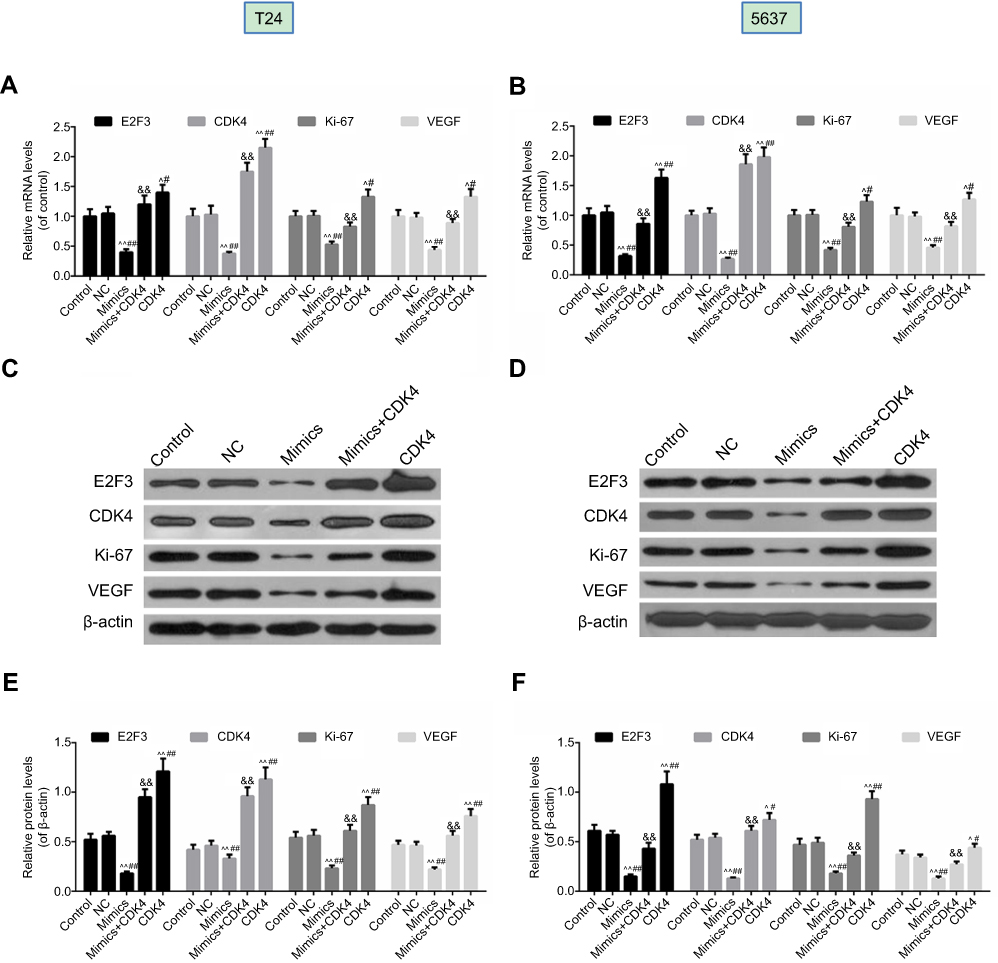 | Figure 7 Overexpression of miRNA-124 inhibited the expression of E2F3, CDK4, Ki-67 and VEGF, which were reversed by CDK4. Reverse transcription-quantitative polymerase chain reaction was used to evaluate the expression levels of E2F3, CDK4, Ki-67 and VEGF in T24 (A) cells and 5637 (B) cells. Western blot was used to determine the expression levels of E2F3, CDK4, Ki-67 and VEGF in T24 (C) cells and 5637 (D) cells. The relative levels of proteins were counted by β-actin as normalization in T24 (E) cells and 5637 (F) cells). ^P<0.05, ^^P<0.01 vs control, #P<0.05, ##P<0.01 vs NC, &&P<0.01 vs mimics. |
Discussion
BC is one of the most common cancers caused by the epithelial lining of the bladder.26 Although surgical treatment has improved the survival rate of many patients, its five-year survival rate is only about 60% due to the easy transfer and invasion of cancer.27,28 Thus, it is urgent to identify new targets for effective treatment of bladder tumors. MicroRNA-124 is involved in the occurrence and development of BC18,29 However, its specific mechanism of action is still unclear. In this study, we aimed to study the mechanism of action of miRNA-124 in BC and provide a promising new treatment for clinical treatment of BC.
In the current study, many miRNAs have been found to be abnormally expressed in BC, playing a role in promoting or inhibiting tumors.30–33 Abnormal expression in miRNA tumor tissues is reflected in two aspects: up-regulation and down-regulation. For example, miR-122 is down-regulated in BC cells, mediating tumor invasion and metastasis,34 and miR-187-5p is up-regulated in BC cells, increasing the risk of tumor recurrence.35 In our study, we discovered that miRNA-124 was down-regulated in BC tissues and cell lines, when compared with matched adjacent non-tumor tissues or normal bladder epithelial cells, respectively. At the same time, the survival rate of patients was tracked clinically, and the data showed that there was a positive association between the expression of miRNA-124 and survival rate. These experimental results suggested that low expression of miRNA-124 was involved in the proliferation of BC, consistent with the previous two studies.36,37 Based on these previous studies and current research observations, it was suggested that down-regulation of miRNA-124 may contribute to the progression of the BC and poor therapeutic effects.
To further confirm the role of miRNA-124 in BC, we established a miRNA-124 overexpressing cell model in T24 cells and 5637 cells and observed that overexpressed miRNA-124 markedly reduced cell viability and angiogenesis rate and inhibited proliferation in T24 cells and 5637 cells. Similarly, studies have found that miRNA-124 overexpression inhibits cell viability, angiogenesis and cell cycle in BC cell lines.18,36,37
It is well known that miRs function by modulating their target genes, and the direct target of miRNA-124 was explored in BC. Bioinformatics prediction, dual luciferase assay and qPCR data confirmed that CDK4 was a direct target gene for miRNA-124. CDK4 is a key member of the cell cycle-related subfamily,38 and it is reported to be involved in many physiological processes, including proliferation and apoptosis.39,40 In addition, CDK4 has been demonstrated to be regulated by post-translational modification of miR and acts downstream of multiple signaling pathways involved in cell proliferation and movement, and the carcinogenic effects of CDK4 have gradually emerged in some common human cancers, including BC.30,41,42 It was further observed that overexpression of CDK4 impaired the suppressive functions of miR-124 on BC cell viability, angiogenesis and proliferation in T24 cells and 5637 cells. Furthermore, CDK4 increased the expression of VEGF that promoted angiogenesis rate. Ki-67, a determining factor for prognosis of BC,43 was increased by CDK4 in mimics+CDK4 group, when compared with mimics. Meanwhile, the expressions of E2F3 and CDK4 were increased in mimics+CDK4 group compared with mimics group. These data demonstrated that miR-124 had suppressive effects on BC cell viability, angiogenesis and proliferation, at least partly, via targeting CDK4. A study has found that miR-124 played an important role in the occurrence and development of BC. Wang’s study found that miR-124 could regulate STAT3-mediated cell migration and apoptosis in bladder cancer.44 It has also been found that miR-124 can regulate BC cell apoptosis or metastasis by targeting CAV1 or DNMT3B.45,46 However, further research is needed on the mechanism by which E2F3 participates in BC. We have enough reason to believe that E2F3 was also likely to participate in the development of BC. For economic reasons, we will delve into it if we have the opportunity.
In the current study, we found that miRNA-124 was downregulated in BC tissues and cell lines, and this low expression was associated with the BC aggressiveness, as well as poor prognosis of patients. Overexpression of miR-124 induced by mimic transfection was observed to inhibit the cells viability, angiogenesis and proliferation. In addition, CDK4 was identified as a target gene of miR-124. Furthermore, overexpression of CDK4 impaired the inhibitory effects of miR-124 on the malignant phenotypes of T24 cells and 5637 cells. Thus, the current study expanded the understanding of the miR-124/CDK4 function in human cancer, at least in BC. Accordingly, these results suggested that miR-124 may be applied as a promising therapeutic candidate for BC treatment.
Ethics approval and consent to participate
Patient consent was written informed consent. All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.No animals were involved in this research.
Acknowledgments
This work was supported by the Project of Nanjing Health Bureau and the Project of Nanjing Medical Science and Technology Development [YKK07041]; the Project of Nanjing Science and Technology Bureau and the Project of Nanjing Science Development [200801080]; the Project of Nanjing Health Bureau and the Project of Nanjing Medical Science and Technology Development [YKK08102]; Nanjing Medical Development Fund and the Project of Nanjing Medical Science and Technology Development [YKK11125]; the Project of Nanjing Science and Technology Bureau and the Project of Nanjing Science Development [201208021]; and the Key Project of Nanjing Municipal Health Bureau and the Project of Nanjing Medical Science and Technology Development [ZKX12021].
Disclosure
The authors report no conflicts of interest in this work.
References
1. Jemal A, Siegel R, Ward E, et al. Cancer statistics, 2008. CA Cancer J Clin. 2008;58(2):71–96. doi:10.3322/CA.2007.0010
2. Yang S, Sun Z, Zhou Q, et al. MicroRNAs, long noncoding RNAs, and circular RNAs: potential tumor biomarkers and targets for colorectal cancer. Cancer Manag Res. 2018;10:2249–2257. doi:10.2147/CMAR.S166308
3. Fan C, Tang Y, Wang J, et al. The emerging role of Epstein-Barr virus encoded microRNAs in nasopharyngeal carcinoma. J Cancer. 2018;9(16):2852–2864. doi:10.7150/jca.25460
4. Bier A, Giladi N, Kronfeld N, et al. MicroRNA-137 is downregulated in glioblastoma and inhibits the stemness of glioma stem cells by targeting RTVP-1. Oncotarget. 2013;4(5):665–676. doi:10.18632/oncotarget.928
5. Deng D, Xue L, Shao N, et al. miR-137 acts as a tumor suppressor in astrocytoma by targeting RASGRF1. Tumour Biol. 2016;37(3):3331–3340. doi:10.1007/s13277-015-4110-y
6. Qiu L, Zhang G-F, Yu L, Wang H-Y, Jia X-J, Wang T-J. Novel oncogenic and chemoresistance-inducing functions of resistin in ovarian cancer cells require miRNAs-mediated induction of epithelial-to-mesenchymal transition. Sci Rep. 2018;8(1):12522. doi:10.1038/s41598-018-30978-6
7. Hon KW, Abu N, Ab Mutalib N-S, Jamal R. miRNAs and lncRNAs as predictive biomarkers of response to FOLFOX therapy in colorectal cancer. Front Pharmacol. 2018;9:846. doi:10.3389/fphar.2018.00846
8. Zhu L, Shu Z, Sun X. Bioinformatic analysis of four miRNAs relevant to metastasis-regulated processes in endometrial carcinoma. Cancer Manag Res. 2018;10:2337–2346.
9. Li Z, Zhou L, Lin C, et al. MiR-302b regulates cell functions and acts as a potential biomarker to predict recurrence in bladder cancer. Life Sci. 2018; 209: 15–23.
10. Li J, Qiu M, An Y, Huang J, Gong C. miR-7-5p acts as a tumor suppressor in bladder cancer by regulating the hedgehog pathway factor Gli3. Biochem Biophys Res Commun. 2018; 503(3):2101–2107.
11. Chen Z, Zhan Y, Chi J, et al. Using microRNAs as novel predictors of urologic cancer survival: an integrated analysis. EBioMedicine. 2018;34:94–107. doi:10.1016/j.ebiom.2018.07.014
12. Enokida H, Yoshino H, Matsushita R, Nakagawa M. The role of microRNAs in bladder cancer. Investig Clin Urol. 2016;57(Suppl 1):S60–S76. doi:10.4111/icu.2016.57.S1.S60
13. Liu X, Liu X, Wu Y, et al. MicroRNAs in biofluids are novel tools for bladder cancer screening. Oncotarget. 2017;8(19):32370–32379. doi:10.18632/oncotarget.16026
14. Mitash N, Tiwari S, Agnihotri S, Mandhani A. Bladder cancer: micro RNAs as biomolecules for prognostication and surveillance. Indian J Urol. 2017;33(2):127–133. doi:10.4103/0970-1591.203412
15. Higuchi F, Uchida S, Yamagata H, et al. Hippocampal MicroRNA-124 enhances chronic stress resilience in mice. J Neurosci. 2016;36(27):7253–7267. doi:10.1523/JNEUROSCI.0319-16.2016
16. Wang G, Liu H, Wei Z, Jia H, Liu Y, Liu J. Systematic analysis of the molecular mechanism of microRNA-124 in hepatoblastoma cells. Oncol Lett. 2017;14(6):7161–7170. doi:10.3892/ol.2017.7131
17. Gao FB. Context-dependent functions of specific microRNAs in neuronal development. Neural Dev. 2010;5:25. doi:10.1186/1749-8104-5-25
18. Zhang, T., Wang J, Zhai X, Li H, Li C, Chang J. MiR-124 retards bladder cancer growth by directly targeting CDK4. Acta Biochim Biophys Sin (Shanghai). 2014;46(12):1072–1079. doi:10.1093/abbs/gmu105
19. Wu J, Weng Y, He F, Liang D, Cai L. LncRNA MALAT-1 competitively regulates miR-124 to promote EMT and development of non-small-cell lung cancer. Anticancer Drugs. 2018;29(7):628–636. doi:10.1097/CAD.0000000000000626
20.
21. Zhang, Y.H., Wang QQ, Li H, Ye T, Gao F, Liu YC. miR-124 radiosensitizes human esophageal cancer cell TE-1 by targeting CDK4. Genet Mol Res. 2016;15(2).
22. Feng T, Xu D, Tu C, et al. MiR-124 inhibits cell proliferation in breast cancer through downregulation of CDK4. Tumour Biol. 2015;36(8):5987–5997. doi:10.1007/s13277-015-3275-8
23. Feng T, Shao F, Wu Q, et al. miR-124 downregulation leads to breast cancer progression via LncRNA-MALAT1 regulation and CDK4/E2F1 signal activation. Oncotarget. 2016;7(13):16205–16216. doi:10.18632/oncotarget.7578
24. Shimizu T, Suzuki H, Nojima M, et al. Methylation of a panel of microRNA genes is a novel biomarker for detection of bladder cancer. Eur Urol. 2013;63(6):1091–1100. doi:10.1016/j.eururo.2012.11.030
25. Hu J, Kong M, Ye Y, Hong S, Cheng L, Jiang L. Serum miR-206 and other muscle-specific microRNAs as non-invasive biomarkers for Duchenne muscular dystrophy. J Neurochem. 2014;129(5):877–883. doi:10.1111/jnc.12662
26. Knowles MA. Molecular pathogenesis of bladder cancer. Int J Clin Oncol. 2008;13(4):287–297. doi:10.1007/s10147-008-0812-0
27. Stein JP, Lieskovsky G, Cote R, et al. Radical cystectomy in the treatment of invasive bladder cancer: long-term results in 1,054 patients. J Clin Oncol. 2001;19(3):666–675. doi:10.1200/JCO.2001.19.3.666
28. Mayr R, Fritsche H-M, Pycha A, Pycha A. Radical cystectomy and the implications of comorbidity. Expert Rev Anticancer Ther. 2014;14(3):289–295. doi:10.1586/14737140.2014.868775
29. Jiao D, Li Z, Zhu M, Wang Y, Wu G, Han X. LncRNA MALAT1 promotes tumor growth and metastasis by targeting miR-124/foxq1 in bladder transitional cell carcinoma (BTCC). Am J Cancer Res. 2018;8(4):748–760.
30. Wan J, Liu H, Yang L, Ma L, Liu J, Ming L. JMJD6 promotes hepatocellular carcinoma carcinogenesis by targeting CDK4. Int J Cancer. 2019;144(10):2489-2500. doi:10.1002/ijc.31816
31. Kutwin P, Konecki T, Borkowska EM, Traczyk-Borszyńska M, Jabłonowski Z. Urine miRNA as a potential biomarker for bladder cancer detection - a meta-analysis. Cent European J Urol. 2018;71(2):177–185. doi:10.5173/ceju.2018.1605
32. Inamoto T, Uehara H, Akao Y, et al. A panel of MicroRNA signature as a tool for predicting survival of patients with urothelial carcinoma of the bladder. Dis Markers. 2018;2018:5468672. doi:10.1155/2018/5468672
33. Zhang L, Feng C, Zhou Y, Zhou Q. Dysregulated genes targeted by microRNAs and metabolic pathways in bladder cancer revealed by bioinformatics methods. Oncol Lett. 2018;15(6):9617–9624. doi:10.3892/ol.2018.8602
34. Guo L, Yin M, Wang Y. CREB1, a direct target of miR-122, promotes cell proliferation and invasion in bladder cancer. Oncol Lett. 2018;16(3):3842–3848. doi:10.3892/ol.2018.9118
35. Li Z, Lin C, Zhao L, et al. Oncogene miR-187-5p is associated with cellular proliferation, migration, invasion, apoptosis and an increased risk of recurrence in bladder cancer. Biomed Pharmacother. 2018;105:461–469. doi:10.1016/j.biopha.2018.05.122
36. Wang X, Wu Q, Xu B, et al. MiR-124 exerts tumor suppressive functions on the cell proliferation, motility and angiogenesis of bladder cancer by fine-tuning UHRF1. FEBS J. 2015;282(22):4376–4388. doi:10.1111/febs.13502
37. Xiong Y, Wang L, Li Y, Chen M, He W, Qi L. The long non-coding RNA XIST interacted with MiR-124 to modulate bladder cancer growth, invasion and migration by targeting Androgen Receptor (AR). Cell Physiol Biochem. 2017;43(1):405–418. doi:10.1159/000480419
38. Malumbres M. Cyclin-dependent kinases. Genome Biol. 2014;15(6):122. doi:10.1186/gb4184
39. Zhang M, Sun G, Shen A, Liu L, Ding J, Peng J. Patrinia scabiosaefolia inhibits the proliferation of colorectal cancer in vitro and in vivo via G1/S cell cycle arrest. Oncol Rep. 2015;33(2):856–860. doi:10.3892/or.2014.3663
40. Song, X., Liu X, Wang H, et al. Combined CDK4/6 and pan-mTOR inhibition is synergistic against intrahepatic cholangiocarcinoma. Clin Cancer Res. 2019;25(1):403-413. doi:10.1158/1078-0432. CCR-18-0284.
41. Yang Y, Zhang L-J, Bai X-G, Xu H-J, Jin Z-L, Ding M. Synergistic antitumour effects of triptolide plus gemcitabine in bladder cancer. Biomed Pharmacother. 2018;106:1307–1316. doi:10.1016/j.biopha.2018.07.083
42. Palumbo A, Lau G, Saraceni M. Abemaciclib: the newest CDK4/6 inhibitor for the treatment of breast cancer. Ann Pharmacother. 2019;53(2):178–185 doi:10.1177/1060028018795146
43. Warli SM, Kadar DD, Siregar GP. Ki-67 expression as a predictive factor of muscle invasion in bladder cancer. Open Access Maced J Med Sci. 2018;6(2):260–262. doi:10.3889/oamjms.2018.088
44. Wang S, Wu G, Han Y, et al. miR-124 regulates STAT3-mediated cell proliferation, migration and apoptosis in bladder cancer. Oncol Lett. 2018;16(5):5875–5881. doi:10.3892/ol.2018.9341
45. Zhou W, He L, Dai Y, Zhang Y, Wang J, Liu B. MicroRNA-124 inhibits cell proliferation, invasion and migration by targeting CAV1 in bladder cancer. Exp Ther Med. 2018;16(4):2811–2820. doi:10.3892/etm.2018.6537
46. Zo RB, Long Z. MiR-124-3p suppresses bladder cancer by targeting DNA methyltransferase 3B. J Cell Physiol. 2018;234(1):464–474. doi:10.1002/jcp.26591
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.