Back to Journals » Journal of Inflammation Research » Volume 18
The Function of the TGFβ Signaling Pathway in Connective Tissue Diseases: From Biology to Clinical Application
Authors Yuan S, Zhu W, Wang Y ![]() , Yu L
, Yu L
Received 10 May 2025
Accepted for publication 14 October 2025
Published 24 October 2025 Volume 2025:18 Pages 14679—14696
DOI https://doi.org/10.2147/JIR.S539571
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Ujjwol Risal
Simin Yuan,1 Wen Zhu,2 Yue Wang,1,2 Lili Yu1,3,4
1Faculty of Chinese Medicine, Macau University of Science and Technology, Macau, People’s Republic of China; 2Jiangsu Province Hospital of Chinese Medicine, Affiliated Hospital of Nanjing University of Chinese Medicine, Nanjing, Jiangsu, People’s Republic of China; 3State Key Laboratory of Mechanism and Quality of Chinese Medicine, Macau University of Science and Technology, Macau, People’s Republic of China; 4Macau University of Science and Technology Zhuhai MUST Science and Technology Research Institute, Zhuhai, Guangdong, People’s Republic of China
Correspondence: Yue Wang; Lili Yu, Email [email protected]; [email protected]
Abstract: Connective tissue diseases (CTDs) are characterized by a diverse array of symptoms, including persistent inflammation, immune system dysfunction, and fibrosis. The transforming growth factor-β (TGFβ) signaling pathway is crucial in fibrosis, dysregulated immune responses, and vascular injury associated with CTDs. TGFβ signaling facilitates the pathological progression of CTDs by modulating fibroblast activation, extracellular matrix accumulation, and immune cell activity. Based on a systematic search of PubMed and Web of Science, this narrative review reveals the dual role of TGFβ signaling in CTD pathogenesis and its therapeutic challenges. This review examines the mechanistic role of the TGFβ signaling pathway in various CTDs, such as systemic sclerosis (SSc), systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and Sjögren’s syndrome (SS). The review further examines the interplay between classical and non-classical pathways of the TGFβ signaling system and its implications in fibrosis and immunomodulation. The clinical applications of TGFβ as a potential therapeutic target are also discussed, with a special focus on treatment of ameliorating CTDs-associated fibrosis and immune abnormalities. These findings underscore the pivotal role of TGFβ signaling in fibrosis and immune regulation, highlighting opportunities for more precise and individualized therapeutic strategies.
Keywords: TGF-β signaling pathway, connective tissue disorders, fibrosis, immunomodulation, therapeutic targets
Introduction
Connective tissue diseases (CTDs) are a group of systemic disorders defined by immune-mediated chronic inflammation.1,2 Common CTDs include conditions such as systemic sclerosis (SSc), systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and Sjögren’s syndrome (SS).3,4 Common characteristics of CTDs include aberrant immune system activation, systemic inflammation, and varied levels of fibrosis, which frequently result in structural and functional impairment of several organs.5,6 While the precise pathophysiology of CTDs remains inadequately elucidated, accumulating data indicates that transforming growth factor-β (TGFβ) is pivotal in their pathogenic mechanisms, particularly in fibrosis and immunological dysregulation.7,8
TGFβ is a multifunctional cytokine essential for regulating cell proliferation, differentiation, migration, and immune responses, with a particularly notable role in tissue repair and immunosuppression.9,10 TGFβ exhibits a dual, context-dependent role in pathophysiology: it is essential for maintaining tissue homeostasis and immune tolerance under physiological conditions, yet its dysregulation drives pathological fibrosis, chronic inflammation, and immune evasion.11 This duality presents a significant therapeutic challenge, as selectively targeting its pathogenic signaling without impairing homeostatic functions remains difficult.12 In CTDs, the overactivation or dysregulation of TGFβ is frequently linked to the onset of fibrosis.12–14 In conditions such as SSc, SS, and RA, abnormal activation of the TGFβ signaling pathway can result in excessive fibroblast activation, resulting in collagen deposition and tissue sclerosis that leads to organ failure.15,16 TGFβ is a crucial regulator of fibrosis formation and a significant contributor to immunological tolerance and immune evasion, hence worsening the clinical symptoms and progression of CTDs.8,17
Due to the significant role of TGFβ in CTDs, interventions targeting this signaling pathway have emerged as a focal point of therapeutic research in recent years. TGFβ inhibitors can effectively impede fibrosis progression, mitigate abnormal immune responses, and enhance organ function.18,19 Strategies such as small molecule TGFβ inhibitors, anti-TGFβ antibodies, and gene therapies have shown potential therapeutic promise in preclinical and clinical studies and offer new targeted intervention options for clinical treatment.20 Nonetheless, the clinical application of TGFβ inhibitors encounters numerous challenges, including concerns over therapeutic specificity, sustained efficacy, and management of adverse effects. Here, we review the mechanism of action, status of clinical application, and future research directions of TGFβ inhibitors in CTDs. Specifically, we evaluate the pathogenic role of TGFβ in various CTDs, analyze the research progress and clinical outcomes of current TGFβ-targeted therapies, and explore potential avenues for therapeutic development. This review aims to provide a theoretical foundation for the optimizing and advancing TGFβ-related treatments in clinical practice while guiding the development of novel pharmaceuticals and personalized therapies.
To provide a thorough overview, we conducted a comprehensive literature search of PubMed and Web of Science using keywords including “TGFβ”, “fibrosis”, and “connective tissue disorders”, with an emphasis on studies that elucidate the underlying pathogenic mechanisms and inform therapeutic approaches.
The Molecular Mechanisms by Which TGFβ Regulates Fibrosis Progression in CTDs
The TGFβ-mediated fibrotic process is the core of the pathology of CTDs.15 This process involves fibroblast activation, extracellular matrix (ECM) remodeling, and modulation of associated molecular pathways.21 Recent studies using single-cell sequencing techniques have elucidated the subpopulation heterogeneity of fibroblasts and their pivotal roles in fibrosis, offering a novel viewpoint on the mechanisms underlying TGFβ-driven fibrosis.
TGFβ Promotes Fibroblast Activation
Fibroblasts are essential effector cells in the fibrotic process.22 Stimulated by TGFβ, fibroblasts differentiate into myofibroblasts, which possess contractile properties and enhanced capability of ECM synthesis.23 Single-cell sequencing revealed that fibroblasts are not a uniform population but comprise several functionally different subpopulations, with certain subpopulations (eg, myofibroblast precursors) exhibiting heightened sensitivity to TGFβ.24 In SSc, TGFβ promotes the differentiation of fibroblast subpopulations into myofibroblasts by activating the Smad2/3 and MAPK pathways.25 These activated fibroblasts exhibit high levels of α-smooth muscle actin (α-SMA) and secrete a large number of ECM components (eg, collagen type I COL1A1, collagen type III COL3A1, and fibronectin FN1), contributing to the fibrotic progression.26 TGFβ also promotes the stability of the fibrotic phenotype in fibroblasts by modulating their epigenetic changes, including DNA methylation and histone acetylation.27 In idiopathic pulmonary fibrosis (IPF), TGFβ prompted the overexpression of DNA methyltransferase (DNMT), which inhibited the expression of fibrosis-suppressor genes (eg, PPARγ), perpetuating the activation of fibroblasts.28
TGFβ Is Involved in ECM Remodeling and Collagen Cross-Linking
ECM remodeling, the hallmark feature of fibrosis, involves the over-synthesis of ECM components and their structural and functional alterations.29 TGFβ facilitates the excessive accumulation of ECM by upregulating the expression of ECM synthesis genes (eg, COL1A1, COL3A1, and FN1).30 Meanwhile, TGFβ regulates the balance of ECM-degrading enzymes [eg, matrix metalloproteinases (MMPs)] and their inhibitors (eg, TIMPs) to inhibit ECM degradation and further exacerbate the process of fibrosis. Collagen cross-linking, a key process in ECM remodeling, is mediated by lysyl oxidase (LOX) and its cognate enzymes (LOXL1-4).31 TGFβ promotes cross-linking of collagen fibrils by up-regulating the expression of the LOX/LOXL enzymes, which enhances the mechanical strength and degradation resistance of ECM.32,33 In SSc and IPF, LOX/LOXL expression is significantly elevated, leading to excessive cross-linking of collagen fibers and hardening of tissues.34 For example, in skin tissues of SSc patients, LOXL2 expression levels were positively correlated with skin thickness and disease severity.35 In IPF, LOXL2-mediated collagen cross-linking not only enhanced the mechanical strength of lung tissue but also further promoted fibroblast activation and ECM deposition, which in turn exacerbated the process of pulmonary fibrosis.36
TGFβ Initiates Fibrosis Signaling Pathway Cascade Responses
The TGFβ-driven fibrotic process relies on the synergistic action of the classical Smad and non-Smad pathways.8 In the classical Smad pathway, after TGFβ binds to the receptor TGFβR1/2, Smad2/3 are phosphorylated and form a complex with Smad4, which enters the nucleus to regulate the expression of pro-fibrotic genes (eg, COL1A1, α-SMA).11,37 This pathway is particularly critical in diseases such as SSc and IPF.38 Among these regulators, overactivation of Smad3 promotes fibroblast-to-myofibroblast transformation and exacerbates ECM deposition.39 Smad7 acts as a negative feedback regulator, limiting signaling overactivation through inhibition of Smad2/3 or degradation of the TGFβ receptor, but its expression is often suppressed in CTDs, leading to increased fibrosis.40
Non-Smad pathways, including MAPK, PI3K/Akt, also play important roles in TGFβ signaling.41 For example, TGFβ in RA promotes synovial fibroblast proliferation and inflammatory factor release through the TAK1-JNK axis.42–44 In SLE, the PI3K/Akt pathway regulates B cell differentiation and autoantibody production.45,46 ERK1/2 activation in IPF promotes lung fibroblast proliferation and ECM deposition.47–49 In addition, TGFβ signaling interacts with other pathways (eg, Wnt/β-catenin, Notch) to further exacerbate fibrosis. For example, the Wnt/β-catenin pathway in SSc enhances Smad3 transcriptional activity and induces COL1A1 and α-SMA expression.50 The Notch pathway in IPF promotes fibronectin expression.51 In summary, the pathological role of TGFβ signaling in CTDs is dependent on an interactive network of multiple pathways and exhibits disease-specific regulatory mechanisms, leading to heterogeneity of fibrosis and immune responses. The role of TGFβ signaling pathway in connective tissue diseases can be seen in Figure 1.
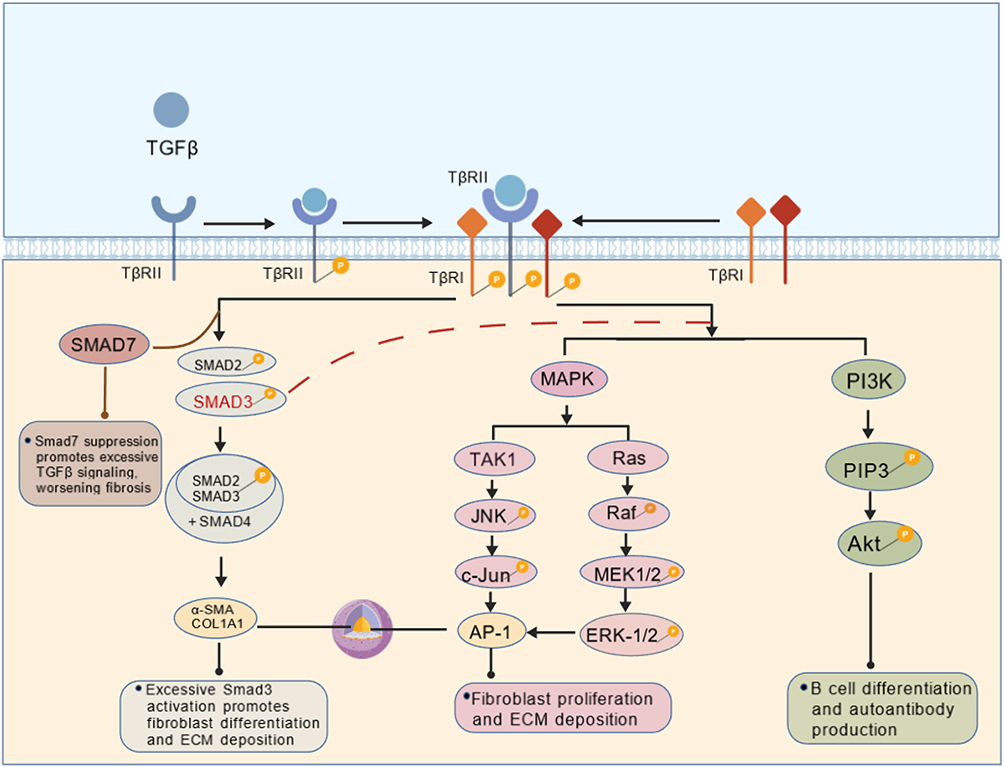 |
Figure 1 Schematic representation of the TGF-β-associated pro-fibrotic signaling pathway in connective tissue diseases.52–54 |
TGFβ Modulates Cell–Cell Interactions for Fibrotic Processes
TGFβ-driven fibrosis also involves multiple cell-to-cell interactions. For example, in SSc, TGFβ promotes fibroblast activation and ECM deposition by inducing endothelial to mesenchymal stromal cell (EndMT) transformation.55 In IPF, TGFβ promotes the progression of pulmonary fibrosis by regulating the interaction between alveolar epithelial cells and fibroblasts.56 In addition, TGFβ is involved in the immunoregulation of fibrosis by regulating the function of immune cells, such as macrophages and T cells. In SSc, TGFβ further promotes fibroblast activation and ECM deposition by inducing macrophage polarization towards M2 type.57
The Crosstalk of TGFβ with the Immune Microenvironment in CTDs
The interaction of the TGFβ signaling pathway with the immune microenvironment plays an important role in the pathological process of CTDs.14,58,59 TGFβ is not only a core driver of fibrosis but also plays a key role in the differentiation, activation, and functional regulation of immune cells.60 By regulating the function of immune cells, such as T cells, macrophages and B cells, TGFβ forms a complex immune microenvironment in CTDs, which in turn affects disease progression and prognosis (Figure 2).
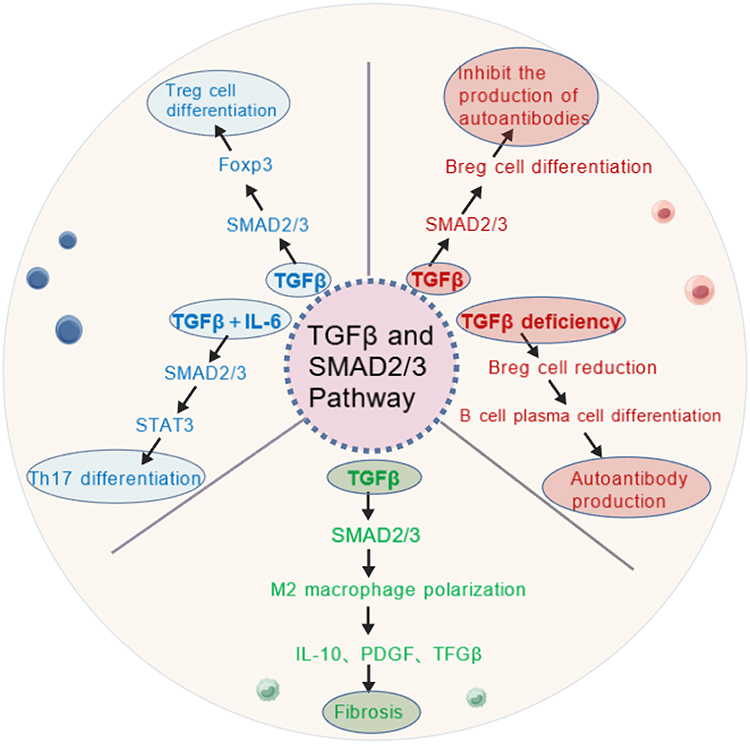 |
Figure 2 A concrete overview of the regulatory mechanisms by which TGFβ modulates immune cells.61–63 |
TGFβ Regulates Th17/Treg Balance
The dual role of TGFβ in T cell differentiation makes it a central molecule in immune regulation.61,64 In normal physiological condition, TGFβ promotes the differentiation of regulatory T cells (Treg), maintains immune tolerance and suppresses autoimmune responses.65 However, in CTDs, dysfunctional TGFβ signaling leads to reduced Treg cell numbers and impaired function, which promotes hyperactivation of helper T cell 17 (Th17) and enhances autoimmune responses.66 In SLE, dysfunction of TGFβ signaling is thought to be the main cause of dysregulation of the Treg/Th17 balance.67 TGFβ induces Foxp3 expression through activation of the Smad2/3 pathway and promotes Treg cell differentiation.68 However, in SLE patients, downregulation of TGFβ signaling or Smad7 overexpression leads to impaired Treg cell function, which further induces overactivation of Th17 cells.69,70 TGFβ synergistically interacts with IL-6 to promote the differentiation of Th17 cells and induces the secretion of proinflammatory factors such as IL-17A and IL-21, thus exacerbating the autoimmune response in SLE. Clinical studies have shown that the proportion of Treg cells in the peripheral blood of SLE patients is lower and the proportion of Th17 cells is higher, and that this trend is positively correlated with disease activity.66 In RA, TGFβ is involved in synovial inflammation and joint destruction by regulating the Th17/Treg balance. TGFβ synergizes with IL-6 to promote Th17 cell differentiation and induces the secretion of IL-17A and IL-22, which further promote synovial fibroblast activation and the progression of joint inflammation.71 In addition, dysfunctional TGFβ signaling leads to impaired immunosuppression of Treg cells, exacerbating the pathological process of RA.
TGFβ in Macrophage Regulation: The Pro-Fibrotic Role of M2-Type Polarization
Macrophages are important immunomodulatory cells during fibrosis, and their phenotypic polarization (M1-type versus M2-type) plays a key role in CTDs.62,72 TGFβ participates in the progression of fibrosis by promoting macrophage polarization to M2-type.73 M2-type macrophages secrete pro-fibrotic factors, such as TGFβ, IL-10, and PDGF, which further promote fibroblast activation and ECM deposition.74 In SSc, TGFβ-induced M2-type macrophage polarization is an important driver of fibrosis.57 TGFβ promotes macrophage polarization toward the M2 type through activation of the Smad2/3 and PI3K/Akt pathways and induces secretion of TGFβ, IL-10 and PDGF.75 These pro-fibrotic factors stimulate fibroblast activation and ECM deposition, driving fibrosis in the skin and visceral organs. Clinical studies have shown that the proportion of M2-type macrophages is significantly higher in peripheral blood and diseased tissues of SSc patients, which is positively correlated with increased skin thickness and disease severity.76 In IPF, TGFβ-induced polarization of M2-type macrophages also plays a key role in pulmonary fibrosis.77 M2-type macrophages exacerbate pulmonary fibrosis by promoting activation of lung fibroblasts and ECM deposition through the secretion of TGFβ and PDGF. M2-type macrophages also inhibit the anti-fibrotic immune response through the secretion of IL-10 and TGFβ, which further contributes to fibrotic process.62,78
TGFβ Regulates B-Cells: Autoantibody Production and Pathology
B cells play a critical role in the autoimmune response in CTDs, and TGFβ participates in the pathological process of the disease by regulating B cell differentiation and function.79,80 TGFβ promotes the differentiation of B cells to regulatory B cells (Breg), which suppresses the autoimmune response.63 However, in CTDs, defective TGFβ signaling function leads to a reduction and impaired function of Breg cells, which in turn promotes the overproduction of autoantibodies.81 In SSc, TGFβ is involved in disease progression by promoting the production of anti-Scl-70 antibodies by B cells. Anti-Scl-70 antibodies are specific autoantibodies in SSc, and their production is closely related to the activation of TGFβ signaling.82 TGFβ promotes the differentiation of B cells to plasma cells through activation of the Smad2/3 pathway and induces the production of anti-Scl-70 antibodies.63 After the formation of immune complexes, it activates the complement system and triggers tissue damage, which in turn exacerbates the fibrotic process of SSc. Clinical studies have shown that patients with anti-Scl-70 antibody-positive SSc usually exhibit more severe skin sclerosis and visceral organ involvement.55 In SLE, downregulation of TGFβ signaling or Smad7 overexpression leads to dysregulation of Breg cell function and promotes autoantibody production, which in turn activates the complement system and exacerbates the pathological process of SLE.83
Advances in Drug Research Targeting TGFβ
The role of the TGFβ signaling pathway in fibrosis and its molecular mechanisms have been analyzed previously. With the deepening understanding of the role of TGFβ in CTDs, therapeutic strategies targeting this pathway have become the focus of research. Advances in pharmacology and molecular biology have propelled a variety of targeted drugs and biologics into preclinical studies and clinical trials, demonstrating the potential for treating CTDs. Next, the current major TGFβ inhibitors in the treatment of CTDs, including small molecule kinase inhibitors, biologics, nucleic acid drugs, gene editing technologies, and natural products, are discussed to assess their clinical translational progress and challenges (Table 1).
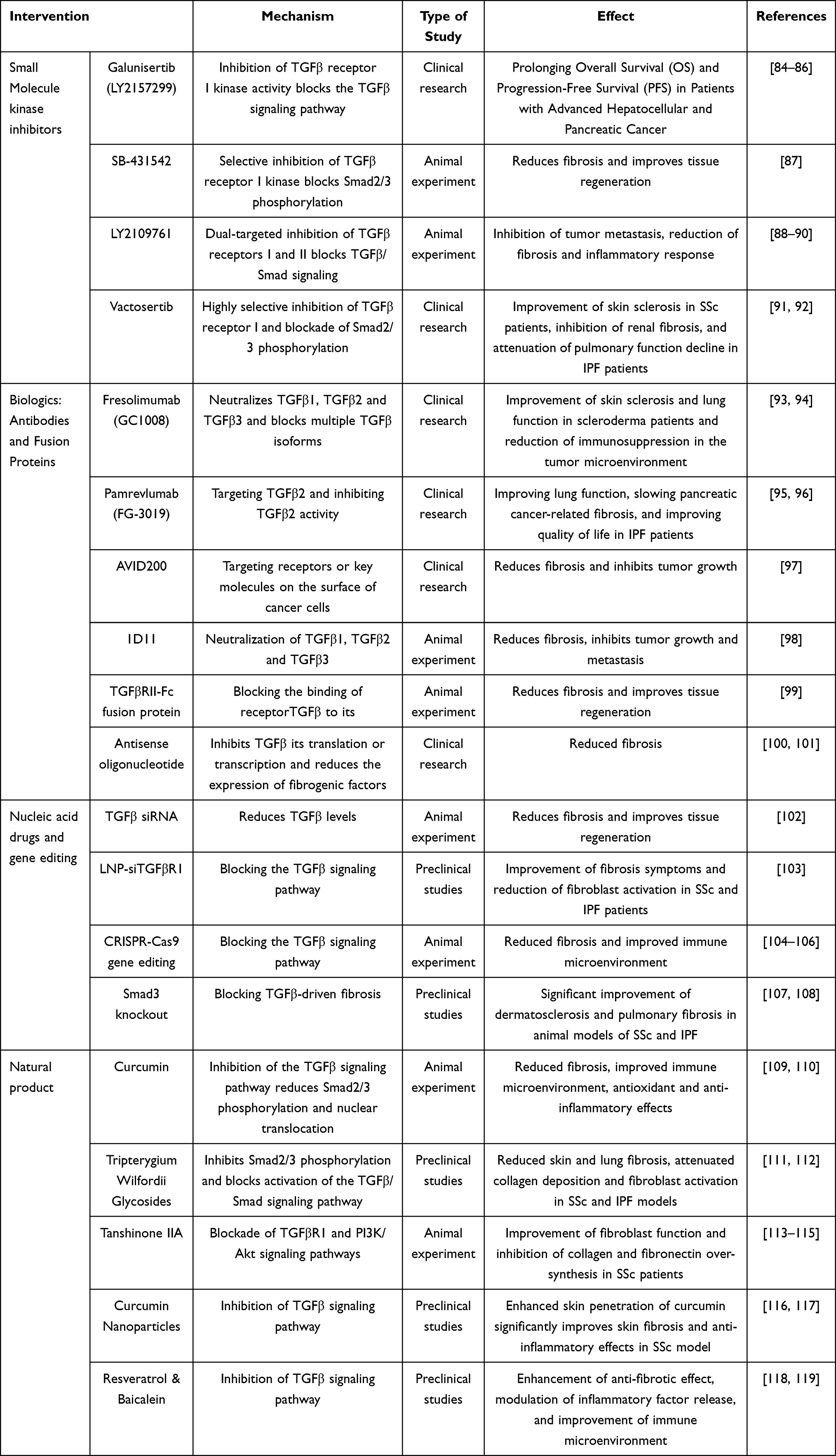 |
Table 1 Summary of Research Advances in Drugs and Therapies Targeting the TGFβ Signaling Pathway |
Small Molecule Kinase Inhibitors
Small molecule kinase inhibitors show significant potential in the treatment of CTDs by targeting the ATP-binding site of Transforming Growth Factor-β Type 1 Receptor (TβR1) and blocking the activation of Smad2/3 signaling pathway.120,121 Galunisertib and Vactosertib, as representative agents, are administered orally and exert their effects through highly selective mechanisms, specifically inhibiting TGF-β1 and TGF-β3-mediated signaling pathways. This targeted inhibition effectively downregulates the expression of pro-fibrotic genes such as COL1A1 and α-SMA.122,123 Preclinical studies have shown that both exhibit significant antifibrotic activity in a variety of fibrosis models.124,125 In clinical trials in the field of CTDs, Galunisertib showed preliminary improvement in skin sclerosis and lung function in SSc and IPF.84,123 Vactosertib, on the other hand, has shown a favorable safety profile and potential to improve fibrosis and disease activity in SSc and IPF.91,126 Both are currently undergoing in-depth validation in clinical trials in SSc and IPF as important directions for the precision treatment of CTDs. Future studies are needed to further evaluate their long-term efficacy and safety to promote their clinical translational applications.
Biological Agents: Antibodies and Fusion Proteins
With the in-depth study of the TGFβ signaling pathway, biologics targeting TGFβ have gradually become a research hotspot, including monoclonal antibodies and receptor fusion proteins, which inhibit the over-activation of the signaling pathway by specifically neutralizing TGFβ or its receptor. For monoclonal antibodies, Fresolimumab (GC1008) is a pan-TGFβ-neutralizing antibody that inhibits the activity of TGFβ1, TGFβ2 and TGFβ3 simultaneously.93 Early clinical trials showed its potential, but its broad inhibition of multiple TGFβ isoforms may trigger side effects such as immunosuppression and tumorigenesis, limiting its application. Pamrevlumab (FG-3019), a monoclonal antibody specifically targeting TGFβ2, has been shown to have a favorable safety profile and anti-fibrosis effects in clinical trials of IPF and pancreatic cancer-related fibrosis, especially in IPF, where it significantly improved lung function and quality of life.93 For receptor fusion proteins, SRK-181 is a TGFβ1-specific trap protein that blocks TGFβ1’s interaction with the receptor.127 Preclinical studies have shown that SRK-181 has a significant antifibrotic effect in a variety of fibrosis models and has little effect on TGFβ2 and TGFβ3 activity, reducing the risk of potential side effects. SRK-181 is now in clinical trials and preliminary results showed its potential to improve fibrosis.127,128
Nucleic Acid Drugs and Gene Editing
The rapid development of nucleic acid drugs and gene editing technologies has provided new therapeutic strategies for targeting the TGFβ signaling pathway. These technologies achieve the regulation of the TGFβ signaling pathway by directly interfering with gene expression or precisely editing the genome, opening up new directions for research in this field.
siRNA
Small interfering RNA (siRNA) technology inhibits the overactivation of the TGFβ signaling pathway by specifically silencing the expression of target genes through the RNA interference (RNAi) mechanism. In therapies targeting the TGFβ signaling pathway, siRNAs usually target TGFβ receptor 1 (TGFβR1) or other key molecules (eg, Smad2/3) to inhibit aberrant signaling.102,129 In recent years, the development of nanoparticle delivery systems (eg, lipid nanoparticles, LNP) has significantly improved the stability and targeting of siRNAs.130 For example, LNP-siTGFβR1 is a lipid nanoparticle-based siRNA delivery system that efficiently delivers siRNA to target cells and specifically silences TGFβR1 expression. In preclinical studies, LNP-siTGFβR1 showed significant anti-fibrotic effects with little side effect on normal tissues. In addition, this technique inhibited fibroblast activation and excessive deposition of ECM, providing new ideas for the treatment of fibrosis-related diseases.131
CRISPR/Cas9 Gene
CRISPR/Cas9 gene editing technology has become a powerful tool for targeting the TGFβ signaling pathway with its efficient and precise genome editing capabilities. The technology achieves gene knockout, insertion or modification by specifically cutting the target gene sequence. In the TGFβ signaling pathway, CRISPR/Cas9 can be used to knock down key molecules such as Smad3 to inhibit the over-activated signaling pathway.104 For example, preclinical studies have shown that fibroblast-specific knockdown of Smad3 significantly inhibited TGFβ-driven fibrotic progression without significant side effects. In addition, CRISPR/Cas9 can be used to edit other key molecules in the TGFβ signaling pathway, such as TGFβR1 or Smad2.105 Fibroblast-specific knockdown of TGFβR1 or Smad2 significantly ameliorated dermatosclerosis and pulmonary fibrosis in preclinical models of SSc and IPF, further validating its potential in the treatment of fibrosis in CTDs.132,133
Natural Products
Natural products and traditional Chinese medicine (TCM) show unique potential in regulating the TGFβ signaling pathway, and their multi-target and multi-pathway mechanisms of action provide new ideas for the treatment of CTDs. In recent years, based on the integrated research of modern pharmacology and nanotechnology, the role of active ingredients of TCM and their complexes in antifibrosis and immunomodulation has been gradually verified scientifically.
Chinese Medicine Monomers
Tripterygium Wilfordii Glycosides is a diterpenoid extracted from Tripterygium wilfordii.111,134 Studies have shown that tretinoin blocks the activation of the TGFβ/Smad signaling pathway by inhibiting the phosphorylation and nuclear translocation of Smad2/3.135 In addition, tretinoin significantly reduced the expression of collagen (COL1A1) and α-SMA, also inhibited the NF-κB signaling pathway to reduce the release of inflammatory factors, demonstrating a dual effect of antifibrotic and anti-inflammatory.136,137
Tanshinone IIA is the main active ingredient extracted from Salvia miltiorrhiza.113 Studies have shown that Tanshinone IIA blocks the over-activation of TGFβ signaling by simultaneously inhibiting the TGFβR1 and PI3K/Akt signaling pathways.138 Its ability to inhibit TGFβR1 phosphorylation and Smad2/3 activation reduces fibroblast to myofibroblast transformation and decreases ECM synthesis.139 In primary fibroblasts from SSc patients, tanshinone IIA significantly reduced COL1A1 and fibronectin expression. These research advances provide an important scientific basis for the role of natural products in regulating the TGFβ signaling pathway and anti-fibrosis.
Compound Chinese Medicines
Classical compound Chinese medicines, such as Xu Fu Zhu Yu Tang, have shown potential in regulating the TGFβ signaling pathway. Studies have shown that Xu Fu Zhu Yu Tang can reduce serum TGFβ1 levels and alleviate fibrosis by inhibiting the TGFβ/Smad signaling pathway and down-regulating LOX/LOXL activity. The advantage of compound Chinese medicine lies in the synergistic effect of its multiple components, which can regulate multiple targets simultaneously, but its complexity and diversity still need to be elucidated by further systematic pharmacological studies.140
Integration of Natural Products and Nanotechnology
Nanotechnology provides new ways to address the low bioavailability and non-specific distribution of natural products. Encapsulation of natural products using nanocarriers (eg, liposomes, polymer nanoparticles) can significantly improve their stability and targeting.141 For example, the skin penetration ability and antifibrotic effect of curcumin were significantly enhanced by nanocarrier loading, and the mechanism involved the inhibition of TGFβR1 and the modulation of ROS signaling pathway.142,143 In addition, natural products such as resveratrol and baicalein have also been enhanced in bioavailability by nanotechnology, showing potential in antifibrosis and anti-inflammation.118 These advances provide new technical support for the clinical application of natural products.
TGF-β Inhibitors in Connective Tissue Diseases
Systemic Sclerosis (SSc)
SSc is a severe connective tissue disease characterized by fibrosis of the skin and internal organs, vasculopathy, and immune dysregulation.82,144 Hyperactivation of the TGFβ signaling pathway is the central driving mechanism of its fibrosis, which is manifested by abnormal fibroblast activation, endothelial cell damage, and immune system dysregulation.145,146 In recent years, therapeutic strategies targeting TGFβ have made significant progress in the management fibrosis and vascular complications. Pamrevlumab (FG-3019), a monoclonal antibody targeting TGFβ2, attenuates dermal fibrosis in SSc mouse models. It was shown that FG-3019 significantly reduced fibrosis induced by angiotensin II (Ang II), with effects similar to CTGF knockdown in fibroblasts. This suggests that FG-3019, as a potential treatment for SSc, may act by inhibiting collagen deposition and myofibroblast accumulation, which are key features of fibrosis.147 Compared with pan-TGFβ inhibitors (eg, Fresolimumab), Pamrevlumab offers safety for long-term use by reducing systemic side effects such as severe infections or thrombocytopenia due to subtype selectivity.
Combination therapies have received much attention as a potential strategy for the treatment of SSc-associated interstitial lung disease (SSc-ILD), especially when used in combination with conventional treatments such as morphenicol (MMF). For example, a Phase II clinical trial (NCT03221257) is investigating the combination of MMF with the antifibrotic drug pirfenidone, showing potential for SSc-ILD.148 The challenge with this type of combination therapy is patient intolerance, especially gastrointestinal side effects. In the management of vascular complications, the TGF-β signaling pathway plays a key role in SSc vascular development and homeostasis maintenance. Studies have shown that BMPR-II mutations may lead to aberrant BMP/Smad signaling and enhance TGF-β/ALK5/Smad2/3 signaling, which in turn triggers vascular smooth muscle cell proliferation and vascular remodeling, leading to complications such as pulmonary hypertension. The vascular wall of patients with SSc has abnormal compositions, and by regulating these compositions, TGF-β contributes to vasculopathy. Preclinical studies have shown that TGF-β antibodies and gene editing techniques attenuate pulmonary vasculopathy, demonstrating the important role of TGF-β signaling in vasculopathy. However, the results of clinical studies have been inconsistent, with some studies failing to show significant clinical benefit, which may be related to inadequate drug dosage or other factors.149 Nonetheless, therapies targeting the TGF-β signaling pathway have potential and need to be further explored in the future in SSc-associated vascular complications.
Systemic Lupus Erythematosus (SLE)
The TGFβ signaling pathway has a dual role in SLE: in physiological state, it maintains immune tolerance by inducing Treg differentiation; in pathological state, its dysregulation leads to defective Treg function and auto-reactive B cell activation, which exacerbates immune imbalance and organ damage. Inhibitors targeting TGFβ show potential in SLE therapy.150,151
In terms of immunomodulation, the pathological effects of TGFβ are mainly characterized by defective Treg function and abnormal B cell activation.152 A phase II clinical trial (NCT04161855) of the pan-TGFβ neutralizing antibody Fresolimumab showed that treatment of patients with moderate-to-severe SLE resulted in a significant reduction in serum anti-dsDNA antibody titers (42% vs 18%, p=0.03) and was associated with an improvement in the disease activity index (SLEDAI). The study demonstrated that Fresolimumab rebuilds immune tolerance by increasing the proportion of functional Treg and reducing Th17 cell infiltration, but its clinical use requires attention to thrombocytopenia and infection risk, suggesting the need for optimization of dosage and patient selection strategies.153
In terms of renoprotection, TGFβ drives the fibrotic process in lupus nephritis (LN) by promoting renal fibroblast activation and podocyte injury.150,154,155 Preclinical studies have shown that the Smad3 inhibitor SIS3 significantly reduces glomerular thylakoid matrix expansion and collagen deposition and decreases urinary protein levels by 60% in an MRL/lpr mouse model.156 In addition, an antisense oligonucleotide sequence targeting TGFβ1 (ISIS 369645) significantly inhibited TGFβ1 expression and reduced tubulointerstitial fibrosis scores in renal biopsy tissues from patients with LN (p<0.01).157 A phase II trial (NCT05292547) evaluated the efficacy of small molecule inhibitor of TGFβR1, LY3200882, in LN treatment, with preliminary data showing that combination therapy reduced the urinary protein/creatinine ratio (UPCR) by up to 35% and improved the renal pathologic activity index.
Rheumatoid Arthritis (RA)
RA is a chronic autoimmune disease characterized by synovitis, bone erosion, and systemic complications, and its pathogenesis involves dysregulation of the immune system, synovial hyperplasia, and bone destruction.158–160 The TGFβ signaling pathway plays a dual role in RA: it is involved in tissue repair in physiological states, and it exacerbates the disease progression by promoting synovial hyperplasia, bone destruction, and systemic fibrosis in pathological states.161,162 In recent years, TGFβ inhibitors have demonstrated potential in controlling local joint damage with systemic complications, and have become an important research direction for RA treatment.
In synovial-targeted therapy, TGFβ drives synovial invasion and bone destruction by activating fibroblast-like synoviocytes (FLS) and osteoclasts.163 Preclinical studies (mouse model of collagen-induced arthritis) demonstrated that Galunisertib (TGFβR1 inhibitor, 5 mg/kg, twice weekly, intra-articular injections) significantly inhibited synovial hyperplasia and bone erosion, with a 62% reduction in the volume of bone erosion compared to the control group (p<0.001), as well as down-regulating the expression of COL1A1 and MMP-3 in synovial tissue. Mechanistic studies showed that Galunisertib inhibited FLS migration and invasion by blocking the TGFβ/Smad2/3 pathway and decreased the RANKL/OPG ratio to inhibit osteoclast differentiation.123 Preliminary data from the Phase I clinical trial (NCT04879810) showed that after 4 weeks of Galunisertib treatment, the joint swelling index (SJC) was reduced by 40%, suggesting a significant potential for its topical application.
In terms of systemic effects, TGFβ plays a central role in RA-associated extra-articular fibrotic complications such as ILD and cardiovascular fibrosis. Serum TGFβ1 levels were positively correlated with HRCT fibrosis scores in patients with RA-ILD (r=0.71, p<0.01).164 Preclinical studies have shown that oral administration of Vactosertib (TGFβR1 inhibitor) inhibits lung fibroblast activation and reduces lung collagen deposition (55% decrease).165 In addition, TGFβ promotes myocardial fibrosis through activation of cardiac fibroblasts, and left ventricular end-diastolic volume (LVEDV) improves more dramatically in RA patients treated with TGFβ inhibitors, suggesting its potential efficacy in cardiovascular fibrosis.166
Sjögren’s Syndrome (SS): TGFβ-Mediated Salivary Gland Fibrosis
SS is an autoimmune disease characterized by salivary gland lymphocyte infiltration and fibrosis. Abnormal activation of the TGFβ signaling pathway is its core pathological mechanism.167 Studies have shown that TGFβ1 expression is significantly upregulated in the salivary glands of SS patients, inducing fibroblast-to-myofibroblast transformation through activation of the Smad2/3 pathway and promoting collagen and α-SMA overexpression.168 Single-cell sequencing revealed the presence of a subpopulation of pro-fibrotic fibroblasts (marker genes: POSTN, CTHRC1) significantly enriched in genes related to the TGFβ pathway (eg, SMAD3, TGFBR1). In addition, TGFβ synergistically promotes the vicious cycle of inflammation and fibrosis by inducing epithelial–mesenchymal transition (downregulation of E-cadherin, upregulation of Vimentin) and modulating Th17/Treg imbalance (2.5-fold increase in Th17 cell infiltration), and M2-type macrophage polarization (40% elevated percentage of CD206+ cells).169 Preclinical studies have shown that blockade of TGFβR1 reverses the fibrotic process and restores salivary secretory function (50% increase in salivary flow rate in a mouse model).170
In terms of targeted therapies, the Smad3 inhibitor SIS3 significantly reduced salivary gland fibrosis and lymphocyte infiltration in a mouse model, while a TGFβ1-neutralizing antibody (1D11) significantly inhibited collagen expression and restored salivary flow rates to 80% of normal levels.171 In clinical trials, topical injections of Galunisertib (phase I trial, NCT04968938) resulted in a 35% increase in salivary production and a 25% reduction in fibrosis scores. A phase II trial (NCT05331053) of AVID200 (TGFβ1/3 trap protein) is exploring the efficacy of its systemic administration on salivary gland function and extra-glandular complications. These studies provide an important basis for targeted therapy for pSS.
Interstitial Lung Disease (ILD)
ILD is a common complication of several CTDs including SSc and RA.172,173 Despite their different primary etiologies, fibroblast activation and ECM remodeling driven by the TGFβ signaling pathway are common core mechanisms of ILD progression.174 In recent years, inhaled therapies targeting TGFβ have emerged as an emerging direction in the treatment of ILD due to their local delivery advantages and potential to reduce systemic toxicity.175
In terms of molecular mechanisms, SSc-ILD and RA-associated interstitial lung disease (RA-ILD) showed significant commonality in fibroblast activation and ECM deposition.176 TGFβ released from alveolar epithelial cell injury induces the transformation of lung fibroblasts into myofibroblasts and secretion of large amounts of collagen (eg, COL1A1, COL3A1) and fibronectin.177 Single-cell sequencing studies demonstrated the presence of specific pro-fibrotic fibroblast subpopulations (eg, THY1+, PDGFRα+) with highly overlapping transcriptional profiles (eg, high expression of LOXL2, POSTN) in the two types of ILD, confirming that TGFβ-driven activation of fibroblasts is a key component of ILD progression. In addition, TGFβ promotes lung fibrosis by regulating macrophage polarization (M2-type) and T-cell differentiation (Th17/Treg imbalance).178
In terms of therapeutic strategies, the TGFβR1 inhibitor dry powder inhaler (PRM-151) offers a new direction for ILD treatment.179 Localized inhalation delivery increases intrapulmonary concentrations of the drug while reducing systemic exposure and systemic side effects.180 Preliminary data from a phase I clinical trial of PRM-151 (NCT05541879) showed that a single inhalation dose (10–100 μg) was well tolerated, with an incidence of mild cough and throat irritation of <15%. Bronchoalveolar lavage fluid (BALF) analysis showed a 50% reduction in TGFβ activity in the treatment group compared to baseline (p=0.01) and a 35% downregulation of COL1A1 mRNA expression. Lung carbon monoxide diffusion (DLCO) increased by an average of 8% in patients in the treatment group after 4 weeks (control group decreased by 2%), suggesting its potential to improve lung function. However, inhaled therapy still faces challenges such as uneven drug distribution and long-term safety, and further optimization is needed.181
Future Challenges and Directions
TGFβ inhibitors have demonstrated potential in the treatment of CTDs, but many challenges remain. Notably, clinical development of pan-TGFβ inhibitors like fresolimumab has yielded mixed results, likely due to the dual homeostatic and pathogenic roles of TGFβ signaling across different tissues and disease contexts. TGFβ has multiple physiological functions, and its prolonged inhibition may interfere with tissue repair and immune homeostasis, increasing the risk of side effects. Therefore, the development of more selective inhibitors such as isoform-specific antibodies or receptor-specific approaches and organ-targeted therapeutic strategies is crucial. For example, in SLE, renal-specific delivery systems (eg, hyaluronic acid-modified liposomes) and combination therapies (eg, with belimumab or JAK inhibitors) reduce systemic side effects and enhance efficacy. In SSc and RA, the combination of TGFβ inhibitors with antifibrotic agents (eg, nidanib) or anti-inflammatory agents (eg, JAK inhibitors) is expected to synergistically inhibit fibrosis and inflammation and optimize therapeutic efficacy. In SS, topical administration (eg, ultrasound-guided nanoparticle delivery) and hydrogel carriers (eg, hyaluronic acid-TGFβ inhibitor couplings) can enhance drug distribution and retention time, subject to monitoring of glandular atrophy and risk of infection, and the combination of B-cell-targeting drugs (eg, rituximab) or JAK inhibitors can further suppress immune infiltration and fibrosis.
This review systematically explores the central mechanisms of the TGFβ signaling pathway in fibrosis and immune dysregulation across various connective tissue diseases while evaluating the potential and limitations of different therapeutic strategies targeting this pathway. Future research should focus on the development of precision stratified therapies, combination strategies and novel delivery systems. Individualized interventions can be achieved by screening high response patient groups (eg, serum TGFβ1, KL-6, or COL3A1 levels in BALF). Meanwhile, nanoparticle- or liposome-encapsulated TGFβ inhibitors can improve targeting and retention time to optimize efficacy. Despite the potential of small molecule kinase inhibitors, biologics, nucleic acid drugs, gene editing technologies, and natural products in targeting the TGFβ signaling pathway, their clinical application still faces obstacles, especially as long-term inhibition may trigger side effects such as immunosuppression and tumorigenesis. Therefore, there is a need to develop more selective inhibitors. Future studies will aim to optimize treatment strategies, enhance efficacy, reduce side effects, and promote the integration of traditional Chinese medicine with modern biotechnology to provide more comprehensive solutions for the treatment of connective tissue diseases.
Summary
The TGFβ signaling pathway, as an important therapeutic target for CTDs, has demonstrated significant clinical potential. However, existing therapeutic approaches still face many challenges in terms of specificity, efficacy sustainability and side effect control. Future research should focus on optimizing existing therapeutic strategies and exploring more precise and personalized therapeutic regimens to improve efficacy, reduce side effects, and promote the application of TGFβ-targeted therapies in more connective tissue diseases. Through precise stratification of treatment, development of selective inhibitors, innovative delivery systems, and optimization of combination therapy strategies, it is expected that more comprehensive and efficient treatment solutions will be provided for patients with CTDs.
Abbreviations
CTDs, Connective tissue diseases; TGFβ, The transforming growth factor-β; TβR1, Transforming Growth Factor-β Type 1 Receptor; SSc, systemic sclerosis; SLE, systemic lupus erythematosus; LN, lupus nephritis; RA, rheumatoid arthritis; SS, Sjögren’s syndrome; IPF, idiopathic pulmonary fibrosis; ILD, Interstitial Lung Disease; SSc-ILD, SSc-associated interstitial lung disease; RA-ILD, RA-associated interstitial lung disease; siRNA, Small interfering RNA; RNAi, RNA interference; ECM, extracellular matrix; α-SMA, α-smooth muscle actin; MMPs, matrix metalloproteinases; FLS, fibroblast-like synoviocytes; EndMT, mesenchymal stromal cell.
Data Sharing Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (grant 82274454).
Disclosure
The authors declare no conflicts of interest.
References
1. Thoreau B, Mouthon L. Pulmonary arterial hypertension associated with connective tissue diseases (CTD-PAH): recent and advanced data. Autoimmun Rev. 2024;23(4):103506. doi:10.1016/j.autrev.2023.103506
2. Turk MA, Hayworth JL, Nevskaya T, Pope JE. Ocular Manifestations in Rheumatoid Arthritis, Connective Tissue Disease, and Vasculitis: a Systematic Review and Metaanalysis. J Rheumatol. 2021;48:25–34. doi:10.3899/jrheum.190768
3. Mulhearn B, Tansley SL, McHugh NJ. Autoantibodies in connective tissue disease. Best Pract Res Clin Rheumatol. 2020;34:101462. doi:10.1016/j.berh.2019.101462
4. Lee CT, Strek ME. The other connective tissue disease-associated interstitial lung diseases: sjogren’s syndrome, mixed connective tissue disease, and systemic lupus erythematosus. Curr Opin Pulm Med. 2021;27:388–395. doi:10.1097/mcp.0000000000000791
5. Martin Calderon L, Pope JE. Precursors to Systemic Sclerosis and Systemic Lupus Erythematosus: from Undifferentiated Connective Tissue Disease to the Development of Identifiable Connective Tissue Diseases. Front Immunol. 2022;13:869172. doi:10.3389/fimmu.2022.869172
6. Kim K, Lee J, Jo YS. Factors for progressive pulmonary fibrosis in connective tissue disease-related interstitial lung disease. Ther Adv Respir Dis. 2023;17:17534666231212301. doi:10.1177/17534666231212301
7. Tzavlaki K, Moustakas A. TGF-β Signaling. Biomolecules. 2020;10. doi:10.3390/biom10030487.
8. Ong CH, Tham CL, Harith HH, Firdaus N, Israf DA. TGF-β-induced fibrosis: a review on the underlying mechanism and potential therapeutic strategies. Eur J Pharmacol. 2021;911:174510. doi:10.1016/j.ejphar.2021.174510
9. Zhang Y, Alexander PB, Wang XF. TGF-β Family Signaling in the Control of Cell Proliferation and Survival. Cold Spring Harb Perspect Biol. 2017;9. doi:10.1101/cshperspect.a022145.
10. Derynck R, Budi EH. Specificity, versatility, and control of TGF-β family signaling. Sci Signal. 2019;12. doi:10.1126/scisignal.aav5183.
11. Deng Z, Fan T, Xiao C, et al. TGF-β signaling in health, disease and therapeutics. Signal Transduct Target Ther. 2024;9. doi:10.1038/s41392-024-01764-w.
12. Massagué J, Sheppard D. TGF-β signaling in health and disease. Cell. 2023;186:4007–4037. doi:10.1016/j.cell.2023.07.036
13. Zhang T, He X, Caldwell L, et al. NUAK1 promotes organ fibrosis via YAP and TGF-β/SMAD signaling. Sci Transl Med. 2022;14:eaaz4028. doi:10.1126/scitranslmed.aaz4028
14. Peng D, Fu M, Wang M, Wei Y, Wei X. Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol Cancer. 2022;21:104. doi:10.1186/s12943-022-01569-x
15. Budi EH, Schaub JR, Decaris M, Turner S, Derynck R. TGF-β as a driver of fibrosis: physiological roles and therapeutic opportunities. J Pathol. 2021;254:358–373. doi:10.1002/path.5680
16. Shimbori C, Bellaye PS, Xia J, et al. Fibroblast growth factor-1 attenuates TGF-β1-induced lung fibrosis. J Pathol. 2016;240:197–210. doi:10.1002/path.4768
17. Meng XM, Nikolic-Paterson DJ, Lan HY. TGF-β: the master regulator of fibrosis. Nat Rev Nephrol. 2016;12:325–338. doi:10.1038/nrneph.2016.48
18. Wang H, Chen M, Sang X, et al. Development of small molecule inhibitors targeting TGF-β ligand and receptor: structures, mechanism, preclinical studies and clinical usage. Eur J Med Chem. 2020;191:112154. doi:10.1016/j.ejmech.2020.112154
19. Larson C, Oronsky B, Carter CA, et al. TGF-beta: a master immune regulator. Expert Opin Ther Targets. 2020;24:427–438. doi:10.1080/14728222.2020.1744568
20. DePeaux K, Rivadeneira DB, Lontos K, et al. An oncolytic virus-delivered TGFβ inhibitor overcomes the immunosuppressive tumor microenvironment. J Exp Med. 2023;220. doi:10.1084/jem.20230053
21. Carthy JM. TGFβ signaling and the control of myofibroblast differentiation: implications for chronic inflammatory disorders. J Cell Physiol. 2018;233:98–106. doi:10.1002/jcp.25879
22. Faust HJ, Cheng TY, Korsunsky I, et al. Adipocyte associated glucocorticoid signaling regulates normal fibroblast function which is lost in inflammatory arthritis. Nat Commun. 2024;15:9859. doi:10.1038/s41467-024-52586-x
23. Vallée A, Lecarpentier Y. TGF-β in fibrosis by acting as a conductor for contractile properties of myofibroblasts. Cell Biosci. 2019;9:98. doi:10.1186/s13578-019-0362-3
24. Hanley CJ, Waise S, Ellis MJ, et al. Single-cell analysis reveals prognostic fibroblast subpopulations linked to molecular and immunological subtypes of lung cancer. Nat Commun. 2023;14:387. doi:10.1038/s41467-023-35832-6
25. Scharenberg MA, Pippenger BE, Sack R, et al. TGF-β-induced differentiation into myofibroblasts involves specific regulation of two MKL1 isoforms. J Cell Sci. 2014;127:1079–1091. doi:10.1242/jcs.142075
26. Chakravarthy A, Khan L, Bensler NP, Bose P, De Carvalho DD. TGF-β-associated extracellular matrix genes link cancer-associated fibroblasts to immune evasion and immunotherapy failure. Nat Commun. 2018;9:4692. doi:10.1038/s41467-018-06654-8
27. Mishra R, Haldar S, Biondi S, Bhari VK, Singh G, Bhowmick. TGF-β controls stromal telomere length through epigenetic modifications. 3 Biotech. 2022;12:290. doi:10.1007/s13205-022-03346-5
28. Epstein Shochet G, Brook E, Bardenstein-Wald B, Shitrit D. TGF-β pathway activation by idiopathic pulmonary fibrosis (IPF) fibroblast derived soluble factors is mediated by IL-6 trans-signaling. Respir Res. 2020;21:56. doi:10.1186/s12931-020-1319-0
29. McKee TJ, Perlman G, Morris M, Komarova SV. Extracellular matrix composition of connective tissues: a systematic review and meta-analysis. Sci Rep. 2019;9:10542. doi:10.1038/s41598-019-46896-0
30. Park HN, Song MJ, Choi YE, Lee DH, Chung JH, Lee ST. LRG1 Promotes ECM Integrity by Activating the TGF-β Signaling Pathway in Fibroblasts. Int J Mol Sci. 2023;24. doi:10.3390/ijms241512445.
31. Laczko R, Csiszar K. Lysyl Oxidase (LOX): functional Contributions to Signaling Pathways. Biomolecules. 2020;10. doi:10.3390/biom10081093.
32. Li H, Zhu X, Cao X, Lu Y, Zhou J, Zhang X. Single-cell analysis reveals lysyl oxidase (Lox)(+) fibroblast subset involved in cardiac fibrosis of diabetic mice. J Adv Res. 2023;54:223–237. doi:10.1016/j.jare.2023.01.018
33. Lu Y, Li H, Chen M, Lin Y, Zhang X. LOX-induced tubulointerstitial fibrosis via the TGF-β/LOX/Snail axis in diabetic mice. J Transl Med. 2025;23:35. doi:10.1186/s12967-024-06056-z
34. Shi X, Young CD, Zhou H, Wang X. Transforming Growth Factor-β Signaling in Fibrotic Diseases and Cancer-Associated Fibroblasts. Biomolecules. 2020;10. doi:10.3390/biom10121666.
35. Yao Y, Findlay A, Stolp J, Rayner B, Ask K, Jarolimek W. Pan-Lysyl Oxidase Inhibitor PXS-5505 Ameliorates Multiple-Organ Fibrosis by Inhibiting Collagen Crosslinks in Rodent Models of Systemic Sclerosis. Int J Mol Sci. 2022;23. doi:10.3390/ijms23105533.
36. Bitterman P. Fibroblast-Matrix Cross-Talk in Idiopathic Pulmonary Fibrosis: cross-Links at the Crossroads. Am J Respir Cell Mol Biol. 2018;58:547–548. doi:10.1165/rcmb.2017-0402ED
37. Wieteska Ł, Taylor AB, Punch E, et al. Structures of TGF-β with betaglycan and signaling receptors reveal mechanisms of complex assembly and signaling. Nat Commun. 2025;16:1778. doi:10.1038/s41467-025-56796-9
38. Mouawad JE, Feghali-Bostwick C. The Molecular Mechanisms of Systemic Sclerosis-Associated Lung Fibrosis. Int J Mol Sci. 2023;24. doi:10.3390/ijms24032963.
39. Stolfi C, Troncone E, Marafini I, Monteleone G. Role of TGF-Beta and Smad7 in Gut Inflammation, Fibrosis and Cancer. Biomolecules. 2020;11. doi:10.3390/biom11010017.
40. Nie YM, Zhou WQ, Niu T, et al. Peptidoglycan isolated from the fruit of Lycium barbarum alleviates liver fibrosis in mice by regulating the TGF-β/Smad7 signaling and gut microbiota. Acta Pharmacol Sin. 2025. doi:10.1038/s41401-024-01454-x
41. Aashaq S, Batool A, Mir SA, Beigh MA, Andrabi KI, Shah ZA. TGF-β signaling: a recap of SMAD-independent and SMAD-dependent pathways. J Cell Physiol. 2022;237:59–85. doi:10.1002/jcp.30529
42. Choi ME, Ding Y, Kim SI. TGF-β signaling via TAK1 pathway: role in kidney fibrosis. Semin Nephrol. 2012;32:244–252. doi:10.1016/j.semnephrol.2012.04.003
43. Totzke J, Scarneo SA, Yang KW, Haystead TAJ. TAK1: a potent tumour necrosis factor inhibitor for the treatment of inflammatory diseases. Open Biol. 2020;10:200099. doi:10.1098/rsob.200099
44. Freeze R, Yang KW, Haystead T, Hughes P, Scarneo S. Delineation of the distinct inflammatory signaling roles of TAK1 and JAK1/3 in the CIA model of rheumatoid arthritis. Pharmacol Res Perspect. 2023;11:e01124. doi:10.1002/prp2.1124
45. Chen X, Zhu L, Xu J, et al. Semaphorin 5A promotes Th17 differentiation via PI3K-Akt-mTOR in systemic lupus erythematosus. Arthritis Res Ther. 2024;26:204. doi:10.1186/s13075-024-03437-z
46. Bacalao MA, Satterthwaite AB. Recent Advances in Lupus B Cell Biology: PI3K, IFNγ, and Chromatin. Front Immunol. 2020;11:615673. doi:10.3389/fimmu.2020.615673
47. Blumer S, Fang L, Chen WC, et al. IPF-Fibroblast Erk1/2 Activity Is Independent from microRNA Cluster 17-92 but Can Be Inhibited by Treprostinil through DUSP1. Cells. 2021;10. doi:10.3390/cells10112836.
48. Dorry S, Perla S, Bennett AM. MAPK Phosphatase-5 is required for TGF-β signaling through a JNK-dependent pathway. bioRxiv. 2024. doi:10.1101/2024.06.27.600976
49. Ye Z, Hu Y. TGF‑β1: gentlemanly orchestrator in idiopathic pulmonary fibrosis (Review). Int J Mol Med. 2021;48. doi:10.3892/ijmm.2021.4965.
50. van Caam A, Aarts J, van Ee T, et al. TGFβ-mediated expression of TGFβ-activating integrins in SSc monocytes: disturbed activation of latent TGFβ? Arthritis Res Ther. 2020;22:42. doi:10.1186/s13075-020-2130-5
51. Fang L, Chen H, Kong R, Que J. Endogenous tryptophan metabolite 5-Methoxytryptophan inhibits pulmonary fibrosis by downregulating the TGF-β/SMAD3 and PI3K/AKT signaling pathway. Life Sci. 2020;260:118399. doi:10.1016/j.lfs.2020.118399
52. Luo K. Signaling Cross Talk between TGF-β/Smad and Other Signaling Pathways. Cold Spring Harb Perspect Biol. 2017;9. doi:10.1101/cshperspect.a022137.
53. Hata A, Chen YG. TGF-β Signaling from Receptors to Smads. Cold Spring Harb Perspect Biol. 2016;8. doi:10.1101/cshperspect.a022061.
54. Zou Y, Dai J, Li J, et al. Role of the TGF‑β/Smad signaling pathway in the transition from acute kidney injury to chronic kidney disease (Review). Int J Mol Med. 2025;56. doi:10.3892/ijmm.2025.5603.
55. Ma J, Sanchez-Duffhues G, Goumans MJ, Ten Dijke P. TGF-β-Induced Endothelial to Mesenchymal Transition in Disease and Tissue Engineering. Front Cell Dev Biol. 2020;8:260. doi:10.3389/fcell.2020.00260
56. Inui N, Sakai S, Kitagawa M. Molecular Pathogenesis of Pulmonary Fibrosis, with Focus on Pathways Related to TGF-β and the Ubiquitin-Proteasome Pathway. Int J Mol Sci. 2021;22. doi:10.3390/ijms22116107.
57. Campitiello R, Soldano S, Gotelli E, et al. The intervention of macrophages in progressive fibrosis characterizing systemic sclerosis: a systematic review. Autoimmun Rev. 2024;23:103637. doi:10.1016/j.autrev.2024.103637
58. Tauriello DVF, Sancho E, Batlle E. Overcoming TGFβ-mediated immune evasion in cancer. Nat Rev Cancer. 2022;22:25–44. doi:10.1038/s41568-021-00413-6
59. Zhao H, Wei J, Sun J. Roles of TGF-β signaling pathway in tumor microenvirionment and cancer therapy. Int Immunopharmacol. 2020;89:107101. doi:10.1016/j.intimp.2020.107101
60. Nixon BG, Gao S, Wang X, Li MO. TGFβ control of immune responses in cancer: a holistic immuno-oncology perspective. Nat Rev Immunol. 2023;23:346–362. doi:10.1038/s41577-022-00796-z
61. Chen W. TGF-β Regulation of T Cells. Annu Rev Immunol. 2023;41:483–512. doi:10.1146/annurev-immunol-101921-045939
62. Jiang Y, Cai R, Huang Y, et al. Macrophages in organ fibrosis: from pathogenesis to therapeutic targets. Cell Death Discov. 2024;10:487. doi:10.1038/s41420-024-02247-1
63. Tamayo E, Alvarez P, Merino R. TGFβ Superfamily Members as Regulators of B Cell Development and Function-Implications for Autoimmunity. Int J Mol Sci. 2018;19. doi:10.3390/ijms19123928.
64. Lebrun JJ. The Dual Role of TGFβ in Human Cancer: from Tumor Suppression to Cancer Metastasis. ISRN Mol Biol. 2012;2012:381428. doi:10.5402/2012/381428
65. Konkel JE, Zhang D, Zanvit P, et al. Transforming Growth Factor-β Signaling in Regulatory T Cells Controls T Helper-17 Cells and Tissue-Specific Immune Responses. Immunity. 2017;46:660–674. doi:10.1016/j.immuni.2017.03.015
66. Wang J, Zhao X, Wan YY. Intricacies of TGF-β signaling in Treg and Th17 cell biology. Cell Mol Immunol. 2023;20:1002–1022. doi:10.1038/s41423-023-01036-7
67. Huang J, Li X, Zhu Q, Wang M, Xie Z, Zhao T. Imbalance of Th17 cells, Treg cells and associated cytokines in patients with systemic lupus erythematosus: a meta-analysis. Front Immunol. 2024;15:1425847. doi:10.3389/fimmu.2024.1425847
68. Sun X, Cui Y, Feng H, Liu H, Liu X. TGF-β signaling controls Foxp3 methylation and T reg cell differentiation by modulating Uhrf1 activity. J Exp Med. 2019;216:2819–2837. doi:10.1084/jem.20190550
69. Koga T. Understanding the pathogenic significance of altered calcium-calmodulin signaling in T cells in autoimmune diseases. Clin Immunol. 2024;262:110177. doi:10.1016/j.clim.2024.110177
70. Tenbrock K, Rauen T. T cell dysregulation in SLE. Clin Immunol. 2022;239:109031. doi:10.1016/j.clim.2022.109031
71. Sarkar S, Cooney LA, Fox DA. The role of T helper type 17 cells in inflammatory arthritis. Clin Exp Immunol. 2010;159:225–237. doi:10.1111/j.1365-2249.2009.04016.x
72. Tseng CC, Sung YW, Chen KY, et al. The Role of Macrophages in Connective Tissue Disease-Associated Interstitial Lung Disease: focusing on Molecular Mechanisms and Potential Treatment Strategies. Int J Mol Sci. 2023;24. doi:10.3390/ijms241511995
73. Oliver MA, Davis XD, Bohannon JK. TGFβ macrophage reprogramming: a new dimension of macrophage plasticity. J Leukoc Biol. 2024;115:411–414. doi:10.1093/jleuko/qiae001
74. Zhang F, Wang H, Wang X, et al. TGF-β induces M2-like macrophage polarization via SNAIL-mediated suppression of a pro-inflammatory phenotype. Oncotarget. 2016;7:52294–52306. doi:10.18632/oncotarget.10561
75. Pang QM, Zhang Q, Wu XC, et al. Mechanism of M2 macrophages modulating astrocyte polarization through the TGF-β/PI3K/Akt pathway. Immunol Lett. 2023;259:1–8. doi:10.1016/j.imlet.2023.05.009
76. Hu M, Yao Z, Xu L, et al. M2 macrophage polarization in systemic sclerosis fibrosis: pathogenic mechanisms and therapeutic effects. Heliyon. 2023;9:e16206. doi:10.1016/j.heliyon.2023.e16206
77. Ge Z, Chen Y, Ma L, Hu F, Xie L. Macrophage polarization and its impact on idiopathic pulmonary fibrosis. Front Immunol. 2024;15:1444964. doi:10.3389/fimmu.2024.1444964
78. Hou J, Shi J, Chen L, et al. M2 macrophages promote myofibroblast differentiation of LR-MSCs and are associated with pulmonary fibrogenesis. Cell Commun Signal. 2018;16:89. doi:10.1186/s12964-018-0300-8
79. Huai G, Markmann JF, Deng S, Rickert CG. TGF-β-secreting regulatory B cells: unsung players in immune regulation. Clin Transl Immunology. 2021;10:e1270. doi:10.1002/cti2.1270
80. Xiao F, Rui K, Shi X, et al. Epigenetic regulation of B cells and its role in autoimmune pathogenesis. Cell Mol Immunol. 2022;19:1215–1234. doi:10.1038/s41423-022-00933-7
81. Tie Y, Tang F, Peng D, Zhang Y, Shi H. TGF-beta signal transduction: biology, function and therapy for diseases. Mol Biomed. 2022;3:45. doi:10.1186/s43556-022-00109-9
82. Volkmann ER, Andréasson K, Smith V. Systemic sclerosis. Lancet. 2023;401:304–318. doi:10.1016/s0140-6736(22)01692-0
83. Rekik R, Smiti khanfir M, Larbi T, et al. Impaired TGF-β signaling in patients with active systemic lupus erythematosus is associated with an overexpression of IL-22. Cytokine. 2018;108:182–189. doi:10.1016/j.cyto.2018.04.011
84. Yingling JM, McMillen WT, Yan L, et al. Preclinical assessment of galunisertib (LY2157299 monohydrate), a first-in-class transforming growth factor-β receptor type I inhibitor. Oncotarget. 2018;9:6659–6677. doi:10.18632/oncotarget.23795
85. Melisi D, Oh DY, Hollebecque A, et al. Safety and activity of the TGFβ receptor I kinase inhibitor galunisertib plus the anti-PD-L1 antibody durvalumab in metastatic pancreatic cancer. J Immunother Cancer. 2021;9. doi:10.1136/jitc-2020-002068
86. Maier A, Peille AL, Vuaroqueaux V, Lahn M. Anti-tumor activity of the TGF-β receptor kinase inhibitor galunisertib (LY2157299 monohydrate) in patient-derived tumor xenografts. Cell Oncol Dordr. 2015;38:131–144. doi:10.1007/s13402-014-0210-8
87. Halder SK, Beauchamp RD, Datta PK. A specific inhibitor of TGF-beta receptor kinase, SB-431542, as a potent antitumor agent for human cancers. Neoplasia. 2005;7:509–521. doi:10.1593/neo.04640
88. Melisi D, Ishiyama S, Sclabas GM, et al. LY2109761, a novel transforming growth factor beta receptor type I and type II dual inhibitor, as a therapeutic approach to suppressing pancreatic cancer metastasis. Mol Cancer Ther. 2008;7:829–840. doi:10.1158/1535-7163.Mct-07-0337
89. Zhang ZH, Miao YY, Ke BL, Liu K, Xu X. LY2109761, Transforming Growth Factor β Receptor Type I and Type II Dual Inhibitor, is a Novel Approach to Suppress Endothelial Mesenchymal Transformation in Human Corneal Endothelial Cells. Cell Physiol Biochem. 2018;50:963–972. doi:10.1159/000494480
90. He X, Guo X, Zhang H, Kong X, Yang F, Zheng C. Mechanism of action and efficacy of LY2109761, a TGF-β receptor inhibitor, targeting tumor microenvironment in liver cancer after TACE. Oncotarget. 2018;9:1130–1142. doi:10.18632/oncotarget.23193
91. Malek E, Rana PS, Swamydas M, et al. Vactosertib, a novel TGF-β1 type I receptor kinase inhibitor, improves T-cell fitness: a single-arm, phase 1b trial in relapsed/refractory multiple myeloma. Res Sq. 2023. doi:10.21203/rs.3.rs-3112163/v1
92. Song KM, Chung DY, Choi MJ, et al. Vactosertib, a Novel, Orally Bioavailable Activin Receptor-Like Kinase 5 Inhibitor, Promotes Regression of Fibrotic Plaques in a Rat Model of Peyronie’s Disease. World J Mens Health. 2020;38:552–563. doi:10.5534/wjmh.190071
93. Morris JC, Tan AR, Olencki TE, et al. Phase I study of GC1008 (fresolimumab): a human anti-transforming growth factor-beta (TGFβ) monoclonal antibody in patients with advanced malignant melanoma or renal cell carcinoma. PLoS One. 2014;9:e90353. doi:10.1371/journal.pone.0090353
94. Moulin A, Mathieu M, Lawrence C, et al. Structures of a pan-specific antagonist antibody complexed to different isoforms of TGFβ reveal structural plasticity of antibody-antigen interactions. Protein Sci. 2014;23:1698–1707. doi:10.1002/pro.2548
95. Richeldi L, Fernández Pérez ER, Costabel U, et al. Pamrevlumab, an anti-connective tissue growth factor therapy, for idiopathic pulmonary fibrosis (PRAISE): a Phase 2, randomised, double-blind, placebo-controlled trial. Lancet Respir Med. 2020;8:25–33. doi:10.1016/s2213-2600(19)30262-0
96. Brenner MC, Krzyzanski W, Chou JZ, et al. FG-3019, a Human Monoclonal Antibody Recognizing Connective Tissue Growth Factor, is Subject to Target-Mediated Drug Disposition. Pharm Res. 2016;33:1833–1849. doi:10.1007/s11095-016-1918-0
97. Mascarenhas J, Migliaccio AR, Kosiorek H, et al. A Phase Ib Trial of AVID200, a TGFβ 1/3 Trap, in Patients with Myelofibrosis. Clin Cancer Res. 2023;29:3622–3632. doi:10.1158/1078-0432.Ccr-23-0276
98. Tabe Y, Shi YX, Zeng Z, et al. TGF-β-Neutralizing Antibody 1D11 Enhances Cytarabine-Induced Apoptosis in AML Cells in the Bone Marrow Microenvironment. PLoS One. 2013;8:e62785. doi:10.1371/journal.pone.0062785
99. Bertolio MS, La Colla A, Carrea A, et al. A Novel Splice Variant of Human TGF-β Type II Receptor Encodes a Soluble Protein and Its Fc-Tagged Version Prevents Liver Fibrosis in vivo. Front Cell Dev Biol. 2021;9:690397. doi:10.3389/fcell.2021.690397
100. Huber-Ruano I, Raventós C, Cuartas I, et al. An antisense oligonucleotide targeting TGF-β2 inhibits lung metastasis and induces CD86 expression in tumor-associated macrophages. Ann Oncol. 2017;28:2278–2285. doi:10.1093/annonc/mdx314
101. Kuespert S, Heydn R, Peters S, et al. Antisense Oligonucleotide in LNA-Gapmer Design Targeting TGFBR2-A Key Single Gene Target for Safe and Effective Inhibition of TGFβ Signaling. Int J Mol Sci. 2020;21. doi:10.3390/ijms21061952.
102. Yang M, Qin C, Tao L, et al. Synchronous targeted delivery of TGF-β siRNA to stromal and tumor cells elicits robust antitumor immunity against triple-negative breast cancer by comprehensively remodeling the tumor microenvironment. Biomaterials. 2023;301:122253. doi:10.1016/j.biomaterials.2023.122253
103. Davies N, Hovdal D, Edmunds N, et al. Functionalized lipid nanoparticles for subcutaneous administration of mRNA to achieve systemic exposures of a therapeutic protein. Mol Ther Nucleic Acids. 2021;24:369–384. doi:10.1016/j.omtn.2021.03.008
104. Tang N, Cheng C, Zhang X, et al. TGF-β inhibition via CRISPR promotes the long-term efficacy of CAR T cells against solid tumors. JCI Insight. 2020;5. doi:10.1172/jci.insight.133977.
105. Alishah K, Birtel M, Masoumi E, et al. CRISPR/Cas9-mediated TGFβRII disruption enhances anti-tumor efficacy of human chimeric antigen receptor T cells in vitro. J Transl Med. 2021;19:482. doi:10.1186/s12967-021-03146-0
106. Asmamaw M, Zawdie B. Mechanism and Applications of CRISPR/Cas-9-Mediated Genome Editing. Biologics. 2021;15:353–361. doi:10.2147/btt.S326422
107. Roberts AB, Tian F, Byfield SD, et al. Smad3 is key to TGF-beta-mediated epithelial-to-mesenchymal transition, fibrosis, tumor suppression and metastasis. Cytokine Growth Factor Rev. 2006;17:19–27. doi:10.1016/j.cytogfr.2005.09.008
108. Lian GY, Wang QM, Mak TS, Huang XR, Yu XQ, Lan HY. Disrupting Smad3 potentiates immunostimulatory function of NK cells against lung carcinoma by promoting GM-CSF production. Cell Mol Life Sci. 2024;81:262. doi:10.1007/s00018-024-05290-4
109. Song K, Peng S, Sun Z, Li H, Yang R. Curcumin suppresses TGF-β signaling by inhibition of TGIF degradation in scleroderma fibroblasts. Biochem Biophys Res Commun. 2011;411:821–825. doi:10.1016/j.bbrc.2011.07.044
110. Datta R, Halder SK, Zhang B. Role of TGF-β signaling in curcumin-mediated inhibition of tumorigenicity of human lung cancer cells. J Cancer Res Clin Oncol. 2013;139:563–572. doi:10.1007/s00432-012-1352-6
111. Zheng W, Mei Y, Chen C, Cai L, Chen H. The effectiveness and safety of Tripterygium wilfordii glycosides combined with disease-modifying anti-rheumatic drugs in the treatment of rheumatoid arthritis: a systematic review and meta-analysis of 40 randomized controlled trials. Phytother Res. 2021;35:2902–2924. doi:10.1002/ptr.6996
112. Lin N, Zhang YQ, Jiang Q, et al. Clinical Practice Guideline for Tripterygium Glycosides/Tripterygium wilfordii Tablets in the Treatment of Rheumatoid Arthritis. Front Pharmacol. 2020;11:608703. doi:10.3389/fphar.2020.608703
113. Zhang L, Lin W, Chen X, Wei G, Zhu H, Xing S. Tanshinone IIA reverses EGF- and TGF-β1-mediated epithelial-mesenchymal transition in HepG2 cells via the PI3K/Akt/ERK signaling pathway. Oncol Lett. 2019;18:6554–6562. doi:10.3892/ol.2019.11032
114. Jiang Y, Bi Y, Zhou L, Zheng S, Jian T, Chen J. Tanshinone IIA inhibits proliferation and migration by downregulation of the PI3K/Akt pathway in small cell lung cancer cells. BMC Complement Med Ther. 2024;24:68. doi:10.1186/s12906-024-04363-y
115. Alam SSM, Samanta A, Uddin F, Ali S, Hoque M. Tanshinone IIA targeting cell signaling pathways: a plausible paradigm for cancer therapy. Pharmacol Rep. 2023;75:907–922. doi:10.1007/s43440-023-00507-y
116. Sandhiutami NMD, Arozal W, Louisa M, Rahmat D, Wuyung PE. Curcumin Nanoparticle Enhances the Anticancer Effect of Cisplatin by Inhibiting PI3K/AKT and JAK/STAT3 Pathway in Rat Ovarian Carcinoma Induced by DMBA. Front Pharmacol. 2020;11:603235. doi:10.3389/fphar.2020.603235
117. Shamsi M, Mohammadzadeh G, Hatami M, Roshanazadeh M, Noor-Behbahani M, Rashidi M. Dual Modulation of Canonical and Non-canonical TGF-β/ROS/Erk1/2 Pathways: synergistic Activation of Nrf-2 and Antioxidant Enzymes (SOD1, GPx, HO-1) by Quercetin Loaded in Solid Lipid Nanoparticles and Curcumin in Atherosclerosis Therapy. Iran J Pharm Res. 2024;23:e151428. doi:10.5812/ijpr-151428
118. Ashrafizadeh M, Najafi M, Orouei S, et al. Resveratrol Modulates Transforming Growth Factor-Beta (TGF-β) Signaling Pathway for Disease Therapy: a New Insight into Its Pharmacological Activities. Biomedicines. 2020;8. doi:10.3390/biomedicines8080261.
119. Fan J, Wei S, Zhang X, et al. Resveratrol inhibits TGF-β1-induced fibrotic effects in human pterygium fibroblasts. Environ Health Prev Med. 2023;28:59. doi:10.1265/ehpm.23-00020
120. Lee HJ. Recent Advances in the Development of TGF-β Signaling Inhibitors for Anticancer Therapy. J Cancer Prev. 2020;25:213–222. doi:10.15430/jcp.2020.25.4.213
121. Goebel GL, Qiu X, Wu P. Kinase-targeting small-molecule inhibitors and emerging bifunctional molecules. Trends Pharmacol Sci. 2022;43:866–881. doi:10.1016/j.tips.2022.04.006
122. Holmgaard RB, Schaer DA, Li Y, et al. Targeting the TGFβ pathway with galunisertib, a TGFβRI small molecule inhibitor, promotes anti-tumor immunity leading to durable, complete responses, as monotherapy and in combination with checkpoint blockade. J Immunother Cancer. 2018;6:47. doi:10.1186/s40425-018-0356-4
123. Herbertz S, Sawyer JS, Stauber AJ, et al. Clinical development of galunisertib (LY2157299 monohydrate), a small molecule inhibitor of transforming growth factor-beta signaling pathway. Drug Des Devel Ther. 2015;9:4479–4499. doi:10.2147/dddt.S86621
124. Peterson JM, Jay JW, Wang Y, et al. Galunisertib Exerts Antifibrotic Effects on TGF-β-Induced Fibroproliferative Dermal Fibroblasts. Int J Mol Sci. 2022;23. doi:10.3390/ijms23126689.
125. Masuda A, Nakamura T, Abe M, et al. Promotion of liver regeneration and anti‑fibrotic effects of the TGF‑β receptor kinase inhibitor galunisertib in CCl4‑treated mice. Int J Mol Med. 2020;46:427–438. doi:10.3892/ijmm.2020.4594
126. Binabaj MM, Asgharzadeh F, Rahmani F, et al. Vactosertib potently improves anti-tumor properties of 5-FU for colon cancer. Daru. 2023;31:193–203. doi:10.1007/s40199-023-00474-y
127. Welsh BT, Faucette R, Bilic S, et al. Nonclinical Development of SRK-181: an Anti-Latent TGFβ1 Monoclonal Antibody for the Treatment of Locally Advanced or Metastatic Solid Tumors. Int J Toxicol. 2021;40:226–241. doi:10.1177/1091581821998945
128. Martin CJ, Datta A, Littlefield C, et al. Selective inhibition of TGFβ1 activation overcomes primary resistance to checkpoint blockade therapy by altering tumor immune landscape. Sci Transl Med. 2020;12. doi:10.1126/scitranslmed.aay8456
129. Zhang J, Chen B, Gan C, Sun H, Zhang J, Feng L. A Comprehensive Review of Small Interfering RNAs (siRNAs): mechanism, Therapeutic Targets, and Delivery Strategies for Cancer Therapy. Int J Nanomed. 2023;18:7605–7635. doi:10.2147/ijn.S436038
130. Hamouda AEI, Filtjens J, Brabants E, et al. Intratumoral delivery of lipid nanoparticle-formulated mRNA encoding IL-21, IL-7, and 4-1BBL induces systemic anti-tumor immunity. Nat Commun. 2024;15:10635. doi:10.1038/s41467-024-54877-9
131. Bai X, Chen Q, Li F, et al. Optimized inhaled LNP formulation for enhanced treatment of idiopathic pulmonary fibrosis via mRNA-mediated antibody therapy. Nat Commun. 2024;15:6844. doi:10.1038/s41467-024-51056-8
132. Yu S, Ericson M, Fanjul A, et al. Genome-wide CRISPR Screening to Identify Drivers of TGF-β-Induced Liver Fibrosis in Human Hepatic Stellate Cells. ACS Chem Biol. 2022;17:918–929. doi:10.1021/acschembio.2c00006
133. Jones DL, Haak AJ, Caporarello N, et al. TGFβ-induced fibroblast activation requires persistent and targeted HDAC-mediated gene repression. J Cell Sci. 2019;132. doi:10.1242/jcs.233486.
134. Na L, Quan J, Wei L, et al. Clinical practice guideline for Tripterygium Glycosides/Tripterygium wilfordii Tablets in treatment of rheumatoid arthritis. Zhongguo Zhong Yao Za Zhi. 2020;45:4149–4153. doi:10.19540/j.cnki.cjcmm.20200710.501
135. Jiang M, Xie Y, Wang P, Du M, Wang Y, Yan S. Research Progress of Triptolide Against Fibrosis. Drug Des Devel Ther. 2024;18:3255–3266. doi:10.2147/dddt.S467929
136. Yang L, Wang Q, Hou Y, et al. The Chinese herb Tripterygium wilfordii Hook F for the treatment of systemic sclerosis-associated interstitial lung disease: data from a Chinese EUSTAR Center. Clin Rheumatol. 2020;39:813–821. doi:10.1007/s10067-019-04784-y
137. Shan Y, Zhao J, Wei K, et al. A comprehensive review of Tripterygium wilfordii hook. f. in the treatment of rheumatic and autoimmune diseases: bioactive compounds, mechanisms of action, and future directions. Front Pharmacol. 2023;14:1282610. doi:10.3389/fphar.2023.1282610
138. Xu L, Zhang Y, Ji N, et al. Tanshinone IIA regulates the TGF-β1/Smad signaling pathway to ameliorate non-alcoholic steatohepatitis-related fibrosis. Exp Ther Med. 2022;24:486. doi:10.3892/etm.2022.11413
139. Zhan CY, Tang JH, Zhou DX, Li ZH. Effects of tanshinone IIA on the transforming growth factor β1/Smad signaling pathway in rat cardiac fibroblasts. Indian J Pharmacol. 2014;46:633–638. doi:10.4103/0253-7613.144933
140. Song SL, Gong ZJ, Zhang QR, Huang TX. Effects of Chinese traditional compound, JinSanE, on expression of TGF-beta1 and TGF-beta1 type II receptor mRNA, Smad3 and Smad7 on experimental hepatic fibrosis in vivo. World J Gastroenterol. 2005;11:2269–2276. doi:10.3748/wjg.v11.i15.2269
141. Ashrafizadeh M, Zarrabi A, Hushmandi K, et al. Toward Regulatory Effects of Curcumin on Transforming Growth Factor-Beta Across Different Diseases: a Review. Front Pharmacol. 2020;11:585413. doi:10.3389/fphar.2020.585413
142. Kunihiro AG, Brickey JA, Frye JB, et al. Curcumin Inhibition of TGFβ signaling in bone metastatic breast cancer cells and the possible role of oxidative metabolites. J Nutr Biochem. 2022;99:108842. doi:10.1016/j.jnutbio.2021.108842
143. Kunihiro AG, Brickey JA, Frye JB, Luis PB, Schneider C, Funk JL. Curcumin, but not curcumin-glucuronide, inhibits Smad signaling in TGFβ-dependent bone metastatic breast cancer cells and is enriched in bone compared to other tissues. J Nutr Biochem. 2019;63:150–156. doi:10.1016/j.jnutbio.2018.09.021
144. Jerjen R, Nikpour M, Krieg T, Denton CP, Saracino AM. Systemic sclerosis in adults. Part I: clinical features and pathogenesis. J Am Acad Dermatol. 2022;87:937–954. doi:10.1016/j.jaad.2021.10.065
145. Ayers NB, Sun CM, Chen SY. Transforming growth factor-β signaling in systemic sclerosis. J Biomed Res. 2018;32:3–12. doi:10.7555/jbr.31.20170034
146. Sun T, Vander Heiden JA, Gao X, et al. Isoform-selective TGF-β3 inhibition for systemic sclerosis. Med. 2024;5:132–147.e137. doi:10.1016/j.medj.2023.12.011
147. Makino K, Makino T, Stawski L, Lipson KE, Leask A, Trojanowska M. Anti-connective tissue growth factor (CTGF/CCN2) monoclonal antibody attenuates skin fibrosis in mice models of systemic sclerosis. Arthritis Res Ther. 2017;19:134. doi:10.1186/s13075-017-1356-3
148. Kawano Y, Chung L. Promise and challenge of systemic sclerosis therapies. Nat Rev Rheumatol. 2021;17:581–582. doi:10.1038/s41584-021-00678-z
149. Tsujino K, Reed NI, Atakilit A, Ren X, Sheppard D. Transforming growth factor-β plays divergent roles in modulating vascular remodeling, inflammation, and pulmonary fibrosis in a murine model of scleroderma. Am J Physiol Lung Cell Mol Physiol. 2017;312:L22–l31. doi:10.1152/ajplung.00428.2016
150. Siegel CH, Sammaritano LR. Systemic Lupus Erythematosus: a Review. JAMA. 2024;331:1480–1491. doi:10.1001/jama.2024.2315
151. Kiriakidou M, Ching CL. Systemic Lupus Erythematosus. Ann Intern Med. 2020;172:Itc81–itc96. doi:10.7326/aitc202006020
152. Robinson GA, Wilkinson MGL, Wincup C. The Role of Immunometabolism in the Pathogenesis of Systemic Lupus Erythematosus. Front Immunol. 2021;12:806560. doi:10.3389/fimmu.2021.806560
153. Yang J, Yang X, Wang L, Li M. B cells control lupus autoimmunity by inhibiting Th17 and promoting Th22 cells. Cell Death Dis. 2020;11:164. doi:10.1038/s41419-020-2362-y
154. Yu C, Li P, Dang X, Zhang X, Mao Y, Chen X. Lupus nephritis: new progress in diagnosis and treatment. J Autoimmun. 2022;132:102871. doi:10.1016/j.jaut.2022.102871
155. Anders HJ, Saxena R, Zhao MH, Parodis I, Salmon JE, Mohan C. Lupus nephritis. Nat Rev Dis Primers. 2020;6:7. doi:10.1038/s41572-019-0141-9
156. Shou J, Cao J, Zhang S, et al. SIS3, a specific inhibitor of smad3, attenuates bleomycin-induced pulmonary fibrosis in mice. Biochem Biophys Res Commun. 2018;503:757–762. doi:10.1016/j.bbrc.2018.06.072
157. Fava A, Rao DA, Mohan C, et al. Urine Proteomics and Renal Single-Cell Transcriptomics Implicate Interleukin-16 in Lupus Nephritis. Arthritis Rheumatol. 2022;74:829–839. doi:10.1002/art.42023
158. Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet. 2016;388:2023–2038. doi:10.1016/s0140-6736(16)30173-8
159. Gravallese EM, Firestein GS, Koscal N, et al. What Is Rheumatoid Arthritis? N Engl J Med. 2024;390:e32. doi:10.1056/NEJMp2310178
160. Radu AF, Bungau SG. Management of Rheumatoid Arthritis: an Overview. Cells. 2021;10. doi:10.3390/cells10112857.
161. Stanford SM, Aleman Muench GR, Bartok B, et al. TGFβ responsive tyrosine phosphatase promotes rheumatoid synovial fibroblast invasiveness. Ann Rheum Dis. 2016;75:295–302. doi:10.1136/annrheumdis-2014-205790
162. Yao Z, Boyce BF. TGFβ priming promotes TNF-induced bone erosion: a promising new target in RA? Nat Rev Rheumatol. 2022;18:617–618. doi:10.1038/s41584-022-00843-y
163. Infante A, Alcorta-Sevillano N, Macías I, et al. Galunisertib downregulates mutant type I collagen expression and promotes MSCs osteogenesis in pediatric osteogenesis imperfecta. Biomed Pharmacother. 2024;175:116725. doi:10.1016/j.biopha.2024.116725
164. Liang M, Matteson EL, Abril A, Distler JHW. The role of antifibrotics in the treatment of rheumatoid arthritis-associated interstitial lung disease. Ther Adv Musculoskelet Dis. 2022;14:1759720x221074457. doi:10.1177/1759720x221074457
165. Park J, Choi J, Cho I, Sheen YY. Radiotherapy-induced oxidative stress and fibrosis in breast cancer are suppressed by vactosertib, a novel, orally bioavailable TGF-β/ALK5 inhibitor. Sci Rep. 2022;12:16104. doi:10.1038/s41598-022-20050-9
166. Khalil H, Kanisicak O, Prasad V, et al. Fibroblast-specific TGF-β-Smad2/3 signaling underlies cardiac fibrosis. J Clin Invest. 2017;127:3770–3783. doi:10.1172/jci94753
167. Maślińska M, Paradowska-Gorycka A, Mańczak M, Kostyra-Grabczak K, Kwiatkowska B. The level of TGF-β in sera of patients with primary Sjögren’s syndrome. Reumatologia. 2019;57:309–314. doi:10.5114/reum.2019.91276
168. Sisto M, Ribatti D, Lisi S. SMADS-Mediate Molecular Mechanisms in Sjögren’s Syndrome. Int J Mol Sci. 2021;22. doi:10.3390/ijms22063203.
169. Ma D, Feng Y, Lin X. Immune and non-immune mediators in the fibrosis pathogenesis of salivary gland in Sjögren’s syndrome. Front Immunol. 2024;15:1421436. doi:10.3389/fimmu.2024.1421436
170. Zou Y, Xiao W, Liu D, et al. Human umbilical cord mesenchymal stem cells improve disease characterization of Sjogren’s syndrome in NOD mice through regulation of gut microbiota and Treg/Th17 cellular immunity. Immun Inflamm Dis. 2024;12:e1139. doi:10.1002/iid3.1139
171. Zhang X, Yun JS, Han D, Yook JI, Kim HS, Cho ES. TGF-β Pathway in Salivary Gland Fibrosis. Int J Mol Sci. 2020;21. doi:10.3390/ijms21239138.
172. Maher TM. Interstitial Lung Disease: a Review. JAMA. 2024;331:1655–1665. doi:10.1001/jama.2024.3669
173. Kondoh Y, Bando M, Kawahito Y, Sato S, Suda T, Kuwana M. Identification and management of interstitial lung disease associated with systemic sclerosis (SSc-ILD), rheumatoid arthritis (RA-ILD), and polymyositis/dermatomyositis (PM/DM-ILD): development of expert consensus-based clinical algorithms. Expert Rev Respir Med. 2024;18:447–456. doi:10.1080/17476348.2024.2374910
174. Cohen ML, Brumwell AN, Ho TC, et al. A fibroblast-dependent TGF-β1/sFRP2 noncanonical Wnt signaling axis promotes epithelial metaplasia in idiopathic pulmonary fibrosis. J Clin Invest. 2024;134. doi:10.1172/jci174598
175. Gao P, Lu Y, Tang K, et al. Ficolin-1 ameliorates pulmonary fibrosis via directly binding to TGF-β1. J Transl Med. 2024;22:1051. doi:10.1186/s12967-024-05894-1
176. Tomos I, Kanellopoulou P, Nastos D, Aidinis V. Pharmacological targeting of ECM homeostasis, fibroblast activation and invasion for the treatment of pulmonary fibrosis. Expert Opin Ther Targets. 2025;29:43–57. doi:10.1080/14728222.2025.2471579
177. Tsukui T, Wolters PJ, Sheppard D. Alveolar fibroblast lineage orchestrates lung inflammation and fibrosis. Nature. 2024;631:627–634. doi:10.1038/s41586-024-07660-1
178. Zhu L, Fu X, Chen X, Han X, Dong P. M2 macrophages induce EMT through the TGF-β/Smad2 signaling pathway. Cell Biol Int. 2017;41:960–968. doi:10.1002/cbin.10788
179. Raghu G, van den Blink B, Hamblin MJ, et al. Effect of Recombinant Human Pentraxin 2 vs Placebo on Change in Forced Vital Capacity in Patients With Idiopathic Pulmonary Fibrosis: a Randomized Clinical Trial. JAMA. 2018;319:2299–2307. doi:10.1001/jama.2018.6129
180. Liu S, Popowski KD, Eckhardt CM, et al. Inhalable Hsa-miR-30a-3p Liposomes Attenuate Pulmonary Fibrosis. Adv Sci. 2025:e2405434. doi:10.1002/advs.202405434.
181. van den Blink B, Dillingh MR, Ginns LC, et al. Recombinant human pentraxin-2 therapy in patients with idiopathic pulmonary fibrosis: safety, pharmacokinetics and exploratory efficacy. Eur Respir J. 2016;47:889–897. doi:10.1183/13993003.00850-2015
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.