Back to Journals » OncoTargets and Therapy » Volume 11
The expression level and prognostic value of HIPK3 among non-small-cell lung cancer patients in China
Authors Liu Y ![]() , Qian L, Yang J, Huang H, Feng J, Li X, Bian T
, Qian L, Yang J, Huang H, Feng J, Li X, Bian T ![]() , Ke H, Liu J, Zhang J
, Ke H, Liu J, Zhang J
Received 28 February 2018
Accepted for publication 2 September 2018
Published 25 October 2018 Volume 2018:11 Pages 7459—7469
DOI https://doi.org/10.2147/OTT.S166878
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jianmin Xu
Yifei Liu,1,* Li Qian,1,* Juanjuan Yang,1 Hua Huang,1 Jia Feng,1 Xiaoli Li,1 Tingting Bian,1 Honggang Ke,2 Jian Liu,3 Jianguo Zhang1
1Department of Pathology, Affiliated Hospital of Nantong University, Nantong, Jiangsu, China; 2Department of Thoracic Surgery, Affiliated Hospital of Nantong University, Nantong, Jiangsu, China; 3Department of Chemotherapy, Affiliated Hospital of Nantong University, Nantong, Jiangsu, China
*These authors contributed equally to this work
Background: Lung cancer is one of the most common malignancies in the world and is at the forefront of causes of all cancer deaths. Identification of new prognostic predictors or therapeutic targets might improve a patient’s survival rate.
Purpose: The Homeodomain interacting protein kinases (HIPKs) function as modulators of cellular stress responses and regulate cell differentiation, proliferation and apoptosis, but the function of HIPK3 is remain unknown.
Patients and methods: We used quantitative real-time polymerase chain reaction (qRT-PCR) and Western blotting methods to detective the expression of HIPK3. A total of 206 samples were obtained from patients and Immunochemical evaluation to determine HIPK3 protein expression. HIPK3 protein levels in in non-small cell lung cancer (NSCLC) were correlated with the clinical characteristics of patients and their 5-year survival rate. In addition, HIPK3 knockdown by specific RNAi promoted cell proliferation, migration, and invasion in A549 and HCC827 cancer cell lines.
Results: The quantitative real-time polymerase chain reaction (qRT-PCR) and Western blotting methods to demonstrate that HIPK3 expression was significantly down-regulated in non-small cell lung cancer (NSCLC) tissues compared with that in normal lung tissues. At the same time, the results of immunohistochemistry assays showed that low expression of HIPK3 was significantly associated with pathology grade; tumor, node, and metastases (TNM) stage; lymph node metastasis; Ki-67 expression; and the 5-year survival rate in NSCLC patients. Univariate analysis revealed that HIPK3 expression, Ki-67 expression, tumor diameter, TNM stage, and age were significantly associated with a poor prognosis. The multivariable analysis illustrated that HIPK3, tumor diameter, TNM, Ki-67 expression, and age had effects on the overall survival of NSCLC patients independently. Kaplan-Meier survival curves revealed that NSCLC patients with a lower HIPK3 expression had a poorer prognosis. In addition, in vivo results also confirmed that HIPK3 over-expression could inhibit tumor growth.
Conclusion: Our findings confirmed that low expression of HIPK3 in NSCLC tissues was significantly correlated with poor survival rates after curative resection. HIPK3 could potentially be used as a valuable biomarker in the prognosis of the survival of NSCLC patients.
Keywords: HIPK3, non-small-cell lung cancer, immunohistochemistry, tumor suppressor gene, prognosis, biomarker
Plain language summary
The expression of HIPK3 mRNA and protein level was significantly downregulated in human non-small-cell lung cancer (NSCLC). Low expression of HIPK3 was significantly associated with pathology grade, TNM stage, lymph node metastasis, Ki-67 expression, and 5-year survival rate in NSCLC patients. HIPK3 silencing promoted invasion and metastasis in NSCLC. HIPK3 could be potentially used as a valuable biomarker for predicting the prognosis of NSCLC patients.
Introduction
According to the statistical analysis released by the American Cancer Society, in 2018, there were ~234,030 newly diagnosed patients with lung cancer, accounting for 13.5% of all newly diagnosed malignancies. In addition, ~154,050 people died from lung cancer each year, accounting for 25.3% of all cancer-related deaths.1 China has not escaped this worldwide trend, and the data being gathered in the country on the cause of death have ranked lung cancer as the first among all malignant tumor deaths for 7 consecutive years. For example, in Beijing, the number of deaths due to lung cancer nearly tripled between 1974 and 2003.
In general, lung cancer cells are classified into two types – non-small-cell lung cancer (NSCLC) and small-cell lung cancer (SCLC).2 NSCLC accounts for 80%–85% of all lung cancers.3 The treatment for lung cancer is mainly surgery in combination with chemotherapy, radiotherapy, immunotherapy, and so on. Despite the advances in cancer treatments, the 5-year survival rate for lung cancer remains <18%4,5 mainly due to the failure of early detection and diagnosis of the disease. In ~85% of patients, the primary lesion will metastasize and invade lymph nodes or other organs at the late stage; at this stage, the treatment is not effective and the prognosis is poor. Therefore, early diagnosis and a comprehensive treatment to prevent and control NSCLC metastasis are critical to reduce mortality.
FAS/HIPK, a member of the HIPK family of genes, was discovered in 1998 by Kim et al6 through the double hybridization of yeast and by obtaining the main cDNA clone of this family member. The family comprises HIPK1, HIPK2, HIPK3, and HIPK4, of which, we focus on HIPK3 in this study.
There have been few studies on HIPK3, especially on its association with lung cancer. In this study, we confirmed a lower expression of HIPK3 in NSCLC tissues than in normal lung tissues and demonstrated that HIPK3 played a role in the proliferation, migration, and invasion of these cancer cells. Our findings suggested that HIPK3 had potential clinical applications as a prognostic predictor or therapeutic target in NSCLC.
Materials and methods
Patients and tissue samples
A total of 206 samples were obtained from patients who had undergone lung resection without immunotherapy or radiation therapy at the Thoracic Surgery Department of the Affiliated Hospital of Nantong University (Nantong, China) between May 2008 and July 2010. Quantitative real-time qRT-PCR was used to investigate 30 patients who underwent curative resection between January 2015 and August 2016 at the same hospital. Follow-up of all patients was completed by May 2017. The tissues that were previously pathologically determined to be NSCLC and matching non-cancerous tissues were obtained from each patient. The procedure was approved by the research ethics committee of the Affiliated Hospital of Nantong University (Nantong, China), and the written informed consent was obtained from each patient.
qRT-PCR analysis
For qRT-PCR analysis, total RNA was isolated from NSCLC samples using TRIzol (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s protocol. Reverse transcription and PCR were performed using a High Capacity cDNA Reverse Transcription Kit and QuantiTect SYBR Green PCR Kit (Thermo Fisher Scientific). The primer sequences for the genes were defined as follows:
HIPK3: forward 5′-ACATTGGAAGAGCATGAGGCAGAGA-3′; reverse 5′-CTGCTGAAAAGCATCACCACAACCA-3′ | |
GAPDH: forward 5′-GCAGGGGGGAGCCAAAAGGGT-3′; reverse 5′-TGGGTGGCAGTGATGGCATGG-3′. |
Immunohistochemical (IHC) evaluation
For statistical analyses, the number of cells showing SYBR green staining on the nuclei and cytoplasm in 10 representative microscopic fields were counted, and the percentage of positive cells was calculated. A semiquantitative assessment of the IHC reactions for HIPK3 was used to score staining results as follows: 0 (negative, ≤5% of cells), 1+ (low, 6%–25%), 2+ (intermediate, 26%–50%), and 3+ (high, ≥51%). The count was recorded as the percentage of positive cells. In one-half of the samples, staining was repeated three times to avoid potential technical errors. The evaluation procedures were independently performed by three experienced pathologists using a multi-head microscope (Precise Instrument Co., Ltd., Beijing, China) to reach a consensus.
In addition to the aforementioned method, another quantitative evaluation method was used. HIPK3 sections were scanned by NanoZoomer slide scanners (NanoZoomer-XR C12000; Hamamatsu, Japan) and observed with NDP.view software (NDP.view2 U12388-01; Hamamatsu). The tumor area was determined as a 20× field (545×577 μm2) viewed by NDP.view in the center. The number of HIPK3 cells was counted in four hotspots (20×) using a computer-automated method (Imagepro plus 6.0; Media Cybernetics Inc., Bethesda, MD, USA), and then mean densities were calculated. The cutoff value of the HIPK3 cell counts was determined by the 5-year survival-specific receiver operating characteristic curve analysis according to the Youden’s index. Then, a low HIPK3 cell count was not more than the cutoff value and a high HIPK3 cell count was greater than the cutoff value.
Cell culture and transfection
A549 and HCC827 cells were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured in Dulbecco’s Modified Eagle’s Medium supplemented with 10% FBS. Cells were transfected with HIPK3 siRNA (Genepharma, Suzhou, China) and their corresponding negative control (siRNA ctrl) at a final concentration of 50 nmol/L using Lipofectamine 2000 Transfection Reagent (Thermo Fisher Scientific) according to the manufacturer’s instructions.
Cell invasion and cell migration
Transwell membranes (8 μm pore size) on 24-well plates were coated with Matrigel (BD Biosciences, San Jose, CA, USA) and used for cell invasion assays. Equal numbers (1×105) of nontransfected cells and cells transfected with siRNA-targeting HIPK3 (siR-HIPK3) or HIPK3 vector were plated in separate wells. The cells were cultured overnight in a serum-free medium before being trypsinized and resuspended at a density of 2×105 cells/mL in a medium containing 2% FBS. The cells were then added to the upper chamber of the incubator with medium containing 20% FBS as the chemoattractant in the lower chamber. Medium containing 2% FBS was added to the lower chamber and used as the control. Matrigel and the cells remaining in the upper chamber were removed using cotton swabs following incubation for 24 hours. The cells on the lower surface of the membrane were fixed with formaldehyde solution and stained with staining solution. The cells in at least five random microscopic fields (×200) were counted and photographed. For the migration assay, the transfected cells were plated and incubated until the formation of 80%–90% confluent monolayer. AP-20 micropipette tips were then used to mimic a scratch wound. The ratio of migratory capacity was determined by measuring the width of the scratch at 0 and 24 hours using bright-field microscopy.
Tumor xenograft model in nude mice
The nude mouse xenograft model was developed, which was xenografted with a human lung cancer cell line, NSCLC A549 cell. This mice model was obtained from Shanghai Animal Experimental Center. Mice were randomly divided into four groups, with four mice in each group, namely the pcDNA3.1 control group (group A), normal saline control group (group B), HIPK3-shRNA group (group C), and HIPK3 overexpression group (group D). All supplied mice were female and weighed 19–20 g. All treatments were administered for 4 weeks.
Western blot analysis
For Western blotting, 50 μg protein was separated by SDS-PAGE and transferred onto polyvinylidene difluoride (PVDF) membranes. The membranes were blocked with 5% nonfat milk for 1 hour and incubated overnight at 4°C with their corresponding primary antibodies (1:1,000 dilution in 5% nonfat milk), followed by incubation with secondary antibodies conjugated with horseradish peroxidase (HRP; 1:8,000 dilution in 5% nonfat milk) for 2 hours at room temperature. The membranes were then detected using an electrogenerated chemiluminescence kit (Thermo Fisher Scientific).
Statistical analyses
All computations were performed using SPSS, Version 24.0 (IBM Corporation, Armonk, NY, USA). All values were expressed as the mean±standard error. HIPK3 expression and clinicopathological features were analyzed using Pearson’s chi-squared test. Univariate and multivariate analyses were performed using the Cox regression model to test independent prognostic factors, and 95% CI was recorded for each marker. Kaplan–Meier analysis and the log-rank test were used to analyze the survival rate. A P-value <0.05 was considered statistically significant.
Results
Decreased HIPK3 expression in NSCLC tissues
The protein and mRNA levels of HIPK3 expression were quantified using Western blotting and qRT-PCR, respectively, in NSCLC tissues and normal tissues. As illustrated in Figure 1, both Western blotting and qRT-PCR showed a visible decrease in HIPK3 expression levels in NSCLC tissues compared with that in normal tissues.
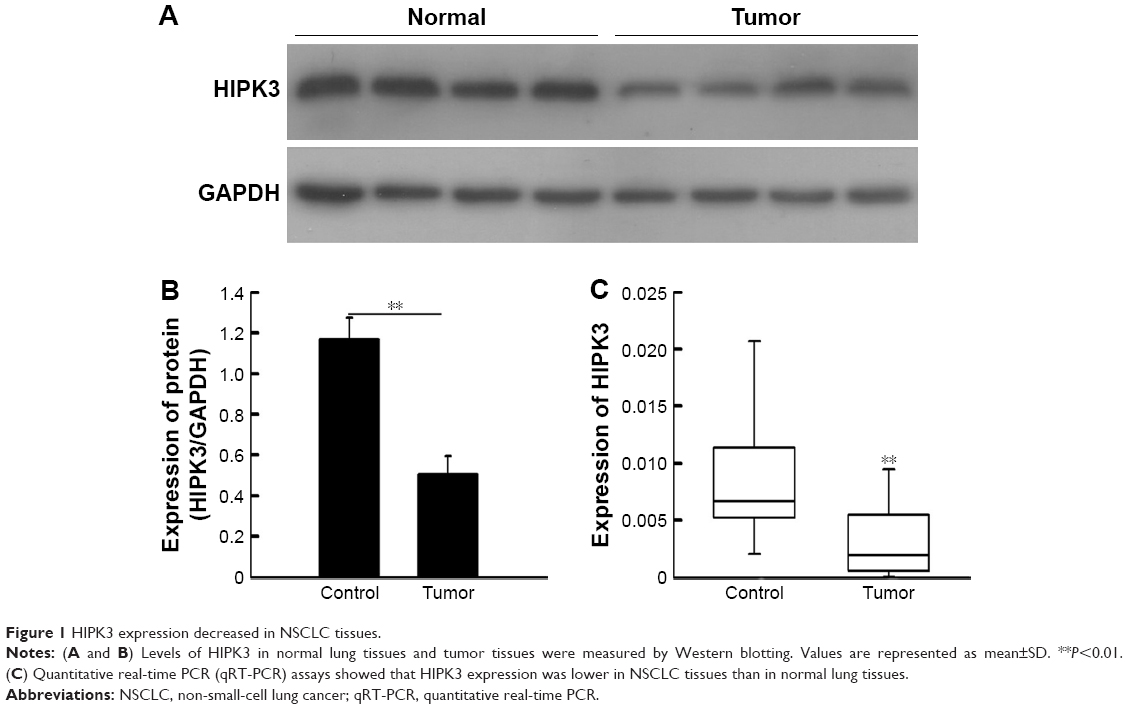 | Figure 1 HIPK3 expression decreased in NSCLC tissues. |
Decreased HIPK3 expression in NSCLC tissues was associated with poor clinicopathological characteristics
IHC was performed in 206 NSCLC tissue samples to investigate the HIPK3 protein expression profile. The cytoplasm and nucleus of the carcinoma cells showed positive immunostaining (Figure 2). Table 1 shows the relationship between the expression of HIPK3 and the clinicopathological characteristics of the tissue samples from 206 NSCLC patients. We identified that HIPK3 protein expression was significantly associated with pathology grade (P=0.004), TNM stage (P=0.002), lymph node metastasis (P=0.010), Ki-67 expression (P=0.015), and 5-year survival rate in NSCLC patients (P=0.001). However, there was no correlation between HIPK3 protein expression and other prognostic factors such as age, sex, tumor diameter, smoking status, and histological type.
 | Figure 2 Representative images of IHC staining of HIPK3 expression in NSCLC tissue samples and benign tumors. |
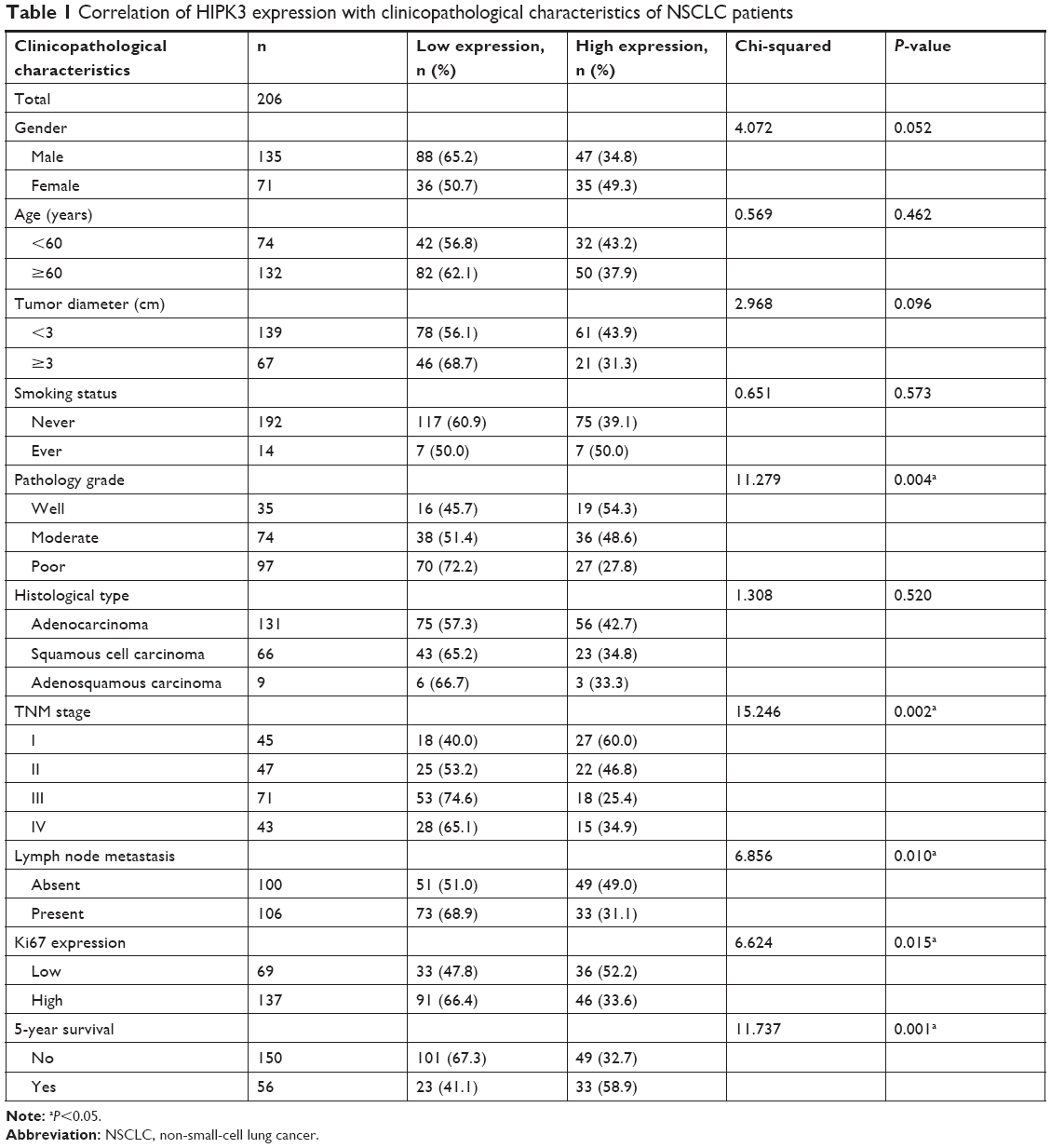 | Table 1 Correlation of HIPK3 expression with clinicopathological characteristics of NSCLC patients |
Decreased HIPK3 protein expression in NSCLC predicted poor survival
Univariate and multivariate analyses were used with the Cox proportional hazards model to identify related prognostic factors for overall survival. The univariate analysis revealed that the following variables were significantly associated with a poor prognosis: HIPK3 expression (P=0.001), Ki-67 expression (P=0.027), tumor size (P<0.001), TNM stage (P<0.001), and age (P=0.029). To verify the accuracy of the results, multivariate analysis was performed using the significant (P<0.05) variables from the univariate analysis. The multivariate analysis revealed that the HIPK3 (P<0.001), tumor size (P<0.001), TNM stage (P<0.001), Ki-67 expression (P=0.027), and age (P=0.006) had independent prognostic value on the overall survival of NSCLC patients (Table 2). Kaplan–Meier survival curves revealed that a poorer prognosis was found in NSCLC patients with lower HIPK3 expression levels (Figure 3).
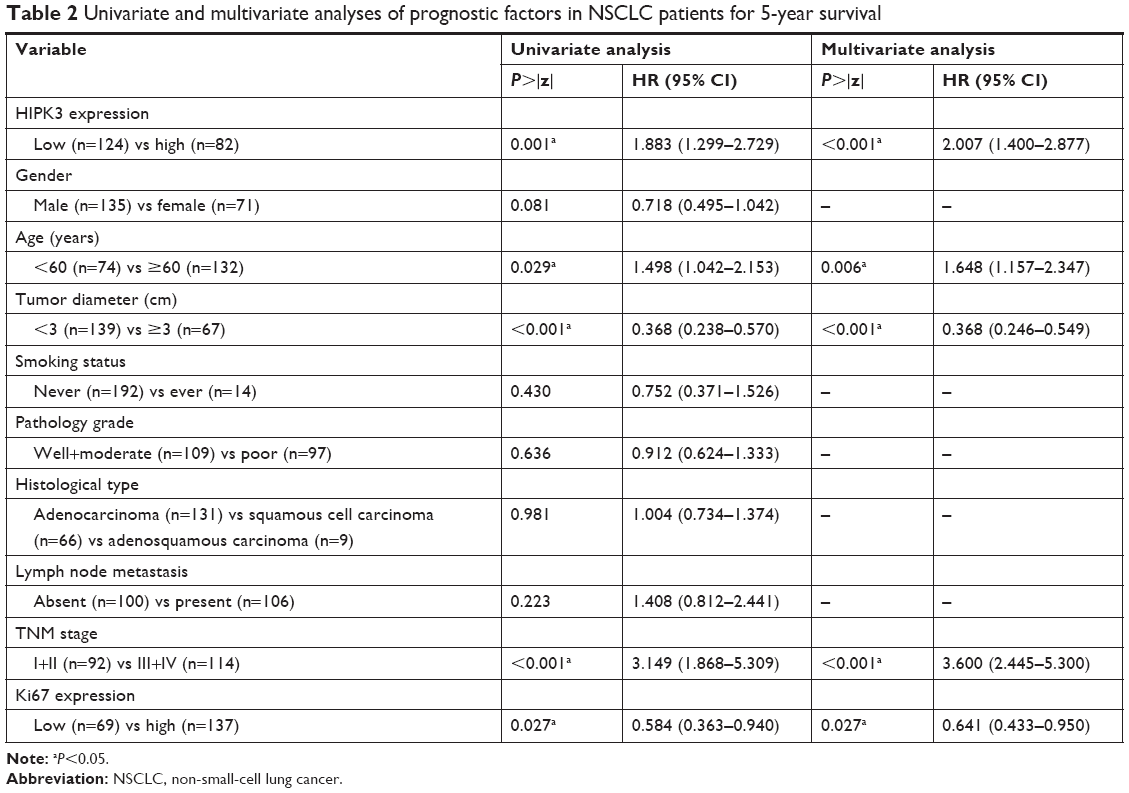 | Table 2 Univariate and multivariate analyses of prognostic factors in NSCLC patients for 5-year survival |
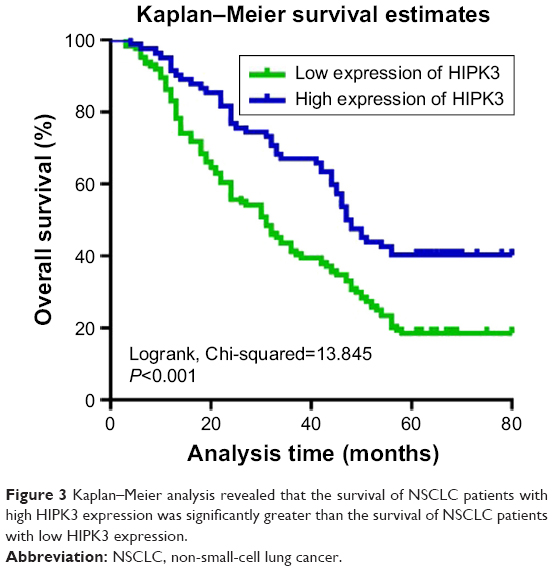 | Figure 3 Kaplan–Meier analysis revealed that the survival of NSCLC patients with high HIPK3 expression was significantly greater than the survival of NSCLC patients with low HIPK3 expression. |
HIPK3 participated in multiple biological processes in NSCLC cells
To further verify the expression of HIPK3 in NSCLC cells in vitro, siR-HIPK3 for knocking down HIPK3 mRNA was synthesized. Western blot analysis showed that siR-HIPK3 reduced HIPK3 protein levels in A549 as shown in Figure 4A and B. As indicated by the results, siR-HIPK3 promoted the migratory ability and invasive capability of the NSCLC cell line. siR-HIPK3 further confirmed the result by enhancing the migratory ability and the invasive capability of both cell lines (Figure 4C and D). To further verify the results, we repeated the experiments in another lung cancer cell line, HCC827. The aforementioned results were confirmed again in HCC827 (Figure 5). Taken together, the results showed that HIPK3 was negatively expressed in the progression and metastasis of NSCLC.
 | Figure 4 siR-HIPK3 enhanced the migration and invasion of A549 cells. |
 | Figure 5 siR-HIPK3 promoted the migration and invasion of HCC827 cells. |
HIPK3 inhibited the proliferation of NSCLC cells in vivo
We further studied the effect of HIPK3 on tumor volume in vivo, and the results showed that the overexpression of HIPK3 inhibited tumor growth and the overexpression of HIPK3-shRNA promoted tumor growth (Figure 6A and B).
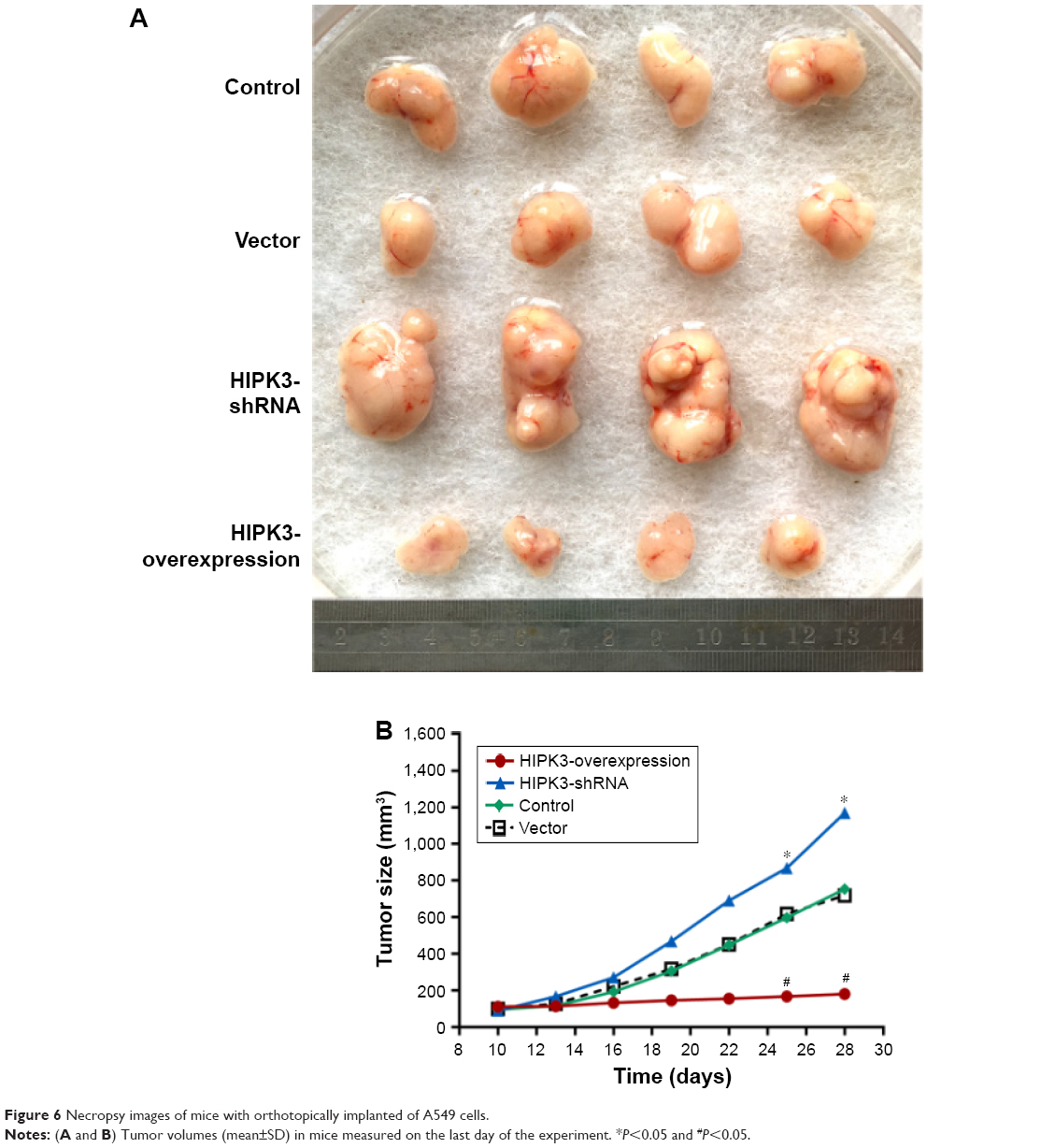 | Figure 6 Necropsy images of mice with orthotopically implanted of A549 cells. |
Discussion
The function of HIPKs is to modulate cellular stress responses and regulate cell differentiation, proliferation, and apoptosis.7 The HIPK family genes comprises HIPK1, HIPK2, and HIPK3, which share similar domain-frustrated structures on serine and threonine residues. Some studies have found that the expression of HIPKs was located mainly in the nucleus and enhanced the inhibitory activity of the natural killer (NK) protein family.8 Studies by Kim et al6 found that HIPKs were involved in a variety of cellular processes and played a role by mediating the phosphorylation of several important protein molecules. They represented a family of highly conserved kinases and were involved in cellular processes including regulation of cell death, proliferation, and differentiation. In addition, it has been reported that these kinases are involved in the regulation of cellular responses to DNA damage.7 The HIPK family not only regulates other proteins in the nucleus, but also regulates those in the cytoplasm and cell membrane and can participate in biological processes, such as cell proliferation, differentiation, and apoptosis, by affecting the phosphorylation of corresponding proteins. The site and mode of action depend on the conditions that the protein encounters after it has been removed from the nucleus.9,10
HIPK1 protein is involved in the biological processes of cell proliferation, activation, and apoptosis by regulating the phosphorylation of many proteins and activation of some signaling pathways.11,12 Other studies have found that HIPK1 protein plays a role in promoting apoptosis to reduce tumor progression. These findings also showed that this protein depends on partner proteins to function. For example, under certain conditions, HIPK1 phosphorylates its partner protein Daxx to redistribute within the cytoplasm by influencing localization and transcriptional repression activities in Daxx cells.13,14 Studies have shown that HIPK1 plays an important role in oncogenesis and that the incidence or size of tumor formation in HIPK1 knockout mice is significantly reduced. This suggests a phenomenon associated with DNA damage–induced apoptosis. These data also suggest that HIPK1 is involved, to some extent, in a reaction to repair the damaged DNA, a process that might be initiated through its interaction with p53.14
As a tumor suppressor gene, HIPK2 is involved in the suppression of the occurrence and metastasis of several tumors. Nodale et al15 found that HIPK2 inhibited the spread of human breast cancer cells by down-regulating vimentin. Similarly, D’Orazi et al16 used RNA interference (RNAi) to study the role of HIPK2 in prostaglandin biosynthesis and found that HIPK2 was inhibited by cytosolic phospholipase A2 (cPLA2) prostaglandin E2 (PGE2) production, thereby inhibiting the occurrence of human colon tumors. Furthermore, Guang et al17 revealed that HIPK2 could inhibit P-catenin–mediated transcription, epidermal stem cell expansion, and skin tumors.
HIPK2 is the most studied member of the HIPK family of genes and is characterized by DNA damage response kinases that have both pro-apoptotic and anti-apoptotic functions to combat genotoxic and oxidative cell damage.18–22 Moreover, He et al23 showed that HIPK4 not only had less sequence homology with HIPKs 1, 2, and 3, but also had a high degree of sequence homology, suggesting that this kinase is highly conserved among species.
In 1998, Kim et al.6 first reported on HIPK3 and that HIPK3 mRNA is widely found in human and mammalian tissues. Northern blot analysis revealed that HIPK3 is expressed as a ~7.5 kb tRNA, whereas in humans, HIPK3 also contains an additional 4.4 kb transcript on chromosome seven. Young et al24 labeled HIPK3 protein with green fluorescent protein (GFP) and observed it in living cells. The GFP spots were first observed in the nucleus and later in the cytoplasm, indicating that HIPK3 protein is first present in the nucleus as a nuclear protein kinase. The HIPK protein family is highly phosphorylated during apoptosis, while purified HIPK3 can phosphorylate NK domain homology transcription factor, suggesting that its family is a group of homologous domain-transforming phosphorylation kinases. Because NK cells are one of the major cells involved in FAS-mediated apoptosis, HIPK3 is an important factor in FAS-mediated apoptosis and is a tumor suppressor gene. In our current study, as illustrated in Figure 1, both Western blotting and qRT-PCR showed a visible decrease in HIPK3 expression levels in NSCLC tissues compared with that in the normal tissue. This result was consistent with the previous research results. Veronique et al25 cloned full-length mouse and human HIPK3 homology and found that 90% of the amino acid sequences were the same. A gene study found that an intact HIPK3 protein molecule was consisted of an NH2-terminal kinase domain and a conserved PEST sequence, which had a COOH-terminus that could coincide with the FADD domain on FAS.
In 2004, Curtin et al26 studied the role of HIPK3 in the apoptotic prostate cancer cell line DU-145. When JNK inhibitor SP600125 was added to DU-145 and detected by Western blotting and flow cytometry, they found that as the JNK levels decreased, HIPK3 and FAS-mediated apoptosis also decreased. After removing the JNK inhibitor, FAS-mediated apoptosis was significantly increased, HIPK3 expression also increased, and there was a change in Caspase-3 expression levels. These results indicated that HIPK3 was associated with FAS-mediated apoptosis. Moller27 also reported HIPK3 expression in the apoptotic human osteosarcoma cell line U20S; however, the clinical application of HIPK3, especially the study of HIPK3 in clinical NSCLC specimens, has not been reported.
In our current study, IHC results demonstrated that low HIPK3 protein expression was significantly associated with pathology grade, TNM stage, lymph node metastasis, Ki-67 expression, and the 5-year survival rate of NSCLC patients. The multivariable analysis revealed that HIPK3, tumor size, TNM stage, Ki-67 expression, and age had independent prognostic effects on the overall survival of patients with NSCLC. Kaplan–Meier survival curves showed that NSCLC patients with higher HIPK3 protein expression had a better prognosis. In addition, HIPK3 knockdown using specific siRNA enhanced the migration and invasion of A549 and HCC827 cells. In vivo experiments in nude mice also demonstrated that HIPK3 over-expression inhibited tumor growth, whereas in contrast, HIPK3-shRNA promoted tumor growth in vivo.
Conclusion
Our findings confirmed that low expression of HIPK3 in NSCLC tissues was significantly correlated with poor survival rates after curative resection. HIPK3 could be used as a valuable biomarker for predicting the prognosis for NSCLC patients.
Ethical approval
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional research committee and with the 1964 Declaration of Helsinki and its later amendments or comparable ethical standards.
Informed consent
Informed consent was obtained from all individual participants included in the study.
Acknowledgments
This study was supported by grants from Six Talent Peaks Project in Jiangsu Province (No WSN-059), the Science Foundation of Nantong City (No MS12015007), Scientific Research Topic of Jiangsu provincial Health and Family Planning Commission (No H201626), and Key Talents of Medical Science in Jiangsu Province (No QNRC2016682).
Disclosure
The authors report no conflicts of interest in this work.
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018, CA: a cancer journal for clinicians. 2018;68:7–30. | ||
Ulahannan SV, Brahmer JR. Antiangiogenic agents in combination with chemotherapy in patients with advanced non-small cell lung cancer. Cancer Invest. 2011;29(4):325–337. | ||
Ramalingam SS, Owonikoko TK, Khuri FR. Lung cancer: new biological insights and recent therapeutic advances. CA Cancer J Clin. 2011;61(2):91–112. | ||
Tanoue LT, Tanner NT, Gould MK, Silvestri GA. Lung cancer screening. Am J Respir Crit Care Med. 2015;191(1):19–33. | ||
Siegel R, Ma J, Zou Z, Jemal A. Cancer statistics, 2014. CA Cancer J Clin. 2014;64(1):9–29. | ||
Kim YH, Choi CY, Lee SJ, Conti MA, Kim Y. Homeodomain-interacting protein kinases, a novel family of co-repressors for homeodomain transcription factors. J Biol Chem. 1998;273(40):25875–25879. | ||
Rinaldo C, Siepi F, Prodosmo A, Soddu S. HIPKs: Jack of all trades in basic nuclear activities. Biochim Biophys Acta. 2008;1783(11):2124–2129. | ||
Chang WH, Liu TC, Yang WK, et al. Amiloride modulates alternative splicing in leukemic cells and resensitizes Bcr-AblT315I mutant cells to imatinib. Cancer Res. 2011;71(2):383–392. | ||
van der Laden J, Soppa U, Becker W. Effect of tyrosine autophosphorylation on catalytic activity and subcellular localisation of homeodomain-interacting protein kinases (HIPK). Cell Commun Signal. 2015;13(3):3. | ||
Akaike Y, Kuwano Y, Nishida K, et al. Homeodomain-interacting protein kinase 2 regulates DNA damage response through interacting with heterochromatin protein 1γ. Oncogene. 2015;34(26):3463–3473. | ||
Rey C, Soubeyran I, Mahouche I, et al. HIPK1 drives p53 activation to limit colorectal cancer cell growth. Cell cycle (Georgetown, Tex.). 2013;12:1879–1891. | ||
Li X, Zhang R, Luo D, et al. Tumor necrosis factor alpha-induced desumoylation and cytoplasmic translocation of homeodomain-interacting protein kinase 1 are critical for apoptosis signal-regulating kinase 1-JNK/p38 activation. J Biol Chem. 2005;280(15):15061–15070. | ||
Ecsedy JA, Michaelson JS, Leder P. Homeodomain-interacting protein kinase 1 modulates Daxx localization, phosphorylation, and transcriptional activity. Mol Cell Biol. 2003;23(3):950–960. | ||
Kondo S, Lu Y, Debbas M, et al. Characterization of cells and gene-targeted mice deficient for the p53-binding kinase homeodomain-interacting protein kinase 1 (HIPK1). Proc Natl Acad Sci U S A. 2003;100(9):5431–5436. | ||
Nodale C, Sheffer M, Jacob-Hirsch J, et al. HIPK2 downregulates vimentin and inhibits breast cancer cell invasion. Cancer Biol Ther. 2012;13(4):198–205. | ||
D’Orazi G, Sciulli MG, di Stefano V, et al. Homeodomain-interacting protein kinase-2 restrains cytosolic phospholipase A2-dependent prostaglandin E2 generation in human colorectal cancer cells. Clin Cancer Res. 2006;12(3 Pt 1):735–741. | ||
Wei G, Ku S, Ma GK, et al. HIPK2 represses beta-catenin-mediated transcription, epidermal stem cell expansion, and skin tumorigenesis. Proc Natl Acad Sci U S A. 2007;104(32):13040–13045. | ||
D’Orazi G, Cecchinelli B, Bruno T, et al. Homeodomain-interacting protein kinase-2 phosphorylates p53 at Ser 46 and mediates apoptosis. Nat Cell Biol. 2002;4(1):11–19. | ||
Hofmann TG, Möller A, Sirma H, et al. Regulation of p53 activity by its interaction with homeodomain-interacting protein kinase-2. Nat Cell Biol. 2002;4(1):1–10. | ||
Zhang Q, Yoshimatsu Y, Hildebrand J, Frisch SM, Goodman RH. Homeodomain interacting protein kinase 2 promotes apoptosis by downregulating the transcriptional corepressor CtBP. Cell. 2003;115(2):177–186. | ||
Zhang J, Pho V, Bonasera SJ, et al. Essential function of HIPK2 in TGFbeta-dependent survival of midbrain dopamine neurons. Nat Neurosci. 2007;10(1):77–86. | ||
Hofmann TG, Glas C, Bitomsky N. HIPK2: A tumour suppressor that controls DNA damage-induced cell fate and cytokinesis, BioEssays: news and reviews in molecular. Cellular and developmental biology. 2013;35:55–64. | ||
He Q, Shi J, Sun H, An J, Huang Y, Sheikh MS. Characterization of Human Homeodomain-interacting Protein Kinase 4 (HIPK4) as a Unique Member of the HIPK Family. Molecular and Cellular Pharmacology. 2010;2:61–68. | ||
Higashimoto Y, Saito S, Tong XH, Hong A, Sakaguchi K, Appella E, Anderson CW. Human p53 is phosphorylated on serines 6 and 9 in response to DNA damage-inducing agents. J Biol Chem. 2000;275:23199–23203. | ||
Venables JP, Bourgeois CF, Dalgliesh C, Kister L, Stevenin J, Elliott DJ. Up-regulation of the ubiquitous alternative splicing factor Tra2beta causes inclusion of a germ cell-specific exon. Human molecular genetics. 2005;14:2289–2303. | ||
Curtin JF, Cotter TG. JNK regulates HIPK3 expression and promotes resistance to Fas-mediated apoptosis in DU 145 prostate carcinoma cells. J Biol Chem. 2004;279:17090–17100. | ||
Moller A, Sirma H, Hofmann TG, et al. PML is required for homeodomain-interacting protein kinase 2 (HIPK2)-mediated p53 phosphorylation and cell cycle arrest but is dispensable for the formation of HIPK domains. Cancer Res. 2003;63:4310–4314. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.