")
Back to Journals » International Journal of Nanomedicine » Volume 14
The effects of graphene and mesenchymal stem cells in cutaneous wound healing and their putative action mechanism
Authors Lasocka I , Jastrzębska E , Szulc-Dąbrowska L, Skibniewski M , Pasternak I , Hubalek Kalbacova M , Skibniewska EM
Received 15 October 2018
Accepted for publication 9 January 2019
Published 1 April 2019 Volume 2019:14 Pages 2281—2299
DOI https://doi.org/10.2147/IJN.S190928
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Thomas Webster
Iwona Lasocka,1 Elżbieta Jastrzębska,2 Lidia Szulc-Dąbrowska,3 Michał Skibniewski,4 Iwona Pasternak,5 Marie Hubalek Kalbacova,6,7 Ewa M Skibniewska1
1Department of Animal Environment Biology, Faculty of Animal Science, Warsaw University of Life Sciences, Warsaw, Poland; 2Department of Medical Biotechnology, Faculty of Chemistry, Warsaw University of Technology, Warsaw, Poland; 3Department of Preclinical Sciences, Faculty of Veterinary Medicine, Warsaw University of Life Sciences, Warsaw, Poland; 4Department of Morphological Sciences, Faculty of Veterinary Medicine, Warsaw University of Life Sciences, Warsaw, Poland; 5Faculty of Physics, Warsaw University of Technology, Warsaw, Poland; 6Institute of Pathological Physiology, 1st Faculty of Medicine, Charles University, Prague, Czech Republic; 7Biomedical Center, Faculty of Medicine in Pilsen, Charles University, Pilsen, Czech Republic
Abstract: This study provides a review of the therapeutic potential of graphene dressing scaffolds and mesenchymal stem cells (MSCs) and their synergistic effects with respect to cutaneous wound healing. This study also considers their putative action mechanism based on the antibacterial, immunomodulating, angiogenic, matrix remodeling effects of materials belonging to the graphene family and MSCs during the wound healing process. In addition, this study discusses the cytocompatibility of graphene, its uses as a platform for skin substitutes, the properties it possesses with respect to providing protection against microbial invasion as well as strategies aimed at minimizing the chance of the occurrence of sepsis. MSCs are capable of secreting several factors that exert a therapeutic impact on reparative processes and tissue regeneration. In light of experiments conducted to date, graphene combined with MSCs appears to have the potential to enhance both the wound healing process and infection control at the injury site.
Keywords: graphene, mesenchymal stem cells, wound, healing
Introduction
Graphene, in combination with mesenchymal stem cells (MSCs), provides a potential clinical application for wound healing purposes. A number of strategies have been advanced to date aimed at enhancing and accelerating the closure of injured tissue in cutaneous wounds, one of which consists of the use of dressing materials containing graphene and derivatives thereof. Moreover, in recent years the attention of a large number of research teams has been devoted to therapy employing MSCs. This study suggests that the synergic effect of a combination of these two approaches may potentially assist in the healing of acute and chronic wounds, which presents a major clinical problem in the fields of both veterinary and human medicine, and with concern to which, due to increasing bacterial resistance, local treatment plays an especially important role. Alternative treatments for hard-to-heal wounds include the application of platelet-rich plasma and cell growth factor preparations, vacuum dressings and other dressings that exhibit antibacterial properties. Deepachitra et al1 demonstrated both in vitro (fibroblasts) and in vivo (rats) that graphene oxide (GO) combined with a collagen-fibrin biofilm can be successfully employed as a dressing material. The treatment of local wounds with MSC applications has gained popularity in recent years as a promising approach for the enhancement of tissue regeneration. It is thought that the therapeutic benefit of MSCs lies principally in the various factors that they secrete such as vascular endothelial growth factor (VEGF), EGF, fibroblast growth factor (FGF), keratinocyte growth factor (KGF), insulin-like growth factor (IGF), platelet-derived growth factor (PDGF), TGF-β, prostaglandin E2 (PGE2), hepatocyte growth factor (HGF), tumor necrosis factor-α (TNF-α), interferon λ (IFNλ) and ILs such as IL-4, IL-6 and IL-10.2–9 All these growth factors and cytokines play an important role in the formation of new blood vessels, cell recruitment, immunomodulation and wound closure. Moreover, MSCs promote direct cell differentiation, proliferation and extracellular matrix (ECM) remodeling.10,11
Cell cultures are employed for both the basic research of many biological processes and for medical applications such as tissue engineering. In the first case, they provide a relatively simple experimental model in contrast to tissues that contain numerous differing cell types. Cell cultures allow for the culturing of specific types of cells and facilitate the study of processes such as cell division, the formation of organelles, protein secretion and differentiation into tissue with specific and determined phenotypes. Intracellular mechanisms, which can be investigated in detail, allow for the closer examination of metabolic processes than would otherwise be possible. With respect to the field of tissue engineering, they enable complicated manipulation leading to the creation of new tissue, which can be maintained and cultivated outside the organism of the donor/recipient. Due to rapid cell growth, in vitro experiments provide information on the process under investigation more quickly than do other methods, thus expediting the obtaining of the final product. Thus, this study suggests that graphene, in combination with MSCs, has significant potential with respect to wound healing and infection control applications.
Cutaneous wound healing
Skin makes up the most extensive organ in the body and has numerous functions including protection against microorganisms. Once this natural barrier is damaged due to injury, burns or systemic dysfunction, the risk of infection increases significantly potentially leading to the occurrence of severe general complications including sepsis.
The cutaneous wound healing process is divided into four distinct phases, ie, hemostasis, inflammation, proliferation and tissue remodeling (Figure 1). It involves cells such as platelets, inflammatory cells, epithelial cells, keratinocytes, fibroblasts, a multitude of cytokines, growth factors and other bioactive molecules as well as interactions with ECM components mediated by integrin receptors and adhesive molecules. Chronic wounds are considered to be those that do not heal within 12 weeks of injury, which usually leads to prolonged pathological inflammation;12–15 thus, the development of methods that accelerate the healing of acute and chronic wounds should make up the primary aim of the wound management process.
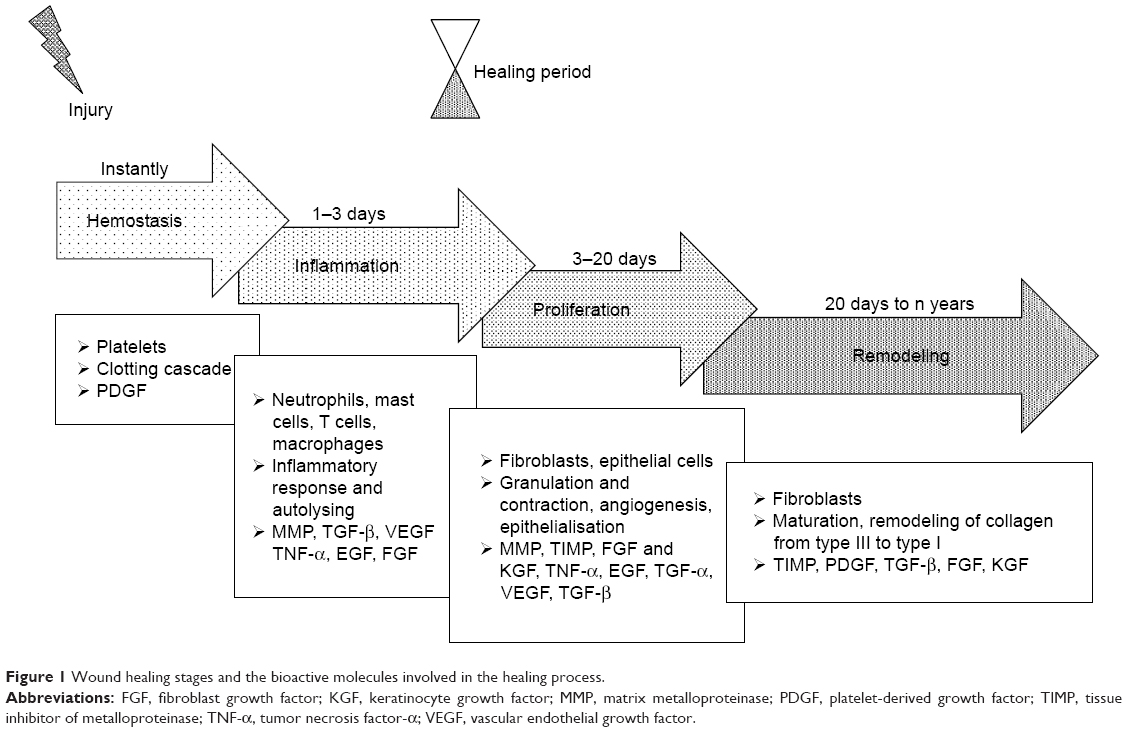 | Figure 1 Wound healing stages and the bioactive molecules involved in the healing process. |
Hemostasis starts immediately following injury due to the constriction of the damaged vessels, which limits the extent of blood loss. This leads to tissue hypoxia and acidosis, which decreases the effect of vasoconstriction and increases the level of vascular permeability for inflammatory cells. Platelets play the most important role at this stage through the formation of a blood clot (coagulation cascade) and the production of multiple signaling molecules such as PDGF, EGF, fibronectin, fibrinogen, histamine, serotonin and the von Willebrand factor.16,17 PDGFs are released from the alpha granules of platelets thus promoting mitogenicity and the chemotaxis of the neutrophils, macrophages, fibroblasts and smooth muscle cells at the wound site.18
The increased infiltration of neutrophils, mast cells, monocytes and T lymphocytes into the wound site occurs during the inflammation stage,19 and TGF-β, TNF-α, EGF, PDGF, VEGF, FGF, IL-1, IL-6, IL-8 and IL-12 are all present in the wound environment at this phase of healing.12,16,19–22 These mediators both control the inflammatory process and modulate epithelialization, collagen accumulation and angiogenesis.20 TGF-β is a potent chemoattractant for immune cells and, as with PDGF, is involved in all the phases of the wound healing process.23,24
First, guided by chemokines, TGF-β and peptides produced by the bacteria present,25 neutrophils begin to migrate into the wound so as to prevent infection.10 However, once the influx of monocytes (the second type of wound-attracted cells) commences, the infiltration of neutrophils begins to decrease. Monocytes are attracted to the wound site by factors such as PDGF and TGF-β as well as by broken-down elastin and collagen products.10 Monocytes undergo phenotypic transformation within the wound so as to form macrophages which are responsible for the further reduction of inflammation and the initiation of the proliferative phase of the healing process.14 The phagocytic role of macrophages, which is based on the removal of destroyed cells and debris from the wound site, is crucial with respect to wound healing. Matrix metalloproteinases (MMPs) released by macrophages, monocytes and lymphocytes are responsible for cleaning the wound of dead and damaged tissue and are secreted under the control of TNF-α, IL-1 and IL-6. Lymphocytes, which appear in the late inflammatory phase, influence both fibroblast proliferation and collagen biosynthesis.12 Oxygen presence is necessary for both the actively proliferating cells and the neutrophil respiratory burst in the wound bed,26 thus requiring the initiation of neovascularization.
The principal cells involved in the proliferation phase consist of fibroblasts which, following stimulation by chemotactic PDGF, EGF and TGF-β gradients, migrate to the location of tissue damage.12,20 Fibroblasts constitute key cells responsible for the initiation of angiogenesis, epithelialization and collagen production. Angiogenesis is essential with concern to maintaining the granulation tissue and is associated with the activity of a large number of molecules (eg, VEGF, FGF, TGF-β and TNF-α).16 Moreover, MMPs promote angiogenesis via the liberation of VEGF,12,27 which stimulates endothelial cell functions such as proliferation, migration, differentiation and survival.18,28 The formation of granulation tissue, the deposition of collagen and angiogenesis occur simultaneously with epithelialization and wound contraction. Fibroblasts secrete collagen type III and fibronectin so as to create mechanically strong tissue.
The fourth and final phase of the wound healing process consists of remodeling, ie, a balance between synthesis and degradation so as to attain well organized tissue. The granulation tissue matures to form a scar. Fibroblasts change to become myofibroblasts and, together with collagen and fibronectin, participate in the wound contraction process. The collagen type I content increases in favor of collagen type III and the fibers become cross-linked and aggregated into the form of fibrillar bundles which affect both the stiffness and tensile strength of the healing tissue.26 FGF plays a very important role in this phase and is produced by keratinocytes, fibroblasts, endothelial cells, smooth muscle cells, chondrocytes and mast cells, some of which are involved in the formation of granulation tissue, epithelialization and tissue remodeling.18 Epithelialization occurs on the surface of the wound; epithelial cells, stimulated by EGF, KGF and TGF-α, migrate and proliferate so as to cover the new tissue. MMPs, with the inhibitors thereof (tissue inhibitor of metalloproteinases [TIMPs]), play a pivotal role in terms of regulating cell migration (keratinocytes, fibroblasts, epithelial and inflammatory cells) in the wound by modifying the wound matrix.22,27 The healing process is complex and long-lasting, and the maximal tensile strength of human wounds (ie, 70% of normal skin) is attained after around 1 year.16
Non-healing (chronic) wounds present a serious problem both for patients themselves and the health care system, and a therapy is urgently required that accelerates the wound healing process, prevents secondary infection and which provides relief to patients. The risk factors of chronic wounds include diabetes, peripheral vascular disease, immunosuppression, acquired immunodeficiency and injury to previously wounded local tissue such as that caused by radiation therapy or certain chemicals.10,26 Impaired wound healing is associated with prolonged inflammation and an imbalance between the production and breakdown of the most important molecules in the wound which may be caused by excessive neutrophil infiltration, an abundance of pro-inflammatory cytokines, ROS, premature cell senescence, defective MSCs or the enhanced activity of wound proteases which degrade PDGF and TGF-β;13,14,21,26 moreover, the oxygen and moisture balance is disrupted. The inflammation phase with respect to normal wound healing lasts up to 7 days, while chronic wounds often stall in this phase and require longer healing periods13 (Figure 2) due to the increased quantity of ECM degradation products which promote inflammation, thus creating a self-perpetuating cycle.21
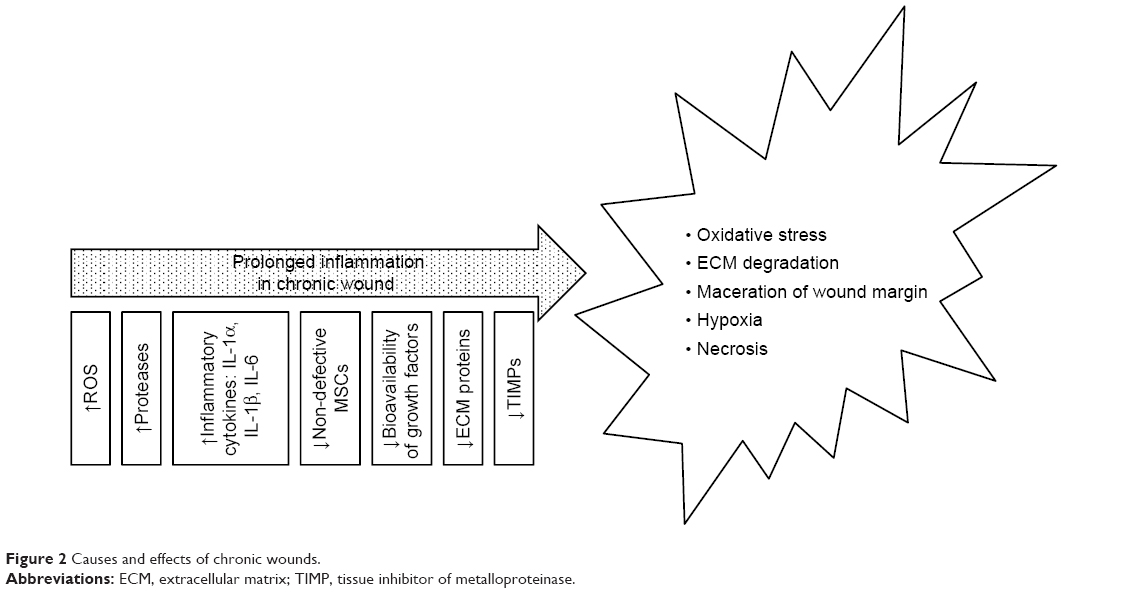 | Figure 2 Causes and effects of chronic wounds. |
Graphene – structure, production methods, characterization techniques and biology-related properties
Graphene is an allotrope of carbon, whose structure consists of a planar sheet of single carbon atoms upon which each atom is bonded to three others densely packed within a honeycomb crystal lattice. In order to create a hexagonal lattice, the carbon must be subjected to sp2 hybridization. Subsequently, overlapped sp2 orbitals create three orbitals which are responsible for the formation of a σ covalent bond (three bonds between the nearest carbon atoms). The fourth bond consists of a π-bond, which is perpendicularly oriented (z-direction, out of the plane).
The potential applications of graphene are highly dependent on the production method employed and the form in which it is obtained (Table 1). However, it has proved difficult to date to obtain an ideal graphene which is both flat and homogeneous,29,30 ie, the creation of an ideal single layer graphene remains a major challenge. In order to fulfill the expectations of engineers, who use graphene for various applications, and scientists who use it in a wide range of research fields, a number of graphene production techniques have been (and continue to be) developed. It is already known that the best quality graphene in terms of structural integrity and electrical properties is obtained by means of the mechanical cleavage of highly oriented pyrolytic graphite.31 Therefore, it will be necessary to evaluate the results of any new graphene production methods via a comparison of the materials produced with the properties of mechanically exfoliated pristine graphene.
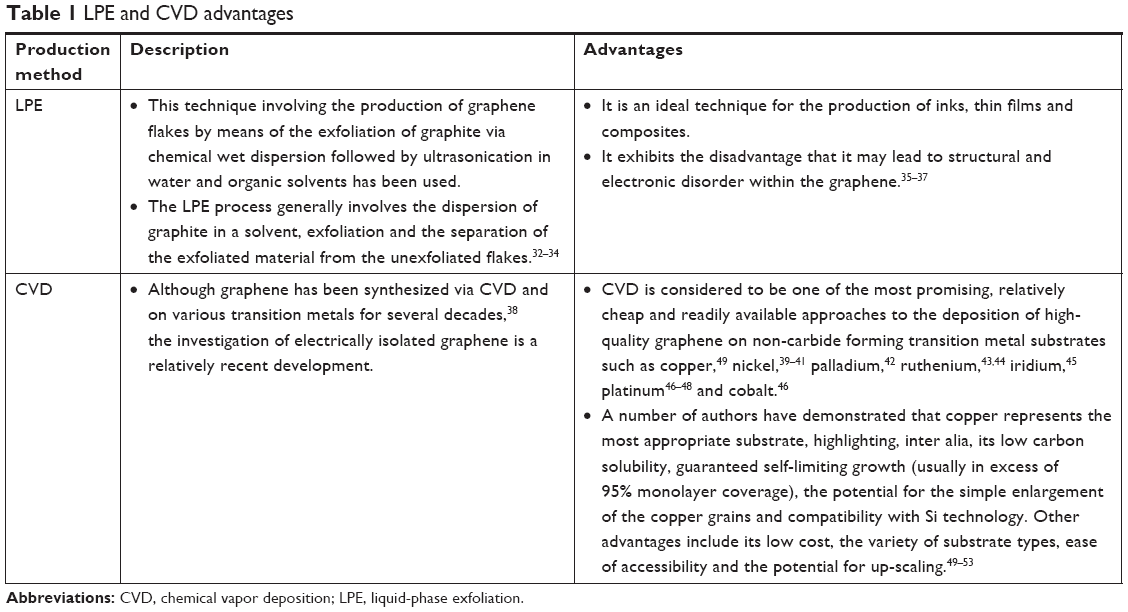 | Table 1 LPE and CVD advantages |
Several strategies are presently being explored aimed at attaining reproducible and scalable graphene on various substrates (Table 1).
Currently, techniques such as scanning electron microscopy (SEM) and Raman spectroscopy are usually employed for the identification of the structure of graphene.54,55 The advantages of these techniques are shown in Table 2. Figure 3 shows a set of SEM images of samples collected simultaneously using two different detectors. The typical features of graphene grown on Cu foil are marked in the images.
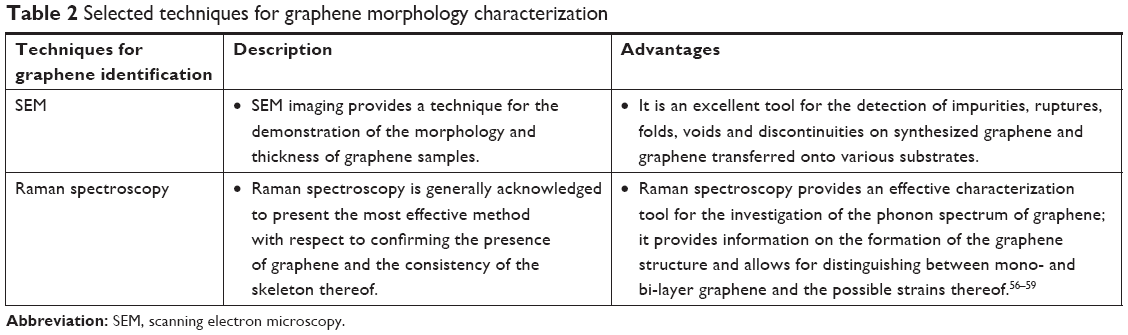 | Table 2 Selected techniques for graphene morphology characterization |
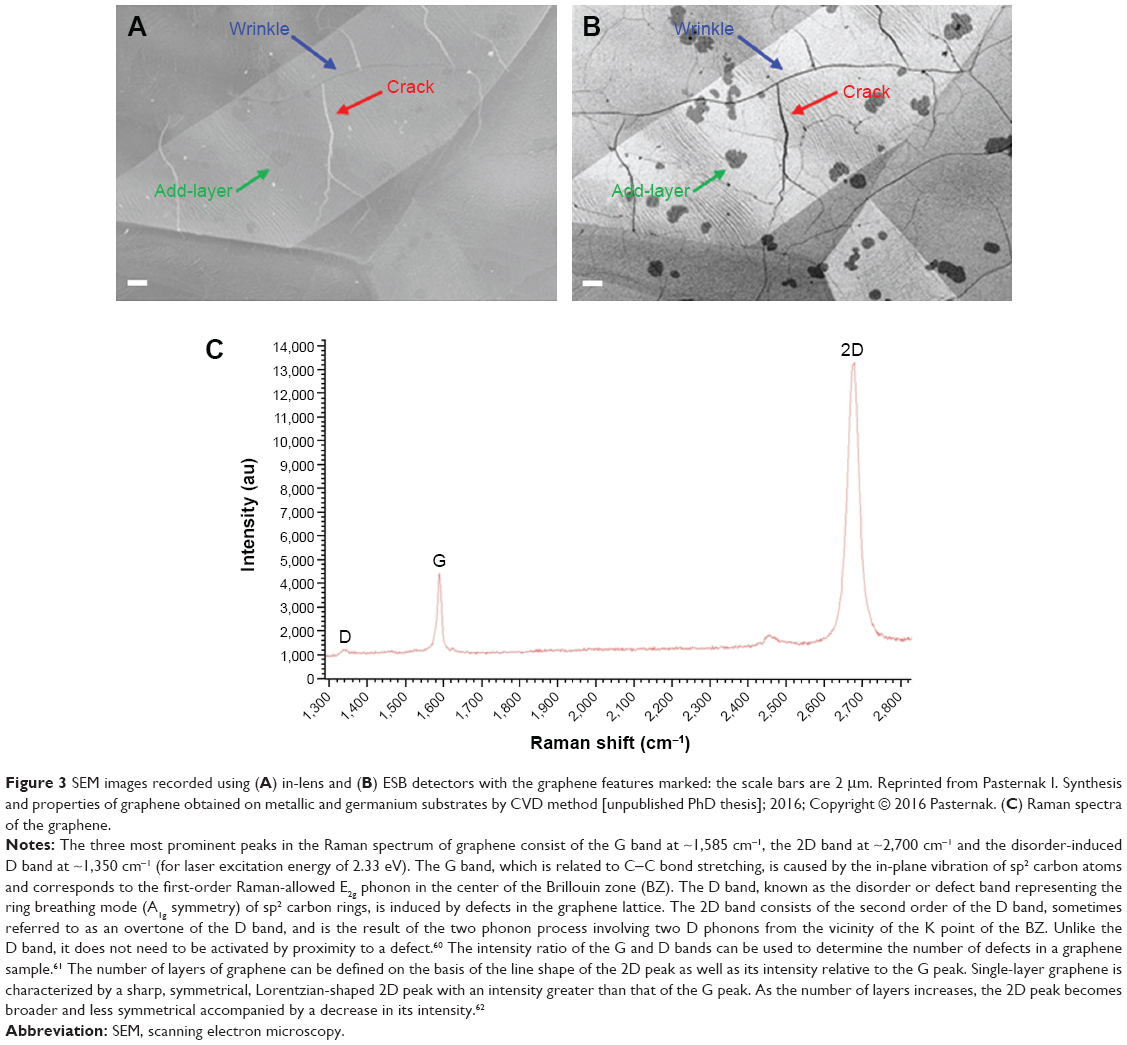 | Figure 3 SEM images recorded using (A) in-lens and (B) ESB detectors with the graphene features marked: the scale bars are 2 μm. Reprinted from Pasternak I. Synthesis and properties of graphene obtained on metallic and germanium substrates by CVD method [unpublished PhD thesis]; 2016; Copyright © 2016 Pasternak. (C) Raman spectra of the graphene. |
Graphene properties with respect to biology (wettability, nanotopography and defects)
Recently published scientific reports and reviews have discussed both the cytotoxic effects of materials belonging to the graphene group and the biosafety of graphene nanotechnologies.64–68 This review, however, focuses on the stimulating properties of graphene and its putative action mechanism, particularly with respect to wound healing and the reduction of infection. First, we distinguished a selection of graphene samples in terms of their physical form, surface chemistry, topography and surface energy; the graphene production method determines its physical form. The chemical vapor deposition (CVD) method depends on the deposition of carbon atoms from hydrocarbon gas on a substrate (eg, copper foil). Graphene films can be transferred onto a wide range of substrates53,55 and are used in both physics69 and biology (eg, surfaces for cell growth).70 While graphene solutions, in which graphene flakes and sheets are formed, are employed in the majority of toxicological studies65,68,71–73 involving the dispersion of graphene in biological fluids, much less attention has been devoted to the biocompatibility of graphene in the form of a monolayer63,70,74–76 which can be used as a scaffold for the transplanting of cells into damaged tissue, especially with respect to acute and chronic wound therapy.
Wettability
Water molecule and protein absorption occurs once the substrate comes into contact with its biological surroundings. The behavior of the substrate in contact with water depends on its hydrophobic and hydrophilic surface properties. GO consists of a highly defective graphene sheet functionalized with oxygen groups (hydroxyl, carboxyl, and epoxy) which evinces high levels of hydrophilicity and protein absorption. Reduced graphene oxide (rGO) is produced via the reduction of GO using high temperatures or chemicals and is considered to have the same favorable level of solubility as GO despite the hydrophobic nature of the pristine form of graphene.77 Huang et al78 indicated that a graphene film produced by means of the dispersing method (in a solvent of tetrahydrofuran, distilled water and dimethylformamide) exhibits reversible hydrophobic and hydrophilic transition in response to UV illumination and dark storage, respectively. Moreover, graphene can be further modulated so as to obtain hydrophobic and hydrophilic surfaces.79
Wettability affects the ability of cell adhesion-mediated proteins to attach to the substrate80 and, consequently, determines cell adhesion.81 Cells attach to the underlying substrate (protein layer) by means of focal contacts, ie, adhesive connections containing a large number of proteins (eg, integrins). The extracellular parts of integrins bind to the ECM, their integral parts anchor integrin into the cell membrane and their intracellular parts bind to the focal adhesion proteins thus forming a physical link between the ECM and the actin cytoskeletal network.63,82,83 Focal complexes that connect cells with the external environment are crucial for the functioning of cellular processes and mechanisms such as mechano-sensoring, spreading, cell migration and proliferation.84 The focal adhesion assembly responds to matrix stiffness,85 a phenomenon that is used by researchers for the qualitative and quantitative analysis of the effect on cellular behavior of the nanotopography of the various substrates used in regenerative medicine.63,79,86–88 James and Tour88 indicated that there are numerous permutations of graphene differentiated in terms both of their physical form and the number of layers. Dai et al30 measured water contact angle dependence on the number of graphene layers and summarized that the wettability of graphene depends on the number of layers, the graphene preparation substrate and its surface chemical composition. They determined that in the case of the presence of more than six layers, the water contact angle value of graphene equaled that of graphite.
Nanotopography
Nanotopography makes up a fundamental factor with respect to the design of biomaterials intended for tissue engineering applications.89 The surface properties of graphene can be described according to its morphology, ie, the presence of wrinkles, fluctuations and N2 adsorption, which alters its mechanical and chemical properties. Wrinkles may arise as the result of thermal stress occurring during the production process or due to the transfer technique employed, ie, it may be influenced by the metal substrate.90,91 Corrugation appearing on transferred surface-grown graphene has been determined at ~2–15 nm in height and ~20–100 nm in width90 or even smaller (1–2 nm).79 Graphene deformations also include ripple formations with a height of up to 1 nm in suspended graphene membranes and the formation of crumples which may be produced via the rapid evaporation of aerosol droplets.92 Roughness caused by the nanostructure of the graphene is capable of changing its hydrophobicity93 and, consequently, its interaction with molecules and cells (eg, focal adhesion, cytoskeleton contraction).
Graphene exhibits an extremely high specific surface area (theoretically 2,630 m2·g−1)30 depending on the preparation method employed and the number of layers, whereas the specific surface areas of the various graphene derivatives range from 600 to 1,600 m2·g−1.94 In conclusion, the various physical forms and chemical structures of graphene derivatives exert differing effects on cells.66
Defects
Each graphene production method leads to the production of differing properties and quality levels and influences the number of defects in the material,55 all of which are capable of affecting the material’s degree of impact on cells and, consequently, its therapeutic effect. Defect-free graphene does not exist; defects in the structure of graphene may arise spontaneously during the production process or may be introduced through changing the properties of the material29,55 which can be identified via the application of Raman spectroscopy (Figure 3).54 While certain defects exert a favorable effect, such as increasing the reactivity of the graphene, others must be eliminated prior to medical application, including hexagons that transform into pentagons (the Stone–Wales defect, SW), single (the absence of one lattice atom) or multiple vacancies, dimensional defects (eg, dislocations – line defects) and defects along the edges.29,54
The influence of graphene on cells and the putative mechanism of this process
The surface properties of graphene allow for its use as a platform for cell adhesion and even induced cell proliferation.75 It has been shown previously that graphene monolayers are non-toxic, stimulate the mitochondrial activity of mouse fibroblasts (L929)63 and potentiate the adhesion and proliferation of osteoblasts and MSCs.70,79,95 In addition, two-dimensional multilayer pristine graphene film has been found to demonstrate good biocompatibility with human stromal fibroblasts.96 Graphene–polycaprolactone composites have also been found to exhibit good biocompatibility employing L929 fibroblasts.97 As mentioned above, the presence of fibroblasts is crucial in the proliferation phase at which time they are recruited into the wound. Confirmation of the biocompatibility of graphene and fibroblasts is essential prior to considering further research on its use in the wound healing process. The use of graphene as a scaffold material exerts an influence on cells by means of its nanotopography; cells are influenced by mechanical forces in the local environment to which they respond via nano-transduction either by adaptation or death. The expected role of graphene in the wound healing process is to mimic the architecture of the native ECM in such a way that proliferation, migration and spatial organization lead to enhanced wound closure, an increase in the strength of the new tissue formed and a reduction in scar formation.
Immunofluorescence staining has revealed that L929 cells created more focal adhesions, and the migration of the cells appeared to be more regular, than on a glass control of similar roughness to graphene.63 Cell migration depends on the roughness of the substrate,82,89 according to which cells that migrate on graphene require a greater amount of energy for the dissolution of old adhesions, tail retraction and the balancing of internal and external forces. Increasing the effective surface energy (associated with moderately rough substrates) enhances the total amount of work per unit area required for full detachment,89 which may result in increased mitochondrial activity and the activation of pathways involved in all the processes concerned with cell migration and other processes essential for cell growth and proliferation. Rho family GPTase are involved in the translation of the signals that regulate the various cellular processes such as cell adhesion, actin cytoskeleton re-organization, polarity, cell growth, proliferation and chemotaxis (Figure 4).98–100 Rougher and stiffer substrates appear to provide better cell scaffolds resulting in enhanced intracellular tension and an increase in the number of focal adhesions and cell proliferation.63,89,100 Focal adhesions play an important role in the transduction of mechanical signals, and a complex network of signaling pathways is involved in the cellular response (Figure 4). Integrin activation leads to the activation of focal adhesion kinase (FAK) and Src kinase. The activation of FAK may lead to enhanced cell proliferation as mediated by extracellular signal-regulated kinases (ERKs) via various signaling pathways.98,100 Moreover, FAK is able to regulate cell migration by means of binding to and promoting the Src-mediated phosphorylation of p130Cas and via the regulation of the RhoA–ROCK pathway.98,100,101 The absence of FAK negatively affects the production of lamellipodia by the cells on the edge of the wound.101
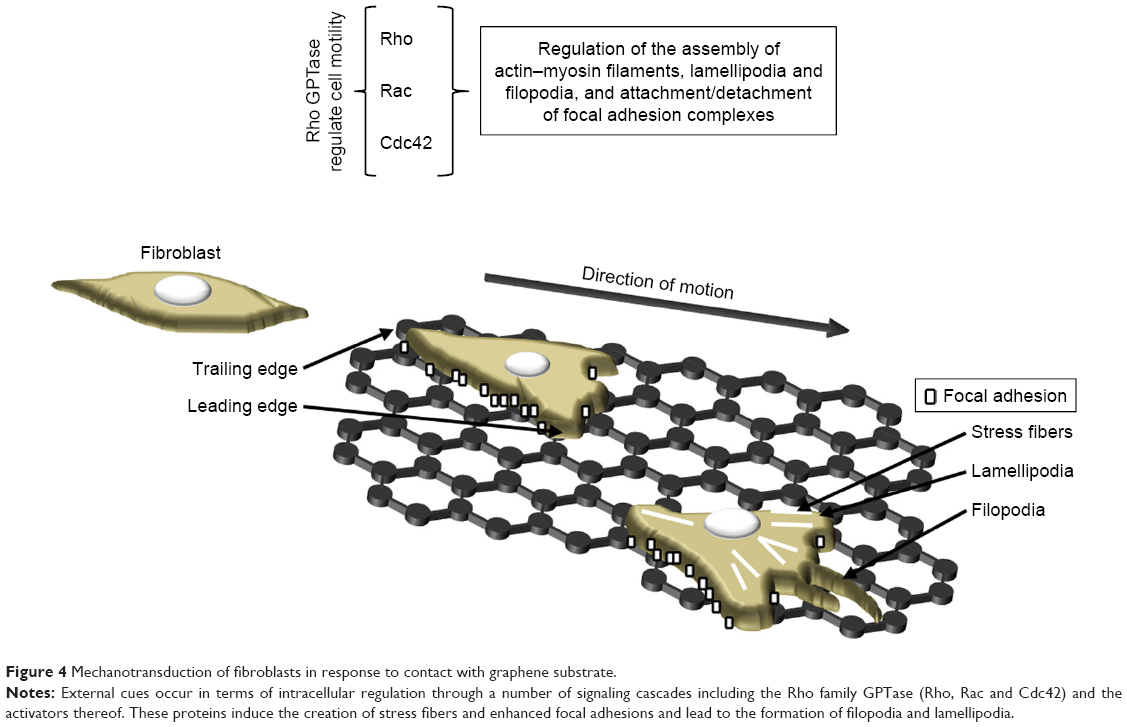 | Figure 4 Mechanotransduction of fibroblasts in response to contact with graphene substrate. |
The actin cytoskeleton is composed of actin filaments and, through combining microfilaments with multiple actin binding proteins, it creates various cellular forms such as cortical actin networks, stress fibers within the cytoplasm, shrinkage rings formed during cytokinesis and surface tabs (lamellipodia, filopodia) in the cells. A strong actin network is required in order to stabilize the cells on the substrate (Figure 4). Marked stress fibers observed in cells cultivated on a graphene scaffold63 indicate a struggle with the substrate architecture and stiffness forces. Kim et al102 and Zhang et al103 suggested that GO micropatterns might provide a suitable cell-guiding substrate for the purposes of tissue engineering and regenerative medicine. Moreover, triangular GO micro-patterns fabricated using meniscus-dragging deposition and photolithography techniques have been determined to enhance the speed, distance and directionality of L929 fibroblasts.102 Due to its ability to guide cells in a specific direction (essential with respect to wound closure and scar formation), the use of a micro-pattern graphene substrate for wound treatment is particularly advantageous. However, Keshavan et al104 showed that the response to identical surface cues (topographical and chemical) is a cell-type dependent mechanism. They noted the higher primary adhesion of Chinese hamster ovary cells on poly-D-lysine coated micro-patterned single-layer graphene (SLG) compared to that on adjacent SLG ablated stripes. However, during the incubation period, the cells were observed to migrate onto the adjacent SLG ablated stripes, which proved a more favorable environment for their subsequent proliferation. Interestingly, these same surfaces resulted in differing primary neuron cell arrangements.105 Osteoblasts have been found to adhere to GO patterns exhibiting highly aligned, oriented and elongated actin filaments which have adapted to the pattern width.103 Moreover, the authors also point out that polarized cells with high cytoskeleton tension and traction forces are capable of overcoming the strong adhesion between cells and GO, thus leading to higher cell contractility and mobility. Following the discovery of the surface patterning of graphene, new tissue engineering applications emerged employing these unique properties. Finally, it has also been shown that materials with unique nano-topographical characteristics – including graphene – offer properties which are similar to those of growth factors.11 To sum up therefore, cells are affected by the mechanical properties of the scaffold, its nano-architecture and chemical signals (eg, growth factors, cytokines and ECM modifying enzymes bound to the graphene). These stimuli lead to tensile, compressive and shear stress which lead to changes in the cell structure and the initiation of signal transduction (eg, FA-Rho GTPase crosstalk) (Figure 4).
One of the additional benefits of graphene consists of its antibacterial activity, which may offer an alternative to the use of antibiotics in the wound healing context. A range of bacteria limitation/death mechanisms initiated by both graphene and its derivatives have been described in the literature14,106–109 depending on the diversity of the various forms of graphene and their chemical properties and the structure of the bacteria wall. The antibacterial activity of graphene materials is associated with membrane stress, which may be accompanied by ROS-dependent110 or -independent oxidative stress.111 Some authors report that the sharp edges of graphene cut through the cell membranes of bacteria thus causing lethal damage to cellular integrity.112,113 A further mechanism consists of the isolation of bacteria through wrapping/trapping them in a sheet-form blanket of graphene thus limiting bacterial access to nutrients.108 Shuai et al proposed GO–Ag nanosystem (polymer scaffold containing 1 wt% GO–1 wt% Ag) with synergistic effect on antibacterial action via combining the capturing effects of GO nanosheets and the killing effects of Ag and showed bacterial inhibition rate >95%.114 Escherichia coli cells were found to lose their cellular integrity accompanied by severe membrane damage following 2.5 hours of incubation with 100 μg mL−1 GO nanosheets;115 moreover, the authors indicated that a large amount of phospholipids were freed from the bacteria cell membranes as a result of interactions between the graphene and lipid molecules. Kurantowicz et al116 determined that 250 μg mL−1 of pristine graphene, GO and rGO consistently inhibited the growth of Salmonella enterica and Listeria monocytogenes by 100%. They further demonstrated that bacterial cells interacted with the sp3-hybrized oxidative group of the GO and distributed themselves over the surface thereof, while the bacterial cells were arranged at the edges of the pristine graphene and rGO. Moreover, they also showed that pristine graphene and rGO exhibit lower levels of antibacterial activity than does GO. On the other hand, Barbolina et al117 pointed out that graphene contaminants are responsible for the reported antibacterial properties rather than graphene alone and concluded that GO purification is crucial in order to ensure the true biological effect of the material. The authors, using highly purified and thoroughly washed GO, failed to discover either bactericidal or bacteriostatic properties over a broad concentration range with concern to planktonic cultures of either E. coli or Staphylococcus aureus.
In addition, the antiviral action of graphene has been demonstrated by Ye et al118 who suggested that this property can be attributed to the unique single-layer structure and negative charge. A non-cytotoxic concentration (6 μg mL−1) of GO was added to PK-15 cells infected with pseudorabies virus and Vero cells infected with porcine epidemic diarrhea virus and was found to suppress both infections. The authors noticed that the GO in the cell culture did not block viral replication and the subsequent spread to neighboring cells, rather the pre-incubation of the viruses with GO induced the significant inhibition of infection. Thus, they suggested that GO inhibits virus infection by inactivating virus particles prior to entering cells. They concluded that the antiviral action mechanism is based on the electrostatic interaction of negatively charged sharp-edged GO with positively charged virus particles, resulting in viral morphology damage (both the envelope and the spikes were destroyed) and subsequent inactivation. Moreover, the authors indicated that both GO and rGO exhibit similar antiviral activity and that the oxygen-containing group is not essential for the initiation of such activity. Song et al119 demonstrated that negatively charged GO efficiently captured the enteric EV71 and H9N2 viruses and that GO surfaces are capable of destabilizing enveloped viruses.
Graphene has also been investigated with respect to hemocompatibility and angiogenic action.65,120–122 GO was shown to exhibit prothrombotic properties which are able to activate Src kinases and induce the release of calcium from intracellular stores; the prothrombotic character was shown to be dependent on the surface charge distribution.123 Jaworski et al,65 based on the results of experiments on chicken embryo red blood cells, demonstrated that different forms of graphene exhibit differing hemocompatibility depending on the production method employed and the surface modification. In addition, Mukherjee et al120 demonstrated the pro-angiogenic activity of graphene and proposed a mechanism based on the intracellular formation of ROS and reactive nitrogen species and the activation of phospho-eNOS and phospho-Akt. Shine et al122 reported that with higher concentrations of graphene (from 0.25% to 1% in the composite), the expression level of angiogenic proteins was enhanced in human mesenchymal stem cells (hMSCs) cultured on calcium silicate/graphene composites. Park et al121 indicated that the incorporation of rGO flakes into MSC spheroids and monolayer cultures promoted the expression of proangiogenic growth factors (VEGF, FGF-2, and HGF) and that the highest expression concerned hybrid spheroids with 5 μg mL−1 rGO flakes. The authors also demonstrated that enhanced cell–ECM interaction through the incorporation of rGO flakes into MSC spheroids leads to an increased amount of VEGF via mediated FN-integrin binding, which leads to the enhanced expression of phosphorylated FAK, phosphorylated ERK and thus VEGF.
Graphene and its derivatives have also been shown to possess immunomodulatory properties depending on their physicochemical features and functionalization.124 These nanocompounds are able to modulate the functions of phagocytic immune cells that participate in supporting the normal wound healing process, including neutrophils,125 macrophages19 and dendritic cells (DCs).126 Neutrophils constitute the first inflammatory cells recruited to the wound tissue from the blood and both act to sterilize the wound via the production of antimicrobial peptides and proteases and to regulate the inflammatory response via the secretion of multiple cytokines and growth factors.125 Recently, it has been determined that primary human neutrophils exposed to small (50–300 nm) and large (10–40 μm) sheets of GO produce neutrophil extracellular traps (NETs), ie, fibrillar networks that contribute toward defense against pathogens (Figure 5).127 The production of NETs was seen to be dependent on the size of the GO sheets and was associated with both ROS production and the influx of Ca2+. Despite the fact that the GO-induced formation of NETs eventually led to neutrophil-cell death (NETosis),127 it represents an important mechanism in terms of immobilizing and killing medically relevant bacteria.128 Macrophages fulfill a large number of beneficial functions with respect to promoting the wound healing process, including the regulation of the inflammatory response, the removal of neutrophils/apoptotic cells, the promotion of angiogenesis, fibroblast proliferation and ECM reorganization.19 In general, the various nanomaterials (graphene family nanomaterials [GFNs]) of the graphene family are cytotoxic in a dose-dependent manner and induce differing types of cell death (apoptosis, autophagy and necrosis) in macrophages.124 However, sub-cytotoxic concentrations of GFNs and the appropriate variations in their physicochemical properties are capable of modulating the immune functions of these cells. It has been shown that a sub-cytotoxic dose of pristine graphene stimulates primary murine macrophages and immortalized macrophages into secreting Th1/Th2 cytokines (IL-1α, IL-6, IL-10, TNF-α and GM-CSF) and chemokines (MCP-1, MIP-1α, MIP-1β and RANTES), most probably due to the toll-like receptor (TLR)-dependent activation of the nuclear factor-κB (NF-κB) signaling pathway.129 (Figure 5). Moreover, GO induces an inflammatory response (together with autophagy) in murine RAW 264.7 macrophages by activating TLRs (TLR4 and TLR9) and their downstream MyD88-, TRAF6- and NF-κB-dependent signaling pathways.130 In addition, GO sheets polarize macrophages toward the M1 phenotype and enhance their pro-inflammatory response in a size-dependent manner. Larger GO sheets exhibit a stronger interaction with the TLR4 plasma membrane, resulting in NF-κB activation and M1 polarization both in vitro and in vivo.131 The M1 immune polarization effect has also been observed with respect to monocytes treated with GO functionalized with amino groups.132 DCs infiltrate wounds quickly following injury and accelerate early wound closure, most likely via the secretion of factors that increase cellular proliferation, granulation tissue formation and angiogenesis.126 GO and other carbonaceous nanoparticles, ie, C60 fullerenes and C60-TRIS fullerenes, have been shown to be taken up by conventional DCs and differentially modulate the antigen presentation ability of these cells (Figure 5). GO only (ie, not fullerenes – a further form of carbon along with graphite and diamond) was found to downregulate intracellular levels of immunoproteasome subunit low molecular mass polypeptide 7 and thus decrease the level of antigen processing in DCs.133 Another study demonstrated that pure GO induces the higher maturation and stronger production of TNF-α and IL-1β in human DCs than does polyvinylpyrrolidone-functionalized GO.134 Moreover, the treatment of DCs with a mixture of GO and a glioma peptide antigen enhances their anti-tumor immune response.135 Taken together, the immune regulatory properties of GFNs are determined largely by the dosage and the variation in their physicochemical properties.
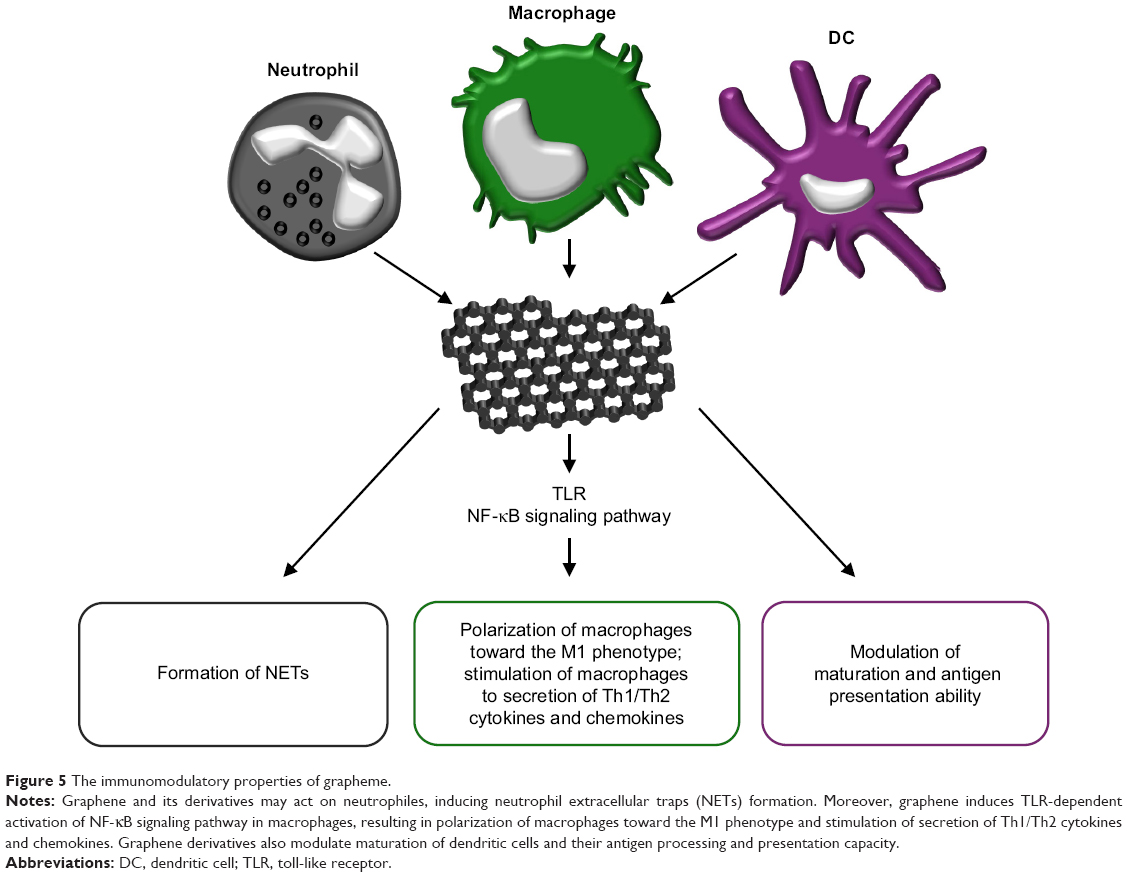 | Figure 5 The immunomodulatory properties of grapheme. |
Graphene has also been used in in vivo studies usually incorporated into composites. A study by Deepachitra et al1 indicated that GO incorporated into a collagen-fibrin biofilm resulted in no adverse effects and enhanced the wound healing process in Wistar rats. In addition, they noticed an increase in the mechanical strength of the composite films with GO and rat healing tissue, thus indicating its potential use as a structural reinforcement material. Zhong et al136 proposed the use of GO as a delivery system for recombinant TIMP-1 (a metalloproteinase inhibitor) and demonstrated the continuous release of TIMP-1 from the GO for up to 40 days. The subcutaneous administration of TIMP-1-GO to rats with experimental wounds has been shown to promote both vascularization and collagen regeneration. Mahmoudi et al15 prepared nanofibers containing GO nanosheets via the electrospinning of chitosan–PVP and demonstrated that GO promoted adhesion and the viability of human skin fibroblast cells, enhanced bactericidal capacity and accelerated the wound healing process in Sprague Dawley rats.
MSCs and their putative mechanism of action in wound repair
The use of MSCs in the treatment of wounds raises great hope for regenerative medicine. MSCs have the capacity for multi-lineage differentiation.83,137,138 They adhere to the surface of the culture vessel, exhibit fibroblast-like morphology and develop into symmetrical colonies. They express such antigens as CD73, CD90 and Cd105 and should not express CD14, D19, CD34, CD45, CD11b, CD79a and HLA-DR surface molecules.3,6,9,139–142 Bone marrow, the umbilical cord, adipose tissue, placenta and cord blood all provide sources of MSCs (Figure 6) containing the therapeutic potential for the treatment of wound healing disorders.143 With respect to normal cutaneous wound healing, MSCs are mobilized from their host sources to the injury site where they support skin repair despite hypoxia and a lack of nutrients.22,144 MSC therapy is dependent on both the sufficient extent of MSC engraftment at the injury site and cell survival within the wound. While autologous MSC transplantation provides a number of reasons for optimism, allogenic MSC transplantation is also feasible since these cells, as with group O red blood cells, are immunologically silent.145,146 A conditioned medium of MSCs (MSC-CM), which includes bioactive molecules secreted by the MSCs in the culture, has also demonstrated regenerative effects with concern to wound healing tissue.2,140,145,147
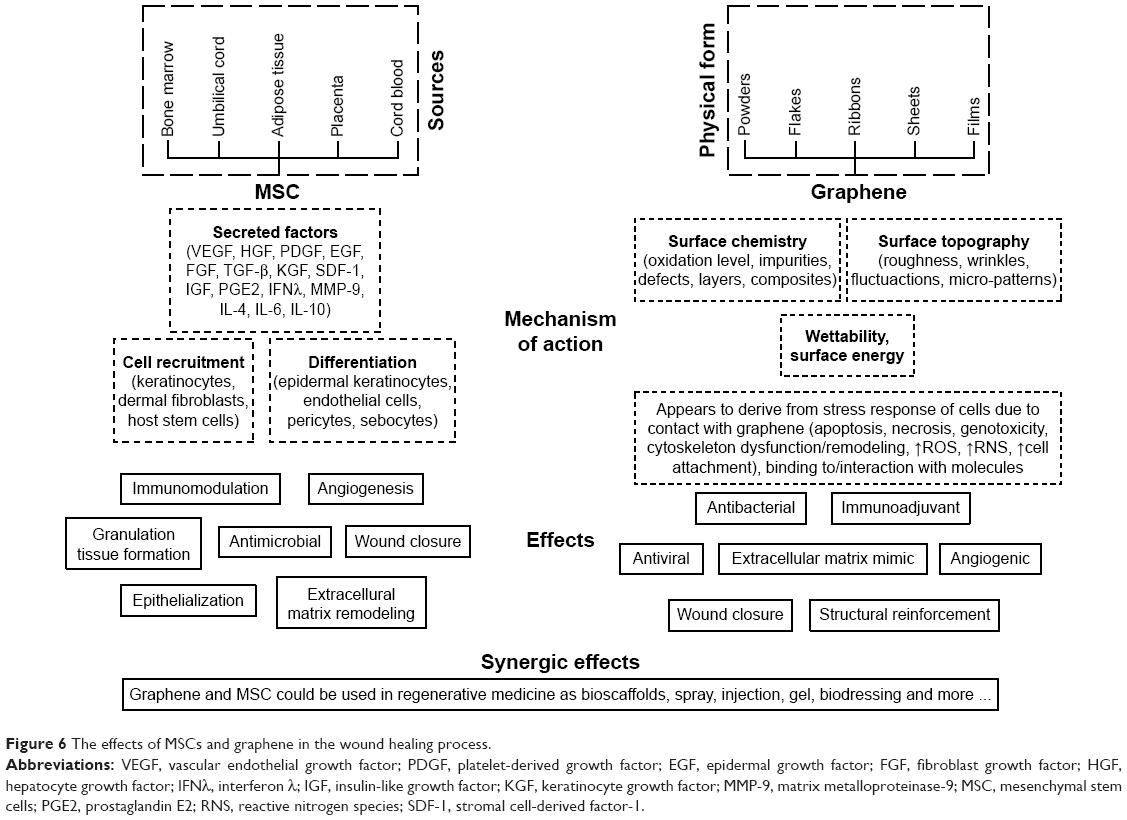 | Figure 6 The effects of MSCs and graphene in the wound healing process. |
One of the most important features regarding the clinical application of MSCs consists of their ability to recruit other cells for the purpose of tissue repair, concerning which differentiation and paracrine signaling have been identified as mechanisms of their action.3,139 It has been shown that MSC-CM enhances wound closure via the acceleration of the in vitro migration of fibroblasts and keratinocytes.147 The wound healing process requires interaction between cells, ECM proteins and biomolecules (growth factors, cytokines and chemokines), and MSCs play a key role in the coordination of individual damaged tissue regeneration processes.11,139 The number of connections with the ECM exerts a significant impact on the strength of the new tissue that replaces lost tissue in the wound. To date, a number of routes have been studied concerning the introduction of MSCs into the organism for wound healing purposes146,148,149 and the most recent study conducted on this theme revealed that the subcutaneous injection of MSCs provided a much more efficient method than intravenous injection with concern to the healing of skin wounds.150 The use of exogenous MSCs also provides a promising strategy with respect to the treatment of non-healing wounds as in the case of those caused by diabetes, vascular insufficiency and several other medical conditions.22,151 While it is believed that MSCs have the therapeutic potential for application with concern to wound-healing disorders, the action mechanism is still not fully understood.143
Although the injection of MSCs into the blood stream leads to healing to a certain extent, the delivery of MSCs on scaffolds provides a significantly more potent therapeutic approach (ie, it is site-specific). Multi-functional scaffolds have the potential to guide the adhesion, growth and differentiation of MSCs so as to form skin-functional and structural tissue. When designing suitable MSC scaffolds, it is important to remember that the physicochemical properties of the biomaterials used may well determine and change the fate of MSCs.138 The stiffness, elasticity, porosity and reactivity of the material may potentially affect cellular behavior through the forces applied, the activation of various molecules during the cell adhesion process and interactions with the scaffold.152–154 Thus, the creation of an effective physical platform will allow for the control of various processes such as the attachment, proliferation and differentiation of MSCs. Moreover, it may also assist in the development of a specific required biological effect via the direction of the behavior of the cell. A detailed knowledge of the signaling mechanism activated by scaffold–cell interactions would reveal the direction of a range of cellular activities thus making it possible to achieve a specific aim via the manipulation of the physiochemical properties of the biomaterial scaffold.
MSC as a producer of bioactive molecules
The secretion of bioactive factors is thought to constitute the principal MSC action mechanism during the wound healing process (Figure 6).7,140,143 The paracrine effect of MSCs is based on the release of growth factors, ILs and other bioactive molecules secreted or packaged into extracellular vesicles or exosomes.8 Growth factors play a pivotal role in the modulation and coordination of cellular processes in all phases of the wound healing process (Figure 1) and the sustained release of these molecules coupled with their bioactivity both stimulate the wound environment and promote wound closure.
The immunomodulatory effects of MSCs are related to the reprogramming of macrophages from type M1 to type M2 anti-inflammatory phenotypes which suppresses the proliferation of T cells, modulates TNF-α production, reduces the NK cell function in the inflammatory phase and lowers the level of IFN-γ activity in the process.8–10,144,155–157 MSCs secrete factors that upregulate the secretion of IL-10 and inhibit neutrophil infiltration into the wound.10 They also secrete IL-10 by themselves.139 MSCs also release TGF-β1 and HGF, which suppresses T cell proliferation, and PGE2, which prevents the differentiation of CD4+ T cells into Th17 cells.157 MSCs also release anti-inflammatory cytokines, eg, IL-4, which is important with respect to chronic wound healing.10 The secretion of IL-6 by MSCs has been demonstrated both in mice and in humans and its dual nature (pro-inflammatory and/or anti-inflammatory effects) has been emphasized.4 MSCs produce IL-6 in a p38MAPK pathway-dependent manner.158 Tamama and Kerpedjieva145 reviewed the relevant literature and summarized that both MSCs themselves and MSC-CM encourage wound repair and that multiple growth factors and cytokines (VEGF, bFGF, IL-6, IL-8) are involved in the MSC-mediated wound healing process.
MSCs promote new vessel formation through the release of VEGF.144 An et al150 revealed that autophagy in MSCs improves cutaneous wound healing via the paracrine secretion of VEGF and the direct phosphorylation of ERK, resulting in the further promotion of the VEGF-induced vascularization of endothelial cells. MSCs have been found to extensively express those factors involved in vessel stabilization, smooth muscle cell migration and matrix remodeling such as TGF-β, PDGF-β and MMP-9 as well as high levels of stromal cell-derived factor-1 (SDF-1) α chemokine, which is known to be involved in the recruitment and retention of proangiogenic macrophages and MSCs themselves.83,159
Wu et al160 revealed that BM-MSC-treated wounds exhibited accelerated wound closure in normal BALB/c mice and diabetic mice compared with fibroblast- or vehicle control medium-treated wounds. Shin et al161 demonstrated that the administration of tonsil-derived MSCs into wound beds significantly promoted the repair of surgical defects in mice. Luo et al162 discovered that MSC-treated wounds exhibited a more regular fiber alignment than did the wounds of the control animals and, moreover, that the former developed both hair follicles and sweat glands.
Li et al140 revealed that the proliferation and migration of dermal fibroblasts was enhanced by MSC-CM; moreover, the activity of the MMPs thereof and the expression of TGF-β3 increased following MSC-CM treatment. The authors also indicated more rapid wound healing and less scarring following the application of MSC-CM in vivo. High levels of TGF-β3 and low levels of TGF-β1 were found in an embryotic wound microenvironment in which scar-free healing generally occurs.163 Hence, the ratio of TGF-β3 to TGF-β1 appears to be an important factor with concern to scar-free wound healing. It is proposed therefore that the anti-scarring potential of TGF-β3 released by MSCs should be employed in medical applications. The benefit of employing MSCs compared to the exogenous administration of TGF-β3 lies in the greater action spectrum associated with the release from the MSCs of a range of other bioactive factors. Scars occur as the result of the excessive amount of the ECM deposited by fibroblasts in the wound bed144 and feature a lack of follicles and nerve endings. In addition, the tensile strength of the new tissue is substantially weaker. MSCs promote anti-scarring properties through the secretion of PGE2, which induces the increased expression of IL-10 via T cells and macrophages.156 The MSC upregulation of IL-10 decreases the expression of the IL-6 and IL-8 necessary for the prevention of the occurrence of an excessive increase in the deposition of collagen in the wound.144,156 The HGF secreted by MSCs acts to modulate fibroblasts via the downregulation of the expression of TGF-β1 which drives both myofibroblast differentiation and the production of collagens types I and III.144,157 HGF also enhances the degradation of the ECM through the upregulation of the fibroblast expression of MMPs.157 MMP-9, ie, one of the MMPs released by MSCs exhibits a high degree of activity against gelatin and degrades other ECM molecules including collagens, laminin and aggrecan.27
The antimicrobial activity of MSCs is based on the secretion of antimicrobial factors such as LL-37 and immune-modulative factors which upregulate the killing of bacteria and the phagocytosis thereof by immune cells.10,22 LL-37 makes up one of the antimicrobial peptides and proteins group – also known as “host defense peptides” and the low production thereof is associated with skin disorders.141
The environment (ie, the tension of oxygen) makes up a further crucial factor, which influences the behavior of MSCs (gene expression, the release of cytokines and other factors). Chen et al2 revealed, based on real-time PCR analysis, that MSCs treated under hypoxic conditions expressed significantly greater amounts of EGF, KGF, IGF-1, VEGF-α and SDF-1 but lower amounts of TGF-β1 than did dermal fibroblasts. In addition, EGF is, for example, an important growth factor with respect to re-epithelialization and the promotion of wound closure. Thus, it is also possible to control the function of MSCs via the triggering of the cultivation conditions.
In summary, MSCs contain a wide range of growth factors and cytokines, which work in synergy to accelerate the wound healing process. The features of the abovementioned bioactive molecules with respect to cell recruitment and MSC differentiation in the context of wound healing will be further described below.
MSC as a coordinator of cell recruitment and differentiation
The therapeutic action of exogenously delivered MSCs and MSC-CM lies in its selective recruitment of host cells to the injury site and the direct differentiation of MSCs. SDF-1 is thought to play an important role in terms of the recruitment of stem cells from bone marrow to the injury site via a CXCR4-dependent mechanism. The enhancement of SDF-1 signaling within injured tissue can also be used to augment cellular transplantation.6,164 MSCs provide support for native cells at the injury site via the secretion of a variety of pro-survival and pro-migratory cytokines and growth factors.165 Chen et al2 found that MSC-CM significantly enhances the migration and proliferation of keratinocytes and endothelial cells; the authors suggested that the various factors released by MSCs (VEGF-α, IGF-1, EGF, KGF, angiopoietin-1, SDF-1, macrophage inflammatory protein-1 alpha and beta and erythropoietin) recruit macrophages and endothelial cells to the wound, thus enhancing the healing process. Lee et al166 demonstrated that MSC-CM harvested under hypoxia promoted fibroblast migration in vitro and dermal wound closure more rapidly than did MSC-CM collected under the normoxic culture condition. Rodriguez-Menocal et al167 revealed that healthy donor MSCs were significantly better than MSCs derived from chronic wound patients in terms of inducing normal fibroblasts to migrate; the authors also indicated that bone marrow-derived MSCs induce fibroblast migration in a dose-dependent manner. Increased fibroblast migration was observed in the presence of MSCs in a low concentration (10% of the population); however, an increased MSC concentration (20% or higher) led to a decrease in the migration of fibroblasts. On the other hand, the attenuated infiltration of inflammatory cells has been observed following the transplantation of tonsil MSCs into mice.161 It might be concluded therefore that the secreted factors of MSCs create a specialized cell recruitment microenvironment and offer great potential with respect to stem cell-based therapies.
Certain evidence suggests that MSCs may also induce tissue regenerative processes through in situ differentiation. When MSCs were cocultured in vitro with keratinocytes they exhibited trans-differentiation to keratinocytes.157 Mishra et al168 demonstrated that a keratinocyte-conditioned medium induced MSC differentiation to dermal myofibroblast-like cells and also enhanced the expression of cytokines including SDF-1, IL-8, IL-6 and CXCL5. The authors also noticed the organization of MSCs around the keratinocytes in vitro and in vivo, which they subsequently compared to the wound repair granulation phase. Sasaki et al169 indicated that MSCs have the capacity to differentiate into multiple skin cell types including keratinocytes, endothelial cells and pericytes in cutaneous wounds in mice following intravenous injection. Wu et al160 demonstrated that the MSC treatment of wounds enhanced the healing process in both normal and diabetic mice. Thus, MSCs play a dual role in wound healing by both producing specific factors and differentiating to specific cell types.
The senescence of MSCs
Cultured primary cells exhibit a limited division number and the aging of MSCs appears to present a major problem with respect to clinical applications, which require a significant number of cells.170–172 Many authors have remarked that later passage MSCs exhibited morphological abnormalities (an increase in cytoplasm granularity and the formation of vacuoles), enlargement and slower proliferation rates.170,171,173–175 They concluded that the long-term culturing of MSCs results in an increase in cell senescence. Turinetto et al171 suggested that it is difficult to predict which passage or number of cell divisions characterize the senescent state of MSCs due to variations in terms both of seeding densities and the time of harvesting. Whitfield et al174 observed a human adult bone marrow stromal cell population during increased in vitro passaging and discovered that the cells increased in size over time; they concluded that the larger MSCs had originated from several different generations and that they had exited the normal cell cycle, thus no longer fulfilling the MSC criterion of exhibiting the capacity for self-renewal in vitro.
Outlook and conclusion
Graphene–MSC dressings present a potentially attractive therapy involving the alteration of the wound environment via both mechanical and chemical stimuli. It is possible to create and alter the trophic functions of MSCs via dynamic ECM–cytoskeletal interactions, cell–cell contacts and soluble and transcription factor signaling,8 and they can be potentiated by means of graphene scaffolds. Kalbacova et al70 were the first to demonstrate that graphene in the monolayer form is non-toxic for MSCs and, moreover, stimulates the growth thereof. Kazantseva et al152 showed that graphene-augmented inorganic nanofiber scaffolds do not impede the normal growth of adipose-derived hMSC; moreover, they are able to both direct the preferential orientation and alter the morphology of MSCs. Shine et al122 showed that hMSCs are capable of uniformly covering calcium silicate/graphene composites and that composites with a higher content of graphene (1%) enhance cell proliferation. Going forward, the greatest challenge for scientists is to produce graphene in a way that closely resembles the nanotopography of the natural ECM of human skin and to achieve a similar biological effect. The direct delivery of MSCs to wounds may induce rapid cell death;143,157 thus, the introduction of MSCs via graphene nanoscaffolds presents a promising alternative delivery method which is capable of minimizing unprogrammed cell death. Li et al176 proposed a 3D graphene foam scaffold loaded with MSCs in connection with wound healing which both exhibited good biocompatibility and promoted the growth and proliferation of MSCs. The authors emphasized that the mechanical properties of graphene foam and MSCs strongly promote integration with the host tissue, which results in enhanced and more rapid wound closure. Li et al176 concluded that a 3D graphene foam together with MSCs synergized so as to promote wound closure via the enhancement of early vascularization accompanied by a reduction in scarring in an animal model, most probably due to the specific electrical properties of 3D graphene foam. Chu et al showed that hybrid scaffold containing in wound healing in diabetic rats.177 The results of experiments conducted to date allow us to conclude that graphene in combination with certain cells has the potential for use in the enhancement of the healing of complicated wounds and that MSCs introduced to wounds directly via graphene scaffolds presents a viable alternative to traditional dressing materials. We believe that the stiffness and nanotopography of cell culture scaffolds like graphene generate the mechanical signals required for the regulation of cell signaling that triggers cell response: migration, proliferation and differentiation.
Acknowledgments
This work was supported by the Warsaw University of Life Sciences (grant number 505-10-072500-P00191-99), the project of National Sustainability Program I No LO1503 provided by the Ministry of Education, Youth and Sports of the Czech Republic, and PROGRES Q26 provided by Charles University, Czech Republic.
Disclosure
The authors report no conflicts of interest in this work.
References
Deepachitra R, Ramnath V, Sastry TP. Graphene oxide incorporated collagen–fibrin biofilm as a wound dressing material. RSC Adv. 2014;4(107):62717–62727. | ||
Chen L, Tredget EE, Wu PYG, Wu Y. Paracrine factors of mesenchymal stem cells recruit macrophages and endothelial lineage cells and enhance wound healing. PLoS One. 2008;3(4):e1886. | ||
Hocking AM, Gibran NS. Mesenchymal stem cells: paracrine signaling and differentiation during cutaneous wound repair. Exp Cell Res. 2010;316(14):2213–2219. | ||
Kyurkchiev D, Bochev I, Ivanova-Todorova E, et al. Secretion of immunoregulatory cytokines by mesenchymal stem cells. World J Stem Cells. 2014;6(5):552–570. | ||
Isakson M, de Blacam C, Whelan D, McArdle A, Clover AJP. Mesenchymal stem cells and cutaneous wound healing: current evidence and future potential. Stem Cells Int. 2015;2015(7):1–12. | ||
Marquez-Curtis LA, Janowska-Wieczorek A, McGann LE, Elliott JA. Mesenchymal stromal cells derived from various tissues: biological, clinical and cryopreservation aspects. Cryobiology. 2015;71(2):181–197. | ||
Duscher D, Barrera J, Wong VW, et al. Stem cells in wound healing: the future of regenerative medicine? A mini-review. Gerontology. 2016;62(2):216–225. | ||
Hofer HR, Tuan RS. Secreted trophic factors of mesenchymal stem cells support neurovascular and musculoskeletal therapies. Stem Cell Res Ther. 2016;7(1):131. | ||
Lee DE, Ayoub N, Agrawal DK. Mesenchymal stem cells and cutaneous wound healing: novel methods to increase cell delivery and therapeutic efficacy. Stem Cell Res Ther. 2016;7:37. | ||
Maxson S, Lopez EA, Yoo D, Danilkovitch-Miagkova A, Leroux MA. Concise review: role of mesenchymal stem cells in wound repair. Stem Cells Transl Med. 2012;1(2):142–149. | ||
Salmasi S, Kalaskar DM, Yoon WW, Blunn GW, Seifalian AM. Role of nanotopography in the development of tissue engineered 3D organs and tissues using mesenchymal stem cells. World J Stem Cells. 2015;7(2):266–280. | ||
Harper D. The physiology of wound healing. Surgery. 2014;32(9):445–450. | ||
Frykberg RG, Banks J. Challenges in the treatment of chronic wounds. Adv Wound Care (New Rochelle). 2015;4(9):560–582. | ||
Parani M, Lokhande G, Singh A, Gaharwar AK. Engineered nanomaterials for infection control and healing acute and chronic wounds. ACS Appl Mater Interfaces. 2016;8(16):10049–10069. | ||
Mahmoudi N, Eslahi N, Mehdipour A, et al. Temporary skin grafts based on hybrid graphene oxide-natural biopolymer nanofibers as effective wound healing substitutes: pre-clinical and pathological studies in animal models. J Mater Sci Mater Med. 2017;28(73):1–13. | ||
Sinno H, Prakash S. Complements and the wound healing cascade: an updated review. Plast Surg Int. 2013;2013(6):1–7. | ||
João de Masi EC, Campos AC, João de Masi FD, Ratti MA, Ike IS, João de Masi RD. The influence of growth factors on skin wound healing in rats. Braz J Otorhinolaryngol. 2016;82(5):512–521. | ||
Barrientos S, Brem H, Stojadinovic O, Tomic-Canic M. Clinical application of growth factors and cytokines in wound healing. Wound Repair Regen. 2014;22(5):569–578. | ||
Koh TJ, DiPietro LA. Inflammation and wound healing: the role of the macrophage. Expert Rev Mol Med. 2011;13:e23. | ||
Olczyk P, Mencner Ł, Komosinska-Vassev K. The role of the extracellular matrix components in cutaneous wound healing. Bio Med Res Int. 2014;747584:1–8. | ||
Zhao R, Liang H, Clarke E, Jackson C, Xue M. Inflammation in chronic wounds. Int J Mol Sci. 2016;17(12):2085. | ||
Kanji S, Das H. Advances of stem cell therapeutics in cutaneous wound healing and regeneration. Mediators Inflamm. 2017;2017:5217967. | ||
Gilbert RWD, Vickaryous MK, Viloria-Petit AM. Signalling by transforming growth factor beta isoforms in wound healing and tissue regeneration. J Dev Biol. 2016;4(2):21. | ||
Gökşen S, Balabanli B, Coşkun-Cevher Ş. Application of platelet derived growth factor-BB and diabetic wound healing: the relationship with oxidative events. Free Radic Res. 2017;51(5):498–505. | ||
Gonzalez AC, Costa TF, Andrade ZA, Medrado AR. Wound healing – a literature review. An Bras Dermatol. 2016;91(5):614–620. | ||
Han G, Ceilley R. Chronic wound healing: a review of current management and treatments. Adv Ther. 2017;34(3):599–610. | ||
Caley MP, Martins VLC, O’Toole EA. Metalloproteinases and wound healing. Adv Wound Care (New Rochelle). 2015;4(4):225–234. | ||
Ke J, Wilgus TA. Vascular endothelial growth factor and angiogenesis in the regulation of cutaneous wound repair. Adv Wound Care (New Rochelle). 2014;3(10):647–661. | ||
Liu L, Qing M, Wang Y, Chen S. Defects in graphene: generation, healing and their effects on the properties of graphene: a review. J Mater Sci Technol. 2015;31(6):599–606. | ||
Dai J-F, Wang G-J, Ma L, Wu C-K. Surface properties of graphene: relationship to graphene-polymer composites. Rev Adv Mater Sci. 2015;40:60–71. | ||
Novoselov KS, Geim AK, Morozov SV, et al. Electric field effect in atomically thin carbon films. Science. 2004;306(5696):666–669. | ||
Stankovich S, Dikin DA, Dommett GHB, et al. Graphene-based composite materials. Nature. 2006;442(7100):282–286. | ||
Hernandez Y, Nicolosi V, Lotya M, et al. High-yield production of graphene by liquid-phase exfoliation of graphite. Nat Nanotechnol. 2008;3(9):563–568. | ||
Lotya M, Hernandez Y, King PJ, et al. Liquid phase production of graphene by exfoliation of graphite in surfactant/water solutions. J Am Chem Soc. 2009;131(10):3611–3620. | ||
Mattevi C, Eda G, Agnoli S, et al. Evolution of electrical, chemical, and structural properties of transparent and conducting chemically derived graphene thin films. Adv Funct Mater. 2009;19(16):2577–2583. | ||
de S, King PJ, Lotya M, et al. Flexible, transparent, conducting films of randomly stacked graphene from surfactant-stabilized, oxide-free graphene dispersions. Small. 2010;6(3):458–464. | ||
Eda G, Chhowalla M. Chemically derived graphene oxide: towards large-area thin-film electronics and optoelectronics. Adv Mater. 2010;22(22):2392–2415. | ||
Karu AE, Beer M. Pyrolytic formation of highly crystalline graphite films. J Appl Phys. 1966;37(5):2179–2181. | ||
Eizenberg M, Blakely JM. Carbon monolayer phase condensation on Ni(111). Surf Sci. 1979;82(1):228–236. | ||
Kim KS, Zhao Y, Jang H, et al. Large-scale pattern growth of graphene films for stretchable transparent electrodes. Nature. 2009;457(7230):706. | ||
Reina A, Jia X, Ho J, et al. Large area, few-layer graphene films on arbitrary substrates by chemical vapor deposition. Nano Lett. 2009;9(1):30–35. | ||
Kwon SY, Ciobanu CV, Petrova V, et al. Growth of semiconducting graphene on palladium. Nano Lett. 2009;9(12):3985–3990. | ||
Marchini S, Günther S, Wintterlin J. Scanning tunneling microscopy of graphene on Ru(0001). Phys Rev B. 2007;76(7):075429. | ||
Sutter PW, Flege J-I, Sutter EA. Epitaxial graphene on ruthenium. Nat Mater. 2008;7(5):406–411. | ||
Coraux J, N’Diaye AT, Busse C, Michely T. Structural coherency of graphene on Ir(111). Nano Lett. 2008;8(2):565–570. | ||
Hamilton JC, Blakely JM. Carbon segregation to single crystal surfaces of Pt, PD and CO. Surf Sci. 1980;91(1):199–217. | ||
Land TA, Michely T, Behm RJ, Hemminger JC, Comsa G. STM investigation of single layer graphite structures produced on Pt(111) by hydrocarbon decomposition. Surf Sci. 1992;264(3):261–270. | ||
Sutter P, Sadowski JT, Sutter E. Graphene growth on Pt. Phys Rev B. 2009;80:245411. | ||
Wang Y, Zheng Y, Xu X, et al. Electrochemical delamination of CVD-grown graphene film: toward the recyclable use of copper catalyst. ACS Nano. 2011;5(12):9927–9933. | ||
Li X, Zhu Y, Cai W, et al. Transfer of large-area graphene films for high-performance transparent conductive electrodes. Nano Lett. 2009;9(12):4359–4363. | ||
Liang X, Sperling BA, Calizo I, et al. Toward clean and crackless transfer of graphene. ACS Nano. 2011;5(11):9144–9153. | ||
Lin W, Xu L, Zwingenberger S, Gibon E, Goodman SB, Li G. Mesenchymal stem cells homing to improve bone healing. J Orthop Translat. 2017;9:19–27. | ||
Ciuk T, Pasternak I, Krajewska A, et al. Properties of chemical vapor deposition graphene transferred by high-speed electrochemical delamination. J Phys Chem C. 2013;117(40):20833–20837. | ||
Eckmann A, Felten A, Mishchenko A, et al. Probing the nature of defects in graphene by Raman spectroscopy. Nano Lett. 2012;12(8):3925–3930. | ||
Skoda M, Dudek I, Jarosz A, Szukiewicz D. Graphene: one material, many possibilities – application difficulties in biological systems. J Nanomater. 2014;2014(6):1–11. | ||
Gupta A, Chen G, Joshi P, Tadigadapa S, Eklund PC. Raman scattering from high-frequency phonons in supported n-graphene layer films. Nano Lett. 2006;6(12):2667–2673. | ||
Ferrari AC, Meyer JC, Scardaci V, et al. Raman spectrum of graphene and graphene layers. Phys Rev Lett. 2006;97(18):189401. | ||
Graf D, Molitor F, Ensslin K, et al. Spatially resolved Raman spectroscopy of single- and few-layer graphene. Nano Lett. 2007;7(2):238–242. | ||
Ds L, Riedl C, Krauss B, von Klitzing K, Starke U, Smet JH. Raman spectra of Epitaxial graphene on sic and of epitaxial graphene transferred to SiO2. Nano Lett. 2008;9:4320–4325. | ||
Zh N, Wang YY, Yu T, Shen ZX. Raman spectroscopy and imaging of graphene. Nano Res. 2008;1(4):273. | ||
Pimenta MA, Dresselhaus G, Dresselhaus MS, Cançado LG, Jorio A, Saito R. Studying disorder in graphite-based systems by Raman spectroscopy. Phys Chem Chem Phys. 2007;9(11):1276–1290. | ||
Wang Y, Ni Z, Yu T, et al. Raman studies of monolayer graphene: the substrate effect. J Phys Chem C. 2008;112(29):10637–10640. | ||
Lasocka I, Szulc-Dąbrowska L, Skibniewski M, et al. Biocompatibility of pristine graphene monolayer: scaffold for fibroblasts. Toxicol In Vitro. 2018;48:276–285. | ||
Syama S, Mohanan PV. Safety and biocompatibility of graphene: a new generation nanomaterial for biomedical application. Int J Biol Macromol. 2016;86:546–555. | ||
Jaworski S, Hinzmann M, Sawosz E, et al. Interaction of different forms of graphene with chicken embryo red blood cells. Environ Sci Pollut Res. 2017;24(27):21671–21679. | ||
Majeed W, Bourdo S, Petibone DM, et al. The role of surface chemistry in the cytotoxicity profile of graphene. J Appl Toxicol. 2017;37(4):462–470. | ||
Pelin M, Fusco L, León V, et al. Differential cytotoxic effects of graphene and graphene oxide on skin keratinocytes. Sci Rep. 2017;7:40572. | ||
Duan G, Zhang Y, Luan B, et al. Graphene-induced pore formation on cell membranes. Sci Rep. 2017;7:42767. | ||
Korenev S, Dishman R, Yebra A, et al. Characterization of graphene stripper foils in 11-MeV cyclotrons. Phys Procedia. 2017;90:369–373. | ||
Kalbacova M, Broz A, Kong J, Kalbac M. Graphene substrates promote adherence of human osteoblasts and mesenchymal stromal cells. Carbon. 2010;48(15):4323–4329. | ||
Wang B, Luo PG, Tackett Ii KN, Ruiz ON, Sun Y-P. Graphene oxides as substrate for enhanced mammalian cell growth. J Nanomater Mol Nanotechnol. 2012;1(2):2. | ||
Gurunathan S, Han JW, Eppakayala V, Dayem AA, Kwon D-N, Kim J-H. Biocompatibility effects of biologically synthesized graphene in primary mouse embryonic fibroblast cells. Nanoscale Res Lett. 2013;8(1):1–13. | ||
Gurunathan S, Woong Han J, Kim E, Kwon D-N, Park J-K, Kim J-H. Enhanced green fluorescent protein-mediated synthesis of biocompatible graphene. J Nanobiotechnology. 2014;12(1):1–16. | ||
Conroy J, Verma NK, Smith RJ, et al. Biocompatibility of pristine graphene monolayers, nanosheets and thin films; 2014. Available from: arxiv.org/abs/1406.2497. Accessed March 4, 2019. | ||
Verdanova M, Rezek B, Broz A, et al. Nanocarbon allotropes – graphene and nanocrystalline diamond – promote cell proliferation. Small. 2016;12(18):2499–2509. | ||
Kim J, Kim S, Jung W. Highly enhanced compatibility of human brain vascular pericyte cells on monolayer graphene. Bioengineered. 2017;8(1):85–91. | ||
McCallion C, Burthem J, Rees-Unwin K, Golovanov A, Pluen A. Graphene in therapeutics delivery: problems, solutions and future opportunities. Eur J Pharm Biopharm. 2016;104:235–250. | ||
Huang C, Xu F, Sun Y. Effects of morphology, tension and vibration on wettability of graphene: a molecular dynamics study. Comput Mater Sci. 2017;139:216–224. | ||
Kalbacova MH, Verdanova M, Broz A, Vetushka A, Fejfar A, Kalbac M. Modulated surface of single-layer graphene controls cell behavior. Carbon. 2014;72:207–214. | ||
Xu LC, Siedlecki CA. Effects of surface wettability and contact time on protein adhesion to biomaterial surfaces. Biomaterials. 2007;28(22):3273–3283. | ||
Rosales-Leal JI, Rodríguez-Valverde MA, Mazzaglia G, et al. Effect of roughness, wettability and morphology of engineered titanium surfaces on osteoblast-like cell adhesion. Colloids Surf A Physicochem Eng Asp. 2010;365(1–3):222–229. | ||
Khalili AA, Ahmad MR. A review of cell adhesion studies for biomedical and biological applications. Int J Mol Sci. 2015;16(8):18149–18184. | ||
Gao C, Peng S, Feng P, Shuai C. Bone biomaterials and interactions with stem cells. Bone Res. 2017;5:17059. | ||
Gupta M, Doss B, Lim CT, Voituriez R, Ladoux B. Single cell rigidity sensing: a complex relationship between focal adhesion dynamics and large-scale actin cytoskeleton remodeling. Cell Adh Migr. 2016;10(5):554–567. | ||
Yeh YC, Ling JY, Chen WC, Lin HH, Tang MJ. Mechanotransduction of matrix stiffness in regulation of focal adhesion size and number: reciprocal regulation of caveolin-1 and β1 integrin. Sci Rep. 2017;7(1):15008. | ||
Jesion I, Skibniewski M, Skibniewska E, et al. Graphene and carbon nanocompounds: biofunctionalization and applications in tissue engineering. Biotechnol Biotechnol Equip. 2015;29(3):415–422. | ||
Feller L, Jadwat Y, Khammissa RAG, Meyerov R, Schechter I, Lemmer J. Cellular responses evoked by different surface characteristics of intraosseous titanium implants. Biomed Res Int. 2015; 2015:171945. | ||
James DK, Tour JM. Graphene: powder, flakes, ribbons, and sheets. Acc Chem Res. 2013;46(10):2307–2318. | ||
Gentile F, Tirinato L, Battista E, et al. Cells preferentially grow on rough substrates. Biomaterials. 2010;31(28):7205–7212. | ||
Liu N, Pan Z, Fu L, Zhang C, Dai B, Liu Z. The origin of wrinkles on transferred graphene. Nano Res. 2011;4(10):996–1004. | ||
Wang W, Yang S, Wang A. Observation of the unexpected morphology of graphene wrinkle on copper substrate. Sci Rep. 2017;7(1):8244. | ||
Deng S, Berry V. Wrinkled, rippled and crumpled graphene: an overview of formation mechanism, electronic properties, and applications. Mater Today. 2015;19(4):198–213. | ||
Huang Y, Chen X, Zhang MQ. Reversible surface wettability conversion of graphene films: optically controlled mechanism. J Mater Sci. 2014;49(8):3025–3033. | ||
Ramakrishna Matte HSS, Subrahmanyam KS, Rao CNR. Synthetic aspects and selected properties of graphene. Nanomater Nanotechnol. 2011;1:5–13. | ||
Kalbacova M, Broz A, Kalbac M. Influence of the fetal bovine serum proteins on the growth of human osteoblast cells on graphene. J Biomed Mater Res A. 2012;100A(11):3001–3007. | ||
Tan XW, Thompson B, Konstantopoulos A, et al. Application of graphene as candidate biomaterial for synthetic keratoprosthesis skirt. Invest Ophthalmol Vis Sci. 2015;56(11):6605–6611. | ||
Sayyar S, Murray E, Thompson BC, Gambhir S, Officer DL, Wallace GG. Covalently linked biocompatible graphene/polycaprolactone composites for tissue engineering. Carbon. 2013;52:296–304. | ||
Provenzano PP, Keely PJ. Mechanical signaling through the cytoskeleton regulates cell proliferation by coordinated focal adhesion and Rho GTPase signaling. J Cell Sci. 2011;124(8):1195–1205. | ||
Hanna S, El-Sibai M. Signaling networks of Rho GTPases in cell motility. Cell Signal. 2013;25(10):1955–1961. | ||
Hoon J, Tan M, Koh C-G. The regulation of cellular responses to mechanical cues by Rho GTPases. Cells. 2016;5(2):E17. | ||
Schaller MD. Cellular functions of FAK kinases: insight into molecular mechanisms and novel functions. J Cell Sci. 2010;123(Pt 7):1007–1013. | ||
Kim S, Kim M, Shin Y, et al. Cell migration according to shape of graphene oxide micropatterns. Micromachines (Basel). 2016;7(10):E186. | ||
Zhang H, Hou R, Xiao P, et al. Single cell migration dynamics mediated by geometric confinement. Colloids Surf B Biointerfaces. 2016;145:72–78. | ||
Keshavan S, Oropesa-Nuñez R, Diaspro A, Canale C, Dante S. Adhesion and migration of CHO cells on micropatterned single layer graphene. 2D Materials. 2017;4(2):025022. | ||
Lorenzoni M, Brandi F, Dante S. Simple and effective graphene laser processing for neuron patterning application. Sci Rep. 2013;3:1954. | ||
Guo X, Mei N. Assessment of the toxic potential of graphene family nanomaterials. J Food Drug Anal. 2014;22(1):105–115. | ||
Hegab HM, Elmekawy A, Zou L, Mulcahy D, Saint CP, Ginic-Markovic M. The controversial antibacterial activity of graphene-based materials. Carbon. 2016;105:362–376. | ||
Zou X, Zhang L, Wang Z, Luo Y. Mechanisms of the antimicrobial activities of graphene materials. J Am Chem Soc. 2016;138(7):2064–2077. | ||
Wang L, Hu C, Shao L. The antimicrobial activity of nanoparticles: present situation and prospects for the future. Int J Nanomedicine. 2017;12:1227–1249. | ||
Gurunathan S, Han JW, Dayem AA, Eppakayala V, Kim JH. Oxidative stress-mediated antibacterial activity of graphene oxide and reduced graphene oxide in Pseudomonas aeruginosa. Int J Nanomedicine. 2012;7:5901–5914. | ||
Song C, Yang C-M, Sun X-F, et al. Influences of graphene oxide on biofilm formation of Gram-negative and Gram-positive bacteria. Environ Sci Pollut Res. 2018;25(3):2853–2860. | ||
Liu S, Zeng TH, Hofmann M, et al. Antibacterial activity of graphite, graphite oxide, graphene oxide, and reduced graphene oxide: membrane and oxidative stress. ACS Nano. 2011;5(9):6971–6980. | ||
Nanda SS, Yi DK, Kim K. Study of antibacterial mechanism of graphene oxide using Raman spectroscopy. Sci Rep. 2016;6(1):1–12. | ||
Shuai C, Guo W, Wu P, et al. A graphene oxide-Ag co-dispersing nanosystem: dual synergistic effects on antibacterial activities and mechanical properties of polymer scaffolds. Chem Eng J. 2018;347(347):322–333. | ||
Tu Y, Lv M, Xiu P, et al. Destructive extraction of phospholipids from Escherichia coli membranes by graphene nanosheets. Nat Nanotechnol. 2013;8(8):594–601. | ||
Kurantowicz N, Sawosz E, Jaworski S, et al. Interaction of graphene family materials with Listeria monocytogenes and Salmonella enterica. Nanoscale Res Lett. 2015;10(1):1–12. | ||
Barbolina I, Woods CR, Lozano N, Kostarelos K, Novoselov KS, Roberts IS. Purity of graphene oxide determines its antibacterial activity. 2D Materials. 2016;3(2):025025. | ||
Ye S, Shao K, Li Z, et al. Antiviral activity of graphene oxide: how sharp edged structure and charge matter. ACS Appl Mater Interfaces. 2015;7(38):21571–21579. | ||
Song Z, Wang X, Zhu G, et al. Virus capture and destruction by label-free graphene oxide for detection and disinfection applications. Small. 2015;11(9–10):1171–1176. | ||
Mukherjee S, Sriram P, Barui AK, et al. Graphene oxides show angiogenic properties. Adv Healthc Mater. 2015;4(11):1722–1732. | ||
Park J, Kim YS, Ryu S, et al. Graphene potentiates the myocardial repair efficacy of mesenchymal stem cells by stimulating the expression of angiogenic growth factors and gap junction protein. Adv Funct Mater. 2015;25(17):2590–2600. | ||
Shine M-Y, Chiang W-H, Chen I-W, Liu W-Y, Chen Y-W. Synergistic acceleration in the osteogenic and angiogenic differentiation of human mesenchymal stem cells by calcium silicate-graphene composites. Mater Sci Eng C. 2017;73:726–735. | ||
Singh SK, Singh MK, Nayak MK, et al. Thrombus inducing property of atomically thin graphene oxide sheets. ACS Nano. 2011;5(6):4987–4996. | ||
Saleem J, Wang L, Chen C. Immunological effects of graphene family nanomaterials. NanoImpact. 2017;5:109–118. | ||
Wilgus TA, Roy S, McDaniel JC. Neutrophils and wound repair: positive actions and negative reactions. Adv Wound Care (New Rochelle). 2013;2(7):379–388. | ||
Vinish M, Cui W, Stafford E, et al. Dendritic cells modulate burn wound healing by enhancing early proliferation. Wound Rep and Reg. 2016;24(1):6–13. | ||
Mukherjee SP, Lazzaretto B, Hultenby K, et al. Graphene oxide elicits membrane lipid changes and neutrophil extracellular trap formation. Chem. 2018;4(2):334–358. | ||
Kaplan MJ, Radic M. Neutrophil extracellular traps: double-edged swords of innate immunity. J Immun. 2012;189(6):2689–2695. | ||
Zhou H, Zhao K, Li W, et al. The interactions between pristine graphene and macrophages and the production of cytokines/chemokines via TLR- and NF-κB-related signaling pathways. Biomaterials. 2012;33(29):6933–6942. | ||
Chen GY, Yang HJ, Lu CH, et al. Simultaneous induction of autophagy and toll-like receptor signaling pathways by graphene oxide. Biomaterials. 2012;33(27):6559–6569. | ||
Ma J, Liu R, Wang X, et al. Crucial role of lateral size for graphene oxide in activating macrophages and stimulating pro-inflammatory responses in cells and animals. ACS Nano. 2015;9(10):10498–10515. | ||
Orecchioni M, Bedognetti D, Newman L, et al. Single-cell mass cytometry and transcriptome profiling reveal the impact of graphene on human immune cells. Nat Commun. 2017;8(1):1109. | ||
Tkach AV, Yanamala N, Stanley S, et al. Graphene oxide, but not fullerenes, targets immunoproteasomes and suppresses antigen presentation by dendritic cells. Small. 2013;9(9–10):1686–1690. | ||
Zhi X, Fang H, Bao C, et al. The immunotoxicity of graphene oxides and the effect of PVP-coating. Biomaterials. 2013;34(21):5254–5261. | ||
Wang W, Li Z, Duan J, Wang C, Fang Y, Yang X-D. In vitro enhancement of dendritic cell-mediated anti-glioma immune response by graphene oxide. Nanoscale Res Lett. 2014;9(1):311. | ||
Zhong C, Shi D, Zheng Y, Nelson PJ, Bao Q. Successive release of tissue inhibitors of metalloproteinase-1 through graphene oxide-based delivery system can promote skin regeneration. Nanoscale Res Lett. 2017;12:533. | ||
Singh A, Singh A, Sen D. Mesenchymal stem cells in cardiac regeneration: a detailed progress report of the last 6 years (2010–2015). Stem Cell Res Ther. 2016;7(1):82. | ||
Kenry, Lee WC, Loh KP, Lim CT. When stem cells meet graphene: opportunities and challenges in regenerative medicine. Biomaterials. 2018;155:236–250. | ||
Al-Shaibani MBH, Wang X-N, Lovat PE, Dickinson AM. Cellular therapy for wounds: applications of mesenchymal stem cells in wound healing. In: Alexandrescu V, editor. Wound Healing – New Insights into Ancient Challenges. London: InTech; 2016. | ||
Li M, Luan F, Zhao Y, et al. Mesenchymal stem cell-conditioned medium accelerates wound healing with fewer scars. Int Wound J. 2017;14(1):64–73. | ||
Alcayaga-Miranda F, Cuenca J, Khoury M. Antimicrobial activity of mesenchymal stem cells: current status and new perspectives of antimicrobial peptide-based therapies. Front Immunol. 2017;8(21507):339. | ||
Lin W-H, Chen T-H, Chang J-K, et al. A direct and polymer-free method for transferring graphene grown by chemical vapor deposition to any substrate. ACS Nano. 2014;8(2):1784–1791. | ||
Hamdan S, Pastar I, Drakulich S, et al. Nanotechnology-driven therapeutic interventions in wound healing: potential uses and applications. ACS Cent Sci. 2017;3(3):163–175. | ||
Nuschke A. Activity of mesenchymal stem cells in therapies for chronic skin wound healing. Organogenesis. 2014;10(1):29–37. | ||
Tamama K, Kerpedjieva SS. Acceleration of wound healing by multiple growth factors and cytokines secreted from multipotential stromal cells/mesenchymal stem cells. Adv Wound Care (New Rochelle). 2012;1(4):177–182. | ||
Zhang J, Huang X, Wang H, et al. The challenges and promises of allogeneic mesenchymal stem cells for use as a cell-based therapy. Stem Cell Res Ther. 2015;6(1):234. | ||
Walter MNM, Wright KT, Fuller HR, Macneil S, Johnson WEB. Mesenchymal stem cell-conditioned medium accelerates skin wound healing: an in vitro study of fibroblast and keratinocyte scratch assays. Exp Cell Res. 2010;316(7):1271–1281. | ||
Eming SA, Martin P, Tomic-Canic M. Wound repair and regeneration: mechanisms, signaling, and translation. Sci Transl Med. 2014;6(265):265sr6. | ||
Steffens D, Mathor MB, Santi BT, Luco DP, Pranke P. Development of a biomaterial associated with mesenchymal stem cells and keratinocytes for use as a skin substitute. Regen Med. 2015;10(8):975–987. | ||
An Y, Liu WJ, Xue P, et al. Autophagy promotes MSC-mediated vascularization in cutaneous wound healing via regulation of VEGF secretion. Cell Death Dis. 2018;9(2):1–14. | ||
Latifi-Pupovci H, Kuçi Z, Wehner S, et al. In vitro migration and proliferation (“wound healing”) potential of mesenchymal stromal cells generated from human CD271+ bone marrow mononuclear cells. J Transl Med. 2015;13(1):315. | ||
Kazantseva J, Ivanov R, Gasik M, Neuman T, Hussainova I. Graphene-augmented nanofiber scaffolds demonstrate new features in cells behaviour. Sci Rep. 2016;6(1):30150. | ||
Jastrzebska E, Tomecka E, Jesion I. Heart-on-a-chip based on stem cell biology. Biosens Bioelectron. 2016;75:67–81. | ||
Brzozka Z, Jastrzebska E. Cardiac Cell Culture Technologies. Cham: Springer International Publishing; 2018. | ||
Ennis WJ, Sui A, Bartholomew A. Stem cells and healing: impact on inflammation. Adv Wound Care (New Rochelle). 2013;2(7):369–378. | ||
Chen D, Hao H, Fu X, Han W. Insight into reepithelialization: how do mesenchymal stem cells perform? Stem Cells Int. 2016;2016:6120173. | ||
Seo BF, Jung S-N. The immunomodulatory effects of mesenchymal stem cells in prevention or treatment of excessive scars. Stem Cells Int. 2016;2016(2):1–8. | ||
Yew TL, Hung YT, Li HY, et al. Enhancement of wound healing by human multipotent stromal cell conditioned medium: the paracrine factors and p38 MAPK activation. Cell Transplant. 2011;20(5):693–706. | ||
Bortolotti F, Ukovich L, Razban V, et al. In vivo therapeutic potential of mesenchymal stromal cells depends on the source and the isolation procedure. Stem Cell Reports. 2015;4(3):332–339. | ||
Wu Y, Chen L, Scott PG, Tredget EE. Mesenchymal stem cells enhance wound healing through differentiation and angiogenesis. Stem Cells. 2007;25(10):2648–2659. | ||
Shin S-C, Seo Y, Park HY, et al. Regenerative potential of tonsil mesenchymal stem cells on surgical cutaneous defect. Cell Death Dis. 2018;9(2):1–12. | ||
Luo G, Cheng W, He W, et al. Promotion of cutaneous wound healing by local application of mesenchymal stem cells derived from human umbilical cord blood. Wound Repair Regen. 2010;18(5):506–513. | ||
Finnson KW, McLean S, Di Guglielmo GM, Philip A. Dynamics of transforming growth factor beta signaling in wound healing and scarring. Adv Wound Care (New Rochelle). 2013;2(5):195–214. | ||
Rennert RC, Sorkin M, Garg RK, Gurtner GC. Stem cell recruitment after injury: lessons for regenerative medicine. Regen Med. 2012;7(6):833–850. | ||
Spees JL, Lee RH, Gregory CA. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res Ther. 2016;7(1):125. | ||
Lee EY, Xia Y, Kim W-S, et al. Hypoxia-enhanced wound-healing function of adipose-derived stem cells: increase in stem cell proliferation and up-regulation of VEGF and bFGF. Wound Repair Regen. 2009;17(4):540–547. | ||
Rodriguez-Menocal L, Salgado M, Ford D, Van Badiavas E. Stimulation of skin and wound fibroblast migration by mesenchymal stem cells derived from normal donors and chronic wound patients. Stem Cells Transl Med. 2012;1(3):221–229. | ||
Mishra PJ, Mishra PJ, Banerjee D. Keratinocyte induced differentiation of mesenchymal stem cells into dermal myofibroblasts: a role in effective wound healing. Int J Transl Sci. 2016;(1):5–32. | ||
Sasaki M, Abe R, Fujita Y, Ando S, Inokuma D, Shimizu H. Mesenchymal stem cells are recruited into wounded skin and contribute to wound repair by transdifferentiation into multiple skin cell type. J Immunol. 2008;180(4):2581–2587. | ||
Estrada JC, Torres Y, Bengurãa A, et al. Human mesenchymal stem cell-replicative senescence and oxidative stress are closely linked to aneuploidy. Cell Death Dis. 2013;4(6):e691. | ||
Turinetto V, Vitale E, Giachino C. Senescence in human mesenchymal stem cells: functional changes and implications in stem cell-based therapy. Int J Mol Sci. 2016;17(7):1164. | ||
Li Y, Wu Q, Wang Y, Li L, Bu H, Bao J. Senescence of mesenchymal stem cells (review). Int J Mol Med. 2017;39(4):775–782. | ||
Wagner W, Horn P, Castoldi M, et al. Replicative senescence of mesenchymal stem cells: a continuous and organized process. PLoS One. 2008;3(5):e2213. | ||
Whitfield MJ, Lee WCJ, Van Vliet KJ. Onset of heterogeneity in culture-expanded bone marrow stromal cells. Stem Cell Res. 2013;11(3):1365–1377. | ||
Kundrotas G, Gasperskaja E, Slapsyte G, et al. Identity, proliferation capacity, genomic stability and novel senescence markers of mesenchymal stem cells isolated from low volume of human bone marrow. Oncotarget. 2016;7(10):10788–10802. | ||
Li Z, Wang H, Yang B, Sun Y, Huo R. Three-dimensional graphene foams loaded with bone marrow derived mesenchymal stem cells promote skin wound healing with reduced scarring. Mater Sci Eng C Mater Biol Appl. 2015;57:181–188. | ||
Chu J, Shi P, Yan W, et al. PEGylated graphene oxide-mediated quercetin-modified collagen hybrid scaffold for enhancement of MSCs differentiation potential and diabetic wound healing. Nanoscale. 2018;10(20):9547–9560. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.