Back to Journals » Journal of Inflammation Research » Volume 9
The association between pro- and anti-inflammatory cytokine polymorphisms and periventricular leukomalacia in newborns with hypoxic-ischemic encephalopathy
Authors Gabriel ML, Del Campo Braojos Braga F, Ribeiro Rodero Cardoso M, Polli Lopes AC, Piatto V, Soares Souza A
Received 6 January 2016
Accepted for publication 3 February 2016
Published 5 May 2016 Volume 2016:9 Pages 59—67
DOI https://doi.org/10.2147/JIR.S103697
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Marta Lúcia Gabriel,1 Fernanda Braojos Braga,1 Mariana Rodero Cardoso,1 Ana Cláudia Lopes,2 Vânia Belintani Piatto,2 Antônio Soares Souza1
1Radiology Department, 2Morphology Department, São José do Rio Preto Medical School, FAMERP, São Paulo, Brazil
Background: Periventricular leukomalacia (PVL) is a frequent consequence of hypoxic-ischemic injury. Functional cytokine gene variants that result in altered production of inflammatory (tumor necrosis factor-alpha [TNF-α] and interleukin-1beta [IL-1β]) or anti-inflammatory (interleukin-10 [IL-10]) cytokines may modify disease processes, including PVL.
Objective: The aim of this study was to evaluate if there is a relationship between the two proinflammatory polymorphisms (TNF-σ-1031T/C and IL-1 β-511C/T) and the anti-inflammatory polymorphism IL-10-1082G/A and PVL risk in Brazilian newborns with and without this injury.
Materials and methods: A cross-sectional case-control study performed at the Neonatal Intensive Care Unit of the Children's Hospital and Maternity of the São José do Rio Preto Medical School (FAMERP). Fifty preterm and term newborns were examined as index cases and 50 term newborns as controls, of both sexes for both groups. DNA was extracted from peripheral blood leukocytes, and the sites that encompassed the three polymorphisms were amplified by polymerase chain reaction-restriction fragment length polymorphism.
Results: Gestational age ranged from 25 to 39 weeks, in the case group, and in the control group it ranged from 38 to 42.5 weeks (P<0.0001). Statistically significant association was found between TNF-α-1031T/C high expression genotype TC (odds ratio [OR], 2.495; 95% confidence interval [CI], 1.10–5.63; P=0.043) as well as between genotypes (TC + CC) (OR, 2.471; 95% CI, 1.10–5.55; P=0.044) and risk of PVL. Statistically significant association was found between IL-1β-511C/T high expression genotypes (CT + TT) (OR, 23.120; 95% CI, 1.31–409.4; P=0.003) and risk of PVL. Statistically significant association between IL-10-1082G/A high expression genotype GG (OR, 0.07407; 95% CI, 0.02–0.34; P<0.0001) as well as between IL-10-1082G high expression allele (OR, 0.5098; 95% CI, 0.29–0.91; P=0,030) and PVL reduced risk was observed. There was a statistically significant association between TC/CT/GA genotype combination and the risk of PVL (OR, 6.469; 95% CI, 2.00–20.92; P=0.001).
Conclusion: There is evidence of an association between the polymorphisms TNF-α-1031T/C, IL-1β-511C/T, and IL-10-1082G/A and PVL risk in this Brazilian newborn population studied.
Keywords: leukomalacia periventricular, tumor necrosis factor-alpha, interleukin-10, interleukin-1beta, polymorphism single nucleotide
Introduction
Periventricular leukomalacia (PVL) is a frequent consequence of hypoxic-ischemic lesion and its occurrence is 0.2 cases per 1,000 births. This injury occurs mostly in premature newborns, which is the main clinicopathologic correlation of hypoxic-ischemic encephalopathy in these cases. Patients with PVL have a higher risk of motor and cognitive delays, seizures, and visual abnormalities, and spastic diplegia as the main sequelae.1–4
PVL has also been recognized in infants born at term (term-born PVL: gestational age >39 weeks), with a distinct clinical condition from that of the preterm population with this lesion. Among children who had neonatal anoxia, PVL was diagnosed in 14%–85% of cases and 30% to 60% of them have spastic diplegia as a consequence.1,5–12
The greatest period of risk for PVL corresponds to the time when oligodendrocyte precursors dominate within cerebral white matter, proliferate and differentiate, prior to active synthesis of myelin sheaths (approximating to 23–32 weeks post-conception in humans). The principal mechanisms responsible for damage to the oligodendrocytes (induced by hypoxia/ischemia) include glutamate excitotoxicity, inflammatory processes mediated by both free radical and cytokines, and by the difficulty of regulation of the oxidative stress caused by lack of production of antioxidant enzymes in oligodendrocytes.2,3,13
The first line of defense to protect against encephalic injuries is the quick response from activated microglia, because of their rapid transformation from a ramified to an amoeboid shape, a process that rapidly happens within minutes to a few hours after the initial injury. Because of this, it has been recognized that activated microglia are related to many encephalon diseases, including PVL.2,14–18
The involvement of cytokines in the pathophysiology of perinatal encephalic injury and pathogenesis of PVL is sustained by epidemiological and clinical data. For normal encephalic development during gestation, and in the physiology of pregnancy, the cytokine network has an extremely important role. In adverse conditions, both astroglia and microglia produce nitric oxide and excitatory amino acids, due to the stimulation of cytokines, in particular tumor necrosis factor (TNF) and interleukin interleukin-1beta (IL-1β), resulting in a neurotoxic effect.3,13,15,19–21
TNF-α is encoded by a highly polymorphic gene, which is located on chromosome 6q. Inherited polymorphisms in its promoter region may contribute to changes in its immune response as well as genetic predisposition to complicated illnesses, such as PVL. The T to C substitution at position -1031 in the TNF-α gene promoter region has been associated with high levels of TNF-α, and ~60% of the difference in the production of TNF-α is genetically determined.21–25
The IL-1β gene is located in the IL-1 cluster on chromosome 2q. IL-1β levels are regulated at transcriptional level and the polymorphism -511C/T in the IL-1β gene promoter region has been found not only to influence levels of IL-1β but also to change the predisposition to several diseases, including PVL. The IL-1β has been considered a key mediator in hypoxic-ischemic encephalic injury. IL-1β and TNF-α stimulate production of other cytokines, endothelial adhesion molecules, and nitric oxide in the central nervous system (CNS).21,22,24,26,27
In contrast, anti-inflammatory cytokines, such as interleukin-10 (IL-10), may have a neuroprotective role in the CNS. IL-10 inhibits IL-1β and endotoxin-induced cytokine production by glial cells.28 The magnitude of IL-10 secretion is partially genetically determined by several polymorphisms in the IL-10 gene promoter region, which is located on chromosome 1q. The polymorphism at -1082 (G to A) is thought to be a major determinant of the magnitude of IL-10 secretion and may influence outcome in several disease states.25,29,30
Therefore, the aim of this study was to evaluate if there is a relationship between the two proinflammatory polymor (TNF-α-1031T/C and IL-1β-511C/T) and the anti-inflammatory polymorphism (IL-10-1082G/A) with PVL risk in Brazilian newborns with and without this injury.
Materials and methods
A cross-sectional case-control study at the Neonatal Intensive Care Unit (NICU) of the Children’s Hospital and Maternity of the São José do Rio Preto Medical School (FAMERP, São Paulo, Brazil), between April and July 2015, was performed.
Among the total of 85 newborns consecutively admitted in the Neonatal Intensive Care Unit, during the study period, 50 newborns were selected (59%), of both sexes, with transfontanellar ultrasonography diagnosis of PVL, which was called the case group. The remaining 35 (41%) newborns were not selected because they had presented one of the exclusion criterion or died during the study period. If applicable, according to the protocol, the infants received therapeutic hypothermia.
For the case selection, the following criteria were considered:
Inclusion criteria (with mandatory presence of the criteria 4 and 5):
- Signs of fetal distress revealed by intrapartum monitoring, such as persistent decelerations and/or sustained fetal bradycardia or the presence of meconium-stained amniotic fluid.
- Apgar score ≤4 or 5 during the first minute of life.
- Need for positive pressure ventilation for at least 2 minutes in order to start breathing.
- Diagnosis of hypoxic-ischemic encephalopathy established by the presence of perinatal asphyxia associated with neurological manifestations due to hypoxia and ischemia (one or more abnormal neurological signs, such as irritability, tremors, altered consciousness, muscle tone changes, and seizures in the first 24 hours of life).31
- Diagnosis of PVL obtained by neuroimaging transfontanellar ultrasonography.
The control group consisted of 50 newborns of both sexes, born in the same period, without hypoxic-ischemic encephalopathy, Apgar score ≥7 during the first minute of life and no alteration to the transfontanellar ultrasonography.
Newborns from both groups with clinical or laboratory suspicion of congenital infection, presence of sepsis and/or meningitis, congenital malformations, presence of seizures unrelated to hypoxic events and of unknown etiology, maternal history of drug abuse, mothers with any infection of STORCH (syphilis, toxoplasmosis, rubella virus, cytomegalovirus, herpes virus) group during pregnancy or seropositivity to HIV, and maternal use of opioid or respiratory depressive drugs administered in the peripartum period were excluded.
Newborn ethnicity was based on skin color (white, nonwhite, black, or mulatto), and all of them were from the same geographic area – São Paulo State, Brazil.
Medical histories were collected with regard to demographic data, clinical presentation, and imaging data. All newborn in the case group were classified per De Vries rating scale,32 in relation to PVL ultrasonographic aspects.
Neuroimaging assessment
Transfontanellar ultrasonographic examination with a 7.5 MHz transducer (SONOS-5500; Philips Medical Systems, Andover, MA, USA) was used to diagnose PVL. Either the presence of a cyst in the periventricular white matter within 7 days of delivery or an increased echodensity in the periventricular white matter within 6 hours of delivery confirmed the diagnosis of PVL.32 The diagnosis of PVL was reconfirmed using magnetic resonance imaging. All the neuroimaging tests were performed in a uniform fashion, and the clinical interpretations were provided by a radiologist specialized in neuroradiology.
Molecular analysis
Genomic DNA was isolated from peripheral blood leukocytes using the Illustra Blood Genomic Prep Mini Spin Kit (GE Healthcare UK Limited™, Little Chalfont Bucks, UK) per the manufacturer’s protocol.
To detect the three polymorphisms −1031T/C, −511C/T, and -1082G/A, nuclear DNA fragments that encompassed the polymorphics sites, respectively, in the TNF-α gene (Gene Bank NCBI RefSeqGene: NG_007462.1), IL-1β gene (Gene Bank NCBI RefSeqGene: AY137079), and IL-10 gene (Gene Bank NCBI RefSeqGene: X78437.2) were amplified by polymerase chain reaction (PCR) and digested, by RFLP, with 5U of their respective restriction enzymes (New England Biolab™; Hertfordshire, UK.) – BbsI for 2.5 hours at 37°C, AvaI for 2.5 hours at 37°C, and BsmFI for 2.5 hours at 65°C.
PCR conditions
Standard PCR was performed in 25 μl, containing 200 ng genomic DNA, 10 pmol of each primer, and FideliTaq™ PCR Master Mix (2×) (GE Healthcare UK Limited™), per the manufacturer’s protocol. Published primers sequences were used to analyze the three polymorphisms −1031T/C, −511C/T, and −1082G/A,21,33,34 and PCR was performed as follows: initial denaturation at 95°C for 2 minutes, followed by 35 cycles of denaturation at 95°C for 15 seconds, annealing at their respective degrees 53°C, 51°C, and 61°C for 30 seconds, and extension at 72°C for 50 seconds, followed by a final extension for 10 minutes at 72°C.
The products from both PCR/RFLP reactions were added to FlashGel™ Loading Dye 5x, analyzed by electrophoresis on a 2.2% agarose FlashGel™ DNA Cassette, and all gels were photodocumented by FlashGel™ Camera (Lonza Group Ltd, Basel, Switzerland).
The −1031T wild-type allele from TNF-α gene was represented by an uncut 150 bp PCR product, and the −1031C polymorphic allele consisted of two fragments at 62 and 88 bp. The −1511C wild-type allele from IL-1β gene consisted of two fragments at 115 and 190 bp, and the −1511T polymorphic allele was represented by an uncut 305 bp PCR product. The -1082G wild-type allele from IL-10 gene consisted of two fragments at 80 and 320 bp, and the −1082A polymorphic allele was represented by an uncut 400 bp PCR product.
To avoid bias in the molecular analysis and final results, all DNA samples were analyzed without the knowledge of the patients’ clinical and ultrasonographic characteristics.
Ethical aspects
The ethics and research protocol were approved by the Research Ethics Committee (FAMERP, São Paulo, Brazil) (#216.753/2013). Before initiation of any procedure, signed informed consent was obtained from all parents.
Statistical analysis
Statistical analyses were performed using Graph Pad Instat, version 3.00 (GraphPad Software Inc, San Diego, CA, USA). The results were previously submitted to descriptive statistics to determine the normal range. The Mann–Whitney test was used for independent samples with nonnormal distribution. Qualitative variables were expressed as a percentage (%) and continuous variables were expressed as a mean and standard deviation. Where applicable, chi-square or Fisher’s exact test was used to compare the variables. The odds ratio (OR) with 95% confidence interval (CI) was used to determine the risk factor for PVL by the polymorphisms analyzed. Results were considered statistically significant when the probability of findings occurring by chance was <5%.
Results
This cross-sectional case-control study included 100 newborns of both sexes. TNF-α, IL-1β, and IL-10 polymorphisms were genotyped in all of them. The case group was composed of 50 newborns with PVL (31 males and 19 females) while 50 newborns (23 males and 27 females) constituted the control group. Gestational age ranged from 25 to 39 weeks, with a median of 31 weeks in case group, and at control group the gestational age ranged from 38 to 42.5 weeks, with a median of 39.1 weeks (P<0.0001). All newborns from both groups had white skin color (100%). Clinical and demographics data are summarized in Table 1.
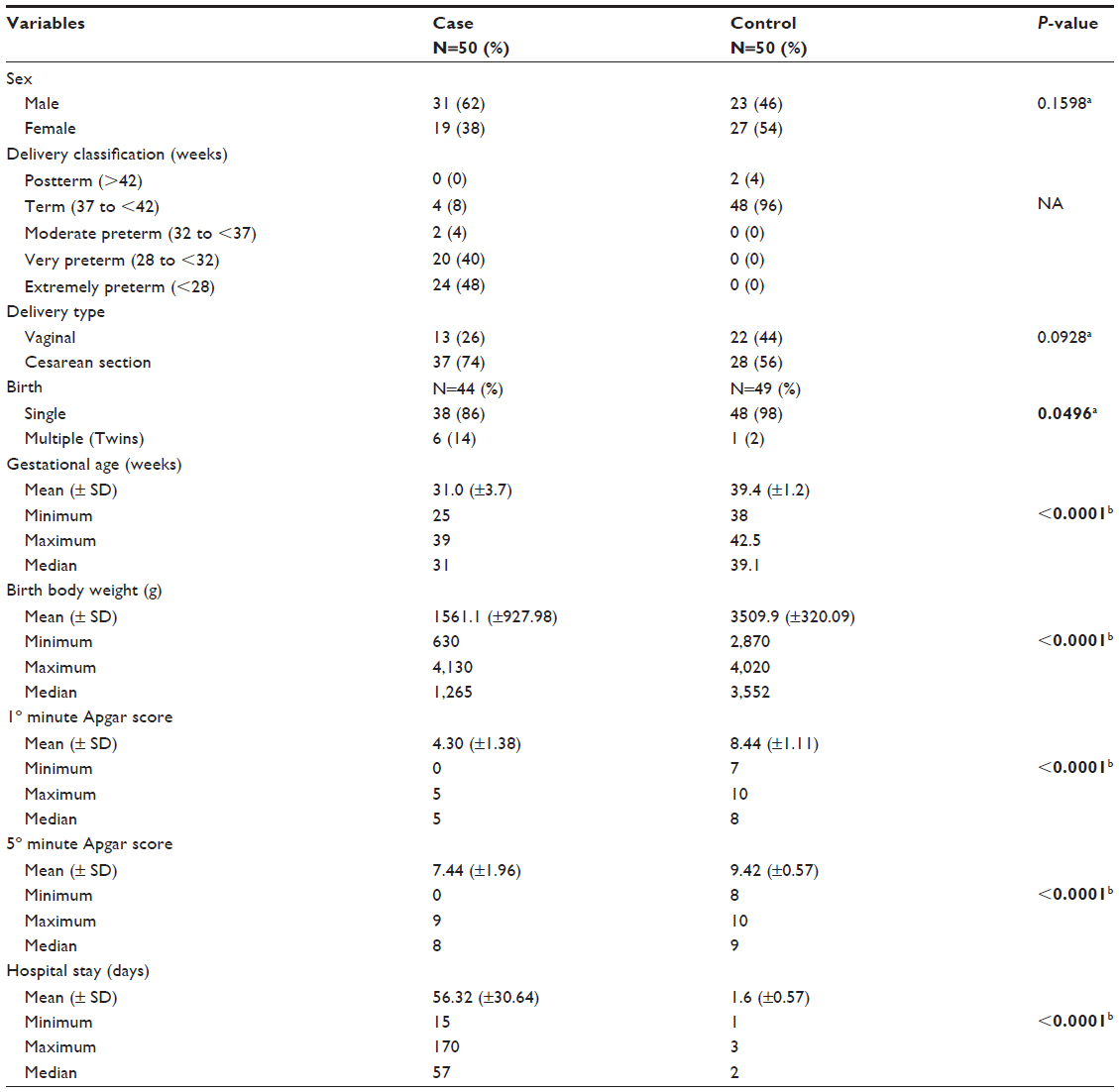 | Table 1 Distribution of the characteristics of case and control groups |
Transfontanellar ultrasonographic assessment
Transfontanellar ultrasonographic data of case group are presented in Table 2.
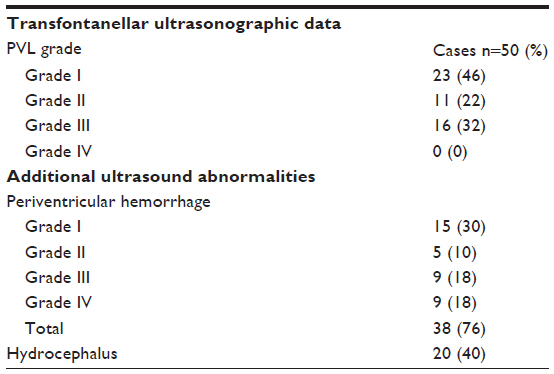 | Table 2 Distribution of transfontanellar ultrasonographic data in the case group |
Cytokine gene polymorphisms assessment
Table 3 shows the genotype and allele distributions for all analyzed polymorphisms in all examined newborns of both case and control groups (N=100). Statistically significant association was found between TNF-α-1031T/C high expression genotype TC (OR, 2.495; 95% CI, 1.10–5.63; P=0.043) as well as between genotypes (TC+CC) (OR, 2.471; 95% CI, 1.10–5.55; P=0.044) and risk of PVL. Statistically significant association between TNF-α-1031T/C high expression genotype TT (OR, 0.408; 95% CI, 0.18–0.91; P=0.044) and PVL reduced risk was also observed. Statistically significant association was found between IL-1β-511C/T high expression genotypes (CT+TT) (OR, 23.120; 95% CI, 1.31–409.4; P=0.003) and risk of PVL. Statistically significant association was found between IL-1β-511C/T high expression genotype CC (OR, 0.04325; 95% CI: 0.002–0.76; P=0.003) and PVL reduced risk. Statistically significant association between IL-10-1082G/A high expression genotype GG (OR, 0.07407; 95% CI, 0.02–0.34; P<0.0001) as well as between IL-10-1082G high expression allele (OR, 0.5098; 95% CI, 0.29–0.91; P=0,030) and PVL reduced risk was observed. Statistically significant association was found between IL-10-1082G/A high expression genotype GA (OR, 13.500; 95% CI, 2.93–62.2; P<0.0001) as well as IL-10-1082A high expression allele (OR, 1.962; 95% CI, 1.10–3.49; P=0.030) and risk of PVL.
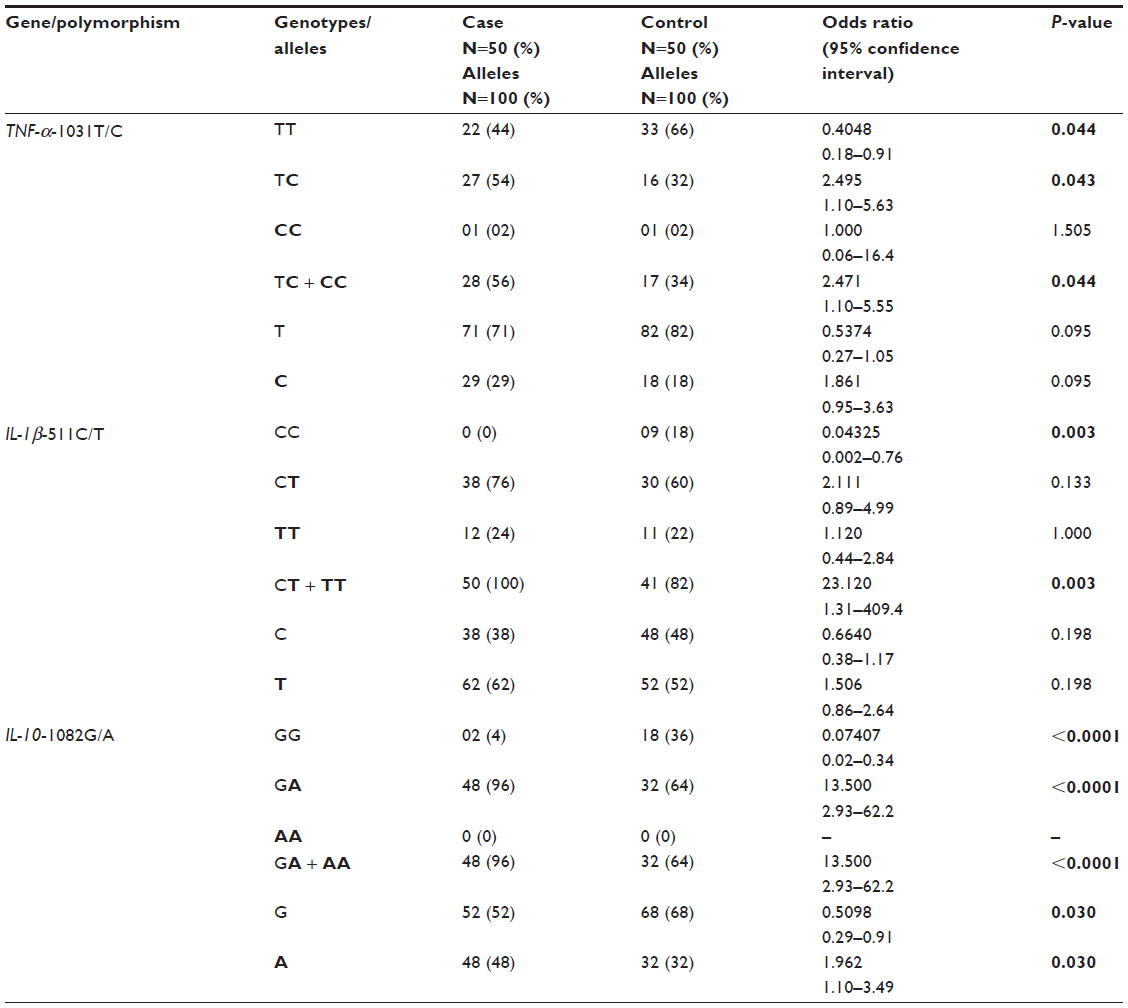 | Table 3 Distribution of genotypes and alleles in case and control groups for the TNF-a-1031T/C, IL-1β-511C/T, and IL-10-1082G/A polymorphisms |
Table 4 shows the distributions among TNF-α-1031T/C, IL-1β-511C/T, and IL-10-1082G/A genotype combinations, in this respective sequence, in all analyzed newborns of both case and control groups (N=100). There was a statistically significant association between TC/CT/GA genotypes combination and the risk of PVL (OR, 6.469; 95% CI, 2.00–20.92; P=0.001). Statistically significant association was found between TT/CC/GA genotypes combination and PVL reduced risk (OR, 0.06778; 95% CI, 0.004–1.24; P=0.027).
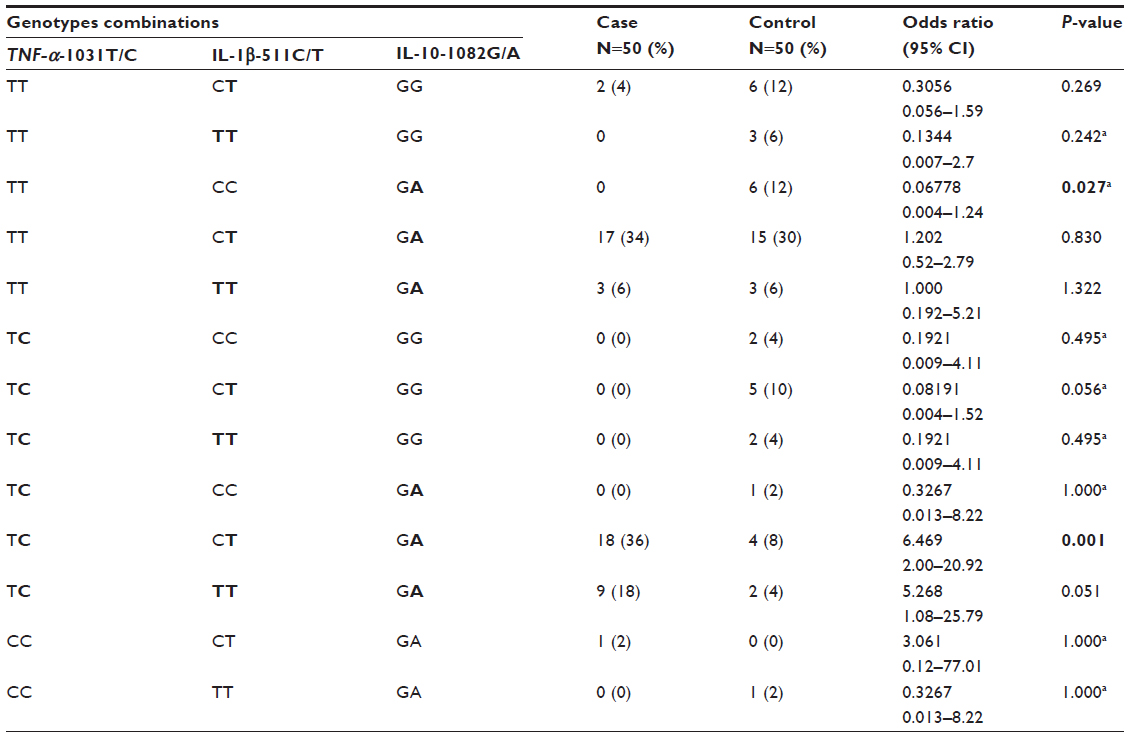 | Table 4 Distribution among TNF-α-1031T/C, IL-1β-511C/T, and IL-10-1082G/A genotype combinations in all analyzed newborns of both case and control groups |
There were no correlation between PVL Grade32 and TNF-α-1031T/C (P=0.7815), IL-1β-511C/T (P=0.7278), and IL-10-1082G/A (P=0.5420) genotypes.
Discussion
The PVL is the major cause of long-term neurological impairments and is preferably associated with the later development of cerebral palsy in children.35
The pathogenesis of ischemic cerebral lesions is due to the participation of inflammatory cascade, resultant from influx of polymorphonuclear leukocytes, followed by monocytes, and by the activation of the microglia, for which the expression of specific adhesion molecules, chemotactic factors, and proinflammatory cytokines is necessary. Cytokines are proteins with a low molecular weight, produced and secreted in the CNS by astrocytes and microglia and also by macrophages, monocytes, fibroblasts, and endothelial cells.3,18,21
Proinflammatory cytokines, particularly TNF-α and IL-1β, may cause encephalic lesion through several different mechanisms, because they have direct neurocytotoxicity, by inhibition of the differentiation of oligodendrocyte precursors and stimulation of oligodendrocyte apoptosis, with consequent vacuolar myelin degeneration. Cytokines may also have effective vasomotor and vaso-occlusive results, and their toxicity may be mediated through alterations in glutamate transport, increasing glutamate-mediated oxidative stress. Conversely, the anti-inflammatory cytokine IL-10 inhibits these effects through an indirect restriction in proinflammatory cytokine synthesis.3,11,22
Functional cytokine gene variants that result in altered production of inflammatory cytokines (TNF-α and IL-1β) or anti-inflammatory (IL-10) may modify disease processes, including PVL. Since the expression of TNF-α and IL-1β is critically linked to the pathogenesis of encephalic injury, genetic variants of these and other cytokines are candidates to modify risk of encephalic lesion in both preterm and term infants.3,20
Therefore, the PVL neuropathological hallmark is a great deal of activated microglia and macrophages that produce proinflammatory cytokines, such as TNF-α and interleukin-1 beta (IL-1β), which directly cause oligodendroglial cell death or worsens white matter lesions.3,20
Based on the known role of both TNF-α and IL-1β cytokines in inflammation, the goal of this study was to determine if the two proinflammatory polymorphisms (TNF-α-1031T/C and IL-1β-511C/T) and the anti-inflammatory polymorphism (IL-10-1082G/A) are associates with PVL risk in Brazilian newborns with and without this injury.
To assess the results of genetic association studies, the case and control groups should have the same ethnic and geographical origins because of the genetic basis of diseases, such as polymorphic configurations, which can differ between regions and populations. The influence of ethnic and geographic diversity can be extensive and should be considered in genetic studies.36,37 In this study, despite the significant miscegenation in Brazil, all newborns consecutively included were from the same geographic area and with white skin color.
The PVL prevalence among the 85 newborns hospitalized during the study period was 59% (1:1.7), a result consistent with the literature (1:1.1 to 1:2.1).1,3,38,39
Since the 1980s, the PVL has also been identified in term newborns (term-born PVL) in studies showing that more than half of all children with cerebral palsy are born at term and that a significant proportion (12%–53%) of infants with neuroimaging data exhibited a pattern of periventricular white matter injury, which is more typically seen in preterm infants1,5–12 Among the 50 cases this study, four male term newborns, because they also had both hypoxic-ischemic encephalopathy and PVL diagnosis, were included (8%). This prevalence was lower than reported at the literature,1,5–12 probably due to the small number of cases.
After molecular analysis, the allelic and genotypic frequencies of polymorphisms between case and control groups newborns were compared. As described, TNF-α cytokine is a central mediator of encephalic injury, and genetic variants of the TNF-α gene may modify risk of similar injuries in the perinatal period, by increased expression of cytokine in the ischemic period. Therefore, TNF-α cytokine has been clearly involved in the pathogenesis of these injuries, including PVL.3,20,21,23,25,40–42
The TNF-α-1031T/C polymorphism analysis in 60 prematurely born children (28 to 32 gestational age weeks) with cerebral palsy, 35 of them with PVL, and 84 controls of the same gestational age, found a significant association between TC and CC genotypes, as well as the polymorphic allele C, and the cerebral palsy risk, including the PVL children.21 In the present study, with a highest PVL case number and with gestational age that ranged from 25 to 39 weeks, similar results were found in which both the TC genotype (intermediate productor inflammatory cytokine) and the same genotype when combined with the CC genotype (high productor inflammatory cytokine) were associated with a significant risk of PVL.
Experimental and clinical evidences implicating IL-1β as another key mediator in hypoxic-ischemic encephalic injury show that IL-1β expression has a genetically determined component. The IL-1β-511T allele has been associated with increased risk for or disease progression in several inflammatory conditions.3,20,43,44
The genotypes CT/TT of polymorphism IL-1β-511C/T carried a significative 4.1-fold PVL risk in studies in PVL preterm newborns.21,41 However, in the present study, these same genotypes carried a higher 23-fold PVL risk.
IL-10 has been considered a neuroprotective anti-inflammatory cytokine, and the -1082G/A polymorphism is associated (A allele) with low IL-10 production.45 Three studies were conducted in preterm newborns with and without PVL, whose gestational age ranged from 26 to 32 weeks, and the sample ranged 18 to 36 newborns, with only one of these studies with term newborns as control group, such as this present study.25,46,47 Results of these two studies showed that the prevalence of PVL was not affected by IL-10-1082G/A genotypes.25,46 However, the third study showed that the GG genotype (anti-inflammatory cytokine high productor) was significantly associated with PVL reduced risk,47 whose data are in agreement with the present study. Interestingly, both studies showed the same number of cases with GG genotype, and PVL in these newborns did not persist for more than 7 days.
The three analyzed polymorphisms in this study were combined, and a statistically significant association was found between TC/CT/GA genotypes and PVL risk, and the TT/CC/GA genotypes combination was significantly associated with PVL reduced risk.
There was no correlation between PVL Grade32 and TNF-α-1031T/C (P=0.7815), IL-1β-511C/T (P=0.7278), and IL-10-1082 G/A (P=0.5420) genotypes. These results are in agreement with studies that performed such a comparison.46,47
Additional genetic studies should be performed to determine the complexities of PVL and develop new approaches for treating this cerebral injury, which can result from the interaction between polymorphic genes associated with inflammatory response and genetic–environmental factors for predicting neonatal encephalic damage and PVL. Such knowledge may enhance the understanding of the pathophysiology of this disorder and facilitate the discovery of the molecular causes of PVL, providing new perspectives on its early diagnosis and stimulating new research. The genetic association with PVL strengthens the proinflammatory polymorphisms TNF-α-1031T/C and IL-1β-511C/T and the anti-inflammatory polymorphism IL-10-1082 G/A hypothesis with regard to PVL pathogenesis.
The potential limitations of these data merit consideration; despite our positive findings, they should be interpreted with caution and must be corroborated by independent and multicenter studies to determine the actual prevalence of the polymorphisms TNF-α-1031T/C, IL-1β-511C/T, and IL-10-1082 G/A and their association with PVL in the Brazilian population. Thus, it is necessary to validate our data in prospective studies with larger PVL cohorts. Further, because our study was conducted in a small sample of Brazilian patients, the extrapolation of the data to other populations should be done with caution.
Conclusion
In conclusion, the present findings indicate that there is evidence of an association between the polymorphisms TNF-α-1031T/C, IL-1β-511C/T, and IL-10-1082G/A and PVL risk in this studied Brazilian newborn population.
Acknowledgments
The authors would like to thank the newborns’ parents whose consent and cooperation made this work possible. This contribution is extremely important in order to allow the following of the scientific research to bring a better future to Brazilian children. The authors would also like to thank the FAPESP FOUNDATION (#2014/16305-8) and FAMERP (BAP 2013/2014; 2014/2015) for their financial support.
Disclosure
The authors report no conflicts of interest in this work.
References
Miller SP, Shevell MI, Patenaude Y, O’Gorman AM. Neuromotor spectrum of periventricular leukomalacia in children born at term. Pediatr Neurol. 2000;23:155–159. | |
Rezaie P, Dean A. Periventricular leucomalacia, inflammation and white matter lesions within the developing nervous system. Neuropathology. 2002;22:106–132. | |
Distefano G, Praticò AD. Actualities on molecular pathogenesis and repairing processes of cerebral damage in perinatal hypoxic-ischemic encephalopathy. Ital J Pediatr. 2010;16;36:63. doi:10.1186/1824-7288-36-63. | |
Ellenberg JH, Nelson KB. The association of cerebral palsy with birth asphyxia: a definitional quagmire. Dev Med Child Neurol. 2013;55:210–216. | |
Lipp-Zwahlen AE, Deonna T, Chrzanowski R, Micheli JL, Calame A. Temporal evolution of hypoxic-ischaemic brain lesions in asphyxiated full-term newborns as assessed by computerized tomography. Neuroradiology. 1985;27(2):138–144. | |
Krägeloh-Mann I, Hagberg B, Petersen D, Riethmüller J, Gut E, Michaelis R. Bilateral spastic cerebral palsy-Pathogenetic aspects from MRI. Neuropediatrics. 1992;23:46–48. | |
Truwit CL, Barkovich AJ, Koch TK, Ferriero DM. Cerebral palsy: MR findings in 40 patients. AJNR Am J Neuroradiol. 1992;13(1):67–78. | |
Krägeloh-Mann I, Petersen D, Hagberg G, Vollmer B, Hagberg B, Michaelis R. Bilateral spastic cerebral palsy-MRI pathology and origin: Analysis from a representative series of 56 cases. Dev Med Child Neurol. 1995;37:379–397. | |
Okumura A, Hayakawa F, Kato T, Kuno K, Watanabe K. MRI findings in patients with spastic cerebral palsy. I: Correlation with gestational age at birth. Dev Med Child Neurol. 1997;39(6):363–368. | |
Hagberg B, Hagberg G, Beckung E, Uvebrant P. Changing panorama of cerebral palsy in Sweden. VIII. Prevalence and origin in the birth year period 1991–1994. Acta Paediatr. 2001;90:271–277. | |
du Plessis AJ, Volpe JJ. Perinatal brain injury in the preterm and term newborn. Curr Opin Neurol. 2002;15:151–157. | |
Wu YW, Croen LA, Shah SJ, Newman TB, Najjar DV. Cerebral palsy in a term population: risk factors and neuroimaging findings. Pediatrics. 2006;118(2):690–697. | |
Pang Y, Campbell L, Zheng B, Fan L, Cai Z, Rhodes P. Lipopolysaccharide- activated microglia induce death of oligodendrocyte progenitor cells and impede their development. Neuroscience. 2010;166:464–475. | |
Barger SW, Basile AS. Activation of microglia by secreted amyloid precursor protein evokes release of glutamate by cystine exchange and attenuates synaptic function. J Neurochem. 2001;76:846–854. | |
Huleihel M, Golan H, Hallak M. Intrauterine infection/inflammation during pregnancy and offspring brain damages: possible mechanisms involved. Reprod Biol Endocrinol. 2004;2:17. | |
Davalos D, Grutzendler J, Yang G, et al. ATP mediates rapid microglial response to local brain injury in vivo. Nat Neurosci. 2005;8:752–758. | |
Minghetti L. Role of inflammation in neurodegenerative diseases. Curr Opin Neurol. 2005;8:315–321. | |
Smith SMC, Mitchell GS, Friedle SA, Sibigtroth CM, Vinit S, Watters JJ. Hypoxia attenuates purinergic P2X receptor-induced inflammatory gene expression in brainstem microglia. Hypoxia (Auckl). 2013;1:1–11. | |
Kadhim H, Tabarki B, Verellen G, De Prez C, Rona AM, Sebire G. Inflammatory cytokines in the pathogenesis of periventricular leukomalacia. Neurology. 2001;56:1278–1284. | |
Lin CY, Chang YC, Wang ST, Lee TY, Lin CF, Huang CC. Altered inflammatory responses in preterm children with cerebral palsy. Ann Neurol. 2010;68(2):204–212. | |
Vidak HK, Ivkovic TC, Jokic M, Spaventi R, Kapitanovic S. The association between proinflammatory cytokine polymorphisms and cerebral palsy in very preterm infants. Cytokine. 2012;58:57–64. | |
Baier RJ. Genetics of perinatal brain injury in the preterm infants. Front Biosci. 2006;11:1371–1387. | |
Gibson CS, MacLennan AH, Goldwater PN, Haan EA, Priest K, Dekker GA; South Australian Cerebral Palsy Research Group. The association between inherited cytokine polymorphisms and cerebral palsy. Am J Obstet Gynecol. 2006;194:674.e1–11. | |
Al-Okaily F, Arfin M, Al-Rashidi S, Al-Balawi M, Al-Asmari A. Inflammation-related cytokine gene polymorphisms in Behçet’s disease. J Inflamm Res. 2015;8:173–180. | |
Nuk M, Orendi K, Rosenberger S, et al. Genetic variations in fetal and maternal tumor necrosis factor-α and interleukin 10: is there an association with preterm birth or periventricular leucomalacia? J Perinatol. 2012;32(1):27–32. | |
Haukim N, Bidwell JL, Smith AJ, et al. Cytokine gene polymorphism in human disease: on-line databases, supplement 2. Genes Immun. 2002;3:313–330. | |
Chen H, Wilkins LM, Aziz N, et al. Single nucleotide polymorphisms in the human interleukin-1B gene affects transcription according to haplotype context. Hum Mol Genet. 2006;15:519–529. | |
Kremlev SG, Palmer C. Interleukin-10 inhibits endotoxin-induced pro-inflammatory cytokines in microglial cell cultures. J Neuroimmunol. 2005;162:71–80. | |
Raedler D, Illi S, Pinto LA, et al. IL10 polymorphisms influence neonatal immune responses, atopic dermatitis, and wheeze at age 3 years. J Allergy Clin Immunol. 2013;131:789–796. | |
An Y, Chen Q, Quan N. Interleukin-1 exerts distinct actions on different cell types of the brain in vitro. J Inflamm Res. 2011;2011(4):11–20. | |
Sarnat HB, Sarnat MS. Neonatal encephalopathy following fetal distress. A clinical and electroencephalographic study. Arch Neurol. 1976;33(10):696–705. | |
de Vries LS, Eken P, Dubowitz LM. The spectrum of leukomalacia using cranial ultrasound. Behav Brain Res. 1992;49:1–6. | |
Turner DM, Williams DM, Sankaran D, Lazarus M, Sinnott PJ, Hutchinson IV. An investigation of polymorphism in the interleukin-10 gene promoter. Eur J Immunogenet. 1997;24:1–8. | |
Rees LE, Wood NA, Gillespie KM, Lai KN, Gaston K, Mathieson PW. The interleukin-10-1082 G/A polymorphism: allele frequency in different populations and functional significance. Cell Mol Life Sci. 2002;59:560–569. | |
Oskoui M, Coutinho F, Dykeman J, Jette N, Pringsheim T. An update on the prevalence of cerebral palsy: a systematic review and meta-analysis. Dev Med Child Neurol. 2013;55:509–519. | |
Ayub Q, Tyler-Smith C. Genetic variation in South Asia: assessing the influences of geography, language and ethnicity for understanding history and disease risk. Brief Funct Genomic Proteomic. 2009;8:395–404. | |
Haber M, Platt DE, Badro DA, et al. Influences of history, geography, and religion on genetic structure: the Maronites in Lebanon. Eur J Hum Genet. 2011;19(3):334–340. | |
Volpe JJ. Cerebral White matter injury of the preterm infant – more common than you think (commentaries). Pediatrics. 2003;112:176–180. | |
Silveira RC, Procianoy RS. Ischemic brain damage in very low birth weight preterm newborn infants. J Pediatr (Rio J). 2005; 81(1 Suppl):23–32. | |
Adcock K, Hedberg C, Loggins J, Kruger TE, Baier RJ. The TNF-alpha -308, MCP-1 -2518 and TGF-beta1 +915 polymorphisms are not associated with the development of chronic lung disease in very low birth weight infants. Genes Immun. 2003;4:420–426. | |
Baier RJ, Loggins J, Yanamandra K. Association of the interleukin-1beta -511 C/T polymorphism with intraventricular hemorrhage and periventricular leukomalacia in ventilated very low birth weight infants. Conference: Canadian Pediatric Society Annual Meeting, At Vancouver, British Columbia, Vol: Paediatr Child Health; 2005;10:25B; doi:10.13140/2.1.4537.9847. | |
Nelson KB, Dambrosia JM, Iovannisci DM, Cheng S, Grether JK, Lammer E. Genetic polymorphisms and cerebral palsy in very preterm infants. Pediatr Res. 2005;57(4):494–499. | |
Froen JF, Munkeby BH, Stray-Pedersen B, Saugstad OD. Interleukin-10 reverses acute detrimental effects of endotoxin-induced inflammation on perinatal cerebral hypoxia-ischemia. Brain Res. 2002;942:87–94. | |
Kimura R, Nishioka T, Soemantri A, Ishida T. Cis-acting effect of the IL1B C-31T polymorphism on IL-1beta mRNA expression. Genes Immun. 2004;5:572–575. | |
Mörmann M, Rieth H, Hua TD, et al. Mosaics of gene variations in the interleukin-10 gene promoter affect interleukin-10 production depending on the stimulation used. Genes Immun. 2004;5:246–255. | |
Yanamandra K, Boggs P, Loggins J, Baier RJ. Interleukin-10 -1082 G/A polymorphism and risk of death or bronchopulmonary dysplasia in ventilated very low birth weight infants. Pediatr Pulmonol. 2005; 39:426–432. | |
Dördelmann M, Kerk J, Dressler F, et al. Interleukin-10 high producer allele and ultrasound-defined periventricular white matter abnormalities in preterm infants: a preliminary study. Neuropediatrics. 2006;37(3):130–136. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.