Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 15
The Association Between Plasma Fatty Acid and Cognitive Function Mediated by Inflammation in Patients with Type 2 Diabetes Mellitus
Authors Shen J, Yu H, Li K, Ding B, Xiao R, Ma W
Received 10 December 2021
Accepted for publication 1 April 2022
Published 6 May 2022 Volume 2022:15 Pages 1423—1436
DOI https://doi.org/10.2147/DMSO.S353449
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Ming-Hui Zou
Jingyi Shen,1,* Huiyan Yu,1,* Kaifeng Li,2 Bingjie Ding,3 Rong Xiao,1 Weiwei Ma1
1School of Public Health, Beijing Key Laboratory of Environmental Toxicology, Capital Medical University, Beijing, 100069, People’s Republic of China; 2Nutrition and Metabolism Research Division, Innovation Center, Heilongjiang Feihe Dairy Co., Ltd., Beijing, 100015, People’s Republic of China; 3Department of Clinical Nutrition, Beijing Friendship Hospital, Capital Medical University, Beijing, 100050, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Weiwei Ma, School of Public Health, Beijing Key Laboratory of Environmental Toxicology, Capital Medical University, Beijing, 100069, People’s Republic of China, Tel/Fax +86-10-83911651, Email [email protected]
Purpose: To verify the mediating role of inflammatory factors in plasma fatty acid-induced changes in cognitive function in patients with type 2 diabetes mellitus (T2DM).
Patients and Methods: In this study, we evaluated the cognitive function of 372 Chinese patients (the average age was 58.00 (52.50, 63.00) years) with T2DM by using the Mini-Mental State Examination (MMSE) and the Montreal Cognitive Assessment (MoCA), with plasma fatty acids measured by gas chromatography analysis and inflammatory cytokines determined by immune turbidimetric analysis and enzyme-linked immunosorbent assay (ELISA) to investigate whether there was a correlation between the plasma fatty acids, inflammatory cytokine levels and cognitive test scores in Chinese patients with T2DM.
Results: We found that the increase of waist circumference and hip circumference might lead to cognitive impairment and induce the inflammatory response. Higher saturated fatty acids (SFAs) levels in plasma were linked to cognitive decline, while higher monounsaturated fatty acids (MUFAs) intake might be a protective factor for cognitive function. In addition, higher levels of plasma n-6 polyunsaturated fatty acids (n-6 PUFAs) stood out as having association with lower cognitive function scores, while higher level of plasma C22:6 n-3 could be a predictor of better cognitive function. In our study, higher SFAs led to higher proinflammatory factor levels. Apart from that, MUFAs and stearoyl-CoA desaturase-18 (SCD-18) were positively related to hypersensitive C-reactive protein (hs-CRP). Meanwhile, higher level of plasma C20:0 could lead to better MMSE delayed recall by reduce the expression of hs-CRP.
Conclusion: Levels of plasma SFAs, C18:3 n-6, and C20:3 n-6 could be a predictor for worse cognitive function, while MUFAs and C22:6 n-3 could be a predictor for better cognitive function. The level of hs-CRP could be a mediator of C20:0 induced the change of cognitive function.
Keywords: cognitive function, inflammatory cytokines, plasma fatty acids, T2DM, the mediation model
Introduction
Diabetes mellitus which exhibited an important influence on the central nervous system, is a group of metabolic diseases in which a person has a high blood glucose level over a prolonged period. According to the report of the International Diabetes Mellitus (IDF),1 there were 537 million living with diabetes in 2021, and the number was predicted to rise to 783 million in 2045, which confirmed that diabetes is one of the largest global health emergencies. The situation is more serious in China. From 2015 to 2017, a nationally representative cross-sectional survey in mainland China was conducted, which showed that the weighted prevalence of diabetes was 11.2% (using the World Health Organization (WHO) criteria).2
The discovery that the attention and memory of patients with diabetes were worse than normal individuals date back to 1922 reported by Miles.3 There were a growing number of findings indicated that diabetes was associated with accelerated cognitive decline, mild cognitive impairment (MCI), Alzheimer’s disease (AD), and dementia.4–7 Cognitive impairment in patients with diabetes was often presented as impaired learning and memory, orientation force, execution, and language skills.8,9 Apart from that, epidemiological researches indicated that the risk to develop dementia in diabetes patients was higher than in normal individuals.10,11 A cohort study showed that the presence of cognitive decline had a 2.5 to a 3.6- fold increase of type 2 diabetes mellitus (T2DM) patients compared with patients without diabetes.12
Several studies suggested that high-fat diet could induce increased obesity13,14 and diabetes,15 and dietary fatty acids could regulate inflammatory responses,16–18 alleviating or aggravating cognition.19 For instance, long-chain saturated fatty acids (LCSFAs), such as palmitic acid (16:0) and stearic acids (18:0), would have adverse effects on glucose metabolism, and increase the risk of T2DM.20 Saturated fatty acids (SFAs) could activate the toll-like receptors (TLRs), especially TLR2 and TLR4,21 then activate nuclear factor kappa B (NF-κB), resulting in inflammatory responses.22 By contrast, monounsaturated fatty acids (MUFAs) and polyunsaturated fatty acids (PUFAs) could reduce the concentration of interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α), and relived inflammatory reaction.18,23,24 However, the role of n-6 PUFAs played in was still controversies. Recent studies have clearly shown that inflammation was considered as a contributor to diabetic-induced neurodegeneration and cognitive impairment.25 Apart from that, dietary fatty acids have strong correlation of cognitive function, and previous studies pointed that higher SFAs could predict worse cognitive function, while MUFAs and n-3 PUFAs were positively related to better cognitive function.26,27 Moreover, the fatty acid profile in the plasma, which reflected short-term effects of diets could be considered as biomarkers for the incident T2DM.28 For instance, researchers found the higher percentages of linoleic acid (C18:2 n-6c) biomarkers of total fatty acids in plasma, the risk of T2DM lower in a study of patients.29 However, the role of inflammation in the progression of fatty acids induced diabetes-associated cognitive decline remains unknown in patients with T2DM.
Therefore, we investigated whether there was a correlation between the plasma fatty acids, inflammatory cytokine levels, and cognitive test scores in Chinese patients with T2DM.
Materials and Methods
Study Design and Study Population
Our cross-sectional study included 372 participants with T2DM, recruited among outpatients from the Department of nutrition Beijing Friendship Hospital, Capital Medical University (Beijing, China). Of all the participants, 214 (57.53%) were males and 158 (42.47%) were females: the average age was 58.00 (52.50, 63.00) years (26–75 years). The study protocol was approved by the ethics committee of Beijing Friendship Hospital, Capital Medical University (Beijing, China) (2015-P2-090-02) and was performed according to the ethical guidelines of the latest Declaration of Helsinki. All participants have signed informed consent and had the right to withdraw from the study at any time for any reason.
Individuals were excluded if they had: (1) severe heart, lung, and kidney dysfunction or malignancy; (2) history of encephalitis, head trauma, and other central nervous diseases; (3) any type of definite mental illness; (4) reading, hearing, or vision impairments; (5) losing self-caring ability.
TADM was diagnosed according to the World Health Organization (WHO) criteria: (1) fasting glucose≥7.0mmol/L; and/or (2) postprandial blood glucose≥11.1 mmol/L; and/or (3) the subjects have been diagnosed T2DM clinically before.2
Cognitive Function Assessments
All of the participants completed the Chinese versions of the Mini-Mental State Examination (MMSE) and the Montreal Cognitive Assessment (MoCA) to provide a general cognitive status.30,31 MMSE scale includes MMSE orientation force (maximum 10 points), MMSE computation (maximum 5 points), MMSE memory (maximum 3 points), MMSE delayed recall (maximum 3 points), MMSE language skills (maximum 9 points), and the total score of MMSE is 30. MOCA scale includes MoCA naming ability (maximum 3 points), MoCA orientation force (maximum 6 points), MoCA delayed recall (maximum 5 points), MoCA abstract thinking (maximum 2 points), MoCA language skills (maximum 3 points), MoCA visual-spatial ability (maximum 5 points) and MoCA attention (maximum 6 points), with a total score of 30 points.
Anthropometric and Laboratory Measurements
Height and weight were measured, and body mass index (BMI) was calculated as weight (kg) divided by height (m) squared. Also, we measured waist circumference and calculated waist-hip ratio. Following overnight fasting, we used a vacuum tube system (Becton-Dickinson) to take a fasting peripheral blood sample (4 mL) from the antecubital vein of each participant and centrifuged the sample at 3000g for 15 minutes to obtain plasma. Finally, plasma samples were stored in the dark at −80°C. Levels of hypersensitive C-reactive protein (hs-CRP) and insulin were assayed using commercial kits. And the 2-hour oral glucose tolerance test (OGTT) was performed.
Fatty Acids Analysis
The fast gas chromatography (GC) analysis was performed to determine the fatty acid compositions in plasma (ug/mL) according to the method of our previous study.19 The detailed steps were as follows. Firstly, after adding 100 ul internal standard in 100 ul plasma samples, KOH methanol solution (1 mL 0.5 mol/L) was then added and the mixture was shaken in a 60 °C water bath for 10 min. Second, 13% BF3-methanol reagent (3 mL) was added in the mixture. The hybrid liquid was kept at 60 °C for 40 min, then was cooled to room temperature. Then the mixture was added in hexane (1.5 mL) and shaken with vortex for 1min immediately. After that, the sodium chloride (2 mL) was added in t, and the mixture was centrifuged at 3000 rpm (15 min), then the upper layer was transferred to a vial for measurement. Finally, fatty acid methyl esters were analyzed using a GC-2010 gas chromatograph (Shimadzu, Japan), equipped with an SP-2560 gas chromatographic column (Supelco, USA).
Measurement of Plasma Inflammatory Cytokines
Each participant’ s inflammatory cytokines (endotoxin (ET), IL-1β, IL-10, NFκBP65, and TNF-α) were measured according to manufacturer instructions using specialized enzyme-linked immunosorbent assay (ELISA) kits (Mlbio, China).
Calculations
Homeostatic model assessment-IR (HOMA-IR) score was calculated using the formula (fasting insulin µIU/mL x fasting glucose (OGTT) mmol/L)/22.5.32 Quantitative sensitivity check index (QUICKI) score was calculated as follows: 1/ (log (fasting insulin µU/mL) + log (fasting glucose (OGTT) mg/dl).33 Desaturase activity including stearoyl-CoA desaturase (SCD), delta-5-desaturase (D5D) and delta-6-desaturase (D6D) were estimated using fatty acid product/precursor ratios as following: SCD-16 = C16:1 n-7/C16:0, SCD-18 = C18:1 n-9/C18:0, D6D = C18:3 n-6/C18:2 n-6 and D5D = C20:4 n-6/C20:3 n-6.34
Statistical Analysis
Data were presented as the mean and standard deviation. Multiple linear regression was used to obtain coefficients between plasma fatty acids, cognition scores, inflammation markers, and biochemistry biomarker. Potential confounders including age, gender, BMI, education, smoking and alcohol habits were also included in the model. Multiple linear regression was carried out using the lm function of the base R program (version 3.5). The simple mediation model was used to evaluate indirect, direct, and total effects between plasma fatty acids, inflammation markers, and cognition scores. A thousand bootstrap samples were drawn to estimate β coefficient indirect, direct, and total effects using Mplus version 7.4.35 Missing values were handled using the full information maximum likelihood implemented within Mplus software. P<0.05 was considered to indicate a statistically significant difference.
Results
The Relationship Between Adiposity Indicators, Biochemistry Biomarkers and Inflammation
In this study, the relationships between adiposity indicators, and biochemistry biomarkers and inflammation were measured after adjusting for confounders. As shown in Table 1, BMI and waist circumference, were negatively associated with QUICKI (P<0.05). At the same time, the results in the Tables 1 and 2 showed that BMI, hip circumference and waist circumference both positively correlated with insulin and hs-CRP (P<0.01).
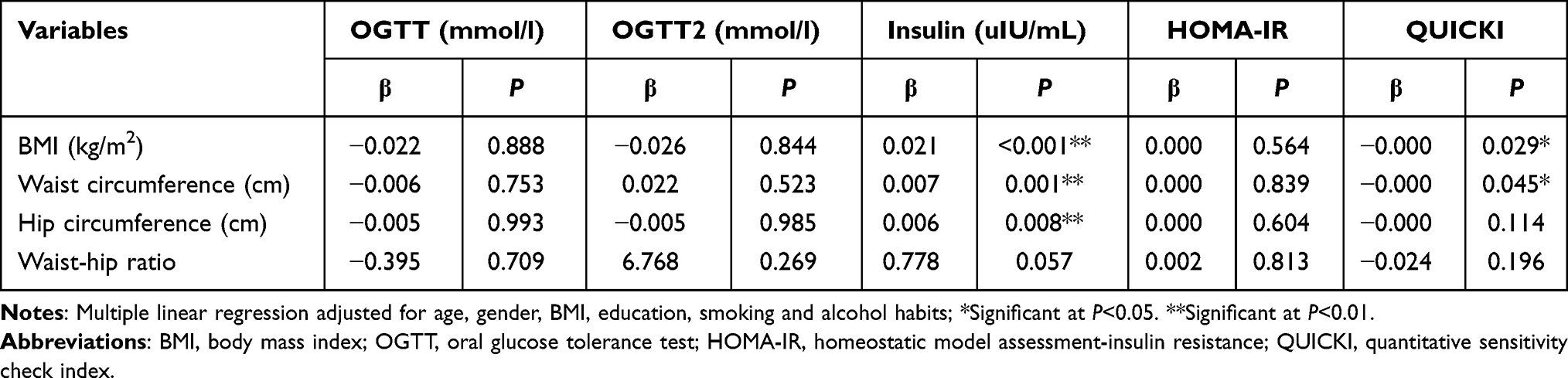 |
Table 1 Multiple Linear Regression Between Adiposity and Biochemistry Biomarkers (n=372) |
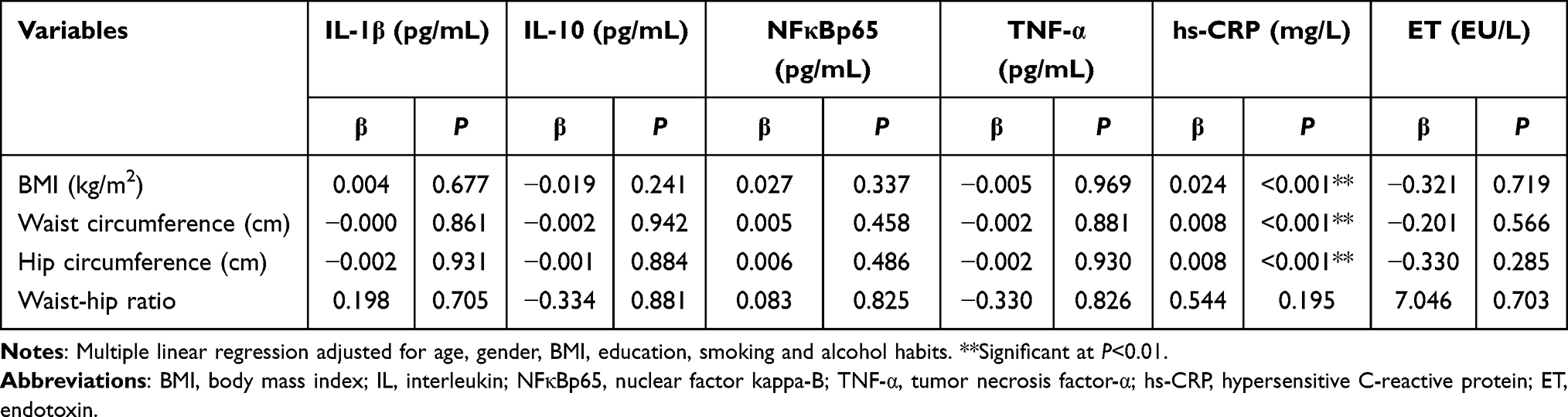 |
Table 2 Multiple Linear Regression Between Adiposity and Inflammation (n=372) |
The Relationship Between Adiposity Indicators and Cognitive Function Score
Then, the impact of adiposity indicators on cognitive function was analyzed. The interdependence was not found between adiposity indicators and scores assessed by MMSE (P>0.05) (Table 3). However, the increase of hip circumference and waist circumference might lead to a drop in the score of language skills in MoCA (P<0.05), and total MoCA score increased concomitantly as hip circumference decreased (Table 4).
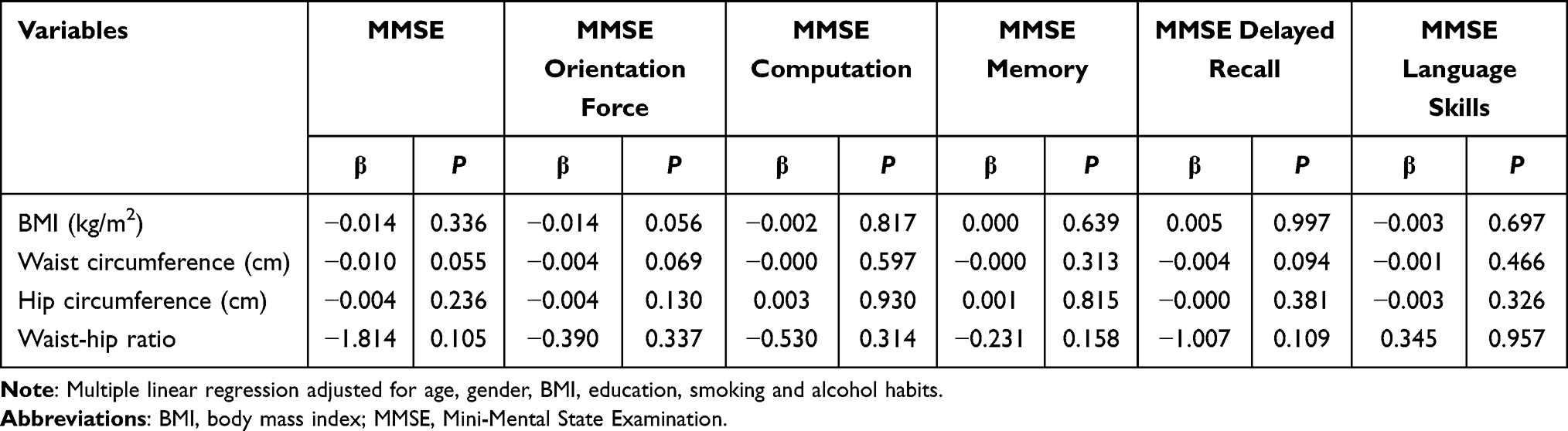 |
Table 3 Multiple Linear Regression Between Cognitive Function Score Measured by MMSE and Adiposity (n=372) |
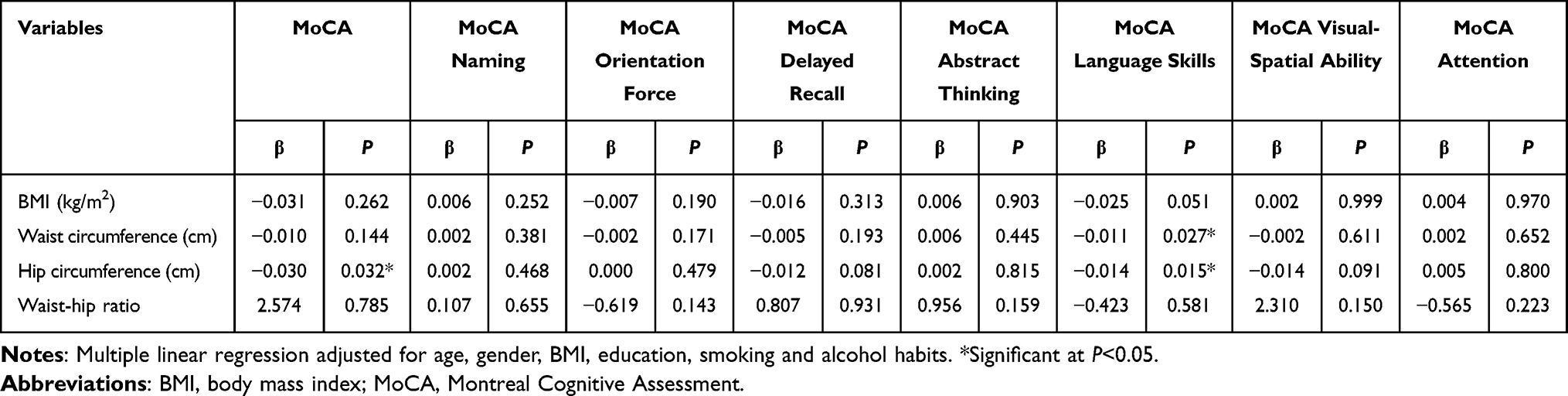 |
Table 4 Multiple Linear Regression Between Cognitive Function Score Measured by MoCA and Adiposity (n=372) |
The Relationship Between Plasma Fatty Acids, Biochemistry Biomarkers, and Inflammation
In our study, we found different plasma fatty acids had different effects on inflammatory responses and biochemistry biomarkers (Tables 5 and 6). Notably, with an increase in levels of plasma fatty acids, the levels of OGTT, OGTT2, insulin and HOMA-IR were also increased (P<0.05), while QUICKI was decreased (P<0.05). Such as, total SFAs, C14:0, C16:0 and C18:3 n-6. However, the level of n-6/n-3 PUFAs was negatively related to OGTT2 (P<0.05). The plasma level of C24:1 was positively related to IL-1β (P<0.05), the C18:0 plasma level was positively related to IL-10 (P<0.05), and plasma levels of total SFAs, C17:0, C18:0, C20:1 and C20:2 were positively related to NFκBp65 (P<0.05). Concomitant with increased plasma levels of C14:0, total MUFAs, C16:1, C18:1 n-9c, total n-6 PUFAs, C18:2 n-6c, C18:3 n-6, C20:3 n-6, C20:4 n-6, total PUFAs, C20:2, SCD-16, SCD-18 and D6D was the increased levels of hs-CRP (P<0.05). However, levels of C16:1, SCD-16 and SCD-18 were negatively related to the plasma level of ET (P<0.05), while the plasma level of C18:0 was positively related to the plasma level of ET (P<0.01).
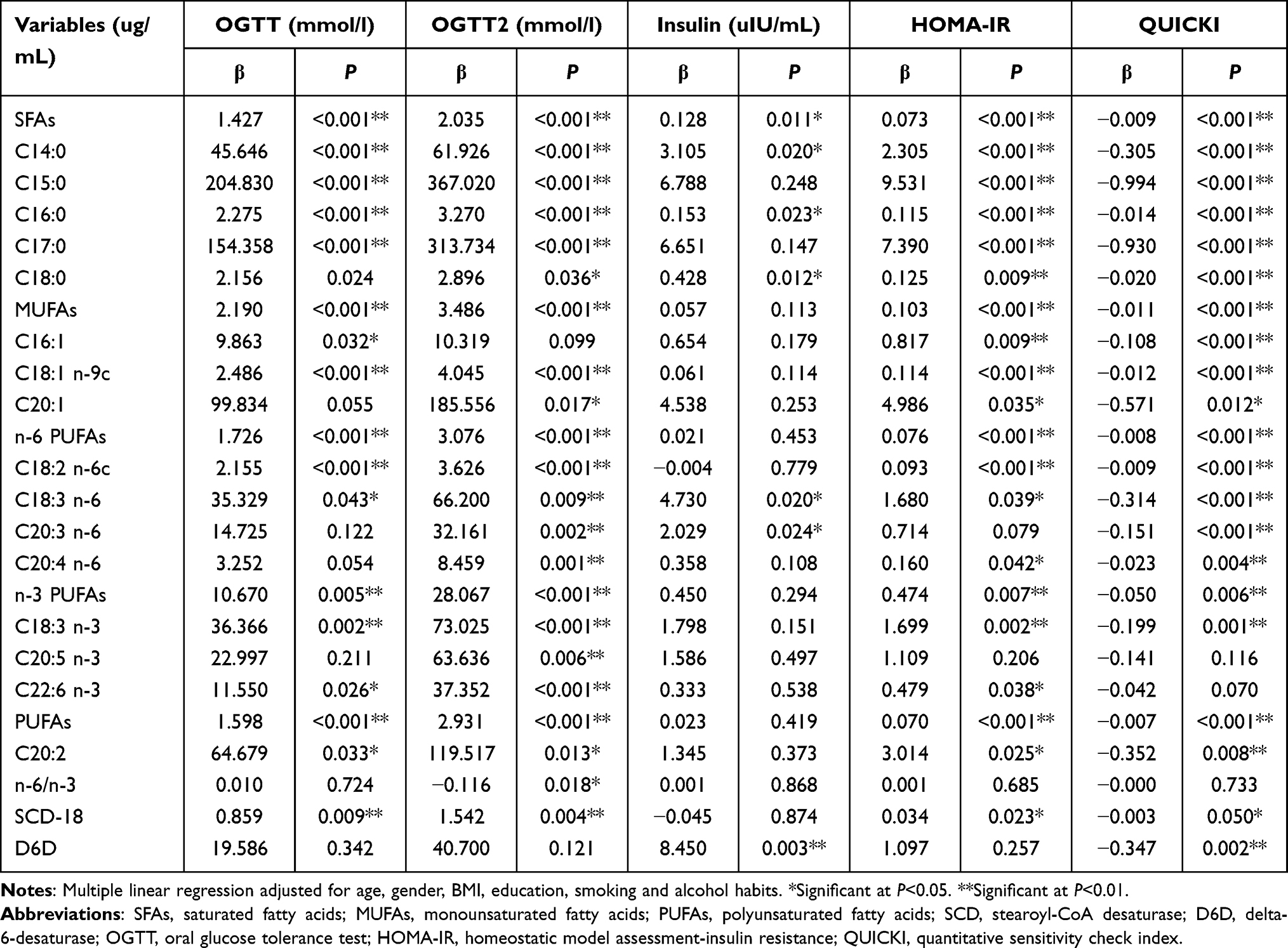 |
Table 5 Multiple Linear Regression Between Plasma Fatty Acids and Biochemistry Biomarkers (n=372) |
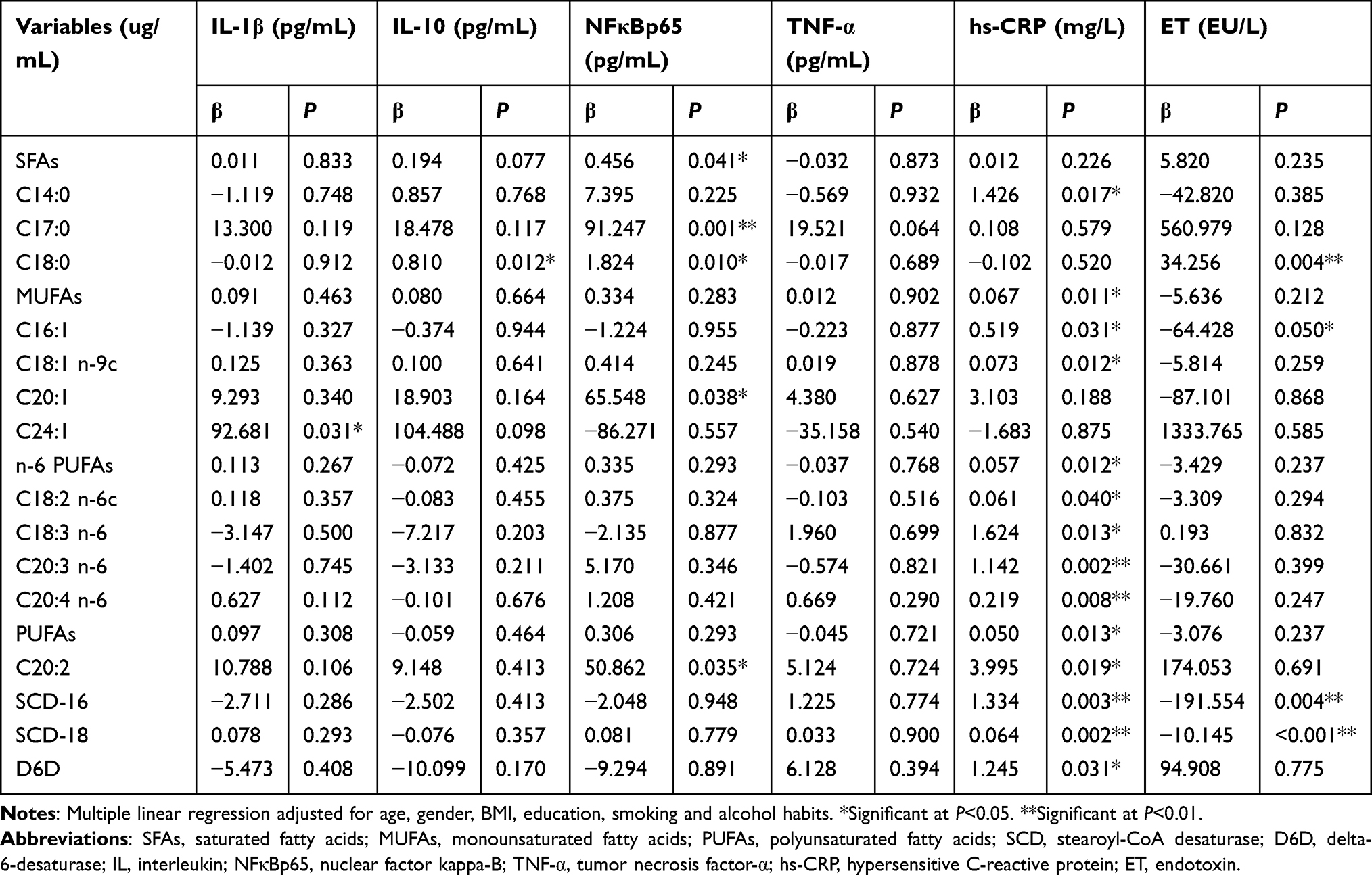 |
Table 6 Multiple Linear Regression Between Plasma Fatty Acids and Inflammation (n=372) |
The Relationship Between Plasma Fatty Acids and Cognitive Function Score
From Table 7, we found higher plasma level of C15:0 was accompanied by higher total MMSE and MMSE delayed recall scores (P<0.05), higher plasma level of C18:0 was accompanied by lower total MMSE and MMSE orientation force scores (P<0.05), and higher plasma level of C22:0 was accompanied by lower MMSE language skills score (P<0.05). In addition, plasma levels of C14:1 and C24:1 were negatively related to MMSE delayed recall and MMSE language skills scores (P<0.05), and the plasma level of C24:1 was also negatively related to total MMSE score (P<0.01). The higher SCD-18 was associated with higher total MMSE and MMSE orientation force scores (P<0.05), higher D6D was associated with lower MMSE delayed recall score (P<0.05), and higher D5D was associated with higher MMSE computation score (P<0.05). The results of Table 8 showed that the plasma level of total SFAs was negatively related to MoCA orientation force (P<0.05). Higher plasma level of C15:0, C20:0, and C18:1 n-9c were accompanied by higher total MoCA and MoCA visual-spatial ability scores (P<0.05). Higher plasma level of C18:0 was accompanied by lower MoCA orientation force and MoCA language skills scores (P<0.01). Higher plasma level of total MUFAs was associated with higher MoCA visual-spatial ability score (P<0.05). Higher plasma level of C24:1 was associated with lower total MoCA, MoCA delayed recall and MoCA visual-spatial ability scores (P<0.05). Higher plasma levels of C18:3 n-6, C20:3 n-6, SCD-16, and D6D were associated with lower MoCA abstract thinking score (P<0.05), while D5D was positively related to MoCA abstract thinking and MoCA attention scores (P<0.05). C22:6 n-3 was positively related to MoCA delayed recall score (P<0.05), n-6/n-3 was positively related to MoCA orientation force score (P<0.05), higher SCD-18 were associated with higher total MoCA, MoCA delayed recall and MoCA language skills scores (P<0.05).
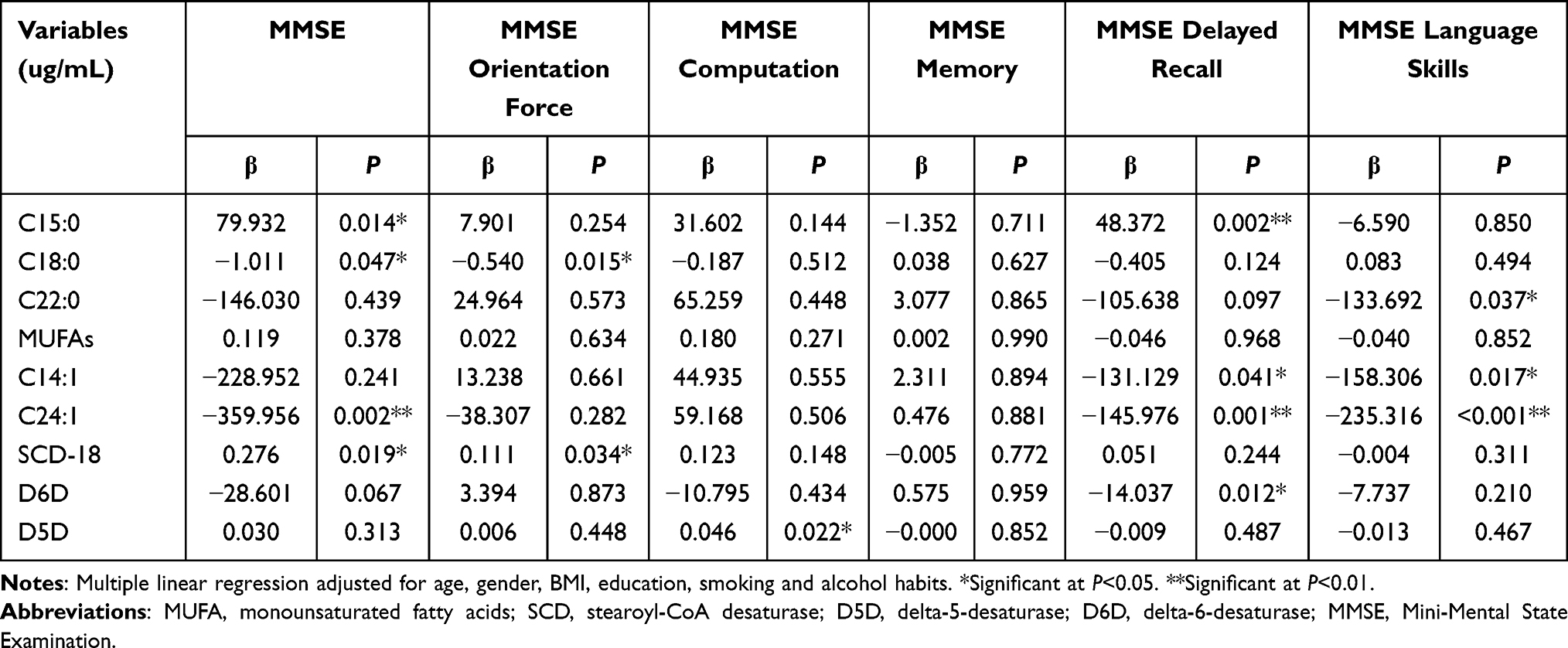 |
Table 7 Multiple Linear Regression Between Plasma Fatty Acids and Cognitive Function Score Measured by MMSE (n=372) |
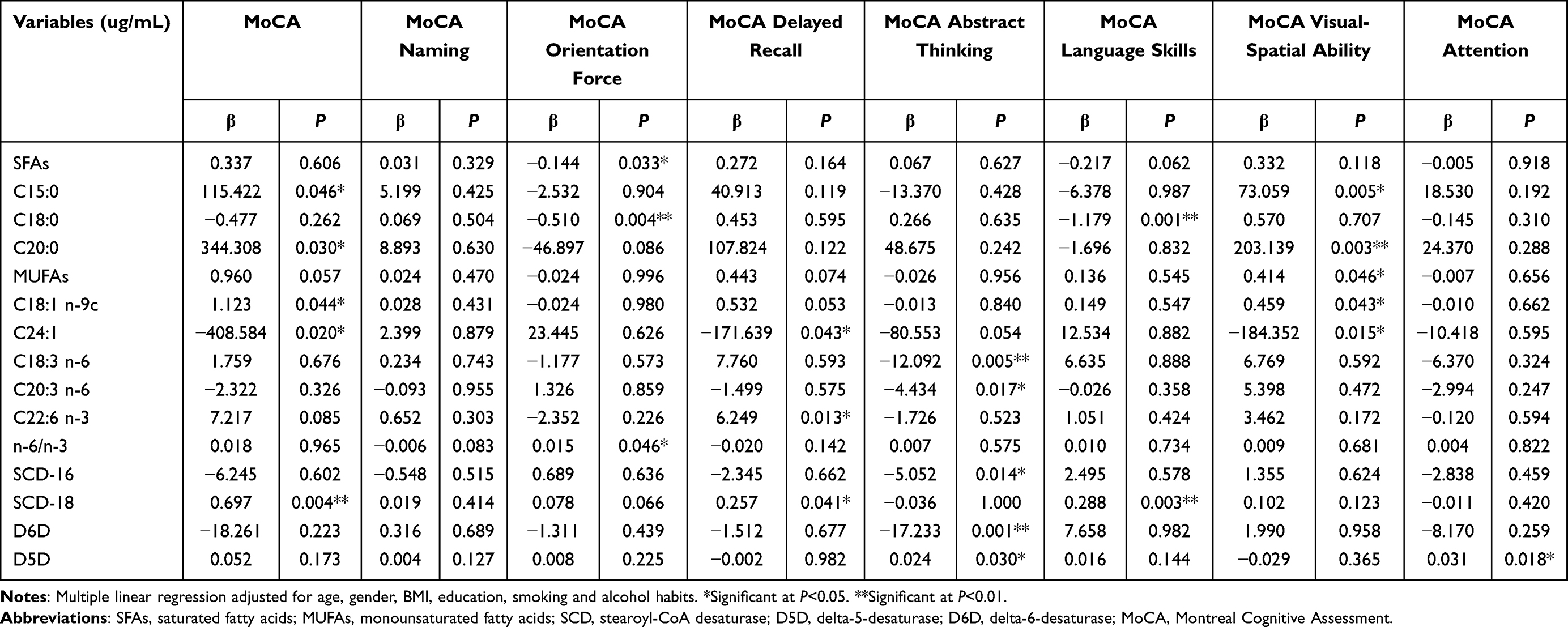 |
Table 8 Multiple Linear Regression Between Plasma Fatty Acids and Cognitive Function Score Measured by MoCA (n=372) |
The Relationship Between Cognitive Function Score, Biochemistry Biomarkers and Inflammation
To verify our hypothesis, we estimated the relationship between cognitive function score, biochemistry biomarkers, and inflammation (Tables 9 and 10). Higher plasma level of IL-1β might lead to lower total MMSE, MMSE orientation force and MMSE delayed recall scores (P<0.01). Accompanying the increase levels of plasma IL-10 was a decreased MoCA language skills score (P<0.05). Elevated level of plasma NFκBp65 was accompanied by lower total MMSE, MMSE computation, MMSE delayed recall, total MoCA and MoCA attention (P<0.05). Higher level of OGTT2 was associated with lower MoCA language skills score (P<0.05), higher level of insulin was associated with lower MMSE delayed recall score (P<0.05) and higher QUICKI was associated with higher MoCA language skills (P<0.05).
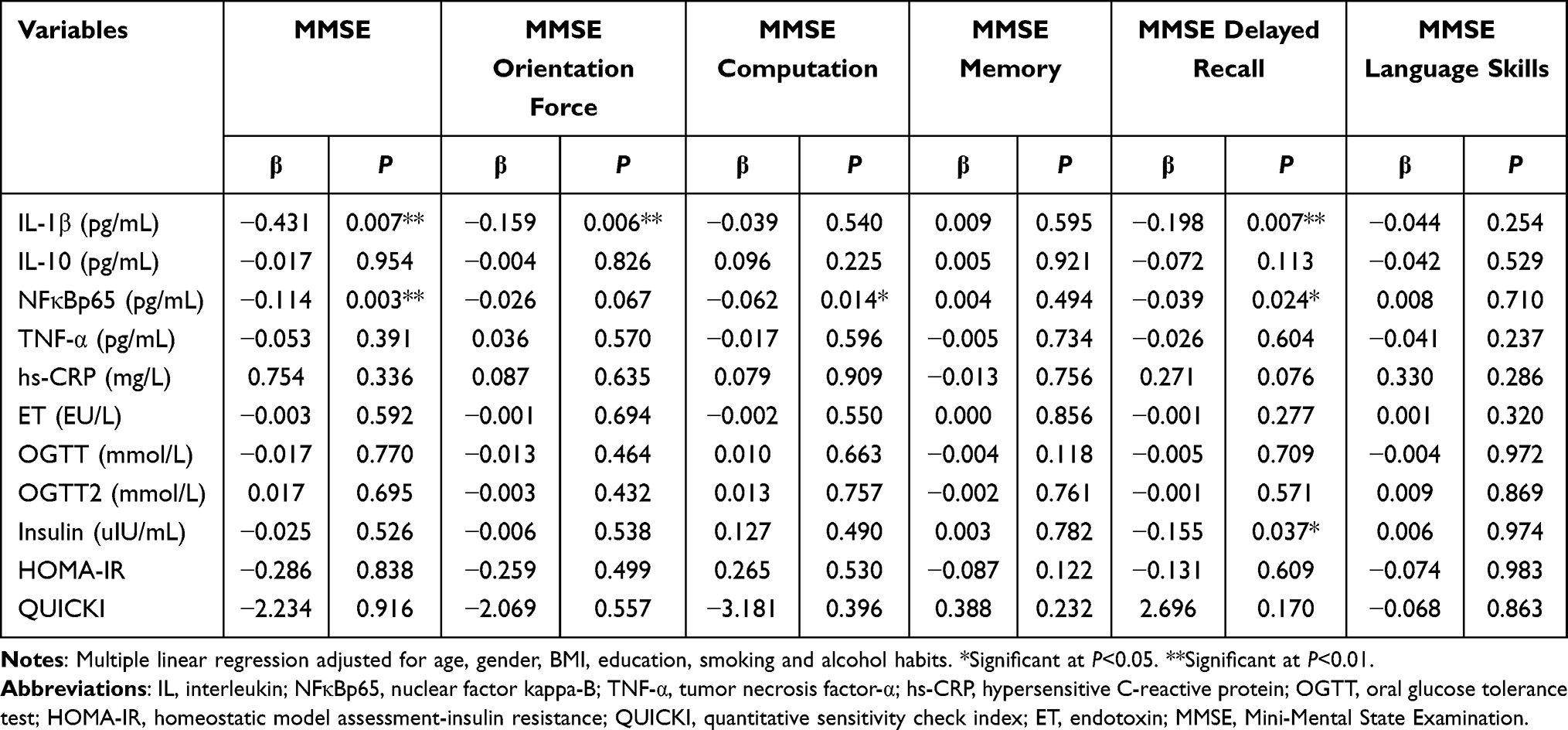 |
Table 9 Multiple Linear Regression Between Cognitive Function Score Measured by MMSE, Inflammation and Biochemistry Biomarkers (n=372) |
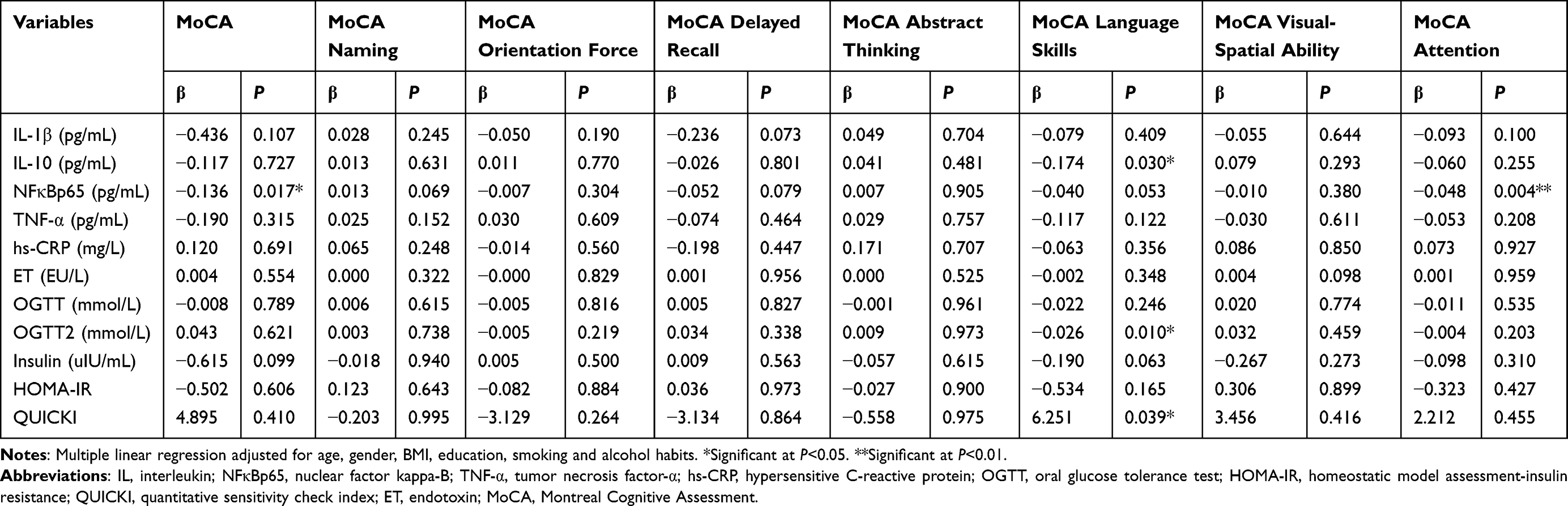 |
Table 10 Multiple Linear Regression Between Cognitive Function Score Measured by MoCA, Inflammation and Biochemistry Biomarkers (n=372) |
Summary of Mediation Models
All the results above showed that certain plasma fatty acids might be the protective or risk factors of the cognitive function in those living with T2DM in our study, which might be achieved by regulating the inflammatory response. After adjusting for age, gender, BMI, education, smoking and drinking habits, the Mplus version 7.4 was used to estimate the mediation effect. From the results shown in the Table 11, the mediation analysis revealed that the level of plasma C15:0 impacted MMSE delayed recall through QUICKI (P<0.05) and the level of plasma C20:0 impacted MMSE delayed recall through hs-CRP (P<0.05). Concretely, higher level of plasma C15:0 could lead to better MMSE delayed recall by reduce QUICKI, with the mediating effect of 11.25% accounting for the total effect. And higher level of plasma C20:0 could lead to better MMSE delayed recall by reduce the expression of hs-CRP, with the mediating effect of 3.06% accounting for the total effect. High level of plasma C18:0 might result in impaired MoCA language skill directly (P>0.05), and high level of plasma C24:1 could result in increased MMSE computation, and decreased MMSE delayed recall directly (P>0.05).
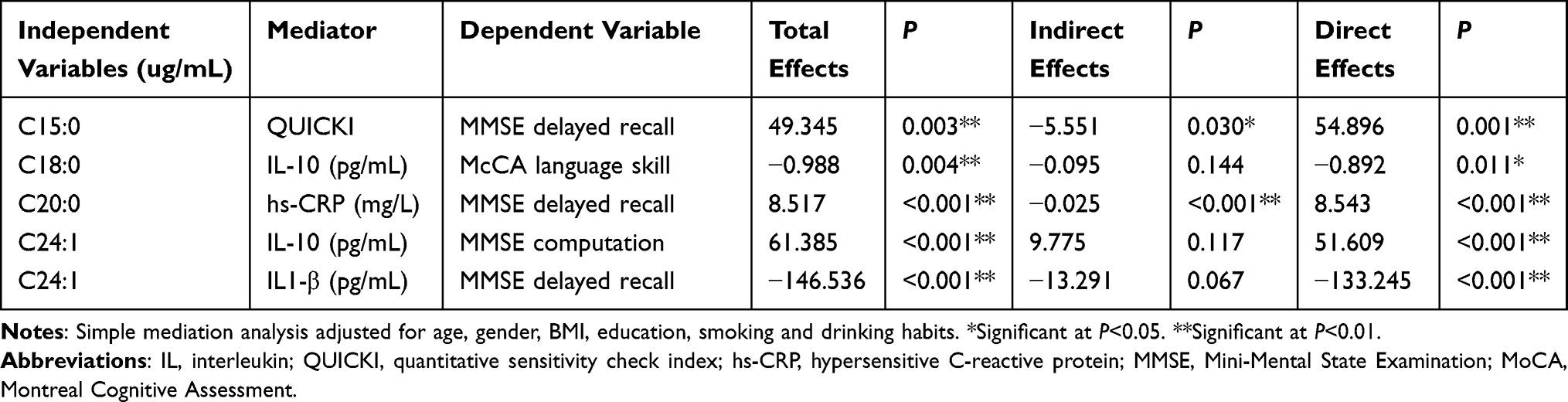 |
Table 11 Summary of Mediation Models |
Discussion
Although some previous studies suggested that diabetes-related inflammation might be one of the biological pathways linked to fatty acids and cognitive decline,36 definite evidence also needed to be verified. Thus, the mediation model was established to examine whether inflammation statistically mediated the relationship between plasma fatty acids and cognition function, which was evaluated by the MMSE and MoCA scales.
Previous studies pointed that overweight and obesity were linked to insulin resistance,37 which was frequently associated with inflammation.38 On the contrary, those who were obese might become insensitive to insulin.39 In the present study, we observed similar results. We found a higher and hs-CRP levels were related to higher BMI, hip circumference and waist circumference, while QUICKI was negatively related to the markers of obesity. Early studies also revealed waist circumference was significantly related to a high prevalence of cognitive dysfunction, which was independent of other related metabolic diseases and lifestyle risk factors, including smoking and drinking.40 Based on neuroimaging findings, those who were obese showed damaged brain structure (brain volume loss and brain atrophy of the grey matter),41,42 which indeed led to cognitive impairment.43 Similarly, in our study, hip circumference and waist circumference were negatively correlated with MoCA language skills scores and the increase in hip circumference might lead to a drop in the total MoCA score. These findings suggested that inflammation from obesity might cause cognitive decline in T2DM.
Most previous studies indicated high SFAs diet might be a risk factor of learning and memory ability, while MUFAs and PUFAs had protective effects of age-related cognitive decline.27,44 Yu45 found high SFAs diet resulted in obesity in mice and subsequently damaged the function of spatial learning and memory in mice. In a follow-up study, a conclusion was drawn by using structured interviews, which indicated high intake of dietary SFAs could aggravate clinical symptoms in AD patients and increase the risk of AD.46 In a prospective population-based study on elder participants with a typical Mediterranean diet, high PUFAs and MUFAs intakes seemed might be a protective measure against age-related cognitive decline.47 n-3 PUFAs could improve cognitive function,48 while the effect on cognitive function of n-6 PUFAs remained controversial.49,50 Some previous studies showed n-6 PUFAs enhanced the tissue inflammatory response,51 and increased the risk of cognitive impairment,52 whereas another study pointed that difference might result from the n-6/n-3 PUFAs ratio in diet.53,54 The similar conclusion was given in our study that the level of plasma SFAs (total SFAs, C18:0 and C22:0) might destroy the orientation force or language skills of patients with T2DM, while C15:1 and C20:0 showed opposite results. And higher MUFAs (total MUFAs and C18:1 n-9c) intake might be a protective factor for cognitive function, except C14:1 and C24:1. We speculated that the effect of plasma fatty acids on cognitive function was not only affected by their levels, but perhaps also related to their proportion of total fatty acids. As for the impact of PUFAs on cognitive functions, higher levels of plasma C18:3 n-6 and C20:3 n-6 were associated with lower MoCA abstract thinking score, while higher C22:6 n-3 and the ratio of n-3: n-6 fatty acids might predict better cognitive function. As reported, the activity of SCD, D5D, D6D was associated with metabolic-related diseases.55 In this study, similarly, SCD-18 and D5D had positive relationships with cognitive function scores, instead, SCD-16 and D6D showed opposite results. SCD converted a portion of 16:0 into palmitoleic (16:1) and 18:0 into oleic acid (18:1), which might contribute to the more abundant of MUFAs species,56 combined the result meant above, MUFAs, especially C18:1 had a stronger protective effect against the damaging effects of SFAs on cognitive function. Arachidonic acid (C20:4 n-6) and g-linoleate (C18:3 n-6) were derived from g-arachidonic acid (C20:3 n-6) and linoleate (C18:2 n-6) through desaturation (D6D and D5D) and elongation,56 which pointed that the activity of D5D and D6D could be changed by the proportion of n-6 PUFAs, and again verified the effects of levels of plasma C18:3 n-6 and C20:3 n-6 on impairing cognitive function. Moreover, previous studies indicated higher D5D was associated with better insulin sensitivity, while lower D6D activity led to a lower risk of insulin resistance,57 which might result in a different effect on cognitive function. And the genotype of related genes might affect the metabolic efficiency of PUFAs and desaturase activity, such as FADS1, which encoded D5D.58
Several types of researches existed that inflammation was involved in the development and/or progression of T2DM.59,60 And the inflammatory response could be impacted by diet fatty acids. High-fat diet might reduce synaptic plasticity and destroy insulin signaling/glucose homeostasis, which activated the innate immune system, including increased inflammatory cytokines, such as IL-6, IL-1β, TNF-α.61 In our study, we found higher levels of plasma SFAs led to higher IL-10, NFκBp65, hs-CRP and ET levels. Another study also showed that a high SFAs diet led to activated protein kinase, TLR4, and higher HOMA-IR values.62 SFAs were reported that contributed to inflammatory response and higher levels of plasma inflammatory factors (eg, IL-1β, IL-6).21,63 While treating the high-fat-diet-fed mice with palmitoleic acid or oleic acid daily by oral gavage, the expression of IL-1β and IL-12 decreased in PPAR-α-knockout mice.24 There was another study showed that n-3 PUFAs supplementation could reduce IL-6 and TNF-α production in T2DM.64 In this research, MUFAs, SCD-16, and SCD-18 were positively related to hs-CRP, while were negatively related ET. That might suggest that MUFAs could reduce ET-related inflammatory response. In addition, higher PUFAs led to higher inflammatory cytokines levels, such as hs-CRP, and NFκBp65. This finding provided evidence that the high-intake of PUFAs could also result in inflammatory responses, which indicated that fatty acid intake should be controlled. These increased inflammatory cytokines could induce activation microglia and result in neuroinflammation,65 accompanied by disrupt neurogenesis and brain structures.66 For example, in a case-control study, the serum levels of hs-CRP, IL-6, and TNF-α in T2DM subjects with MCI were significantly higher than in type 2 diabetic patients.67 Consistently, we showed that higher IL-1β, IL-10 and NFκBp65 levels were related to lower cognitive function scores.
To verify the effects of fatty acids on inflammation and cognition function, the mediating effect analysis was conducted. The mediation analysis revealed that higher levels of plasma C15:0 and C20:0 could lead to better MMSE delayed recall by reducing QUICKI and the expression of hs-CRP, respectively. As previous study reported, as a biomarker for inflammation, hs-CRP was generally associated with increased risk of poor cognitive outcomes.68,69 That was consistent with the findings of our present study.
There were still some limitations in our study. We needed to expand the sample size and choose different sources of participants instead of limited to hospital patients and health adults should be included in the study as a control. Apart from that, erythrocyte membrane fatty acid components of T2DM should be tested to evaluate the long-term intake of dietary fatty acid.
Conclusion
In conclusion, our study supported the hypothesis that plasma fatty acid could influence cognitive function by regulating inflammation, which suggested that plasma fatty acids could be evaluated as a potential indicator of cognitive function decline.
Abbreviations
AD, Alzheimer’s disease; BMI, body mass index; D5D, delta-5-desaturase; D6D, delta-6-desaturase; ELISA, enzyme-linked immunosorbent assay; LCSFAs, long chain saturated fatty acids; HOMA-IR, homeostatic model assessment-IR; hs-CRP, hypersensitive C-reactive protein; IDF, International Diabetes Mellitus; IL, interleukin; ET, endotoxin; MCI, mild cognitive impairment; MMSE, Mini-Mental State Examination; MoCA, Montreal Cognitive Assessment; MUFAs, monounsaturated fatty acids; NF-κB, nuclear factor kappa B; OGTT, oral glucose tolerance test; PUFAs, polyunsaturated fatty acids; QUICKI, quantitative sensitivity check index; SCD, stearoyl-CoA desaturase; SFAs, saturated fatty acids; T2DM, type 2 diabetes mellitus; TLRs, toll-like receptors; TNFα, tumor necrosis factor-α.
Data Sharing Statement
The data that support the findings of this study are available on request from the corresponding author. The manuscript has been posted to the preprint server Research Square, the link was as follows ” DOI:10.21203/rs.3.rs-577863/v3”.
Ethics Statement
We confirm that the data accessed in this study complies with relevant data protection and privacy legislation.
Ethics Approval and Informed Consent
This research was approved by the Ethics Committee of the Beijing Friendship Hospital, Capital Medical University (Beijing, China) (2015-P2-090-02).
Acknowledgments
Thanks for all the participants in this study.
Funding
This work was supported by grants from the Beijing Natural Science Foundation (NO. 7222242) and the National Natural Science Foundation of China (NO. 81472982).
Disclosure
The authors report no conflicts of interest in this work.
References
1. IDF Diabetes Atlas. International diabetes mellitus [homepage on the internet] 2021; 2021. Available from: https://diabetesatlas.org/.
2. Li Y, Teng D, Shi X, et al. Prevalence of diabetes recorded in mainland China using 2018 diagnostic criteria from the American Diabetes Association: national cross sectional study. BMJ. 2020;369:m997. doi:10.1136/bmj.m997
3. Miles WR, Root HF. Psychologic tests applied to diabetic patients. Arch Intern Med. 1922;30:767–777. doi:10.1001/archinte.1922.00110120086003
4. Nunley KA, Rosano C, Ryan CM, et al. Clinically relevant cognitive impairment in middle-aged adults with childhood-onset Type 1 diabetes. Diabetes Care. 2015;38(9):1768–1776. doi:10.2337/dc15-0041
5. Rawlings AM, Sharrett AR, Schneider AL, et al. Diabetes in midlife and cognitive change over 20 years: a cohort study. Ann Intern Med. 2014;161:785–793. doi:10.7326/m14-0737
6. Biessels GJ, Deary IJ, Ryan CM. Cognition and diabetes: a lifespan perspective. Lancet Neurol. 2008;7:184–190. doi:10.1016/s1474-4422(08)70021-8
7. Li JQ, Tan L, Wang HF, et al. Risk factors for predicting progression from mild cognitive impairment to Alzheimer’s disease: a systematic review and meta-analysis of cohort studies. J Neurol Neurosurg Psychiatry. 2016;87:476–484. doi:10.1136/jnnp-2014-310095
8. Zheng F, Yan L, Yang Z, et al. HbA (1c), diabetes and cognitive decline: the English longitudinal study of ageing. Diabetologia. 2018;61:839–848. doi:10.1007/s00125-017-4541-7
9. Sink KM, Divers J, Whitlow CT, et al. Cerebral structural changes in diabetic kidney disease: African American-Diabetes Heart Study MIND. Diabetes Care. 2015;38:206–212. doi:10.2337/dc14-1231
10. Morris JK, Vidoni ED, Honea RA, et al. Impaired glycemia increases disease progression in mild cognitive impairment. Neurobiol Aging. 2014;35:585–589. doi:10.1016/j.neurobiolaging.2013.09.033
11. Zhang DA, Lam V, Chu V, et al. Type 2 diabetes with comorbid depression in relation to cognitive impairment: an opportunity for prevention? Mol Neurobiol. 2018;55:85–89. doi:10.1007/s12035-017-0719-8
12. Nooyens AC, Baan CA, Spijkerman AM, et al. Type 2 diabetes and cognitive decline in middle-aged men and women: the Doetinchem Cohort Study. Diabetes Care. 2010;33:1964–1969. doi:10.2337/dc09-2038
13. Hammad SS, Jones PJ. Dietary fatty acid composition modulates obesity and interacts with obesity-related genes. Lipids. 2017;52:803–822. doi:10.1007/s11745-017-4291-9
14. Namekawa J, Takagi Y, Wakabayashi K, et al. Effects of high-fat diet and fructose-rich diet on obesity, dyslipidemia and hyperglycemia in the WBN/Kob-Lepr (fa) rat, a new model of type 2 diabetes mellitus. J Vet Med Sci. 2017;79:988–991.
15. Barrière DA, Noll C, Roussy G, et al. Combination of high-fat/high-fructose diet and low-dose streptozotocin to model long-term type-2 diabetes complications. Sci Rep. 2018;8(1):424. doi:10.1038/s41598-017-18896-5
16. Pimentel GD, Lira FS, Rosa JC, et al. Intake of trans fatty acids during gestation and lactation leads to hypothalamic inflammation via TLR4/NFκBp65 signaling in adult offspring. J Nutr Biochem. 2012;23(3):265–271. doi:10.1016/j.jnutbio.2010.12.003
17. Finucane OM, Lyons CL, Murphy AM, et al. Monounsaturated fatty acid-enriched high-fat diets impede adipose NLRP3 inflammasome-mediated IL-1β secretion and insulin resistance despite obesity. Diabetes. 2015;64:2116–2128. doi:10.2337/db14-1098.
18. Duca FA, Sakar Y, Covasa M. The modulatory role of high fat feeding on gastrointestinal signals in obesity. J Nutr Biochem. 2013;24:1663–1677. doi:10.1016/j.jnutbio.2013.05.005
19. Fan R, Zhao L, Ding BJ, et al. The association of blood non-esterified fatty acid, saturated fatty acids, and polyunsaturated fatty acids levels with mild cognitive impairment in Chinese population aged 35–64 years: a cross-sectional study. Nutr Neurosci. 2019:1–13. doi:10.1080/1028415x.2019.1610606
20. Mozaffarian D. Saturated fatty acids and type 2 diabetes: more evidence to re-invent dietary guidelines. Lancet Diabetes Endocrinology. 2014;2(10):770–772. doi:10.1016/s2213-8587(14)70166-4
21. Hwang DH, Kim JA, Lee JY. Mechanisms for the activation of Toll-like receptor 2/4 by saturated fatty acids and inhibition by docosahexaenoic acid. Eur J Pharmacol. 2016;785:24–35. doi:10.1016/j.ejphar.2016.04.024
22. Gérard P. Gut microbiota and obesity. Cell Mol Life Sci. 2016;73(1):147–162. doi:10.1007/s00018-015-2061-5
23. Cao H, Gerhold K, Mayers JR, et al. Identification of a lipokine, a lipid hormone linking adipose tissue to systemic metabolism. Cell. 2008;134:933–944. doi:10.1016/j.cell.2008.07.048
24. Souza CO, Teixeira AA, Lima EA, et al. Palmitoleic acid (n-7) attenuates the immunometabolic disturbances caused by a high-fat diet independently of PPARα. Mediators Inflamm. 2014;2014:582197. doi:10.1155/2014/582197
25. Duarte AI, Candeias E, Alves IN, et al. Liraglutide protects against brain Amyloid-β (1–42) accumulation in female mice with early Alzheimer’s disease-like pathology by partially rescuing oxidative/nitrosative stress and inflammation. Int J Mol Sci. 2020;21(5):1746. doi:10.3390/ijms21051746
26. Janssen CI, Kiliaan AJ. Long-chain polyunsaturated fatty acids (LCPUFA) from genesis to senescence: the influence of LCPUFA on neural development, aging, and neurodegeneration. Prog Lipid Res. 2014;53:1–17. doi:10.1016/j.plipres.2013.10.002
27. Solfrizzi V, Custodero C, Lozupone M, et al. Relationships of dietary patterns, foods, and micro- and macronutrients with Alzheimer’s disease and late-life cognitive disorders: a systematic review. J Alzheimer’s Dis. 2017;59:815–849. doi:10.3233/jad-170248
28. Lankinen MA, Stančáková A, Uusitupa M, et al. Plasma fatty acids as predictors of glycaemia and type 2 diabetes. Diabetologia. 2015;58(11):2533–2544. doi:10.1007/s00125-015-3730-5
29. Wu JHY, Marklund M, Imamura F, et al. Omega-6 fatty acid biomarkers and incident type 2 diabetes: pooled analysis of individual-level data for 39 740 adults from 20 prospective cohort studies. Lancet Diabetes Endocrinology. 2017;5:965–974. doi:10.1016/s2213-8587(17)30307-8
30. Ding B, Xiao R, Ma W, et al. The association between macronutrient intake and cognition in individuals aged under 65 in China: a cross-sectional study. BMJ open. 2018;8:e018573. doi:10.1136/bmjopen-2017-018573
31. Dong Y, Yean Lee W, Hilal S, et al. Comparison of the Montreal cognitive assessment and the mini-mental state examination in detecting multi-domain mild cognitive impairment in a Chinese sub-sample drawn from a population-based study. Int Psychogeriatr. 2013;25(11):1831–1838. doi:10.1017/s1041610213001129
32. Matthews DR, Hosker JP, Rudenski AS, et al. Homeostasis model assessment: insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia. 1985;28:412–419. doi:10.1007/bf00280883
33. Katz A, Nambi SS, Mather K, et al. Quantitative insulin sensitivity check index: a simple, accurate method for assessing insulin sensitivity in humans. J Clin Endocrinol Metab. 2000;85(7):2402–2410. doi:10.1210/jcem.85.7.6661
34. Li K, Brennan L, McNulty BA, et al. Plasma fatty acid patterns reflect dietary habits and metabolic health: a cross-sectional study. Mol Nutr Food Res. 2016;60(9):2043–2052. doi:10.1002/mnfr.201500711
35. Muthén BO, Muthén LK, Asparouhov T. Regression and Mediation Analysis Using Mplus. Los Angeles, CA: Muthén & Muthén; 2016.
36. Sharma G, Parihar A, Talaiya T, et al. Cognitive impairments in type 2 diabetes, risk factors and preventive strategies. J Basic Clin Physiol Pharmacol. 2020;31. doi:10.1515/jbcpp-2019-0105
37. Kahn BB, Flier JS. Obesity and insulin resistance. J Clin Invest. 2000;106(4):473–481. doi:10.1172/jci10842
38. Parvaresh Rizi E, Baig S, Shabeer M, et al. Meal rich in carbohydrate, but not protein or fat, reveals adverse immunometabolic responses associated with obesity. Nutr J. 2016;15(1):100. doi:10.1186/s12937-016-0219-0
39. McLean FH, Campbell FM, Langston RF, et al. A high-fat diet induces rapid changes in the mouse hypothalamic proteome. Nutr Metab. 2019;16(1):26. doi:10.1186/s12986-019-0352-9
40. Hou Q, Guan Y, Yu W, et al. Associations between obesity and cognitive impairment in the Chinese elderly: an observational study. Clin Interv Aging. 2019;14:367–373. doi:10.2147/cia.S192050
41. Allen PJ, Batra P, Geiger BM, et al. Rationale and consequences of reclassifying obesity as an addictive disorder: neurobiology, food environment and social policy perspectives. Physiol Behav. 2012;107:126–137. doi:10.1016/j.physbeh.2012.05.005
42. Alosco ML, Gunstad J. The negative effects of obesity and poor glycemic control on cognitive function: a proposed model for possible mechanisms. Curr Diab Rep. 2014;14(6):495. doi:10.1007/s11892-014-0495-z
43. Gardarsdottir M, Sigurdsson S, Aspelund T, et al. Atrial fibrillation is associated with decreased total cerebral blood flow and brain perfusion. Ep Europace. 2018;20(8):1252–1258. doi:10.1093/europace/eux220
44. Dohrmann DD, Putnik P, Bursać Kovačević D, et al. Japanese, Mediterranean and Argentinean diets and their potential roles in neurodegenerative diseases. Food Res Int. 2019;120:464–477. doi:10.1016/j.foodres.2018.10.090
45. Yu H, Bi Y, Ma W, et al. Long-term effects of high lipid and high energy diet on serum lipid, brain fatty acid composition, and memory and learning ability in mice. Int J Dev Neurosci. 2010;28(3):271–276. doi:10.1016/j.ijdevneu.2009.12.001
46. Laitinen MH, Ngandu T, Rovio S, et al. Fat intake at midlife and risk of dementia and Alzheimer’s disease: a population-based study. Dement Geriatr Cogn Disord. 2006;22(1):99–107. doi:10.1159/000093478
47. Solfrizzi V, Colacicco AM, D’Introno A, et al. Dietary intake of unsaturated fatty acids and age-related cognitive decline: a 8.5-year follow-up of the Italian Longitudinal Study on Aging. Neurobiol Aging. 2006;27(11):1694–1704. doi:10.1016/j.neurobiolaging.2005.09.026
48. Solfrizzi V, Frisardi V, Seripa D, et al. Mediterranean diet in predementia and dementia syndromes. Curr Alzheimer Res. 2011;8:520–542. doi:10.2174/156720511796391809
49. MacDonald-Wicks L, McEvoy M, Magennis E, et al. Dietary long-chain fatty acids and cognitive performance in older Australian adults. Nutrients. 2019;11. doi:10.3390/nu11040711
50. Kalmijn S, Feskens EJ, Launer LJ, et al. Polyunsaturated fatty acids, antioxidants, and cognitive function in very old men. Am J Epidemiol. 1997;145:33–41. doi:10.1093/oxfordjournals.aje.a009029
51. Grimble RF, Tappia PS. Modulation of pro-inflammatory cytokine biology by unsaturated fatty acids. Z Ernahrungswiss. 1998;37(Suppl 1):57–65.
52. Bigornia SJ, Scott TM, Harris WS, et al. Prospective associations of erythrocyte composition and dietary intake of n-3 and n-6 PUFA with measures of cognitive Function. Nutrients. 2018;10(9):1253. doi:10.3390/nu10091253
53. Dong Y, Xu M, Kalueff AV, et al. Dietary eicosapentaenoic acid normalizes hippocampal omega-3 and 6 polyunsaturated fatty acid profile, attenuates glial activation and regulates BDNF function in a rodent model of neuroinflammation induced by central interleukin-1β administration. Eur J Nutr. 2018;57:1781–1791. doi:10.1007/s00394-017-1462-7
54. Stavrinou PS, Andreou E, Aphamis G, et al. The effects of a 6-month high dose omega-3 and omega-6 polyunsaturated fatty acids and antioxidant vitamins supplementation on cognitive function and functional capacity in older adults with mild cognitive impairment. Nutrients. 2020;12(2):325. doi:10.3390/nu12020325
55. Bonafini S, Giontella A, Tagetti A, et al. Fatty acid profile and desaturase activities in 7–10-year-old children attending primary school in Verona south district: association between palmitoleic acid, SCD-16, Indices of adiposity, and blood Pressure. Int J Mol Sci. 2020;21(11):3899. doi:10.3390/ijms21113899
56. Kopf T, Schaefer HL, Troetzmueller M, et al. Influence of fenofibrate treatment on triacylglycerides, diacylglycerides and fatty acids in fructose fed rats. PLoS One. 2014;9:e106849. doi:10.1371/journal.pone.0106849
57. Astarita G, Jung KM, Vasilevko V, et al. Elevated stearoyl-CoA desaturase in brains of patients with Alzheimer’s disease. PLoS One. 2011;6:e24777. doi:10.1371/journal.pone.0024777
58. Obukowicz M, Welsch D, Salsgiver W, et al. Novel, selective delta6 or delta5 fatty acid desaturase inhibitors as antiinflammatory agents in mice. Lipids. 1999;34(Suppl):S149. doi:10.1007/bf02562269
59. Chawla A, Nguyen KD, Goh YP. Macrophage-mediated inflammation in metabolic disease. Nat Rev Immunol. 2011;11:738–749. doi:10.1038/nri3071
60. Donath MY, Shoelson SE. Type 2 diabetes as an inflammatory disease. Nat Rev Immunol. 2011;11(2):98–107. doi:10.1038/nri2925
61. Dutheil S, Ota KT, Wohleb ES, et al. High-fat diet induced anxiety and anhedonia: impact on brain homeostasis and inflammation. Neuropsychopharmacology. 2016;41:1874–1887. doi:10.1038/npp.2015.357
62. Tamer F, Ulug E, Akyol A, et al. The potential efficacy of dietary fatty acids and fructose induced inflammation and oxidative stress on the insulin signaling and fat accumulation in mice. Food Chem Toxicol. 2020;135:110914. doi:10.1016/j.fct.2019.110914
63. Granholm AC, Bimonte-Nelson HA, Moore AB, et al. Effects of a saturated fat and high cholesterol diet on memory and hippocampal morphology in the middle-aged rat. J Alzheimers Dis. 2008;14:133–145. doi:10.3233/jad-2008-14202
64. O’Mahoney LL, Matu J, Price OJ, et al. Omega-3 polyunsaturated fatty acids favourably modulate cardiometabolic biomarkers in type 2 diabetes: a meta-analysis and meta-regression of randomized controlled trials. Cardiovasc Diabetol. 2018;17:98. doi:10.1186/s12933-018-0740-x
65. Guillemot-Legris O, Muccioli GG. Obesity-induced neuroinflammation: beyond the hypothalamus. Trends Neurosci. 2017;40:237–253. doi:10.1016/j.tins.2017.02.005
66. Hao S, Dey A, Yu X, et al. Dietary obesity reversibly induces synaptic stripping by microglia and impairs hippocampal plasticity. Brain Behav Immun. 2016;51:230–239. doi:10.1016/j.bbi.2015.08.023
67. Zheng M, Chang B, Tian L, et al. Relationship between inflammatory markers and mild cognitive impairment in Chinese patients with type 2 diabetes: a case-control study. BMC Endocr Disord. 2019;19(1):73. doi:10.7326/M14-0737
68. Lien CY, Lu CH, Chang CC, et al. Correlation between hypovitaminosis D and nutritional status with the severity of clinical symptoms and impaired cognitive function in patients with Parkinson’s disease. Acta neurologica Taiwanica. 2021;30(2):63–73.
69. John A, Rusted J, Richards M, et al. Accumulation of affective symptoms and midlife cognitive function: the role of inflammation. Brain Behav Immun. 2020;84:164–172. doi:10.1016/j.bbi.2019.11.021
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.