")
Back to Journals » International Journal of Nanomedicine » Volume 19
The Application of Nanoparticles Targeting Cancer-Associated Fibroblasts
Authors Huang Q, Ge Y, He Y, Wu J , Tong Y, Shang H, Liu X, Ba X, Xia D, Peng E, Chen Z , Tang K
Received 12 November 2023
Accepted for publication 23 March 2024
Published 8 April 2024 Volume 2024:19 Pages 3333—3365
DOI https://doi.org/10.2147/IJN.S447350
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor R.D.K. Misra
Qiu Huang,* Yue Ge,* Yu He, Jian Wu, Yonghua Tong, Haojie Shang, Xiao Liu, Xiaozhuo Ba, Ding Xia, Ejun Peng, Zhiqiang Chen, Kun Tang
Department of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, 430030, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhiqiang Chen; Kun Tang, Email [email protected]; [email protected]
Abstract: Cancer-associated fibroblasts (CAF) are the most abundant stromal cells in the tumor microenvironment (TME), especially in solid tumors. It has been confirmed that it can not only interact with tumor cells to promote cancer progression and metastasis, but also affect the infiltration and function of immune cells to induce chemotherapy and immunotherapy resistance. So, targeting CAF has been considered an important method in cancer treatment. The rapid development of nanotechnology provides a good perspective to improve the efficiency of targeting CAF. At present, more and more researches have focused on the application of nanoparticles (NPs) in targeting CAF. These studies explored the effects of different types of NPs on CAF and the multifunctional nanomedicines that can eliminate CAF are able to enhance the EPR effect which facilitate the anti-tumor effect of themselves. There also exist amounts of studies focusing on using NPs to inhibit the activation and function of CAF to improve the therapeutic efficacy. The application of NPs targeting CAF needs to be based on an understanding of CAF biology. Therefore, in this review, we first summarized the latest progress of CAF biology, then discussed the types of CAF-targeting NPs and the main strategies in the current. The aim is to elucidate the application of NPs in targeting CAF and provide new insights for engineering nanomedicine to enhance immune response in cancer treatment.
Keywords: nanoparticles, cancer therapy, cancer-associated fibroblasts, drug delivery, cancer, nanomedicine
Background
In recent years, several barriers to effective tumor eradication, including the formation of distant metastasis, angiogenesis, drug resistance, and immunosuppression, have been identified. Consequently, there has been an increasing interest in the classification of different cells within the tumor parenchyma; these endeavors have led to the emergence of the tumor microenvironment (TME) concept.1–6 Characterization of the TME has drawn attention to the deleterious role of cancer-associated fibroblasts (CAFs) in this context.7 Under normal conditions, fibroblasts maintain a quiescent state and mainly contribute to the secretion of collagen, which supports the formation and integrity of the extracellular matrix (ECM), the fundamental tissue scaffold.8,9 In the context of wound healing, fibroblasts become activated to facilitate tissue repair before undergoing apoptosis.10 Tumors are often likened to “cancerous wounds”11 that fail to heal and continuously stimulate fibroblasts through the secretion of cytokines such as TGF-β,12 preventing them from returning to their resting state and promoting tumor growth. Recent investigations, however, have uncovered the presence of cancer-restraining CAF (rCAF) subtypes, which have tumor suppressor properties.13 CAFs play significant roles in tumor proliferation, metastasis, chemotherapy resistance, cancer metabolism, immunotherapy resistance, and immune evasion,14 which will be comprehensively discussed in this review. Consequently, CAF-targeting therapies have emerged; however, despite promising preclinical findings, the desired levels of efficacy have yet to be achieved and the clinical implementation of these therapies remains limited.15
The past few decades have hailed rapid advancements in nanotechnology, which have facilitated the development and delivery of targeted drugs for cancer therapy.16,17 However, results from clinical trials of anti-CAF drugs, such as vismodegib, navitoclax, and sibrotuzumab, have highlighted their uncontrollable toxicity and unstable action.7,18 Drug encapsulation in nanoparticles (NPs) promises to overcome these challenges. NPs can optimize drug properties, reduce degradation rates in the body, improve solubility, enhance targeting capabilities, and improve overall drug efficacy.19 Various types of NPs are available, including those constructed from lipids, metals, polymers, biological macromolecules, and cell membranes; each type has distinct properties and characteristics, which can be matched to a specific application. Moreover, coupling CAF-specific peptides or antibodies to the surface of NPs enables efficient targeting and drug release within these cells.20,21
This review provides an overview of the characteristics of CAFs and their interactions with immune cells in the TIME. Furthermore, we present a comprehensive summary of nanomaterial selection and construction strategies, which are being used in the development of NPs for CAF-targeted immunotherapy.
TME and Cancer Therapy
Numerous studies have established the significance of the TME in tumor progression, which can be as important as the genetic and epigenetic alteration of tumor cells.22,23 The TME provides an external view of the changes occurring within tumor cells and evolves dynamically throughout tumor development. During the initial stages of cancer development, innate immune cells such as natural killer (NK) cells, macrophages, and dendritic cells (DCs) infiltrate the tumor and mount a rapid immune response.24,25 In addition, antigen-presenting cells, prime and stimulate lymphocytes, which are essential for tumor eradication. Tumor cells employ various mechanisms to evade or impair anti-tumor T cell activity; in addition, they recruit various pro-tumor immune cells, including myeloid-derived suppressor cells (MDSCs), regulatory T cells (Tregs), CAFs, macrophages, and neutrophils. The subsequent increase in the ratio of pro-tumor to anti-tumor immune cells contributes to tumor progression. Given the limitations of conventional tumor treatment approaches, attention has shifted toward immunotherapy as a potent means of anti-tumor eradication. Over the past few decades, diverse forms of cancer immunotherapy have been explored.26 Current immunotherapy strategies encompass: 1) oncolytic viral therapy, which uses transgenic viruses to infect tumors and establish a pro-inflammatory TME to enhance anti-tumor immune responses;27 2) tumor vaccines, which use tumor-associated antigens to prime tumor-specific T cells;28 3) cytokine therapy, which modulates cytokine patterns to activate the immune system;29 4) adoptive immune cell transfer, which involves the reinfusion of autologous, existing or genetically-engineered (eg, CAR-T cells) anti-tumor immune cells into the patient;30 and 5) immune checkpoint inhibitor (ICI) therapy.31 Immunotherapy has emerged as an effective and well-tolerated treatment approach in clinical practice. However, the development of immunotherapy resistance poses a significant challenge. To address this issue, attention have been refocused on specific immune and stromal cells within the TME. Consequently, targeted drugs have been developed to remodel the TME and mitigate immunotherapy resistance.26 Meanwhile, advancements in nanotechnology have optimized immunotherapy via the use of nanoscale drug carriers. This innovative approach enhances the efficacy of targeted drugs and provides a promising direction for overcoming immunotherapy resistance.32
CAFs Characteristics
CAFs Biology
Fibroblasts are the most abundant stromal cells in normal tissues; they play a crucial role in maintaining ECM stability and structural integrity.8 They typically originate from mesenchymal stem cells (MSCs) and remain in a quiescent state as individual cells primarily located near the basement membrane. Fibroblasts are activated during processes such as wound repair, tissue inflammation, and fibrosis, before subsequently undergoing apoptosis or returning to a resting state upon completion of the healing and remodeling process.33 During tumor development, a prolonged state of chronic inflammation, often referred to as the “tumor wound”, arises.11 This sustained state of injury and repair leads to the accumulation of numerous complex cytokines within the TME, triggering persistent fibroblast activation. These hyperactivated fibroblasts, otherwise known as CAFs, are characterized by heightened proliferation, increased migration, and resistance to apoptosis.34 In addition, CAFs produce substantial amounts of ECM components and proteases, including collagen, matrix metalloproteinases, and plasmin.35 This accumulation of ECM building blocks, in turn, increases tumor stiffness, which impedes immune cell infiltration, promotes immunosuppression, and facilitates tumor immune evasion.36 CAFs also secrete numerous pro-tumor factors such as vascular endothelial growth factor (VEGF), which stimulates angiogenesis and promotes tumor proliferation and metastasis.37 Moreover, CAFs significantly contribute to the remodeling of cancer metabolism,38 particularly lipid metabolism39 and mitochondrial oxidative stress.40 Additionally, CAFs can induce resistance to chemotherapy or immunotherapy via the release of autocrine exosomes and the creation of self-established physical barriers within the tumor.41–44 Understanding the functions of CAFs in tumor tissues is crucial for the effective design and implementation of NP-based, CAF-targeting therapeutic strategies.
CAFs Origin
There is mounting evidence indicating that CAFs do not arise from a single cell source but rather represent a complex and heterogeneous cell population.45–47 Moreover, CAFs differ between tumors arising in different tissues and displaying tissue-specific characteristics.7 The majority of CAFs originate from fibroblasts in normal tissues with a particularly high fibroblast content, such as the breast, lung, and colorectal tissues. Additionally, innate stellate cells of the pancreas48 and liver49 are common sources of CAFs; upon activation these cells differentiate into CAFs with a muscle-like phenotype, characterized by the expression of α-smooth muscle actin (α-SMA).50 And bone marrow (BM)-derived fibroblasts can be recruited to the TME and transform into CAFs. Moreover, both BM- and tissue-derived MSCs serve as precursor cells for normal fibroblasts and can directly differentiate into CAFs.51–54 Finally, other cell types, such as epithelial or endothelial cells, adipocytes, and perivascular cells, can undergo metaplasia or trans-differentiation to transform into CAFs.55–57 The transition processes of CAFs are portrayed in the Figure 1.
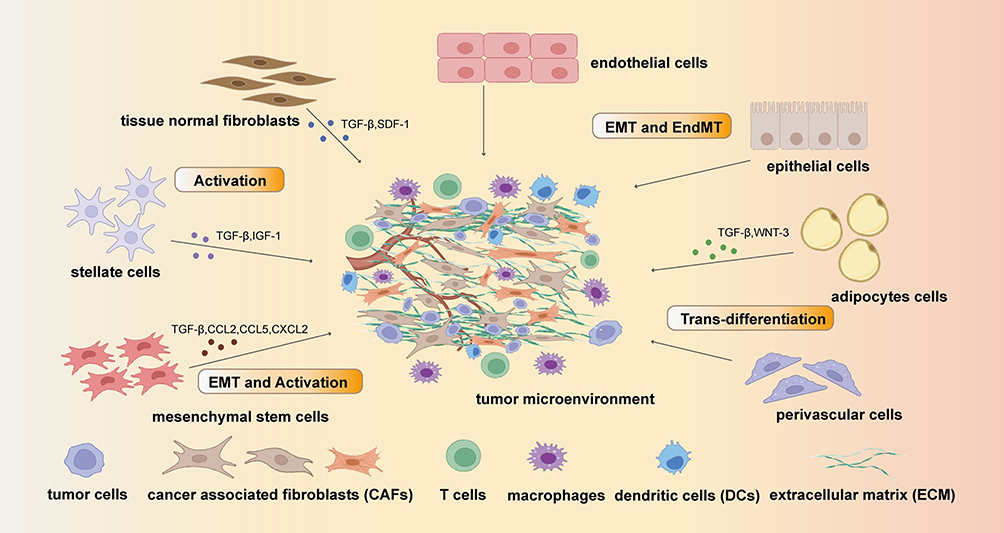 |
Figure 1 The origin of cancer associated fibroblast (CAFs): the main original cell of CAFs are tissue normal fibroblasts, stellate cells, mesenchymal stem cells, endothelial cells, epithelial cells, adipocytes cells and perivascular cells. These cells can transform into CAFs by Epithelial–mesenchymal transition (EMT) pathway and Endothelial-to-mesenchymal transition (EndMT) or be activated by cytokines including TGF-β, SDF-1, WNT-3, IGF-3, CCL2, CCL5 and CXCL2. |
CAFs Heterogeneity
The heterogeneity of CAFs is influenced by their diverse origins and their spatial relationship with tumor cells within the TME.58–60 Distinct types of CAFs often vary in their surface markers and biological characteristics.50 Initially, CAFs were categorized into the myofibroblastic (mCAF) and inflammatory (iCAF) CAF subtypes.47 However, the advancement of single-cell sequencing technology has facilitated the exploration of CAF subtypes and heterogeneity within the TME in more detail. Current research on the classification of CAFs primarily focuses on connective-tissue-rich tumors, such as those of breast and pancreatic cancers. Despite having different tissue specificities, CAFs demonstrate broad similarities.61 The three main CAF subtypes are: mCAFs, pro-inflammatory and immune regulatory CAFs (iCAFs), and antigen-presenting CAFs (apCAFs).59,62 mCAFs remodel the ECM and influence tumor tissue rigidity; they are therefore characterized by the expression of genes associated with collagen formation and smooth muscle contraction (eg, α-SMA).63 mCAF subtypes can be further classified based on the surface expression of functional molecules (eg, TPM1,64 POSTN,65 ACTA2, and MYL966), which is used to identify their origin and ability to promote tumor growth. Moreover, the different mCAF subtypes can morph into each other during different stages of tumor progression by varying their surface molecule expression. In breast cancer, a significant proportion of CAFs originate from PDPN+ CAFs (pCAFs), and turn into the Fbn1+Mfap5+ and Acta2+Thbs2+ CAFs in the later stages.67 Meanwhile, in non-small cell lung cancer (NSCLC), the COL13A1+ and COL14A1+ CAFs (with strong epithelial-mesenchymal transition [EMT] characteristics) dominate in the early stages of disease but later transition into the ECM37+ or ACTA2+ subtypes.68,69 The immunomodulatory properties of iCAFs in the TME depend on the organ they reside in and the associated environmental factors. Tumor cells drive iCAFs to perturb certain elements of the TIME. Simultaneously, iCAFs modulate tumor cells through the secretion of their own substances, thereby enhancing immunosuppression and immune evasion.70,71 iCAFs express relatively low surface α-SMA levels, and are classified into various subtypes based on specific molecular marker combinations.47 For instance, iCAF subtypes can be designated as pCAFs or S100A4+CAF (sCAFs).67 Additionally, CAF subtypes can be distinguished based on the variation in the secretion of certain factors. Moreover, the iCAF and mCAF phenotypes are highly dynamic and can undergo interconversion. Commonly secreted mediators include leukemia inhibitory factor, interleukin (IL)-6, IL-8, IL-11, CXCL, and CCL family members; these mediators are implicated in pathways such as the Hedgehog, IL1R1/JAK/STAT, NF-κB, and classical complement pathways.59,70,72–74 iCAFs have been reported to inhibit the infiltration of DCs, B cells, and T cells, increase the recruitment of MDSCs and Tregs, and promote the polarization of M2 macrophages.59 The discovery of apCAFs indicates that not all CAFs have pro-tumor properties;46 however, the formation of these CAFs largely depends on whether they originate from endothelial or epithelial cells. Moreover, apCAFs typically occur in the late stage of cancer or in individuals with compromised immune function. Overall, few studies have characterized this CAF subtype, and its prevalence, formation, and antigen presenting functions remain unclear. Beyond the three common classifications of CAFs, metabolic CAFs (meCAF, involved in metabolism) and vascular CAFs (vCAFs, involved in angiogenesis) have also been identified.60,65 Despite a substantial body of research addressing CAF classification, no universally accepted categorization method exists. Therefore, considerable strides still need to be made to comprehensively classify CAF subtypes. The example of the subtypes of CAFs are shown in the Figure 2.
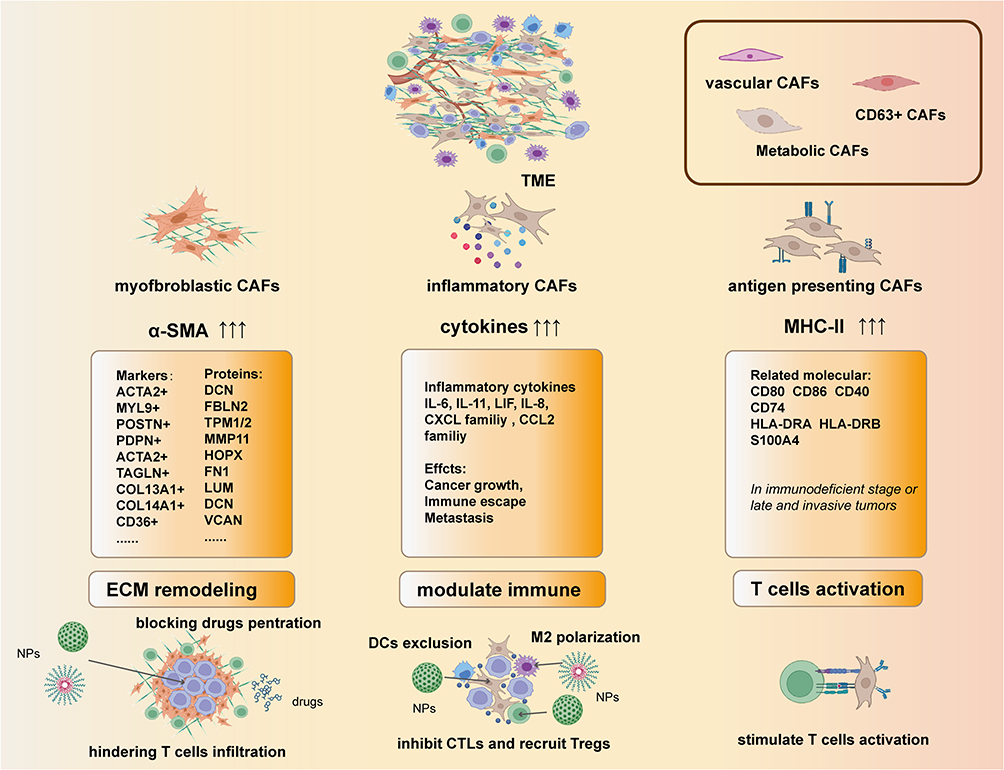 |
Figure 2 The heterogeneity of CAFs: the main subtypes of CAFs are defined by their features. In the classification of mCAFs, the overexpressed genes such as ACTA2, CD36 and PDPN are define as the subtype of mCAFs. And subtypes of the iCAFs with different functions are define by their secreted cytokines. The way of defining apCAFs are similar to myCAFs but the markers are usually human leukocyte antigens and cluster of differentiations (CD). |
The Role of CAFs in the TME
CAFs and Tumor Cells
The interactions between CAFs and tumor cells are mutual. Tumor cells persist creating an education process to recruit and active CAFs consistently. This process is not only controlled by a wild range of tumor-secreted factors (eg, TGF-β, PDGF and IL-6) but also rely on some stimulation and the change of metabolite composition in the TME.7 For instance, the accumulation of reactive oxygen species triggered by local hypoxia can induce the expression of CXCL12 and hypoxia inducible factor 1α to promote the transition of mCAFs.75–77 In the study of Kitamura et al, they found that cancer lactate can be reused by CAFs. CAFs can use monocarboxylate transporter 1 to utilize the lactate to enhance TCA cycle and upregulate the expression of IL-6.78 Activated CAFs can evolve along with the tumor cells and attach great importance in affecting multiple malignant behaviors. In breast cancer and prostate cancer, it has been reported that CAFs can boost the tumor growth and induce tumorigenesis and this effect is mainly due to the interaction between CXCL12 (secreted by CAFs) and CXC-chemokine receptor 4 (CXCR4, expressed on tumor cells).79,80 Rapid tumor growth demands enough nutrients and oxygen so new vessel formation in the TME is necessary. CAFs-derived amounts of pro-angiogenesis factors including WNT2, VEGF, FGF2 and PDGFA can initiate and stimulate this procedure.81 Notably, CXCL12 secreted by CAFs can recruit the vesicular endothelial cells into TME and accelerate vessel formation.7,79,82 CAFs-derived TGF-β and CXCL12 can lead EMT in tumor cells, which contribute to invasion and metastasis. Moreover, CAFs can create gaps in ECM, basement membrane and vascular wall which make it favorable for tumor invasion and metastasis.83 Recent numerous researches have also proved that CAFs are indispensable in the resistance of cancer therapy. CAFs are capable of remolding ECM by secreting collagen and tenascin C (TNC) to form dense physical barriers which can prevent chemotherapeutic and immunotherapeutic drugs from penetrating into the deep parts of tumor.7,14,18 Moreover, CAFs-derived factors can increase the resistance ability of tumor cells. For instance, CAFs can cause resistance to cisplatin-based chemotherapy. Cisplatin can increase TGF-β level to activate CAFs to produce IL-6, which can induce EMT and avoid apoptosis.84 And CXCL12/CXCR4 axis mentioned above can activate Wnt/β-catenin pathway to induce cisplatin resistance.82 Moreover, Xingbo Long et al showed that bladder cancer cells can promote the transformation of CAFs, which then upregulate the IGF1/ERβ pathway in bladder cancer cells to increase their ability of cisplatin resistance.85 In general, CAFs and tumor cells can co-evolve along with cancer progression and the mutual interaction may result in adverse outcomes. This is why CAFs has become the promising target for cancer therapy during recent years. What’s more, eliminating CAFs and tumor cells simultaneously is more important and effective than merely targeting CAFs or tumor cells.
Crosstalk Between CAFs and Immune Cells
CAFs and T Cells
T lymphocytes are the most important adaptive immune cells in the TME. They exert an adaptive immune response by recognizing specific tumor-related antigens.74–77 However, prolonged antigen exposure leads to the upregulation of immune checkpoint proteins (ICPs) on the surface of T cells, which impairs their ability to mount an effective immune response and results in reduced proliferation, loss of memory function, and exhaustion.78,79 CAFs play a significant role in inducing T cell exhaustion and have a potent capacity to induce the differentiation of Tregs. Notably, Sakai et al observed that the infiltration of Foxp3+ CD4+ T cells was accompanied by increased numbers of WNT2+ FGFR2+ CAFs.80 In vitro experiments have also demonstrated a significant increase in Tregs within the peripheral blood mononuclear cell (PBMC) population in the presence of CAFs.81 Nonetheless, the relationship between CAFs and Treg infiltration in tumors remains unclear, with some studies suggesting that partial CAF depletion actually promotes Treg infiltration into the tumor, thereby promoting tumor progression.82,83 The influence of CAFs on CD8+ T cells is also multifaceted. First, CAFs can effectively exclude CD8+ T cells from the tumor core, causing their marginalization.84 Second, CAFs can impede the activity and proliferation of T cells. In vitro studies have demonstrated that co-culturing CAFs with PBMCs significantly reduced CD8+ T cell numbers and their ability to secrete cytotoxic molecules.81 CD8+ T cells and CAFs also show great relation in metabolic regulation. Weimin Wang. et al reported that CAFs can produce cysteine and glutathione to lead resistance to platinum-based therapy. While CD8+ T cells can control the release through IFN-γ.86 Additionally, CAFs themselves can express ICPs such as PD-L1, which induce ICP expression on the surface of CD8+ T cells and exacerbates their exhaustion. Notably, LRRC15+ CAFs have been positively associated with the expression of multiple exhaustion molecules, including TIM3, LAG3, and CD39, on T cells.85,86 (Figure 3)
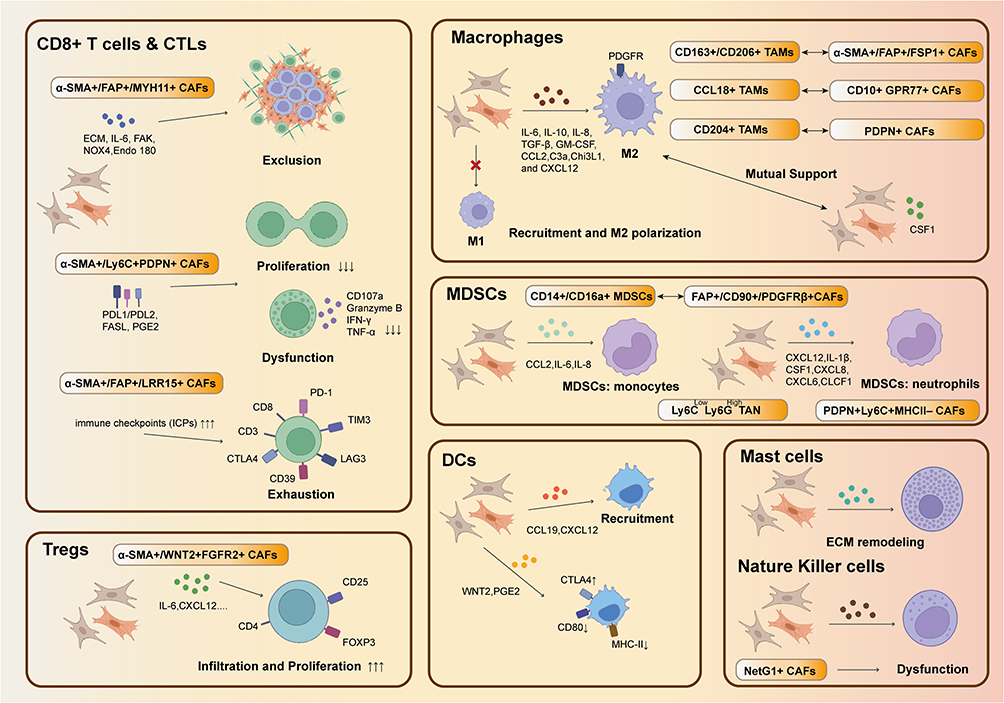 |
Figure 3 Crosstalk of CAFs and immune cells: CAFs can inhibit CD8+ T cells by inducing exclusion, dysfunction and exhaustion and they can also inhibit their proliferation while induce the proliferation of Tregs. CAFs can induce M2 polarization while M2 can also enhance the function of CAFs. This kind mutual support also can be found in the interaction of CAFs and MDSCs. CAFs also can disrupt the functions of DCs and NK cells while recruit mast cells. |
CAFs and TAMs
Macrophages are the most abundant and extensively studied innate immune cells in the TME. Two distinct phenotypes of macrophages exist: M1 macrophages have anti-tumor properties, while M2 macrophages exhibit pro-tumor characteristics.87 CAFs and macrophages behave similarly within tumors, exerting their functions through autocrine and paracrine signaling mechanisms.88–90 For instance, the KRAS pathway induces M2 macrophage polarization and enhances the capacity of CAFs to secrete IL-6 and CXCL1.91 mCAFs and macrophages can communicate extensively with each other within the TME.92 Moreover, the presence and functions of tumor-associated macrophages (TAMs) and CAFs appear to be intertwined across various tumor types. In breast cancer, the presence of CD163+ and CD206+ TAMs positively correlate with that of α-SMA+ CAFs.89 Similarly, CD204+ TAMs in lung cancer exhibit concordant infiltration patterns with FDPN+ CAFs.93,94 The co-localization of TAMs and CAFs often implies drug resistance and immunosuppression, implying that these cell types can synergistically induce adverse effects. CAF-derived cytokines and chemokines (eg, IL-6, IL-10, GM-CSF, and CXCL12) recruit monocytes into the TME, facilitating their polarization into M2 TAMs.36,95–97 In gastric cancer, CAFs promote the transformation of M2 macrophages through periostin expression.98 Moreover, CAFs exacerbate immunosuppression in breast cancer by secreting chitinase-3-like 1, which attracts M2 TAMs and leads to CD8+ T cell depletion.99 In turn, TAMs maintain persistent CAF activation, which further contributes to breast cancer tissue fibrosis and sclerosis through the activation of the TGF-β signaling pathway.100 Additionally, macrophages cause MSCs to adopt CAF-like characteristics in vitro by upregulating markers such as α-SMA, FAP, and vimentin.53 The intricate interplay among CAF, TAMs, and tumor cells facilitates tumor initiation and progression (Figure 3).
CAFs and MDSCs
MDSCs contribute significantly to the formation of the immunosuppressive TIME. Tumor-associated cytokines drive the differentiation of immature myeloid cells into MDSCs, which share biological similarities and exhibit close associations with CAFs.101 In lung cancer, a positive correlation has been observed between the expression of MDSC- (eg, CD14 and CD16) and CAF- (eg, FAP and PDGFR-β) associated surface markers, implying the co-infiltration of CAFs and MDSCs.102 Likewise, CAFs can recruit MDSCs or influence their functions through various pathways. For example, CAFs promote MDSC infiltration and increase the expression of adhesion molecules on MDSCs by secreting IL-6, IL-8, and CCL2.96,103,104 Moreover, CAFs play a role in MDSC differentiation, for instance, by inducing the expression of APOE and TREM-2 expression in MDSCs, thereby enhancing their ability to suppress CD8+ T cells92 (Figure 3).
CAFs and DCs
As professional antigen presenting cells, DCs prime T cells to recognize and eliminate tumors.105 CAFs have been found to downregulate the expression of MHC class I and II molecules on the DC surface, thereby impeding T cell priming.106 In addition, CAFs interfere with the immunomodulatory function of DCs, establishing an immunosuppressive environment. Conversely, CAFs can recruit DCs to the tumor site via the CXCL12 and CCL19/CCR7 axes.107 In colon cancer, the secretion of WNT2 by CAFs significantly hinders the activation and maturation of DCs.108 Moreover, CAFs can inhibit DC function by promoting the expression of ICPs, such as CTLA-4, on the DC surface106 (Figure 3).
CAFs and Cancer Therapy
As previously mentioned, most CAFs inhibit the pro-tumor immune response within the TME. Therefore, over the past few years, a considerable number of immunotherapeutic approaches targeting CAFs have been developed (Table 1); some of these have successfully entered the preclinical and clinical trial stages.109 Three main CAF-targeting strategies have been employed: CAF elimination, CAF inhibition, and CAF modulation.61 Unlike most other immune cells, CAFs do not have a well-defined molecular signature; thus, current studies have mainly targeted the molecules FAP and α-SMA, which are highly expressed on the CAF surface.50,110 FAP targeting using FAP inhibitors (eg, PT630111 and sibrotuzumab112), DNA cancer vaccines,113,114 and CAR-T cells115 has been relatively well documented. Moreover, a dual-specificity antibody drug targeting CD137 and FAP (RO7122290) is being investigated in clinical trials.116 These drugs and therapeutic methods have all demonstrated similar tumor-inhibiting effects. However, results from a recent Phase II trial of an FAP-specific monoclonal antibody (PT100) showed no clinical benefits.117,118 Moreover, FAP-based oncolytic adenoviral vaccines are merely at the preclinical stage of development; this may due to concerns regarding their low efficacy and stability. Similarly, because CAR-T cell therapy is seldomly used in solid tumor, FAP-specific CAR-T cell therapy is unlikely to be applied to the treatment of all tumor types. Furthermore, not all CAFs express FAP, making it challenging to completely eliminate tumors by targeting FAP-positive cells. Moreover, as FAP is also expressed on healthy cells, targeting this molecule can result in off-target toxicity and adverse effects. Thus, FAP-specific CAR-T cell therapy is likely to induce cytotoxicity,119 while an α-SMA-targeting approach will likely increase Treg infiltration.120 Therefore, restoring the stromal balance in tumor tissue rather than eliminating CAFs altogether has been put forward. And this can be achieved using TGF-β pathway inhibitors, such as rizaben and galunisertib.121–123 However, these inhibitors often induce side effects such as neutropenia, liver toxicity, and thrombosis. In addition, vitamin A (eg, all-trans retinoic acid [ATRA]124) and vitamin D (eg, calcipotriol125) analogs have been used to interfere with CAF activation.126,127 When used in combination with chemotherapy and radiotherapy, these inhibitors can effectively enhance the anti-tumor immune response within the TME and achieve tumor growth inhibition. Unfortunately, these approaches are still in the preclinical stage and may harm T cell immunity.128 Recently, Li et al have investigated the positive role of calcipotriol in tumor metastasis and recurrence after adjuvant photothermal therapy (PTT).129,130 Targeting important pathways within CAFs is another research focus. Examples of pathway inhibitors include AMD3100,131 an inhibitor of the CXCL12-CXCR4 pathway; ruxolitinib,132 a JAK pathway inhibitor; and sonidegib,133 IPI-926,134 and GDC-0449 (vismodegib),135 inhibitors of the Hedgehog pathway. In preclinical studies, AMD3100 has shown promising results when used in combination with ICIs; however, precisely how it modulates the TME remains unclear.131 In a phase II clinical trial of pancreatic cancer, ruxolitinib has demonstrated expected outcomes, with beneficial effects primarily observed in patients with high C-reactive protein levels.132 Despite this partial response, ruxolitinib is still regarded as a promising drug. GDC-0449, an inhibitor of the Hedgehog ligand SHH, did not confer benefits when used in combination with gemcitabine for pancreatic cancer; in addition, over half of the patients experienced significant drug toxicity.135
As described above, targeting CAFs therapies are currently unresolved challenges in clinical trials. Moreover, the efficacy of individual CAF-targeting methods is limited, while the use of combination therapies often leads to more adverse drug reactions and increases treatment complexity. The emergence of nanotechnology has improved the precision of drug delivery while optimizing drug release mechanisms.17,20 Additionally, the unique properties of NPs bring forth diverse treatment modalities, such as PTT.20 Immune therapies utilizing specifically tailored nanomaterials have demonstrated remarkable results in the treatment of various tumors.16 Therefore, exploring the value of using NPs in targeting tumors and CAFs has been the new research orientation.
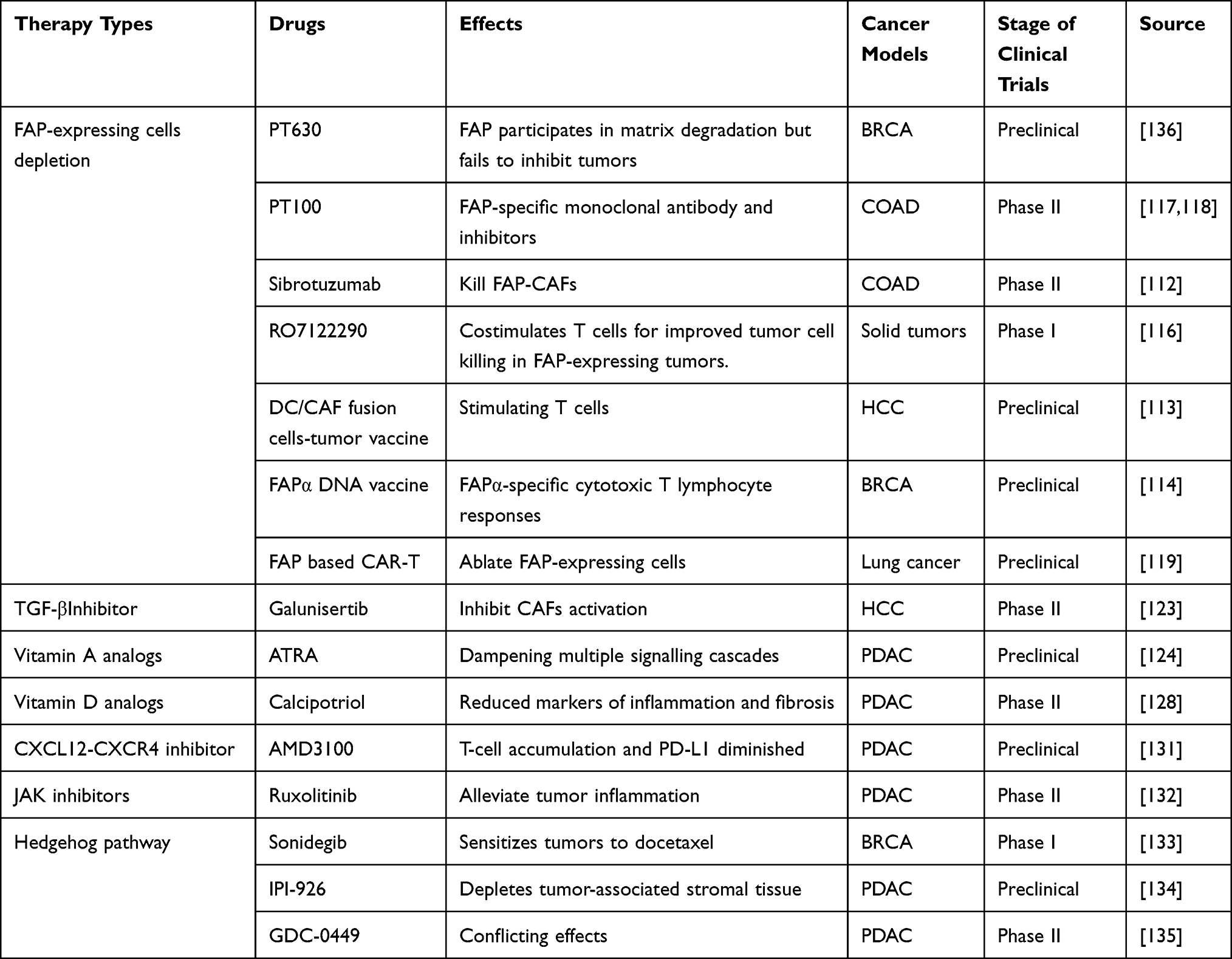 |
Table 1 Drugs for Targeting CAFs in Preclinical and Clinical Trials |
The Advantages and Functions of NPs
In the past few decades, medical nanotechnology has developed rapidly, and many inorganic biologically inert materials have been applied to nanomedicines. Compared with common drug molecules, nanomedicines have better performance in targeting, stability, long-term efficacy, versatility and safety.137 The advantages of nanomedicines benefit from their functions. 1) NPs have a stealth effect, that is, they are in a state of immune escape so they will not be quickly cleared in the body. The surface charge of NPs can make them not be cleared in the glomerular filtration membrane, especially for some small molecules. Neutral and hydrophilic polymers can improve the stability and reduce biological interactions in Vivo, and thus prolong the retention time in tissues and blood. The application of polyethylene glycol (PEG) as a modification group is the best proof.138 Bio-derived NPs can also bring this stealth effect. Nanomedicine based on red blood cell (RBC) membrane can improve the stability owing to the low immunogenicity and similar characteristics to RBC in blood. Albumin has become a good drug carrier because its size and charge characteristics are not filtered by glomeruli. Recently, CD47 has been found as a sign of ‘don ‘t eat me ‘signal to macrophages, which has also led to an increasing number of modified with CD47-modified NPs to achieve stealth effects.138 2) NPs provide the basis for the design of multifunctional drugs, which can accurately target tumor. Ph response strategy and redox response strategy are the main methods. They can take advantage of the specific Ph changes in tumor tissue and the specific overactive redox environment. More interestingly, these can bring the cutting effect of PEG, which is conducive to the release and uptake of drugs. At the same time, NPs modified with cell-specific markers can achieve the expected precise targeting effect. For example, nanoparticles modified with macrophage surface marker CD80, CD86 and other molecules can accurately locate macrophages, integrate into the cell.137 As discussed in this review, NPs modified with CAF surface marker FAP show a high targeting effect on CAF. 3) Some effects of NPs can be triggered in vitro. Light, magnetic field, and ultrasound can trigger effects such as self-luminescence (commonly used in surgical tracers) and photothermal effects (metal nanoparticles, carbon-based nanoparticles, nano-polymers). This feature makes it more versatile, and in the application, doctors can more actively grasp the timing of treatment.137
In cancer therapy, nanomedicines need to enter the tissue through blood vessels, then infiltrate into it to release, and eventually be taken up by targeted cells. Macromolecular drugs can infiltrate into solid tumor tissues through blood vessels and accumulate in tumor tissues, which is the enhanced permeability and retention (EPR) effect. This effect is thought to be fundamental for the design of nanomedicine and it is affected by the size, shape, softness and surface properties.139,140 In the research from Shrey Sindhwani, they thought that the way of entering tumors by processing endothelial cells is more common than EPR effect for NPs141 And there is also a concern that the EPR effect may cause an increased risk of metastasis.142 Moreover, EPR effect is limited in clinical applications due to its heterogeneity which is caused by the individual differences between patients.140 Although there exist some controversies, enhancing EPR effect is still an effective method to improve curative effect in the current. CAF is the barrier for NPs to enter tissue due to its effect on the ECM (CAF makes the tumor become solid, which is not conducive to the penetration of drugs, and impair the diffusion of drugs. CAF can also absorb drugs, which will cause a decrease in drug concentration.7 Therefore, the depletion of CAFs in cancer therapy can be considered as a way to enhance the EPR effect from the biological perspective. However, this method can bring risks because that eliminating CAFs may promote tumor progress, which is similar to the risk from EPR effect. So, there also exist many researches on disrupting the functions of CAFs and inhibiting its activation. We have summarized the examples to show how NPs are applied for targeting CAFs in Table 2. The types, the targeting sites, the loaded drugs and the targeting strategies are listed. All of these will be discussed thoroughly in the following sections.
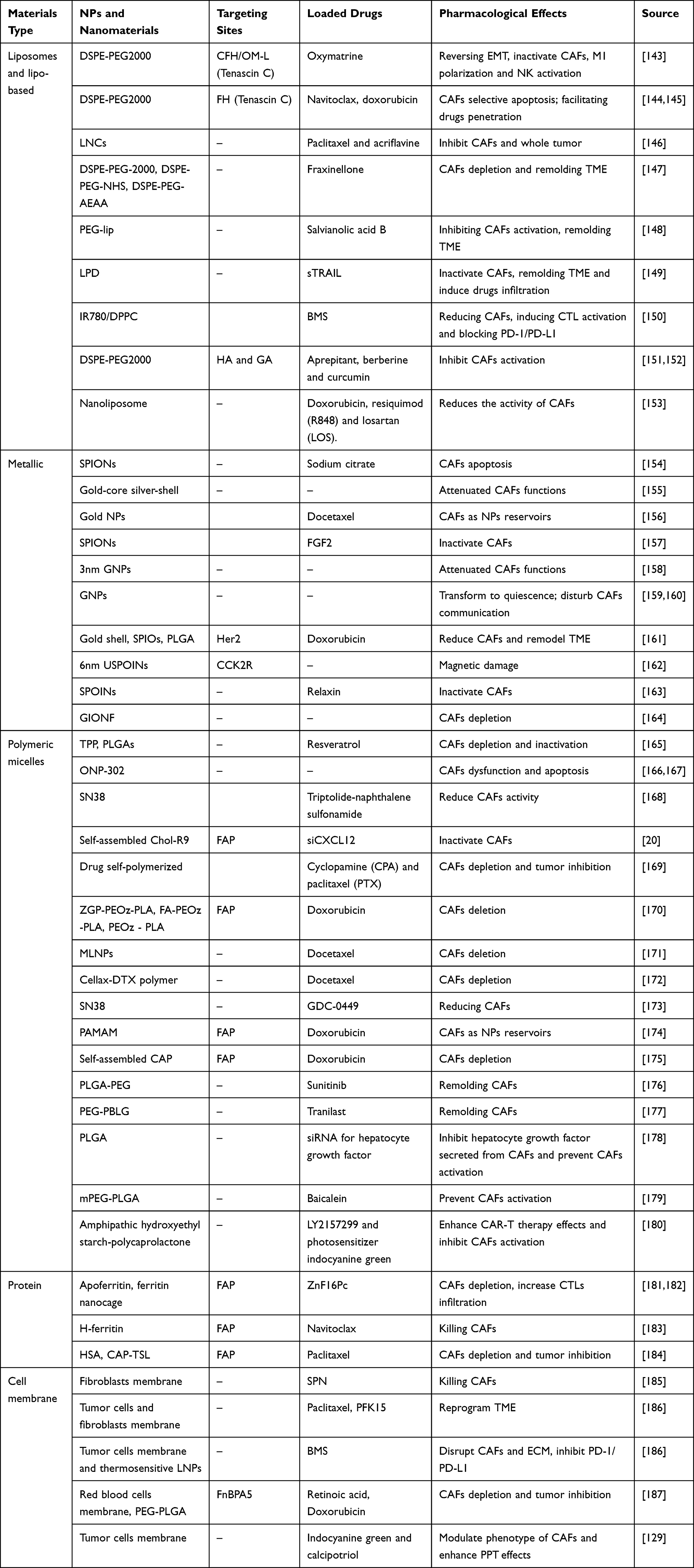 |
Table 2 Constructed NPs and Their Effects for Targeting CAFs |
NPs Targeting CAFs in Cancer Therapy
Drug Delivery Systems for Targeting CAFs
Liposomes and Lipids
Liposomes, which are characterized by high biocompatibility, sustained release, and effective immune cell targeting, have revolutionized drug delivery.188 Owing to its exceptional hydrophilicity and hydrophobicity, PEG is frequently employed as a linker to connect drugs and liposomes. Recent studies have explored the potential use of PEG in the development of CAF-targeting NPs.189 The targeting efficiency of liposomes can be further improved by attaching peptides or antibodies that recognize specific biomarkers.190 While FAP is commonly used as a CAF biomarker for guiding NPs,191 TNC,192 which is expressed in the ECM, has also been increasingly used to target CAFs. The FH and CFH peptides exhibit high binding affinity for TNC;193 thus, by conjugating these targeting moieties to NPs, the precise targeting of CAFs can be achieved. Moreover, optimized liposome-based NPs, such as lipid nano-capsules, have demonstrated even higher efficiency than conventional liposomes.194 Additionally, DSPE-PEG2000, known for their stable chemistry, can serve not only as raw materials for liposome synthesis but also as efficient drug carriers. Conjugation of DSPE-PEG2000 to liposomes extends the half-lives of these delivery systems.144,145 Several traditional Chinese medicines, including oxymatrine,143 salvianolic acid B (SAB),148 and fraxinellone (Frax)147 have also shown great potential as fibroblast modulators when conjugated to DSPE-PEG2000. Multifunctional lipo-based NPs are also developing currently. For instance, thermosensitive lipid NPs can be constructed using the photothermal agent IR-780 and the phospholipid Dipalmitoylphosphatidylcholine (DPPC) (Figure 4).150 Furthermore, the potential of using liposome complexes with polymeric micelles or biomacromolecules in targeting CAFs has been investigated. Notably, lipid-coated protamine DNA (LPD) complexes exhibit exceptional transfection efficiency, making them suitable for the delivery of drugs that regulate intracellular metabolism and pathways. Indeed, loading LPD complexes with TNF-related factors has been shown to effectively inhibit CAFs and induce their apoptosis.149
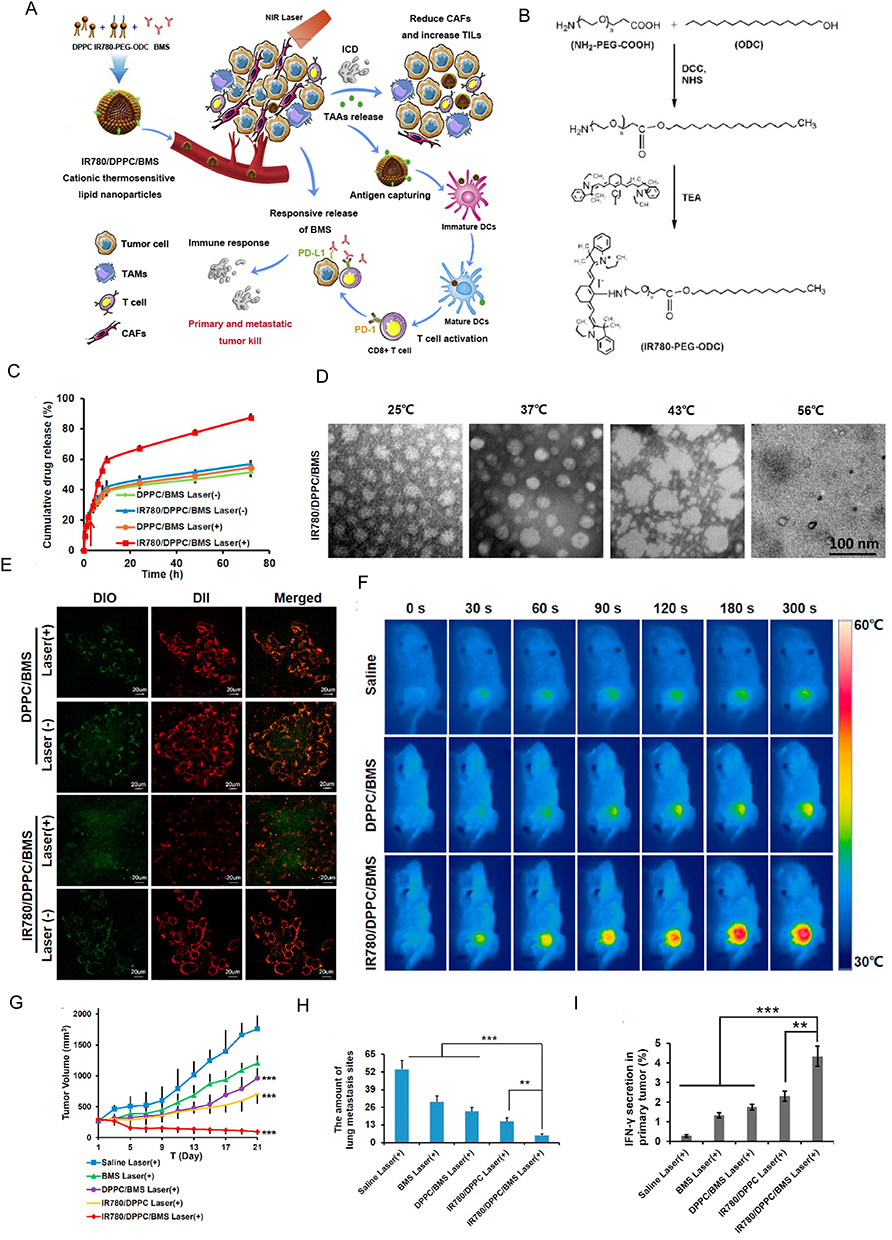 |
Figure 4 Design, characteristics and effects of IR780/DPPC/BMS. (A) Scheme of IR780/DPPC/BMS. (B) Synthetic route of IR780-PEG-ODC. (C) In vitro NIR-triggered drug release profiles. (D) Transmission electron microscope (TEM) images of IR780/DPPC/BMS at different temperatures. (E) Cellular uptake and responsive drug release in 4T1 tumor cells. (F) In vivo photothermal effect. (G) Tumor volumes of 4T1 tumor-xenografted mice in different treatment groups. (H) The quantitative analysis of pulmonary metastatic nodules in various treatment groups. (I) Detection of released cytokines in primary tumor after synergistic immuno-photothermal therapy. *:P<0.05; **:P<0.01; ***P<0.001. Reprinted from Cancer Letters, volume 522, Tan Y-N, Li Y-P, Huang J-D, et al. Thermal-sensitive lipid nanoparticles potentiate anti-PD therapy through enhancing drug penetration and T lymphocytes infiltration in metastatic tumor. 238–254, Copyright 20221, with permission from Elservier.150 |
Metals and Metal Oxides
Metallic NPs and their derivatives possess several advantageous properties, including sustained release, local heat generation, high photothermal sensitivity, high affinity, and improved irradiation effects. These unique characteristics enable them to effectively target CAFs and remodel the TME.162,195,196 Among the types of metallic NPs, gold NPs (GNPs) and iron NPs are the most commonly used. CAFs exhibit a 7-fold higher rate of GNP uptake and a 3-fold longer rate of GNP retention than tumor cells (Figure 5).197 The therapeutic effect of GNPs can be further enhanced by combination with docetaxel (DTX); this may be because GNPs sensitize CAFs to external factors and promote drug absorption. Thus, GNPs not only serve as ideal carriers for CAF-targeted therapies but also remodel CAFs into drug repositories, leading to sustained effects. In vitro studies have demonstrated that GNPs can reduce radiotherapy resistance in tumor and CAF co-culture models by suppressing CAF activity.198 Additionally, GNPs promote the transition of CAFs from an activated to a resting state; this may occur through the regulation of cellular lipid metabolism and the disruption of communication between tumor cells and CAFs via altered soluble factor secretion.159,160 The functional properties of GNPs are influenced by their size; 3 nm is considered as the optimal GNP diameter.158 Furthermore, multifunctional and optimized GNPs-related NPs have recently been developed. Hybrid gold and silver NPs, with a core-shell structure, have been designed to modulate CAFs even more effectively than conventional GNPs.155 In these particles, gold acts as the Trojan-horse for precise silver delivery, which in turn effectively interferes with CAF-mediated communication pathways.199 Gold-shell-based photothermal NPs also enhance the CAF killing ability of GNPs and promote targeted drug release.163 Among the iron-based NPs, superparamagnetic iron oxide nanoparticles (SPOINs) have demonstrated exceptional in vivo compatibility.156,162,200 Citrate-coated SPOINs can be easily absorbed by CAFs within 15–30 minutes of treatment.154 Due to their magnetic properties, SPOINs can induce the mechanical destruction of CAFs under low-frequency magnetic conditions,201 with smaller SPOINs proving most effective. Additionally, when exposed to alternating current magnetic fields, SPOINs can be used as a thermal therapy and induce apoptosis in CAFs.154 Finally, SPOINs have a large surface area and are richly decorated by various functional groups, which serve as favorable drug attachment sites.156 As such, this type of NP can enhance the half-life, solubility, and therapeutic effects of drugs. Notably, biomolecules such as relaxin202 and fibroblast growth factor (Figure 6) 157 have been successfully conjugated to SPOINs to inhibit CAF differentiation and activation, ultimately reducing the extent of fibrosis in the TME. Moreover, the combination of gold and iron architectures enhances the CAF-targeting efficiency of NPs constructed using either metal alone. For instance, the iron-oxide-modified GNPs are preferentially absorbed by CAFs, where they act as a thermal source to effectively kill CAFs, reduce tumor stiffness, and ultimately lead to complete tumor elimination.163
 |
Figure 5 Docetaxel (DTX)-Mediated Uptake and Retention of Gold Nanoparticles (GNPs) in Tumor Cells and in CAFs. (A) Schematic diagram of functionalized GNP. (B) TEM of GNPs. (C) Zeta Potentials for pure GNPs. Scheme of transportation of GNPs in cells (D) and effects of DTX on cell division (E). Confocal microscopy images of GNPs transportation (F) and effects of DTX on cell division (G). (H) Effect of DTX on GNP. Retention of NPs in the presence of DTX in tumor cells (I) and CAFs (J). (L) Retention of GNPs in tumor cells and CAFs (K) post recovery. *:P<0.05. Reprinted from Alhussan A, Bromma K, Perez MM, et al. Docetaxel-mediated uptake and retention of gold nanoparticles in tumor cells and in cancer-associated fibroblasts. Cancers (Basel). 2021;13(13):3157. Creative Commons.197 |
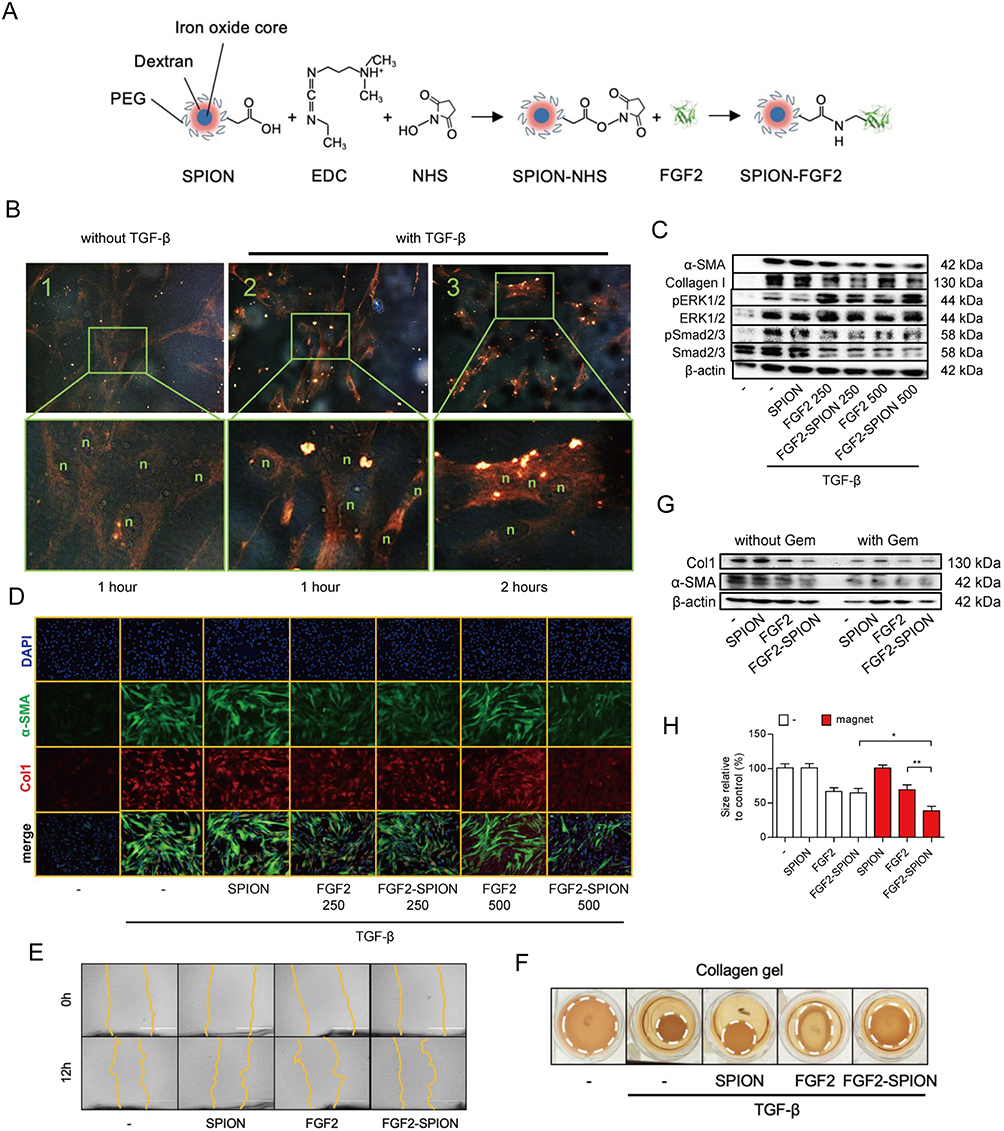 |
Figure 6 Fibroblast growth factor 2 (FGF2) engineered SPIONs attenuate tumor stroma and potentiate the effect of chemotherapy in pancreatic tumor. (A) Schematic representation of conjugation of FGF2 to SPION. (B) Binding of FGF2-SPION on non-activated human pancreatic stellate cells (hPSCs) and activated hPSCs. Western blots demonstrating the differentiation-related biomarkers (C) and representative immunofluorescence images showing the effect of FGF2 and FGF2-SPION on the protein expression of α-SMA and col-1 in TGF-β-activated hPSCs (D). Effect of FGF2-SPION on hPSCs migration (E) and contractility (F). (G) Effect of FGF2-SPIONs on the tumor stroma and gemcitabine efficacy in 3D heterospheroids. (H) Magnetic driven iron oxide accumulation. *P<0.05; **P<0.01. Reprinted from Mardhian DF, Vrynas A, Storm G, et al. FGF2 engineered SPIONs attenuate tumor stroma and potentiate the effect of chemotherapy in 3D heterospheroidal model of pancreatic tumor. Nanotheranostics. 2020;4(1):26–39. Creative Commons.157 |
Polymeric Micelles
In recent years, polymeric micelles have been rapidly adapted as NPs for use in cancer therapy. Their significant advantage lies in their ability to penetrate into cells independently of the properties of their drug cargo.164,203,204 Poly(lactic-co-glycolic acid) (PLGA) is one of the commonly used nanomaterials.205 An example is ONP-302, a PLGA-based NP specifically designed to target TAMs and CAFs. With a diameter ranging from 400 to 800 nm and a negative surface charge of 35 to 50 mV, ONP-302 selectively targets CAFs, inhibits their cancer-promoting function, and induces their apoptosis.166,167 PLGA-like polymers are commonly employed to construct multi-response and sequential delivery NPs (Figure 7).165,206 FAP serves as the molecular target, while pH-responsive groups like folic acid, benzimidazole, and β-cyclodextrin are conjugated to PLGA-like polymers to form the complete CAF-modulating agents.170 Certain molecules, such as amphiphilic peptides, 9-arginine, and PEG5K-P(MMESSN38)5K (PSN38) (Figure 8), possess both hydrophilic and hydrophobic properties.168,173 These molecules have the ability to self-assemble into carrier cages when exposed to the appropriate medium. While these NPs exhibit excellent drug-loading properties, the addition of cell localization groups, such as FAP, is often necessary. Other polymers that have been reportedly used for CAF targeting include a complex of carboxymethylcellulose and DTX, which preferentially accumulates in CAFs and causes their depletion.172 Micellar-like NPs synthesized from poly (ethylene glycol)-b-poly(lactide)-co-poly(N3-α-ε-caprolactone) demonstrate similar effects.171 Furthermore, the positive surface charge of cationic poly(amidoamine) (PAMAM) enables better drug penetration into CAFs.174 Finally, the self-assembly of anti-CAF agents and anti-neoplastic drugs into NPs has been identified as a promising approach for cancer therapy.175
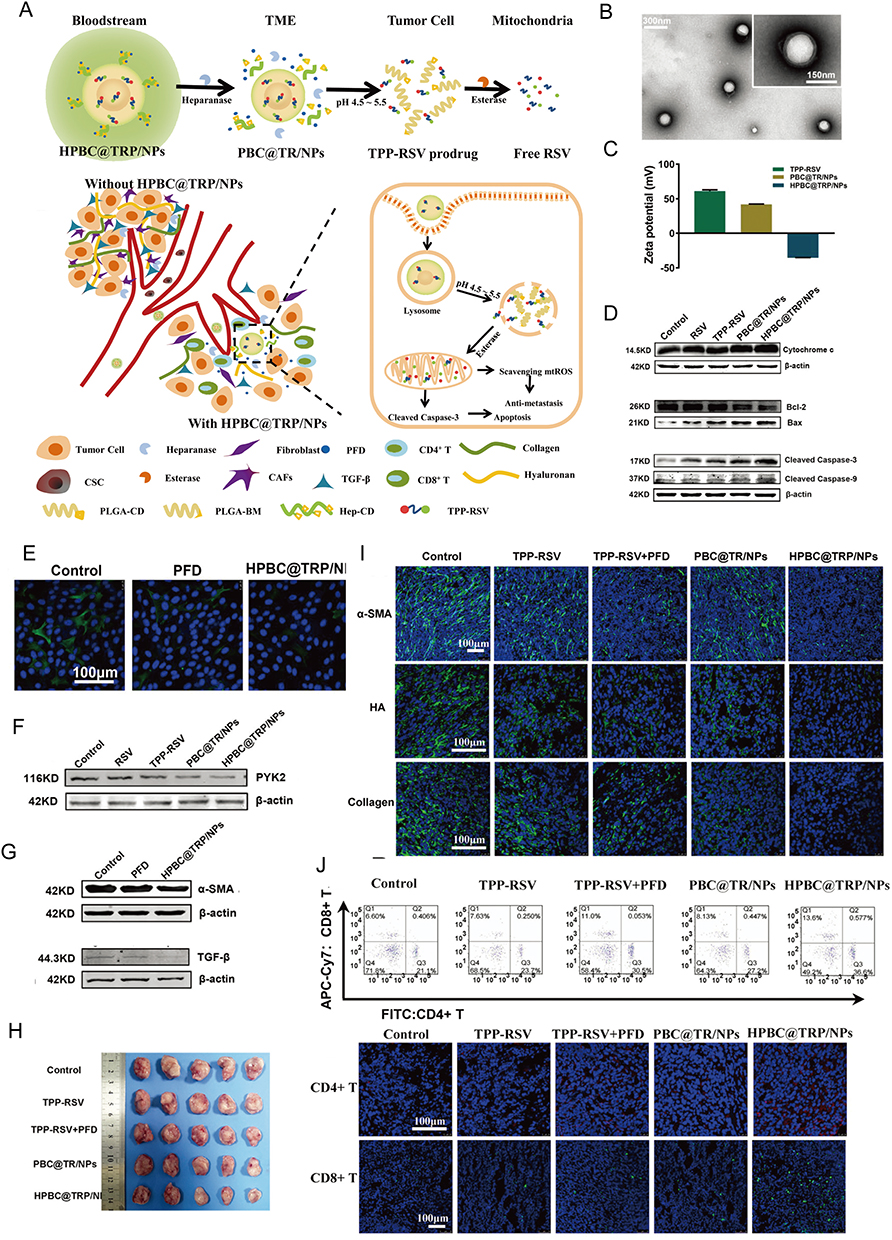 |
Figure 7 On-demand responsive nanoplatform mediated targeting of CAFs and down-regulating mtROS-PYK2 signaling for antitumor metastasis. (A) Disintegration of the on-demand successively responsive HPBC@TRP/NPs and its therapeutic mechanism. (B) TEM images of HPBC@TRP/NPs. (C) Zeta potential reversal characteristic. (D) Evaluation of the relative cell viability and mechanism of the apoptosis signal pathway. (E) CLSM images of α-SMA activated CAFs in 4T1/NIH3T3 cells treated with PFD and HPBC@TRP/NPs. Down-regulation of the mitochondrial oxidation system and cancer associated fibroblasts in vitro: Western blot protein expression levels of PYK2 (F), α-SMA (G), and TGF-β (G) in 4T1/NIH3T3 co-cultured cells. (H) Enhanced in vivo therapeutic effect. (I) Normalization of the tumor ECM to remodel TME. (J) Modulation of TIME in 4T1/NIH3T3 tumors after treatments. Reprinted from Zuo T, Zhang J, Yang J, et al. On-demand responsive nanoplatform mediated targeting of CAFs and down-regulating mtROS-PYK2 signaling for antitumor metastasis. Biomater Sci. 2021;9(5):1872–1885. 2021 © Royal Society of Chemistry.165 |
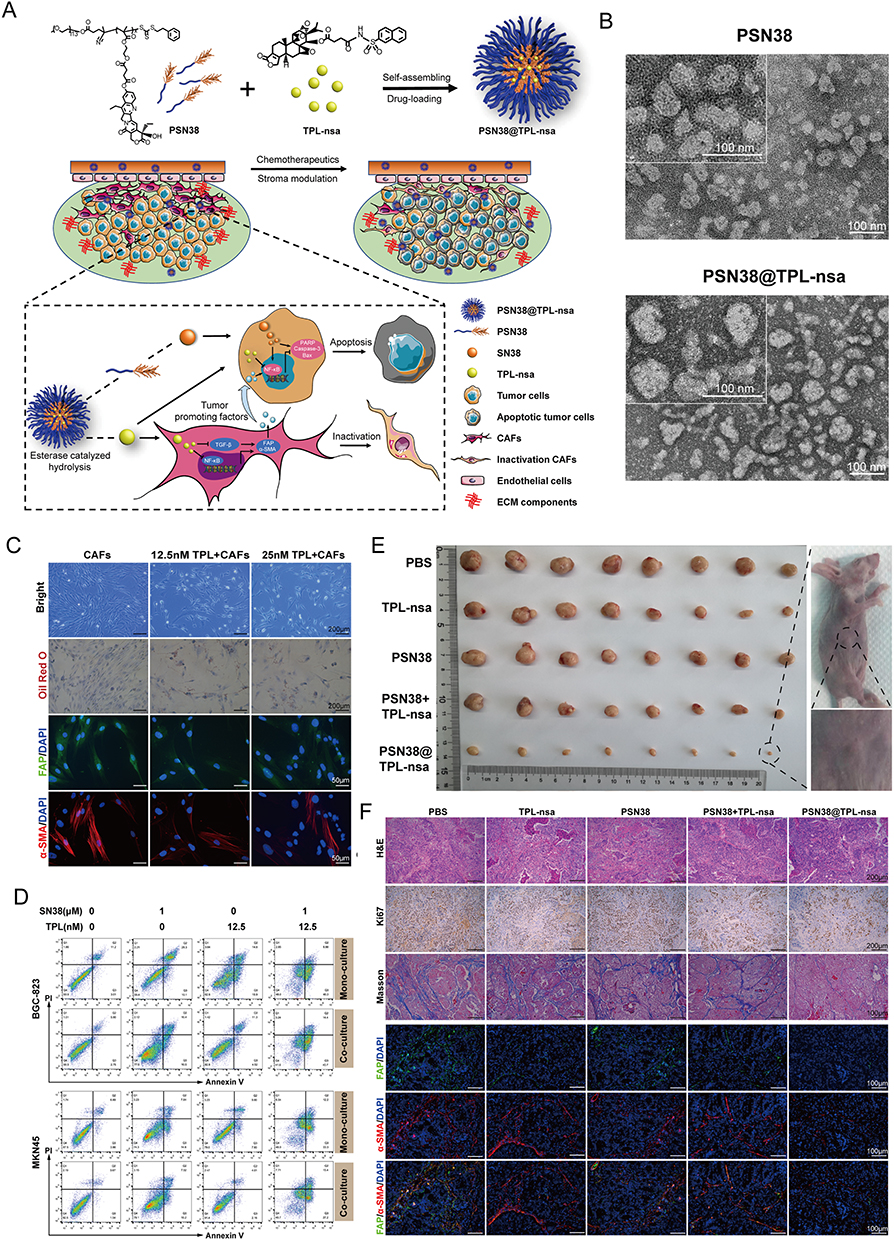 |
Figure 8 Prodrug polymeric micelles integrating CAFs and synergistic chemotherapy for gastric cancer (GCs). (A) Schematic overview of the composition and synergistic mechanisms. (B) TEM images of PSN38 and PSN38@TPL-nsa NPs. (C) Low dose of TPL inactivated CAFs and inhibited CAFs. (D) Apoptotic analysis of BGC-823 and MKN45 cells after the treatment of TPL, SN38 or a combination of SN38 and TPL for 24 h in the co-culture and mono-culture model respectively. (E) The antitumor efficiency of PSN38@TPL-nsa in GC PDX model. (F) PSN38@TPL-nsa remodeled GC microenvironment in vivo. Reprinted from Zheng S, Wang J, Ding N, et al. Prodrug polymeric micelles integrating cancer-associated fibroblasts deactivation and synergistic chemotherapy for gastric cancer. J Nanobiotechnology. 2021;19(1):381. Creative Commons.168 |
Proteins
Ferritin and albumin are the primary protein types utilized in the construction of NPs for CAF targeting. These proteins possess similar chemical characteristics and are natural biomacromolecules that elicit minimal immunogenic reactions.207–209 The presence of amino acid residues within these protein-based NPs provides multiple binding sites for drug loading, and their amphoteric nature allows for pH-responsive behavior within the TME.210 Ferritin and albumin have been employed as nanocarrier cages, to which FAP -specific single-chain variable fragments have been conjugated to target CAFs (Figure 9).181,184 These NPs have been recently modified to encapsulate ZnF16Pc, a common photosensitizer, for use in photodynamic therapy (PDT).181,182 These NPs not only selectively eradicate CAFs to halt tumor progression but also activate T cells and enhance the efficacy of ICI therapy.
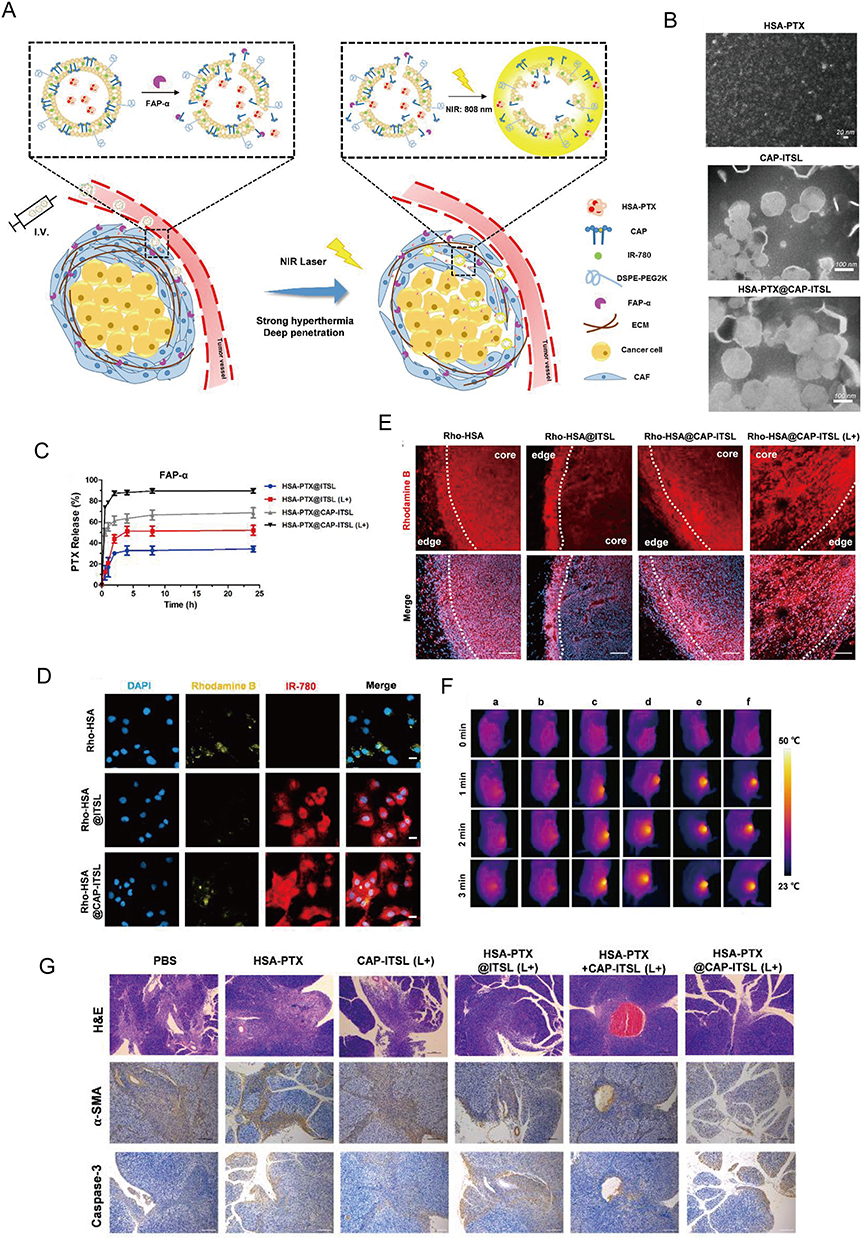 |
Figure 9 Targeting CAFs by dual-responsive lipid-albumin NPs to enhance drug perfusion for pancreatic tumor therapy. (A) Diagram of dual-responsive lipid-albumin NPs (HSA-PTX@CAP-ITSL) (B) TEM images of HSA-PTX, CAP-ITSL and HSA-PTX@CAP-ITSL. (C) The PTX release profiles of different formulations incubated with FAP-α. (D) cellular uptake after TGF-β activated NIH 3T3 cells incubated with different formulations. (E) Rhodamine B distribution of different treatment groups. (F) IR thermal images of subcutaneous tumor-bearing mice in different treatment groups.(G) H&E staining, α-SMA staining and Caspase-3 staining of tumor tissues. Reprinted from J Control Release, volume 321, Yu Q, Qiu Y, Li J, et al. Targeting cancer-associated fibroblasts by dual-responsive lipid-albumin nanoparticles to enhance drug perfusion for pancreatic tumor therapy. 564–575, Copyright 2020, with permission from Elsevier.184 |
Cell Membranes
Cell membranes have recently gained significant attention as NP biomaterials as they closely resemble the cellular components of the TME. Cell membrane surfaces exhibit a greater diversity of receptors and target molecules than the surfaces of synthetic NPs, allowing them to more effectively simulate the biological characteristics and functions of their source cells.211–213 Currently, the application of CAF-targeting cell membrane NPs is not widespread, and the main sources of cell membranes include RBCs, tumor cells, and fibroblasts. RBC-based NPs can protect peptides used in cell targeting from degradation.187 For instance, the FnBPA5 peptide exhibits a strong affinity for CAFs and the ECM, but its rapid metabolism poses a challenge. Encapsulating the external peptide segment of PLGA-PEG-FnBPA5 within RBC-based NPs enhances its targeting ability.187 The homology between the CAF membranes of NPs and those of their target cells makes them ideal for modulating CAFs.185,214 Moreover, when used as adjuvants, tumor cell membranes can activate immune responses to enhance the efficacy of anti-fibrotic drugs.186 Furthermore, the hybridization of the two types of cell membranes was more effective than using a single type, as it resulted in the targeted damage of both CAFs and tumor cells in the TME (Figure 10).215
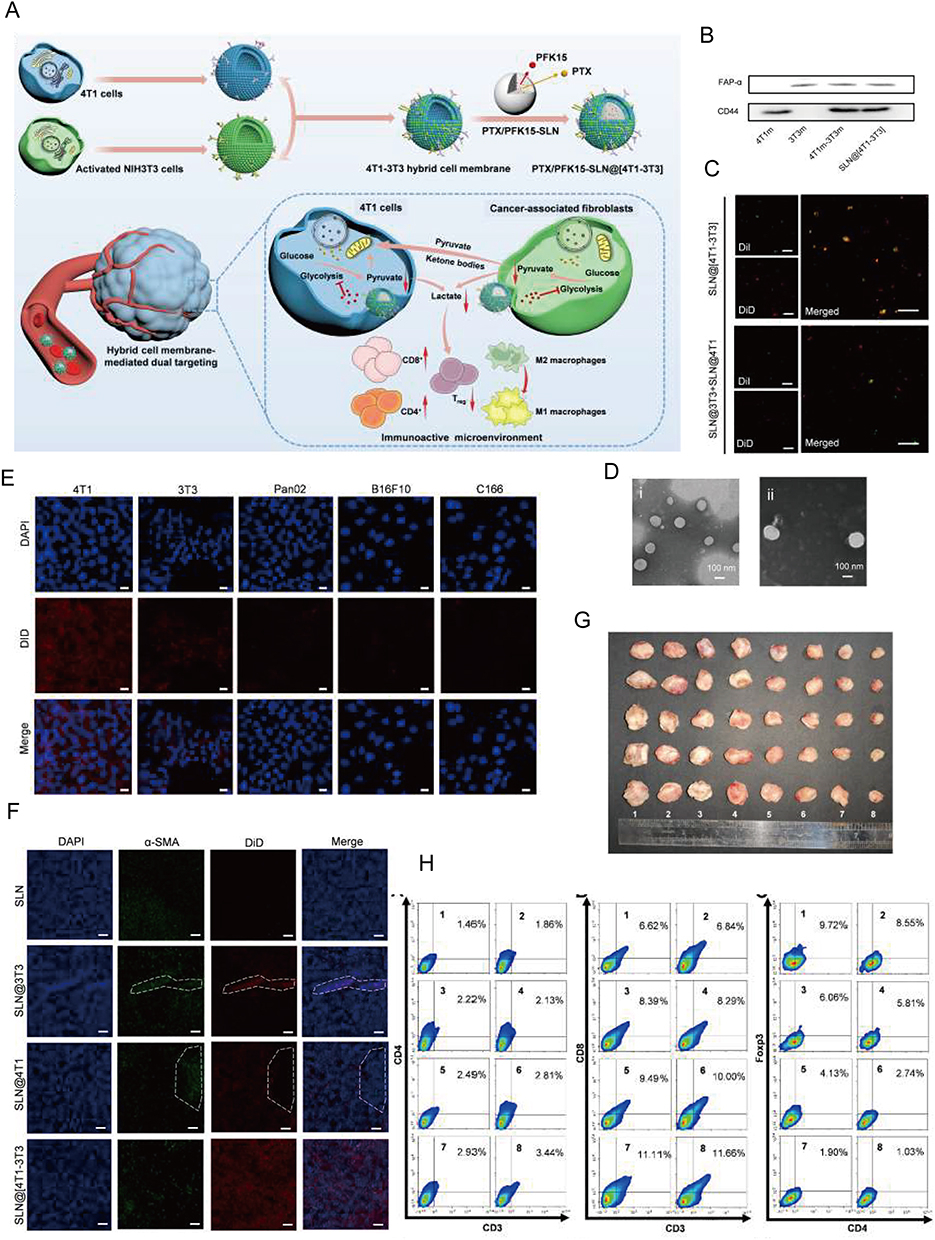 |
Figure 10 Metabolic Reprogramming by Dual-targeting Biomimetic NPs for Enhanced Tumor Chemo-Immunotherapy. (A) Scheme of PTX/PFK15-SLN@[4T1-3T3] NPs. (B) Western blot assay of 4T1 membrane (4T1m), 3T3 membrane (3T3m), 4T1-3T3 membrane (4T1m-3T3m), and SLN@[4T1-3T3] NPs for 4T1 membrane marker CD44and 3T3 membrane marker FAP-α. (C) CLSM images of the SLN@[4T1-3T3] NPs and a physical mixture of SLN@4T1 NPs and SLN@3T3 NPs. (D) Representative TEM images of PTX/PFK15-SLN and PTX/PFK15-SLN@[4T1-3T3] NPs. (E) CLSM images of 4T1, 3T3, B16F10, Pan02 and C166 cells stained with DAPI and cultured with DiD-labeled SLN@[4T1-3T3] NPs. (F) Representative CLSM images in tumor sites. The dotted line inside indicated CAFs. (G) Antitumor effect evaluation in vivo. (H) Representative flow cytometric images of the tumor CD4+ T cells, CD8+ T cells and Treg cells (gated by CD25). Reprinted from Acta Biomater, volume 148, Zang S, Huang K, Li J, et al. Metabolic reprogramming by dual-targeting biomimetic nanoparticles for enhanced tumor chemo-immunotherapy. 181–193, Copyright 2022, with permission from Elsevier.215 |
Strategies of NPs Construction for Targeting CAFs
CAF Depletion
Modifying the NPs surface with molecules specific for CAF surface markers represents the most common method of CAF targeting. For instance, targeting FAP, a surface marker expressed by 90% of CAFs, to deliver drug payloads to CAFs has been shown to significantly inhibit tumor growth.216 In a recent study, Shin, H designed antigenic peptide epitopes derived from the FAP protein and conjugated them to lipid NPs containing CpG adjuvants which is FAPPEP-SLNPs and it can be acted as a nano-vaccine, effectively activating T cell immunity and eliminating FAP-positive CAFs.217 And in the research of Guo et al, they synthesized dipeptide Z-glycine-proline to target FAP to eliminate CAFs barriers.218 Additionally, TNC, a component of the ECM expressed by CAFs,144,192 has been targeted using a stable peptide called FH.143 Some studies have also proposed targeting α-SMA. Depletion of α-SMA+ CAFs in an α-SMA-thymidine kinase hybrid transgenic mouse model resulted in the inhibition of tumor angiogenesis but also increased hypoxia, subsequently inducing EMT and stemness. Furthermore, targeting α-SMA has been shown to increase Treg infiltration into the tumor and exacerbate immunosuppression in the TME.219,220 Although the direct use of α-SMA-targeting immunotherapeutic drugs has not achieved the desired results, their value should not be dismissed in the context of NPs-based therapeutics.169,219 NPs can be used to deliver CAF-sensitive drugs rather than directly targeting α-SMA.169 Furthermore, NPs can accommodate a complex formulation of both anti-tumor and anti-CAF drugs169 to remove the side effect of merely targeting CAFs.221
The direct NP-mediated elimination of CAFs typically uses one of two strategies. The first strategy is using NPs to load non-specific cytotoxic drugs, and then conjugate the targeting molecules such as CAF surface biomarkers to NPs surface. The second strategy encompasses anti-fibrotic drugs, which exhibit a high affinity for CAFs but require modification and encapsulation within NPs due to their pharmacokinetic limitations. In some solid tumors with high infiltration of CAFs, the EPR effect will decrease. Depletion of CAFs can “loosen” the TME to assist nanoparticles enter the deep tumor tissue. Both mentioned strategies can improve their effect through the enhanced EPR effect provided by CAF depletion.
In the first strategy, the CAF-targeting NPs can reduce fibrosis, improving the penetration of anti-tumor drugs into the tumor tissue and suppressing CAF function. Additionally, the anti-tumor drugs can themselves eliminate CAFs, leading to favorable therapeutic outcomes. Several promising candidates, which achieve both tumor inhibition and CAF depletion, have been identified; these include acriflavine (ACF),146 paclitaxel (PTX) (Figure 10),215 DTX,172 and navitoclax (Nav).183 LNC-encapsulated PTX and ACF have been shown to inhibit tumor growth in vitro.146 Moreover, FAP-guided thermosensitive liposome-coated human serum albumin-PTX NPs can induce hyperthermia in the tumor stroma to enhance the release of chemotherapy drugs to eliminate CAFs (Figure 9).184 Self-polymerizing micelles containing cyclopamine (CPA) and PTX can reshape the TME by regulating Hedgehog signaling.169,222 NPs generated using the cellax-DTX polymer (with a size of 120 nm), specifically accumulate in α-SMA+ CAFs, leading to their depletion. This strategy reduces the density of the TME, consequently increasing tissue perfusion. Furthermore, cellax-DTX NPs have been shown to deplete TAMs, thereby disrupting the lines of communication between TAMs and CAFs, subsequently inducing CAF depletion.172 Nav promotes the apoptosis of CAFs by preventing the binding of the pro-apoptotic molecules BAX and BAK to BCL-2.223,224 Although the application of naked Nav is limited by its toxicity and hydrophobic nature,225 the use of Nav encapsulated in FAP-labeled H-ferritin (HFn) cages has demonstrate promising anti-CAF effects in vivo.183 The action of such FAP-labeled HFn cages has now been validated in various studies. For instance, ZnF16Pc can be encapsulated within these cages to induce PDT.181,182 In another study exploring the nano-drug loading of Nav, a TNC-linked peptide was used as a targeting ligand, resulting in similar levels of efficacy.144 Moreover, although the co-delivery of Nav and doxorubicin (DOX) does not enhance therapeutic efficacy, it allows for a reduction in the dosage of DOX, thereby alleviating its side effects. The use of FAP- and folate-conjugated dual-responsive NPs loaded with DOX has shown excellent therapeutic efficacy by facilitating endolysosomal escape170 and removing stromal barriers via CAF elimination; the mechanism of CAF depletion may rely on the downregulation of α-SMA expression on CAFs and their subsequent inhibition. The absence of side effects may be attributed to the fact that NP-mediated anti-tumor effects may indirectly influence α-SMA expression on CAFs. In breast cancer, Her2-DOX-SPIONPs@PLGA@Au induces a photothermal effect, which reduces CAF infiltration, promotes DOX release, and enhances immunogenic cell death.161 Retinoic acid (RA), a vitamin A metabolite, has been shown to disrupt Golgi apparatus function in CAFs.226 Encapsulation of the RA-DOX complex within FnBPA5-RBC-PEG-PLGA promotes CAF targeting, inhibits the Golgi functions of CAF, and facilitates DOX penetration.187 DOX NPs loaded with sulfated hyaluronic acid and mifepristone operate via a similar mechanism to the previously mentioned NPs.227 The incorporation of triptolide (TPL), an active ingredient of Tripterygium wilfordii, in PSN38@TPL-nsa can inhibit CAFs, reversing their pro-tumor functions and increasing their sensitivity to chemotherapy by inhibiting the NF-κB pathway168 (Figure 8). PFK15, an inhibitor of glycolysis, was loaded into hybrid membrane NPs composed of tumor and fibroblast cell membranes. The resulting NPs could interfere with glucose metabolism in both cell types, thereby promoting the infiltration of CD8 T cells into the tumor (Figure 10).215 Additionally, fibroblast membrane NPs can be loaded with photosensitive materials to achieve similar effects.185 For instance, in the case of IR780/DPPC/BMS, the IR780 component induces PTT, DPPC facilitates thermosensitive release, and BMS inhibits the PD-1/PD-L1 pathway. When exposed to radiation, this system can disrupt ECM integrity and CAF function. Of note, this effect does not specifically target CAFs, but rather facilitates the capture of tumor-associated antigens and enhances the sustained release of PD-1 inhibitors via the PTT-induced degradation of the ECM (Figure 4).150,228
In the second strategy, the NPs or the encapsulated drugs themselves exhibit strong CAF-targeting and -modulating abilities. For example, 3-nm GNPs are easily absorbed by CAFs and downregulate key CAF markers (eg, N-cadherin, α-SMA, vimentin, FSP-1, and S100 family proteins) to inhibit CAF activation and motility. Additionally, GNPs can reduce the CAF-mediated secretion of HGF, IL-6, IL-8, TGF-β1, and PDGF-α, thereby disrupting the communication between CAFs and tumor cells.158 Additionally, the use of GNPs modified to contain iron oxide (GOINFs) has been validated. These NPs function as efficient nano-heaters and outperform GNPs in the near-infrared (NIR) spectral range.163 Similar to gold, iron exhibits a pronounced thermal effect and can be effectively delivered to CAFs. Following a brief exposure to an alternating magnetic field, SPIONs can induce caspase 8-dependent apoptosis in CAFs.154 Furthermore, a relatively novel, promising approach using ultra-small iron oxide NPs exploits the magnetic properties of iron to mechanically damage both tumors and CAFs.201 Other non-metallic materials with similar CAF-eliminating capabilities also exist. For instance, PLGA-derived ONP-302 can delay tumor growth and reverse the exhausted and immunosuppressive phenotypes of CD8+ T cells.166 In addition, ONP-302 induces the polarization of macrophages toward the M1 subtype, inhibits the expression of pro-tumor genes in CAFs, and promotes CAF apoptosis. The NP-mediated delivery of small interfering RNA has also emerged as an effective strategy for targeting CAFs. CAFs secrete CXCL12 to retain their immunosuppressive properties and facilitate tumor growth and migration via the activation of the CXCL12/CXCR4 pathway. Therefore, utilizing the PNP/siCXCL12/monoclonal antibody approach to effectively block CXCL12 expression in CAFs could be a promising strategy to reduce tumor growth and angiogenesis.20
Inhibiting CAF Activation
Using NPs to revert activated CAFs back to a resting state represents a feasible CAF-modulating strategy. However, the presence of cytokines, such as TGF-β1, IL-10, and IL-6, within the TME can persistently activate CAFs and significantly reduce the efficacy of CAF-targeting drugs.9 Therefore, it is crucial for NPs formulations to maintain optimal drug concentrations in the tumor and elicit long-lasting effects. Many studies have focused on targeting TGF-β1, the main inducer of fibroblast activation.12 In the recent research of Zheng et al, they found that mPEG-PLGA loaded baicalein could inhibit CAFs activation through blocking TGF-β signaling pathway.179 Moreover, in another study by Mardhian et al, the SPOIN-mediated delivery of fibroblast growth factor 2 has been shown to inhibit the pSmad2/3 pathway and activate ERK1/2 signaling, thus, effectively interfering with TGF-β1-induced CAF activation. This in turn reduced α-SMA and collagen-1 expression in CAFs and inhibited the formation of new CAFs via the differentiation of stellate cells (Figure 6).157 Moreover, the liposomal delivery of oxymatrine can restore the ECM and collagen content of the TME to normal levels, downregulate HMGB1, and inactivate stellate cells, consequently reducing the TGF-β1 concentration.143 Similarly, PEG-encapsulated SAB inhibits TGF-β1 secretion, which may increase the expression of tumor suppressor factors such as CXCL9 and CXCL10, recruit CD4+ T cells, CD8+ T cells, and M1 macrophages, while reducing the infiltration of MDSCs, Tregs, and M2 macrophages.148 CAFs may be the cause of limited therapeutic effects of CAR-T in solid tumors. So, Tang et al utilized amphipathic hydroxyethyl starch-polycaprolactone to load TGF-β inhibitor (LY2157299) and photosensitizer indocyanine green. This co-loaded NPs can enhance the CAR-T effects and maybe a promising application.180
Moreover, anti-tumor drugs are often used to reduce CAF activation. Pirfenidone, a TGF-β1 antagonist, is commonly used in the treatment of idiopathic pulmonary fibrosis. In a recent study, Zuo et al developed pirfenidone-containing HPBC@TRP/NPs, which allow pirfenidone to effectively reach the TME without degradation in the peripheral tissues and rapidly exert its therapeutic effects. Simultaneously, another core drug, resveratrol, enters the cells and is released through lysosomes, where it activates the mtROS-PYK2 pathway to induce mitochondria-driven apoptosis (Figure 7).165 In the study of Li et al, they used glycyrrhetinic acid (GA) to target hepatocellular carcinoma and HA to target hepatic stellate cells, and then constructed GA&HA-modified liposomes to load aprepitant (APR) and curcumin (CUR), which is CUR-APR/HA&GA-LPs. In vivo and in vitro, CUR-APR/HA&GA-LPs can prevent hepatic stellate cells transforming into CAFs.151 In another study from Wu et al, they utilized the same construction methods in Li et al study and showed that the berberine also presented the same effects of targeting CAFs as APR.152
NPs can improve drug delivery into cells. This approach not only enables the targeting of CAFs but also ensures optimal drug doses are delivered to the tumor site.198 GNPs serve as a prime example in this regard. GNPs (with a size of 20 nm) not only suppress the expression of CAF markers (specifically α-SMA and fibronectin), diminishing their responsiveness to activators, but also disrupt intercellular communication by interfering with TGF-β1, TSP1, PDGF, and uPA release from tumor cells.159 Additionally, GNPs promote the expression of FASN, SREBP2, and FABP3 in their target cell, thereby increasing the cellular lipid content and causing the cell to adopt a resting phenotype.159 Another noteworthy example involves the use of an LPD-coated secretable TNF-related apoptosis-inducing ligand (sTRAIL). This construct exhibits stable expression and release within CAFs, inducing apoptosis in adjacent cancer nests while concurrently inhibiting CAF activation.149
Disrupting CAF Function
This type of NPs mainly targets CAF indirectly, causing non-specific cellular damage, which affects CAF functions and products. To date, this approach has proved more effective than others in reversing the immunosuppressive state of the TME. For instance, Frax (with a diameter of 145 nm) is absorbed by both tumor cells and CAFs. Once in the cells, Frax reduces the expression of CUGBP1, thereby interfering with the TGF-β1 and IFN-γ pathways and attenuating tumor fibrosis.147 Additionally, Frax reduces the amounts of immunosuppressive cells such as MDSCs in the TME and enhances the infiltration of CD8 T cells. Moreover, the action of Frax can be further boosted by combining it with tumor vaccines. The presence of surface molecules, such as CXCR4 and CD44, on PLGA NPs coated with tumor cell membranes inhibits the migration of tumor cells toward CAFs and promotes the infiltration of T cells, particularly IFNγ+CD8+ T cells into the tumor.229 By utilizing hyperthermia, FAP-guided, thermosensitive, liposome-coated human serum albumin -PTX NPs selectively target the tumor stroma and eliminate CAFs, facilitating the release of chemotherapy drugs (Figure 9).184 In the study of Panagi et al, they developed tranilast-loaded micelles, which can be uptake by CAFs thus inhibiting the expression of TGF-β. In the breast cancer model constructed by 4T1 and EO771, tranilast-loaded micelles can reduce the content of HA and collagen, which induce CD8 T cells penetrate.177 Another study reported the development of a NIR-responsive, membrane-coated PD-1/PD-L1 inhibitor; its primary function was to enhance the capture of tumor-associated antigens by DCs and activate T cells, while its secondary function was to eliminate CAFs via PTT.186 Polymer-coated, sunitinib-containing NPs enhance the infiltration of T cells into the tumor while reducing the proportions of Tregs and MDSCs in the TME. They also remodel the TME by exerting effects on CAFs, collagen, and blood vessels, as well as altering the cytokine composition of the TME by facilitating the transition from a T helper (Th)2 to a Th1 immune response.176 In the recent study from Li et al, they chose losartan to reduce the activity of CAFs. And they used nanoliposome to co-load resiquimod, DOX and losartan to construct the multifunctional NPs to remodel TME.153
CAFs contribute to tumor development and drug resistance by activating the Hedgehog signaling pathway in a paracrine manner. In a study by Wang et al the use of SN38-coated NPs containing the Hedgehog pathway inhibitor GDC-0449 effectively suppressed the expression of key drug resistance molecules (eg, GLI-1 and UGT1A) while reducing collagen content within tumor tissues.173 Additionally, Kovacs et al have discovered that gold-silver-gold composite NPs possess potent anti-metastatic properties and can inhibit the pro-tumor effects of CAFs, ultimately reducing the migration and invasiveness of tumor cells.155 In colorectal cancer, CAFs can secrete hepatocyte growth factor to promote tumor progression. Based on this, Shen et al designed siRNA to interfere the expression of hepatocyte growth factor and used PLGA to deliver the siRNA. When combined with chemotherapy, the novel PLGA-siRNA NPs can overcome the chemotherapy-induced hepatocyte growth factor upregulation and then inhibit the CAFs activation. This loop showed great potential of targeting CAFs to alleviate chemotherapy resistance.178 There also exist an approach which is inserting molecules for disrupting CAFs function on the surface of NPs rather using NPs to load them. Liu et al inserted integrin β1 inhibiting peptide FINIII4 on their PLGA loaded DOX NPs. This made the NPs can respond to FAP and induce the combination of FINIII4 and integrin β1 to disrupt the functions of CAFs to decrease the density of tumor.230
Conclusions and Future Perspectives
Although we had gained a considerable understanding of CAFs, many questions remain unanswered. The incomplete understanding of CAF biology may be a contributing factor to the occurrence of many unexpected outcomes in clinical studies targeting CAFs.7 Consequently, many studies investigating the use of NPs as CAF-targeted therapies are still someway away from being adopted into the clinic.18 Currently, there is no explicit consensus on the tumor models used to study CAFs. The construction of these models faces challenges such as the selection of an appropriate cellular source for CAF generation (the most commonly used cells are embryonic fibroblasts231 and fibroblasts from original tissue sources232), the method of model establishment (eg, xenograft models or spontaneous tumor models),9 and the most physiologically relevant tumor cell to CAF ratios (existing studies suggest tumor cells to fibroblast ratios ranging from 1:1 to 1:10).9,231–233 The cancer type is also an important consideration in studies of NPs-mediated CAF targeting. It is therefore crucial to choose research models that are optimized for the study of a specific cancer type. During the process of tumor development CAFs exhibit considerable heterogeneity, particularly in terms of their cellular origins and surface marker expression.9 Thus, the optimal approach involves inducing tumor formation in situ, as it better aligns with the requirements of studying CAFs and replicates the natural occurrence of tumors. Furthermore, there is room for improvement in the way the cellular CAF sources are handled during the study of cell-membrane-encapsulated NPs. For instance, in the research of Li et al and Zang et al, the source of CAFs current is TGF-β-treated NIH/3T3 cells, which is evidently inaccurate.185,215 The primary purpose of cell membrane encapsulation is to mimic the target cell, thereby increasing the targeting efficacy of the therapeutic approach. However, using highly artificial cell membranes may impact the therapeutic efficacy of NPs.
Currently, the main CAF-targeting NPs are constructed using lipids, metals, and various polymers. These materials have been extensively studied and have shown considerable success in CAF-targeted therapy studies. However, in recent years, emerging materials such as hydrogels,234 nanoenzymes,235 and metal or carbon rods have gained increasing attention.236 These nanomaterials offer certain advantages over existing ones and have shown promising results in targeting tumor cells. Future research will explore whether these more optimized nanomaterials meet the diverse therapeutic requirements. Increasingly, specific targeting ligands (eg, FAP20,154,164), nanomaterials, and drugs are being combined to form complexes. However, this singular approach resembles that of drug-modified nanomaterials and does not fully exploit the unique properties of nanomaterials. Consequently, these nanoscale particle drugs eventually enter systemic circulation and undergo hepatobiliary metabolism, highlighting the importance of addressing how NPs can be engineered to evade rapid clearance from the body.165 Recently, Zuo et al developed the HPBC@TRP/NPs system, which effectively protects the core drug-loaded NPs, thereby achieving improved therapeutic efficacy.165
Current strategies for NPs construction primarily focus on eliminating CAFs. However, previous basic and clinical trials have shown that normalizing the tumor stroma and TME may limit tumor occurrence and progression more effectively than direct CAF elimination.112,117,118 Therefore, the future design of NPs targeting CAFs should place more emphasis on inhibiting CAF activation and interfering with CAF functions.15 Consequently, compounds such as vitamin A/D and CXCL12-CXCR4/JAK pathway inhibitors are promising CAF-modulating drug candidates, as these drugs have already shown considerable efficacy in targeting CAFs in the clinical trials setting.124,125,131,132 Additionally, strategies that simultaneously target CAFs and kill tumor cells have been proven more effective than drugs that target each cell type individually. Therefore, in the future development of NPs targeting CAF, the principle should be centered around killing tumor cells, while using CAF- targeting as complementary approach. Targeting CAFs inhibits tumor progression, while killing tumor cells promotes the normalization of the tumor stroma and the TME; thus, targeting both cell types simultaneously represents a synergistic approach. Currently, the main approach for targeting CAFs is through the surface modification of NPs with FAP- or TNC-binding molecules.143,157 However, FAP is not a universal CAF marker; moreover, CAF markers can vary at different stages of tumor development. Therefore, a better approach would be to use NPs encapsulated in a CAF membrane. Additionally, the therapeutic benefits of multifunctional NPs that eliminate tumor cells and target CAFs would be more significant than those derived from combination therapies. The development of a more specialized repertoire of CAF-targeting NPs will undoubtedly lead to superior therapeutic outcomes.197
In summary, the NPs-mediated targeting of CAFs has achieved remarkable outcomes, indicating that their application will likely form an essential component of tumor immunotherapy. However, before CAF-targeting NPs enter clinical studies, important aspects, including the toxicity and metabolic properties of the nanomaterials used, need to be considered.237 In the future, the development of CAF-targeting drugs will be driven by advancements in cutting-edge research techniques such as single-cell sequencing, bispecific antibodies, flow cytometry, and high-resolution imaging. Concurrently, nanomaterials used for the construction of CAF-targeting NPs will continue be optimized in accordance with the therapeutic requirements. This highlights the need for more extensive collaboration between experts in the fields of materials science and biomedicine.
Abbreviation
CAFs, cancer-associated fibroblasts; rCAFs, cancer-restraining CAF; myCAF, myofibroblastic CAFs; iCAF, inflammatory CAFs; apCAF, antigen-presenting CAFs; meCAF, metabolic CAFs; vCAF, vascular CAFs; EMT, epithelial-mesenchymal transition; TME, tumor microenvironment; TIME, tumor immune microenvironment; ECM, extracellular matrix; NPs, nanoparticles; NK, natural killer cells; DCs, dendritic cells; MDSCs, myeloid-derived suppressor cells; Tregs, regulatory T cells; VEGF, vascular endothelial growth factor; α-SMA, α-smooth muscle actin; MSCs, mesenchymal stem cells; LIF, Leukemia Inhibitory Factor; ICPs, immune checkpoint proteins; ICI, immune checkpoint inhibitor; PBMCs, peripheral blood mononuclear cells; TAMs, tumor-associated macrophages; BM, bone marrow; NSCLC, non-small cell lung cancer; IL, interleukin; ATAR, all-trans retinoic acid; PTT, photothermal therapy; PDT, photodynamic therapy; PEG, polyethylene glycol; FAP, fibroblast activation protein-α; TNC, tenascin C; SAB, salvianolic acid B; Frax, fraxinellone; LPD, lipid-coated protamine DNA; GNP, Gold nanoparticle; DTX, docetaxel; SPOIN, superparamagnetic iron oxide nanoparticle; PLGA, Poly(lactic-co-glycolic acid); EPR, enhanced permeability and retention effect; PSN38: PEG5K-P(MMESSN38)5K; PAMAM, the positive surface charge of cationic poly(amidoamine); RBC, red blood cells; ACF, acriflavine; PTX, paclitaxel; Nav, navitoclax; CPA, cyclopamine; DOX, doxorubicin; TPL, triptolide; NIR, near-infrared; sTRAIL, secretable TNF-related apoptosis-inducing ligand; DPPC, dipalmitoylphosphatidylcholine; APR, aprepitant; CUR, curcumin.
Data Sharing Statement
No data was used in the research and all related researches were referenced.
Acknowledgments
Our research was funded by the National Natural Science Foundation of China (No.82170779, No.82270804), 2019 Wuhan Yellow Crane Talent Program (Outstanding Young Talents), and the Tongji Hospital (HUST) Foundation for Excellent Young Scientist (No.2020YQ15).
Qiu Huang and Yue Ge are joint first authors for and contributed equally to this study. Zhiqiang Chen and Kun Tang are shared corresponding authors for this study.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Quail DF, Joyce JA. Microenvironmental regulation of tumor progression and metastasis. Nat Med. 2013;19(11):1423–1437. doi:10.1038/nm.3394
2. Hinshaw DC, Shevde LA. The tumor microenvironment innately modulates cancer progression. Cancer Res. 2019;79(18):4557–4566. doi:10.1158/0008-5472.CAN-18-3962
3. Chen F, Zhuang X, Lin L, et al. New horizons in tumor microenvironment biology: challenges and opportunities. BMC Med. 2015;13:45. doi:10.1186/s12916-015-0278-7
4. Baldominos P, Barbera-Mourelle A, Barreiro O, et al. Quiescent cancer cells resist T cell attack by forming an immunosuppressive niche. Cell. 2022;185(10):1694–1708.e19. doi:10.1016/j.cell.2022.03.033
5. Erin N, Grahovac J, Brozovic A, et al. Tumor microenvironment and epithelial mesenchymal transition as targets to overcome tumor multidrug resistance. Drug Resist Updat. 2020;53:100715. doi:10.1016/j.drup.2020.100715
6. Kim HJ, Ji YR, Lee YM. Crosstalk between angiogenesis and immune regulation in the tumor microenvironment. Arch Pharm Res. 2022;45(6):401–416. doi:10.1007/s12272-022-01389-z
7. Chen X, Song E. Turning foes to friends: targeting cancer-associated fibroblasts. Nat Rev Drug Discov. 2019;18(2):99–115. doi:10.1038/s41573-018-0004-1
8. Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. 2016;16(9):582–598. doi:10.1038/nrc.2016.73
9. Rimal R, Desai P, Daware R, et al. Cancer-associated fibroblasts: origin, function, imaging, and therapeutic targeting. Adv Drug Deliv Rev. 2022;189:114504. doi:10.1016/j.addr.2022.114504
10. Bainbridge P. Wound healing and the role of fibroblasts. J Wound Care. 2013;22(8):407–412. doi:10.12968/jowc.2013.22.8.407
11. Dvorak HF, Underhill LH, Dvorak HF. Tumors: wounds that do not heal. Similarities between tumor stroma generation and wound healing. N Engl J Med. 1986;315(26):1650–1659. doi:10.1056/NEJM198612253152606
12. Peng D, Fu M, Wang M, et al. Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol Cancer. 2022;21(1):104. doi:10.1186/s12943-022-01569-x
13. Mizutani Y, Kobayashi H, Iida T, et al. Meflin-positive cancer-associated fibroblasts inhibit pancreatic carcinogenesis. Cancer Res. 2019;79(20):5367–5381. doi:10.1158/0008-5472.CAN-19-0454
14. Biffi G, Tuveson DA. Diversity and biology of cancer-associated fibroblasts. Physiol Rev. 2021;101(1):147–176. doi:10.1152/physrev.00048.2019
15. Chen Y, McAndrews KM, Kalluri R. Clinical and therapeutic relevance of cancer-associated fibroblasts. Nat Rev Clin Oncol. 2021;18(12):792–804. doi:10.1038/s41571-021-00546-5
16. Bockamp E, Rosigkeit S, Siegl D, et al. Nano-enhanced cancer immunotherapy: immunology encounters nanotechnology. Cells. 2020;9(9):2102. doi:10.3390/cells9092102
17. Pérez-Herrero E, Fernández-Medarde A. Advanced targeted therapies in cancer: drug nanocarriers, the future of chemotherapy. Eur J Pharm Biopharm. 2015;93:52–79. doi:10.1016/j.ejpb.2015.03.018
18. Liu M, Song W, Huang L. Drug delivery systems targeting tumor-associated fibroblasts for cancer immunotherapy. Cancer Lett. 2019;448:31–39. doi:10.1016/j.canlet.2019.01.032
19. Farokhzad OC, Langer R. Impact of nanotechnology on drug delivery. ACS Nano. 2009;3(1):16–20. doi:10.1021/nn900002m
20. Lang J, Zhao X, Qi Y, et al. Reshaping prostate tumor microenvironment to suppress metastasis via cancer-associated fibroblast inactivation with peptide-assembly-based nanosystem. ACS Nano. 2019;13(11):12357–12371. doi:10.1021/acsnano.9b04857
21. Zhen Z, Tang W, Wang M, et al. Protein nanocage mediated fibroblast-activation protein targeted photoimmunotherapy to enhance cytotoxic T cell infiltration and tumor control. Nano Lett. 2017;17(2):862–869. doi:10.1021/acs.nanolett.6b04150
22. Pitt JM, Marabelle A, Eggermont A, et al. Targeting the tumor microenvironment: removing obstruction to anticancer immune responses and immunotherapy. Ann Oncol. 2016;27(8):1482–1492. doi:10.1093/annonc/mdw168
23. Hanahan D, Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. 2012;21(3):309–322. doi:10.1016/j.ccr.2012.02.022
24. Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013;14(10):1014–1022. doi:10.1038/ni.2703
25. Coussens LM, Zitvogel L, Palucka AK. Neutralizing tumor-promoting chronic inflammation: a magic bullet? Science. 2013;339(6117):286–291. doi:10.1126/science.1232227
26. Zhang Y, Zhang Z. The history and advances in cancer immunotherapy: understanding the characteristics of tumor-infiltrating immune cells and their therapeutic implications. Cell Mol Immunol. 2020;17(8):807–821. doi:10.1038/s41423-020-0488-6
27. Russell SJ, Peng KW, Bell JC. Oncolytic virotherapy. Nat Biotechnol. 2012;30(7):658–670. doi:10.1038/nbt.2287
28. van der Bruggen P, Traversari C, Chomez P, et al. A gene encoding an antigen recognized by cytolytic T lymphocytes on a human melanoma. Science. 1991;254(5038):1643–1647. doi:10.1126/science.1840703
29. Berraondo P, Sanmamed MF, Ochoa MC, et al. Cytokines in clinical cancer immunotherapy. Br J Cancer. 2019;120(1):6–15. doi:10.1038/s41416-018-0328-y
30. Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer. Science. 2015;348(6230):62–68. doi:10.1126/science.aaa4967
31. Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12(4):252–264. doi:10.1038/nrc3239
32. Peng S, Xiao F, Chen M, et al. Tumor-microenvironment-responsive nanomedicine for enhanced cancer immunotherapy. Adv Sci (Weinh). 2022;9(1):e2103836. doi:10.1002/advs.202103836
33. D’Arcangelo E, Wu NC, Cadavid JL, et al. The life cycle of cancer-associated fibroblasts within the tumour stroma and its importance in disease outcome. Br J Cancer. 2020;122(7):931–942. doi:10.1038/s41416-019-0705-1
34. Räsänen K, Vaheri A. Activation of fibroblasts in cancer stroma. Exp Cell Res. 2010;316(17):2713–2722. doi:10.1016/j.yexcr.2010.04.032
35. Piersma B, Hayward MK, Weaver VM. Fibrosis and cancer: a strained relationship. Biochim Biophys Acta Rev Cancer. 2020;1873(2):188356. doi:10.1016/j.bbcan.2020.188356
36. Mao X, Xu J, Wang W, et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021;20(1):131. doi:10.1186/s12943-021-01428-1
37. Wan X, Guan S, Hou Y, et al. FOSL2 promotes VEGF-independent angiogenesis by transcriptionnally activating Wnt5a in breast cancer-associated fibroblasts. Theranostics. 2021;11(10):4975–4991. doi:10.7150/thno.55074
38. Li Z, Sun C, Qin Z. Metabolic reprogramming of cancer-associated fibroblasts and its effect on cancer cell reprogramming. Theranostics. 2021;11(17):8322–8336. doi:10.7150/thno.62378
39. Gong J, Lin Y, Zhang H, et al. Reprogramming of lipid metabolism in cancer-associated fibroblasts potentiates migration of colorectal cancer cells. Cell Death Dis. 2020;11(4):267. doi:10.1038/s41419-020-2434-z
40. Xi L, Peng M, Liu S, et al. Hypoxia-stimulated ATM activation regulates autophagy-associated exosome release from cancer-associated fibroblasts to promote cancer cell invasion. J Extracell Vesicles. 2021;10(11):e12146. doi:10.1002/jev2.12146
41. Li C, Teixeira AF, Zhu H-J, et al. Cancer associated-fibroblast-derived exosomes in cancer progression. Mol Cancer. 2021;20(1):154. doi:10.1186/s12943-021-01463-y
42. Nicolas AM, Pesic M, Engel E, et al. Inflammatory fibroblasts mediate resistance to neoadjuvant therapy in rectal cancer. Cancer Cell. 2022;40(2):168–184.e13. doi:10.1016/j.ccell.2022.01.004
43. Kieffer Y, Hocine HR, Gentric G, et al. Single-cell analysis reveals fibroblast clusters linked to immunotherapy resistance in cancer. Cancer Discov. 2020;10(9):1330–1351. doi:10.1158/2159-8290.CD-19-1384
44. Zhao P, Tian Y, Lu Y, et al. Biomimetic calcium carbonate nanoparticles delivered IL-12 mRNA for targeted glioblastoma sono-immunotherapy by ultrasound-induced necroptosis. J Nanobiotechnology. 2022;20(1):525. doi:10.1186/s12951-022-01731-z
45. Costa A, Kieffer Y, Scholer-Dahirel A, et al. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell. 2018;33(3):463–479.e10. doi:10.1016/j.ccell.2018.01.011
46. Elyada E, Bolisetty M, Laise P, et al. Cross-species single-cell analysis of pancreatic ductal adenocarcinoma reveals antigen-presenting cancer-associated fibroblasts. Cancer Discov. 2019;9(8):1102–1123. doi:10.1158/2159-8290.CD-19-0094
47. Öhlund D, Handly-Santana A, Biffi G, et al. Distinct populations of inflammatory fibroblasts and myofibroblasts in pancreatic cancer. J Exp Med. 2017;214(3):579–596. doi:10.1084/jem.20162024
48. Omary MB, Lugea A, Lowe AW, et al. The pancreatic stellate cell: a star on the rise in pancreatic diseases. J Clin Invest. 2007;117(1):50–59. doi:10.1172/JCI30082
49. Yin C, Evason KJ, Asahina K, et al. Hepatic stellate cells in liver development, regeneration, and cancer. J Clin Invest. 2013;123(5):1902–1910. doi:10.1172/JCI66369
50. Nurmik M, Ullmann P, Rodriguez F, et al. In search of definitions: cancer-associated fibroblasts and their markers. Int J Cancer. 2020;146(4):895–905. doi:10.1002/ijc.32193
51. Whiteside TL. Exosome and mesenchymal stem cell cross-talk in the tumor microenvironment. Semin Immunol. 2018;35:69–79. doi:10.1016/j.smim.2017.12.003
52. Zhu Q, Zhang X, Zhang L, et al. The IL-6-STAT3 axis mediates a reciprocal crosstalk between cancer-derived mesenchymal stem cells and neutrophils to synergistically prompt gastric cancer progression. Cell Death Dis. 2014;5(6):e1295. doi:10.1038/cddis.2014.263
53. Zhang Q, Chai S, Wang W, et al. Macrophages activate mesenchymal stem cells to acquire cancer-associated fibroblast-like features resulting in gastric epithelial cell lesions and malignant transformation in vitro. Oncol Lett. 2019;17(1):747–756. doi:10.3892/ol.2018.9703
54. Shi Y, Du L, Lin L, et al. Tumour-associated mesenchymal stem/stromal cells: emerging therapeutic targets. Nat Rev Drug Discov. 2017;16(1):35–52. doi:10.1038/nrd.2016.193
55. Jotzu C, Alt E, Welte G, et al. Adipose tissue-derived stem cells differentiate into carcinoma-associated fibroblast-like cells under the influence of tumor-derived factors. Anal Cell Pathol (Amst). 2010;33(2):61–79. doi:10.1155/2010/695162
56. Verginadis II, Avgousti H, Monslow J, et al. A stromal integrated stress response activates perivascular cancer-associated fibroblasts to drive angiogenesis and tumour progression. Nat Cell Biol. 2022;24(6):940–953. doi:10.1038/s41556-022-00918-8
57. Bhome R, Emaduddin M, James V, et al. Epithelial to mesenchymal transition influences fibroblast phenotype in colorectal cancer by altering miR-200 levels in extracellular vesicles. J Extracell Vesicles. 2022;11(5):e12226. doi:10.1002/jev2.12226
58. Mhaidly R, Mechta-Grigoriou F. Fibroblast heterogeneity in tumor micro-environment: role in immunosuppression and new therapies. Semin Immunol. 2020;48:101417. doi:10.1016/j.smim.2020.101417
59. Lavie D, Ben-Shmuel A, Erez N, et al. Cancer-associated fibroblasts in the single-cell era. Nat Cancer. 2022;3(7):793–807. doi:10.1038/s43018-022-00411-z
60. Bartoschek M, Oskolkov N, Bocci M, et al. Spatially and functionally distinct subclasses of breast cancer-associated fibroblasts revealed by single cell RNA sequencing. Nat Commun. 2018;9(1):5150. doi:10.1038/s41467-018-07582-3
61. Liu T, Han C, Wang S, et al. Cancer-associated fibroblasts: an emerging target of anti-cancer immunotherapy. J Hematol Oncol. 2019;12(1):86. doi:10.1186/s13045-019-0770-1
62. Lakins MA, Ghorani E, Munir H, et al. Cancer-associated fibroblasts induce antigen-specific deletion of CD8 +T Cells to protect tumour cells. Nat Commun. 2018;9(1):948. doi:10.1038/s41467-018-03347-0
63. Darby IA, Zakuan N, Billet F, et al. The myofibroblast, a key cell in normal and pathological tissue repair. Cell Mol Life Sci. 2016;73(6):1145–1157. doi:10.1007/s00018-015-2110-0
64. Miyazaki Y, Oda T, Inagaki Y, et al. Adipose-derived mesenchymal stem cells differentiate into heterogeneous cancer-associated fibroblasts in a stroma-rich xenograft model. Sci Rep. 2021;11(1):4690. doi:10.1038/s41598-021-84058-3
65. Wang Y, Liang Y, Xu H, et al. Single-cell analysis of pancreatic ductal adenocarcinoma identifies a novel fibroblast subtype associated with poor prognosis but better immunotherapy response. Cell Discov. 2021;7(1):36. doi:10.1038/s41421-021-00271-4
66. Peng J, Sun B-F, Chen C-Y, et al. Single-cell RNA-seq highlights intra-tumoral heterogeneity and malignant progression in pancreatic ductal adenocarcinoma. Cell Res. 2019;29(9):725–738. doi:10.1038/s41422-019-0195-y
67. Friedman G, Levi-Galibov O, David E, et al. Cancer-associated fibroblast compositions change with breast cancer progression linking the ratio of S100A4(+) and PDPN(+) CAFs to clinical outcome. Nat Cancer. 2020;1(7):692–708. doi:10.1038/s43018-020-0082-y
68. Kim N, Kim HK, Lee K, et al. Single-cell RNA sequencing demonstrates the molecular and cellular reprogramming of metastatic lung adenocarcinoma. Nat Commun. 2020;11(1):2285. doi:10.1038/s41467-020-16164-1
69. Lambrechts D, Wauters E, Boeckx B, et al. Phenotype molding of stromal cells in the lung tumor microenvironment. Nat Med. 2018;24(8):1277–1289. doi:10.1038/s41591-018-0096-5
70. Monteran L, Erez N. The dark side of fibroblasts: cancer-associated fibroblasts as mediators of immunosuppression in the tumor microenvironment. Front Immunol. 2019;10:1835. doi:10.3389/fimmu.2019.01835
71. Erez N, Truitt M, Olson P, et al. Cancer-associated fibroblasts are activated in incipient neoplasia to orchestrate tumor-promoting inflammation in an NF-kappaB-dependent manner. Cancer Cell. 2010;17(2):135–147. doi:10.1016/j.ccr.2009.12.041
72. Biffi G, Oni TE, Spielman B, et al. IL1-Induced JAK/STAT signaling is antagonized by TGFβ to Shape CAF heterogeneity in pancreatic ductal adenocarcinoma. Cancer Discov. 2019;9(2):282–301. doi:10.1158/2159-8290.CD-18-0710
73. Kwon OJ, Zhang Y, Li Y, et al. Functional heterogeneity of mouse prostate stromal cells revealed by Single-Cell RNA-Seq. iScience. 2019;13:328–338. doi:10.1016/j.isci.2019.02.032
74. Steele NG, Biffi G, Kemp SB, et al. Inhibition of hedgehog signaling alters fibroblast composition in pancreatic cancer. Clin Cancer Res. 2021;27(7):2023–2037. doi:10.1158/1078-0432.CCR-20-3715
75. Toullec A, Gerald D, Despouy G, et al. Oxidative stress promotes myofibroblast differentiation and tumour spreading. EMBO Mol Med. 2010;2(6):211–230. doi:10.1002/emmm.201000073
76. Zhang X, He C, He X, et al. HIF-1 inhibitor-based one-stone-two-birds strategy for enhanced cancer chemodynamic-immunotherapy. J Control Release. 2023;356:649–662. doi:10.1016/j.jconrel.2023.03.026
77. Zhang X, He C, Xiang G. Engineering nanomedicines to inhibit hypoxia-inducible Factor-1 for cancer therapy. Cancer Lett. 2022;530:110–127. doi:10.1016/j.canlet.2022.01.012
78. Kitamura F, Semba T, Yasuda-Yoshihara N, et al. Cancer-associated fibroblasts reuse cancer-derived lactate to maintain a fibrotic and immunosuppressive microenvironment in pancreatic cancer. JCI Insight. 2023;8(20). doi:10.1172/jci.insight.163022
79. Orimo A, Gupta PB, Sgroi DC, et al. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell. 2005;121(3):335–348. doi:10.1016/j.cell.2005.02.034
80. Olumi AF, Grossfeld GD, Hayward SW, et al. Carcinoma-associated fibroblasts direct tumor progression of initiated human prostatic epithelium. Cancer Res. 1999;59(19):5002–5011. doi:10.1186/bcr138
81. Unterleuthner D, Neuhold P, Schwarz K, et al. Cancer-associated fibroblast-derived WNT2 increases tumor angiogenesis in colon cancer. Angiogenesis. 2020;23(2):159–177. doi:10.1007/s10456-019-09688-8
82. Zhang F, Cui J-Y, Gao H-F, et al. Cancer-associated fibroblasts induce epithelial-mesenchymal transition and cisplatin resistance in ovarian cancer via CXCL12/CXCR4 axis. Future Oncol. 2020;16(32):2619–2633. doi:10.2217/fon-2020-0095
83. Glentis A, Oertle P, Mariani P, et al. Cancer-associated fibroblasts induce metalloprotease-independent cancer cell invasion of the basement membrane. Nat Commun. 2017;8(1):924. doi:10.1038/s41467-017-00985-8
84. Shintani Y, Fujiwara A, Kimura T, et al. IL-6 secreted from cancer-associated fibroblasts mediates chemoresistance in NSCLC by increasing epithelial-mesenchymal transition signaling. J Thorac Oncol. 2016;11(9):512–536. doi:10.1016/j.jtho.2016.05.025
85. Long X, Xiong W, Zeng X, et al. Cancer-associated fibroblasts promote cisplatin resistance in bladder cancer cells by increasing IGF-1/ERβ/Bcl-2 signalling. Cell Death Dis. 2019;10(5):375. doi:10.1038/s41419-019-1581-6
86. Wang W, Kryczek I, Dostál L, et al. Effector T cells abrogate stroma-mediated chemoresistance in ovarian cancer. Cell. 2016;165(5):1092–1105. doi:10.1016/j.cell.2016.04.009
87. Yang M, McKay D, Pollard JW, et al. Diverse Functions of Macrophages in Different Tumor Microenvironments. Cancer Res. 2018;78(19):5492–5503. doi:10.1158/0008-5472.CAN-18-1367
88. Zhou X, Franklin RA, Adler M, et al. Circuit design features of a stable two-cell system. Cell. 2018;172(4):744–757.e17. doi:10.1016/j.cell.2018.01.015
89. Gok Yavuz B, Gunaydin G, Gedik ME, et al. Cancer associated fibroblasts sculpt tumour microenvironment by recruiting monocytes and inducing immunosuppressive PD-1+ TAMs. Sci Rep. 2019;9(1):3172. doi:10.1038/s41598-019-39553-z
90. Buechler MB, Fu W, Turley SJ. Fibroblast-macrophage reciprocal interactions in health, fibrosis, and cancer. Immunity. 2021;54(5):903–915. doi:10.1016/j.immuni.2021.04.021
91. Velez-Delgado A, Donahue KL, Brown KL, et al. Extrinsic KRAS signaling shapes the pancreatic microenvironment through fibroblast reprogramming. Cell Mol Gastroenterol Hepatol. 2022;13(6):1673–1699. doi:10.1016/j.jcmgh.2022.02.016
92. Timperi E, Gueguen P, Molgora M, et al. Lipid-associated macrophages are induced by cancer-associated fibroblasts and mediate immune suppression in breast cancer. Cancer Res. 2022;82(18):3291–3306. doi:10.1158/0008-5472.CAN-22-1427
93. Suzuki J, Aokage K, Neri S, et al. Relationship between podoplanin-expressing cancer-associated fibroblasts and the immune microenvironment of early lung squamous cell carcinoma. Lung Cancer. 2021;153:1–10. doi:10.1016/j.lungcan.2020.12.020
94. Sakai T, Aokage K, Neri S, et al. Link between tumor-promoting fibrous microenvironment and an immunosuppressive microenvironment in stage I lung adenocarcinoma. Lung Cancer. 2018;126:64–71. doi:10.1016/j.lungcan.2018.10.021
95. Cho H, Seo Y, Loke KM, et al. Cancer-stimulated CAFs enhance monocyte differentiation and protumoral TAM Activation via IL6 and GM-CSF secretion. Clin Cancer Res. 2018;24(21):5407–5421. doi:10.1158/1078-0432.CCR-18-0125
96. Stadler M, Pudelko K, Biermeier A, et al. Stromal fibroblasts shape the myeloid phenotype in normal colon and colorectal cancer and induce CD163 and CCL2 expression in macrophages. Cancer Lett. 2021;520:184–200. doi:10.1016/j.canlet.2021.07.006
97. Zhang A, Qian Y, Ye Z, et al. Cancer-associated fibroblasts promote M2 polarization of macrophages in pancreatic ductal adenocarcinoma. Cancer Med. 2017;6(2):463–470. doi:10.1002/cam4.993
98. Li X, Sun Z, Peng G, et al. Single-cell RNA sequencing reveals a pro-invasive cancer-associated fibroblast subgroup associated with poor clinical outcomes in patients with gastric cancer. Theranostics. 2022;12(2):620–638. doi:10.7150/thno.60540
99. Cohen N, Shani O, Raz Y, et al. Fibroblasts drive an immunosuppressive and growth-promoting microenvironment in breast cancer via secretion of Chitinase 3-like 1. Oncogene. 2017;36(31):4457–4468. doi:10.1038/onc.2017.65
100. Maller O, Drain AP, Barrett AS, et al. Tumour-associated macrophages drive stromal cell-dependent collagen crosslinking and stiffening to promote breast cancer aggression. Nat Mater. 2021;20(4):548–559. doi:10.1038/s41563-020-00849-5
101. Gabrilovich DI. Myeloid-Derived Suppressor Cells. Cancer Immunol Res. 2017;5(1):3–8. doi:10.1158/2326-6066.CIR-16-0297
102. Xiang H, Ramil CP, Hai J, et al. Cancer-associated fibroblasts promote immunosuppression by inducing ROS-Generating Monocytic MDSCs in lung squamous cell carcinoma. Cancer Immunol Res. 2020;8(4):436–450. doi:10.1158/2326-6066.CIR-19-0507
103. Zhang R, Qi F, Zhao F, et al. Cancer-associated fibroblasts enhance tumor-associated macrophages enrichment and suppress NK cells function in colorectal cancer. Cell Death Dis. 2019;10(4):273. doi:10.1038/s41419-019-1435-2
104. Zhao Q, Huang L, Qin G, et al. Cancer-associated fibroblasts induce monocytic myeloid-derived suppressor cell generation via IL-6/exosomal miR-21-activated STAT3 signaling to promote cisplatin resistance in esophageal squamous cell carcinoma. Cancer Lett. 2021;518:35–48. doi:10.1016/j.canlet.2021.06.009
105. Wculek SK, Cueto FJ, Mujal AM, et al. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. 2020;20(1):7–24. doi:10.1038/s41577-019-0210-z
106. Cheng JT, Deng Y-N, Yi H-M, et al. Hepatic carcinoma-associated fibroblasts induce IDO-producing regulatory dendritic cells through IL-6-mediated STAT3 activation. Oncogenesis. 2016;5(2):e198. doi:10.1038/oncsis.2016.7
107. Berzaghi R, Tornaas S, Lode K, et al. Ionizing radiation curtails immunosuppressive effects from cancer-associated fibroblasts on dendritic cells. Front Immunol. 2021;12:662594. doi:10.3389/fimmu.2021.662594
108. Huang TX, Tan X-Y, Huang H-S, et al. Targeting cancer-associated fibroblast-secreted WNT2 restores dendritic cell-mediated antitumour immunity. Gut. 2022;71(2):333–344. doi:10.1136/gutjnl-2020-322924
109. Zhao P, Zhang Y, Li W, et al. Recent advances of antibody drug conjugates for clinical applications. Acta Pharm Sin B. 2020;10(9):1589–1600. doi:10.1016/j.apsb.2020.04.012
110. Ziani L, Chouaib S, Thiery J. Alteration of the antitumor immune response by cancer-associated fibroblasts. Front Immunol. 2018;9:414. doi:10.3389/fimmu.2018.00414
111. Santos AM, Jung J, Aziz N, et al. Targeting fibroblast activation protein inhibits tumor stromagenesis and growth in mice. J Clin Invest. 2009;119(12):3613–3625. doi:10.1172/JCI38988
112. Scott AM, Wiseman G, Welt S, et al. A Phase I dose-escalation study of sibrotuzumab in patients with advanced or metastatic fibroblast activation protein-positive cancer. Clin Cancer Res. 2003;9(5):1639–1647.
113. Qian L, Tang Z, Yin S, et al. Fusion of dendritic cells and cancer-associated fibroblasts for activation of anti-tumor cytotoxic T lymphocytes. J Biomed Nanotechnol. 2018;14(10):1826–1835. doi:10.1166/jbn.2018.2616
114. Xia Q, Zhang -F-F, Geng F, et al. Anti-tumor effects of DNA vaccine targeting human fibroblast activation protein α by producing specific immune responses and altering tumor microenvironment in the 4T1 murine breast cancer model. Cancer Immunol Immunother. 2016;65(5):613–624. doi:10.1007/s00262-016-1827-4
115. Kraman M, Bambrough PJ, Arnold JN, et al. Suppression of antitumor immunity by stromal cells expressing fibroblast activation protein-alpha. Science. 2010;330(6005):827–830. doi:10.1126/science.1195300
116. Cancer Discovery. FAP-Targeted 4-1BB agonist tamps down advanced tumors. Cancer Discov. 2023;13(8):Of2. doi:10.1158/2159-8290.CD-NB2023-0044
117. Hofheinz RD, al-Batran S-E, Hartmann F, et al. Stromal antigen targeting by a humanised monoclonal antibody: an early phase II trial of sibrotuzumab in patients with metastatic colorectal cancer. Onkologie. 2003;26(1):44–48. doi:10.1159/000069863
118. Cunningham CC. Talabostat. Expert Opin Investig Drugs. 2007;16(9):1459–1465. doi:10.1517/13543784.16.9.1459
119. Roberts EW, Deonarine A, Jones JO, et al. Depletion of stromal cells expressing fibroblast activation protein-α from skeletal muscle and bone marrow results in cachexia and anemia. J Exp Med. 2013;210(6):1137–1151. doi:10.1084/jem.20122344
120. Özdemir BC, Pentcheva-Hoang T, Carstens JL, et al. Depletion of carcinoma-associated fibroblasts and fibrosis induces immunosuppression and accelerates pancreas cancer with reduced survival. Cancer Cell. 2015;28(6):831–833. doi:10.1016/j.ccell.2015.11.002
121. Wei Y, Kim TJ, Peng DH, et al. Fibroblast-specific inhibition of TGF-β1 signaling attenuates lung and tumor fibrosis. J Clin Invest. 2017;127(10):3675–3688. doi:10.1172/JCI94624
122. Mariathasan S, Turley SJ, Nickles D, et al. TGFβ attenuates tumour response to PD-L1 blockade by contributing to exclusion of T cells. Nature. 2018;554(7693):544–548. doi:10.1038/nature25501
123. Faivre S, Santoro A, Kelley RK, et al. Novel transforming growth factor beta receptor I kinase inhibitor galunisertib (LY2157299) in advanced hepatocellular carcinoma. Liver Int. 2019;39(8):1468–1477. doi:10.1111/liv.14113
124. Carapuça EF, Gemenetzidis E, Feig C, et al. Anti-stromal treatment together with chemotherapy targets multiple signalling pathways in pancreatic adenocarcinoma. J Pathol. 2016;239(3):286–296. doi:10.1002/path.4727
125. Sherman MH, Yu R, Engle D, et al. Vitamin D receptor-mediated stromal reprogramming suppresses pancreatitis and enhances pancreatic cancer therapy. Cell. 2014;159(1):80–93. doi:10.1016/j.cell.2014.08.007
126. Ene-Obong A, Clear AJ, Watt J, et al. Activated pancreatic stellate cells sequester CD8+ T cells to reduce their infiltration of the juxtatumoral compartment of pancreatic ductal adenocarcinoma. Gastroenterology. 2013;145(5):1121–1132. doi:10.1053/j.gastro.2013.07.025
127. Ishihara S, Inman DR, Li W-J, et al. Mechano-signal transduction in mesenchymal stem cells induces prosaposin secretion to drive the proliferation of breast cancer cells. Cancer Res. 2017;77(22):6179–6189. doi:10.1158/0008-5472.CAN-17-0569
128. Gorchs L, Ahmed S, Mayer C, et al. The vitamin D analogue calcipotriol promotes an anti-tumorigenic phenotype of human pancreatic CAFs but reduces T cell mediated immunity. Sci Rep. 2020;10(1):17444. doi:10.1038/s41598-020-74368-3
129. Li X, Yong T, Wei Z, et al. Reversing insufficient photothermal therapy-induced tumor relapse and metastasis by regulating cancer-associated fibroblasts. Nat Commun. 2022;13(1):2794. doi:10.1038/s41467-022-30306-7
130. Wang X, Dai G, Jiang G, et al. A TMVP1-modified near-infrared nanoprobe: molecular imaging for tumor metastasis in sentinel lymph node and targeted enhanced photothermal therapy. J Nanobiotechnology. 2023;21(1):130. doi:10.1186/s12951-023-01883-6
131. Feig C, Jones JO, Kraman M, et al. Targeting CXCL12 from FAP-expressing carcinoma-associated fibroblasts synergizes with anti-PD-L1 immunotherapy in pancreatic cancer. Proc Natl Acad Sci U S A. 2013;110(50):20212–20217. doi:10.1073/pnas.1320318110
132. Hurwitz HI, Uppal N, Wagner SA, et al. Randomized, double-blind, phase ii study of ruxolitinib or placebo in combination with capecitabine in patients with metastatic pancreatic cancer for whom therapy with gemcitabine has failed. J Clin Oncol. 2015;33(34):4039–4047. doi:10.1200/JCO.2015.61.4578
133. Cazet AS, Hui MN, Elsworth BL, et al. Targeting stromal remodeling and cancer stem cell plasticity overcomes chemoresistance in triple negative breast cancer. Nat Commun. 2018;9(1):2897. doi:10.1038/s41467-018-05220-6
134. Olive KP, Jacobetz MA, Davidson CJ, et al. Inhibition of Hedgehog signaling enhances delivery of chemotherapy in a mouse model of pancreatic cancer. Science. 2009;324(5933):1457–1461. doi:10.1126/science.1171362
135. Kim EJ, Sahai V, Abel EV, et al. Pilot clinical trial of hedgehog pathway inhibitor GDC-0449 (vismodegib) in combination with gemcitabine in patients with metastatic pancreatic adenocarcinoma. Clin Cancer Res. 2014;20(23):5937–5945. doi:10.1158/1078-0432.CCR-14-1269
136. Huang Y, Simms AE, Mazur A, et al. Fibroblast activation protein-α promotes tumor growth and invasion of breast cancer cells through non-enzymatic functions. Clin Exp Metastasis. 2011;28(6):567–579. doi:10.1007/s10585-011-9392-x
137. Li J, Kataoka K. Chemo-physical strategies to advance the in vivo functionality of targeted nanomedicine: the next generation. J Am Chem Soc. 2021;143(2):538–559. doi:10.1021/jacs.0c09029
138. Wen P, Ke W, Dirisala A, et al. Stealth and pseudo-stealth nanocarriers. Adv Drug Deliv Rev. 2023;198:114895. doi:10.1016/j.addr.2023.114895
139. Ikeda-Imafuku M, Wang LL-W, Rodrigues D, et al. Strategies to improve the EPR effect: a mechanistic perspective and clinical translation. J Control Release. 2022;345:512–536. doi:10.1016/j.jconrel.2022.03.043
140. Sun R, Xiang J, Zhou Q, et al. The tumor EPR effect for cancer drug delivery: current status, limitations, and alternatives. Adv Drug Deliv Rev. 2022;191:114614. doi:10.1016/j.addr.2022.114614
141. Sindhwani S, Syed AM, Ngai J, et al. The entry of nanoparticles into solid tumours. Nat Mater. 2020;19(5):566–575. doi:10.1038/s41563-019-0566-2
142. Peng F, Setyawati MI, Tee JK, et al. Nanoparticles promote in vivo breast cancer cell intravasation and extravasation by inducing endothelial leakiness. Nat Nanotechnol. 2019;14(3):279–286. doi:10.1038/s41565-018-0356-z
143. Guo J, Zeng H, Shi X, et al. A CFH peptide-decorated liposomal oxymatrine inactivates cancer-associated fibroblasts of hepatocellular carcinoma through epithelial-mesenchymal transition reversion. J Nanobiotechnology. 2022;20(1):114. doi:10.1186/s12951-022-01311-1
144. Chen B, Wang Z, Sun J, et al. A tenascin C targeted nanoliposome with navitoclax for specifically eradicating of cancer-associated fibroblasts. Nanomedicine. 2016;12(1):131–141. doi:10.1016/j.nano.2015.10.001
145. Chen B, Dai W, Mei D, et al. Comprehensively priming the tumor microenvironment by cancer-associated fibroblast-targeted liposomes for combined therapy with cancer cell-targeted chemotherapeutic drug delivery system. J Control Release. 2016;241:68–80. doi:10.1016/j.jconrel.2016.09.014
146. Fourniols T, Bastien E, Canevat A, et al. Inhibition of colorectal cancer-associated fibroblasts by lipid nanocapsules loaded with acriflavine or paclitaxel. Int J Pharm. 2020;584:119337. doi:10.1016/j.ijpharm.2020.119337
147. Hou L, Liu Q, Shen L, et al. Nano-delivery of fraxinellone remodels tumor microenvironment and facilitates therapeutic vaccination in desmoplastic melanoma. Theranostics. 2018;8(14):3781–3796. doi:10.7150/thno.24821
148. Chen Y, Hu M, Wang S, et al. Nano-delivery of salvianolic acid B induces the quiescence of tumor-associated fibroblasts via interfering with TGF-beta1/Smad signaling to facilitate chemo- and immunotherapy in desmoplastic tumor. Int J Pharm. 2022;623:121953. doi:10.1016/j.ijpharm.2022.121953
149. Miao L, Liu Q, Lin CM, et al. Targeting tumor-associated fibroblasts for therapeutic delivery in desmoplastic tumors. Cancer Res. 2017;77(3):719–731. doi:10.1158/0008-5472.CAN-16-0866
150. Tan Y-N, Li Y-P, Huang J-D, et al. Thermal-sensitive lipid nanoparticles potentiate anti-PD therapy through enhancing drug penetration and T lymphocytes infiltration in metastatic tumor. Cancer Letters. 2021;522:238–254. doi:10.1016/j.canlet.2021.09.031
151. Li Y, Wu J, Lu Q, et al. GA&HA-modified liposomes for co-delivery of aprepitant and curcumin to inhibit drug-resistance and metastasis of hepatocellular carcinoma. Int J Nanomedicine. 2022;17:2559–2575. doi:10.2147/IJN.S366180
152. Wu J, Qi C, Wang H, et al. Curcumin and berberine co-loaded liposomes for anti-hepatocellular carcinoma therapy by blocking the cross-talk between hepatic stellate cells and tumor cells. Front Pharmacol. 2022;13:961788. doi:10.3389/fphar.2022.961788
153. Li X, Luo Y, Huang Z, et al. Multifunctional liposomes remodeling tumor immune microenvironment for tumor chemoimmunotherapy. Small Methods. 2023;7(5):e2201327. doi:10.1002/smtd.202201327
154. Ferraz FS, Lopez JL, Lacerda SM, et al. Biotechnological approach to induce human fibroblast apoptosis using superparamagnetic iron oxide nanoparticles. J Inorg Biochem. 2020;206:111017.
155. Kovacs D, Igaz N, Marton A, et al. Core-shell nanoparticles suppress metastasis and modify the tumour-supportive activity of cancer-associated fibroblasts. J Nanobiotechnology. 2020;18(1):18. doi:10.1186/s12951-020-0576-x
156. Dilnawaz F, Singh A, Mohanty C, et al. Dual drug loaded superparamagnetic iron oxide nanoparticles for targeted cancer therapy. Biomaterials. 2010;31(13):3694–3706. doi:10.1016/j.biomaterials.2010.01.057
157. Mardhian DF, Vrynas A, Storm G, et al. FGF2 engineered SPIONs attenuate tumor stroma and potentiate the effect of chemotherapy in 3D heterospheroidal model of pancreatic tumor. Nanotheranostics. 2020;4(1):26–39. doi:10.7150/ntno.38092
158. Xia C, Pan J, Wang J, et al. Functional blockade of cancer-associated fibroblasts with ultrafine gold nanomaterials causes an unprecedented bystander antitumoral effect. Nanoscale. 2020;12(38):19833–19843. doi:10.1039/D0NR04682E
159. Hossen MN, Rao G, Dey A, et al. Gold nanoparticle transforms activated cancer-associated fibroblasts to quiescence. ACS Appl Mater Interfaces. 2019;11(29):26060–26068. doi:10.1021/acsami.9b03313
160. Zhang Y, Elechalawar CK, Hossen MN, et al. Gold nanoparticles inhibit activation of cancer-associated fibroblasts by disrupting communication from tumor and microenvironmental cells. Bioact Mater. 2021;6(2):326–332. doi:10.1016/j.bioactmat.2020.08.009
161. Zheng D, Wan C, Yang H, et al. Her2-targeted multifunctional nano-theranostic platform mediates tumor microenvironment remodeling and immune activation for breast cancer treatment. Int J Nanomedicine. 2020;15:10007–10028. doi:10.2147/IJN.S271213
162. Dadfar SM, Roemhild K, Drude NI, et al. Iron oxide nanoparticles: diagnostic, therapeutic and theranostic applications. Adv Drug Deliv Rev. 2019;138:302–325. doi:10.1016/j.addr.2019.01.005
163. Nicolas-Boluda A, Vaquero J, Laurent G, et al. Photothermal depletion of cancer-associated fibroblasts normalizes tumor stiffness in desmoplastic cholangiocarcinoma. ACS Nano. 2020;14(5):5738–5753. doi:10.1021/acsnano.0c00417
164. Wang Y, Xia H, Chen B, et al. Rethinking nanoparticulate polymer-drug conjugates for cancer theranostics. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2023;15(1):e1828. doi:10.1002/wnan.1828
165. Zuo T, Zhang J, Yang J, et al. On-demand responsive nanoplatform mediated targeting of CAFs and down-regulating mtROS-PYK2 signaling for antitumor metastasis. Biomater Sci. 2021;9(5):1872–1885. doi:10.1039/D0BM01878C
166. Donthireddy L, Vonteddu P, Murthy T, et al. ONP-302 nanoparticles inhibit tumor growth by altering tumor-associated macrophages and cancer-associated fibroblasts. Journal of Cancer. 2022;13(6):1933–1944. doi:10.7150/jca.69338
167. Getts DR, Terry RL, Getts MT, et al. Therapeutic inflammatory monocyte modulation using immune-modifying microparticles. Sci Transl Med. 2014;6(219):219ra7. doi:10.1126/scitranslmed.3007563
168. Zheng S, Wang J, Ding N, et al. Prodrug polymeric micelles integrating cancer-associated fibroblasts deactivation and synergistic chemotherapy for gastric cancer. J Nanobiotechnology. 2021;19(1):381. doi:10.1186/s12951-021-01127-5
169. Zhao J, Wang H, Hsiao C-H, et al. Simultaneous inhibition of hedgehog signaling and tumor proliferation remodels stroma and enhances pancreatic cancer therapy. Biomaterials. 2018;159:215–228. doi:10.1016/j.biomaterials.2018.01.014
170. Zhou C, Hu X, Liu Q, et al. Stromal barrier-dismantled nanodrill-like and cancer cell-targeted pH-responsive polymeric micelles for further enhancing the anticancer efficacy of doxorubicin. ACS Biomater Sci Eng. 2021;7(12):5690–5705. doi:10.1021/acsbiomaterials.1c01131
171. Monteiro PF, Gulfam M, Monteiro CJ, et al. Synthesis of micellar-like terpolymer nanoparticles with reductively-cleavable cross-links and evaluation of efficacy in 2D and 3D models of triple negative breast cancer. J Control Release. 2020;323:549–564. doi:10.1016/j.jconrel.2020.04.049
172. Ernsting MJ, Hoang B, Lohse I, et al. Targeting of metastasis-promoting tumor-associated fibroblasts and modulation of pancreatic tumor-associated stroma with a carboxymethylcellulose-docetaxel nanoparticle. J Control Release. 2015;206:122–130. doi:10.1016/j.jconrel.2015.03.023
173. Wang L, Liu X, Zhou Q, et al. Terminating the criminal collaboration in pancreatic cancer: nanoparticle-based synergistic therapy for overcoming fibroblast-induced drug resistance. Biomaterials. 2017;144:105–118. doi:10.1016/j.biomaterials.2017.08.002
174. Hou L, Chen D, Hao L, et al. Transformable nanoparticles triggered by cancer-associated fibroblasts for improving drug permeability and efficacy in desmoplastic tumors. Nanoscale. 2019;11(42):20030–20044. doi:10.1039/C9NR06438A
175. Ji T, Zhao Y, Ding Y, et al. Transformable peptide nanocarriers for expeditious drug release and effective cancer therapy via cancer-associated fibroblast activation. Angew Chem Int Ed Engl. 2016;55(3):1050–1055. doi:10.1002/anie.201506262
176. Huo M, Zhao Y, Satterlee AB, et al. Tumor-targeted delivery of sunitinib base enhances vaccine therapy for advanced melanoma by remodeling the tumor microenvironment. J Control Release. 2017;245:81–94. doi:10.1016/j.jconrel.2016.11.013
177. Panagi M, Mpekris F, Chen P, et al. Polymeric micelles effectively reprogram the tumor microenvironment to potentiate nano-immunotherapy in mouse breast cancer models. Nat Commun. 2022;13(1):7165. doi:10.1038/s41467-022-34744-1
178. Shen W, Yao P-A, Li W, et al. Cancer-associated fibroblast-targeted nanodrugs reshape colorectal tumor microenvironments to suppress tumor proliferation, metastasis and improve drug penetration. J Mater Chem B. 2023;11(9):1871–1880. doi:10.1039/D2TB02253B
179. Zheng F, Luo Y, Liu Y, et al. Nano-baicalein facilitates chemotherapy in breast cancer by targeting tumor microenvironment. Int J Pharm. 2023;635:122778. doi:10.1016/j.ijpharm.2023.122778
180. Tang Y, Yao W, Hu H, et al. TGF-beta blocking combined with photothermal therapy promote tumor targeted migration and long-term antitumor activity of CAR-T cells. Mater Today Bio. 2023;20:100615. doi:10.1016/j.mtbio.2023.100615
181. Zhou S, Zhen Z, Paschall AV, et al. FAP-targeted photodynamic therapy mediated by ferritin nanoparticles elicits an immune response against cancer cells and cancer associated fibroblasts. Adv Funct Mater. 2021;31(7). doi:10.1002/adfm.202007017
182. Li L, Zhou S, Lv N, et al. Photosensitizer-encapsulated ferritins mediate photodynamic therapy against cancer-associated fibroblasts and improve tumor accumulation of nanoparticles. Mol Pharm. 2018;15(8):3595–3599. doi:10.1021/acs.molpharmaceut.8b00419
183. Sitia L, Bonizzi A, Mazzucchelli S, et al. Selective Targeting of Cancer-Associated Fibroblasts by Engineered H-Ferritin Nanocages Loaded with Navitoclax. Cells. 2021;10(2):328. doi:10.3390/cells10020328
184. Yu Q, Qiu Y, Li J, et al. Targeting cancer-associated fibroblasts by dual-responsive lipid-albumin nanoparticles to enhance drug perfusion for pancreatic tumor therapy. J Control Release. 2020;321:564–575. doi:10.1016/j.jconrel.2020.02.040
185. Li J, Zhen X, Lyu Y, et al. Cell membrane coated semiconducting polymer nanoparticles for enhanced multimodal cancer phototheranostics. ACS Nano. 2018;12(8):8520–8530. doi:10.1021/acsnano.8b04066
186. Tan YN, Huang J-D, Li Y-P, et al. Near-infrared responsive membrane nanovesicles amplify homologous targeting delivery of anti-PD immunotherapy against metastatic tumors. Adv Healthc Mater. 2022;11(6):e2101496. doi:10.1002/adhm.202101496
187. Zhao T, Zhang R, He Q, et al. Partial ligand shielding nanoparticles improve pancreatic ductal adenocarcinoma treatment via a multifunctional paradigm for tumor stroma reprogramming. Acta Biomater. 2022;145:122–134. doi:10.1016/j.actbio.2022.03.050
188. Yingchoncharoen P, Kalinowski DS, Richardson DR. Lipid-based drug delivery systems in cancer therapy: what is available and what is yet to come. Pharmacol Rev. 2016;68(3):701–787. doi:10.1124/pr.115.012070
189. Saw PE, Park J, Lee E, et al. Effect of PEG pairing on the efficiency of cancer-targeting liposomes. Theranostics. 2015;5(7):746–754. doi:10.7150/thno.10732
190. Sonju JJ, Dahal A, Singh SS, et al. Peptide-functionalized liposomes as therapeutic and diagnostic tools for cancer treatment. J Control Release. 2021;329:624–644. doi:10.1016/j.jconrel.2020.09.055
191. Zhao L, Chen J, Pang Y, et al. Fibroblast activation protein-based theranostics in cancer research: a state-of-The-art review. Theranostics. 2022;12(4):1557–1569. doi:10.7150/thno.69475
192. Yilmaz A, Loustau T, Salomé N, et al. Advances on the roles of tenascin-C in cancer. J Cell Sci. 2022;135(18). doi:10.1242/jcs.260244
193. Kim MY, Kim OR, Choi YS, et al. Selection and characterization of tenascin C targeting peptide. Mol Cells. 2012;33(1):71–77. doi:10.1007/S10059-012-2214-4
194. Huynh NT, Passirani C, Saulnier P, et al. Lipid nanocapsules: a new platform for nanomedicine. Int J Pharm. 2009;379(2):201–209. doi:10.1016/j.ijpharm.2009.04.026
195. Hu H, Xu Q, Mo Z, et al. New anti-cancer explorations based on metal ions. J Nanobiotechnology. 2022;20(1):457. doi:10.1186/s12951-022-01661-w
196. Singh P, Pandit S, Mokkapati VRSS, et al. Gold nanoparticles in diagnostics and therapeutics for human cancer. Int J Mol Sci. 2018;19(7):1979. doi:10.3390/ijms19071979
197. Alhussan A, Bromma K, Perez MM, et al. Docetaxel-mediated uptake and retention of gold nanoparticles in tumor cells and in cancer-associated fibroblasts. Cancers (Basel). 2021;13(13):3157. doi:10.3390/cancers13133157
198. Alhussan A, Palmerley N, Smazynski J, et al. Potential of gold nanoparticle in current radiotherapy using a co-culture model of cancer cells and cancer associated fibroblast cells. Cancers (Basel). 2022;14(15):3586. doi:10.3390/cancers14153586
199. Park EJ, Yi J, Kim Y, et al. Silver nanoparticles induce cytotoxicity by a Trojan-horse type mechanism. Toxicol In Vitro. 2010;24(3):872–878. doi:10.1016/j.tiv.2009.12.001
200. Santhosh PB, Ulrih NP. Multifunctional superparamagnetic iron oxide nanoparticles: promising tools in cancer theranostics. Cancer Lett. 2013;336(1):8–17. doi:10.1016/j.canlet.2013.04.032
201. Lopez S, Hallali N, Lalatonne Y, et al. Magneto-mechanical destruction of cancer-associated fibroblasts using ultra-small iron oxide nanoparticles and low frequency rotating magnetic fields. Nanoscale Adv. 2022;4(2):421–436. doi:10.1039/D1NA00474C
202. Mardhian DF, Storm G, Bansal R, et al. Nano-targeted relaxin impairs fibrosis and tumor growth in pancreatic cancer and improves the efficacy of gemcitabine in vivo. J Control Release. 2018;290:1–10. doi:10.1016/j.jconrel.2018.09.031
203. Ghosh B, Biswas S. Polymeric micelles in cancer therapy: state of the art. J Control Release. 2021;332:127–147. doi:10.1016/j.jconrel.2021.02.016
204. Masood F. Polymeric nanoparticles for targeted drug delivery system for cancer therapy. Mater Sci Eng C Mater Biol Appl. 2016;60:569–578. doi:10.1016/j.msec.2015.11.067
205. Acharya S, Sahoo SK. PLGA nanoparticles containing various anticancer agents and tumour delivery by EPR effect. Adv Drug Deliv Rev. 2011;63(3):170–183. doi:10.1016/j.addr.2010.10.008
206. Liu J, He S, Luo Y, et al. Tumor-microenvironment-activatable polymer nano-immunomodulator for precision cancer photoimmunotherapy. Adv Mater. 2022;34(8):e2106654. doi:10.1002/adma.202106654
207. Sun X, Hong Y, Gong Y, et al. Bioengineered ferritin nanocarriers for cancer therapy. Int J Mol Sci. 2021;22:13.
208. Truffi M, Fiandra L, Sorrentino L, et al. Ferritin nanocages: a biological platform for drug delivery, imaging and theranostics in cancer. Pharmacol Res. 2016;107:57–65. doi:10.1016/j.phrs.2016.03.002
209. Kang S, Oltrogge LM, Broomell CC, et al. Controlled assembly of bifunctional chimeric protein cages and composition analysis using noncovalent mass spectrometry. J Am Chem Soc. 2008;130(49):16527–16529. doi:10.1021/ja807655t
210. Belletti D, Pederzoli F, Forni F, et al. Protein cage nanostructure as drug delivery system: magnifying glass on apoferritin. Expert Opin Drug Deliv. 2017;14(7):825–840. doi:10.1080/17425247.2017.1243528
211. Choi B, Park W, Park S-B, et al. Recent trends in cell membrane-cloaked nanoparticles for therapeutic applications. Methods. 2020;177:2–14. doi:10.1016/j.ymeth.2019.12.004
212. Zhen X, Cheng P, Pu K. Recent advances in cell membrane-camouflaged nanoparticles for cancer phototherapy. Small. 2019;15(1):e1804105. doi:10.1002/smll.201804105
213. Chen HY, Deng J, Wang Y, et al. Hybrid cell membrane-coated nanoparticles: a multifunctional biomimetic platform for cancer diagnosis and therapy. Acta Biomater. 2020;112:1–13. doi:10.1016/j.actbio.2020.05.028
214. Wang L, Wu M, Pan Y, et al. Sequential targeting biomimetic nano platform for enhanced mild photothermal therapy and chemotherapy of tumor. Comput Struct Biotechnol J. 2023;21:2780–2791. doi:10.1016/j.csbj.2023.04.024
215. Zang S, Huang K, Li J, et al. Metabolic reprogramming by dual-targeting biomimetic nanoparticles for enhanced tumor chemo-immunotherapy. Acta Biomater. 2022;148:181–193. doi:10.1016/j.actbio.2022.05.045
216. Hamson EJ, Keane FM, Tholen S, et al. Understanding fibroblast activation protein (FAP): substrates, activities, expression and targeting for cancer therapy. Proteomics Clin Appl. 2014;8(5–6):454–463. doi:10.1002/prca.201300095
217. Shin H, Kim Y, Jon S. Nanovaccine displaying immunodominant T cell epitopes of fibroblast activation protein is effective against desmoplastic tumors. ACS Nano. 2023;17(11):10337–10352. doi:10.1021/acsnano.3c00764
218. Guo C, Zhang W, Zhang Q, et al. Novel dual CAFs and tumour cell targeting pH and ROS dual sensitive micelles for targeting delivery of paclitaxel to liver cancer. Artif Cells Nanomed Biotechnol. 2023;51(1):170–179. doi:10.1080/21691401.2023.2193221
219. Rhim AD, Oberstein P, Thomas D, et al. Stromal elements act to restrain, rather than support, pancreatic ductal adenocarcinoma. Cancer Cell. 2014;25(6):735–747. doi:10.1016/j.ccr.2014.04.021
220. Rajurkar M, De Jesus-Monge WE, Driscoll DR, et al. The activity of Gli transcription factors is essential for Kras-induced pancreatic tumorigenesis. Proc Natl Acad Sci U S A. 2012;109(17):E1038–47. doi:10.1073/pnas.1114168109
221. Cheng Y, Zou J, He M, et al. Spatiotemporally controlled Pseudomonas exotoxin transgene system combined with multifunctional nanoparticles for breast cancer antimetastatic therapy. J Control Release. 2023;367:167–83.
222. Tu Y, Zhang W, Fan G, et al. Paclitaxel-loaded ROS-responsive nanoparticles for head and neck cancer therapy. Drug Deliv. 2023;30(1):2189106. doi:10.1080/10717544.2023.2189106
223. Souers AJ, Leverson JD, Boghaert ER, et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat Med. 2013;19(2):202–208. doi:10.1038/nm.3048
224. Chen J, Jin S, Abraham V, et al. The Bcl-2/Bcl-XL/Bcl-w Inhibitor, Navitoclax, enhances the activity of chemotherapeutic agents in vitro and in vivo. Mol Cancer Ther. 2011;10(12):2340–2349. doi:10.1158/1535-7163.MCT-11-0415
225. Gandhi L, Camidge DR, Ribeiro de oliveira M, et al. Phase I study of Navitoclax (ABT-263), a novel Bcl-2 family inhibitor, in patients with small-cell lung cancer and other solid tumors. J Clin Oncol. 2011;29(7):909–916. doi:10.1200/JCO.2010.31.6208
226. Luo J, Zhang P, Zhao T, et al. Golgi apparatus-targeted chondroitin-modified nanomicelles suppress hepatic stellate cell activation for the management of liver fibrosis. ACS Nano. 2019;13(4):3910–3923. doi:10.1021/acsnano.8b06924
227. Liu H, Wu J, Qi C, et al. pH-responsive sulfated hyaluronic acid nanoparticles targeting tumor cells and CAFs for the treatment of breast cancer. Recent Pat Anti-Cancer Drug Disc. 2023;18(2):200–210. doi:10.2174/1574892817666220510123820
228. Zou C, Tang Y, Zeng P, et al. cRGD-modified nanoparticles of multi-bioactive agent conjugate with pH-sensitive linkers and PD-L1 antagonist for integrative collaborative treatment of breast cancer. Nanoscale Horiz. 2023;8(7):870–886. doi:10.1039/D2NH00590E
229. Jin J, Krishnamachary B, Barnett JD, et al. Human cancer cell membrane-coated biomimetic nanoparticles reduce fibroblast-mediated invasion and metastasis and induce T-cells. ACS Appl Mater Interfaces. 2019;11(8):7850–7861. doi:10.1021/acsami.8b22309
230. Liu Z, Ji P, Liu H, et al. FNIII14 peptide-enriched membrane nanocarrier to disrupt stromal barriers through reversing CAFs for augmenting drug penetration in tumors. Nano Lett. 2023; 23(21):9963–71.
231. Ni C, Lou X, Yao X, et al. ZIP1(+) fibroblasts protect lung cancer against chemotherapy via connexin-43 mediated intercellular Zn(2+) transfer. Nat Commun. 2022;13(1):5919. doi:10.1038/s41467-022-33521-4
232. Ford K, Hanley CJ, Mellone M, et al. NOX4 inhibition potentiates immunotherapy by overcoming cancer-associated fibroblast-mediated CD8 T-cell exclusion from tumors. Cancer Res. 2020;80(9):1846–1860. doi:10.1158/0008-5472.CAN-19-3158
233. Huang H, Wang Z, Zhang Y, et al. Mesothelial cell-derived antigen-presenting cancer-associated fibroblasts induce expansion of regulatory T cells in pancreatic cancer. Cancer Cell. 2022;40(6):656–673.e7. doi:10.1016/j.ccell.2022.04.011
234. Correa S, Grosskopf AK, Lopez Hernandez H, et al. Translational applications of hydrogels. Chem Rev. 2021;121(18):11385–11457. doi:10.1021/acs.chemrev.0c01177
235. Zeng Z, Zhang C, Li J, et al. Activatable polymer nanoenzymes for photodynamic immunometabolic cancer therapy. Adv Mater. 2021;33(4):e2007247. doi:10.1002/adma.202007247
236. Wang X, Ihara S, Li X, et al. Rod-scale design strategies for immune-targeted delivery system toward cancer immunotherapy. ACS Nano. 2019;13(7):7705–7715. doi:10.1021/acsnano.9b01271
237. Yang Y, Guo J, Huang L. Tackling TAMs for cancer immunotherapy: it’s nano time. Trends Pharmacol Sci. 2020;41(10):701–714. doi:10.1016/j.tips.2020.08.003
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.