Back to Archived Journals » Botanics: Targets and Therapy » Volume 6
The anticancer properties of phytochemical extracts from Salvia plants
Authors Jiang Y, Zhang L, Rupasinghe V
Received 19 October 2015
Accepted for publication 15 January 2016
Published 30 March 2016 Volume 2016:6 Pages 25—44
DOI https://doi.org/10.2147/BTAT.S98610
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Ayse Kuruuzum-Uz
Yuanyuan Jiang,1,2 Li Zhang,2 HP Vasantha Rupasinghe1,3
1Department of Environmental Sciences, Faculty of Agriculture, Dalhousie University, Truro, NS, Canada; 2College of Science, Sichuan Agricultural University, Yaan, Sichuan, People's Republic of China; 3Department of Pathology, Faculty of Medicine, Dalhousie University, Halifax, NS, Canada
Abstract: Salvia species have been used as traditional medicine in many countries for a long time for health benefits. More importantly, in recent decades, the extracts of Salvia species have been shown to exhibit significant anticancer effects in vitro and in vivo on a wide range of cancer types. Therefore, this review provides a systematic summary of the anticancer profile and the underlying mechanisms of the extracts from Salvia species, which reveals the potential of these species, especially Salvia miltiorrhiza and Salvia officinalis, to be used as natural anticancer agents or auxiliary agents and bring new insights for further research and development of the genus Salvia.
Keywords: Salvia, chemoprevention, anticancer, phytochemicals, sage, Danshen
Introduction
Cancer, a major obstacle to human health and well-being, is still one of the main “killers” leading to death, whether in developed or developing countries. Based on the latest statistics on trends in cancer incidence and mortality worldwide, the International Agency for Research on Cancer reported in 2012 the worldwide burden of cancer rose to an estimated 14 million new cases per year, a figure expected to rise to 22 million annually within the next 2 decades.1 The social and economic burden due to cancer incidence is enormous especially in the developing countries which have more than 60% of the world’s total cases of cancer and account for ~70% of the world’s cancer deaths because of the lack of early detection and access to treatments.1,2 Thus, cancer remains a major health problem worldwide, and effective prevention and treatment of cancer remain imperative.3
Over the years, anticancer agents have been derived from both natural as well as synthetic chemicals. However, synthetic drugs need to pass through a labyrinth of regulatory stages before their recommendation could be made for public use and pose a great risk for unexpected toxic side effects, which affect and limit their use. In contrast, natural compounds from dietary sources perceived as nontoxic are more appealing than those from nondietary sources. As well, humans have naturally acquired the ability to consume them without documented side effects. Due to the cost-effectiveness and wide human acceptance, natural agents, such as phytochemicals, attract more scientific research interest.4,5 From around the 1940s to the early 2010s, of the small molecule anticancer drugs, more than 70% are nonsynthetic, with almost 50% being either natural products or derived from natural products,6 such as the widely used anticancer agents taxanes, vinca alkaloids, and camptothecin class of compounds. Research indicated that different cancers have different causes, respond differently to treatments, and are consequently more or less curable. In most cases, the outcome of therapy is very different for different cancers, and there is a large diversity in the spectrum of mutations in individual cancer patients.7,8 Recently, a lot of bioactive components have been shown to modulate cell signaling pathways to mediate the anticancer effect, such as Notch, Wnt, mitogen-activated protein kinase (MAPK), and phosphatidylinositol 3-kinase (PI3K)/Akt pathways.9–13 Activated PI3K phosphorylates phosphatidylinositol (4,5)-bisphosphate by converting to phosphatidylinositol (3,4,5)-triphosphate, leading to the recruitment of Akt to the plasma membrane, where it is phosphorylated and activated. Akt is a key kinase downstream of PI3K in the signaling pathway.14 PI3K/Akt shows cross-talks with other pathways that regulate cell growth or survival, such as the MAPK pathway. Its constitutive activation is found in various types of cancers.15 Therefore, screening and investigating the anticancer property of phytochemicals and making clear the mechanism of actions will benefit the finding and developing of more potentially effective anticancer agents.
The genus Salvia (belonging to the family Lamiaceae) consists of nearly 1,000 species and has undergone marked species extinction in three regions of the world: Central and South America (500 spp.), Central Asia/Mediterranean (250 spp.), and Eastern Asia (90 spp.).16 The Salvia species, usually referred to as “sage”, are among the important aromatic spice plants worldwide.17 The name of the genus, Salvia, is derived from the Latin salvere, in reference to the curative properties of the plant, which means it has a historical reputation of being recognized as a medicinal herb.18 The main phytochemical components of Salvia species are flavonoids and terpenoids. Some of the Salvia species have been used worldwide as traditional herbal medicine as well as flavoring spices due to the presence of naturally occurring phytochemicals.19 The aerial parts of these plants usually contain flavonoids and triterpenoids as well as the volatile compounds, such as monoterpenoids, while in the roots, the main compounds are diterpenoids. Several abietane diterpenes with aromatic or quinoid C-rings were isolated from Salvia species, many of which possessed biological activities.18 Salvia miltiorrhiza (Figure 1A) is one of the most recognized medicinal plants and its root (“Danshen”, Figure 1B) has been widely used as traditional herbal medicine in clinics in the People’s Republic of China, Korea, Japan, and other Asian countries for the treatment of various microcirculatory disturbance-related diseases, such as cardiac-cerebral vascular disease, liver dysfunction, and diabetic vascular complication.20 Tanshinones are quite well known abietane diterpenes, first isolated from S. miltiorrhiza. Salvia officinalis (Figure 1C), another important sage widely used in Europe for both culinary and medicinal preparations,21,22 is reported to have the highest amount of essential oil in leaves compared to the other species of Salvia, which exhibits potent antioxidant capacity, as well as anticancer, antispasmodic, antimicrobial, anti-inflammatory, and carminative and mucolytic activities.18,19,23,24 Many other species of the genus Salvia have also been studied for their composition and most of the phytochemicals are with potential anticancer properties (Table 1). In recent years, the potential use of Salvia as a new anticancer agent has been recognized.25–28 Therefore, in this article, the anticancer property of phytochemicals from Salvia plants including their mechanism of action is reviewed.
 | Figure 1 Salvia miltiorrhiza (A); the roots of S. miltiorrhiza (Danshen) (B); and Salvia officinalis (C). |
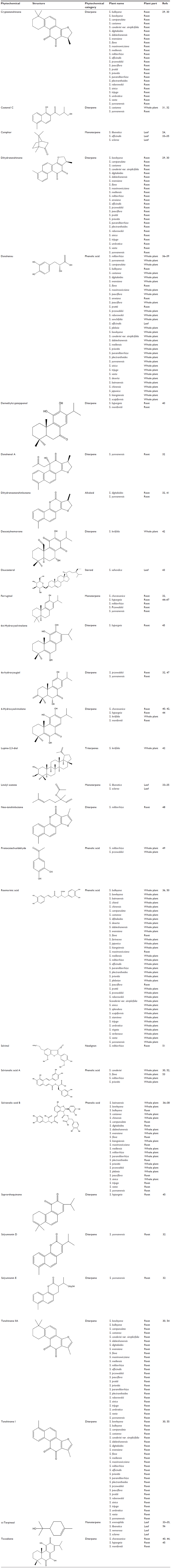 | Table 1 Phytochemicals with potential anticancer properties in Salvia genus |
Evidence of genus Salvia as a potential therapeutic source for various cancers
Breast cancer
According to the data from World Cancer Report 2014, breast cancer has become the most common cancer affecting women worldwide, accounting for 25.2% of the total cancer.19 The major risk factors for breast cancer were classified into three major categories: reproductive (hormone exposure), genetic, and environmental.57 Furthermore, characterizing the expression of three receptors, namely, the estrogen receptor (ER), progesterone receptor, and the human epidermal growth factor receptor 2 (HER2), has been an important part of the standard assessment of breast tumors in current clinical practice.7 Since hormone replacement therapy was considered to increase the risk of breast cancer,58,59 investigation of other possible alternatives has gained importance.
Phytoestrogens are naturally occurring plant compounds that are capable of eliciting estrogen-like properties through binding to ER and other mechanisms.60 Epidemiological evidence suggests a lower incidence of breast cancer in Asia relative to that in Western countries and this relationship is due to the high consumption of phytoestrogen-rich foods (soybean and their products) in Asia. Although the relationship between phytoestrogen intake and breast cancer risk is not yet conclusive in epidemiology, ample evidence shows that phytoestrogen-rich plants have a positive effect in decreasing breast tumor incidence. The effect of phytoestrogen on breast cancer is reported to mediate through multiple signaling pathways including ataxia telangiectasia and Rad3-related, Wnt/β-catenin, PI3K/Akt, nuclear factor-κB (NF-kB), p53, insulin like growth factor 1, MAPK, ER-mediated and BRCA-mediated.61–65
Liposoluble compounds (such as tanshinone IIA [1,6,6-trimethyl-8,9-dihydro-7H-naphtho[1,2-g][1]benzofuran-10,11-dione] and tanshinone I [1,6-dimethylphenanthro[1,2-b]furan-10,11-dione]) rich in S. miltiorrhiza and related species are regarded as having phytoestrogenic effects with antiproliferation of human breast cancer MCF-7 mediated by ER.66 HER2 is a receptor tyrosine kinase and is involved in signal transduction pathways leading to tumor cell proliferation. Yang et al67 have found that Danshen extract strongly inhibited the proliferation of both MCF-7 vector control cells and HER2-overexpressed MCF-7 cells via a blockade of cell cycle entry into the S phase, which was mediated by the downregulation of Akt phosphorylation and an upregulation of p27 (a cell-cycle inhibitor). Meanwhile, the extract has downregulated the tyrosine phosphorylation of HER2 in MCF-7 cells. Nevertheless, MCF-7 cells were more resistant to the Danshen-induced inhibition of Akt phosphorylation and p27 upregulation; that is to say, the overexpression of HER2 could confer resistance to Danshen in MCF-7 cells. The apoptosis of eukaryotic cells is mainly introduced by death receptor-induced external apoptotic pathway, mitochondria-induced inner apoptotic pathway, Granzyme B-induced apoptosis pathway, and endoplasmic reticulum stress-induced apoptotic pathway.68 For ER-negative human breast cancer cells BT-20, tanshinone IIA may be inducing ER stress and MAPK pathway to induce apoptosis and inhibit proliferation resulting in an increased protein expression of caspase-12, GADD153, phospho-p38, phosphor-JNK, caspase-3, and Bax, but decreased Bcl-xl and phospho-extracellular signal-regulated kinase expression.54 Meanwhile, tanshinone IIA inhibited the proliferation of MDA-MB-231 cells and decreased the protein expression of phosphatidylethanolamine conjugated microtubule-associated protein 1A/1B-light chain 3 (LC3-II) and Erb-B2 (also known as HER2) in vitro. When effected on an MDA-MB-231 xenograft animal model, tanshinone IIA resulted in a reduction in tumor size and weight with the downregulation of NF-κB p65 and upregulation of caspase-3 compared to the control group.69 While compared with tanshinone IIA, tanshinone I was more sensitive in inhibiting the growth of MCF-7 and especially MDA-MB-231 cell lines in part by downregulation of Aurora A expression and function. Therefore, Aurora A perhaps was an important functional target of tanshinone I action. Therefore, tanshinone I has the potential to be developed as an effective and safe agent for the therapy and prevention of breast cancer in further investigation as well as with potent antiangiogenic activity and minimal side effects in vivo.70 Phosphatase and tensin homolog deleted on chromosome 10 (PTEN) is an antagonist of PI3K signaling by dephosphorylating phosphatidylinositol (3,4,5)-triphosphate at position D3 to generate phosphatidylinositol (4,5)-bisphosphate.71 Daucosterol, a β-sitosterol glycoside, is isolated from Salvia sahendica. Research indicated that daucosterol inhibited cell proliferation by inducing apoptosis. The inhibition of cell growth and survival was 52% when the cells were treated with 20 μM of daucosterol for 24 hours by 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay and the possible cellular mechanisms are PI3K/Akt inhibition by upregulating PTEN and triggering of reactive oxygen species production, which induces mitochondrial oxidative stress and subsequent release of cytochrome c followed by caspase-3 activation and apoptosis.43 Metastasis is a complicated multistep process that involves interactions between cancer cells and their surrounding microenvironment, and angiogenesis is a critical step in tumor growth and metastasis.72,73 Vascular endothelial growth factor (VEGF) is a major mediator of angiogenesis in cancer, and overexpression of VEGF in cancer tissues was found associated with tumor growth and metastasis. The expression of VEGF is induced under hypoxic conditions in which the alpha subunit of hypoxia inducible factor-1 plays an important role.74 The ethanolic crude extract of Salvia triloba inhibited the expression of VEGF and the hypoxia inducible factor-1α at mRNA level in MCF-7 cells under both normoxic and hypoxic conditions.75
Paclitaxel, a plant-derived chemotherapy agent for breast cancer, has been widely used since it was available in clinics. However, chemotherapy resistance greatly restricts the use of first-line chemotherapeutics paclitaxel. Transgelin 2 may mediate paclitaxel resistance by activating PI3K/Akt signaling pathway to suppress human breast cancer paclitaxel resistance cell (MCF-7/PTX) apoptosis. Research has indicated that salvianolic acid A, one of the important phenolic compounds found in many Salvia species,30 can reverse paclitaxel resistance through suppressing transgelin 2 expression by involving attenuation of PI3K/Akt pathway activation and adenosine-triphosphate binding cassette transporter upregulation.52 Choi et al49 have indicated that protocatechualdehyde, a polyphenol in S. miltiorrhiza and related species,76,77 suppressed proliferation of breast cancer cells (MCF-7 and MDA-MB-231) via an estrogen-dependent manner.49 In addition, neo-tanshinlactone isolated from S. miltiorrhiza was active against ER-positive human breast cancer cell lines (MCF-7 and ZR-75-1) (with half maximal inhibitory concentration [IC50] values of 0.6 and 0.3 μg/mL, respectively) and HER-2-overexpressing breast cancer cell line (SK-BR-3, HER-2+), but was inactive against two ER cell lines (MDA-MB-231 and HS 587-T) (with IC50>10 μg/mL).48 Furthermore, the extracts of S. africana-caerulea, S. africana-lutea, S. albicaulis, S. aurita, S. chamelaeagnea, S. diserma, S. dolomitica, S. garipensis, S. lanceola, S. muirii, S. namaensis, S. radula, S. repens, S. runcinata, S. schlechteri, S. stenophylla, and S. verbenaca, the Salvia species used in traditional medicine in South Africa, showed cytotoxic effects on human breast cancer cells (MCF-7) with IC50 values ranging between 9.7 and 43.7 μg/mL.78
Lung cancer
Lung cancer is the most common cancer in men and the third most common in women. It is one of the most aggressive human cancers, with a 5-year overall survival of 10%–15%. Adenocarcinoma, squamous cell carcinoma, small cell carcinoma, and large cell carcinoma are the four major histological types of lung cancer.1 In a previous research, taxodione and 6α-hydroxysalvinolone isolated from Salvia hypargeia have been reported to possess cytotoxic response in human lung cancer.45 Recently, it was found that danshenol A and 6α-hydroxysugiol from Salvia yunnanensis also exhibit substantial cytotoxicity in human lung cancer cell line NCI-H460.32
It is reported that tanshinones in S. miltiorrhiza and related species have specific effects on lung cancer cell lines.55,79 Tanshinone I, tanshinone IIA, and cryptotanshinone reduced the cell viability when exposed to highly invasive human lung adenocarcinoma cell line CL1-5. However, tanshinone I and cryptotanshinone had no effect on cell apoptosis that is different from tanshinone IIA. Meanwhile, among these compounds, tanshinone I significantly inhibited lung cancer cell migration, invasion, and gelatinase activity in vitro; reduced metastasis, angiogenesis, and tumorigenesis in vivo which may occur through platelet-derived growth factor β and its downstream pathways; and suppressed the expression of angiogenic factor interleukin-8 through NF-κB and interleukin-8 activator protein-1 pathways.79 When exposed to non-small cell lung cancer (H1299, H23, A549), the three components have the ability to inhibit the cell proliferation in vitro, with tanshinone I being the most potent, via cell cycle arrest and apoptosis induction which is different from the effect on CL1-5. Cryptotanshinone and tanshinone I inhibited cell proliferation by arresting cell cycle at S phase, but tanshinone IIA arrested cell cycle at G2/M phase. In a mouse model, tanshinone I significantly reduced the final tumor weight associated with inducing cancer cell apoptosis, inhibiting cancer cell proliferation, and reducing angiogenesis, without significant adverse effect on food intake or body weight. Aurora A, a member of a novel oncogenic family of mitotic serine/threonine kinases, may be an important molecular target for tanshinone I action against lung cancer.55 Recently, Xie et al80 have found that tanshinone IIA may suppress A549 proliferation (IC50 is 145.3 μmol/L, treatment for 24 hours, cell counting kit [CCK] method), induce apoptosis and cell cycle arrest at the S phase, and decrease the expression of VEGF and VEGF receptor 2 (VEGFR2). The molecular mechanisms of action of tanshinone IIA in the VEGF/VEGFR signaling pathway could be through tanshinone IIA binding to the VEGFR2 kinase domain, blocking the downstream pathways of VEGF/VEGFR after VEGF combined with VEGFR2 in vascular endothelial cells. In addition, research indicated that salvianolic acid A can significantly decrease A549 cell growth (cell growth was mostly blocked at G1 phase) and promote partial apoptosis. When treated on A549 cell with 100 μg/mL salvianolic acid A for 24 hours, the inhibitory ratio of cell growth was ~90% by the MTT assay. Salvianolic acid A treatment increased mitochondrial membrane permeability and increased the cleaved caspase-3 protein level at molecular level, as well as upregulated phosphatase and tensin homolog [PTEN] protein level and downregulated Akt phosphorylation which may explain the possible mechanisms in salvianolic acid A-induced apoptosis in A549 cells.81
Prostate cancer
Globally, prostate cancer is the second most frequently diagnosed cancer and the fifth most common cause of cancer death among men, with an estimated 1.1 million new cases (15% of all cancers in men) and 0.3 million cancer deaths (7% of all cancer deaths in men) in 2012.1 Current therapeutic modalities for prostate cancer usually have variable effectiveness and develop metastasis and drug resistance associated with high toxicity to normal tissues. Therefore, searching for more effective and safe agents for the chemoprevention of prostate cancer remains the top priority in prostate cancer research.82
In an earlier research, 6α-hydroxysalvinolone and taxodione isolated from S. hypargeia were found to have cytotoxic activity in a hormone-dependent human prostate cancer cell LNCaP.45 The methanol crude extracts of Salvia menthaefolia, Salvia sclarea, Salvia dominica, Salvia spinos, and Salvia palestina exhibited antiproliferative activity against prostate cancer cell MDA Pca2b.83 6-Hydroxysalvinolone, deacetylnemorone, and lupine-2,3-diol isolated from Salvia leriifolia exhibited a potent antiproliferative activity against the prostate cancer cell PC-3.42 Danshenol A isolated from S. yunnanensis exhibited substantial cytotoxicity on PC-3.32 As for the action mechanism, Won et al84 showed that tanshinone IIA induces p53 activation and mitochondrial dysfunction, leading to caspase-9/caspase-3 mediated apoptosis, as well as appears to inhibit PI3K/Akt survival pathway to contribute to apoptosis induction signaling. Gong et al82 have made a research on the effect of bioactive tanshinones inhibiting the growth of prostate cancer cells. When treated on androgen-sensitive LNCaP and androgen-independent PC-3 and DU-145 human prostate cancer cells using tanshinones (including cryptotanshinone, tanshinone I and tanshinone IIA), tanshinone I was the most potent compound to inhibit cell growth and especially for PC-3. Among them, tanshinone I, the most potent one in apoptosis induction of PC-3, also had the ability to inhibit the growth of DU-145 prostate tumor in mice associated with induction of apoptosis, decrease of proliferation, inhibition of angiogenesis, and downregulation of Aurora A, whereas it did not alter food intake or body weight. Therefore, Aurora A was considered to play an important function in prostate cancer cell growth and may be a functional target of tanshinones. Interestingly, cryptotanshinone and tanshinone I arrested the cell cycle progression at S phase, whereas tanshinone IIA did it at G2/M phases. Recently, Atmaca and Bozkurt85 have shown that the methanolic extract of S. triloba was found to be cytotoxic in both PC-3 and DU-145 cells. IC50 values (treatment for 72 hours) of the extract in PC-3 and DU-145 cells are 287±8 and 456±15 μg/mL, respectively. PC-3 cells were more sensitive to the extract than DU-145 cells with changes in angiogenic cytokines, and the VEGF level was significantly decreased by 2.5-fold compared with the control when treated with 287 μg/mL of the extract for 72 hours in PC-3 cells. Those results strengthen the evidence for the potential use of the genus Salvia as a natural resource of antiprostate cancer agents.
Colorectal cancer
Colorectal cancer represents almost 10% of the global cancer incidence burden in 2012, and is the third most common cancer in men (an estimated 746,000 cases) and the second most common in women (614,000 cases). Colorectal cancer is the fourth most common cause of death from cancer worldwide, with an estimated 694,000 deaths.1 Saprorthoquinone and taxodione isolated from S. hypargeia have been reported for their cytotoxic activity on human colon cancer (Col 2) more than a decade ago.45 The crude methanolic extracts from S. menthaefolia have exhibited the in vitro growth inhibitory activity on colorectal adenocarcinoma (WiDr and HT-29) with IC50 values of 89.6 and 95.3 μg/mL, respectively.83 The extracts of S. africana-caerulea, S. africana-lutea, S. albicaulis, S. aurita, S. chamelaeagnea, S. diserma, S. lanceola, S. muirii, S. namaensis, S. radula, S. repens, S. runcinata, S. schlechteri, S. stenophylla, and S. verbenaca have shown cytotoxic effects on human colon cancer cells (HT-29) with IC50 values ranging between 17.1 and 57.0 μg/mL.78 Tanshinone IIA and tanshinone I have an effect on the growth inhibition and apoptotic induction in human colon cancer Colo 205 cells. While tanshinone IIA arrested Colo-205 cells in the G1 phase, tanshinone I arrested Colo 205 cells in the G0/G1 phase. Tanshinone IIA induces apoptosis in Colo 205 cells through both mitochondrial-mediated intrinsic and Fas-mediated extrinsic caspase cell-death pathways, and tanshinone I induces apoptosis in Colo 205 cells through both mitochondrial-mediated intrinsic cell-death pathways and p21-mediated G0/G1 cell cycle arrest.86,87
An epidemiological study has shown that nutrition is a key factor in modulating sporadic colorectal carcinoma risk. Aromatic plants of the genus Salvia have been attributed many medicinal properties, including anticolorectal cancer activity. When treated on the two human colon carcinoma-derived cell lines HCT15 and CO115, the water extracts of Salvia fruticosa and S. officinalis have a significant inhibition of HCT15 cell proliferation, while the extracts and rosmarinic acid (the main phenolic compound of these extracts) all significantly induced apoptosis in a concentration-dependent manner in both HCT15 and CO115 cells. However, they did not affect PI3K/Akt pathway but inhibited the MAPK/extracellular signal-regulated kinase pathway in the Kirsten rat sarcoma viral oncogene homolog mutated HCT15 cell line and not in the V-raf murine sarcoma viral oncogene homolog B1 mutated CO115 cell which maybe results from an inhibition of Kirsten rat sarcoma viral oncogene homolog, upstream to V-raf murine sarcoma viral oncogene homolog B1.11
Multidrug resistance, the resistance of cancer cells to a variety of structurally unrelated chemotherapeutic agents following exposure to a single cytotoxic compound, is a major factor in the failure of many forms of chemotherapy.88 There are specific correlations between cell membrane transporters or pumps and a drug-resistant phenotype. The overexpression of a glycoprotein, termed P-glycoprotein (P-gp), is associated with “reduced drug permeation” in multidrug-resistant cells, which mediates the efflux of cytotoxic drugs out of cancer cells by functioning as an efflux pump. Therefore, the selection of P-gp-inhibitor cytotoxic agent represents a logical approach to overcome multidrug resistance in cancer chemotherapy.29,89 Among the main tanshinones of tanshinone I, tanshinone IIA, cryptotanshinone, dihydrotanshinone, and miltirone, only cryptotanshinone and dihydrotanshinone decreased digoxin efflux ratio in a concentration-dependent manner, indicating their inhibitory effects on P-gp function, whereas tanshinone I, tanshinone IIA, and miltirone had no inhibitory effects in Caco-2 cells (human colorectal cancer cells). Moreover, both cryptotanshinone and dihydrotanshinone could potentiate the cytotoxicity of doxorubicin and irinotecan in P-gp overexpressing colon cancer cells (SW620/Ad-300). The mechanistic studies revealed that these two tanshinones increased intracellular accumulation of the P-gp substrate anticancer drugs, presumably by downregulating P-gp mRNA and protein levels, and inhibiting P-gp ATPase activity.29 5-Fluorouracil, one of the chemotherapeutic medicines for colon cancer, has low efficacy and resistance in therapy.90 The Colo 205 xenograft model cotreated with tanshinone IIA plus 5-fluorouracil caused a reduction in the xenograft tumor volumes and decreased the protein expression of P-gp and LC3-II expression compared to 5-fluorouracil alone.91 Therefore, tanshinone IIA has the potential to be used as the adjuvant agent in the chemotherapy of colon cancer.
Linalyl acetate, α-terpineol, and camphor, the main components of the essential oil from Lebanese sage (Salvia libanotica), synergistically induced cell arrest and apoptosis resulting in the inhibition of the growth of two isogenic human colon cancer cell lines HCT-116 (p53+/+ and p53–/–). In addition, by the treatment of protocatechualdehyde, cyclin D1 promoter activity was significantly inhibited and poly adenosine diphosphate (ADP)-ribose polymerase cleavage was dramatically increased in human colorectal cancer cells (HCT116 and SW480). The latest research has indicted that exposure of protocatechualdehyde activated the levels of activating transcription factor 3 (ATF3) protein and mRNA in HCT116 and SW480 cells. However, ATF3 overexpression enhanced protocatechualdehyde-mediated cleavage of poly ADP-ribose polymerase.92,93 Transcriptional induction of the ATF3 gene has been related to the downregulation of cyclin D1 transcription.94 Meanwhile, ATF3 knockdown inhibited apoptosis and cell viability. Therefore, protocatechualdehyde possesses cell viability inhibition and induction of apoptosis activity in human colorectal cancer cells (HCT116 and SW480) resulting in ATF3 expression and transcriptional activation.93
Liver cancer
Liver cancer represents 6% and 9% of the global cancer incidence and mortality burden, respectively. With an estimated 746,000 deaths in 2012, it is the second most common cause of death from cancer worldwide.1 In most populations, the major histological type of primary liver neoplasm is hepatocellular carcinoma.95
Existing studies have demonstrated that reduced glutathione (GSH) depletion is associated with mitochondrial dysfunction and induction of apoptosis, which is related to an increase in reactive oxygen species generation in cells; therefore, the balance of GSH and glutathione disulfide (GSSG) was considered to be very important to aerobic organisms.96,97 Earlier research has found that aqueous extract from S. miltiorrhiza has strongly inhibited the proliferation of hepatocellular carcinoma cells (HepG2) and caused apoptotic cell death in vitro, which is related to the depletion of intracellular GSH and reduction of mitochondrial membrane potential.98 The tanshinones (cryptotanshinone, tanshinone I, tanshinone IIA, and dihydrotanshinone) have the ability to decrease cell viability of HepG2 cells. However, only tanshinone IIA induced apoptosis in HepG2 cells without inducing changes in the GSH/GSSG ratio, an indicator of oxidative stress, despite the observed augmentation in GSH and oxidized GSSG, and the others showed lower efficacy in inducing apoptosis despite inducing changes in intracellular GSH/GSSG ratio in the HepG2 cells.96 In addition, research has indicated that tanshinones have cytotoxic effects on doxorubicin-resistant human liver cancer cells. Cryptotanshinone suppressed doxorubicin efflux, a process mediated by P-gp, in P-gp-overexpressed R-HepG2 (a subclone of HepG2) cells. Compared with cryptotanshinone, dihydrotanshinone, and tanshinone, tanshinone IIA provided the best synergism with doxorubicin.99 Recently, Jeon et al100 have declared that tanshinone IIA induces apoptotic cell death in HepG2 cells by the inhibition of cytochrome P450 2J2, which is highly expressed in human tumors and carcinoma cell lines. And in vivo, HepG2 cell-based tumor growth was significantly decreased in a dose-dependent manner after the treatment with tanshinone IIA.
The alcohol extract of Salvia chinensis can inhibit the growth and angiogenesis of tumor in H22 bearing mice while significantly reducing the express of VEGF and microvessel density.101 The total flavonoids from S. chinensia (95% ethanolic extract) induced HepG2 and Huh-7 (human hepatocellular cancer cell) cell apoptosis by suppressing cellular NF-κB signaling. In vivo, the extract (total flavonoids) is able to induce H22 cell apoptosis without detectable toxic impacts on normal hepatocytes and vital organs of tumor-bearing mice.102
The failure of the immune system either to recognize or mediate destruction of tumor cells which results in the development and progression of tumor, however, could be corrected by immunotherapy. The ideal cancer treatment is not only to cause destruction of local tumors, but also to activate a systemic antitumor immunity.103 Therefore, the development of immunotherapies holds great potential for cancer treatment.104 There are several studies on polysaccharide antitumor immunology mechanism, and it is generally believed that improving the host’s immune function is one of its antitumor mechanisms.105 The typical polysaccharides isolated from plants show anticancer activity thought to be through activating the host immune response.105–108 Recently, a new polysaccharide SMP-W1 purified from commercial S. miltiorrhiza crude polysaccharide had an antiproliferative effect on hepatocellular carcinoma H22 cells in vitro, and simultaneously, significantly inhibited tumor growth and improved the immune response in tumor-bearing mice.109 In vivo, polysaccharides extracted by water method at 100°C from S. chinensis were able to stimulate an antitumor immune response in mice transplanted with H22 cells by alleviating tumor transplantation-induced CD4+ T-cell apoptosis and dysregulation of serum cytokine profiles. Most importantly, PSSC have no toxic effects on tumor-bearing animals compared with the cisplatin treatment group.110 These researches provide novel insights into the application of Salvia species extracts as a potential adjuvant reagent stimulating anticancer immune response in clinical hepatocellular carcinoma treatment.
Other type of cancers
Earlier research has indicated that oil extract of S. libanotica displayed strong growth inhibitory effects on a mouse papilloma-derived cell line (SP-l) following 24 hours of treatment with an estimated IC50 of 50 μg/mL, and the growth inhibition was due to cytostatic and not cytotoxic effects.34 Melanoma is an aggressive form of skin cancer that has limited therapeutic options,111 with more than 232,000 new cases and ~55,000 deaths in 2012.1 Research has indicated that essential oils of Salvia bracteata and Salvia rubifolia from Lebanon exhibited an inhibitory effect on the human melanoma cells M14 through induction of apoptotic cell death. The essential oil of S. rubifolia was significantly more effective than the essential oil of S. bracteata that maybe resulted from the difference in the concentration of sesquiterpenes.112 Russo et al24 have shown that essential oil of S. officinalis caused the inhibition of cell growth and induction of apoptosis cell death when treated on human melanoma cell lines (A375, M14, and A2058). Because of the influence of environmental and pedoclimatic conditions on the qualitative chemical composition of essential oil, there is difference in the anticancer activity among the essential oils of S. officinalis collected from different places. α- And β-thujone isomers associated with the synergism of other compounds present in the essential oil such as camphor may be related to the potential anticancer activity of the essential oils. In addition, danshensu (β-(3,4-dihydroxyphenyl)-d-lactic acid), one of the major water-soluble components of Danshen, has antitumor activity in B16F10 melanoma via affecting tumor angiogenesis and tumor invasion by downregulation of the expression of VEGF and matrix metalloproteinases-2 and -9.113
The data indicated that stomach cancer is the fifth most common cancer worldwide in 2012. Most stomach cancers are gastric carcinomas, which are malignant epithelial neoplasms.1 Despite varied treatment strategies, the control of this cancer at an advanced stage remains challenging. Therefore, the development of novel therapeutic strategies or agents to improve the life expectancy of gastric cancer patients is also an urgent need.114 Tanshinone IIA can induce gastric cancer cell (MKN45 and SGC7901 cell lines) growth inhibition and apoptosis in vitro and in a gastric cancer mouse xenograft model, and it not only causes cell cycle arrest in the G2/M phase but also triggers the intrinsic apoptotic signaling pathway.115 SMPA, another neutral polysaccharide from the roots of S. miltiorrhiza, significantly enhanced the immune function of rats with N-methyl-N’-nitro-nitrosoguanidine-induced gastric cancer.114
Data from low-resource countries show that cervical cancer is still often the most common cancer among women.1 Tanshinone IIA purified from Danshen extract can induce apoptosis in HeLa cells through mitotic arrest at M phase without the disruption of microtubule structure in interphase cells, and induced apoptosis through the mitochondria-dependent pathway. In addition, it could trigger the mitotic arrested cells to enter apoptosis faster than vincristine or taxol.116 Pan et al117 have found that tanshinone IIA isolated from Danshen powder strongly inhibits the growth of HeLa cells through interfering in the process of microtubule assembly, leading to G2/M phase arrest and subsequent apoptosis. The interaction network analysis of 12 proteins suggested that tanshinone IIA treatment on HeLa cells regulates the expressions of proteins involved in apoptotic processes, spindle assembly, and p53 activation, including vimentin, maspin, α- and β-tubulin, and glucose-regulated protein 75. Meanwhile, salyunnanin E isolated from S. yunnanensis exhibits substantial cytotoxicity in HeLa cell with an IC50 value of 0.86 μmol/L.32 Human papilloma viruses (HPVs) are small double-stranded circular DNA viruses. Expression of HPV E5, E6, and E7 oncoproteins can alter multiple signaling pathways to cause cancer.118 Unfortunately, infection with high-risk HPV, such as 16 and 18, plays a major role in the incidence of cervical cancer.119 When treated on CaSki (which contains an integrated HPV16 genome as well as sequences related to HPV18, wt p53, and wt pRb), tanshinone IIA was found to downregulate expression of HPV E6 and E7 genes and modulate associated proteins E6AP and E2F1, as well as cause S phase cell cycle arrest and p53-mediated apoptosis. In vivo, tanshinone IIA (injection, 30 mg/kg in 100 μL) resulted in over 66% reduction in tumor volume of cervical cancer xenograft in athymic nude mice (CaSki cell injection).120 Therefore, tanshinone IIA has the potential to be a therapeutic agent for cervical cancer, as well as other HPV-related cancers.
Taxodione, ferruginol, and 6-hydroxysalvinolone isolated form Salvia chorassanica exhibited cytotoxic and apoptotic effectiveness against the leukemic cancer cells K562 and HL-60.44 Rosmarinic acid-induced cytotoxicity toward CCRF-CEM and CEM/ADR5000 (P-gp overexpressing, multidrug-resistant cell line) leukemic cancer cells with IC50 values of 14.6±1.58 and 44.5±5.3 μM, respectively. Rosmarinic acid binds to inhibitor κB kinase-β and decreases nuclear p65 translocation. The inactivation of NF-κB transcriptional activity may then lead to the deregulation of related gene.121 In addition, increased reactive oxygen species generation upon chemotherapeutic agents is related to mitochondrial DNA mutations, resulting in resistance of leukemia cells to DNA-damaging agents,122 and because rosmarinic acid induced necrosis and apoptosis through DNA damage in a reactive oxygen species-independent manner; therefore, the occurrence of multidrug resistance might be avoided by rosmarinic acid treatment.121 Recently, Zhang et al123 indicated that treatment with tanshinone IIA significantly prolonged the survival of acute promyelocytic leukemia bearing mice by inhibiting the proliferation and triggering apoptosis of acute promyelocytic leukemia cells.
In addition, tanshinone IIA has an effect of inhibiting the growth of human pancreatic cancer cell (PANC-1 cell line) and human ovarian cancer cells (A2780 and SKOV3 cell lines) in a concentration- and time-dependent manner and induce cell apoptosis.124–126 Acetone extract of S. hypargeia roots, among 16 Salvia extracts, showed the highest activity against the human ovarian cancer cell (A2780 cell line). 6-Hydroxysalvinolone and demethylcryptojaponol purified from S. hypargeia were found to be active against A2780 cell line with IC50 values of 3.9 and 1.2 μg/mL.40 Salvianolic acid B, one of polyphenols in Salvia species,30 significantly decreased the incidence of squamous carcinoma and inhibited angiogenesis during the process.37 When treated on human nasopharyngeal carcinoma cells (C666-1 cell line), salvianolic acid B inhibited the cell growth with an IC50 value of 80±6.8 μg/mL and induced cell apoptosis.38
In fact, there are many Salvia species that contain similar bioactive phytochemical profiles. Most of these species are grown in locations similar to that of S. miltiorrhiza and are of great development and utilization value. Therefore, the existing data on anticancer activity of known phytochemicals, such as tanshinones, salvianolic acid, and rosmarinic acid of Salvia species, can be used as important biomarkers in the selection and development of additional Salvia species. In addition, the current knowledge can also be used to investigate anticancer effects of phytochemicals with similar structure and synthesize analogues for screening against cancer. It seems that the high lipophilicity, short half-life, and low bioavailability of tanshinone IIA are some limitations in clinical application. Further research is required for reducing the cost of raw material through mass production of Salvia, green extraction techniques for the bioactives, and synthesis and assessment of structurally modified anticancer phytochemicals. Therefore, the development of phytogenic anticarcinogens still requires further investigation for exploring their fullest prospects.
Conclusion
As the largest genus in the Lamiaceae or Labiatae family, Salvia species have made a great contribution to human health all around the world. Evidence provided from laboratory studies supports the cancer preventive role of dietary intake of Salvia species against many types of cancers by interacting with signaling pathways, such as ER-mediated, MAPK/extracellular signal-regulated kinase, NF-κB, VEGF/VEGFR, PTEN/PI3K/Akt, p53, and mitochondria-dependent pathways. This paper has reviewed the recent advancements in the exploration of Salvia species in relation to anticancer effects, illustrating their potential as natural anticancer agents, and revealed phytochemicals in Salvia species exhibiting potential in cancer prevention and complementing conventional cancer treatments. However, the data collected by in vitro assays that reveal the potential of those extracts to be used as the anticancer candidates also should be tested by in vivo assays. Therefore, collaborated research and efforts of cancer research experts and oncologists in the world are still needed to demonstrate the safety, quality, and efficacy in systematic experimental animal models and human clinical trials before accepting phytochemicals of Salvia in clinic as anticancer agents or auxiliary agents based on existing research achievements.
Acknowledgments
This work is supported by China Scholarship Council (CSC), the Science and Technology Program Project Fund of Sichuan Province (No 2014FZ0056), and the Agriculture Science Technology Achievement Transformation Fund of Sichuan Province (No 14NZ0018).
Disclosure
The authors report no conflicts of interest in this work.
References
Stewart BW, Wild CP. World Cancer Report 2014. Lyon: World Health Organization Press; 2014. | |
Parker L. The impact of the environment on cancer genomics. In: Dellaire G, Berman JN, Arceci RJ, editors. Cancer Genomics. Chapter 26. Elsevier; 2014:449–465. | |
Heyninck K, Haegeman G, Goel A. Cancer chemoprevention by dietary polyphenols. In: Watso RR, Preedy VR, Zibadi S, editors. Polyphenols in Human Health and Disease. Chapter 90. London: Academic Press; 2014:1199–1216. | |
Mukhtar H. Chemoprevention: making it a success story for controlling human cancer. Cancer Lett. 2012;326:123–127. | |
Gupta P, Wright SE, Kim SH, Srivastava SK. Phenethyl isothiocyanate: A comprehensive review of anti-cancer mechanisms. Biochim Biophys Acta. 2014;1846:405–424. | |
Newman DJ, Cragg GM. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J Nat Prod. 2012;75:311–335. | |
Bydoun M, Marcato P, Dellaire G. Breast cancer genomics. Cancer Genomics. 2014;13:213–232. | |
Eggermont AMM, Caldas C, Ringborg U, Medema R, Tabernero J, Wiestler O. Cancer core Europe: A consortium to address the cancer care-cancer research continuum challenge. Eur J Cancer. 2014;50:2745–2746. | |
Yuan X, Wu H, Xu H, et al. Notch signaling: An emerging therapeutic target for cancer treatment. Cancer Lett. 2015;369:20–27. | |
Masuda M, Sawa M, Yamada T. Therapeutic targets in the Wnt signaling pathway: Feasibility of targeting TNIK in colorectal cancer. Pharmacol Ther. 2015;156:1–9. | |
Xavier CP, Lima CF, Fernandes-Ferreira M, Pereira-Wilson C. Salvia fruticosa, Salvia officinalis, and rosmarinic acid induce apoptosis and inhibit proliferation of human colorectal cell lines: the role in MAPK/ERK pathway. Nutr Cancer. 2009;61:564–571. | |
Fruman DA, Rommel C. PI3K and cancer: lessons, challenges and opportunities. Nat Rev Drug Discov. 2014;13:140–156. | |
Erstad DJ, Cusack JC. Targeting the NF-kB pathway in cancer therapy. Surg Oncol Clin N Am. 2013;22:705–746. | |
Mabuchi S, Kuroda H, Takahashi R, Sasano T. The PI3K/AKT/mTOR pathway as a therapeutic target in ovarian cancer. Gynecol Oncol. 2015;137:173–179. | |
Houede N, Pourquier P. Targeting the genetic alterations of the PI3K-AKT-mTOR pathway: its potential use in the treatment of bladder cancers. Pharmacol Ther. 2015;145:1–18. | |
Walker JB, Sytsma KJ, Treutlein J, Wink M. Salvia (lamiaceae) is not monophyletic: implications for the systematics, radiation, and ecological specializations of Salvia and tribe mentheae. Am J Bot. 2004;91:1115–1125. | |
Orhan IE, Senol FS, Ozturk N, Akaydin G, Sener B. Profiling of in vitro neurobiological effects and phenolic acids of selected endemic Salvia species. Food Chem. 2012;132:1360–1367. | |
Spiridon EK. Sage: The genus Salvia (Medicinal and Aromatic Plants-Industrial Profiles). London: Harwood Academic Publishers: Stewart; 2000:318. | |
Hamidpour M, Hamidpour R, Hamidpour S, Shahlari M. Chemistry, pharmacology, and medicinal property of sage (Salvia) to prevent and cure illnesses such as obesity, diabetes, depression, dementia, lupus, autism, heart disease, and cancer. J Tradit Complement Med. 2014;4:82–88. | |
Lu SP, Ting ZP. Composition identification of Salvia extracts and testing of its inhibiting myocytes cell death caused by hypoxia/reoxygenation. Carbohydr Polym. 2011;86:621–624. | |
Li MH, Li QQ, Zhang C, et al. An ethnopharmacological investigation of medicinal Salvia plants (Lamiaceae) in China. Acta Pharmaceut Sin B. 2013;3:273–280. | |
Lu YR, Foo LY. Flavonoid and phenolic glycosides from Salvia officinalis. Phytochemistry. 2000;55:263–267. | |
Martins N, Barros L, Santos-Buelga C, Henriques M, Silva S, Ferreira IC. Evaluation of bioactive properties and phenolic compounds in different extracts prepared from Salvia officinalis L. Food Chem. 2015;170:378–385. | |
Russo A, Formisano C, Rigano D, et al. Chemical composition and anticancer activity of essential oils of Mediterranean sage (Salvia officinalis L.) grown in different environmental conditions. Food Chem Toxicol. 2013;55:42–47. | |
Wang X, Morris-Natschke SL, Lee KH. New developments in the chemistry and biology of the bioactive constituents of Tanshen. Med Res Rev. 2007;27:133–148. | |
Zhang Y, Jiang PX, Ye M, Kim SH, Jiang C, Lü JX. Tanshinones: sources, pharmacokinetics and anti-cancer activities. Int J Mol Sci. 2012;13:13621–13666. | |
Chen X, Guo J, Bao J, Lu J, Wang Y. The anticancer properties of Salvia miltiorrhiza Bunge (Danshen): a systematic review. Med Res Rev. 2014;34:768–794. | |
Zhang WW, Lu YL. Advances in studies on antitumor activities of compounds in Salvia miltiorrhiza. Zhongguo Zhong Yao Za Zhi. 2010;35: 389–392. | |
Hu T, To KK, Wang L, et al. Reversal of P-glycoprotein (P-gp) mediated multidrug resistance in colon cancer cells by cryptotanshinone and dihydrotanshinone of Salvia miltiorrhiza. Phytomedicine. 2014;21:1264–1272. | |
Li MH, Li QQ, Liu YZ, et al. Pharmacophylogenetic study on plants of genus Salvia L. from China. Chin Herb Med. 2013;5: 164–181. | |
Pan ZH, Li Y, Wu XD, et al. Norditerpenoids from Salvia castanea Diels f. pubescens. Fitoterapia. 2012;83:1072–1074. | |
Wu CY, Liao Y, Yang ZG, et al. Cytotoxic diterpenoids from Salvia yunnanensis. Phytochemistry. 2014;106:171–177. | |
Itani WS, El-Banna SH, Hassan SB, Larsson RL, Bazarbachi A, Gali-Muhtasib HU. Anticolon cancer components from Lebanese sage (Salvia libanotica) essential oil: mechanistic basis. Cancer Biol Ther. 2014;7:1765–1773. | |
Gali-Muhtasib HU, Affara NI. Chemopreventive effects of sage oil on skin papillomas in mice. Phytomedicine. 2000;7:129–136. | |
DŽamić A, Soković M, Ristić M, Grujić-Jovanović S, Vukojević J, Marin PD. Chemical composition and antifungal activity of Salvia sclarea (Lamiaceae) essential oil. Arch Biol Sci. 2008;60: 233–237. | |
Li MH, Chen JM, Peng Y, Xiao PG. Distribution of phenolic acids in Chinese Salvia plants. World Sci Technol Mod Tradit Chin Med. 2008;10:46–52. Chinese. | |
Yang Y, Zou DR, Zhou ZT. Chemopreventive effect of salvianolic acid B on oral carcinogenesis. J Shanghai Jiaotong Univ (Medical Science). 2006;26:1122–1126. Chinese. | |
Liu X, Tang CZ, Wang L, Lin SX, Wan RQ, Chen J. Suppression of growth of human nasopharyngeal carcinoma cells by salvianolic acid B in vitro. J Guangzhou Univ Tradit Chinese Med. 2011;28:163–166, 218. Chinese. | |
Li MH, Chen JM, Peng Y, Wu QL, Xiao PG. Investigation of Danshen and related medicinal plants in China. J Ethnopharmacol. 2008;120:419–426. | |
Topcu G, Turkmen Z, Schilling JK, Kingston DGI, Pezzuto JM, Ulubelen A. Cytotoxic activity of some Anatolian Salvia extracts and isolated abietane diterpenoids. Pharm Biol. 2008;46:180–184. | |
Xu G, Yang J, Wang YY, et al. Diterpenoid constituents of the roots of Salvia digitaloides. J Agric Food Chem. 2000;58:12157–12161. | |
Choudhary MI, Hussain A, Adhikari A, et al. Anticancer and α-chymotrypsin inhibiting diterpenes and triterpenes from Salvia leriifolia. Phytochem Lett. 2013;6:139–143. | |
Esmaeili MA, Farimani MM. Inactivation of PI3K/Akt pathway and upregulation of PTEN gene are involved in daucosterol, isolated from Salvia sahendica, induced apoptosis in human breast adenocarcinoma cells. S Afr J Bot. 2014;93:37–47. | |
Tayarani-Najaran Z, Mousavi SH, Tajfard F, et al. Cytotoxic and apoptogenic properties of three isolated diterpenoids from Salvia chorassanica through bioassay-guided fractionation. Food Chem Toxicol. 2013;57:346–351. | |
Ulubelen A, Topçu G, Chai HB, Pezzuto JM. Cytotoxic activity of triterpenoids isolated from Salvia hypargeia. Pharm Biol. 1999;37:148–151. | |
Fu YJ, Shi T, Wang JA. Content determination of ferruginol in Salvia miltiorrhiza by HPLC and study on the influential factors. Chin Pharm. 2014;25:3690–3692. Chinese. | |
Yang LX. Studies on the Chemical Constituents and Bioactivities of Salvia Przewalskii Maxim [MD thesis]. Beijing: Union Medical College; 2011. Chinese. | |
Wang XH, Bastow KF, Sun CM, et al. Isolation, structure elucidation, total synthesis, and anti-breast cancer activity of neo-tanshinlactone from Salvia miltiorrhiza. J Med Chem. 2004;47: 5816–5819. | |
Choi J, Jiang X, Jeong JB, Lee SH. Anticancer activity of protocatechualdehyde in human breast cancer cells. J Med Food. 2014;17: 842–848. | |
Tepe, B. Antioxidant potentials and rosmarinic acid levels of the methanolic extracts of Salvia virgata (Jacq), Salvia staminea (Montbret and Aucher ex Bentham) and Salvia verbenaca (L.) from Turkey. Bioresour Technol. 2008;99:1584–1588. | |
Chang JY, Chang CY, Kuo CC, Chen LT, Wein YS, Kuo YH. Salvinal, a novel microtubule inhibitor isolated from Salvia miltiorrhizae Bunge (Danshen), with antimitotic activity in multidrug-sensitive and -resistant human tumor cells. Mol Pharmacol. 2004;65:77–84. | |
Cai JX, Chen SY, Zhang WP, et al. Salvianolic acid a reverses paclitaxel resistance in human breast cancer MCF-7 cells via targeting the expression of transgelin 2 and attenuating PI3 K/Akt pathway. Phytomedicine. 2014;21:1725–1732. | |
Lu YR, Foo LY. Polyphenolics of Salvia. Phytochemistry. 2002;59: 117–140. | |
Yan MY, Chien SY, Kuo SJ, Chen DR, Su CC. Tanshinone IIA inhibits BT-20 human breast cancer cell proliferation through increasing caspase 12, GADD153 and phospho-p38 protein expression. Int J Mol Med. 2012;29:855–863. | |
Li YL, Gong Y, Li LL, M. Abdolmaleky H, Zhou JR. Bioactive tanshinone I inhibits the growth of lung cancer in part via down regulation of Aurora A function. Mol Carcinog. 2013;52:535–543. | |
Rajabi Z, Ebrahimi M, Farajpour M, Mirza M, Ramshini H. Compositions and yield variation of essential oils among and within nine Salvia species from various areas of Iran. Ind Crops Prod. 2014;61:233–239. | |
Ban KA, Godellas CV. Epidemiology of breast cancer. Surg Oncol Clin N Am. 2014;23:409–422. | |
Cuzick J. Hormone replacement therapy and the risk of breast cancer. Eur J Cancer. 2008;44:2344–2349. | |
Moreira AC, Silva AM, Santos MS, Sardão VA. Phytoestrogens as alternative hormone replacement therapy in menopause: What is real, what is unknown. J Steroid Biochem Mol Biol. 2014;143:61–71. | |
Swann R, Perkins KA, Velentzis LS, et al. The DietCompLyf study: a prospective cohort study of breast cancer survival and phytoestrogen consumption. Maturitas. 2013;75:232–240. | |
Simmen RCM, Rahal OM, Montales MTE, et al. Soy foods: towards the development of novel therapeutics for breast cancer. In: Cho WCS, editor. Cancer Chemoprevention and Treatment by Diet Therapy. Chapter 5. Dordrecht: Springer; 2013:121–140. | |
Maggiolini M, Bonofiglio D, Marsico S, et al. Estrogen receptor α mediates the proliferative but not the cytotoxic dose-dependent effects of two major phytoestrogens on human breast cancer cells. Mol Pharmacol. 2001;60:595–602. | |
Vauzour D, Vafeiadou K, Rice-Evans C, Cadenas E, Spencer JP. Inhibition of cellular proliferation by the genistein metabolite 5,7,3’,4’-tetrahydroxyisoflavone is mediated by DNA damage and activation of the ATR signaling pathway. Arch Biochem Biophys. 2007;468: 159–166. | |
Cherdshewasart W, Panriansaen R, Picha P. Pretreatment with phytoestrogen-rich plant decreases breast tumor incidence and exhibits lower profile of mammary ERα and ERβ. Maturitas. 2007;58:174–181. | |
Hsieh CJ, Kuo PL, Hsu YC, et al. Arctigenin, a dietary phytoestrogen, induces apoptosis of estrogen receptor-negative breast cancer cells through the ROS/p38 MAPK pathway and epigenetic regulation. Free Radic Biol Med. 2014;67:159–170. | |
Zhao PW, Niu JZ, Wang DW, Chen JX, Wang LQ. Phytoestrogenic effects of six active components in Chinese medicine. Chin Pharmaceut J. 2007;42:1852–1855. Chinese. | |
Yang W, Ju JH, Jeon MJ, Han X, Shin I. Danshen (Salvia miltiorrhiza) extract inhibits proliferation of breast cancer cells via modulation of Akt activity and p27 level. Phytother Res. 2010;24:198–204. | |
Li M, Lin J. The apoptotic pathways and their mechanisms. Int J Obstetr Gynaecol. 2014;41:103–107. | |
Su CC, Chien SY, Kuo SJ, Chen YL, Cheng CY, Chen DR. Tanshinone IIA inhibits human breast cancer MDA-MB-231 cells by decreasing LC3-II, Erb-B2 and NF-kappaBp65. Mol Med Rep. 2012;5:1019–1022. | |
Gong Y, Li YL, Abdolmaleky HM, Li LL, Zhou JR. Tanshinones inhibit the growth of breast cancer cells through epigenetic modification of aurora A expression and function. PLoS One. 2012;7:1–11. | |
Chalhoub N, Baker SJ. PTEN and the PI3-kinase pathway in cancer. Ann Rev Phytopathol. 2009;4:127–150. | |
Chen JJW, Peck K, Hong TM, et al. Global analysis of gene expression in invasion by a lung cancer model. Cancer Res. 2001;61: 5223–5230. | |
Folkman J, Shing Y. Angiogenesis. J Biol Chem. 1992;267: 10931–10934. | |
Pugh CW, Ratcliffe PJ. Regulation of angiogenesis by hypoxia: role of the HIF system. Nat Med. 2003;9:677–684. | |
Zihlif1 M, Afifi F, Abu-Dahab R, et al. The antiangiogenic activities of ethanolic crude extracts of four Salvia species. BMC Complement Altern Med. 2013;13:358–367. | |
Gu M, Wang X, Su Z, Ouyang F. One-step separation and purification of 3,4-dihydroxyphenyllactic acid, salvianolic acid B and protocatechualdehyde from Salvia miltiorrhiza Bunge by high-speed counter-current chromatography. J Chromatogr A. 2007;1140:107–111. | |
Wu ZJ, Ouyang MG, Yang CR. Polyphenolic constituents of Salvia przewalskii. Acta Bot Yunnan. 1999;21:512–516. | |
Kamatou GPP, Van Zyl RL, Davids H, Van Heerden FR, Lourens ACU, Viljoen AM. Antimalarial and anticancer activities of selected South African Salvia species and isolated compounds from S. radula. S Afr J Bot. 2008;74:238–243. | |
Lee CY, Sher HF, Chen HW, et al. Anticancer effects of tanshinone I in human non-small cell lung cancer. Mol Cancer Ther. 2008;7:3527–3538. | |
Xie J, Liu JH, Liu H, et al. The antitumor effect of tanshinone IIA on anti-proliferation and decreasing VEGF/VEGFR2 expression on the human non-small cell lung cancer A549 cell line. Acta Pharmaceut Sin B. 2015;5:554–563. | |
Bi L, Chen JP, Yuan XJ, Jiang ZQ, Chen WP. Salvianolic acid A positively regulates PTEN protein level and inhibits growth of A549 lung cancer cells. Biomed Rep. 2013;1:213–217. | |
Gong Y, Li YL, Lu Y, et al. Bioactive tanshinones in Salvia miltiorrhiza inhibit the growth of prostate cancer cells in vitro and in mice. Int J Cancer. 2011;129:1042–1052. | |
Fiore G, Nencini C, Cavallo F, et al. In vitro antiproliferative effect of six Salvia species on human tumor cell lines. Phytother Res. 2006;20:701–703. | |
Won SH, Lee HJ, Jeong SJ, et al. Tanshinone IIA induces mitochondria dependent apoptosis in prostate cancer cells in association with an inhibition of phosphoinositide 3-kinase/AKT pathway. Biol Pharm Bull. 2010;33:1828–1834. | |
Atmaca H, Bozkurt E. Apoptotic and anti-angiogenic effects of Salvia triloba extract in prostate cancer cell lines. Tumor Biol. Epub 2015 Oct 12. DOI: 10.1007/s13277-015-4208-2. | |
Su CC, Chen GW, Lin JG. Growth inhibition and apoptosis induction by tanshinone I in human colon cancer Colo 205 cells. Int J Mol Med. 2008;22:613–618. | |
Su CC, Chen GW, Kang JC, Chan MH. Growth inhibition and apoptosis induction by tanshinone IIA in human colon adenocarcinoma cells. Planta Med. 2008;74:1357–1362. | |
Persidis A. Cancer multidrug resistance. Nat Biotechnol. 1999;17: 94–95. | |
Szakacs G, Paterson JK, Ludwig JA, Booth-Genthe C, Gottesman MM. Targeting multidrug resistance in cancer. Nat Rev Drug Discov. 2006;5:219–234. | |
Sheng YP, Guo WJ. Mechanisms of resistance to 5-FU and how to overcome them. J Int Oncol. 2006;33:502–505. | |
Su CC. Tanshinone IIA potentiates the efficacy of 5-FU in Colo205 colon cancer cells in vivo through down-regulation of P-gp and LC3-II. Exp Ther Med. 2012;3:555–559. | |
Jeong JB, Lee SH. Protocatechualdehyde possesses anti-cancer activity through down-regulating cyclin D1 and HDAC2 in human colorectal cancer cells. Biochem Biophys Res Commun. 2013;430:381–386. | |
Lee JR, Lee MH, Eo HJ, et al. The contribution of activating transcription factor 3 to apoptosis of human colorectal cancer cells by protocatechualdehyde, a naturally occurring phenolic compound. Arch Biochem Biophys. 2014;64:203–210. | |
James CG, Woods A, Underhill TM, Beier F. The transcription factor ATF3 is upregulated during chondrocyte differentiation and represses cyclin D1 and A gene transcription. BMC Mol Biol. 2006;7:30–41. | |
Bosetti C, Turati F, La Vecchia C. Hepatocellular carcinoma epidemiology. Best Pract Res Clin Gastroenterol. 2014;28:753–770. | |
Lee WY, Chiu LC, Yeung JH. Cytotoxicity of major tanshinones isolated from Danshen (Salvia miltiorrhiza) on HepG2 cells in relation to glutathione perturbation. Food Chem Toxicol. 2008;46:328–338. | |
Macho A, Hirsch T, Marzo I, et al. Glutathione depletion is an early and calcium elevation is a late event of thymocyte apoptosis. J Immunol. 1997;158:4612–4619. | |
Liu J, Shen HM, Ong CN. Salvia miltiorrhiza inhibits cell growth and induces apoptosis in human hepatoma HepG2 cells. Cancer Lett. 2000;153:85–93. | |
Lee WYW, Cheung CCM, Liu KWK, et al. Cytotoxic effects of tanshinones from Salvia miltiorrhiza on doxorubicin-resistant human liver cancer cells. J Nat Prod. 2010;73:854–859. | |
Jeon YJ, Kim JS, Hwang GH, et al. Inhibition of cytochrome P450 2J2 by tanshinone IIA induces apoptotic cell death in hepatocellular carcinoma HepG2 cells. Eur J Pharmacol. 2015;764:480–488. | |
Liu F, Liu JX, Ren JG, Li JM, Li HH. Effect of Salvia cheinensis extraction on angiogenesis of tumor. Zhongguo Zhong Yao Za Zhi. 2012;37:1285–1288. Chinese. | |
Xiang MX, Su HW, Hu YJ, Hu Y, Yang TM, Shu GG. Chemical composition of total flavonoids from Salvia chinensia Benth and their pro-apoptotic effect on hepatocellular carcinoma cells: potential roles of suppressing cellular NF-κB signaling. Food Chem Toxicol. 2013;62:420–426. | |
Wu F, Wang ZB, Lu P, et al. Activated anti-tumor immunity in cancer patients after high intensity focused ultrasound ablation. Ultrasound Med Biol. 2004;30:1217–1222. | |
Zhao DX, Li ZJ, Zhang Y, et al. Enhanced antitumor immunity is elicited by adenovirus-mediated gene transfer of CCL21 and IL-15 in murine colon carcinomas. Cell Immunol. 2014;289:155–161. | |
Inatsuka C, Yang Y, Gad E, Rastetter L, Disis ML, Lu H. Gamma delta T cells are activated by polysaccharide K (PSK) and contribute to the anti-tumor effect of PSK. Cancer Immunol Immunother. 2013;62:1335–1345. | |
Zhang Y, Li S, Wang X, Zhang L, Cheung PCK. Advances in lentinan: Isolation, structure, chain conformation and bioactivities. Food Hydrocoll. 2011;25:196–206. | |
Fan LP, Ding SD, Ai LZ, Deng KQ. Antitumor and immunomodulatory activity of water-soluble polysaccharide from Inonotus obliquus. Carbohydr Polym 2012;90:870–874. | |
Park SD, Lai YS, Kim CH. Immunopotentiating and antitumor activities of the purified polysaccharides from Phellodendron chinese SCHNEID. Life Sci. 2004;75:2621–2632. | |
Liu L, Jia J, Zeng G, et al. Studies on immunoregulatory and anti-tumor activities of a polysaccharide from Salvia miltiorrhiza Bunge. Carbohydr Polym. 2013;92:479–483. | |
Shu GG, Zhao WH, Yue L, Su HW, Xiang MX. Antitumor immunostimulatory activity of polysaccharides from Salvia chinensis Benth. J Ethnopharmacol. 2015;168:237–247. | |
Kaushik G, Kwatra D, Subramaniam D, Jensen RA, Anant S, Mammen JM. Honokiol affects melanoma cell growth by targeting the AMP-activated protein kinase signaling pathway. Am J Surg. 2014;208:995–1002. | |
Cardile V, Russo A, Formisano C, et al. Essential oils of Salvia bracteata and Salvia rubifolia from Lebanon: Chemical composition, antimicrobial activity and inhibitory effect on human melanoma cells. J Ethnopharmacol. 2009;126:265–272. | |
Zhang LJ, Chen L, Lu Y, et al. Danshensu has anti-tumor activity in B16F10 melanoma by inhibiting angiogenesis and tumor cell invasion. Eur J Pharmacol. 2010;643:195–201. | |
Wang N, Yang J, Lu J, et al. A polysaccharide from Salvia miltiorrhiza Bunge improves immune function in gastric cancer rats. Carbohydr Polym. 2014;111:47–55. | |
Chen J, Shi DY, Liu SL, Zhong L. Tanshinone IIA induces growth inhibition and apoptosis in gastric cancer in vitro and in vivo. Oncol Rep. 2012;27:523–528. | |
Zhou L, Chan WK, Xu N, et al. Tanshinone IIA, an isolated compound from Salvia miltiorrhiza Bunge, induces apoptosis in HeLa cells through mitotic arrest. Life Sci. 2008;83:394–403. | |
Pan TL, Hung YC, Wang PW, et al. Functional proteomic and structural insights into molecular targets related to the growth inhibitory effect of tanshinone IIA on HeLa cells. Proteomics. 2010;10: 914–929. | |
Chen JZ. Signaling pathways in HPV-associated cancers and therapeutic implications. Rev Med Virol. 2015;25:24–53. | |
Doorbar J, Egawa N, Griffin H, Kranjec C, Murakami I. Human papillomavirus molecular biology and disease association. Rev Med Virol. 2015;25:2–23. | |
Munagala R, Aqil F, Jeyabalan J, Gupta RC. Tanshinone IIA inhibits viral oncogene expression leading to apoptosis and inhibition of cervical cancer. Cancer Lett. 2015;356:536–546. | |
Wu CF, Hong CL, Klauck SM, Lin YL, Efferth T. Molecular mechanisms of rosmarinic acid from salvia miltiorrhiza in acute lymphoblastic leukemia cells. J Ethnopharmacol. 2015;176:55–68. | |
Carew JS, Zhou Y, Albitar M, Carew JD, Keating MJ, Huang P. Mitochondrial DNA mutations in primary leukemia cells after chemotherapy: clinical significance and therapeutic implications. Leukemia. 2003;17:1437–1447. | |
Zhang KJ, Li J, Meng WT, Xing HY, Yang YM. Tanshinone IIA inhibits acute promyelocytic leukemia cell proliferation and induces their apoptosis in vivo. Blood Cells Mol Dis. 2016;56:46–52. | |
Wang Y, Li Q, Fan ZZ, et al. Tanshinone IIA induces apoptosis of pancreatic cancer cells via the SAPK/JNK signal pathway. Word Chin J Digestol. 2011;19:1028–1033. | |
Zhu M, Chen P, Shi XY, Kong X, Guo LX, Zheng F. The effect of inhibition and inducing apoptosis of tanshinone IIA on human ovarian cancer A2780 cell lines. J Emerg Tradit Chin Med. 2009;18:596–597. Chinese. | |
Zhuang YY, Wang HL, Du RT, Zhang C. The studies of apoptosis effect and its mechanisms of tanshinone IIA on human ovarian cancer cell SKOV3. Int J Obstet Gynaecol. 2011;38:328–331. |
 © 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2016 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.