Back to Journals » International Journal of Nanomedicine » Volume 20
Tendon Tissue Engineering: Pathophysiological Mechanism and Bioengineering Therapy of Tendinopathy
Authors Chen B, Zhao Y, Guo S, Tang C, Xu B, Zhou M, Chen Q, Ma L, Lyu J, Guo L, Wang Y ![]()
Received 2 July 2025
Accepted for publication 16 September 2025
Published 15 October 2025 Volume 2025:20 Pages 12529—12571
DOI https://doi.org/10.2147/IJN.S550439
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kamakhya Misra
Bin Chen,1,* Yingqi Zhao,2,* Shizhen Guo,1 Chuyue Tang,1 Baoyun Xu,1 Mei Zhou,1 Qianbo Chen,1 Lin Ma,1 Jingtong Lyu,1 Lin Guo,1 Yunjiao Wang1
1Department of Orthopeadics/Sports Medicine Center, Southwest Hospital, Third Military Medical University, Chongqing, People’s Republic of China; 2Department of Orthopedics, Chongqing Hospital of The Chinese People’s Armed Police Force, Chongqing, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lin Guo, Department of Orthopeadics/Sports Medicine Center, State Key Laboratory of Trauma, Burn and Combined Injury, Southwest Hospital, Third Military Medical University, Gaotanyan Street. 30, Shapingba District, Chongqing, 400038, People’s Republic of China, Email [email protected] Yunjiao Wang, Department of Orthopeadics/Sports Medicine Center, State Key Laboratory of Trauma, Burn and Combined Injury, Southwest Hospital, Third Military Medical University, Gaotanyan Street. 30, Shapingba District, Chongqing, 400038, People’s Republic of China, Email [email protected]
Abstract: Tendinopathy afflicts many professional athletes and the elderly. However, due to the unique cellular and histological composition of tendons, healing is frequently unsatisfactory. The clinical physical therapy and surgical interventions often fail to meet patient expectations. In recent years, bioengineering technology has undergone rapid development, with a significant number of studies in the biological field focusing on bioengineering technology to explore emerging treatments for diseases. Therefore, bioengineering technology has the potential to become an important part of future tendon healing therapies. The present article will describe the sources of scaffolds, biological factors and bioengineering strategies, with a focus on their current applications in laboratory and clinical contexts.
Keywords: Tendinopathy, bioengineering, pathophysiology, scaffolds, cells, cytokines
Introduction
Tendons represent dense connective tissues responsible for linking bones and muscles. Moreover, they facilitate the storage and transmission of forces to the skeletal system during movement. Tendinopathy is defined as a broad spectrum of clinical tendon disorders, which are characterised by the presence of pain, swelling, loss of function and impaired performance. It is one of the most common musculoskeletal disorders on a global scale, affecting a wide range of individuals, including athletes, workers and the elderly. Tendon rupture is the most prevalent acute tendon injury and occurs predominantly during sporting activities. Chronic tendon injuries represent a prevalent clinical problem, primarily attributable to repetitive mechanical loading and advanced age.1 Individuals afflicted with tendinopathy encounter a diminished quality of life, a condition precipitated by discomfort and functional impairment.2 The healing response following tendon injury is often delayed and ineffective due to the hypocellular and oligovascular nature of tendon tissue. Some theories have been advanced in an attempt to elucidate the mechanism of tendon healing failure. The earliest theory proposed that mechanical factors lead to insufficient tendon repair and progressive cell death after injury to the tendon, which promote degeneration of the tendon. It has been hypothesised that inflammatory factors play a role in the development and progression of tendinopathy. It is hypothesised that vascular ingrowth and neurogenic inflammation may also be contributing factors to the development of pain and functional impairment in the affected tendon.3 Currently, common treatments for chronic tendon injuries include immobilization, physical therapy, nonsteroidal anti-inflammatory drugs and local glucocorticoid injections. On the other hand, tendon ruptures generally necessitate surgical intervention. Whilst the aforementioned treatments have been shown to enhance the function of the diseased tendon and alleviate pain, it remains challenging to restore the highly organised extracellular matrix of the tendon to its pre-injury state. This challenge gives rise to the possibility of re-injury, and the repaired tendon tissue is often unable to achieve the same level of function as it did prior to injury.4,5 By combining cell biology and materials science, tendon tissue engineering is expected to be a promising therapeutic strategy to promote injury tendon repair. Materials commonly used in tendon tissue engineering to construct scaffolds, biological factors that can be incorporated into the scaffolds and bioengineering strategies that can improve the properties of biomaterials to fit the specific structure of the tendon will be described later, respectively.
The Composition and Structure of Tendon
Tendons connect muscle to bone and are important organs for transmitting force in the musculoskeletal system. Tendon is a highly organized low-cell connective tissue composed of highly oriented fibers formed in a layered structure. Tropocollagen, a triple helical peptide chain, accumulates into microfibrils. These microfibrils then form collagen fibrils, which then form subfibres. The final step in this process is the formation of fibres, which then form fascicles. The fascicles are wrapped by the epitenon, forming form the tendon. When epitenon extends between the fascicles, it is called the endotenon. The endotenon is a thin mesh of connective tissue covering each fascicle. The outer tendon is a loose sheath of connective tissue containing the tendon’s vascular, lymphatic, and nerve supply. The connective tissue also reduces friction during movement6,7 (Figure 1).
 |
Figure 1 Composition and structure of tendon. TDSCs from three sources and tendon ECM constitute the tendon microenvironment together. The tendon ECM is mainly composed of type I collagen and type III collagen. Other components help maintain the homeostasis of the tendon microenvironment. |
The tendon is a kind of oligocellular tissue, mainly composed of type I collagen and water with very few cells. The extracellular matrix constitutes a significant component of tendon. The dry weight of tendon constitutes approximately 30% of its total mass, with collagen accounting for 70% to 80% of this dry weight. The predominant types of collagen in tendon include type I and type III.8 Type I collagen constitutes 95% of the total collagen in tendon. The tendon’s triple-helix structure consists of two α1 chains and one α2 chain, organised into a highly hierarchical arrangement. This structural organisation enables the tendon to stretch and absorb forces in response to mechanical stimuli.9 Type II collagen is present in low concentrations in tendons and is found in high concentrations in the area surrounding the tendon-bone junction. Type II collagen may help maintain the flexibility and resistance of tendons after injury.10 Type III collagen has been shown to produce fibres of a smaller size and lower organisation. It is imperative for the alignment and ductility of the initial collagen fibrils.11 Type IV collagen is found mostly in the basement membrane of tendon vessels.12 Type V collagen constitutes the fundamental structural element of type I collagen fibrils, serves as the template for their formation, and plays a pivotal role in regulating fibre diameter.13 Type VI collagen may be involved in the attachment of cells to extracellular matrix (ECM) molecules. Type X collagen is abundantly found in fibrocartilage, tendon-bone junctions. Collagen types XII and XIV play a role in the regulation of collagen fibrils ogenesis by providing specific molecular bridges between collagen fibrils and other matrix components. Collagen type XII stabilizes collagen fibrils structure during development or healing, whereas collagen type XIV restricts the diameter of collagen fibrils.14 Elastin fibers are located within the collagen fiber network and comprise approximately 1–2% of the dry mass of the tendon, allowing tissue repair after ligament stretch and deformation under loading.15
Tenoblasts and tenocytes are responsible for 90–95% of the cellular component of tendons and are arranged along the long axis of the tendon. In addition to the aforementioned cells, the presence of synoviocytes, chondrocytes, endothelial cells, pericytes, and tendon-derived stem cells (TDSCs) has also been identified. Tenocytes are a type of fibroblast that is found in the tendon. These cells are specialised in maintaining and repairing the tendon, and they can respond to external signals in the environment. This response involves the remodelling of the microenvironment according to stress and strain. The process of remodelling is mediated by matrix metalloproteinases (MMPs) and tissue inhibitor of matrix metalloproteinases (TIMPs).16–18 Tenoblasts cells are defined as immature tendon cells that retain the capacity for division, thereby enabling the repair of minor tissue damage. It has been established that both tenoblasts and tenocytes possess the capacity to synthesize collagens and other ECM components, in addition to being able to catabolize metabolic proteins. Therefore, both play an important role in the assembly and remodeling of ECM.19 TDSCs have colony-forming ability, self-renewal capacity and multidifferentiation potential. They can differentiate into tendon cells, adipocytes, chondrocytes, and osteoblasts.20 The absence of specific markers in TDSCs complicates the determination of their precise origin and location. Furthermore, given the disparities in cell source and cell isolation procedures, there is a divergence in the expression of markers for TDSCs.21 An early study proposed CD44, CD146 and Stro1 as surface markers of TDSCs.22 Nestin was identified by scRNA-seq as a key marker of TDSCs. Nestin+ cells have been identified as playing a pivotal role in the endogenous tendon injury repair process. Inhibition of nestin expression has been demonstrated to impede tendon healing, resulting in a randomised arrangement of cells and misalignment of collagenous progenitor fibres.23 The cell population of Tppp3+ Pdgfra+ was defined as TDSCs by Tyler Harvey et al. They demonstrated by inducible lineage tracing that Tppp3+ cells can generate new tendon cells and self-renew upon injury.23 Walia et al summarized three potential sources of TDSCs: tendon bundles, epitendon and perivascular.24
The non-collagenous ECM of tendons consists of proteoglycans (PGs), glycosaminoglycans (GAGs) and glycoproteins.3 Proteoglycans are the most abundant nonfibrous proteins in tendons. Proteoglycans are usually divided into large proteoglycans such as versicans, aggrecans and small leucine-rich proteoglycans (SLRP). Among them, such as versicans and aggrecans, provide resistance to compression by increasing the water content of the tendon and facilitate the diffusion of nutrients and metabolites. Decorins, as the main proteoglycans of tendon, belong to SLRP, which also includes biglycans, lumicans and fibromodulin. SLRP are involved in stabilising collagen fibrils and regulating the diameter of collagen fibrils during tendon development. Biglycans and lumicans play a major role at the early stages, while decorins and fibromodulin play a major role at the later stages. Decorins and biglycans may have complementary roles.14 Glycoproteins mainly include fibronectin, laminin, platelet-responsive proteins, lubricin and tendonogenic protein.25,26 Fibronectin and laminin mostly assist in the connection of the vessel wall and basement membrane to other components.27 It is evident that platelet-responsive proteins (TSPs) play a pivotal role in the regulation of vascularization. These proteins have the capacity to influence the interaction between cells and proteins, as well as the interaction among proteins within the ECM.28 It has been hypothesised that lubricin may promote inter-bundle sliding, and that tendonogenic protein C is involved in tendon adaptation to compressive mechanical forces.29 Tendon regulatory proteins are markers of mature tendon cells.30 Hyaluronic acid (HA), an integral component of GAG, has been demonstrated to enhance tendon cell activity and adhesion, in addition to regulating ECM synthesis and proliferation.31 Recent findings have revealed that, in a manner analogous to that observed in hard matrices, stem cells exhibit a propensity to differentiate towards the bone lineage. Similarly, in soft matrices, stem cells demonstrate a tendency to differentiate into adipocytes. The stiffness of the ECM in tendons has been shown to play a crucial role in the tendonogenic differentiation of stem cells.32 It is evident that the composition and structure of the ECM provide a biomechanically active physical scaffold for tendons to perform the mechanical function of force transmission. In addition, the biochemical microenvironment that is facilitated by the ECM is conducive to tissue development, healing and regeneration, thereby ensuring tendon biological function and maintaining homeostasis.33
Risk Factors for Tendinopathy
Among the extrinsic factors that have been demonstrated to be significantly associated with tendinopathy, the deleterious effect of high loading or the application of relatively small loads over many repetitive cycles on tendons has been found to be of particular significance. This has been observed in professional athletes participating in sports such as running, basketball, field hockey and volleyball.34–36 Tennis elbow is more common in tennis players. The overall incidence of tennis elbow has been reported to be anywhere from 35% to 51%.37,38 Long-term highly repetitive movements or poor workplace ergonomics at work are also high-risk factors for tendinopathy. Rates as high as 18% and 41% have been reported in spine surgeons and coal miners, respectively.39 The use of statins, fluoroquinolones and hormone replacement therapy have negative effects on tendons.25
Intrinsic factors, including age, nutrition, anatomical abnormalities, muscle weakness and high body mass index (BMI), are significantly associated with tendinopathy.40 There are reports suggesting that metabolic disorders, including obesity, diabetes mellitus, hypercholesterolemia and hyperuricemia, may potentially result in tendon damage.41,42 Genetic factors have been demonstrated to play a role in the development of tendinopathy. Variants in the genes TNC and COL5A1, which encode important structural components of tendons, have been shown to be contributing factors in the development of Achilles tendinopathy.43,44
The Pathophysiology of Tendinopathy
The healing process after a tendon injury ideally involves three successive, yet overlapping, stages. ① During the inflammatory phase, the initial response to injury is characterised by the influx of inflammatory cells into the affected area. Within the initial 24-hour period, monocytes and macrophages were predominant, engulfing necrotic tissue. Tendon-derived cells migrate to the wound, and the tissue begins to synthesize type III collagen.45 Many chemokines are released to mediate the inflammatory response, stimulate the proliferation of fibroblasts and tendon cells, and stimulate the angiogenesis process.7 ② During the proliferation phase, the accumulation of fibroblasts and type III collagen reached the peak, and the concentration of water and glycosaminoglycan in the tissue remained high.45 ③ During the remodelling phase, collagen fibres began to orient longitudinally along the tendon’s long axis. The ratio of type III to type I collagen, collagen cross-linking, and concentrations of glycosaminoglycans, water, and DNA returned to normal levels.46 MMPs have been identified as pivotal factors in the process of tendon remodelling. MMP-9 and MMP-13 have been shown to be exclusively involved in collagen degradation, while MMP-2, MMP-3 and MMP-14 have been demonstrated to be involved not only in collagen degradation, but also in collagen remodelling.47 However, the process of tendon self-repair can take years, and the repaired tissue has weaker biomechanics than the original, uninjured tendon.48 To prevent tendon injury and promote repair, it is important to understand the factors associated with tendinopathy. The following will introduce the various factors that influence the development and progression of tendinopathy (Figure 2).
 |
Figure 2 The main signaling pathways leading to the pathogenesis and progression of tendinopathy. Abbreviations: PGE2, Prostaglandin E2; cAMP, cyclic Adenosine Monophosphate; PKA, Protein Kinase A; CEBP, CCAAT/enhancer-binding protein; IGF-1, Insulin-like Growth Factor 1; CREB, cAMP-response element binding protein; IL-1β, Interleukin-1 beta; ERK1/2, Extracellular-regulated kinase 1/2; BMP2, Bone Morphogenetic Protein 2; Smad, Sma and Mad related protein; PPARγ2, Peroxisome Proliferator-Activated Receptor gamma 2; TGFβ1, Transforming Growth Factor Beta 1; mTORC1, mechanistic target of rapamycin complex 1; NFκ B, Nuclear Factor kappa-B; Wnt5b, wingless-type MMTV integration site family, member 5B; JNK, c-Jun N-terminal kinase; Wnt5a, wingless-type MMTV integration site family, member 5A; RhoA, Ras homolog gene family member A; ROCK, Rho-associated coiled-coil containing protein kinase; CXCL13, C-X-C motif Chemokine Ligand13; CXCR5, C-X-C chemokine receptor type 5; HMGB1, High Mobility Group Box 1; RAGE, Receptor for Advanced Glycation End Products; β-Catenin, Catenin beta; EGR1, Early Growth Response 1. |
Inflammation and the immune system. A study has revealed significant infiltration of mast cells and macrophages in samples of early human tendinopathy.49 Macrophages play a key role in regulating inflammation and tissue repair.50 A recent spatial transcriptomics study demonstrated that the increased presence of macrophages in the diseased tendon resulted in alterations to the microenvironment of TDSCs, thereby promoting their differentiation into bone and cartilage formation. Concurrently, this process impeded the tendon’s normal healing process.51 Macrophages have been observed to promote cell proliferation and collagen deposition; however, their presence may also result in a reduction in the ultimate tensile strength of the Achilles tendon.52 Macrophages are now generally classified into two distinct types: M1-type and M2-type macrophages.53 Despite the fact that certain researchers have utilised single-cell analysis to demonstrate the existence of numerous subtypes of macrophages, the majority of studies in the domain of tendon bioengineering tend to simply divide macrophages into M1 and M2 subtypes.54,55 The initial phase of tendinopathy is characterised by the infiltration of M1 macrophages. It was observed to be at a higher concentration within 14 weeks, up to 18 times the normal concentration. The M1 macrophage phenotype has been observed to engulf debris and apoptotic cells during the early phase of injury. However, this process has also been shown to result in the damage of surrounding healthy tissues and the release of pro-inflammatory cytokines, such as IL-1, IL-6, IL-12 and tumour necrosis factor-α (TNF-α).56–59 The M2 macrophage phenotype manifested predominantly in the middle and late stages of tendon injury, which was observed to increase significantly after 28 days of injury.56 The M2 macrophage phenotype has been shown to secrete TGF-β1, a key factor in promoting the cartilage differentiation of mesenchymal stem cell (MSC), thereby enhancing the tendon-to-bone healing process. However, it is important to note that TGF-β1 also plays a role in the formation of tendon adhesion by recruiting MSC to form fibroblasts. The secretion of TGF-β1 by the M2 macrophage phenotype also contributes to the termination of the inflammatory response during tendon healing.60–62 The secretion of exosome by M2 macrophage phenotype has been demonstrated to stimulate FAP differentiation, thus reducing excessive fibre deposition and fat accumulation.63 Despite the capacity of M2 macrophage phenotype to stimulate the synthesis of new matrix, the resulting tissue exhibits disparities in terms of structure, composition and material properties when compared to normal tendons.64 The presence of mast cells in the injured tendon, in conjunction with their capacity to secrete nerve growth factor (NGF), has the potential to induce the growth of nerves into the healing tendon. This process, in turn, can result in the occurrence of neurogenic inflammation and the subsequent secretion of vascular endothelial growth factor (VEGF), which plays a pivotal role in the promotion of neovascularization.65 Alim hypothesised that, following a tendon injury, mast cells receive transmitters secreted by nerve endings. These mast cells then secrete mediators that can further activate neurons, creating a cycle that leads to persistent inflammation and pain in tendinopathy. The glutamate signalling process may play an important role in this cycle.66 A number of inflammatory factors have also been identified as contributing to the development of tendinopathy. An earlier study mentioned that pro-inflammatory factors secreted by infiltrating macrophages in diseased tendon tissue may lead to matrix destruction by expressing and accumulating MMPs. TNF-α promotes the production of PGE2, MMP-1, MMP-3, MMP-8 and MMP-13 by activating the IL-21 signalling pathway. This leads to pain and ECM breakdown.67 Some studies have observed the upregulation of IL-17 and IL-33 in tendinopathic tissues, while IL-17 and IL-33 increased expression of type III collagen, which may inhibit the conversion of type III collagen to type I collagen.68,69 It is thought that the upregulation of TGF-β1 in diseased tendons promotes the upregulation of plasminogen activator inhibitor-1 (PAI-1). PAI-1 inhibits the action of tissue-type plasminogen activator (tPA) and urokinase plasminogen activator (uPA), which leads to scar formation in the tendon.70
Mechanical injury. Theories positing the primary cause of tendon injury include the accumulation of micro-damage in collagen fibres of the tendon, precipitated by excessive utilisation or recurrent stretching of the tendon during exertion or physical activity. The up-regulation of MMPs has been identified as a potential contributor to collagen fibre degradation. The formation of micro-damage may induce tendon cells to release MMPs, which further degrade the tendon matrix, thus preventing effective repair of the micro-damage.71–73 A report mentions that patients with tendinopathy often make inadequate adjustments to their mechanics in order to maintain function and minimise pain. Examples include reducing ankle dorsiflexion and limiting ankle and foot activity. These functional biomechanical defects lead to excessive, repetitive overload of the Achilles tendon, which further aggravates tendinopathy.74 C S Bestwick et alproposed that the maximum tensile load of the tendon may result in ischemia, and that the subsequent recovery of normal perfusion may enhance the production of reactive oxygen species (ROS). High temperature during tendon movement may also stimulate mitochondria to produce ROS, damage and induce apoptosis of TDSCs.75 It has been previously documented that both the synthesis and degradation of tendon collagen are elevated following exercise, with collagen degradation being predominant during the initial 24–36 hours and synthesis being predominant during the subsequent 36–72 hours. It has been demonstrated that repetitive training in which the rest period is insufficient can result in a net degradation of the tendon matrix. This, in turn, can lead to tendon injury.76 The occurrence of acute tendon rupture and chronic tendinopathy is greatly increased by the accumulation of micro-damage to tendon collagen fibres.77,78 Overloading may also lead to differentiation of TDSCs in the non-tenogenic direction as well as degenerative changes that impede tendon healing, which may be induced by high levels of PEG2, while Chen et al suggested that overload-induced calcific tendinopathy is regulated by the mTORC1 signaling pathway.79,80 It cannot be ignored that some studies have also suggested that appropriate mechanical stimulation can help heal tendon injuries.81
Metabolic factors. Diabetic patients tend to be in a pro-inflammatory state, which will likely cause chronic inflammation leading to tendinopathy. Kwan et al mentioned that the pro-catabolic response of healthy tendon cells is attenuated in high glucose environments, which may lead to inflammation after prolonged exposure of tendon tissues to microinjuries and result in degenerative tendon changes.82 Within a high glucose milieu, the proliferation of TDSCs is suppressed, while apoptosis is elevated and cellular autophagy is impeded.83–85 An earlier study noted that type III collagen expression was downregulated in injured tendons of diabetic rats, which may lead to impaired tendon tissue healing and reduced biomechanical properties.86,87 In tendons from diabetic patients, accumulation of advanced glycosylation end products(AGEs) in the ECM of tendon cells impedes collagen fiber slippage and promotes aging with heterotopic ossification of tendon tissue.88,89 Shivam et al found that AGEs inhibit mitochondrial function of tendon cells and inhibit their proliferation, limiting the ability of tendon cells to synthesize the ECM and disrupting tendon homeostasis.90 AGEs also increased ROS expression.91 The accumulation of ROS will interfere with cell function, leading to the loss of self-renewal ability and stemness of TDSCs. Furthermore, it is predicted that this process may also induce apoptosis or promote osteogenic differentiation of TDSCs.92–94 And the damaged tendon has a lower resistance to oxidative stress relative to the healthy tendon, which may make the damaged tendon more susceptible to ROS damage.95 Hypercholesterolemia also reduces tendon biomechanical properties and impairs healing by upregulating pro-inflammatory factors and interfering with MMPs.96
Degenerative changes. Recent research has revealed that damaged fetal tendons of sheep have the capacity for complete regeneration. Protein analysis has demonstrated that fetal secreted proteins exhibit a substantial discrepancy with those of adult sheep. Furthermore, the inflammatory response in adult sheep tendons is predominantly oriented towards defence, as opposed to tissue remodelling, in comparison to that observed in the fetus.97 Kristen showed that SCX-profile cells in neonatal mouse tendons play a key role in the post-tendon injury. Whilst complete functional recovery and tendon-specific differentiation are of key importance, it has been demonstrated that SCX lineage cells in adults differentiate towards cartilage after injury to the tendon. Furthermore, in the absence of tendon cells, exogenous αSMA cells will persist and cause scarring.98 With aging, tissues in adult individuals gradually lose the possibility of complete regeneration after tissue injury.98 In addition to replicative senescence, persistent inflammation has also been demonstrated to induce senescence. This may be due to decreased immune function, increased oxidative stress and focal death due to inflammation.99–102 Kannus et al conducted a study of tendon specimens from 891 patients, revealing that 97% of the subjects exhibited degenerative changes.103 Concurrently, the mRNA expression levels of SCX and TNMD in degenerative tendons were significantly reduced, and the distribution of type I collagen was reduced while the distribution of type III collagen was relatively increased, which could lead to the down-regulation of biomechanical properties of tendons.104,105 Disruption of tendon microstructure also increases with aging.106 The above factors lead to a greater tendency for tendon microdamage and tendinopathy. Cellular senescence produces a senescence-associated secretory phenotype (SASP), which encompasses a range of interleukins, inflammatory factors, chemokines, protease and growth factors such as IL-6, IL-8, and CXCL1, etc. The SASP contributes to the maintenance of cellular senescence, with evidence suggesting a role for autocrine and paracrine functions in this process. For instance, the presence of IL-6 and IL-8 has been demonstrated to mediate cell cycle arrest.107 The family of cytokines known as transforming growth factor-beta (TGF-β) has been observed to induce normal cellular senescence via paracrine secretion.108 SASP also promotes chronic inflammation, induces inflammatory infiltration of tendons and leads to senescence of immune cells, which diminishes the body’s ability to regulate inflammation and clear senescent cells.109 Aging also induces senescence and depletion of the stem cell pool. The proliferative and migratory capacities of senescent TDSCs are down-regulated, which may lead to impaired tendon healing.110 Senescence of TDSCs also leads to up-regulation of aberrant differentiation and inability to maintain their differentiated state, which may affect the healing of tendon tissue.111,112 A study revealed that, in the aftermath of tendon injury, adipocyte infiltration around the injured tissue manifested at a higher frequency in aged rats than in young rats.113 A number of studies have observed that aging tendons appear to accumulate more fat.114–116 However, one study found that senescence inhibited the lipogenic differentiation of TDSCs by down-regulating PPARγ signaling, which may represent that fat accumulation in senescent tendons is not associated with abnormal differentiation of TDSCs.117
The combination of these reasons, as well as genetic factors and drug use interferes with the healing of tendon tissue, making it difficult to convert the damaged tendon tissue from a randomly arranged type III collagen morphology to a regularly arranged type I collagen morphology, and preventing the restoration of biomechanical and tissue-secondary morphology in healthy tendons.118
Current Clinical Therapy for Tendinopathy
The prevailing clinical approach to tendinopathy is currently focused on the management of symptoms, with surgical intervention typically reserved as a definitive measure. The utilisation of non-steroidal anti-inflammatory drugs (NSAIDs) is conventionally employed to alleviate short-term (7–14 days) pain arising from tendinopathy.119–121 Corticosteroids injection has been utilised in the treatment of tendinopathy for decades. Its primary function is the alleviation of short-term symptoms, with a duration of efficacy spanning up to approximately six weeks.122,123 Furthermore, PRP injection has been observed to contribute to the alleviation of pain.124 In addition, topical glyceryl trinitrate treatment has been demonstrated to be efficacious in the management of short-term pain (6 months) in patients diagnosed with tendinopathy. A subsequent follow-up investigation revealed that the beneficial effects of the treatment were sustained for a period of three years in patients who had previously undergone local trinitrate treatment. This finding suggests that topical glyceryl trinitrate treatment may facilitate tendon healing in addition to providing analgesia.125,126 Tendon injury has been observed to be accompanied by the growth of new blood vessels. As demonstrated by clinical trials, the injection of sclerosing polidocanol into the neovascularization area can be an effective treatment.127,128 The utilisation of physical therapy methods, including ultrasound treatment, eccentric exercise, low-intensity laser therapy, shock wave therapy and low-frequency electrical therapy, is frequently employed to alleviate symptoms. However, the efficacy of these methods remains frequently ambiguous.129–135
It is evident that existing therapies are accompanied by a certain degree of adverse effects. For instance, the utilisation of NSAIDs has been demonstrated to elevate the likelihood of gastrointestinal complications.136 It has been documented that the utilisation of corticosteroid injections may result in Achilles tendon rupture. It is not considered to be the case that corticosteroids injected into the affected area can help reduce the risk of further surgery being required.137–139 In cases where non-surgical treatment has failed, surgical intervention is considered a last resort for the management of tendinopathy. Surgical interventions for tendinopathy encompass the excision of the degenerative tendon, the removal of adhesions surrounding the tendon, the decompression of the tendon and the implementation of multiple longitudinal tenotomies. However, it should be noted that surgical procedures can often be challenging to achieve patient satisfaction.136,140,141 In summary, bioengineered scaffolds that can provide the appropriate biomechanical strength and biological signals to guide the repair process within a short timeframe after injury represent a new potential treatment.
Commonly Used Carrier and Scaffold Materials
The bioengineering of tendon is based on the development of biomaterials and biological factors. They are integrated by various bio-engineering technologies to better repair the damaged parts of the body(Figure 3). The biomechanical strength of a scaffold is an important factor in its application to tendon repair. The uniaxial tension method is generally used in the study to obtain the Young’s modulus and stiffness of the scaffolds. The ultimate tensile strength and maximum load are obtained by applying uniaxial tension until fracture occurs.142 The performance of bioengineering carriers or scaffolds varies according to the properties of the materials they are made from. The following summarises the advantages, disadvantages and application directions of the materials commonly used in various tendon bioengineering, divided into natural and synthetic materials (Table 1).
 |
Figure 3 Biomaterials, biological factors and bioengineering techniques commonly used in tendon bioengineering. (a) Biomaterials commonly used in tendon tissue engineering. (b) Biological factors commonly used in tendon tissue engineering. (c) Bioengineering techniques commonly used in tendon tissue engineering. Abbreviations: PGA, polyglycolic acid; PLA, Polylactic Acid; ADSCs, adipose-derived stem cells; EVs, extracellular vesicles; BMSCs, bone marrow derived mesenchymal stem cells; TDSCs, tendon-derived stem cells; PRP, platelet rich plasma; PDGF, platelet-derived growth factor; VEGF, vascular endothelial growth factor; IGF, insulin-like growth factor; TGF-β, transforming growth factor β; BMP, bone morphogenetic protein; FGF-2, basic fibroblast growth factor. |
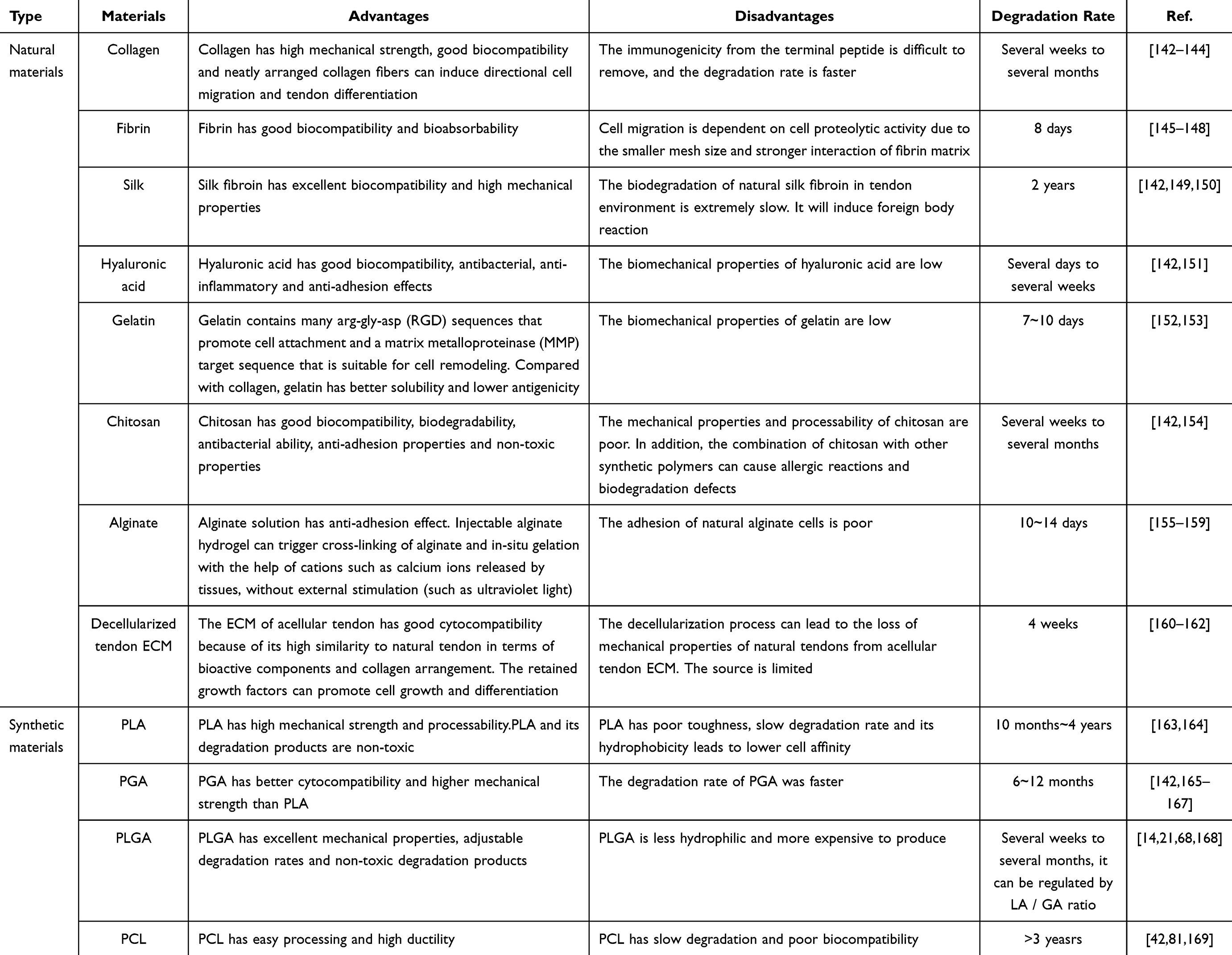 |
Table 1 Advantages and disadvantages of natural materials and synthetic materials |
Natural Materials
Natural biomaterials such as collagen, fibrin, silk and HA are widely used as carriers and scaffolds in tendon tissue engineering because of their high biocompatibility and promotion of cell attachment and differentiation.
Collagen
Collagen is the most abundant protein in the human body and type I is the most abundant and studied for biomedical applications. Collagen is the main component of tendons and therefore the first natural material used to make tendon scaffolds. It has good cellular and cytokine binding sites, which support cellular activity. Uniaxial stretching of collagen scaffolds that have been mechanically stimulated and treated promotes tendon regeneration.170 The most commonly used collagen is derived from animal tissues. To ensure immunogenicity, the collagen must be physically or chemically cross-linked to remove antigens and pathogens. This process weakens the collagen’s mechanical properties and accelerates its degradation.171 Therefore, it is necessary to blend type I collagen with other synthetic or biopolymers to enhance its mechanical strength.172 Qian et al utilized collagen in combination with filipin to create scaffolds in a rabbit tendinopathy model, which ensured the mechanical properties of the scaffolds and improved healing at the tendon-bone interface.173 In addition to producing hybrid scaffolds, studies have developed electrochemically aligned collagen. This process uses an electric field to arrange collagen molecules in an orderly fashion, thereby enhancing the material’s mechanical properties to a level comparable with that of normal tendons.174 Collagen can also be used to coat the scaffolds to increase their biocompatibility. Yuan et al produced poly(lactic acid) (PLA) scaffolds with collagen and chondroitin sulphate coatings using electrostatic spinning. These scaffolds exhibited excellent alignment and tensile mechanics, enabling them to better mimic natural tendons. Furthermore, the human bone marrow mesenchymal stem cells on the scaffolds demonstrated a higher rate of cell spreading and proliferation. The expression of SCX and TNMD also appeared to be increased175 (Figure 4a). Some studies have also explored sources of collagen other than animal tissue. Dafna et al attempted to increase the yield of collagen fibres by extracting them from coral tissue. They implanted the extracted collagen fibres into alginate hydrogel and experiments in rats revealed that the scaffold was biocompatible, safe and promoted tissue repair.57
Figure 4 Continued. Figure 4 Application of common biomaterials in tendon regeneration. (a) PLA scaffold coated with collagen and chondroitin sulfate is used to repair rat Achilles tendon; *p <0.05, **p < 0.01, “normal” refers to native rat Achilles tendon. (b) The nanofiber scaffold combined with GelMA and SF promoted tendon tissue regeneration in vivo; SG: SF+GelMA; the dotted squares indicate the lesion area; **p<0.01. (c) PLA-dicumarol conjugates-electrospun nanofiber membrane (ENM) (PCD) reduces the deposition of collagen type III through the Cx43/TGFβ/Smad3 signalling pathway and alleviates peritendinous adhesions; *p<0.05, **p<0.01, ***p<0.001. (d) Alkaline chitosan-coated PLGA significantly reduced the inflammatory response in vivo caused by degradation products; *p<0.05, **p<0.01. Reproduced with permission.

Fibrin
Fibrin is a biopolymer similar to collagen. It is highly biocompatible, but has poor mechanical strength. It is therefore usually used in combination with other, stronger scaffolds.166 Zhao et al found that knitted poly(L-lactic-co-glycolic acid) (PLGA) has a porous structure that allows tissue to grow inwards, and that its mechanical strength compensates for the shortcomings of fibrin in this respect. They therefore incorporated fibrin gel into knitted PLGA scaffolds, which effectively promoted cell attachment and growth within the scaffolds, as fibrin can serve as an effective delivery matrix.146 It has been mentioned that fibronectin gels exhibit better biological, structural and mechanical properties in cell-based tendon tissue engineering constructs compared to collagen gels, and may be a better alternative to collagen in improving the biocompatibility of scaffolds.176 More recent studies have explored the use of platelet-rich fibrin in tendons. This material has a denser fibrin network and a slower degradation rate. It also contains growth factors that may benefit cell proliferation, inflammation and ECM deposition.177 Daigo et al used platelet-rich fibrin scaffolds in a rabbit model to repair patellar tendon defects. The ultimate failure load and stiffness of the scaffold group were found to be relatively higher after 12 weeks, and the ultimate breaking load of the repair group reached 78% of that of the healthy control group of the same age at 20 weeks.178 Wong et al prepared a platelet-rich fibrin gel and implanted it into an Achilles tendon defect in rabbits. This was found to improve tendon healing and collagen deposition in the repaired tissue, as determined by imaging and histological observations. Imaging and histology revealed that the scaffold improved tendon healing, and that the collagen fibres of the repaired tissue were cross-linked and aligned along the long axis of the tendon.179
Silk
Silk is one of the strongest fibrous proteins in nature and consists of 60–80% of sericin and 15–35% of sericin. Compared with other natural materials, sericin has high tensile properties, and it possesses excellent biocompatibility, adjustable biodegradability, and mechanical properties.180 Chen et al utilized sericin to fabricate well-arranged scaffolding structures. The ordered scaffolds promoted the proliferation and tendon-like differentiation of the attached stem cells.181 Compared with pure sericin scaffolds, incorporating gelatin methacryloyl (GelMA) into sericin scaffolds enhanced cell attachment and tendon repair. It also promoted the proliferation of MSCs, the production of vascular endothelial growth factor (VEGF) and the expression of tendon-related genes. This provides an advantageous microenvironment for stem cell growth182 (Figure 4b). As a bioscaffold material, sericin proteins exhibit excellent mechanical strength and high thermal stability. They can also improve the adhesion and proliferation of human fibroblasts. Furthermore, sericin proteins possess inherent tendinopathy repair properties. Following the local injection of an aqueous filaggrin solution into a rat model of tendinopathy, filaggrin was found to promote tendon injury repair by enhancing tendon cell activity and proliferation, upregulating the expression of SCX, TNMD, TNC and core proteoglycans, enhancing collagen production and attenuating oxidative damage. Filaggrin was also found to possess anti-nociceptive and anti-allergic effects.183
Hyaluronic
Hyaluronic acid (HA) is a glycosaminoglycan mainly found in the ECM. HA has unique viscoelasticity and good biocompatibility and non-immunogenicity, and it has also been suggested that it has some anti-inflammatory properties.184,185 HA can enhance the activity of tendon-derived cells and significantly induce the expression of type I collagen, without increasing the expression of type III collagen.186 HA degrades in a very short time. Therefore HA requires chemical modification to enhance biodegradability.187 Honda et al tried direct injection of HA into completely torn rabbit infraspinatus tendon. Four and eight weeks after surgery, the ultimate load-to-failure was significantly higher in the HA group than in the control group (45.61 ± 9.0 N vs 32.42 ± 9.4 N at 4 weeks, 90.7 ± 16.0 N vs 66.97 ± 10.0 N at 8 weeks).188 Many applications of HA have been motivated by the anti-adhesion effect of tendons. Chen et al suggested that mixing HA with platelet-rich plasma and adding HA to the anti-adhesion membrane could reduce adhesions by decreasing the entry of foreign fibroblasts into the tendon injury. In this study, functional assays and histological observations revealed that the composition of HA mixed with platelet-rich plasma at a certain ratio assisted by nanofibrous core shells reduced tendon adhesion formation and inflammation and promoted tendon healing.189 Miescher et al implanted HA into DegraPol scaffolds and found that, compared to scaffolds without HA, adhesion was significantly reduced by around 50% after HA was added, converging with healthy tendons.190
Gelatin
Gelatin is a naturally occurring polymer produced by the partial hydrolysis of collagen. It is less antigenic than natural collagen and has modifiable functional groups due to the absence of specific amino acids, such as tyrosine, tryptophan and phenylalanine, which mediate immunogenic responses.191 As a product of denatured collagen, gelatin resembles natural collagen chains, and its lower cost compared to purified collagen, as well as its greater stability to organic solvent solubility, are properties that make gelatin suitable for industrial-scale scaffold manufacturing.192 However, like collagen, gelatin has the disadvantage of weak mechanical properties.193 In tendon tissue engineering, gelatin can be used to coat scaffolds or be composite-processed with scaffold materials. This improves the biocompatibility of the scaffold, supports the delivery of cells and proteins, and promotes collagen production and reduces inflammation, facilitating tissue healing.194 A study by Lim et al suggests that mixing polycaprolactone (PCL) with gelatin and using the electrostatic spinning technique produces aligned nanofibres with properties similar to natural tissue collagen, as well as diameters similar to the collagen protofibres found in natural tissues. This improves the biocompatibility of the resulting scaffold.191 Gelatin provides a cell adhesion matrix when compounded with a bioinert hydrogel system.195 Tough hydrogels are hydrogels that sacrifice biocompatibility and degradation rates for improving mechanical properties and are often difficult to apply directly in bioengineering. Yuan et al prepared gelatin-based tough hydrogels into which gelatin was implanted and excellent biocompatibility, in vivo self-degradation, and resemblance to natural tissue constituents were obtained.196 The gelatin derivative GelMA hydrogel is a commonly used material for tendon bioengineering. The biological properties of GelMA are favourable for cell attachment, proliferation and spreading. The photocrosslinking properties of GelMA also enable microfabrication of the hydrogel, producing a unique morphology and 3D structure.152 Yang et al integrated GelMA into PCL scaffolds to address the poor bioactivity of PCL scaffolds due to their lack of cell attachment and tissue integration.192 Another study found that encapsulating PCL scaffolds within GelMA hydrogels provided a suitable microenvironment that enhanced the tendon differentiation of TDSCs.197
Chitosan
Chitosan (CS) is derived from chitin, the second most abundant natural polysaccharide in the shells of organisms such as crabs, lobsters, turtles, shrimps, and insects. CS is a natural material that is biodegradable, non-toxic and biocompatible. It has potent antioxidant and antimicrobial properties, and degradation of CS results in the production of harmless aminosaccharides that can be fully absorbed by the body. CS is a cationic polymer and can therefore be combined with other synthetic or negatively charged natural materials to form composites, or with negatively charged mucins on cell membranes to promote adhesion.198,199 Hydrogels are commonly used in tendon tissue engineering. However, tendon movement can cause the hydrogel to displace and fragment. Freedman et al therefore achieved adhesion to tissues by incorporating CS on one side of the hydrogel.200 In addition to being used to promote adhesion, CS can also be used to resist tendon adhesion. The positively charged surface of CS may lead to electrostatic repulsion, and the ability of fibroblasts to adhere to the electrostatically spun nanofibrous membrane synthesized by PCL and CS was therefore reduced, thus synergizing the physical barrier effect of the nanofibrous membrane to reduce tendon adhesion postoperatively.201 The anti-adhesion effect of CS is not only due to its physical properties, but also to its biological properties that give it an anti-adhesion effect. The biological properties of CS also contribute to its anti-adhesion activity, which may be due to the down-regulation of acetylated p65 and p53 expression in tendon by increasing SIRT 1 expression.202 CS is also biocompatible. By adding CS to collagen and polycaprolactone hybrid hydrogels, the chemical structure of CS is structurally similar to glycosaminoglycans, and the combination of collagen makes the scaffolds compositionally similar to the extracellular matrix, which enhances the scaffolds’ biocompatibility and exhibits a slow degradation rate.203
Sodium Alginate
Sodium alginate (SA) is derived from brown algae found in the ocean. It has a structure similar to glycosaminoglycans and is both hydrophilic and water-soluble. It thickens under neutral conditions and forms hydrogels in the presence of multivalent cations. Therefore, it has been proposed to promote in situ gelation of injectable alginate hydrogels with the help of calcium ions released from tissues rather than through external stimulation.155 SA is important in bioengineering due to its biocompatibility as well as non-toxicity, mild gelation properties, and affordability.204 Whether SA is compounded with other materials to prepare hydrogels or SA is cross-linked with other materials and then wet-spun to construct fiber-aligned scaffolds can enhance its biocompatibility.205,206 Although SA hydrogels are beneficial in restoring the histological properties of damaged tendons, the biomechanical strength and tissue continuity that hydrogels can provide make them unsuitable for use in cases of tendon tears. Therefore, loading SA hydrogel onto PCL electrospun fibre scaffolds can optimise the hydrophobicity of PCL with the help of the hydrogel, while maintaining the biomechanical strength provided by the PCL. This is an effective solution.207 It has also been suggested that SA aqueous solution has the effect of preventing adhesions after tendon surgery.156 Jayasree et al used SA coated on scaffolds composed of PCL nanofibers and reduced protein adsorption which may lead to lower cell and fibrous tissue attachment, therefore the anti-adhesion utility of SA needs further investigation.208
Decellularized Tendon ECM
Decellularised ECM is a low-immunogenic scaffold material because cells, nucleic acids and antigens are removed, while ECM components such as collagen, elastin and proteoglycans, as well as a variety of cytokines, are retained. This reduces foreign body reactions, inflammation and immune responses after implantation.209 Natural decellularized tendon ECM is a suitable option for tendon defect reconstruction. A key advantage of using natural decellularized tendon ECM for repair is that the typical arrangement of type I collagen fiber bundles in decellularized tendon ECM promotes stem cell recruitment, tendon differentiation of precursor cells for tendon repair.210 Tao et al prepared dense tendon anti-adhesion membranes using decellularised bovine tendon-derived ECM. These membranes reduced tendon adhesions when applied to injured tendons and promoted tendon cell proliferation. This may be due to the release of growth factors that were retained during processing.211 Nodoka et al mentioned that the use of decellularized ECM from tendon-muscle junctions(TMJ) induced MSCs to express TMJ marker genes and proteins.212 It was also found that loading senescent TDSCs into young decellularized tendon ECM decreased the senescence-associated β-galactosidase activity of senescent TDSCs and upregulated the expression of stem cell markers.213 However, the dense collagenous structure of the decellularised ECM in tendon tissue hinders cellular infiltration. It is therefore possible to dissolve the ECM to form a solution and combine it with a variety of biomaterials. This compensates for the lack of mechanical properties of the ECM by combining it with biomaterials that have high mechanical strength.214
Gene Vectors
Viral vectors, such as adenoviruses, adeno-associated viruses, retroviruses and lentiviruses, are the most commonly used vectors for gene delivery. These vectors offer high transfection efficiency and stable gene expression.215 Patrick Bolt et al used an adenovirus vector to deliver the BMP-14 gene to a rat model of an Achilles tendon tear. Two weeks later, there were more tendon cells in the virus-transfected healing site, and the tensile strength was 70% higher than in the control group.216 Sys Hasslund et al delivered growth and differentiation factor 5 (GDF-5) to a mouse model of flexor tendon plasty via a recombinant adeno-associated virus. In vivo experiments showed that lower doses of GDF-5 were more effective at inhibiting adhesion without affecting repair strength.217 However, viral vectors have several disadvantages, including immunogenicity, insertion mutagenesis, carcinogenicity and a limited capacity for gene packaging. Consequently, alternative delivery systems such as plasmids, exosomes, inorganic nanoparticles and liposomes have been proposed.218 Genbin Wu et aldelivered the TGF-β1 gene silencing plasmid into the chicken tendon defect model. The plasmid prevents tendon adhesion and promotes tendon function repair by silencing the TGF-β1 gene.219 Although non-viral vectors show great potential, further development is needed to overcome problems such as biological barriers and poor transfection efficiency.
Synthetic Materials
Compared to natural materials, synthetic materials have controlled degradation rates, can be fabricated into complex shapes, and offer better cell attachment, delivery of soluble molecules, and greater mechanical strength. Furthermore, synthetic materials can be produced at a low cost in large quantities and have a longer shelf life.220 But synthetic materials have limited biocompatibility and are not favorable for cell proliferation and adhesion.221,222
Polylactic Acid (PLA)
PLA is a polymer produced by the chemical reaction of biofermented lactic acid. Due to their non-toxicity, biodegradability and bioabsorbability, PLA and its copolymers have received much attention in biomedicine. PLA has a high tensile strength and Young’s modulus. It also has good flexural strength and superior cell adhesion to PGA, PLGA and PCL.8 However, the low toughness, slow degradation rate and hydrophobicity of PLA limit the use of PLA.223 Therefore PLA needs to be blended or copolymerized with components to achieve the desired properties.166 The defects caused by the hydrophobicity of PLA can be improved with the help of collagen, which is highly biocompatible. In one study, collagen yarn was composite prepared with PLA yarn, the former provided excellent biological properties to improve tissue healing and inward cell growth, while the latter provided the required mechanical properties, the hybrid scaffolds could reach a tensile strength of 354.0±36.0N. The cell coverage on the scaffolds was significantly higher than that on the pure PLA yarn, and the cell attachment pattern tended to be similar to that of the natural attachment on the ECM.224 Coating the PLA scaffolds with a chitosan-collagen hydrogel increased their biocompatibility, and tendon cells were observed to attach and spread well on the coated scaffolds. An outer alginate gel coating was also applied to prevent peritendinous adhesions and reduce protein adsorption on the coated scaffolds compared to the uncoated ones.225 Adhesions after tendon surgery can hinder tendon gliding, but anti-adhesive nanofibrous membranes can be used to improve this. Hadda et al prepared a nanocomposite membrane by blending polylactide (PLA) with polyether urethane (PEU), which has a suitable degradation rate, biocompatibility and mechanical properties. Mechanical tests showed that the copolymer membrane had suitable ductility.226 Another study used PLA coupled with biscoumarin to fabricate nanofibre membranes. This reduced tissue adhesion and fibroblast activity, inhibited TGFβ production, and had no effect on the biomechanical strength of the repaired tendon227 (Figure 4c). However, it has been suggested that the degradation of polyesters such as PLA in vivo can cause a foreign body reaction. The PLA barrier can also lead to the formation of peritendon granulomas and adhesions. Therefore, Liu et al prepared ibuprofen-loaded PLA fibrous membranes using electrostatic spinning. This enhanced the anti-inflammatory and anti-adhesion effects by reducing macrophage infiltration.228
Polyglycolic Acid (PGA)
PGA is a hydrophilic polyester with a higher mechanical strength than PLA. Its high stiffness makes it suitable for use in tendon-bone healing, and it is primarily used to make porous scaffolds and sutures. Its high initial cell growth rate, relatively fast degradation rate and high porosity favor intercellular contacts at high cell densities, which will promote high synthesis rates of extracellular matrix.165 Earlier, Yokoya et al suggested that PGA is suitable for the repair of tendon-bone junctions.229 Romeo et al developed a nanofibre scaffold polymerised using PGA and PLCL, and the ultimate load of sheep infraspinatus tendons treated with this scaffold increased significantly. After 12 weeks, the ultimate failure load was almost 75% of that of a non-surgical tendon. Collagen fibres were also observed extending to the calcified fibrocartilage area and adhering to the humerus.230 However, the high degradation rate of PGA leads to a rapid decrease in mechanical strength 2–4 weeks after implantation, and its degradation product, ethanoic acid, leads to an increase in local acid concentration, which can lead to tissue damage.166,231
Poly(L-Lactic-Co-Glycolic Acid) (PLGA)
PLGA, as a copolymer of PLA and PGA, improves control over the degradation rate by allowing customization of the degradation rate by varying the PLA:PGA ratio. However, pure PLGA has poor hydrophilic, mechanical and bioactive properties, so it is often blended with other biomaterials. Altman et al blended it with filipin proteins used to increase hydrophilicity in order to provide the necessary mechanical properties. Biological studies have shown the scaffolds to be biocompatible, resorbable and able to support inward cell growth and tissue remodeling.232 Making electrospun nanofibers from blends of PLGA and small intestinal submucosa can also improve the hydrophilicity of PLGA.233 Loading the surface of PLGA scaffolds with a hydroxyapatite mineralized coating can improve the strength and stiffness of PLGA scaffolds without compromising ductility and toughness.234 However, most biodegradable aliphatic polyesters such as PLA, PGA, and PLGA suffer from the problem of aseptic inflammation due to their acidic degradation products. Therefore, Shen et al attempted to coat alkaline chitosan on PLGA scaffolds in the expectation that chitosan would act as an acid neutralizer. It was found that although the cell adhesion ability of PLGA scaffolds decreased after the addition of alkaline chitosan, cell migration and collagen secretion could still be promoted, while at the same time the scaffolds significantly reduced the recruitment of inflammatory cells and the formation of foreign body giant cells235 (Figure 4d).
Polycaprolactone (PCL)
PCL is bioabsorbable and nontoxic to cells or organisms and unlike PGA and PLA, PCL has high ductility but low tensile strength and Young’s modulus. PCL degrades three times more slowly than PLA. Its hydrophobicity typically results in poor cell attachment and inadequate tissue integration. PLC scaffolds for tendon repair are generally combined with electrostatic spinning technology.8 Yang et al incorporated gelatin hydrogel into electrospun PCL scaffolds. The hydrogel provided a highly hydrated environment similar to that of natural tissues and enabled the homogeneous distribution of cells encapsulated within it. As components of natural ECM, collagen and gelatin mimicked natural tissue microenvironments to a certain extent, improving the biocompatibility of PCL scaffolds.192 In another study, a sheep membrane enriched with collagen, cytokines (such as TGF-β1, bFGF, VEGF and PDGF) and other active ingredients was selected to mimic the tendon sheath. To improve the mechanical strength of the membrane and confer moderate hydrophobicity, PCL nanofibres were coated on both surfaces to construct a composite membrane that mimicked the structure of the tendon sheath. This effectively isolated the exogenous adherent tissues and promoted endogenous tendon healing.236 PCL can also be used as constructed nanoparticles to control the sustained release of drugs.237
Hydrogel Materials
Hydrogels are three-dimensional networks with a high water content, composed of hydrophilic polymers. They are similar to natural ECM in terms of their high biocompatibility and the fact that their physical, chemical and biological properties can be adjusted according to the needs of a variety of sources, such as collagen, chitosan, HA, alginate, gelatin and elastin, from natural sources, as well as polyethylene glycol (PEG) and polyvinyl alcohol (PVA) from synthetic sources. The advantages and disadvantages of each type of hydrogel are related to its source. Hydrogels from natural materials have insufficient mechanical strength and it is difficult to control degradation time, while hydrogels from synthetic materials have relatively low biocompatibility.238
Drug Carrier Hydrogel
In bioengineering treatments for tendinopathy, hydrogel materials are typically used for drug delivery and scaffold fabrication. When delivering drugs, the poor mechanical strength and adhesion of the hydrogel must be considered, as well as whether it can be used as a drug carrier to release drugs over a long period of time and remain undamaged in the affected area during tendon activities. For instance, a hydrogel system could be employed to release non-steroidal anti-inflammatory drugs (NSAIDs) over an extended period at the site of tendon injury via a single injection, thereby controlling inflammation and pain while promoting tissue regeneration.239 Benjamin et al prepared a tough viscous hydrogel consisting mainly of alginate, acrylamide, and chitosan. This hydrogel was found to exhibit significantly improved adhesion in both in vitro and in vivo experiments. It was shown to adhere to affected areas for at least three weeks and release drugs on demand for over one week. Furthermore, the hydrogel was capable of releasing drugs for over one year. In vivo experiments, it was found to adhere to the affected area for at least 3 weeks, release drug on demand for more than 1 week, and resist mechanical breakdown caused by tissue movement. The hydrogel reduced friction during tendon movement, alleviating tissue adhesion around the diseased tendon. An increase in M2-polarised macrophages and tendon stem cells was also observed, contributing to improved tendon healing.200 On the other hand, Cai et al proposed a self-healing, MMP-2-degradable gelatin methacryloyl microsphere formulation containing Smad3 siRNA-loaded nanoparticles and a hyaluronic acid (HA) hydrogel. An increase in MMP-2 following tendon injury would promote the release of Smad3 siRNA, reducing the adhesion of peritendinous tissue without affecting the tendon’s mechanical properties. Even if the tendon movement leads to the rupture of the self-healing hydrogel, the self-healing hydrogel can restore the original structure after pressure reduction to minimize the rupture-induced inflammation as well as adhesions.240 Similarly, Ren et al designed an immediate self-healing hydrogel that can adapt to irregular tendon wounds, fitting tightly into them. It is equipped with double encapsulation of microspheres and hydrogel to ensure the slow release of basic fibroblast growth factor (bFGF). This significantly reduces inflammation and facilitates collagen I secretion, promoting tendon healing in an injury model.241
Scaffold Hydrogel
Hydrogels are also often used as scaffolds due to their generally excellent bioactivity on their own. Peyman et al prepared hydrogels consisting of polyglycerol-functionalised reduced graphene oxide and polyglycerol-functionalised molybdenum disulfide, which were hybridised in a gelatin matrix. Experiments demonstrated that the group treated with the scaffold had a better clinical tendon score, and that the presence of sulphur and molybdenum in the hybridised scaffolds may inhibit the inflammatory response and alleviate peritendinous tissue adhesion during the healing process. Furthermore, the scaffold’s small pore size prevented exogenous fibroblasts from entering the injured area, while allowing growth factors such as VEGF to pass through. This stimulated the healing process, reducing inflammation and adhesions. Accelerated tendon healing and more regular tissue morphology were also observed, and it was found that tendons treated with the scaffold possessed higher biomechanical properties. However, the toxicity of the scaffold was not evaluated.242 Xu et al developed a hydrogel consisting of bioactive glass and SA. The bioactive glass promotes M2 macrophage polarisation and enhances angiogenesis, thereby promoting tendon healing. Meanwhile, the SA alleviates the pain caused by the high local pH resulting from the bioactive glass. Application of this scaffold improves tendon histological morphology and tensile properties, with no adverse effects due to the osteogenic properties of the bioactive glass. The osteogenic properties of bioactive glass resulted in heterotopic ossification.155 However, hydrogels have poor mechanical properties compared to natural tendons. Fibre-reinforced hydrogel composites (FRHCs) have been proposed to improve tensile strength, stiffness and toughness by incorporating reinforcing fibres made from natural or synthetic materials into the hydrogel matrix. Their biocompatibility allows FRHCs to recruit TDSCs, promote their differentiation into tendon tissue and modulate the immune environment. The aligned fibres can also mimic the arrangement of natural tendon tissue to guide collagen formation and promote tendon healing with better morphology. However, the mechanical strength of the FRHC is still inferior to that of natural tendons. FRHC only increased the tensile modulus of hydrogels from 10^(−3) to 10¹ MPa, whereas the tensile modulus of pure fibre scaffolds was around 10¹ to 10² MPa and that of human tendons was around 80 MPa.243,244 It is worth mentioning that He Zhu et al’s directional annealing casting (DAC) technique transformed the polyvinyl alcohol (PVA) hydrogel from a disordered porous structure into a highly oriented fibrous structure. This increases its ultimate tensile strength to 43.5 MPa.245 Despite the excellent biocompatibility demonstrated by hydrogel materials, there is still a need to further enhance the mechanical properties of hydrogel materials to suit the needs of tendon as a mechanically transferable tissue.
Commonly Used Biological Factors
Loading cells and cytokines onto the scaffold will help to further mimic the natural structure of the tendon. Delivering genes to organisms through various carriers is also a commonly used bioengineering therapy. The following will briefly summarize the common cells, cytokines, genes and exosomes in tendon bioengineering.
Cells
An important aspect to consider for tendon tissue engineering is the cell source. The more common ones are mesenchymal stem cells, tendon-derived cells, dermal fibroblasts, myogenic stem cells, periosteal progenitor cells and induced pluripotent stem cells. Direct injection of cells for tendinopathy has been shown to be effective.16 However, direct cell injection will bring the problem of low cell survival. Applying bioengineering technology to prepare scaffolds to deliver cells may be a better solution.246
Tendon-Derived Stem Cells
Tendon-derived stem cells(TDSCs) are ideal for the treatment of tendinopathies. TDSCs can replenish local tendon cell populations, stimulate the production of growth factors and ECM proteins including type I collagen to promote tissue repair and regeneration. TDSCs have been attempted to be used for tendon repair since early times. Ni et al delivered TDSCs to rat patellar tendon window defects via fibrin glue solution. At 4 weeks, staining revealed improved cell alignment, collagen fiber alignment. Significantly higher ultimate stress and Young’s modulus than the undelivered cell group.247 A number of scaffolds have also been developed for delivery of TDSCs. Ning et al developed decellularized tendon hydrogel for loading TDSCs, which retained more stromal cell-derived factor-1(SDF-1) and fibromodulin(Fmod) compared to collagen gel and significantly promoted TDSCs migration and tendon differentiation.248 Fibrovascular scarring, fat accumulation, and heterotopic ossification are often present after tendon injury, so Zhang et al proposed a RADA nanofiber hydrogel loaded with TDSCs to give the repaired tendon a better macroscopic appearance and improved stromal organization, reduced adipocyte accumulation and vascularization, and attenuated heterotopic ossification formation at 8 weeks postoperatively249 (Figure 5a). The presence of collagen chaotic arrangement in the scar tissue formed after tendon injury weakens the mechanical properties and function of the injured tendon. Electrostatic spinning technology is often used to prepare neatly aligned nanofibers to mimic the anisotropic hierarchical structure of natural tendon tissues and to guide the orientation and differentiation of TDSCs with the formation of regular ECM. Yang et al found that oriented electrostatically spun fiber yarns PCLs of 50 μm in diameter could promote tendonogenic differentiation of TDSCs. In vivo experiments in rats revealed that bundled fiber yarns promoted tendon repair in vivo by inducing the formation and orientation of neo-collagenous tissues, demonstrating the potential of this scaffold as a tendon mimicking multiscale scaffold.250 TDSCs also have anti-inflammatory effects. Mao et al used small intestinal submucosal (SIS) scaffolds to deliver TDSCs to Achilles tendon defects in rats. SIS provided a large amount of collagenous tissue for Achilles tendon regeneration, which provided a suitable cellular environment for TDSCs. TDSCs enhanced tendon regeneration and anti-adhesion ability by regulating inflammatory responses together with SIS.251 However, TDSCs show phenotypic loss during in vitro culture, posing many obstacles to the expansion and application of TDSCs.252 Many have explored factors that may contribute to the maintenance of stem cell phenotype, such as cytokines, physical microenvironment.32,253,254 Since TDSCs therapies tend to be autologous in origin, they are limited in number and may be less effective if patients have risk factors associated with tendinopathy.255,256 Therefore stable access to TDSCs remains to be explored.
Figure 5 Continued. Figure 5 Application of common biological factors in tendon regeneration. (a) RADA hydrogel loaded with human TDSCs promoted the repair of rat patellar tendon and reduced heterotopic ossification at 8 weeks; in the staining pictures black arrows indicate adipocytes and green arrows indicate blood vessels; in the Micro-CT green arrows indicate ectopic calcification; *p<0.05, ***p<0.001. (b) BMSCs-loaded PLGA scaffolds enhanced tendon-bone healing in a rabbit rotator cuff repair model; *p<0.05. (c) PRP and HA were delivered to the rabbit flexor tendon rupture model through PCL shell, which reduced tendon adhesion formation and inflammation and promoted tendon healing; arrows indicate adhesion tissues surrounding the tendon; the suture site, tendon and residual membrane are identified as S, T and M, respectively. (d) TDSC-Exos can promote the early healing of injured tendon in rat tendon defect model through HA scaffold; N: normal area; W: wound area; *p<0.05, **p<0.01. Reproduced with permission.

Tendon Cells
Tendon cells are the major cells within tendon tissue and have an important role in maintaining the homeostasis of the tendon ECM. Improvements in tendinopathy with autologous tendon cell therapy have been reported in animal and clinical trials. For example, an experiment in which autologous tendon cells were delivered via a scaffold in a sheep model of tendinopathy found that tendon cells improved histologic morphology, promoted proteoglycan production, and biomechanically approximated the uninjured tendon.257 Another clinical trial in which autologous tendon cells were injected into patients with gluteal tendinopathy found that some growth factors, such as FGFb, TGFβ and PDGFα, were upregulated after injection, which induced tendon cell proliferation and matrix production. And resulted in a significant and sustained improvement in clinical scores at 24 months after injection.258 However, tendon cells, like TDSCs, suffer from phenotypic loss during in vitro expansion.259 The application of autologous tendon cells is hampered by their limited cell number, poorer proliferative capacity, and higher risk of morbidity in the donor area.260
Mesenchymal Stem Cells
Mesenchymal stem cells(MSCs) have high proliferative and self-renewal capacity and can differentiate into tendon cells under specific induction conditions in vivo or in vitro. Secondly, MSCs have a paracrine effect, which can secrete cytokines and extracellular vesicles to neighboring cells to promote the formation of blood vessels in damaged tissues, enhance the tendon formation ability of tendon stem cells, and attenuate the inflammatory response of damaged tissues. Meanwhile, MSCs have a wide range of sources: bone marrow, adipose tissue, placenta, umbilical cord and other tissues. Bone marrow and adipose-derived stem cells have been recognized as suitable alternatives to tendon stem cells.2 Burk et al found more improvement in lameness scores and ultrasound scores after injection of bone marrow-derived MSCs(BMSCs) into naturally equine tendinopathic tendon compared to saline-injected group.261 Clinical studies have also suggested that the use of BMSCs in the treatment of chronic patellar tendinopathy resulted in improvements in clinical manifestations and tendon structure.262 Chen delivered 3D printed PLGA scaffolds loaded with BMSCs to a rabbit rotator cuff injury. The presence of BMSCs significantly increased the mRNA expression levels of type I and III collagen, tenogenic proteins and disaccharide proteoglycans. It also increased the diameter of collagen and improved the biomechanical properties of the injured tendon263 (Figure 5b). However the application of BMSCs may result in increased alkaline phosphatase activity leading to heterotopic ossification. BMSCs need to be collected by drilling through the iliac crest, whereas adipose-derived stem cells(ADSCs) can be obtained in large quantities by less invasive methods without any ethical issues and it has been found that the addition of ADSCs in the early stages of tendon healing has been found to inhibit heterotopic ossification and can increase the I/III collagen ratio.264 Bortolazzo et al utilized ADSCs in conjunction with microcurrent stimulation in an attempt to repair tendon injuries.265 It has been proposed that ADSCs cell sheets can be used to promote tendon-bone healing.266,267 Wu et al prepared HA, collagen and SA into scaffolds loaded with ADSCs by 3D printing. The synergistic effect of slow-release BMP-2 effectively promoted the differentiation of ADSCs into osteogenic, tendinogenic and chondrogenic lineages. Regeneration of tissue structure and recovery of mechanical properties were promoted.268 However, ectopic fat deposition was found in animal experiments using ADSCs therapy and ADSCs obtained from obese donors were found to exhibit excessive immune responses and reduced differentiation potential.4,269
Dermal Fibroblasts
Dermal fibroblasts have a spindle morphology and ECM similar to that of tendon cells. They are easier to obtain and to culture than tendon cells. In an animal study, dermal fibroblast therapy was found to exhibit histological and biomechanical properties similar to those of tendon, about 75% of the strength of natural tendon. Lee et al compared the ability of human dermal fibroblasts and tendon cells to promote the healing of rotator cuff tears and concluded that dermal fibroblasts showed cellular properties comparing to those of tendon cells.270 Rhee et al proposed the use of a combination of HA and dermal fibroblasts to prevent retear after rotator cuff repair. HA synergized with dermal fibroblasts to increase collagen fiber density and collagen fiber continuity and tendon-bone interface maturation in the injured rotator cuff.271 Treatment with autologous dermal fibroblasts has also been found to improve pain and risk of re-tear in patients in clinical trials.272 However, the use of fibroblasts may pose a problem of scarring.273
Embryonic Stem Cells and Induced Pluripotent Stem Cells
Embryonic stem cells (ESCs) are derived from the inner cell mass of blastocysts. Due to their strong differentiation ability, ESCs are considered an important cell source for regenerative medicine. In vitro animal experiments have revealed that ESCs can exhibit significant up-regulation of tendon-related genes, including Tnmd, platelet reactive protein 4, tendonogenic protein C and type I collagen. In vivo experiments also showed that ESCs improved tendon tissue repair. However, the application of ESCs involves social and ethical issues, which can be avoided by induced pluripotent stem cells(iPSCs). IPSCs are derived from differentiated somatic cells and it has been found that applying mechanical loads on iPSCs promotes the tendon differentiation of iPSCs. Zhang et al attempted to utilize the physical properties of induced iPSCs to induce tendon differentiation and they fabricated well-aligned chitosan-based cells by stabilized jet electrostatic spinning. They fabricated well-aligned chitosan-based microfiber scaffolds by stabilized jet electrostatic spinning to regulate the orientation and differentiation of iPSCs and their in vivo study revealed that they improved the structure and mechanical properties of injured tendons. However, the lower differentiation efficiency of iPSCs compared to ESCs and the risk of tumors such as teratoma formation limit the clinical application of iPSCs, probably due to the fact that iPSCs retain some of the epigenetic changes.274–277
Cytokines
Cytokines are compounds or molecules that are synthesized and secreted by cells to exert an effect on organisms, tissues, or other cells, interacting with and regulating cellular activity. Cytokines are often combined in bioengineering with scaffolds, which act as carriers and provide both physical and mechanical support. Often cytokine therapy combined with a suitable biological scaffold is superior to a single injection of the same cytokine therapy.221 Currently, scaffolds of various natural or synthetic materials can be manipulated by manipulating their physicochemical properties to provide different release kinetics, depending on the desired release profile. It has recently been found that combined delivery of cytokines with different spatial and temporal distributions in composite scaffolds enhances tissue regeneration compared to long-term release of single factors.278
Platelet Rich Plasma (PRP)
PRP, an autologous human plasma preparation with increased platelet concentration produced by centrifugation of a larger volume of blood, is thought to potentially assist in the post-injury repair of tendon tissue through a variety of growth factors, including many cytokines such as TGF-β1, PDGF, bFGF, VEGF, EGF, IGF-1, and hepatocyte growth factor(HGF). PRP has been found to promote tendon cell proliferation and improve collagen production and COL1A1:COL3A1.279 Chen concluded that PRP injections can relieve the pain of tendinopathy.280 PRP preparations can be further categorized into leukocyte-rich PRP and leukocyte-poor PRP. After summarizing recent in vivo and in vitro experiments, Liu et al concluded that leukocyte-rich PRP helps to promote type I collagen synthesis without promoting type III collagen synthesis, which may help to restore the natural structure of tendons. And since tendons in the inflammatory phase require the involvement of inflammatory cells, early administration of leukocyte-rich PRP promotes tendon healing better than late administration.281 Whereas the current study suggests that leukocyte-poor PRP may contribute to tendon recovery after rotator cuff repair, it does not otherwise demonstrate particular advantages.282–284 In order to prolong the release of PRP in vivo, Zhang et al proposed the use of gelatin sponges for prolonging the release of bioactive factors in PRP, which promoted the expression of the osteogenic genes ALP, OCN and RUNX2. In combination with gelatin sponges PRP promoted a clearer junction at the tendon-osteochondral junction and more regular fibrocartilaginous transition zones, as compared to the control group. Histologic scores were also higher. Unfortunately, the study lacked long-term observation and did not find biomechanical benefits of the combined gelatin sponge on the affected area in the short term.285 And with the development of electrostatic spinning technology, nanofiber membranes have become an option for controlled drug release, Chen et al injected PRP into nanofiber core shells to mimic the secretion of synovial fluid and growth factors in normal tendon sheaths189 (Figure 5c). However, the help of PRP on injury healing is controversial, for example, a clinical follow-up found no difference between PRP and controls, these controversies may be related to the different ways of preparing PRP, more studies are needed to confirm the exact efficacy of PRP in tendinopathy.286,287
Platelet-Derived Growth Factor (PDGF)
During the inflammatory response phase PDGF attracts inflammatory cells such as neutrophils and macrophages and promotes debris breakdown and phagocytosis. During the healing phase PDGF attracts tendon cells and fibroblasts to migrate to the injury site and synthesize ECM.288 In vitro experiments have found that PDGF stimulates tendon cell proliferation and ECM synthesis and an in vivo experiment confirmed that delivery of PDGF to the injury site promoted tendon cell proliferation and type I collagen expression.289 PDGF-BB immobilized on nanofiber scaffolds has also been studied to promote the proliferation and tendonogenic differentiation of ADSCs.290
Vascular Endothelial Growth Factor (VEGF)
VEGF has been found to have the potential to enhance Achilles tendon healing and increase tensile strength in both animal and human experiments, which could possibly be related to the stimulation of angiogenesis by VEGF. It has also been mentioned that VEGF has an inhibitory effect on the lipogenic differentiation of TDSCs and reduces the accumulation of adipocytes in tendons of aged rats.113 However, some studies have found that the area of increased pain at the injury is closely related to the neovascularization system in the tissue. More experiments are needed to explore whether VEGF is suitable for tendon repair at this time.16
Insulin-Like Growth Factor (IGF)
IGF-1 plays a key role in tendon healing by promoting mitosis and migration of tendon cells, increasing collagen and proteoglycan production during the remodeling phase, and increasing the mechanical strength of the healing tendon, as well as inhibiting local inflammation. IGF-1 may play a regulatory role through the PI3K/AKT and ERK pathways.291 Rieber et al used an electrostatic spinning technique to prepare a three-layer structured polymer degrapol tube with an IGF-1-containing middle layer, which promoted the up-regulation of ki-67 and TNMD in rabbit tendon cells in an in vitro assay.292 However, the current clinical trials did not show a more significant effect and more in vivo studies are needed to explore the application of IGF.293,294
Transforming Growth Factor β (TGF-β)
TGF-β is released immediately after tendon injury and promotes macrophage and fibroblast chemotaxis while possibly acting as an inhibitor of excessive inflammation. TGF-β1 has been found to promote secretion of fibronectin and collagen by MSCs. It has also been mentioned that TGF-β1 and cytoskeletal signaling synergistically promote tendon differentiation of fibroblasts, thereby promoting tendon healing.295 However, high levels of TGF-β1 can lead to increased adhesions. Therefore, Wu et al used scaffolds loaded with a silencing TGF-β1 plasmid delivered into a chicken tendon defect model to prevent tendon adhesions.219 TGF-β1, TGF-β2, and TGF-β3 of the TGF-β family increase the secretion of collagen types I and III, which is currently thought to lead to excessive scar tissue formation. But blocking TGF-β1 reduced the mechanical strength of tendons at the site of injury. Another study inhibited ECM production and tendon repair after interfering with TGFβ2 expression using miR-378a.293,296 Therefore, the relationship between tendon repair and tendon scarring needs to be considered when regulating TGF-β.
Bone Morphogenetic Protein (BMP)
BMP-2 is one of the most important osteogenic growth factors. It also induces the formation of type I collagen, which enhances tendon-bone healing and increases its biomechanical strength. However, when high doses of BMP-2 are used to obtain a therapeutic effect it may lead to ectopic bone formation. Therefore Arvinius et al prepared SA-chitin scaffolds to protect BMP-2 from enzymatic degradation and to allow controlled release of BMP-2. The scaffold-treated group possessed tendon-bone junctions that more closely resembled the natural morphology and had better biomechanical properties.297 Han et al combined the use of BMP2 and platelet-rich fibrin to treat anterior cruciate ligament reconstruction in rats, promoting the upregulation of signals associated with bone formation and tendon regeneration.298 In animal experiments, BMP13 injection was found to assist in the repair of injured tendon tissue by promoting the formation of highly organized type I collagen fibers and significantly enhanced mechanical strength. The importance of BMP signaling for tendon differentiation was mentioned in a zebrafish model by Niu et al.5 However, the tenogenic induction of BMP may need to be combined with mechanical stimulation.299
Basic Fibroblast Growth Factor (bFGF or FGF-2)
Basic fibroblast growth factor(bFGF) has been found to be expressed after tendon injury. bFGF stimulates the proliferation and differentiation of vascular endothelial cells during the inflammatory phase to promote neointima formation and assists in the conversion of collagen type III to collagen type I during the remodeling phase by stimulating the secretion of collagen type III from fibroblasts. bFGF has been found to increase the biomechanical strength of tendons after topical bFGF treatment in an animal model of injury. In animal injury models, it was found that local treatment with bFGF may increase the biomechanical strength of tendon healing and accelerate tendon-bone remodeling.300 bFGF can stimulate the proliferation, self-renewal and tendon differentiation of BMSCs, which are commonly used in tendon repair. bFGF stimulates proliferation, self-renewal and tendon differentiation of BMSCs commonly used for tendon repair. Stimulation of stem cell proliferation and tendon differentiation by implanting bFGF in scaffolds has been studied to enhance the production of tendon-specific proteins, providing biological clues for maintaining the tenogenic phenotype.301 However, excess bFGF can lead to tendon adhesions as well as scar tissue formation.302,303
Genes
During tendon repair, genes that promote tendon healing are not produced immediately after injury and genes that promote adhesion and scar formation are overexpressed. Gene therapy seeks to modify or manipulate the expression of genes in order to achieve better healing of the tendon with the formation of fewer adhesions. Gene therapy allows for the continuous production of endogenous proteins and is more widely applicable than cell or cytokine therapy, which requires repeated infusions.218 Vectors currently used to deliver genes generally include adenoviruses, lentiviruses, retroviruses and adeno-associated viruses. However, gene therapy is still a work in progress and tends to be expensive and risky with the use of viral vectors.7
Promoting Tendon Healing
Due to scar repair, tendon injuries often do not reach the level of healthy tendons and are at risk of re-rupture. There have been many studies exploring the delivery of genes to promote tendon repair as well as biomechanical levels after repair. Usman et al injected double stranded miR-210 directly into rat Achilles tendon injury repairs. At 2 weeks, the treatment group had significantly more collagen fibers than the control group in terms of diameter and ultimate destructive loading. At 12 weeks, the treatment group possessed more parallel and dense collagen fibers.304 A study promoted tendon generation after retroviral delivery of long chain non-coding RNA H19 to human TDSCs and accelerated tendon healing in a mouse tendon defect model.305 Tang et al delivered the bFGF gene using an adenovirus-associated vector to a surgical repair model of the toe flexor tendon in chickens. bFGF gene-delivered tendons showed significantly greater ultimate strength than control tendons at 2 and 8 weeks, with no increase in adhesion formation. In a similar experiment, they found that delivery of the VEGF gene resulted in a slightly later healing-promoting response and stronger healing than the bFGF gene.306 Heterotopic ossification after tendon injury impedes tendon healing. Geng et al injected lentivirus-loaded miR-337-3p into the affected area of a rat model of collagenase-induced tendinopathy and regulated the osteogenic differentiation of TDSCs by targeting NOX4-JNK signaling and IRS1-ERK signaling, which ameliorated heterotopic ossification and promoted the healing of the affected area healing.307 Modifying the gene expression of cells in vitro before delivery in vivo is also a commonly used bioengineering protocol.308–310 Chen et al overexpressed SCX from human embryonic stem cell-derived MSCs and loaded them onto knitted silk and collagen sponge composite scaffolds. The overexpression of SCX enhanced the tendon lineage differentiation and promoted tendon regeneration in vivo.311
Inhibition of Tendon Adhesion
Adhesion is another important problem that arises during tendon repair. Adhesion will result in hindering tendon sliding. In one study, delivery of miRNA plasmid inhibiting TGF-β1 expression by adenovirus-associated vector to transverse finger flexor tendons of chickens reduced adhesions but decreased tendon healing strength.312 Cyclooxygenase(COX) is a key pro-inflammatory factor in the inflammatory microenvironment after tendon injury, so Sun et al delivered siRNA using PLGA nanoparticles loaded with extracted M2 macrophage membranes on the surface to inhibit COX expression. The treatment reduced inflammatory response and significantly improved tendon adhesion in a rat model of tendon and rotator cuff injuries.313 Hasslund et al delivered growth differentiation factor 5(GDF-5) via recombinant adeno-associated virus to reconstructed phalangeal flexor tendons in mice, which improved joint flexion function without compromising the strength of the repair within a certain dose range.217
Exosomes
All cell types can produce exosomes across biological membranes, which play a role in intercellular communication and homeostatic regulation. Exosomes are bilayer vesicles with diameters ranging from 30 to 150 nm containing proteins, lipids, mRNA, non-coding RNA and DNA. Exosomes have been used as drug delivery vehicles and are immunocompatible, non-cytotoxic and non-mutagenic in comparison to other existing vectors. The molecular components in exosomes vary and are influenced by a variety of factors and the function of exosomes depends largely on the content of the molecular components, which is closely related to the nature of their parental cells.314 The use of exosomes for the treatment of tendon disorders offers a number of advantages over cellular therapies, including low immunogenicity, no need to maintain cell viability, elimination of the risk of uncontrolled proliferation and differentiation of the implanted cells and no need to consider the requirements for graft traits, survival before and after treatment.315
MSCs-Derived Exosomes
Due to the wide range of MSC sources, many MSC-derived exosomes related experiments have been carried out. Yao et al found that miR-29a-3p in human umbilical cord MSC-derived exosomes promoted the expression of tendon markers and increased collagen production through the PTEN/mTOR/TGF-β1 signaling cascade, which accelerated tendon healing process and improved tendon biomechanical properties.316 Other studies have focused on modulating the inflammatory response in tendinopathy to improve tendon healing. Shi et al delivered exosomes derived from BMSCs to rat patellar tendon defects via fibrin glue. The exosomes inhibited the production of apoptotic cells and pro-inflammatory macrophages, increased the expression of anti-inflammatory mediators IL-10 and IL-4, and up-regulated the expression of SCX and TNMD, which ultimately led to the improvement of tendon morphology.317 M2 macrophage polarization to inhibit inflammation in tendinopathy has the potential to improve the histological and biofunctional evaluation of tendon. Xu et al used bioactive glass to induce MSC-derived exosomes to regulate M2 macrophage polarization to activate anti-inflammatory effects and increase angiogenesis, which was histologically and biomechanically superior to uninduced MSC exosomes.315 Song et al used melatonin to modulate the circadian rhythm of ADSCs and found that it increased the production of secretory extracellular vesicles and enhanced the inflammatory regulation of ADSCs, and inhibited M1 macrophage polarization.318 Wu et al also found that ADSC exosomes from young mice had higher NAMPT content compared with ADSC exosomes from aged mice, which could be mediated by NAMPT/SIR. They can promote macrophage phagocytosis and M2 polarization through the NAMPT/SIRT1/NF-κb p65/NLRP3 pathway. They also enhanced cell viability and alleviated tendon cell senescence through the NAMPT/SIRT1/PPARγ/PGC-1α pathway.319 The alleviation of tendon senescence would be expected to improve tendon repair.
TDSCs-Derived Exosomes
It has been found that exosomes from TDSCs can balance matrix synthesis and degradation in injured tendons and promote tendon repair by decreasing the expression of MMP-3 and increasing the expression of TIMP-3 and Col-1a 1.320 Song et al delivered exosomes derived from TDSCs to tendon defects via HA scaffolds and found that the exosomes could protect tendon cells from oxidative stress damage and promoted tendon cell proliferation and migration through miR-144-3p in them. Increased expression of type I collagen production and tendon-specific markers were also observed in the injured tissues. Better fiber alignment and histological scores were obtained321 (Figure 5d). Micro-RNAs have demonstrated considerable roles in tendon repair, but instead of directly intervening in the cells with miRNAs, An et al used lentivirally transfected overexpressed mir-337-3p exosomes from TDSCs promoted tendon cell proliferation and migration and prevented tendon cell apoptosis by regulating CASP3.322 However, there are relatively few studies on exosomes of TDSCs, which are limited by the isolation and characterization of TDSCs.
Exosomes from Other Sources
In addition to MSC and TDSCs-derived exosomes, studies have explored other sources of exosomes for assisting in tendon repair. Graça et al cultured human TDSCs using platelet-derived exosomes and found that platelet-derived exosomes increased the expression of markers of tendon generation and promoted extracellular matrix remodeling and synthesis of anti-inflammatory mediators.323 Another study also found that platelet-derived exosomes attenuated TDSCs iron death in rats through activation of AMPK phosphorylation and downstream nuclear factor erythroid 2-related factor 2(Nrf 2)/glutathione peroxidase 4 (GPX4) signaling pathway.324 A study also utilized platelet-derived exosomes loaded with recombinant Yap 1 protein delivered to TDSCs, which significantly protected TDSCs from oxidative stress-induced stemness loss and SASP via the NF-κB signaling pathway.325 Jin et al found that anti-aging signaling is rich in EV secreted by human exfoliated deciduous dentin stem cells (SHED-Exos). SHED-Exos regulated histone methylation and inhibited NF-κB to reverse tendon stem cell aging. Local delivery of SHED-Exos reduced senescent cells and ectopic bone formation.326 Rao Chen et alalso mentioned that the use of dendritic cell-derived exosomes (DEX) to repair the mouse model of Achilles tendon rupture can induce the transformation of M1 macrophages into M2 macrophages by activating the PI3K / AKT pathway. It has been demonstrated through rigorous research that DEXs have the capacity to stimulate the synthesis of type I collagen and to impede the synthesis of type III collagen within the tissue. This process has been shown to result in an acceleration in the healing process of tendons. Moreover, the inflammatory microenvironment was found to be improved by DEX treatment, as evidenced by a reduction in the levels of various cytokines (IL-1β, IL-4, IL-6, TNF-α, and IFN-γ).
Although exosomes show good therapeutic potential for tendinopathies, the requirement for exosomes to persist in the injured area and maintain effective concentrations, as well as the synergistic generation of a favorable microenvironment for tendon repair in conjunction with other biological factors, such as cells and cytokines, requires the support of other bioengineered materials and methods.
Commonly Used Bioengineering Strategies
Cell Slices
Cell sheet technology is a scaffold-free tissue engineering technique. In tissue engineering, the delivery of cells disrupts intercellular and cell-ECM interactions, resulting in less than optimal survival and uneven distribution of implanted cells. Cell sheet technology, however, uses a degradable substrate to carry the cells, which maintains intercellular junctions and maximizes the ECM components without disrupting them, resulting in a more homogeneous distribution of cells and avoiding the immunogenicity issues that need to be considered when using conventional scaffolding materials. Cell sheet transplantation has been reported to have a higher cell survival rate within 12 months compared to cell injection. Cell sheets can also be stacked to form three-dimensional structures. The most widely studied is the thermosensitive cell sheet, in which the substrate changes from hydrophobic to hydrophilic when the temperature falls below a critical temperature, which causes the cells to detach from the substrate. Electrosensitive cell sheets can be used to control cell adhesion and detachment by applying an electrical potential. Photosensitive cell sheets are generally irradiated by UV light for that century of cell adhesion and detachment. There are also PH-sensitive cell sheets, magnetic field-sensitive cell sheets, ROS-sensitive cell sheets, etc.327,328
In an earlier study exploring the delivery of temperature-sensitive TDSCs cell sheets to Achilles tendon defects in rats, the tendon defect area implanted with TDSCs cell sheets had better organized collagen fibers compared to the disorganized collagen fibers in the control group. Transmission electron microscopy observations revealed the presence of longitudinally aligned collagen fibers and thick collagen fibrils in the TDSCs cell sheet implantation group. The implanted group also possessed a higher limiting Loading329 (Figure 6a). Liu et al demonstrated in a canine rotator cuff tendon bone healing model that treatment with bone marrow mesenchymal stem cell slices combined with engineered tendon-fibrocartilage-bone composites had higher histological scores compared to treatment with engineered tendon-fibrocartilage-bone composites alone. More neo-fibrocartilage formation and collagen fibre deposition at the tendon-bone interface were observed. The ultimate destructive load and ultimate stress were elevated.330 A similar study was done by Tang et al.331 Zhao et al also used bone marrow mesenchymal stem cell sheets for repairing Achilles tendon defects in rats in conjunction with a composite scaffold loaded with bFGF in order to increase the amount of stem cells implanted to ensure the integrity of the cellular surroundings. The scaffold co-treatment group showed higher expression of tendon marker genes and a significant increase in biomechanical strength.146 Gonçalves et al attempted to apply magnetic field-sensitive stem cell sheets to tendon repair, in which magnetically induced particles were implanted to respond to the magnetic field and provide directional biomechanical stimulation of the cells to remotely modulate the guidance of the cells.332 Another study explored the role of magnetic field sensitive cell sheets in the regulation of tendon inflammation, using IL-1β to induce inflammation produced by magnetically assisted cell sheets could be used to study inflammatory events in tendon cells in vitro, while pulsed electromagnetic field stimulation may have promoted the expression of anti-inflammatory genes through the MAPK pathway.333 However potential safety issues of stem cells such as the risk of inducing cancer have delayed their clinical translational application. Therefore, Yao et al proposed decellularized tendon-derived stem cell sheets. The decellularized tendon-derived stem cell sheet treatment group showed similar histological morphology, collagen levels and growth factor levels as the tendon-derived stem cell sheet treatment group, demonstrating its potential to be used as a cell-free alternative to its cell-free counterpart in the repair of tendon injuries.334 Cell sheet technology has many advantages over traditional direct cell injections, but the high cost of cell sheet production limits its wide-scale application.335
Figure 6 Continued. Figure 6 Application of common tissue engineering techniques in tendon regeneration. (a) TDSC sheet effectively promoted tendon remodeling after implantation in rat tendon defects. (b) 3D printed bionic multi-layer scaffold is helpful to promote the healing of large-scale rotator cuff tendon defects in rabbit models; the boxes in the upper panel of each group are displayed at higher magnification in the lower panel; T: tendon; B: bone. (c) The nanofiber yarn made by electrospinning can simulate the arrangement and grading structure of natural tendon tissue, which is helpful to promote tendon repair in rats; N: neo-collgen; NFY: NFY-50; *p<0.05. (d) The curled nanofiber scaffold prepared by electrospinning promotes tendon-bone interface regeneration in a rabbit model of giant rotator cuff tear; *: significant difference compared with NS, P<0.05; **: significant difference compared with NS, P<0.01. Reproduced with permission. Abbreviations: NTBI, natural tendon-to-bone interface; NS, nanofiber scaffold; CNS, crimped nanofiber scaffold.

3D Printing
3D printing is a bioengineering technology based on printing inks, controlled by computers, which can rapidly design and fabricate materials and allow for precise control of structures at the micron scale. 3D printing allows for the incorporation of proteins, growth factors, drugs, DNA and other biochemical signals with cells. And even introduces cellular density gradients to produce scaffolds with intricate and reproducible properties. It is also capable of controlling porosity and pore size, ultimately creating complex tissues that mimic natural tissues. There are many types of 3D printing, such as inkjet-based bioprinting, which helps to maintain cell viability but is less likely to achieve the mechanical properties of tendons. Light-curing-based bioprinting, which can provide high-precision scaffolds but the laser beams can lead to cellular damage. Fused deposition-based bioprinting is also more common, but its lower precision limits its ability to mimic natural tendon Structure.336–338
In order to construct tendon-like 3D structures, there have been many studies exploring 3D printed scaffolds applied to tendon repair, such as composite scaffolds based on tendon decellularized ECM hydrogel, composite scaffolds based on PLA and graphene, composite scaffolds based on collagen glycosaminoglycan and acrylonitrile butadiene styrene(ABS), composite scaffolds based on GelMA hydrogel and glycosaminoglycan HA based composite scaffolds.339–342 Jiang et al designed PLGA, collagen and fibronectin based scaffolds. They also compared monolayer PLGA scaffolds with 3D scaffolds. It is clear that the scaffolds with 3D structures have better mechanical properties and are more capable of supporting the proliferation and differentiation of loaded cells343 (Figure 6b). In order to construct 3D scaffolds with high porosity while controlling the fiber orientation in the scaffolds and further improving the precision of the scaffolds, Zhang et al attempted to construct scaffolds using electrohydrodynamic 3D printing using PCL as the basic printing ink, modified with F127 to increase the biocompatibility of scaffolds.344
The tendon-bone interface has complex gradient characteristics. Chae et al developed scaffolds with gradient characteristics similar to natural TBIs in terms of structure, composition and cellular heterogeneity by taking advantage of 3D printing’s ability to generate predetermined complex layered structures.345 Neubauer also tried to mimic the gradient between tendon bone with the help of a recombinant spider silk hydrogel with fluorapatite. The study used recombinant spider silk hydrogel with fluorapatite to try to simulate gradients between tendon bones. This study achieved adjustable gradient characteristics by encapsulating fibroblasts in recombinant spider silk-fluorapatite hydrogel after using recombinant spider silk hydrogel compounded with fluorapatite to generate a mineralization gradient.346
The applications of 3D printing in tendons go beyond scaffold development. Since 3D printing can mimic real tendons, it can be used to study the mechanical and physical properties of tendons. Monteiro et al developed a 3D tendon microphysiology system using 3D printing with porcine tendon decellularized ECM hydrogel and human adipose-derived stem cells to observe cellular crosstalk during tendon development, physiology, and pathology.347 Balestri et al suggested that most studies focused on a single interface, whereas injuries often involved multiple interfaces. In order to study the bone-tendon-muscle interface simultaneously, they constructed a 3D in vitro model of bone-tendon-muscle with collagen-agarose-based composite hydrogel and hydroxyapatite through 3D printing to study disease, cancer, aging, and drug effects at the interface.348
However, 3D printing is difficult to print at high resolution on the nanoscale. Therefore it is often difficult to mimic the nanoscale extracellular matrix properties of some tissues.349 In contrast, nanoscale orientable fibers prepared by electrostatic spinning technology are ideally suited to mimic the neatly aligned collagen fiber structure in natural tendons.
Electrostatic Spinning
Electrostatic spinning, a technique that uses an electric field to create fibers from a solution, is commonly used to develop nanofiber scaffolds.350 Scaffolds made from nanofibers have the advantage of high surface area to volume ratio, small pore size and large porosity.351 Currently electrostatic spinning technology has enabled the fabrication of fibrous scaffolds with smaller diameters. Fibers produced by electrostatic spinning have diameters in the range of 50–1000 nm, which is two to three orders of magnitude smaller than fibers made by conventional processes and is closer to the diameter scale of the ECM collagen fibers of natural tendons, which has shown excellent in vitro performance in terms of adhesion, spreading and tendon differentiation of stem cells.231 Many natural and synthetic biomaterials are widely used in electrostatic spinning to produce oriented fiber surfaces, nanofiber yarns and multilayer scaffolds with anisotropic mechanical properties. PLA, PCL and PLGA are the more commonly used materials to fabricate scaffolds for electrostatic spinning due to their biomechanical properties.175,301,352 Yang used PCL to fabricate oriented fibers for induced TDSCs elongation and tendon differentiation250 (Figure 6c). Lim et al attempted to co-mingle PCL and gelatin for electrostatic spinning to mimic collagenous protofibrils of natural tendons.191 Yi et al prepared highly oriented, high-strength SF fibers using a stabilized jet electrostatic spinning method and supported the adhesion, migration and growth of cells in an oriented manner along the fiber axis.353
It has been mentioned that collagen protofibrils in natural tendons exhibit a wavy morphology along the fibre axis. In order to mimic the morphology of natural tendons even further, wavy nanofibrous scaffolds made of Polydioxanone(PPDO) and filipin protein composite or polyester urethane urea(PEUU) and gelatin composite have been proposed. Such convoluted microstructures mechanically and morphologically mimic natural tendon tissue, which can promote tendon cell proliferation and also improve tendon differentiation of loaded ADSCs260,354 (Figure 6d). Another study also incorporated porous properties in addition to obtaining a wavy structure to increase the effect of cell infiltration into the scaffold.355 Another study mimicked the tubular structure of tendons by fabricating nanofiber bundles and their surrounding nanofiber shells with adjustable flexibility to more closely match the properties of natural tendons.356
Nanofibrous membranes prepared by electrostatic spinning have shown promising results in the prevention of tissue adhesions. Nanofibrous membranes can form an effective physical barrier with a structure that prevents the infiltration of fibroblasts around the affected area without affecting the entry of growth factors and nutrients, and therefore, nanofibrous membranes have been studied as drug carriers to control the release of drugs.189,201 Dong et al developed electrospun PCL/SF fiber nanomembranes for anti-inflammatory purposes and used immunomodulatory extracellular matrix for surface modification to mitigate scaffold-induced foreign body reaction.357 Abdulmalik et al co-mingled PCL and cellulose acetate to produce electrospun nanofibers to protect the drug from premature release and degradation.358 A recent study proposed a bilayer positively charged electrostatically spun fibrous membrane, which was prepared by electrostatic spinning of MMP-2 responsive GelMA solution containing COX-2 siRNA complexes into nanofibrous membranes and modulating the surface charge by the naturally occurring cationic polymer chitosan. Thus siRNA was able to be delivered unidirectionally into the peritendinous tissue, both increasing gene utilization and avoiding interference with fragile endogenous tendon healing.359
Blending synthetic materials with natural materials is a way to balance mechanical strength and biocompatibility, but the rapid degradation of natural components may lead to loss of mechanical integrity of electrospun scaffolds.8 And the electrostatic spinning technique is difficult to develop large volume scaffold structures.360 Therefore, more research is still needed to ensure that the final biomechanics meet the requirements and to break through the volume limitations in order to adapt to a wider range of tendon injury conditions.
Clinical Challenges
Many studies have shown the considerable potential of integrating scaffolds with various biological factors for tendon repair. However, it must be emphasised that most of these studies are conducted in a laboratory environment and that only a few have been successfully commercialised or entered clinical trials.8 Although cell sheet technology can retain the microenvironment of cell growth more effectively than other methods, reducing production costs remains challenging. This suggests that there is still some way to go before this technology can be used in clinical practice. The time-consuming nature of the printing process is one of the challenges that needs to be overcome before 3D printing can be produced on a standardised basis.361 Another limitation of using 3D printing for clinical applications is the difficulty of reproducing the obtained shape.337 While 3D printing can be used to create complex scaffold systems, the large diameter of the fibres produced means that it is usually not as effective as electrospinning in simulating the hierarchical structure of natural tendon tissue.232 Electrospinning can produce fibres on the nanoscale, and the diameter of collagen fibrils is approximately 100 nm. This makes electrospinning the most widely used technology in tendon tissue engineering. One of the obstacles to the standardization of electrospinning is its low production efficiency. In addition, the use of toxic solvents such as chloroform, due to their high volatility and inhalation toxicity, poses serious health and environmental risks. At present, the solubility of environmentally friendly solvents to polymers is limited and needs to be further developed. Although the polymers used in electrospinning such as PLA and PCL can be degraded into non-toxic by-products under natural conditions, these fibers usually have limited strength, elasticity and uniformity compared to conventional polymers.362
Currently, most tendon tissue engineering materials are used primarily in experimental animals. Therefore, it is important to consider the differences between animal models and human diseases. As most animals are supported by their limbs, their forelimbs have more weight-bearing functions than human limbs. Therefore, in the animal rotator cuff injury model, animals will withstand greater mechanical stress than humans.363 The treadmill model commonly used in overload studies activates the animal stress response through forced exercise, which may interfere with tendinopathy inflammation levels.364 The most commonly used model of diabetic tendinopathy is the intravenous or intraperitoneal injection of streptozotocin (STZ) to induce type 1 diabetes. However, type 2 diabetes is more prevalent in clinical practice. The pathogenesis of tendinopathy caused by different types of diabetes may differ.365 The fundamental flaws in most tendinopathy models are the use of acute interventions to replicate chronic processes and inadequate management of postoperative mechanical load. These issues can result in discrepancies between animal models and clinical diseases.363 Therefore, while these bioengineering technologies have shown promise in laboratory settings, their application to standardised production requires further research and collaboration.
Summary and Future Perspectives
Tendinopathy is one of the sports system injuries that plague many professional athletes and laborers. The pathophysiological mechanisms of tendinopathy are complex. This article summarizes four main factors that contribute to the development of tendinopathy and impede the healing of injury tendons. Repetitive or excessive mechanical stress is now generally recognized as the main risk factor for tendinopathy. Whereas most of the body tissues and organs have an inflammatory response involved in their repair after injury. Metabolic diseases such as diabetes are increasingly affecting people’s health due to lifestyle changes and are also important players in tendinopathy. Tendinopathies caused by aging are also gaining attention with the aging society and tendon senescence does not only occur in aging individuals, but may also be induced when tissue injury occurs. A number of mechanisms culminate in the three most common manifestations of tendinopathy: scar repair, heterotopic ossification and fat accumulation. The latter is evidence of failed tendon repair and disrupts the tendon’s natural morphology. This affects its biomechanics, causing discomfort and reducing functionality and quality of life. Current conventional therapies cannot restore the natural morphology of the tendon, and there is a risk of tendinopathy recurrence. However, tissue engineering techniques could improve current therapies.
Cells and various cell-derived biological substances such as cytokines, genes, and exosomes are the cornerstones of biological activity, and many therapies have been developed to deliver these substances by injection alone. However, biological activity is often not driven by a single factor, but rather by a combination of biological substances. Therefore, bioengineering that has the ability to integrate and deliver various biological substances will have the potential to further mimic biological activity and more powerfully guide tissue and organ activity in the desired direction. However, many currently developed scaffold materials have the problem of being difficult to meet the high biomechanical performance needs of tendons. Although hydrogels have good biocompatibility and currently show broad bioengineering uses, they lack sufficient biomechanical properties to assist tendons in transmitting muscle-to-bone forces. In contrast, fiber scaffolds with high mechanical properties have relatively low biocompatibility. Although FRHCs enhance the mechanical properties of hydrogels by incorporating fibers with high mechanical properties, they still fall short of those of natural tendons. Therefore, we hope that new hybrid materials will be developed. The rapid development of AI technology nowadays may help us to find more suitable material ratios or processing parameters.
Aging and inflammation are important factors in the development of tendinopathy. Many materials have been explored to target these two factors in the hope of inhibiting and reversing the progression of tendinopathy (Table 2). Mechanical stress injuries are the largest contributor to the development of tendinopathy. But there has been little bioengineering research into mechanical stress injuries. Electrostatic spinning technology shows promise for future scaffolding materials that assist tendons in transmitting forces between muscle and bone to alleviate excessive mechanical loading on the injured tendon. Metabolic diseases such as diabetes mellitus can contribute to the development of tendinopathy. However, tendinopathy due to diabetes mellitus is not widely recognised as a complication of the disease, and few studies have explored bioengineering treatments for diabetes-induced tendinopathy. With further research on the pathophysiological processes following tendon injury and the development of bioengineered materials, future bioengineering will hopefully produce scaffolds that more closely resemble the properties of natural tendons and find more appropriate intervention options, providing more therapeutic choices for future tendinopathy patients.
 |
Table 2 Exploration on the treatment of the pathophysiological mechanism of tendinopathy |
Funding
Chongqing Natural Science Foundation (cstc2021jcyj-msxmX0703, cstc2021jcyj-bsh0247), Doctoral Foundation Program (CSTB2022BSXM-JCX0018), Chongqing health commission(2022MSX007), Medical youth talent support program (YXQN202401).
Disclosure
Bin Chen and Yingqi Zhao are co-first authors for this study. The authors report no conflicts of interest in this work.
References
1. Leong DJ, Sun HB. Mesenchymal stem cells in tendon repair and regeneration: basic understanding and translational challenges. Ann N Y Acad Sci. 2016;1383(1):88–96. doi:10.1111/nyas.13262
2. Youngstrom DW, LaDow JE, Barrett JG. Tenogenesis of bone marrow-, adipose-, and tendon-derived stem cells in a dynamic bioreactor. Connect Tissue Res. 2016;57(6):454–465. doi:10.3109/03008207.2015.1117458
3. Millar NL, Silbernagel KG, Thorborg K, et al. Tendinopathy. Nat Rev Dis Primer. 2021;7(1):1. doi:10.1038/s41572-020-00234-1
4. Qin Y, Ge G, Yang P, et al. An update on adipose‐derived stem cells for regenerative medicine: where challenge meets opportunity. Adv Sci. 2023;10(20). doi:10.1002/advs.202207334
5. Niu X, Subramanian A, Hwang TH, Schilling TF, Galloway JL. Tendon cell regeneration is mediated by attachment site-resident progenitors and BMP signaling. Curr Biol. 2020;30(17):3277–3292.e5. doi:10.1016/j.cub.2020.06.016
6. Sharma P, Maffulli N. Tendon injury and tendinopathy: healing and repair. JBJS. 2005;87(1):187–202.
7. Lipman K, Wang C, Ting K, Soo C, Zheng Z. Tendinopathy: injury, repair, and current exploration. Drug Des Devel Ther. 2018;12:591–603. doi:10.2147/DDDT.S154660
8. Shiroud Heidari B, Ruan R, Vahabli E, et al. Natural, synthetic and commercially-available biopolymers used to regenerate tendons and ligaments. Bioact Mater. 2023;19:179–197. doi:10.1016/j.bioactmat.2022.04.003
9. Gaut L, Duprez D. Tendon development and diseases. WIREs Dev Biol. 2016;5(1):5–23. doi:10.1002/wdev.201
10. Akamatsu FE, Teodoro WR, Itezerote AM. Photobiomodulation therapy increases collagen II after tendon experimental injury. Histol Histopathol. 2021;36(6):663–674. doi:10.14670/HH-18-330
11. Li HY, Hua YH. Achilles tendinopathy: current concepts about the basic science and clinical treatments. BioMed Res Int. 2016;2016:1–9. doi:10.1155/2016/6492597
12. von der Mark K. Localization of collagen types in tissues. Int Rev Connect Tissue Res. 1981;9:265–324. doi:10.1016/b978-0-12-363709-3.50012-7
13. Linsenmayer TF, Fitch JM, Birk DE. Heterotypic collagen fibrils and stabilizing collagens: controlling elements in corneal morphogenesis?. Ann N Y Acad Sci. 1990;580(1):143–160. doi:10.1111/j.1749-6632.1990.tb17926.x
14. Banos CC, Thomas AH, Kuo CK. Collagen fibrillogenesis in tendon development: current models and regulation of fibril assembly. Birth Defects Res. 2008;84(3):228–244. doi:10.1002/bdrc.20130
15. Henninger HB, Underwood CJ, Romney SJ, Davis GL, Weiss JA. Effect of elastin digestion on the quasi-static tensile response of medial collateral ligament: elastin mechanics in ligament. J Orthop Res. 2013;31(8):1226–1233. doi:10.1002/jor.22352
16. Walden G, Liao X, Donell S, Raxworthy MJ, Riley GP, Saeed A. A clinical, biological, and biomaterials perspective into tendon injuries and regeneration. Tissue Eng Part B Rev. 2017;23(1):44–58. doi:10.1089/ten.teb.2016.0181
17. Lewis JS. Rotator cuff tendinopathy. Br J Sports Med. 2009;43(4):236–241. doi:10.1136/bjsm.2008.052175
18. Citro V, Clerici M, Boccaccini AR, Della Porta G, Maffulli N, Forsyth NR. Tendon tissue engineering: an overview of biologics to promote tendon healing and repair. J Tissue Eng. 2023;14:20417314231196275. doi:10.1177/20417314231196275
19. Sharma P, Maffulli N. Tendinopathy and tendon injury: the future. Disabil Rehabil. 2008;30(20–22):1733–1745. doi:10.1080/09638280701788274
20. Wang Y, He G, Tang H, et al. Aspirin promotes tenogenic differentiation of tendon stem cells and facilitates tendinopathy healing through regulating the GDF7/Smad1/5 signaling pathway. J Cell Physiol. 2020;235(5):4778–4789. doi:10.1002/jcp.29355
21. Lui PPY. Markers for the identification of tendon-derived stem cells in vitro and tendon stem cells in situ – update and future development. Stem Cell Res Ther. 2015;6(1):106. doi:10.1186/s13287-015-0097-y
22. Ruzzini L, Abbruzzese F, Rainer A, et al. Characterization of age-related changes of tendon stem cells from adult human tendons. Knee Surg Sports Traumatol Arthrosc. 2014;22(11):2856–2866. doi:10.1007/s00167-013-2457-4
23. Yin Z, Jie HJ, Yang L, et al. Single-cell analysis reveals a nestin+ tendon stem/progenitor cell population with strong tenogenic potentiality. Sci Adv. 2016;2(11):e1600874. doi:10.1126/sciadv.1600874
24. Harvey T, Flamenco S, Fan CM. A Tppp3+Pdgfra+ tendon stem cell population contributes to regeneration and reveals a shared role for PDGF signalling in regeneration and fibrosis. Nat Cell Biol. 2019;21(12):1490–1503. doi:10.1038/s41556-019-0417-z
25. Walia B, Huang AH. Tendon stem progenitor cells: understanding the biology to inform therapeutic strategies for tendon repair. J Orthop Res. 2019;37(6):1270–1280. doi:10.1002/jor.24156
26. Winnicki K, Ochała-Kłos A, Rutowicz B, Pękala PA, Tomaszewski KA. Functional anatomy, histology and biomechanics of the human Achilles tendon — a comprehensive review. Ann Anat - Anat Anz. 2020;229:151461. doi:10.1016/j.aanat.2020.151461
27. Jozsa L, Lehto M, Kannus P, et al. Fibronectin and laminin in Achilles tendon. Acta Orthop Scand. 1989;60(4):469–471. doi:10.3109/17453678909149322
28. Adams JC. THROMBOSPONDINS: multifunctional regulators of cell interactions. Ann Rev Cell Dev Biol. 2001;17(1):25–51. doi:10.1146/annurev.cellbio.17.1.25
29. De Palma L, Marinelli M, Memè L, Pavan M. Immunohistochemistry of the enthesis organ of the human achilles tendon. Foot Ankle Int. 2004;25(6):414–418. doi:10.1177/107110070402500609
30. Docheva D, Hunziker EB, Fässler R, Brandau O. Tenomodulin is necessary for tenocyte proliferation and tendon maturation. Mol Cell Biol. 2005;25(2):699–705. doi:10.1128/MCB.25.2.699-705.2005
31. Oliva F, Marsilio E, Asparago G, Frizziero A, Berardi AC, Maffulli N. The impact of hyaluronic acid on tendon physiology and its clinical application in tendinopathies. Cells. 2021;10(11):3081. doi:10.3390/cells10113081
32. Zhang Y, Zhang E, Qin T, et al. Matrix stiffness-mediated tenogenesis of tendon stem/progenitor cells via integrin-αm for tendon regeneration. Biochem Biophys Res Commun. 2023;678:90–96. doi:10.1016/j.bbrc.2023.08.007
33. Screen HRC, Berk DE, Kadler KE, Ramirez F, Young MF. Tendon functional extracellular matrix. J Ortho Res. 2015;33(6):793–799. doi:10.1002/jor.22818
34. Florit D, Pedret C, Casals M, Malliaras P, Sugimoto D, Rodas G. Incidence of tendinopathy in team sports in a multidisciplinary sports club over 8 seasons. J Sports Sci Med. 2019;18(4):780–788. PMID: 31827363 ; PMCID: PMC6873129..
35. Kakouris N, Yener N, Fong DTP. A systematic review of running-related musculoskeletal injuries in runners. J Sport Health Sci. 2021;10(5):513–522. doi:10.1016/j.jshs.2021.04.001
36. Bertelsen ML, Hulme A, Petersen J, et al. A framework for the etiology of running‐related injuries. Scand J Med Sci Sports. 2017;27(11):1170–1180. doi:10.1111/sms.12883
37. Carroll R. Tennis elbow: incidence in local league players. Br J Sports Med. 1981;15(4):250–256. doi:10.1136/bjsm.15.4.250
38. Abrams GD, Renstrom PA, Safran MR. Epidemiology of musculoskeletal injury in the tennis player. Br J Sports Med. 2012;46(7):492–498. doi:10.1136/bjsports-2012-091164
39. Hopkins C, Fu SC, Chua E, et al. Critical review on the socio-economic impact of tendinopathy. Asia-Pac J Sports Med Arthrosc Rehabil Technol. 2016;4:9–20. doi:10.1016/j.asmart.2016.01.002
40. Riley G. The pathogenesis of tendinopathy. A molecular perspective. Rheumatology. 2004;43(2):131–142. doi:10.1093/rheumatology/keg448
41. Scott A, Zwerver J, Grewal N, et al. Lipids, adiposity and tendinopathy: is there a mechanistic link? Critical review. Br J Sports Med. 2015;49(15):984–988. doi:10.1136/bjsports-2014-093989
42. Abate M, Schiavone C, Salini V, Andia I. Occurrence of tendon pathologies in metabolic disorders. Rheumatology. 2013;52(4):599–608. doi:10.1093/rheumatology/kes395
43. Vaughn NH, Stepanyan H, Gallo RA, Dhawan A. Genetic factors in tendon injury: a systematic review of the literature. Orthop J Sports Med. 2017;5(8):232596711772441. doi:10.1177/2325967117724416
44. September AV, Schwellnus MP, Collins M. Tendon and ligament injuries: the genetic component. Br J Sports Med. 2007;41(4):241–246. doi:10.1136/bjsm.2006.033035
45. Sharma P, Maffulli N. Biology of tendon injury: healing, modeling and remodeling. J Musculoskelet Neuronal Interact. 2006;6(2):181–190. PMID: 16849830.
46. Lin TW, Cardenas L, Soslowsky LJ. Biomechanics of tendon injury and repair. J Biomech. 2004;37(6):865–877. doi:10.1016/j.jbiomech.2003.11.005
47. Oshiro W, Lou J, Xing X, Tu Y, Manske PR. Flexor tendon healing in the rat: a histologic and gene expression study. J Hand Surg. 2003;28(5):814–823. doi:10.1016/S0363-5023(03)00366-6
48. Docheva D, Müller SA, Majewski M, Evans CH. Biologics for tendon repair. Adv Drug Deliv Rev. 2015;84:222–239. doi:10.1016/j.addr.2014.11.015
49. Millar NL, Hueber AJ, Reilly JH, et al. Inflammation is present in early human tendinopathy. Am J Sports Med. 2010;38(10):2085–2091. doi:10.1177/0363546510372613
50. Tsuzaki M, Guyton G, Garrett W, et al. IL‐1β induces COX2, MMP‐1, ‐3 and ‐13, ADAMTS‐4, IL‐1β and IL‐6 in human tendon cells. J Orthop Res. 2003;21(2):256–264. doi:10.1016/S0736-0266(02)00141-9
51. Fu W, Yang R, Li J. Single-cell and spatial transcriptomics reveal changes in cell heterogeneity during progression of human tendinopathy. BMC Biol. 2023;21(1):132. doi:10.1186/s12915-023-01613-2
52. De La Durantaye M, Piette AB, Van Rooijen N, Frenette J. Macrophage depletion reduces cell proliferation and extracellular matrix accumulation but increases the ultimate tensile strength of injured Achilles tendons. J Orthop Res. 2014;32(2):279–285. doi:10.1002/jor.22504
53. Yang QQ, Zhang L, Zhou YL, Tang JB. Morphological changes of macrophages and their potential contribution to tendon healing. Colloid Surf B Biointerfaces. 2022;209:112145. doi:10.1016/j.colsurfb.2021.112145
54. Martinez FO. Macrophage activation and polarization. Front Biosci. 2008;13(13):453. doi:10.2741/2692
55. Lehner C, Spitzer G, Gehwolf R, et al. Tenophages: a novel macrophage-like tendon cell population expressing CX3CL1 and CX3CR1. Dis Model Mech. 2019:
56. Sugg KB, Lubardic J, Gumucio JP, Mendias CL. Changes in macrophage phenotype and induction of epithelial‐to‐mesenchymal transition genes following acute Achilles tenotomy and repair. J Orthop Res. 2014;32(7):944–951. doi:10.1002/jor.22624
57. Marsolais D, Cǒté CH, Frenette J. Neutrophils and macrophages accumulate sequentially following Achilles tendon injury. J Orthop Res. 2001;19(6):1203–1209. doi:10.1016/S0736-0266(01)00031-6
58. Koh TJ, DiPietro LA. Inflammation and wound healing: the role of the macrophage. Expert Rev Mol Med. 2011;13:e23. doi:10.1017/S1462399411001943
59. Chamberlain CS, Leiferman EM, Frisch KE, et al. The influence of macrophage depletion on ligament healing. Connect Tissue Res. 2011;52(3):203–211. doi:10.3109/03008207.2010.511355
60. Wang L, Li S, Xiao H, et al. TGF-β1 derived from macrophages contributes to load-induced tendon-bone healing in the murine rotator cuff repair model by promoting chondrogenesis. Bone Jt Res. 2023;12(3):219–230. doi:10.1302/2046-3758.123.BJR-2022-0368.R1
61. Li Y, Wang X, Hu B, et al. Neutralization of excessive levels of active TGF-β1 reduces MSC recruitment and differentiation to mitigate peritendinous adhesion. Bone Res. 2023;11(1):24. doi:10.1038/s41413-023-00252-1
62. Fadok VA, Bratton DL, Konowal A, Freed PW, Westcott JY, Henson PM. Macrophages That have ingested apoptotic cells in vitro inhibit proinflammatory cytokine production through autocrine/paracrine mechanisms involving TGF-, PGE2, and PAF. J Clin Invest. 1998;101(4):890–898. doi:10.1172/JCI1112
63. Liu M, Ng M, Phu T, et al. Polarized macrophages regulate fibro/adipogenic progenitor (FAP) adipogenesis through exosomes. Stem Cell Res Ther. 2023;14(1):321. doi:10.1186/s13287-023-03555-6
64. Sunwoo JY, Eliasberg CD, Carballo CB, Rodeo SA. The role of the macrophage in tendinopathy and tendon healing. J Orthop Res. 2020;38(8):1666–1675. doi:10.1002/jor.24667
65. Leon A, Levi-Montalcinit R, Dal Toso R. Mast cells synthesize, store, and release nerve growth factor. Proc Natl Acad Sci USA. 1994;91(9):3739–3743. doi:10.1073/pnas.91.9.3739
66. Alim A, Peterson M, Pejler G. Do mast cells have a role in tendon healing and inflammation? Cells. 2020;9(5):1134. doi:10.3390/cells9051134
67. D’Addona A, Maffulli N, Formisano S, Rosa D. Inflammation in tendinopathy. Surgeon. 2017;15(5):297–302. doi:10.1016/j.surge.2017.04.004
68. Millar NL, Gilchrist DS, Akbar M, et al. MicroRNA29a regulates IL-33-mediated tissue remodelling in tendon disease. Nat Commun. 2015;6(1):6774. doi:10.1038/ncomms7774
69. Millar NL, Akbar M, Campbell AL, et al. IL-17A mediates inflammatory and tissue remodelling events in early human tendinopathy. Sci Rep. 2016;6(1):27149. doi:10.1038/srep27149
70. Li H, Luo S, Wang H. The mechanisms and functions of TGF-β1 in tendon healing. Injury. 2023;54(11):111052. doi:10.1016/j.injury.2023.111052
71. Thornton GM, Shao X, Chung M, et al. Changes in mechanical loading lead to tendonspecific alterations in MMP and TIMP expression: influence of stress deprivation and intermittent cyclic hydrostatic compression on rat supraspinatus and Achilles tendons. Br J Sports Med. 2010;44(10):698–703. doi:10.1136/bjsm.2008.050575
72. Nakama LH, King KB, Abrahamsson S, Rempel DM. Evidence of tendon microtears due to cyclical loading in an in vivo tendinopathy model. J Orthop Res. 2005;23(5):1199–1205. doi:10.1016/j.orthres.2005.03.006
73. Bell R, Boniello MR, Gendron NR, Flatow EL, Andarawis‐Puri N. Delayed exercise promotes remodeling in sub‐rupture fatigue damaged tendons. J Orthop Res. 2015;33(6):919–925. doi:10.1002/jor.22856
74. Sorosky B, Press J, Plastaras C, Rittenberg J. The practical management of achilles tendinopathy. Clin J Sport Med. 2004;14(1):40–44. doi:10.1097/00042752-200401000-00007
75. Bestwick CS, Maffulli N. Reactive oxygen species and tendinopathy: do they matter? Br J Sports Med. 2004;38(6):672–674. doi:10.1136/bjsm.2004.012351
76. Magnusson SP, Langberg H, Kjaer M. The pathogenesis of tendinopathy: balancing the response to loading. Nat Rev Rheumatol. 2010;6(5):262–268. doi:10.1038/nrrheum.2010.43
77. Wren TAL, Lindsey DP, Beaupré GS, Carter DR. Effects of creep and cyclic loading on the mechanical properties and failure of human achilles tendons. Ann Biomed Eng. 2003;31(6):710–717. doi:10.1114/1.1569267
78. Abate M, Gravare-Silbernagel K, Siljeholm C, et al. Pathogenesis of tendinopathies: inflammation or degeneration? Arthritis Res Ther. 2009;11(3):235. doi:10.1186/ar2723
79. Zhang C, Zhu J, Zhou Y, Thampatty BP, Wang JHC. Tendon Stem/progenitor cells and their interactions with extracellular matrix and mechanical loading. Stem Cells Int. 2019;2019(1):3674647.
80. Zhang J, Wang JH. Production of PGE 2 increases in tendons subjected to repetitive mechanical loading and induces differentiation of tendon stem cells into non‐tenocytes. J Orthop Res. 2010;28(2):198–203. doi:10.1002/jor.20962
81. Zeng Y, Wang C, Lei J, et al. Spatiotemporally responsive cascade bilayer microneedles integrating local glucose depletion and sustained nitric oxide release for accelerated diabetic wound healing. Acta Pharm Sin B. 2024;14(11):5037–5052. doi:10.1016/j.apsb.2024.06.014
82. Kwan CK, Fu SC, Yung PS-H, et al. A high glucose level stimulate inflammation and weaken pro-resolving response in tendon cells – a possible factor contributing to tendinopathy in diabetic patients. Asia-Pac J Sports Med Arthrosc Rehabil Technol. 2020;19:1–6. doi:10.1016/j.asmart.2019.10.002
83. Ahmed AS, Schizas N, Li J, et al. Type 2 diabetes impairs tendon repair after injury in a rat model. J Appl Physiol. 2012;113(11):1784–1791. doi:10.1152/japplphysiol.00767.2012
84. Ahmed AS, Li J, Abdul AMD, et al. Compromised neurotrophic and angiogenic regenerative capability during tendon healing in a rat model of Type-II diabetes. PLoS One. 2017;12(1):e0170748. doi:10.1371/journal.pone.0170748
85. Song FC, Yuan JQ, Zhu MD, et al. High glucose represses the proliferation of tendon fibroblasts by inhibiting autophagy activation in tendon injury. Biosci Rep. 2022;42(3). doi:10.1042/BSR20210640
86. Ackermann PW, Hart DA. Metabolic Influences on Risk for Tendon Disorders. Vol. 920. Springer International Publishing;2016. doi:10.1007/978-3-319-33943-6
87. Lin YC, Li YJ, Rui YF, et al. The effects of high glucose on tendon-derived stem cells: implications of the pathogenesis of diabetic tendon disorders. Oncotarget. 2017;8(11):17518–17528. doi:10.18632/oncotarget.15418
88. Xu L, Xu K, Wu Z, et al. Pioglitazone attenuates advanced glycation end products‐induced apoptosis and calcification by modulating autophagy in tendon‐derived stem cells. J Cell Mol Med. 2020;24(3):2240–2251. doi:10.1111/jcmm.14901
89. Li Y, Fessel G, Georgiadis M, Snedeker JG. Advanced glycation end-products diminish tendon collagen fiber sliding. Matrix Biol. 2013;32(3–4):169–177. doi:10.1016/j.matbio.2013.01.003
90. Patel SH, Yue F, Saw SK, et al. Advanced glycation end-products suppress mitochondrial function and proliferative capacity of achilles tendon-derived fibroblasts. Sci Rep. 2019;9(1):12614. doi:10.1038/s41598-019-49062-8.
91. Yoshikawa T, Mifune Y, Inui A, et al. Influence of diabetes-induced glycation and oxidative stress on the human rotator cuff. Antioxidants. 2022;11(4):743. doi:10.3390/antiox11040743
92. Benson RT, McDonnell SM, Knowles HJ, Rees JL, Carr AJ, Hulley PA. Tendinopathy and tears of the rotator cuff are associated with hypoxia and apoptosis. J Bone Joint Surg Br. 2010;92-B(3):448–453. doi:10.1302/0301-620X.92B3.23074
93. Chen H, Ge H-A, Wu G-B, et al. Autophagy prevents oxidative stress-induced loss of self-renewal capacity and stemness in human tendon stem cells by reducing ROS accumulation. Cell Physiol Biochem. 2016;39(6):2227–2238. doi:10.1159/000447916
94. Shen H, Cheng L, Zheng Q, Liu W, Wang Y. Scavenging of reactive oxygen species can adjust the differentiation of tendon stem cells and progenitor cells and prevent ectopic calcification in tendinopathy. Acta Biomater. 2022;152:440–452. doi:10.1016/j.actbio.2022.09.007
95. Yoshida K, Itoigawa Y, Wada T. Association of superoxide‐induced oxidative stress with rotator cuff tears in human patients. J Orthop Res. 2020;38(1):212–218. doi:10.1002/jor.24472
96. Taylor B, Cheema A, Soslowsky L. Tendon pathology in hypercholesterolemia and familial hypercholesterolemia. Curr Rheumatol Rep. 2017;19(12):76. doi:10.1007/s11926-017-0704-2
97. Ribitsch I, Bileck A, Aldoshin AD, et al. Molecular mechanisms of fetal tendon regeneration versus adult fibrous repair. Int J Mol Sci. 2021;22(11):5619. doi:10.3390/ijms22115619
98. Howell K, Chien C, Bell R, et al. Novel model of tendon regeneration reveals distinct cell mechanisms underlying regenerative and fibrotic tendon healing. Sci Rep. 2017;7(1):45238. doi:10.1038/srep45238
99. Li J, Wang X, Yao Z, et al. NLRP3‐dependent crosstalk between pyroptotic macrophage and senescent cell orchestrates trauma‐induced heterotopic ossification during aberrant wound healing. Adv Sci. 2023;10(19):2207383. doi:10.1002/advs.202207383
100. Zhu X. Inflammation, epigenetics, and metabolism converge to cell senescence and ageing: the regulation and intervention. Signal Transduct Target Ther. 2021;6(1):245.
101. Calcinotto A, Kohli J, Zagato E, Pellegrini L, Demaria M, Alimonti A. Cellular senescence: aging, cancer, and injury. Physiol Rev. 2019;99(2):1047–1078. doi:10.1152/physrev.00020.2018
102. Singh A, Schurman SH, Bektas A, et al. Aging and inflammation. Cold Spring Harb Perspect Med. 2024;14(6):a041197. doi:10.1101/cshperspect.a041197
103. Kannus P, Józsa L. Histopathological changes preceding spontaneous rupture of a tendon. A controlled study of 891 patients. J Bone Joint Surg Am. 1991;73(10):1507–1525. doi:10.2106/00004623-199173100-00009
104. Sugiyama Y, Naito K, Goto K, et al. Effect of aging on the tendon structure and tendon‑associated gene expression in mouse foot flexor tendon. Biomed Rep. 2019. doi:10.3892/br.2019.1200
105. Buckley MR, Evans EB, Matuszewski PE, et al. Distributions of types I, II and III collagen by region in the human supraspinatus tendon. Connect Tissue Res. 2013;54(6):374–379. doi:10.3109/03008207.2013.847096
106. Li Y, Dai G, Shi L, et al. The potential roles of tendon stem/progenitor cells in tendon aging. Curr Stem Cell Res Ther. 2019;14(1):34–42. doi:10.2174/1574888X13666181017112233
107. Kuilman T, Michaloglou C, Vredeveld LCW, et al. Oncogene-induced senescence relayed by an interleukin-dependent inflammatory network. Cell. 2008;133(6):1019–1031. doi:10.1016/j.cell.2008.03.039
108. Acosta JC, Banito A, Wuestefeld T, et al. A complex secretory program orchestrated by the inflammasome controls paracrine senescence. Nat Cell Biol. 2013;15(8):978–990. doi:10.1038/ncb2784
109. Li X, Li C, Zhang W, Wang Y, Qian P, Huang H. Inflammation and aging: signaling pathways and intervention therapies. Signal Transduct Target Ther. 2023;8(1):239. doi:10.1038/s41392-023-01502-8
110. Arnesen SM, Lawson MA. Age-related changes in focal adhesions lead to altered cell behavior in tendon fibroblasts. Mech Ageing Dev. 2006;127(9):726–732. doi:10.1016/j.mad.2006.05.003
111. Huang K, Chen C, Dong S, et al. Differentiation ability of tendon-derived stem cells and histological characteristics of rotator cuff remnant on the greater tuberosity degenerated with age and chronicity. Arthrosc J Arthrosc Relat Surg. 2022;38(4):1037–1048. doi:10.1016/j.arthro.2021.09.027
112. Bitman-Lotan E, Orian A. Nuclear organization and regulation of the differentiated state. Cell Mol Life Sci. 2021;78(7):3141–3158. doi:10.1007/s00018-020-03731-4
113. Lai F, Wang J, Tang H, et al.
114. Lai F, Tang H, Wang J. Effects of aging on the histology and biochemistry of rat tendon healing. BMC Musculoskelet Disord. 2021;22(1). doi:10.1186/s12891-021-04838-w
115. Chaudhury S, Dines JS, Delos D, Warren RF, Voigt C, Rodeo SA. Role of fatty infiltration in the pathophysiology and outcomes of rotator cuff tears. Arthritis Care Res. 2012;64(1):76–82. doi:10.1002/acr.20552
116. Zhou Z, Akinbiyi T, Xu L, et al. Tendon‐derived stem/progenitor cell aging: defective self‐renewal and altered fate. Aging Cell. 2010;9(5):911–915. doi:10.1111/j.1474-9726.2010.00598.x
117. Lai F, Wang J, Tang H, et al. Adipogenic differentiation was inhibited by downregulation of PPARγ signaling pathway in aging tendon stem/progenitor cells. J Orthop Surg. 2021;16(1):614. doi:10.1186/s13018-021-02720-y
118. Fu SC, Rolf C, Cheuk YC, Lui PP, Chan KM. Deciphering the pathogenesis of tendinopathy: a three-stages process. BMC Sports Sci Med Rehabil. 2010;2(1):30. doi:10.1186/1758-2555-2-30
119. Pattanittum P, Turner T, Green S, Buchbinder R. Non-steroidal anti-inflammatory drugs (NSAIDs) for treating lateral elbow pain in adults. Cochrane Database Syst Rev. 2013;2013(5). doi:10.1002/14651858.CD003686.pub2
120. Mazières B, Rouanet S, Guillon Y, Scarsi C, Reiner V. Topical ketoprofen patch in the treatment of tendinitis: a randomized, double blind, placebo controlled study. J Rheumatol. 2005;32(8):1563–1570.
121. Petri M, Hufman SL, Waser G, Cui H, Snabes MC, Verburg KM. Celecoxib effectively treats patients with acute shoulder tendinitis/bursitis. J Rheumatol. 2004;31(8):1614–1620.
122. Smidt N, Van Der Windt DA, Assendelft WJ, Devillé WL, Korthals-de Bos IB, Bouter LM. Corticosteroid injections, physiotherapy, or a wait-and-see policy for lateral epicondylitis: a randomised controlled trial. Lancet. 2002;359(9307):657–662. doi:10.1016/S0140-6736(02)07811-X
123. Hay EM, Paterson SM, Lewis M, Hosie G, Croft P. Pragmatic randomised controlled trial of local corticosteroid injection and naproxen for treatment of lateral epicondylitis of elbow in primary care. BMJ. 1999;319(7215):964–968. doi:10.1136/bmj.319.7215.964
124. Arirachakaran A, Sukthuayat A, Sisayanarane T, Laoratanavoraphong S, Kanchanatawan W, Kongtharvonskul J. Platelet-rich plasma versus autologous blood versus steroid injection in lateral epicondylitis: systematic review and network meta-analysis. J Orthop Traumatol. 2016;17(2):101–112. doi:10.1007/s10195-015-0376-5
125. Paoloni JA, Appleyard RC, Nelson J, Murrell GAC. Topical Glyceryl trinitrate application in the treatment of chronic supraspinatus tendinopathy: a randomized, double-blinded, placebo-controlled clinical trial. Am J Sports Med. 2005;33(6):806–813. doi:10.1177/0363546504270998
126. Paoloni JA, Murrell GAC. Three-year follow up study of topical glyceryl trinitrate treatment of chronic noninsertional achilles tendinopathy. Foot Ankle Int. 2007;28(10):1064–1068. doi:10.3113/FAI.2007.1064
127. Zeisig E, Öhberg L, Alfredson H. Sclerosing polidocanol injections in chronic painful tennis elbow‐promising results in a pilot study. Knee Surg Sports Traumatol Arthrosc. 2006;14(11):1218–1224. doi:10.1007/s00167-006-0156-0
128. Alfredson H, Öhberg L. Neovascularisation in chronic painful patellar tendinosis—promising results after sclerosing neovessels outside the tendon challenge the need for surgery. Knee Surg Sports Traumatol Arthrosc. 2005;13(2):74–80. doi:10.1007/s00167-004-0549-x
129. D’Vaz AP, Ostor AJK, Speed CA, et al. Pulsed low-intensity ultrasound therapy for chronic lateral epicondylitis: a randomized controlled trial. Rheumatology. 2006;45(5):566–570. doi:10.1093/rheumatology/kei210
130. Reza Nourbakhsh M, Fearon FJ. An alternative approach to treating lateral epicondylitis. A randomized, placebo-controlled, double-blinded study. Clin Rehabil. 2008;22(7):601–609. doi:10.1177/0269215507088447
131. Rompe JD, Maffulli N. Repetitive shock wave therapy for lateral elbow tendinopathy (tennis elbow): a systematic and qualitative analysis. Br Med Bull. 2007;83(1):355–378. doi:10.1093/bmb/ldm019
132. Rompe JD, Furia J, Maffulli N. Eccentric loading versus eccentric loading plus shock-wave treatment for midportion achilles tendinopathy: a randomized controlled trial. Am J Sports Med. 2009;37(3):463–470. doi:10.1177/0363546508326983
133. Rompe JD, Nafe B, Furia JP, Maffulli N. Eccentric loading, shock-wave treatment, or a wait- and-see policy for tendinopathy of the main body of tendo achillis: a randomized controlled trial. Am J Sports Med. 2007;35(3):374–383. doi:10.1177/0363546506295940
134. Jonsson P, Alfredson H. Superior results with eccentric compared to concentric quadriceps training in patients with jumper’s knee: a prospective randomised study. Br J Sports Med. 2005;39(11):847–850. doi:10.1136/bjsm.2005.018630
135. Bjordal JM, Lopes-Martins RAB, Iversen VV. A randomised, placebo controlled trial of low level laser therapy for activated Achilles tendinitis with microdialysis measurement of peritendinous prostaglandin E2 concentrations. Br J Sports Med. 2006;40(1):76–80. doi:10.1136/bjsm.2005.020842
136. Andres BM, Murrell GAC. Treatment of tendinopathy: what works, what does not, and what is on the horizon. Clin Orthop. 2008;466(7):1539–1554. doi:10.1007/s11999-008-0260-1
137. Jones JG. Achilles tendon rupture following steroid injection. J Bone Joint Surg Am. 1985;67(1):170. doi:10.2106/00004623-198567010-00025
138. Chechick A, Amit Y, Israeli A, Horoszowski H. Recurrent rupture of the achilles tendon induced by corticosteroid injection. Br J Sports Med. 1982;16(2):89–90. doi:10.1136/bjsm.16.2.89
139. Mohamadi A, Chan JJ, Claessen FMAP, Ring D, Chen NC. Corticosteroid injections give small and transient pain relief in rotator cuff tendinosis: a meta-analysis. Clin Orthop. 2017;475(1):232–243. doi:10.1007/s11999-016-5002-1
140. Morberg P, Jerre R, Swärd L, Karlsson J. Long‐term results after surgical management of partial Achilles tendon ruptures. Scand J Med Sci Sports. 1997;7(5):299–303. doi:10.1111/j.1600-0838.1997.tb00157.x
141. Westin O, Nilsson Helander K, Grävare Silbernagel K, Samuelsson K, Brorsson A, Karlsson J. Patients with an Achilles tendon re-rupture have long-term functional deficits in function and worse patient-reported outcome than primary ruptures. Knee Surg Sports Traumatol Arthrosc. 2018;26(10):3063–3072. doi:10.1007/s00167-018-4952-0
142. Silva M, Ferreira FN, Alves NM, Paiva MC. Biodegradable polymer nanocomposites for ligament/tendon tissue engineering. J Nanobiotechnology. 2020;18(1):23. doi:10.1186/s12951-019-0556-1
143. Malafaya PB, Silva GA, Reis RL. Natural–origin polymers as carriers and scaffolds for biomolecules and cell delivery in tissue engineering applications☆. Adv Drug Deliv Rev. 2007;59(4–5):207–233. doi:10.1016/j.addr.2007.03.012
144. Kishore V, Bullock W, Sun X, Van Dyke WS, Akkus O. Tenogenic differentiation of human MSCs induced by the topography of electrochemically aligned collagen threads. Biomaterials. 2012;33(7):2137–2144. doi:10.1016/j.biomaterials.2011.11.066
145. Hubbell JA. Materials as morphogenetic guides in tissue engineering. Curr Opin Biotechnol. 2003;14(5):551–558. doi:10.1016/j.copbio.2003.09.004
146. Zhao T, Qi Y, Xiao S, et al. Integration of mesenchymal stem cell sheet and bFGF-loaded fibrin gel in knitted PLGA scaffolds favorable for tendon repair. J Mater Chem B. 2019;7(13):2201–2211. doi:10.1039/C8TB02759E
147. Hankemeier S, Hurschler C, Zeichen J, et al. Bone marrow stromal cells in a liquid fibrin matrix improve the healing process of patellar tendon window defects. Tissue Eng Part A. 2009;15(5):1019–1030. doi:10.1089/ten.tea.2008.0046
148. Li S, Dan X, Chen H, et al. Developing fibrin-based biomaterials/scaffolds in tissue engineering. Bioact Mater. 2024;40:597–623. doi:10.1016/j.bioactmat.2024.08.006
149. Ni T, Senthil‐Kumar P, Dubbin K, et al. A photoactivated nanofiber graft material for augmented Achilles tendon repair. Lasers Surg Med. 2012;44(8):645–652. doi:10.1002/lsm.22066
150. Thurber AE, Omenetto FG, Kaplan DL. In vivo bioresponses to silk proteins. Biomaterials. 2015;71:145–157. doi:10.1016/j.biomaterials.2015.08.039
151. Zhai P, Peng X, Li B, Liu Y, Sun H, Li X. The application of hyaluronic acid in bone regeneration. Int J Biol Macromol. 2020;151:1224–1239. doi:10.1016/j.ijbiomac.2019.10.169
152. Yue K, Trujillo-de Santiago G, Alvarez MM, Tamayol A, Annabi N, Khademhosseini A. Synthesis, properties, and biomedical applications of gelatin methacryloyl (GelMA) hydrogels. Biomaterials. 2015;73:254–271. doi:10.1016/j.biomaterials.2015.08.045
153. Catarino M, Castro F, Macedo JP, et al. Mechanisms of degradation of collagen or gelatin materials (hemostatic sponges) in oral surgery: a systematic review. Surgeries. 2024;5(3):532–548. doi:10.3390/surgeries5030043
154. Chen E, Yang L, Ye C. An asymmetric chitosan scaffold for tendon tissue engineering: in vitro and in vivo evaluation with rat tendon stem/progenitor cells. Acta Biomater. 2018;73:377–387. doi:10.1016/j.actbio.2018.04.027
155. Xu H, Zhu Y, Xu J, et al. Injectable bioactive glass/sodium alginate hydrogel with immunomodulatory and angiogenic properties for enhanced tendon healing. Bioeng Transl Med. 2023;8(1):e10345. doi:10.1002/btm2.10345
156. Namba J, Shimada K, Saito M, Murase T, Yamada H, Yoshikawa H. Modulation of peritendinous adhesion formation by alginate solution in a rabbit flexor tendon model. J Biomed Mater Res B Appl Biomater. 2007;80B(1):273–279. doi:10.1002/jbm.b.30594
157. Pawar SN, Edgar KJ. Alginate derivatization: a review of chemistry, properties and applications. Biomaterials. 2012;33(11):3279–3305. doi:10.1016/j.biomaterials.2012.01.007
158. Boontheekul T, Kong HJ, Mooney DJ. Controlling alginate gel degradation utilizing partial oxidation and bimodal molecular weight distribution. Biomaterials. 2005;26(15):2455–2465. doi:10.1016/j.biomaterials.2004.06.044
159. Ashton RS, Banerjee A, Punyani S, Schaffer DV, Kane RS. Scaffolds based on degradable alginate hydrogels and poly(lactide-co-glycolide) microspheres for stem cell culture. Biomaterials. 2007;28(36):5518–5525. doi:10.1016/j.biomaterials.2007.08.038
160. Wang S, Wang Y, Song L, et al. Decellularized tendon as a prospective scaffold for tendon repair. Mater Sci Eng C. 2017;77:1290–1301.
161. Lovati AB, Bottagisio M, Moretti M. Decellularized and engineered tendons as biological substitutes: a critical review. Stem Cells Int. 2016;2016(1):7276150.
162. Li J, Wen M, Zhang S, et al. Research progress on the preparation and application of decellularized tendons. Curr Issues Mol Biol. 2025;47(4):251.
163. Farah S, Anderson DG, Langer R. Physical and mechanical properties of PLA, and their functions in widespread applications — a comprehensive review. Adv Drug Deliv Rev. 2016;107:367–392. doi:10.1016/j.addr.2016.06.012
164. Chen CC, Chueh JY, Tseng H, Huang HM, Lee SY. Preparation and characterization of biodegradable PLA polymeric blends. Biomaterials. 2003;24(7):1167–1173. doi:10.1016/S0142-9612(02)00466-0
165. Lu L, Zhu X, Valenzuela RG, Currier BL, Yaszemski MJ. Biodegradable polymer scaffolds for cartilage tissue engineering. Clin Orthop. 2001;391:S251–S270. doi:10.1097/00003086-200110001-00024
166. Sabir MI, Xu X, Li L. A review on biodegradable polymeric materials for bone tissue engineering applications. J Mater Sci. 2009;44(21):5713–5724. doi:10.1007/s10853-009-3770-7
167. Hajiali H, Shahgasempour S, Naimi-Jamal MR, Peirovi H. Electrospun PGA/gelatin nanofibrous scaffolds and their potential application in vascular tissue engineering. Int J Nanomed. 2011;6:2133–2141. doi:10.2147/IJN.S24312
168. Zhao W, Li J, Jin K, Liu W, Qiu X, Li C. Fabrication of functional PLGA-based electrospun scaffolds and their applications in biomedical engineering. Mater Sci Eng C. 2016;59:1181–1194. doi:10.1016/j.msec.2015.11.026
169. Siddiqui N, Asawa S, Birru B, Baadhe R, Rao S. PCL-based composite scaffold matrices for tissue engineering applications. Mol Biotechnol. 2018;60(7):506–532. doi:10.1007/s12033-018-0084-5
170. Subramanian G, Stasuk A, Elsaadany M, Yildirim-Ayan E. Effect of uniaxial tensile cyclic loading regimes on matrix organization and tenogenic differentiation of adipose‐derived stem cells encapsulated within 3D collagen scaffolds. Stem Cells Int. 2017;2017. doi:10.1155/2017/6072406
171. Tang Y, Wang Z, Xiang L, Zhao Z, Cui W. Functional biomaterials for tendon/ligament repair and regeneration. Regen Biomater. 2022;9:rbac062. doi:10.1093/rb/rbac062
172. Maghdouri-White Y, Petrova S, Sori N. Electrospun silk–collagen scaffolds and BMP-13 for ligament and tendon repair and regeneration. Biomed Phys Eng Exp. 2018;4(2):025013. doi:10.1088/2057-1976/aa9c6f
173. Qian S, Wang Z, Zheng Z, Ran J, Zhu J, Chen W. A collagen and silk scaffold for improved healing of the tendon and bone interface in a rabbit model. Med Sci Monit. 2019;25:269–278. doi:10.12659/msm.912038
174. Islam A, Bohl MS, Tsai AG, Younesi M, Gillespie R, Akkus O. Biomechanical evaluation of a novel suturing scheme for grafting load-bearing collagen scaffolds for rotator cuff repair. Clin Biomech. 2015;30(7):669–675. doi:10.1016/j.clinbiomech.2015.05.007
175. Yuan H, Li X, Lee MS, et al. Collagen and chondroitin sulfate functionalized bioinspired fibers for tendon tissue engineering application. Int J Biol Macromol. 2021;170:248–260. doi:10.1016/j.ijbiomac.2020.12.152
176. Breidenbach AP, Dyment NA, Lu Y, et al. Fibrin gels exhibit improved biological, structural, and mechanical properties compared with collagen gels in cell-based tendon tissue-engineered constructs. Tissue Eng Part A. 2015;21(3–4):438–450. doi:10.1089/ten.tea.2013.0768
177. Barbon S, Stocco E, Macchi V, et al. Platelet-rich fibrin scaffolds for cartilage and tendon regenerative medicine: from bench to bedside. Int J Mol Sci. 2019;20(7):1701. doi:10.3390/ijms20071701
178. Matsunaga D, Akizuki S, Takizawa T, Omae S, Kato H. Compact platelet-rich fibrin scaffold to improve healing of patellar tendon defects and for medial collateral ligament reconstruction. The Knee. 2013;20(6):545–550. doi:10.1016/j.knee.2013.04.018
179. Wong CC, Huang YM, Chen CH, Lin FH, Yeh YY, Bai MY. Cytokine and growth factor delivery from implanted platelet-rich fibrin enhances rabbit achilles tendon healing. Int J Mol Sci. 2020;21(9):3221. doi:10.3390/ijms21093221
180. Kim SH, Hong H, Ajiteru O, et al. 3D bioprinted silk fibroin hydrogels for tissue engineering. Nat Protoc. 2021;16(12):5484–5532. doi:10.1038/s41596-021-00622-1
181. Chen J, Mo Q, Sheng R, et al. The application of human periodontal ligament stem cells and biomimetic silk scaffold for in situ tendon regeneration. Stem Cell Res Ther. 2021;12(1):596. doi:10.1186/s13287-021-02661-7
182. Xue Y, Kim H, Lee J, et al. Co‐electrospun silk fibroin and gelatin methacryloyl sheet seeded with mesenchymal stem cells for tendon regeneration. Small. 2022;18(21):2107714. doi:10.1002/smll.202107714
183. Micheli L, Parisio C, Lucarini E, et al. Restorative and pain-relieving effects of fibroin in preclinical models of tendinopathy. Biomed Pharmacother. 2022;148:112693. doi:10.1016/j.biopha.2022.112693
184. Kogan G, Šoltés L, Stern R, Gemeiner P. Hyaluronic acid: a natural biopolymer with a broad range of biomedical and industrial applications. Biotechnol Lett. 2006;29(1):17–25. doi:10.1007/s10529-006-9219-z
185. Greenberg DD, Stoker A, Kane S, Cockrell M, Cook JL. Biochemical effects of two different hyaluronic acid products in a co-culture model of osteoarthritis. Osteoarthritis Cartilage. 2006;14(8):814–822. doi:10.1016/j.joca.2006.02.006
186. Osti L, Berardocco M, Di Giacomo V, Di Bernardo G, Oliva F, Berardi AC. Hyaluronic acid increases tendon derived cell viability and collagen type I expression in vitro: comparative study of four different Hyaluronic acid preparations by molecular weight. BMC Musculoskelet Disord. 2015;16(1):284. doi:10.1186/s12891-015-0735-7
187. Cristino S, Grassi F, Toneguzzi S, et al. Analysis of mesenchymal stem cells grown on a three‐dimensional HYAFF 11®‐based prototype ligament scaffold. J Biomed Mater Res A. 2005;73A(3):275–283. doi:10.1002/jbm.a.30261
188. Honda H, Gotoh M, Kanazawa T, et al. Hyaluronic acid accelerates tendon-to-bone healing after rotator cuff repair. Am J Sports Med. 2017;45(14):3322–3330. doi:10.1177/0363546517720199
189. Chen CH, Chen SH, Chen SH, et al. Hyaluronic acid/platelet rich plasma-infused core-shell nanofiber membrane to prevent postoperative tendon adhesion and promote tendon healing. Int J Biol Macromol. 2023;231:123312. doi:10.1016/j.ijbiomac.2023.123312
190. Miescher I, Schaffner N, Rieber J, et al. Hyaluronic acid/PEO electrospun tube reduces tendon adhesion to levels comparable to native tendons – an in vitro and in vivo study. Int J Biol Macromol. 2024;273:133193. doi:10.1016/j.ijbiomac.2024.133193
191. Lim WL, Chowdhury SR, Ng MH, Law JX. Physicochemical properties and biocompatibility of electrospun polycaprolactone/gelatin nanofibers. Int J Environ Res Public Health. 2021;18(9):4764. doi:10.3390/ijerph18094764
192. Yang G, Lin H, Rothrauff BB, Yu S, Tuan RS. Multilayered polycaprolactone/gelatin fiber-hydrogel composite for tendon tissue engineering. Acta Biomater. 2016;35:68–76. doi:10.1016/j.actbio.2016.03.004
193. Rice JJ, Martino MM, De Laporte L, Tortelli F, Briquez PS, Hubbell JA. Engineering the regenerative microenvironment with biomaterials. Adv Healthc Mater. 2013;2(1):57–71. doi:10.1002/adhm.201200197
194. Zhang Q, Qiao Y, Li C, et al. Chitosan/gelatin-tannic acid decorated porous tape suture with multifunctionality for tendon healing. Carbohydr Polym. 2021;268:118246. doi:10.1016/j.carbpol.2021.118246
195. No YJ, Tarafder S, Reischl B, et al. High-strength fiber-reinforced composite hydrogel scaffolds as biosynthetic tendon graft material. ACS Biomater Sci Eng. 2020;6(4):1887–1898. doi:10.1021/acsbiomaterials.9b01716
196. Yuan X, Zhu Z, Xia P, et al. Tough gelatin hydrogel for tissue engineering. Adv Sci. 2023;10(24):2301665.
197. Yang Q, Li J, Meng H, et al. Coaxial electrospun nanofibrous membranes as dual‐functional biomimetic tendon sheath for tendon repair and anti‐peritendinous adhesion. Advanced Healthcare Materials. 2025;14(1):2402074.
198. Islam M, Shahruzzaman M, Biswas S, Nurus Sakib M, Rashid TU. Chitosan based bioactive materials in tissue engineering applications-A review. Bioact Mater. 2020;5(1):164–183. doi:10.1016/j.bioactmat.2020.01.012
199. Abd El-Hack ME, El-Saadony MT, Shafi ME, et al. Antimicrobial and antioxidant properties of chitosan and its derivatives and their applications: a review. Int J Biol Macromol. 2020;164:2726–2744. doi:10.1016/j.ijbiomac.2020.08.153
200. Freedman BR, Kuttler A, Beckmann N, et al. Enhanced tendon healing by a tough hydrogel with an adhesive side and high drug-loading capacity. Nat Biomed Eng. 2022;6(10):1167–1179. doi:10.1038/s41551-021-00810-0
201. Fakhraei O, Alimohammadi M, Moradi A, et al. Nanofibrous polycaprolactone/chitosan membranes for preventing postsurgical tendon adhesion. J Biomed Mater Res B Appl Biomater. 2022;110(6):1279–1291. doi:10.1002/jbm.b.34999
202. Chen Q, Lu H, Yang H. Chitosan prevents adhesion during rabbit flexor tendon repair via the sirtuin 1 signaling pathway. Mol Med Rep. 2015;12(3):4598–4603.
203. Far BF. Synthesis and characterization of chitosan/collagen/polycaprolactone hydrogel films with enhanced biocompatibility and hydrophilicity for artificial tendon applications. Int J Biol Macromol. 2023;253:127448.
204. Tomić S, Babić Radić MM, Vuković JS, Filipović VV, Nikodinovic-Runic J, Vukomanović M. Alginate-based hydrogels and scaffolds for biomedical applications. Mar Drugs. 2023;21(3):177. doi:10.3390/md21030177
205. Hojabri M, Tayebi T, Kasravi M. Wet-spinnability and crosslinked fiber properties of alginate/hydroxyethyl cellulose with varied proportion for potential use in tendon tissue engineering. Int J Biol Macromol. 2023;240:124492. doi:10.1016/j.ijbiomac.2023.124492
206. Song W, Ma Z, Wang X, et al. Macroporous granular hydrogels functionalized with aligned architecture and small extracellular vesicles stimulate osteoporotic tendon‐to‐bone healing. Adv Sci. 2023;10(34). doi:10.1002/advs.202304090
207. Yao Z, Qian Y, Jin Y, et al. Biomimetic multilayer polycaprolactone/sodium alginate hydrogel scaffolds loaded with melatonin facilitate tendon regeneration. Carbohydr Polym. 2022;277:118865. doi:10.1016/j.carbpol.2021.118865
208. Jayasree A, Kottappally Thankappan S, Ramachandran R, et al. Bioengineered braided micro–nano (multiscale) fibrous scaffolds for tendon reconstruction. ACS Biomater Sci Eng. 2019;5(3):1476–1486. doi:10.1021/acsbiomaterials.8b01328
209. Solis-Cordova J, Edwards JH, Fermor HL, Riches P, Brockett CL, Herbert A. Characterisation of native and decellularised porcine tendon under tension and compression: a closer look at glycosaminoglycan contribution to tendon mechanics. J Mech Behav Biomed Mater. 2023;139:105671. doi:10.1016/j.jmbbm.2023.105671
210. Lohan A, Kohl B, Meier C, Schulze-Tanzil G. Tenogenesis of decellularized porcine achilles tendon matrix reseeded with human tenocytes in the nude mice xenograft model. Int J Mol Sci. 2018;19(7):2059. doi:10.3390/ijms19072059
211. Tao M, Liang F, He J, et al. Decellularized tendon matrix membranes prevent post-surgical tendon adhesion and promote functional repair. Acta Biomater. 2021;134:160–176. doi:10.1016/j.actbio.2021.07.038
212. Iwasaki N, Roldo M, Karali A, Blunn G. In vitro development of a muscle-tendon junction construct using decellularised extracellular matrix: effect of cyclic tensile loading. Biomater Adv. 2024;161:213873. doi:10.1016/j.bioadv.2024.213873
213. Jiang D, Xu B, Gao P. Effects of young extracellular matrix on the biological characteristics of aged tendon stem cells. Adv Clin Exp Med. 2018;27(12):1625–1630.
214. Huang S, Rao Y, Zhou M, et al. Engineering an extracellular matrix-functionalized, load-bearing tendon substitute for effective repair of large-to-massive tendon defects. Bioact Mater. 2024;36:221–237. doi:10.1016/j.bioactmat.2024.02.032
215. Ilaltdinov AW, Gong Y, Leong DJ, et al. Advances in the development of gene therapy, noncoding RNA, and exosome‐based treatments for tendinopathy. Ann N Y Acad Sci. 2021;1490(1):3–12. doi:10.1111/nyas.14382
216. Bolt P, Clerk AN, Luu HH, et al. BMP-14 gene therapy increases tendon tensile strength in a rat model of achilles tendon injury. J Bone Jt Surg. 2007;89(6):1315–1320. doi:10.2106/JBJS.F.00257
217. Hasslund S, Dadali T, Ulrich-Vinther M, Søballe K, Schwarz EM, Awad HA. Freeze-dried allograft-mediated gene or protein delivery of growth and differentiation factor 5 reduces reconstructed murine flexor tendon adhesions. J Tissue Eng. 2014;5. doi:10.1177/2041731414528736
218. Jin J, Yang QQ, Zhou YL. Non-viral delivery of gene therapy to the tendon. Polymers. 2022;14(16):3338. doi:10.3390/polym14163338
219. Wu G, Sun B, Zhao C, et al. Three-dimensional tendon scaffold loaded with TGF-β1 gene silencing plasmid prevents tendon adhesion and promotes tendon repair. ACS Biomater Sci Eng. 2021;7(12):5739–5748. doi:10.1021/acsbiomaterials.1c00747
220. Eltom A, Zhong G, Muhammad A. Scaffold techniques and designs in tissue engineering functions and purposes: a review. Adv Mater Sci Eng. 2019;2019:1–13. doi:10.1155/2019/3429527
221. Gross G, Hoffmann A. Therapeutic strategies for tendon healing based on novel biomaterials, factors and cells. Pathobiology. 2013;80(4):203–210. doi:10.1159/000347059
222. Patel A, Zaky SH, Schoedel K, et al. Design and evaluation of collagen-inspired mineral-hydrogel nanocomposites for bone regeneration. Acta Biomater. 2020;112:262–273. doi:10.1016/j.actbio.2020.05.034
223. Murariu M, Dubois P. PLA composites: from production to properties. Adv Drug Deliv Rev. 2016;107:17–46. doi:10.1016/j.addr.2016.04.003
224. Xie Y, Zhang F, Akkus O, King MW. A collagen/PLA hybrid scaffold supports tendon‐derived cell growth for tendon repair and regeneration. J Biomed Mater Res B Appl Biomater. 2022;110(12):2624–2635. doi:10.1002/jbm.b.35116
225. Deepthi S, Nivedhitha sundaram M, Deepti Kadavan J, Jayakumar R. Layered chitosan-collagen hydrogel/aligned PLLA nanofiber construct for flexor tendon regeneration. Carbohydr Polym. 2016;153:492–500. doi:10.1016/j.carbpol.2016.07.124
226. Zebiri H, Chammas E, Pompee C, Chammas M, Garric X. Synthesis of PLA–poly(ether urethane)–PLA copolymers and design of biodegradable anti-adhesive membranes for orthopaedic applications. J Mater Chem B. 2021;9(3):832–845. doi:10.1039/D0TB02545C
227. Li Y, Hu C, Hu B, et al. Sustained release of dicumarol via novel grafted polymer in electrospun nanofiber membrane for treatment of peritendinous adhesion. Adv Health Mater. 2023;12(15). doi:10.1002/adhm.202203078
228. Liu S, Chen H, Wu T, et al. Macrophage infiltration of electrospun polyester fibers. Biomater Sci. 2017;5(8):1579–1587. doi:10.1039/C6BM00958A
229. Yokoya S, Mochizuki Y, Nagata Y, Deie M, Ochi M. Tendon-bone insertion repair and regeneration using polyglycolic acid sheet in the rabbit rotator cuff injury model. Am J Sports Med. 2008;36(7):1298–1309. doi:10.1177/0363546508314416
230. Romeo A, Easley J, Regan D, et al. Rotator cuff repair using a bioresorbable nanofiber interposition scaffold: a biomechanical and histologic analysis in sheep. J Shoulder Elbow Surg. 2022;31(2):402–412. doi:10.1016/j.jse.2021.07.018
231. Wu S, Zhou R, Zhou F, Streubel PN, Chen S, Duan B. Electrospun thymosin Beta-4 loaded PLGA/PLA nanofiber/ microfiber hybrid yarns for tendon tissue engineering application. Mater Sci Eng C. 2020;106:110268. doi:10.1016/j.msec.2019.110268
232. Criscenti G, Longoni A, Di Luca A, et al. Triphasic scaffolds for the regeneration of the bone–ligament interface. Biofabrication. 2016;8(1):015009. doi:10.1088/1758-5090/8/1/015009
233. Kim K, Kim H, Do S, Kim H. Potential of aligned electrospun PLGA/SIS blended nanofibrous membrane for tendon tissue engineering. Polymers. 2023;15(10):2313. doi:10.3390/polym15102313
234. Kolluru PV, Lipner J, Liu W, et al. Strong and tough mineralized PLGA nanofibers for tendon-to-bone scaffolds. Acta Biomater. 2013;9(12):9442–9450. doi:10.1016/j.actbio.2013.07.042
235. Shen Y, Tu T, Yi B, et al. Electrospun acid-neutralizing fibers for the amelioration of inflammatory response. Acta Biomater. 2019;97:200–215. doi:10.1016/j.actbio.2019.08.014
236. Liu C, Tian S, Bai J, et al. Regulation of ERK1/2 and SMAD2/3 pathways by using multi-layered electrospun PCL–amnion nanofibrous membranes for the prevention of post-surgical tendon adhesion. Int J Nanomed. 2020;15:927–942. doi:10.2147/IJN.S231538
237. Ye J, Wang Y, Zeng W, et al. Local injection of rapamycin‐loaded Pcl‐Peg nanoparticles for enhanced tendon healing in rotator cuff tears via simultaneously reducing fatty infiltration and drug toxicity. Adv Healthc Mater. 2023:2300612. doi:10.1002/adhm.202300612
238. Annabi N, Tamayol A, Uquillas JA, et al. 25th anniversary article: rational design and applications of hydrogels in regenerative medicine. Adv Mater. 2014;26(1):85–124. doi:10.1002/adma.201303233
239. Kim J, Seo BB, Hong KH, Kim SE, Kim YM, Song SC. Long-term anti-inflammatory effects of injectable celecoxib nanoparticle hydrogels for Achilles tendon regeneration. Acta Biomater. 2022;144:183–194. doi:10.1016/j.actbio.2022.03.033
240. Cai C, Zhang X, Li Y, et al. Self‐healing hydrogel embodied with macrophage‐regulation and responsive‐gene‐silencing properties for synergistic prevention of peritendinous adhesion. Adv Mater. 2022;34(5). doi:10.1002/adma.202106564
241. Ren Z, Duan Z, Zhang Z, Fu R, Zhu C, Fan D. Instantaneous self-healing and strongly adhesive self-adaptive hyaluronic acid-based hydrogel for controlled drug release to promote tendon wound healing. Int J Biol Macromol. 2023;242:125001. doi:10.1016/j.ijbiomac.2023.125001
242. Barzegar PEF, Mohammadi Z, Sattari S, et al. Graphene-MoS2 polyfunctional hybrid hydrogels for the healing of transected Achilles tendon. Biomater Adv. 2022;137:212820. doi:10.1016/j.bioadv.2022.212820
243. Iii RNK, Huang AH, Baker BM. Augmentation of tendon and ligament repair with fiber‐reinforced hydrogel composites. Adv Health Mater. 2024;13(29):2400668.
244. Snedeker JG, Foolen J. Tendon injury and repair – a perspective on the basic mechanisms of tendon disease and future clinical therapy. Acta Biomater. 2017;63:18–36. doi:10.1016/j.actbio.2017.08.032
245. Zhu H, Wang C, Yang Y, et al. High-strength mechanically gradient hydrogels via physical crosslinking for tendon-mimetic tissue repair. NPJ Flex Electron. 2025;9(1):53. doi:10.1038/s41528-025-00430-7
246. Hamdi H, Planat-Benard V, Bel A, et al. Epicardial adipose stem cell sheets results in greater post-infarction survival than intramyocardial injections. Cardiovasc Res. 2011;91(3):483–491. doi:10.1093/cvr/cvr099
247. Ni M, Lui PPY, Rui YF, et al. Tendon‐derived stem cells (TDSCs) promote tendon repair in a rat patellar tendon window defect model. J Orthop Res. 2012;30(4):613–619. doi:10.1002/jor.21559
248. Ning LJ, Zhang YJ, Zhang YJ, et al. Enhancement of migration and tenogenic differentiation of macaca mulatta tendon-derived stem cells by decellularized tendon hydrogel. Front Cell Dev Biol. 2021;9:651583. doi:10.3389/fcell.2021.651583
249. Zhang H, Dai Y, Long H, et al. Tendon stem/progenitor cell-laden nanofiber hydrogel enhanced functional repair of patellar tendon. Tissue Eng Part A. 2023;29(5–6):150–160. doi:10.1089/ten.tea.2022.0183
250. Yang Q, Li J, Su W, et al. Electrospun aligned poly(ε-caprolactone) nanofiber yarns guiding 3D organization of tendon stem/progenitor cells in tenogenic differentiation and tendon repair. Front Bioeng Biotechnol. 2022;10:960694. doi:10.3389/fbioe.2022.960694
251. Mao X, Yao L, Li M, et al. Enhancement of tendon repair using tendon-derived stem cells in small intestinal submucosa via M2 macrophage polarization. Cells. 2022;11(17):2770. doi:10.3390/cells11172770
252. Tan Q, Lui PPY, Rui YF. Effect of in vitro passaging on the stem cell-related properties of tendon-derived stem cells—implications in tissue engineering. Stem Cells Dev. 2012;21(5):790–800. doi:10.1089/scd.2011.0160
253. Liu H, Zhu S, Zhang C, et al. Crucial transcription factors in tendon development and differentiation: their potential for tendon regeneration. Cell Tissue Res. 2014;356(2):287–298. doi:10.1007/s00441-014-1834-8
254. Zhang H, Liu MF, Liu RC, Shen WL, Yin Z, Chen X. Physical microenvironment-based inducible scaffold for stem cell differentiation and tendon regeneration. Tissue Eng Part B Rev. 2018;24(6):443–453. doi:10.1089/ten.teb.2018.0018
255. Chen Z, Chen P, Zheng M. Challenges and perspectives of tendon-derived cell therapy for tendinopathy: from bench to bedside. Stem Cell Res Ther. 2022;13(1). doi:10.1186/s13287-022-03113-6
256. Rui YF, Lui PPY, Li G, Fu SC, Lee YW, Chan KM. Isolation and characterization of multipotent rat tendon-derived stem cells. Tissue Eng Part A. 2010;16(5):1549–1558. doi:10.1089/ten.tea.2009.0529
257. Roßbach BP, Gülecyüz MF, Kempfert L, et al. Rotator cuff repair with autologous tenocytes and biodegradable collagen scaffold: a histological and biomechanical study in sheep. Am J Sports Med. 2020;48(2):450–459. doi:10.1177/0363546519892580
258. Bucher TA, Ebert JR, Smith A, et al. Autologous tenocyte injection for the treatment of chronic recalcitrant gluteal tendinopathy. Orthop J Sports Med. 2017;5(2). doi:10.1177/2325967116688866
259. Holladay C, Abbah SA, O’Dowd C, Pandit A, Zeugolis DI. Preferential tendon stem cell response to growth factor supplementation: growth factor supplementation of tendon stem cell culture. J Tissue Eng Regen Med. 2016;10(9):783–798. doi:10.1002/term.1852
260. Wu S. Tendon-bioinspired wavy nanofibrous scaffolds provide tunable anisotropy and promote tenogenesis for tendon tissue engineering. Mater Sci. 2021;2021:1.
261. Burk J, Wittenberg-Voges L, Schubert S, Horstmeier C, Brehm W, Geburek F. Treatment of naturally occurring tendon disease with allogeneic multipotent mesenchymal stromal cells: a randomized, controlled, triple-blinded pilot study in horses. Cells. 2023;12(21):2513. doi:10.3390/cells12212513
262. Soler R, Rodas G, Rius-Tarruella J, et al. Safety and efficacy of bone marrow–derived mesenchymal stem cells for chronic patellar tendinopathy (with gap >3 mm) in patients: 12-month follow-up results of a phase 1/2 clinical trial. Orthop J Sports Med. 2023;11(9):23259671231184400. doi:10.1177/23259671231184400
263. Chen P, Cui L, Fu SC, et al. The 3D-printed PLGA scaffolds loaded with bone marrow-derived mesenchymal stem cells augment the healing of rotator cuff repair in the rabbits. Cell Transplant. 2020;29:0963689720973647
264. Jiang L, Lu J, Chen Y, et al. Mesenchymal stem cells: an efficient cell therapy for tendon repair (Review). Int J Mol Med. 2023;52(2):70. doi:10.3892/ijmm.2023.5273
265. O. Bortolazzo F, D. Lucke L, De oliveira fujii L, et al. Microcurrent and adipose‐derived stem cells modulate genes expression involved in the structural recovery of transected tendon of rats. FASEB J. 2020;34(8):10011–10026. doi:10.1096/fj.201902942RR
266. Song W, Zhang D, Wu D, et al. Cryopreserved adipose-derived stem cell sheets. Am J Sports Med. 2023;51(8):2005–2017. doi:10.1177/03635465231171682
267. Shin MJ, Shim IK, Kim DM, et al. Engineered cell sheets for the effective delivery of adipose-derived stem cells for tendon-to-bone healing. Am J Sports Med. 2020;48(13):3347–3358. doi:10.1177/0363546520964445
268. Wu Z, Yang J, Chong H, et al. 3D-printed biomimetic scaffolds loaded with ADSCs and BMP-2 for enhanced rotator cuff repair. J Mater Chem B. 2024;12(47):12365–12377. doi:10.1039/D4TB01073F
269. Uysal AC, Mizuno H. Tendon regeneration and repair with adipose derived stem cells. Curr Stem Cell Res Ther. 2010;999(999):1–7. doi:10.2174/157421019941450888X
270. Lee JH, Kim YH, Rhee SM, et al. Rotator cuff tendon healing using human dermal fibroblasts: histological and biomechanical analyses in a rabbit model of chronic rotator cuff tears. Am J Sports Med. 2021;49(13):3669–3679. doi:10.1177/03635465211041102
271. Rhee SM, Jeon S, Han J, et al. The effect of combining hyaluronic acid and human dermal fibroblasts on tendon healing. Am J Sports Med. 2023;51(12):3243–3250. doi:10.1177/03635465231191779
272. Gaspar D, Spanoudes K, Holladay C, Pandit A, Zeugolis D. Progress in cell-based therapies for tendon repair. Adv Drug Deliv Rev. 2015;84:240–256. doi:10.1016/j.addr.2014.11.023
273. Berry DP, Harding KG, Stanton MR, Jasani B, Ehrlich PH. Human wound contraction: collagen organization, fibroblasts, and myofibroblasts. Plast Reconstr Surg. 1998;102(1):132–134. doi:10.1097/00006534-199807000-00020
274. Polo JM, Liu S, Figueroa ME, et al. Cell type of origin influences the molecular and functional properties of mouse induced pluripotent stem cells. Nat Biotechnol. 2010;28(8):848–855. doi:10.1038/nbt.1667
275. Yuan Z, Yu H, Long H, et al. Stem cell applications and tenogenic differentiation strategies for tendon repair. Stem Cells Int. 2023;2023:1–15. doi:10.1155/2023/3656498
276. Bavin EP, Smith O, Baird AEG, Smith LC, Guest DJ. Equine induced pluripotent stem cells have a reduced tendon differentiation capacity compared to embryonic stem cells. Front Vet Sci. 2015;2. doi:10.3389/fvets.2015.00055
277. Zhang C, Yuan H, Liu H. Well-aligned chitosan-based ultrafine fibers committed teno-lineage differentiation of human induced pluripotent stem cells for Achilles tendon regeneration. Biomaterials. 2015;53:716–730. doi:10.1016/j.biomaterials.2015.02.051
278. Vo TN, Kasper FK, Mikos AG. Strategies for controlled delivery of growth factors and cells for bone regeneration. Adv Drug Deliv Rev. 2012;64(12):1292–1309. doi:10.1016/j.addr.2012.01.016
279. Chalidis B, Givissis P, Papadopoulos P, Pitsilos C. Molecular and biologic effects of Platelet-Rich Plasma (PRP) in ligament and tendon healing and regeneration: a systematic review. Int J Mol Sci. 2023;24(3):2744. doi:10.3390/ijms24032744
280. Chen X, Jones IA, Park C, Vangsness CT. The efficacy of platelet-rich plasma on tendon and ligament healing: a systematic review and meta-analysis with bias assessment. Am J Sports Med. 2018;46(8):2020–2032. doi:10.1177/0363546517743746
281. Liu X, Li Y, Shen L, Yan M. Leukocyte and Platelet-Rich Plasma (L-PRP) in tendon models: a systematic review and meta-analysis of in vivo/in vitro studies. Evid Based Complement Alternat Med. 2022;2022:1–15. doi:10.1155/2022/5289145
282. Hurley ET, Colasanti CA, Anil U, et al. The effect of platelet-rich plasma leukocyte concentration on arthroscopic rotator cuff repair: a network meta-analysis of randomized controlled trials. Am J Sports Med. 2021;49(9):2528–2535. doi:10.1177/0363546520975435
283. Jiang G, Wu Y, Meng J. Comparison of leukocyte-rich platelet-rich plasma and leukocyte-poor platelet-rich plasma on achilles tendinopathy at an early stage in a rabbit model. Am J Sports Med. 2020;48(5):1189–1199. doi:10.1177/0363546520906142
284. Zhang C, Cai YZ, Wang Y. Injection of leukocyte-poor platelet-rich plasma for moderate-to-large rotator cuff tears does not improve clinical outcomes but reduces retear rates and fatty infiltration: a prospective, single-blinded randomized study. Arthrosc J Arthrosc Relat Surg. 2022;38(8):2381–2388.e1. doi:10.1016/j.arthro.2022.02.007
285. Zhang M, Zhang L. Effect of autologous platelet-rich plasma and gelatin sponge for tendon-to-bone healing after rabbit anterior cruciate ligament reconstruction.
286. Kesikburun S, Tan AK, Yılmaz B, Yaşar E, Yazıcıoğlu K. Platelet-rich plasma injections in the treatment of chronic rotator cuff tendinopathy: a randomized controlled trial with 1-year follow-up. Am J Sports Med. 2013;41(11):2609–2616. doi:10.1177/0363546513496542
287. Le ADK, Enweze L, DeBaun MR, Dragoo JL. Current clinical recommendations for use of platelet-rich plasma. Curr Rev Musculoskelet Med. 2018;11(4):624–634. doi:10.1007/s12178-018-9527-7
288. Evrova O, Buschmann J. In vitro and in vivo effects of PDGF-BB delivery strategies on tendon healing: a review. Eur Cell Mater. 2017;34:15–39. doi:10.22203/eCM.v034a02
289. Thomopoulos S, Zaegel M, Das R, et al. PDGF‐BB released in tendon repair using a novel delivery system promotes cell proliferation and collagen remodeling. J Orthop Res. 2007;25(10):1358–1368. doi:10.1002/jor.20444
290. Perikamana SKM, Lee J, Kim EM, Byun H, Lee S, Shin H. Harnessing biochemical and structural cues for tenogenic differentiation of adipose derived stem cells (ADSCs) and development of an in vitro tissue interface mimicking tendon-bone insertion graft. Acta Biomater. 2018;74:464–477. doi:10.1016/j.actbio.2018.05.035
291. Disser NP, Sugg KB, Talarek JR, Sarver DC, Rourke BJ, Mendias CL. Insulin‐like growth factor 1 signaling in tenocytes is required for adult tendon growth. FASEB J. 2019;33(11):12680–12695. doi:10.1096/fj.201901503R
292. Rieber J, Meier-Bürgisser G, Miescher I, et al. Bioactive and elastic emulsion electrospun DegraPol tubes delivering IGF-1 for tendon rupture repair. Int J Mol Sci. 2023;24(12):10272. doi:10.3390/ijms241210272
293. Wang Y, Li J. Current progress in growth factors and extracellular vesicles in tendon healing. Int Wound J. 2023;20(9):3871–3883. doi:10.1111/iwj.14261
294. Miescher I, Rieber J, Calcagni M, Buschmann J. In vitro and in vivo effects of IGF-1 delivery strategies on tendon healing: a review. Int J Mol Sci. 2023;24(3):2370. doi:10.3390/ijms24032370
295. Wang (王文波) W, Li (李洁) J, Wang (王科云) K, et al. Induction of predominant tenogenic phenotype in human dermal fibroblasts via synergistic effect of TGF-β and elongated cell shape. Am J Physiol-Cell Physiol. 2016;310(5):C357–C372. doi:10.1152/ajpcell.00300.2015
296. Liu Y, Feng L, Xu J, et al. MiR-378a suppresses tenogenic differentiation and tendon repair by targeting at TGF-β2. Stem Cell Res Ther. 2019;10(1):108. doi:10.1186/s13287-019-1216-y
297. Arvinius C, Civantos A, Rodríguez-Bobada C, et al. Enhancement of in vivo supraspinatus tendon–to-bone healing with an alginate-chitin scaffold and rhBMP-2. Injury. 2021;52(1):78–84. doi:10.1016/j.injury.2020.11.019
298. Han L. Sustained BMP-2 release and platelet rich fibrin synergistically promote tendon-bone healing after anterior cruciate ligament reconstruction in rat. European Review for Medical & Pharmacological Sciences. 2019;23(20):1.
299. Williams LA, Bhargav D, Diwan AD. Unveiling the Bmp13 enigma: redundant morphogen or crucial regulator? Int J Biol Sci. 2008;318–329. doi:10.7150/ijbs.4.318
300. Zhao J, Zhao S, Zhao J, et al. Biological augmentation of rotator cuff repair using bFGF-loaded electrospun poly(lactide-co-glycolide) fibrous membranes. Int J Nanomed. 2014:2373. doi:10.2147/IJN.S59536
301. Darshan TG, Chen CH, Kuo CY, et al. Development of high resilience spiral wound suture-embedded gelatin/PCL/heparin nanofiber membrane scaffolds for tendon tissue engineering. Int J Biol Macromol. 2022;221:314–333. doi:10.1016/j.ijbiomac.2022.09.001
302. Sahoo S, Toh SL, Goh JCH. A bFGF-releasing silk/PLGA-based biohybrid scaffold for ligament/tendon tissue engineering using mesenchymal progenitor cells. Biomaterials. 2010;31(11):2990–2998. doi:10.1016/j.biomaterials.2010.01.004
303. Lu J, Jiang L, Chen Y, et al. The functions and mechanisms of basic fibroblast growth factor in tendon repair. Front Physiol. 2022;13:852795. doi:10.3389/fphys.2022.852795
304. Usman MA, Nakasa T, Shoji T, et al. The effect of administration of double stranded MicroRNA-210 on acceleration of Achilles tendon healing in a rat model. J Orthop Sci. 2015;20(3):538–546. doi:10.1007/s00776-015-0709-5
305. Lu YF, Liu Y, Fu WM, et al. Long noncoding RNA H19 accelerates tenogenic differentiation and promotes tendon healing through targeting miR-29b-3p and activating TGF-b1 signaling. FASEB J. 2017;31(3):954–964.
306. Tang JB, Zhou YL, Wu YF, Liu PY, Wang XT. Gene therapy strategies to improve strength and quality of flexor tendon healing. Expert Opin Biol Ther. 2016;16(3):291–301. doi:10.1517/14712598.2016.1134479
307. Geng Y, Zhao X, Xu J, et al. Overexpression of mechanical sensitive miR-337-3p alleviates ectopic ossification in rat tendinopathy model via targeting IRS1 and Nox4 of tendon-derived stem cells. J Mol Cell Biol. 2020;12(4):305–317. doi:10.1093/jmcb/mjz030
308. Lee JY, Zhou Z, Taub PJ, et al. BMP-12 treatment of adult mesenchymal stem cells in vitro augments tendon-like tissue formation and defect repair in vivo. PLoS One. 2011;6(3):1.
309. Tan SL, Chan CK, Ahmad TS, et al. Growth differentiation factor 5-induced mesenchymal stromal cells enhance tendon healing. Tissue Eng C Method. 2024;30(10):431–442.
310. Otabe K, Nakahara H, Hasegawa A, et al. Transcription factor Mohawk controls tenogenic differentiation of bone marrow mesenchymal stem cells in vitro and in vivo. J Orthop Res. 2015;33(1):1–8. doi:10.1002/jor.22750
311. Chen X, Yin Z, Chen JL, et al. Scleraxis -overexpressed human embryonic stem cell–derived mesenchymal stem cells for tendon tissue engineering with knitted silk-collagen scaffold. Tissue Eng Part A. 2014;20(11–12):1583–1592. doi:10.1089/ten.tea.2012.0656
312. Giatsidis G. Gene Therapy in Reconstructive and Regenerative Surgery. Springer International Publishing; 2018. doi:10.1007/978-3-319-78957-6
313. Sun J, Ju F, Jin J, et al. M2 macrophage membrane‐mediated biomimetic‐nanoparticle carrying COX‐siRNA targeted delivery for prevention of tendon adhesions by inhibiting inflammation. Small. 2023;19(33). doi:10.1002/smll.202300326
314. Zhang Y, Ju W, Zhang H, Mengyun L, Shen W, Chen X. Mechanisms and therapeutic prospects of mesenchymal stem cells-derived exosomes for tendinopathy. Stem Cell Res Ther. 2023;14(1):307. doi:10.1186/s13287-023-03431-3
315. Xu H, Zhu Y, Hsiao AWT, et al. Bioactive glass-elicited stem cell-derived extracellular vesicles regulate M2 macrophage polarization and angiogenesis to improve tendon regeneration and functional recovery. Biomaterials. 2023;294:121998. doi:10.1016/j.biomaterials.2023.121998
316. Yao Z, Li J, Xiong H. MicroRNA engineered umbilical cord stem cell-derived exosomes direct tendon regeneration by mTOR signaling. J Nanobiotechnol. 2021;19(1). doi:10.1186/s12951-021-00906-4
317. Shi Z, Wang Q, Jiang D. Extracellular vesicles from bone marrow-derived multipotent mesenchymal stromal cells regulate inflammation and enhance tendon healing. J Transl Med. 2019;17(1):211. doi:10.1186/s12967-019-1960-x
318. Song W, Guo Y, Liu W, et al. Circadian rhythm‐regulated ADSC‐derived sEVs and a triphasic microneedle delivery system to enhance tendon‐to‐bone healing. Adv Mater. 2024. doi:10.1002/adma.202408255
319. Wu G, Su Q, Li J, et al. NAMPT encapsulated by extracellular vesicles from young adipose-derived mesenchymal stem cells treated tendinopathy in a “One-Stone-Two-Birds” manner. J Nanobiotechnol. 2023;21(1):7. doi:10.1186/s12951-022-01763-5
320. Wang Y, He G, Guo Y, et al. Exosomes from tendon stem cells promote injury tendon healing through balancing synthesis and degradation of the tendon extracellular matrix. J Cell Mol Med. 2019;23(8):5475–5485. doi:10.1111/jcmm.14430
321. Song K. Exosomes from tendon derived stem cells promote tendon repair through miR-144-3p-regulated tenocyte proliferation and migration. 2022.
322. An Q, Zhou Z, Xu C, Xiao Q. Exosomes derived from mir-337-3p over-expressing tendon stem cells protect against apoptosis of tenocytes via targeting caspase3. BMC Musculoskelet Disord. 2024;25(1):561. doi:10.1186/s12891-024-07691-9
323. Graça AL, Domingues RMA, Calejo I, Gómez-Florit M, Gomes ME. Therapeutic effects of platelet-derived extracellular vesicles in a bioengineered tendon disease model. Int J Mol Sci. 2022;23(6):2948. doi:10.3390/ijms23062948
324. Chen D, Tang Q, Song W, He Y. Platelet-derived exosomes alleviate tendon stem/progenitor cell senescence and ferroptosis by regulating AMPK/Nrf2/GPX4 signaling and improve tendon-bone junction regeneration in rats. J Orthop Surg. 2024;19(1):382. doi:10.1186/s13018-024-04869-8
325. Lu J, Yang X, He C, et al. Rejuvenation of tendon stem/progenitor cells for functional tendon regeneration through platelet-derived exosomes loaded with recombinant Yap1. Acta Biomater. 2023;161:80–99. doi:10.1016/j.actbio.2023.02.018
326. Jin S, Wang Y, Wu X, et al. Young exosome bio‐nanoparticles restore aging‐impaired tendon stem/progenitor cell function and reparative capacity. Adv Mater. 2023;35(18):2211602. doi:10.1002/adma.202211602
327. Li M, Ma J, Gao Y, Yang L. Cell sheet technology: a promising strategy in regenerative medicine. Cytotherapy. 2019;21(1):3–16. doi:10.1016/j.jcyt.2018.10.013
328. Zhou S, Yang R, Zou Q, et al. Fabrication of tissue-engineered bionic urethra using cell sheet technology and labeling by ultrasmall superparamagnetic iron oxide for full-thickness urethral reconstruction. Theranostics. 2017;7(9):2509–2523. doi:10.7150/thno.18833
329. Komatsu I, Wang JHC, Iwasaki K, Shimizu T, Okano T. The effect of tendon stem/progenitor cell (TSC) sheet on the early tendon healing in a rat Achilles tendon injury model. Acta Biomater. 2016;42:136–146. doi:10.1016/j.actbio.2016.06.026
330. Liu Q, Yu Y, Reisdorf RL, et al. Engineered tendon-fibrocartilage-bone composite and bone marrow-derived mesenchymal stem cell sheet augmentation promotes rotator cuff healing in a non-weight-bearing canine model. Biomaterials. 2019;192:189–198. doi:10.1016/j.biomaterials.2018.10.037
331. Tang Y, Chen C, Liu F, et al. Structure and ingredient-based biomimetic scaffolds combining with autologous bone marrow-derived mesenchymal stem cell sheets for bone-tendon healing. Biomaterials. 2020;241:119837. doi:10.1016/j.biomaterials.2020.119837
332. Gonçalves AI, Rodrigues MT, Gomes ME. Tissue-engineered magnetic cell sheet patches for advanced strategies in tendon regeneration. Acta Biomater. 2017;63:110–122. doi:10.1016/j.actbio.2017.09.014
333. Vinhas A, Gonçalves AI, Rodrigues MT, Gomes ME. Human tendon-derived cell sheets created by magnetic force-based tissue engineering hold tenogenic and immunomodulatory potential. Acta Biomater. 2021;131:236–247. doi:10.1016/j.actbio.2021.06.036
334. Yao S, Liang Z, Lee YW, Yung PSH, Lui PPY. Bioactive decellularized tendon-derived stem cell sheet for promoting graft healing after anterior cruciate ligament reconstruction. Am J Sports Med. 2023;51(1):66–80. doi:10.1177/03635465221135770
335. Egami M, Haraguchi Y, Shimizu T, Yamato M, Okano T. Latest status of the clinical and industrial applications of cell sheet engineering and regenerative medicine. Arch Pharm Res. 2014;37(1):96–106. doi:10.1007/s12272-013-0299-8
336. Truby RL, Lewis JA. Printing soft matter in three dimensions. Nature. 2016;540(7633):371–378. doi:10.1038/nature21003
337. Ruiz-Alonso S, Lafuente-Merchan M, Ciriza J, Saenz-del-Burgo L, Pedraz JL. Tendon tissue engineering: cells, growth factors, scaffolds and production techniques. J Control Release. 2021;333:448–486. doi:10.1016/j.jconrel.2021.03.040
338. Alhaskawi A, Zhou H, Dong Y, et al. Advancements in
339. Silva M, Gomes S, Correia C, et al. Biocompatible 3D-printed tendon/ligament scaffolds based on polylactic acid/graphite nanoplatelet composites. Nanomaterials. 2023;13(18):2518. doi:10.3390/nano13182518
340. Mozdzen LC, Rodgers R, Banks JM, Bailey RC, Harley BAC. Increasing the strength and bioactivity of collagen scaffolds using customizable arrays of 3D-printed polymer fibers. Acta Biomater. 2016;33:25–33. doi:10.1016/j.actbio.2016.02.004
341. Zhang Y, Lei T, Tang C, et al. 3D printing of chemical-empowered tendon stem/progenitor cells for functional tissue repair. Biomaterials. 2021;271:120722. doi:10.1016/j.biomaterials.2021.120722
342. Zhao F, Cheng J, Zhang J, et al. Comparison of three different acidic solutions in tendon decellularized extracellular matrix bio-ink fabrication for 3D cell printing. Acta Biomater. 2021;2021:1.
343. Jiang X. 3D bioprinting of multilayered scaffolds with spatially differentiated ADMSCs for rotator cuff tendon-to-bone interface regeneration. Appl Mater Today. 2022;2022:1.
344. Zhang H, Pei Z, Wang C, Li M, Zhang H, Qu J. Electrohydrodynamic 3D printing scaffolds for repair of achilles tendon defect in rats. Tissue Eng Part A. 2021;27(19–20):1343–1354. doi:10.1089/ten.tea.2020.0290
345. Chae S, Yong U, Park W, et al. 3D cell-printing of gradient multi-tissue interfaces for rotator cuff regeneration. Bioact Mater. 2023;19:611–625. doi:10.1016/j.bioactmat.2022.05.004
346. Neubauer VJ, Hüter F, Wittmann J, et al. Flow simulation and gradient printing of fluorapatite- and cell-loaded recombinant spider silk hydrogels. Biomolecules. 2022;12(10):1413. doi:10.3390/biom12101413
347. Monteiro RF, Bakht SM, Gomez-Florit M, et al. Writing 3D in vitro models of human tendon within a biomimetic fibrillar support platform. ACS Appl Mater Interfaces. 2023;15(44):50598–50611. doi:10.1021/acsami.2c22371
348. Balestri W, Hickman GJ, Morris RH, Hunt JA, Reinwald Y. Triphasic 3D in vitro model of bone-tendon-muscle interfaces to study their regeneration. Cells. 2023;12(2):313. doi:10.3390/cells12020313
349. Wang M, Favi P, Cheng X, et al. Cold atmospheric plasma (CAP) surface nanomodified 3D printed polylactic acid (PLA) scaffolds for bone regeneration. Acta Biomater. 2016;46:256–265. doi:10.1016/j.actbio.2016.09.030
350. Li Z, Xie MB, Li Y, Ma Y, Li JS, Dai FY. Recent progress in tissue engineering and regenerative medicine. J Biomater Tissue Eng. 2016;6(10):755–766. doi:10.1166/jbt.2016.1510
351. Rinoldi C, Kijeńska‐Gawrońska E, Khademhosseini A, Tamayol A, Swieszkowski W. Fibrous systems as potential solutions for tendon and ligament repair, healing, and regeneration. Adv Healthc Mater. 2021;10(7):2001305. doi:10.1002/adhm.202001305
352. Moffat KL, Levine WN, Spalazzi JP, Doty SB, Levine WN, Lu HH. Novel nanofiber-based scaffold for rotator cuff repair and augmentation. Tissue Eng A. 2009;15(1):115–126. doi:10.1089/ten.tea.2008.0014
353. Yi B, Zhang H, Yu Z, Yuan H, Wang X, Zhang Y. Fabrication of high performance silk fibroin fibers via stable jet electrospinning for potential use in anisotropic tissue regeneration. J Mater Chem B. 2018;6(23):3934–3945. doi:10.1039/C8TB00535D
354. Wang L, Zhu T, Kang Y, et al. Crimped nanofiber scaffold mimicking tendon-to-bone interface for fatty-infiltrated massive rotator cuff repair. Bioact Mater. 2022;16:149–161. doi:10.1016/j.bioactmat.2022.01.031
355. Zhao J, Zhang D, Lan Q, et al. Tendon decellularized matrix modified fibrous scaffolds with porous and crimped microstructure for tendon regeneration. ACS Appl Bio Mater. 2024;7(7):4747–4759. doi:10.1021/acsabm.4c00565
356. Schynkel L, Meeremans M, Meyer AA, et al. Cell guiding multicomponent nanoyarn tendon scaffolds with tunable morphology and flexibility. ACS Appl Mater Interfaces. 2023;15(36):42241–42250. doi:10.1021/acsami.3c08241
357. Dong L, Li L, Song Y, et al. MSC-derived immunomodulatory extracellular matrix functionalized electrospun fibers for mitigating foreign-body reaction and tendon adhesion. Acta Biomater. 2021;133:280–296. doi:10.1016/j.actbio.2021.04.035
358. Abdulmalik S, Gallo J, Nip J, et al. Nanofiber matrix formulations for the delivery of exendin-4 for tendon regeneration: in vitro and in vivo assessment. Bioact Mater. 2023;25:42–60. doi:10.1016/j.bioactmat.2023.01.013
359. Liu J. Unidirectional gene delivery electrospun fibrous membrane via charge repulsion for tendon repair. Bioact Mater. 2024;2024:1.
360. Khorshidi S, Solouk A, Mirzadeh H, et al. A review of key challenges of electrospun scaffolds for tissue-engineering applications: challenges regarding electrospun scaffolds: a review. J Tissue Eng Regen Med. 2016;10(9):715–738. doi:10.1002/term.1978
361. Bishop ES, Mostafa S, Pakvasa M, et al. 3-D bioprinting technologies in tissue engineering and regenerative medicine: current and future trends. Genes Dis. 2017;4(4):185–195. doi:10.1016/j.gendis.2017.10.002
362. Cho Y, Baek JW, Sagong M, Ahn S, Nam JS, Kim I. Electrospinning and nanofiber technology: fundamentals, innovations, and applications. Adv Mater. 2025;37(28):2500162. doi:10.1002/adma.202500162
363. Zhang G, Zhou X, Hu S, Jin Y, Qiu Z. Large animal models for the study of tendinopathy. Front Cell Dev Biol. 2022;10:1031638. doi:10.3389/fcell.2022.1031638
364. Moraska A, Deak T, Spencer RL, Roth D, Fleshner M. Treadmill running produces both positive and negative physiological adaptations in Sprague-Dawley rats. Am J Physiol-Regul, Integr Comp Physiol. 2000;279(4):R1321–R1329. doi:10.1152/ajpregu.2000.279.4.R1321
365. Mukohara S, Mifune Y, Inui A, et al. In vitro and in vivo tenocyte-protective effectiveness of dehydroepiandrosterone against high glucose-induced oxidative stress. BMC Musculoskelet Disord. 2021;22(1):519. doi:10.1186/s12891-021-04398-z
366. Wang S, Xiao Y, Tian J, et al. Targeted macrophage CRISPR‐Cas13 mRNA editing in immunotherapy for tendon injury. Adv Mater. 2024;2024:1.
367. Chen M, Zou F, Wang P, et al. Dual‐barb microneedle with JAK/STAT inhibitor‐loaded nanovesicles encapsulation for tendinopathy. Adv Healthc Mater. 2024;13(28):2401512. doi:10.1002/adhm.202401512
368. Shen H, Yoneda S, Abu Y, Guilak F, Gelberman RH. Stem cell‐derived extracellular vesicles attenuate the early inflammatory response after tendon injury and repair. J Orthop Res. 2020;38(1):117–127. doi:10.1002/jor.24406
369. Chen M, Li Y, Xiao L. AQP1 modulates tendon stem/progenitor cells senescence during tendon aging. Cell Death Dis. 2020;11(3). doi:10.1038/s41419-020-2386-3
370. Chen L, Wang GD, Liu JP, et al. miR-135a modulates tendon stem/progenitor cell senescence via suppressing ROCK1. Bone. 2015;71:210–216. doi:10.1016/j.bone.2014.11.001
371. Cai Z, Xin Z, Wang H, Wang C, Liu X. Extracellular vesicle‐contained thrombospondin 1 retards age‐related degenerative tendinopathy by rejuvenating tendon stem/progenitor cell senescence. Small. 2024;2400598. doi:10.1002/smll.202400598
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.