Back to Journals » Drug Design, Development and Therapy » Volume 15
Targeting Protein Neddylation to Inactivate Cullin-RING Ligases by Gossypol: A Lucky Hit or a New Start?
Received 12 October 2020
Accepted for publication 16 December 2020
Published 6 January 2021 Volume 2021:15 Pages 1—8
DOI https://doi.org/10.2147/DDDT.S286373
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Qing Yu,1– 3 Yi Sun3
1Department of Head and Neck Surgery, The Cancer Hospital of the University of Chinese Academy of Sciences (Zhejiang Cancer Hospital), Institute of Basic Medicine and Cancer (IBMC), Chinese Academy of Science, Hangzhou, Zhejiang, People’s Republic of China; 2Key Laboratory of Head & Neck Cancer Translational Research of Zhejiang Province, Hangzhou, Zhejiang, People’s Republic of China; 3Cancer Institute of the Second Affiliated Hospital and Institute of Translational Medicine, Zhejiang University School of Medicine, Hangzhou, Zhejiang, People’s Republic of China
Correspondence: Yi Sun
Cancer Institute of the Second Affiliated Hospital and Institute of Translational Medicine, Zhejiang University School of Medicine, 268 Kaixuan Road, Hagnzhou, Zheijiang, People’s Republic of China
Tel +86 571 86971812
Fax +86 571 88981576
Email [email protected]
Abstract: Cullin-RING E3 ligases (CRLs) are the largest family of E3 ubiquitin ligases, responsible for about 20% of the protein degradation by the ubiquitin-proteasome system (UPS). Given their vital roles in multiple cellular processes, and over-activation in many human cancers, CRLs are validated as promising targets for anti-cancer therapies. Activation of CRLs requires cullin neddylation, a process catalysed by three neddylation enzymes. Recently, our group established an AlphaScreen-based in vitro cullin neddylation assay and employed it for high-throughput screening to search for small-molecule inhibitors targeting cullin neddylation. During our pilot screen, gossypol, a natural product extracted from cottonseeds, was identified as one of the most potent neddylation inhibitors of cullin-1 and cullin-5. We further demonstrated that gossypol blocks cullin neddylation by binding to cullin-1/-5 to inactivate CRL1/5 ligase activity, leading to accumulation of MCL-1 and NOXA, the substrates of CRL1 and CRL5, respectively. The combination of gossypol and an MCL-1 inhibitor synergistically enhanced the anti-proliferative effect in multiple human cancer cell lines. Our study unveiled a rational combination of two previously known inhibitors of the Bcl-2 family for enhanced anti-cancer efficacy and identified a novel activity of gossypol as an inhibitor of CRL1 and CRL5 E3s, thus providing a new possibility in the development of novel CRL inhibitors for anti-cancer therapy.
Keywords: anti-cancer drug, cullin-RING E3 ligases, natural product, high-throughput screen, neddylation, small-molecule inhibitors
Introduction
In mammalian cells, the ubiquitin-proteasome system (UPS) is responsible for more than 80% of the degradation of cellular proteins, which plays a major role in maintaining cellular homeostasis.1 Proteins doomed for degradation by UPS will first be subjected to ubiquitylation, a process to label the target protein with poly-ubiquitin chains before being recognized by the proteasome.2 Ubiquitylation is a three-step enzymatic reaction, sequentially catalyzed by three enzymes, an E1 activating enzyme (NAE), an E2 conjugating enzyme, and an E3 ligase.3–5 Structural and biological studies have revealed the core function of E3 ligases as not only scaffolds to hold the multienzyme complex but also recruiters for selecting specific substrates.6
In mammalian cells, the cullin-RING E3 ligases (CRLs) are the largest family of E3 ubiquitin ligases, consisting of four subunits: scaffold cullins (with 8 family members: CUL-1, −2, −3, −4A, −4B, −5, −7, −9); adaptor proteins (with many members), substrate receptors (with many members), and the RING proteins (with two family members of RBX1 and RBX2/SAG).6 CRLs are responsible for about 20% of the UPS-mediated protein degradation,7 thus playing vital roles in cells as the substrates of CRLs control multiple major cellular processes such as cell cycle progression, apoptosis, DNA replication and repair, and signal transduction among many others.8
Importantly, activation of CRLs requires cullin neddylation, a process of attaching the ubiquitin-like molecule, NEDD8 into the lysine residue of a given substrate.9 Like ubiquitylation, neddylation is also catalysed by three enzymes: E1 NEDD8 activating enzyme (NAE), E2 NEDD8 conjugate enzymes (with two family members: UBE2M and UBE2F), and E3 NEDD8 ligase (with over 10 members).10 In many types of human cancers, a variety of CRL components as well as neddylation enzymes were overexpressed, leading to overactivation of CRLs.9 Thus, targeting neddylation enzymes to inactivate CRLs is a sound strategy. MLN4924, a small molecular inhibitor of NAE, was discovered in 2009 and is currently at several Phase II clinical trials for anti-cancer application.7 However, MLN4924 has intrinsic cytotoxicity due to its inhibition of neddylation E1 to block the entire neddylation process and subsequent inactivation of the entire family of CRLs. Thus, the discovery of more potent small molecules that selectively target individual CRLs with an anticipated low cytotoxicity is in high demand.10
Recently our group has established an AlphaScreen-based in vitro cullin neddylation assay.11 Using this robust and powerful assay, we performed high-throughput screening (HTS) for small-molecule inhibitors that inhibits cullin-5 neddylation. In a pilot experiment where diversified libraries of ~17,000 compounds were screened, gossypol, a natural product extracted from cottonseeds, was identified as one of the most potent neddylation inhibitors of cullin-5, as well as cullin-1. Our further investigations not only uncovered an unreported target for gossypol but also provided a rational strategy of drug combination to enhance the anti-cancer efficacy of this century-old drug. Here we review the background behind our study, discuss the biological significance of our findings, and then propose some perspectives for future investigation to clarify the mode of gossypol action as a CRL inhibitor and its potential application for other human diseases.
Cullin-RING Ligase: A Good Anti-Cancer Target, but a Bad Screen Target
The history of UPS-targeting drugs began two decades ago when Goldberg and his colleagues curiously generated proteasome inhibitors for treating muscle wasting.12 Their efforts eventually led to the success of Bortezomib (Velcade), the first-in-class proteasome inhibitor approved by the food and drug administration (FDA) for treating multiple myeloma (MM) in the U.S.13 Now bortezomib and its successors (eg, carfilzomib and ixazomib) are included in the therapeutic guidelines for treatment of multiple myeloma14 with a total annual sale >$300 million in 2018 in the U.S.15
Although the success of bortezomib validates UPS as a promising therapeutic target, it does not mask the fact that pan-proteasome inhibitors carry an intrinsically high level of cytotoxicity due to general inhibition of the entire protein degradation machinery.16 Thus, the upstream E3 ligases with a total of ~600 in the human genome have been considered as a more ideal target in the UPS, given their key roles in determining the specificity of substrate proteins for ubiquitylation.10,17 For example, Nurix is a growing company focusing on E3-targeting therapeutics with two major pipelines on E3 enhancers and E3 inhibitors. In July of 2020, Nurix successfully launched their $200-million initial public offering (IPO) on Nasdaq, showing the market confidence of bringing novel E3 ligases-targeted drugs into the clinics.18
CRLs are the largest family of E3 ligases and have been validated as attractive anti-cancer targets.9,19 The activity of CRLs depends on neddylation, a three-step enzymatic cascade resembling ubiquitylation, with its own NEDD8 E1, E2s, and E3s.9 In 2009, MLN4924 became the first reported neddylation inhibitor, specifically targeting NEDD8 E1.7 MLN4924 showed potent inhibitory effect against all cullin neddylation both in vitro and in vivo, and showed significant anti-tumour efficacy in multiple cancer cell lines as well as in mouse models.7 To date, MLN4924 has entered 38 related phase I/II clinical trials, of which 27 are still ongoing (https://clinicaltrials.gov). The success of MLN4924 further validated that targeting cullin neddylation is a feasible strategy. However, the pan-inhibition of the entire neddylation reaction, involving inactivation of both cullins and non-cullin substrates, has intrinsically associated cytotoxicity due to its non-specific nature.10 Direct inhibition of individual CRLs is expected to have higher selectivity with fewer side effects.
The discovery of such small molecule inhibitors, however, has been met by significant challenge. Up until now, no small molecule inhibitor has been reported to specifically target any given CRL. Traditional small molecule inhibitors, such as kinase inhibitors, are often designed to bind to a specific pocket in the target protein20 but for CRLs, such a druggable pocket is often lacking.21,22 The interactions among the components of CRLs as well as between enzymes and substrates are transient in nature and largely based on the protein–protein interactions (PPIs), often with flat and large contact surfaces.21,22 Furthermore, the reaction of ubiquitylation and neddylation are both dynamic cascades. Numerous conformational changes synergistically occur within the sophisticated CRL complex.23 Any change in conformation or binding mode would cause alteration in enzymatic activity, not to mention difficulty in optimization of assay conditions containing multiple proteins, including ubiquitin, E1, E2, E3 (with multiple components) and the substrate in the right amount and proper ratio. Thus, it is extremely hard to establish traditional high-throughput screens for direct CRL inhibitors. The lack of a binding pocket among these PPI interactions also makes virtual screening a difficult task.
AlphaScreen-Based in vitro Neddylation Assay: The Advantages
It appears that it is more practical to screen for indirect inhibitors of CRLs via targeting neddylation with blockage of cullin neddylation as a readout. To this end, we explored several in vitro HTS techniques, including FRET24 and ELISA,25 and eventually optimized the AlphaScreen® (PerkinElmer Inc.) assay to conduct HTS for small molecules that inhibit cullin-5 neddylation as the readout. This fully optimized AlphaScreen-based HTS assay for neddylation inhibitors has two major advantages:
- It is based on a dynamic enzymatic reaction. Instead of simply introducing one pair of interacting proteins, as shown in most reported AlphaScreen-based assays, we assembled a five-element reaction mixture to set up a functional neddylation machinery (ie, a NEDD8, a NEDD8 E1, a NEDD8 E2 (UBE2F), and co-expressed SAG (E3) and cullin-5 (substrate)). Once ATP is added, the reaction starts to produce neddylated cullin-5 and any small molecules that stop such a reaction will be captured as a positive hit.
- It is a modular reaction mixture with replaceable components. By simply replacing the NEDD8 E2 (UBE2F by UBE2M) and E3/Substrate (SAG/CUL5 by RBX1/CUL1), a new assay is readily available for identifying inhibitors targeting different cullin neddylation. And this feature has helped us in setting up a series of orthogonal experiments to classify positive hits with different targets in a high-throughput format.
Using this model, we performed a pilot screen with a diversified library containing 1.7K compounds. Gossypol was identified and later validated as a positive hit. The follow-up confirmation experiments showed that gossypol indeed binds CUL5, as determined by cellular thermal shift assays (CETSA) and mutagenesis assays, and inhibits CUL5 neddylation in both pre-charged in vitro enzymatic assays and in vivo cell culture settings in multiple lines of cancer cells.11
To the best of our knowledge, we are the first to report the inhibitory effect of gossypol against cullin neddylation. Thus, gossypol is the first class of natural product that inhibits cullin neddylation by targeting the neddylation E2–E3 interaction or neddylation E3, leading to inactivation of CRL5.
Gossypol: A Potential Anti-Cancer Drug Candidate with a Variety of Mechanisms
Gossypol is found in the seeds and root bark of certain cotton plants (Gossypium).26 The natural gossypol is a racemic mixture consists of two enantiomers, (+)-gossypol and (-)-gossypol (also known as AT-101).27 Gossypol was initially found to have male antifertility effects in the1980s and has been used as a male oral contraceptive in China since then.26 In addition, gossypol was also found to have anti-viral, anti-parasitic, and anti-inflammatory activity.28 More recently, a number of studies have shown that gossypol has potent anti-tumour effects against multiple cancer cell lines.28 However, the exact mode of action of gossypol remains unclear.
Back in 2003, Kitada et al identified gossypol as an antagonist of the B-cell lymphocyte/leukemia-2 (Bcl-2) family of proteins through a screening of a small library of natural polyphenols.29 The follow-up studies demonstrated that gossypol structurally behaves as a BH3-mimetic which can bind to Bcl-2 and Bcl-xl30,31 and later AT-101 [(-)-gossypol] became one of the first orally available Bcl-2 inhibitor that has entered the clinical trials.32 Up until now, AT-101 has been investigated in more than 20 phase I/II and 1 Phase III clinical trial(s) as a single agent or in combination with other agents against multiple types of cancer (https://clinicaltrials.gov). Two phase II trials are still ongoing in China (CTR20150540, CTR20150541).
Although AT-101 showed potent response in a few trials,33 the high level of cytotoxicity and low or moderate efficacy in many trials limits its further application in the clinic.34 While few studies have shown that gossypol was orally applicable and well tolerated when applied in combination with other therapies,34–37 most studies of gossypol as a single-agent reported considerable side effect, including hematologic toxicities and gastrointestinal motility.38–43 Of note, some of the results in clinical trials showed that Bcl-2 protein levels were not significantly changed in peripheral blood mononuclear cells (PBMCs) of AT-101 treated patients, indicating AT-101 may not always be “on-target” in vivo.34
On the other hand, the Bcl-2 family proteins may not be the major targets of AT-101 in some patients. Indeed, gossypol has been reported to affect many other proteins or pathways, such as inhibiting mitosis, cyclin D1, and Rb,44 decreasing ATP synthesis related with aldehyde dehydrogenase,45 suppressing the mTOR/p70S6K1 signalling pathway,46 downregulating phospholipase A2 to induce endoplasmic reticulum stress,47 and interacting with intracellular copper to generate reactive oxygen species (ROS).48
Furthermore, the polyphenol structure of gossypol might contribute to its multi-target properties in cells, and thus has received a great deal of attention from medicinal chemists.49 Several groups have synthesized a series of polyphenol derivatives of gossypol with similar or improved anti-cancer efficacy,49,50 while the others have attempted to improve its solubility with a set of modifications.51,52 None of these approaches, however, has significantly improved the efficacy of gossypol as an anticancer agent, nor for understanding its diversified mechanisms of action.
Rational Design for Drug Combination
Paradoxically, while gossypol was found to inhibit CRL5, as well as CRL1 in multiple cancer cell lines, unlike MLN4924, it failed to cause accumulation of some of the most well-known cullin substrates (eg, p21, p27, HIF1, CDT1, and NRF2).19 Rather, in our assay conditions, gossypol selectively caused the accumulation of NOXA and MCL-1, two Bcl-2 family protein known to have close interactions.53 Indeed, other groups have also shown gossypol could increase NOXA and MCL-1 protein levels in different cancer cell lines.54,55 However, no mechanistic evidence was provided to explain why this pair of mutually restrictive proteins are both increased at the protein levels in response to gossypol treatment. We have previously reported that NOXA is a substrate of CUL5 and is strictly controlled via CUL5-mediated K11-linked polyubiquitylation.56 And MCL-1 has also been reported to be tightly regulated by ubiquitylation via at least six E3 ligases,57 including SCFFBW7 and SCFβ-TrCP, two CRL1 ligases.58,59 Our findings therefore provided a mechanistic explanation in which gossypol inactivates both CUL5-SAG E3 ligases (CRL5) and CUL1-RBX1 E3 ligases (CRL1) by blocking CUL5/CUL1 neddylation, leading to suppressed degradation of NOXA and MCL-1 (Figure 1A–C). As a pro-survival protein, MCL-1 is known to be associated with resistance to Bcl-2 inhibitors.60–63 We therefore rationally combined gossypol with an MCL-1 inhibitor S6384564 and found a synergetic effect against the growth and survival of multiple lines of human cancer cells (Figure 1D).
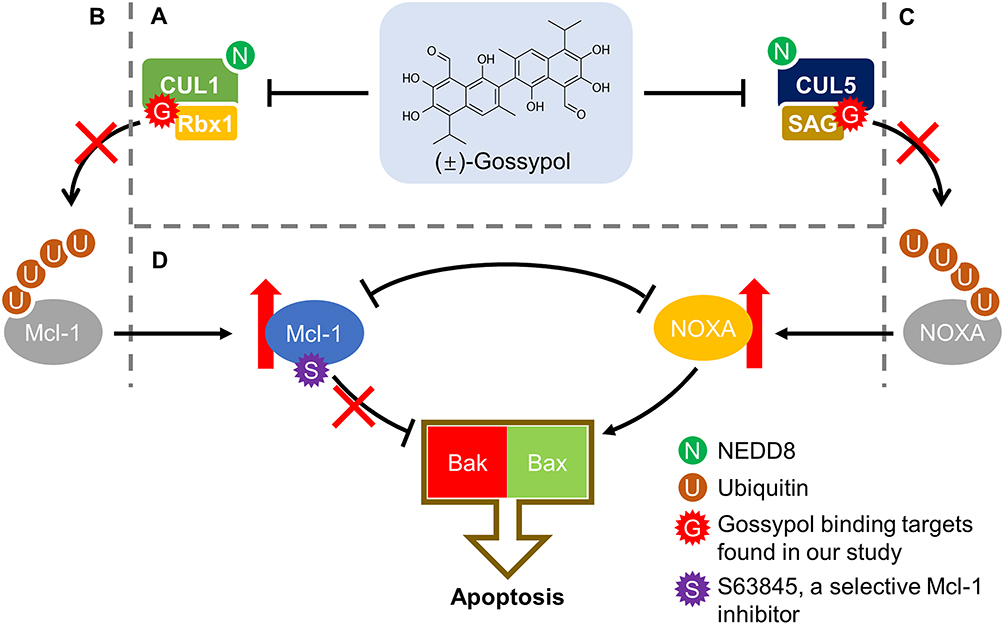 |
Figure 1 Gossypol targets both cullin neddylation and Bcl-2 family proteins. Our study uncovered the inhibitory effect of gossypol against the neddylation of CUL1/CUL5 (A), blocking the degradation of MCL-1 (B) and NOXA (C), two reported substrates of CUL1 and CUL5, respectively. Consequently, MCL-1 and NOXA proteins were accumulated, but the opposite effects of pro-survival (by MCL-1) or pro-apoptosis (by NOXA) compromise the anti-cancer activity of gossypol. The rational combination of gossypol and MCL-1 inhibitor significantly sensitized multiple lines of cancer cells to gossypol, providing a mechanism-based strategy of drug combination for enhanced efficacy (D). |
Future Perspectives
The low efficacy and unmanageable adverse events of gossypol in multiple clinical trials limited its use as a single anti-cancer agent. Fortunately, our findings may help further exploit its therapeutic potentials. In general, the fact that CRLs is a new physical target of gossypol provides a novel mechanism of gossypol action as an anticancer agent. On the other hand, gossypol-induced accumulation of anti-apoptotic MCL-1 (in addition to pro-apoptotic NOXA) may confer enhanced survival of cancer cells as a side effect. Below we proposed potential future studies, based upon our finding that gossypol is a neddylation inhibitor.
- Why and how gossypol selectively affects NOXA and MCL1, but not the other substrates of CUL5 and CUL1 or substrates of other cullins. It will be informative to compare the binding affinity between gossypol and different Cullin-RING E3 ligases, but a definitive answer will rely on the co-crystallization of gossypol with either CUL-5 or CUL-1, which, however, is technically difficult due to poor solubility of gossypol.
- Gossypol has been shown to bind to at least three anti-apoptotic proteins, MCL-1, Bcl-2 and Bcl-xL (Figure 1D).31 What is the binding affinity between gossypol-CUL1/5 vs gossypol-Bcl-2 family members? How much gossypol-CUL1/5 binding and inhibition of CRL1/5 contributes to the biological activity of gossypol as an anti-cancer agent? It should be testable both in vitro cell culture and in vivo mouse xenograft models, using cancer cells with depletion of both CUL1 and CUL5.
- Given the unique polyphenol structure, gossypol has its limitations for further chemical modification.49 However, it might still be worth the effort to structurally dissect manifold bioactivities of gossypol, which is essential to develop more selective compounds against relevant targets.
- Since gossypol has shown its bioactivities in multiple diseases, gossypol as a neddylation inhibitor may have new therapeutic application beyond cancer. For example, gossypol was reported to possess antiviral properties against enveloped viruses, including HIV-1 and HSV-2,65 but the exact mechanism is unclear. Given that CRL5 has been demonstrated as a key E3 ligase hijacked by virus during infection and thus validated as a promising target for anti-virus therapy,66 it would be intriguing to investigate whether inactivation of CRL5 is indeed the mechanism of gossypol action as an anti-viral agent.
In conclusion, for the first time, we identified gossypol as a neddylation inhibitor through an AlphaScreen-based HTS for small molecule inhibitors of CUL5 neddylation. We demonstrated that gossypol inhibits cullin neddylation by targeting the neddylation E2-E3 using both biochemical and cell biological assays. Our study provides a sound strategy to combine gossypol with an MCL-1 inhibitor for enhanced anti-cancer efficacy and elucidate a potential mechanism by which gossypol inhibits viral replication via CRL5 inactivation.
Funding
This work was supported by the National Key R&D Program of China (2016YFA0501800 to Y.S.), and the National Natural Science Foundation of China (81572718 and 81630076 to Y.S.).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Collins GA, Goldberg AL. The logic of the 26S proteasome. Cell. 2017;169(5):792–806. doi:10.1016/j.cell.2017.04.023
2. Hershko A, Ciechanover A. The ubiquitin system. Annu Rev Biochem. 1998;67:425–479. doi:10.1146/annurev.biochem.67.1.425
3. Ye Y, Rape M. Building ubiquitin chains: E2 enzymes at work. Nat Rev Mol Cell Biol. 2009;10:755–764. doi:10.1038/nrm2780
4. Schulman BA, Wade Harper J, Harper JW. Ubiquitin-like protein activation by E1 enzymes: the apex for downstream signalling pathways. Nat Rev Mol Cell Biol. 2009;10:319–331. doi:10.1038/nrm2673
5. Deshaies RJ, Joazeiro CAP. RING domain E3 ubiquitin ligases. Annu Rev Biochem. 2009;78:399–434. doi:10.1146/annurev.biochem.78.101807.093809
6. Zheng N, Shabek N. Ubiquitin ligases: structure, function, and regulation. Annu Rev Biochem. 2017;86:129–157. doi:10.1146/annurev-biochem-060815-014922
7. Soucy TA, Smith PG, Milhollen MA, et al. An inhibitor of NEDD8-activating enzyme as a new approach to treat cancer. Nature. 2009;458:732–736. doi:10.1038/nature07884
8. Introduction. Advances in experimental medicine and biology; 2020.
9. Zhou L, Zhang W, Sun Y, Jia L. Protein neddylation and its alterations in human cancers for targeted therapy. Cell Signal. 2018;44:92–102. doi:10.1016/j.cellsig.2018.01.009
10. Yu Q, Jiang Y, Sun Y. Anticancer drug discovery by targeting cullin neddylation. Acta Pharm Sin B. 2020;10(5):746–765. doi:10.1016/j.apsb.2019.09.005
11. Yu Q, Hu Z, Shen Y, et al. Gossypol inhibits cullin neddylation by targeting SAG-CUL5 and RBX1-CUL1 complexes. Neoplasia. 2020;22(4):179–191. doi:10.1016/j.neo.2020.02.003
12. Goldberg AL. Development of proteasome inhibitors as research tools and cancer drugs. J Cell Biol. 2012;199(4):583–588. doi:10.1083/jcb.201210077
13. Raab MS, Podar K, Breitkreutz I, Richardson PG, Anderson KC. Multiple myeloma. Lancet. 2009;374(9686):324–339. doi:10.1016/S0140-6736(09)60221-X
14. Mikhael J, Ismaila N, Cheung MC, et al. Treatment of multiple myeloma: ASCO and CCO joint clinical practice guideline. J Clin Oncol. 2019;37(14):1228–1263. doi:10.1200/JCO.18.02096
15. Park JE, Miller Z, Jun Y, Lee W, Kim KB. Next-generation proteasome inhibitors for cancer therapy. Transl Res. 2018;198:1–16. doi:10.1016/j.trsl.2018.03.002
16. Manasanch EE, Orlowski RZ. Proteasome inhibitors in cancer therapy. Nat Rev Clin Oncol. 2017;14(7):417–433. doi:10.1038/nrclinonc.2016.206
17. Bulatov E, Valiullina A, Sayarova R, Rizvanov A. Promising new therapeutic targets for regulation of inflammation and immunity: RING-type E3 ubiquitin ligases. Immunol Lett. 2018;202:44–51. doi:10.1016/j.imlet.2018.08.001
18. Crawford DR, Amstad PA, Foo DD, Cerutti PA. Constitutive and phorbol-myristate-acetate regulated antioxidant defense of mouse epidermal JB6 cells. Mol Carcinog. 1989;2(3):136–143.
19. Zhao Y, Morgan MA, Sun Y. Targeting neddylation pathways to inactivate Cullin-RING ligases for anti-cancer therapy. Antioxid Redox Signal. 2014;21(17):2383–2400. doi:10.1089/ars.2013.5795
20. Ferguson FM, Gray NS. Kinase inhibitors: the road ahead. Nat Rev Drug Discov. 2018;17:353–376.
21. Sheng C, Dong G, Miao Z, Zhang W, Wang W. State-of-the-art strategies for targeting protein-protein interactions by small-molecule inhibitors. Chem Soc Rev. 2015;44:8238–8259. doi:10.1039/C5CS00252D
22. Blundell TL, Burke DF, Chirgadze D, et al. Protein-protein interactions in receptor activation and intracellular signalling. Biol Chem. 2000;381:955–959. doi:10.1515/BC.2000.117
23. Baek K, Krist DT, Prabu JR, et al. NEDD8 nucleates a multivalent cullin-RING-UBE2D ubiquitin ligation assembly. Nature. 2020;578(7795):461–466. doi:10.1038/s41586-020-2000-y
24. Wu K, Chong RA, Yu Q, et al. Suramin inhibits cullin-RING E3 ubiquitin ligases. Proc Natl Acad Sci. 2016;113(14):E2011–E2018. doi:10.1073/pnas.1601089113
25. Ceccarelli DF, Tang X, Pelletier B, et al. An allosteric inhibitor of the human Cdc34 ubiquitin-conjugating enzyme. Cell. 2011;145(7):1075–1087. doi:10.1016/j.cell.2011.05.039
26. Qian SZ, Wang ZG. Gossypol: a potential antifertility agent for males. Annu Rev Pharmacol Toxicol. 1984;24:329–360. doi:10.1146/annurev.pa.24.040184.001553
27. Benvenuto M, Mattera R, Sticca JI, et al. Effect of the BH3 mimetic polyphenol (-)-Gossypol (AT-101) on the in vitro and in vivo growth of malignant mesothelioma. Front Pharmacol. 2018;9:1269. doi:10.3389/fphar.2018.01269
28. Keshmiri-Neghab H, Goliaei B. Therapeutic potential of gossypol: an overview. Pharm Biol. 2014;52(1):124–128. doi:10.3109/13880209.2013.832776
29. Kitada S, Leone M, Sareth S, Zhai D, Reed JC, Pellecchia M. Discovery, characterization, and structure-activity relationships studies of proapoptotic polyphenols targeting B-cell lymphocyte/leukemia-2 proteins. J Med Chem. 2003;46(20):4259–4264. doi:10.1021/jm030190z
30. Vogler M. Targeting BCL2-proteins for the treatment of solid tumours. Adv Med. 2014;2014:943648. doi:10.1155/2014/943648
31. Wang G, Nikolovska-Coleska Z, Yang C-Y, et al. Structure-based design of potent small-molecule inhibitors of anti-apoptotic Bcl-2 proteins. J Med Chem. 2006;49(21):6139–6142. doi:10.1021/jm060460o
32. Han Z, Liang J, Li Y, He J. Drugs and clinical approaches targeting the antiapoptotic protein: a review. Biomed Res Int. 2019;2019:1212369. doi:10.1155/2019/1212369
33. Mehner M, Kubelt C, Adamski V, Schmitt C, Synowitz M, Held-Feindt J. Combined treatment of AT101 and demethoxycurcumin yields an enhanced anti-proliferative effect in human primary glioblastoma cells. J Cancer Res Clin Oncol. 2020;146(1):117–126. doi:10.1007/s00432-019-03107-7
34. Stein MN, Goodin S, Gounder M, et al. A Phase I study of AT-101, a BH3 mimetic, in combination with paclitaxel and carboplatin in solid tumors. Invest New Drugs. 2020;38(3):855–865. doi:10.1007/s10637-019-00807-2
35. Schelman WR, Mohammed TA, Traynor AM, et al. A phase I study of AT-101 with cisplatin and etoposide in patients with advanced solid tumors with an expanded cohort in extensive-stage small cell lung cancer. Invest New Drugs. 2014;32(2):295–302. doi:10.1007/s10637-013-9999-7
36. Ready N, Karaseva NA, Orlov SV, et al. Double-blind, placebo-controlled, randomized Phase 2 study of the proapoptotic agent AT-101 plus docetaxel, in second-line non-small cell lung cancer. J Thorac Oncol. 2011;6(4):781–785. doi:10.1097/JTO.0b013e31820a0ea6
37. Van Poznak C, Seidman AD, Reidenberg MM, et al. Oral gossypol in the treatment of patients with refractory metastatic breast cancer: a phase I/II clinical trial. Breast Cancer Res Treat. 2001;66:239–248. doi:10.1023/A:1010686204736
38. Xie H, Yin J, Shah MH, et al. A phase II study of the orally administered negative enantiomer of gossypol (AT-101), a BH3 mimetic, in patients with advanced adrenal cortical carcinoma. Invest New Drugs. 2019;37(4):755–762. doi:10.1007/s10637-019-00797-1
39. Stein MN, Hussain M, Stadler WM, et al. A Phase II study of AT-101 to overcome Bcl-2-mediated resistance to androgen deprivation therapy in patients with newly diagnosed castration-sensitive metastatic prostate cancer. Clin Genitourin Cancer. 2016;14:22–27. doi:10.1016/j.clgc.2015.09.010
40. Sonpavde G, Matveev V, Burke JM, et al. Randomized phase II trial of docetaxel plus prednisone in combination with placebo or AT-101, an oral small molecule Bcl-2 family antagonist, as first-line therapy for metastatic castration-resistant prostate cancer. Ann Oncol. 2012;23:1803–1808. doi:10.1093/annonc/mdr555
41. Baggstrom MQ, Qi Y, Koczywas M, et al. A phase II study of AT-101 (Gossypol) in chemotherapy-sensitive recurrent extensive-stage small cell lung cancer. J Thorac Oncol. 2011;6:1757–1760. doi:10.1097/JTO.0b013e31822e2941
42. Suk Heist R, Fain J, Chinnasami B, et al. Phase I/II study of AT-101 with topotecan in relapsed and refractory small cell lung cancer. J Thorac Oncol. 2010;5:1637–1643. doi:10.1097/JTO.0b013e3181e8f4dc
43. Liu G, Kelly WK, Wilding G, Leopold L, Brill K, Somer B. An open-label, multicenter, phase I/II study of single-agent AT-101 in men with castrate-resistant prostate cancer. Clin Cancer Res. 2009;15(9):3172–3176. doi:10.1158/1078-0432.CCR-08-2985
44. Ligueros M, Jeoung D, Tang B, Hochhauser D, Reidenberg MM, Sonenberg M. Gossypol inhibition of mitosis, cyclin D1 and Rb protein in human mammary cancer cells and cyclin-D1 transfected human fibrosarcoma cells. Br J Cancer. 1997;76(1):21–28. doi:10.1038/bjc.1997.330
45. Rekha GK, Sladek NE. Inhibition of human class 3 aldehyde dehydrogenase, and sensitization of tumor cells that express significant amounts of this enzyme to oxazaphosphorines, by the naturally occurring compound gossypol. Adv Exp Med Biol. 1997;414:133–146.
46. Yang D, Qu J, Qu X, et al. Gossypol sensitizes the antitumor activity of 5-FU through down-regulation of thymidylate synthase in human colon carcinoma cells. Cancer Chemother Pharmacol. 2015;76(3):575–586. doi:10.1007/s00280-015-2749-0
47. Soderquist RS, Danilov AV, Eastman A. Gossypol increases expression of the pro-apoptotic BH3-only protein NOXA through a novel mechanism involving phospholipase A2, cytoplasmic calcium, and endoplasmic reticulum stress. J Biol Chem. 2014;289(23):16190–16199. doi:10.1074/jbc.M114.562900
48. Zubair H, Azim S, Khan HY, et al. Mobilization of intracellular copper by gossypol and apogossypolone leads to reactive oxygen species-mediated cell death: putative anticancer mechanism. Int J Mol Sci. 2016;17(5):973. doi:10.3390/ijms17060973
49. Lu Y, Li J, Dong C-E, Huang J, Zhou H-B, Wang W. Recent advances in gossypol derivatives and analogs: a chemistry and biology view. Future Med Chem. 2017;9:1243–1275. doi:10.4155/fmc-2017-0046
50. Tang G, Nikolovska-Coleska Z, Qiu S, Yang C-Y, Guo J, Wang S. Acylpyrogallols as inhibitors of antiapoptotic Bcl-2 proteins. J Med Chem. 2008;51(4):717–720. doi:10.1021/jm701358v
51. Yan F, Cao XX, Jiang HX, et al. A novel water-soluble gossypol derivative increases chemotherapeutic sensitivity and promotes growth inhibition in colon cancer. J Med Chem. 2010;53:5502–5510. doi:10.1021/jm1001698
52. Ionov M, Gordiyenko N, Olchowik E, et al. The immobilization of gossypol derivative on N-polyvinylpyrrolidone increases its water solubility and modifies membrane-active properties. J Med Chem. 2009;52:4119–4125. doi:10.1021/jm9002507
53. Guikema JE, Amiot M, Eldering E. Exploiting the pro-apoptotic function of NOXA as a therapeutic modality in cancer. Expert Opin Ther Targets. 2017;21:767–779. doi:10.1080/14728222.2017.1349754
54. Albershardt TC, Salerni BL, Soderquist RS, et al. Multiple BH3 mimetics antagonize antiapoptotic MCL1 protein by inducing the endoplasmic reticulum stress response and up-regulating BH3-only protein NOXA. J Biol Chem. 2011;286:24882–24895. doi:10.1074/jbc.M111.255828
55. Meng Y, Tang W, Dai Y, et al. Natural BH3 mimetic (-)-gossypol chemosensitizes human prostate cancer via Bcl-xL inhibition accompanied by increase of Puma and Noxa. Mol Cancer Ther. 2008;7:2192–2202. doi:10.1158/1535-7163.MCT-08-0333
56. Zhou W, Xu J, Li H, et al. Neddylation E2 UBE2F promotes the survival of lung cancer cells by activating CRL5 to degrade NOXA via the K11 linkage. Clin Cancer Res. 2017;23:1104–1116. doi:10.1158/1078-0432.CCR-16-1585
57. Wu X, Luo Q, Liu Z. Ubiquitination and deubiquitination of MCL1 in cancer: deciphering chemoresistance mechanisms and providing potential therapeutic options. Cell Death Dis. 2020;11(7):556. doi:10.1038/s41419-020-02760-y
58. Wertz IE, Kusam S, Lam C, et al. Sensitivity to antitubulin chemotherapeutics is regulated by MCL1 and FBW7. Nature. 2011;471(7336):110–114. doi:10.1038/nature09779
59. Ding Q, He X, Hsu J-M, et al. Degradation of Mcl-1 by beta-TrCP mediates glycogen synthase kinase 3-induced tumor suppression and chemosensitization. Mol Cell Biol. 2007;27:4006–4017. doi:10.1128/MCB.00620-06
60. Choudhary GS, Al-Harbi S, Mazumder S, et al. MCL-1 and BCL-xL-dependent resistance to the BCL-2 inhibitor ABT-199 can be overcome by preventing PI3K/AKT/mTOR activation in lymphoid malignancies. Cell Death Dis. 2015;6:e1593. doi:10.1038/cddis.2014.525
61. Mazumder S, Choudhary GS, Al-Harbi S, Almasan A. Mcl-1 phosphorylation defines ABT-737 resistance that can be overcome by increased NOXA expression in leukemic B cells. Cancer Res. 2012;72(12):3069–3079. doi:10.1158/0008-5472.CAN-11-4106
62. Yamaguchi R, Perkins G. Mcl-1 levels need not be lowered for cells to be sensitized for ABT-263/737-induced apoptosis. Cell Death Dis. 2011;2:e227. doi:10.1038/cddis.2011.109
63. Yecies D, Carlson NE, Deng J, Letai A. Acquired resistance to ABT-737 in lymphoma cells that up-regulate MCL-1 and BFL-1. Blood. 2010;115(16):3304–3313. doi:10.1182/blood-2009-07-233304
64. Kotschy A, Szlavik Z, Murray J, et al. The MCL1 inhibitor S63845 is tolerable and effective in diverse cancer models. Nature. 2016;538:477–482. doi:10.1038/nature19830
65. Tai-Shun L, Schinazi RF, Zhu J, et al. Anti-HIV-1 activity and cellular pharmacology of various analogs of gossypol. Biochem Pharmacol. 1993;46(2):251–255. doi:10.1016/0006-2952(93)90411-O
66. Zhao Y, Xiong X, Sun Y. Cullin-RING ligase 5: functional characterization and its role in human cancers. Semin Cancer Biol. 2020;67:61–79. doi:10.1016/j.semcancer.2020.04.003
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.