Back to Journals » Infection and Drug Resistance » Volume 19
Targeting Horizontal Gene Transfer to Combat Antimicrobial Resistance: A Review of Mechanisms, Drivers, and Multi-Omics Strategies
Authors Tang L ![]() , Yang W, Yang L, Lv Y, Zhang J
, Yang W, Yang L, Lv Y, Zhang J
Received 18 December 2025
Accepted for publication 24 March 2026
Published 2 April 2026 Volume 2026:19 589962
DOI https://doi.org/10.2147/IDR.S589962
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Lili Tang,1 Wei Yang,1 Lingqi Yang,1 You Lv,2 Jian Zhang1
1Department of Pharmacy, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China; 2Information Center of West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China
Correspondence: Jian Zhang, Department of Pharmacy, West China Hospital, Sichuan University, Chengdu, 610041, People’s Republic of China, Email [email protected]
Abstract: The widespread dissemination of antibiotic resistance genes in bacteria primarily relies on horizontal gene transfer (HGT), a phenomenon that has profound implications for global healthcare and animal husbandry. Therefore, elucidating the key mechanisms of HGT is crucial for controlling the global spread of resistance genes. Horizontal gene transfer can occur not only through classical pathways such as conjugation, transformation, and transduction but also involves non-classical mechanisms including gene transfer agents, outer membrane vesicles, and nanotubes. This process is mediated by various mobile genetic elements, such as plasmids, bacteriophages, transposons, integrons, integrative and conjugative elements (ICEs), and integrative and mobilizable elements (IMEs). HGT is typically regulated by a combination of host-specific intrinsic factors and external environmental conditions. To address the spread of resistance, numerous detection and prevention tools targeting this mechanism have been developed. This article focuses on the process of HGT and its associated mobile genetic elements, systematically analyzes key factors influencing this process, summarizes sequencing and bioinformatic technologies used for monitoring HGT, and explores prevention strategies informed by genomic, proteomic, and metabolomic approaches. The aim is to provide a theoretical foundation and practical guidance for the control of drug-resistant bacteria.
Keywords: HGT, ARGs, MGEs, control of drug-resistant bacteria
Introduction
As an ancient cornerstone of Earth’s ecosystems, bacteria have developed unparalleled environmental adaptability through their 3.5-billion-year evolutionary journey, forming complex communities from deep-sea hydrothermal vents to human mucosal surfaces.1 This ubiquitous presence endows bacteria with a dual role: while commensal species maintain host metabolic homeostasis, pathogenic species such as Mycobacterium tuberculosis and Streptococcus pneumoniae cause approximately 7.7 million annual deaths, accounting for 13.4% of global mortality and representing the second-largest health threat after cardiovascular diseases.2,3 The 20th-century discovery of antibiotics temporarily shifted the balance of power between humans and pathogens—for instance, β-lactams irreversibly bind penicillin-binding proteins (PBPs) to block peptidoglycan cross-linking, while quinolones inhibit DNA gyrase to trigger chromosomal fragmentation.4,5 These “chemical weapons” dramatically reduced infection-related mortality. Yet beneath this triumph lurked a crisis: excessive antibiotic use in medical, livestock, and agricultural sectors accelerated bacterial evolution, enabling their survival through the development of resistance.6–8
The essence of antibiotic treatment failure lies in bacterial adaptive countermeasures against chemical intervention, where adaptive changes propagate vertically or horizontally among bacteria.9,10 HGT enables resistance genes like blaNDM−1 to jump between species via plasmids at frequencies much higher than those of vertical inheritance, directly fueling the global spread of multidrug-resistant (MDR) superbugs.11 Carbapenem-resistant Acinetobacter baumannii (CRAB) disseminates through conjugative plasmids in intensive care units (ICUs), elevating sepsis mortality to 74%, while vancomycin-resistant enterococci (VRE) contaminate medical devices, triggering hospital outbreaks.12,13 Modeling projections indicate that without effective intervention, antimicrobial resistance (AMR)-related annual deaths will exceed 10 million by 2050, surpassing current cancer mortality.14,15
HGT manifests through diverse mechanisms: classical pathways include transformation, conjugation, and transduction, while non-classical routes encompass gene transfer agents (GTAs), outer membrane vesicles (OMVs), and nanotubes.16–24 Transformation efficiency is governed by the expression of comEC, conjugation involves nanoscale channel assembly via type IV secretion systems (TraA protein), with the oriT site of plasmids initiating single-stranded DNA transfer under the catalysis of relaxase (NikB).25–27 Transduction requires the integration of phage genes into the host genome.28 GTAs can modify the functions of the host through genomic insertion, while OMVs and nanotubes mediate the entry of resistance gene-containing DNA and plasmids into recipient bacteria, enhancing horizontal transfer efficiency.29–32 Throughout this process, key genetic elements—plasmids, integrons, transposons, ICEs, IMEs, and prophages—can be detected in environmental samples via high-throughput sequencing coupled with bioinformatic analysis.33–39 The efficiency of HGT is modulated by host states and environmental factors.40,41 Targeted interventions at genetic and protein levels can mitigate clinical resistance events, exemplified by CRISPR-Cas9 systems that cleave resistance plasmids and chemical inhibitors that disrupt HGT-associated proteins to block cellular uptake and chromosomal integration.42–44
Consequently, this review systematically delineates the mechanisms of HGT and associated genetic elements, investigates factors influencing HGT efficiency, summarizes genomic and bioinformatic applications in resistance gene identification and outbreak prediction, and explores high-efficacy human interventions against HGT—ultimately charting a strategic roadmap for humanity’s scientific campaign against drug-resistant bacteria.
The Mechanisms of HGT
Horizontal transfer of antibiotic resistance genes (ARGs) plays a pivotal role in bacterial evolution under antibiotic selection pressure, driving the emergence of untreatable “superbugs” by concentrating multiple ARGs within single cells. Understanding HGT mechanisms is therefore crucial for combating resistant bacteria. Classical and non-classical pathways facilitate HGT, disseminating ARGs between bacteria.
The Classical Mechanisms of HGT
The classical mechanisms of HGT comprise transformation, conjugation, and transduction.45–47 Transformation represents the oldest HGT mechanism, wherein donor bacteria release 7–50 kb DNA fragments during cell death, active secretion, or specific growth phases.48 The recipient uptakes extracellular DNA (eDNA), which may integrate into chromosomes via homologous recombination—a process demanding stringent sequence complementarity. Only DNA fragments with allelic counterparts in the recipient’s genome can integrate, while completely heterologous sequences are degraded by nucleases after uptake, positioning the recipient bacterium as the key actor in this process.49,50 The Type VI Secretion System (T6SS) critically enhances HGT efficiency by enabling donor cells to secrete DNA, deliver virulence factors, and lyse neighboring cells.51–53 In naturally transformable bacteria, DNA uptake involves proteins encoded by the com regulon: exogenous double-stranded DNA (dsDNA) enters through ComEC/ComA transmembrane channels as single-stranded DNA (ssDNA) in a 3′-5′ direction. Internalized ssDNA binds DNA processing protein A (DprA), which recruits RecA recombinase to facilitate homology search and strand invasion. RecA then mediates homologous recombination between the ssDNA and recipient DNA, forming hybrid duplexes.25,54–56 Environmental ARGs primarily originate from actively secreted or cell death-derived eDNA. Although 99% of eDNA degrades within 7 days in natural settings, 11 of 12 priority antibiotic-resistant pathogens acquire eARGs via natural transformation in clinical environments. Moreover, soil eARG concentrations increase 20-fold with 5-fold higher resistant bacterial counts, underscoring transformation’s significant role in environmental resistance evolution.57,58
Conjugation facilitates ARG transfer through direct cell contact mediated by large (>30 kb) dsDNA plasmids. This mechanism was first demonstrated in 1946 by Joshua Lederberg and Edward Tatum using bacterial auxotrophs to prevent revertant interference.59 Classical F-plasmid-driven conjugation occurs intra- and interspecifically via three modes: F⁺×F−, Hfr×F−, and F′×F−. F⁺ denotes bacteria with cytoplasmic F-plasmids, F− lacks F-factors, Hfr results from F-factor chromosomal integration, and F′ carries F-factors recombined with host genes. The latter two modes transfer partial chromosomal DNA to recipients.60–62 The F-plasmid structure includes oriT (transfer origin), tra (transfer gene cluster encoding conjugation proteins, constituting 1/3 of plasmid length), par (partition genes), ssb (single-strand binding protein), RepFIC/RepFIB (replication origins), Tn1000 (transposon), and finO/sraB (regulatory genes).63,64 Conjugative ARG transfer predominates in Enterobacteriaceae like Escherichia coli (E. coli), Klebsiella, Salmonella, Shigella, and Citrobacter.65–69 Broad-host-range plasmids (eg., IncP-1) enable cross-genera transfer between Gram-negative proteobacteria and Gram-positive Bacillus species,70–72 making conjugation the most extensively studied HGT type.
Transduction introduces >100 kb DNA fragments into bacterial chromosomes via viruses.73,74 Bacteriophages serve as primary vectors, operating through lytic or lysogenic cycles. Lytic phages replicate within hosts, lysing cells to release progeny virions that infect new bacteria.75,76 Lysogenic phages integrate into specific host genomic sites, remaining dormant until induced to enter lytic cycles. Upon host lysis, phage particles may package bacterial DNA fragments, transferring these genes to subsequent hosts during infection.77 Transduction occurs via generalized (any host gene packaged post-lysis) or specialized (only host genes adjacent to integrated phage DNA transferred) mechanisms.78
The Nonclassical Mechanisms of HGT
Beyond classical mechanisms, bacterial DNA transfer also occurs through gene transfer agents (GTAs), outer membrane vesicles (OMVs), and pilus-like nanotubular structures.79,80 GTAs are hypothesized to originate from ancestral phages, sharing structural features such as capsids and tails, yet predominantly carrying non-self DNA for horizontal transfer rather than for self-replication—thus representing a “domesticated” viral vector. Upon genomic insertion, GTAs can modulate host bacterial motility, quorum sensing, exopolysaccharide synthesis, and biofilm formation, playing pivotal roles in bacterial and archaeal evolution.81–85 GTAs differ from viruses in four key aspects: they package random host DNA fragments for delivery to compatible recipients, exhibit fragmented genomes at multiple chromosomal loci, cannot fully package their own encoding genes, and replicate vertically with the host genome.82 Three distinct GTA lineages are recognized: Type I (α-proteobacteria; model host: Rhodobacter spp)., Type II (α-proteobacteria; model host: Bartonella spp)., and Type III (spirochetes; model host: Brachyspira hyodysenteriae).86 The GTA of Rhodobacter capsulatus (RcGTA) exemplifies a hybrid mechanism where donor DNA is packaged in transducing phage-like particles while recipient cells employ natural transformation machinery for DNA uptake, facilitating environmental adaptation and biofilm maintenance.87,88
Outer membrane vesicles, spherical lipid nanostructures released by bacteria, are ubiquitous in both Gram-positive and Gram-negative species.89 They were first observed in E. coli (1965), later documented in Brucella through electron microscopy and SDS-PAGE analysis (1987), and confirmed in Staphylococcus aureus (S. aureus) (2009).31,90,91 OMVs carry virulence factors, plasmids, adhesins, toxins, and nucleic acids, enabling functions like intercellular communication, immune modulation, and virulence dissemination. Their role in HGT—termed vesiduction—involves transferring DNA fragments >30 kbp (single-copy prokaryotic sequences).31,79,92–94 OMV-encapsulated DNA exhibits enhanced stability and transfer efficiency due to protection from exonucleases and resistance to environmental dilution.95 Marchant et al demonstrated OMV-mediated transfer of ΔtolR (bacterial envelope maintenance) and ΔdegS (σE activation for misfolded proteins) genes from Salmonella Typhi to polymyxin B-susceptible bacteria.96 Qiao et al achieved functional transfer of the nirS gene via OMVs, demonstrating that E. coli DH5α released OMVs containing recombinant pET28a-nirS-EGFP plasmids. This yielded a transformation frequency of 2.76×106 CFU/g and active nitrite reductase expression (18.16 U/mL) in recipient E. coli BL21 cells at 200 μg OMV concentrations.97 Federica et al further established OMVs as key vectors for Klebsiella pneumoniae to disseminate β-lactam resistance plasmids across species including E. coli, Salmonella enterica, Pseudomonas aeruginosa, and Burkholderia cepacia.98
Nanotube-mediated HGT, discovered by Dubey et al in adjacent Bacillus subtilis (B. subtilis) cells, facilitates intercellular transfer of nutrients, electrons, toxins, and genetic material.32,99,100 These structures exhibit species-specific dimensions: Thermococcus species produce micron-length nanotubes often filled with extracellular vesicles, while the hyperthermophilic crenarchaeon Pyrodictium forms nanotubes with only 25 nm.101 Antibiotic stress significantly increases nanotube production in pathogens like Salmonella Typhi, Enterococcus faecalis, Proteus spp., and Klebsiella spp.102 Crucially, Dubey demonstrated the co-transfer of chloramphenicol acetyltransferase (cat) and erythromycin resistance methylase (erm) genes between B. subtilis strains via nanotubes, generating dual-resistant bacteria, with cross-genera transfer observed in E. coli and S. aureus.99
Mobile Genetic Elements (MGEs) Associated with HGT
Plasmids
Plasmids, as mobile genetic elements, participate in genetic material transfer across multiple horizontal gene transfer (HGT) forms and represent the most crucial mobile elements. Unique to prokaryotes, these extrachromosomal DNA molecules replicate independently within cells or integrate into bacterial chromosomes for replication. Their classification into incompatibility (Inc) groups stems from the inability of plasmids from the same group to coexist stably within a single bacterial cell. Plasmids sharing an Inc group typically possess identical or similar replication/partitioning systems; however, when two such plasmids coexist, interference in replication occurs, leading to the loss of one plasmid during cell division. This results in uneven plasmid distribution within bacterial populations, where strains carrying environmentally advantageous plasmids gain survival benefits. Plasmids serve as reservoirs for ARGs, with 5% of neonatal gut bacteria carrying ARG-harboring plasmids.103 They carry key resistance genes, exemplified by plasmid-borne colistin resistance genes (mcr), which accelerate the emergence of colistin resistance in multidrug-resistant Gram-negative bacteria, posing a significant global health threat. In late 2015, Chinese researchers identified the first plasmid-mediated colistin resistance gene, mcr-1, in E. coli. This gene encodes a PmrC-like pEtN transferase that modifies lipid A by transferring pEtN from the cell membrane, conferring colistin resistance. Subsequently, nine mcr-1 homologs (mcr-2 to mcr-10) were discovered worldwide.104–114 Current research indicates that nearly all colistin resistance in E. coli is mediated by plasmid-borne mcr-1, while mcr genes can be carried by diverse plasmids. For instance, mcr-1-bearing IncI2-like and IncX4 plasmids have disseminated globally, and mcr-3 has been identified on various plasmid types including IncHI2, IncA/C, and IncFII/FIB, with IncHI2 being predominant.115,116 The lipid A modification mediated by these genes reduces the negative charge of the bacterial outer membrane, consequently enhancing resistance to antimicrobial peptides (AMPs) produced by intestinal epithelial cells (β-defensins 1–3 and LL37) and Paneth cells (α-defensins 5 and 6).106,117,118
Transposons and Integrons
Transposons, as mobile DNA sequences within genomes, comprise two categories: Class I retrotransposons replicate via RNA intermediates and integrate into genomes using reverse transcriptase, while Class II DNA transposons employ a “cut-and-paste” mechanism catalyzed by transposase and feature terminal inverted repeats (IRs). These elements stably insert into host genomes, carrying foreign genes ranging from several kb to tens of kb, enabling large resistance gene fragments from external DNA to integrate into host chromosomes for stable expression. For instance, the chromosomally integrated Tn1549 transposon in vancomycin-resistant Enterococcus faecium ST117 harbors a highly mutated vanB2 operon.119 Integrons capture and express exogenous genes, particularly accelerating resistance gene integration and dissemination among bacteria under antibiotic pressure. Classified into five types based on IntI integrase amino acid sequences, Class 1 integrons are most prevalent and notorious, distributed widely across Gram-negative and Gram-positive bacteria including E. coli, Pseudomonas, Salmonella, Corynebacterium, Streptococcus, Enterococcus, and Staphylococcus. They associate with diverse resistance gene cassettes such as aadA (streptomycin resistance), bla (β-lactam resistance), dfrA (trimethoprim resistance), and sul1 (sulfonamide resistance). This integron family achieves mobility through association with Tn402-like transposons and Tn3 family elements (eg., Tn21, Tn1696).120,121 Class 1 integrons were detected in 84/91 multidrug-resistant (MDR) and extensively drug-resistant (XDR) Pseudomonas aeruginosa isolates from burn wound infections in Bangladesh.122 Classes 2 and 3 integrons exhibit lower clinical detection rates in Gram-negative bacteria, mediating resistance to trimethoprim, streptothricin, streptomycin, chloramphenicol, erythromycin, and β-lactams.123–125 Class 4 and 5 integrons are specifically associated with Vibrio cholerae and Vibrio salmonicida, respectively.126,127
ICEs and IMEs
Integrative and conjugative elements (ICEs) and integrative and mobilizable elements (IMEs) are composite genetic units that carry multiple functional modules enabling autonomous excision and integration within bacterial genomes, serving as critical vectors for disseminating acquired antibiotic resistance genes. ICEs contain core genes including int (encoding an integrase, typically a tyrosine recombinase that recognizes chromosomal target sites for site-specific recombination), xis (encoding an excisionase that directs recombination orientation), a type IV secretion system (T4SS, facilitating ICE transfer between bacteria), and nic/oriT (the relaxase recognition site initiating ICE replication and conjugation). Normally integrated into the host chromosome with conjugation genes silenced, ICEs excise to form double-stranded DNA circles upon induction. The T4SS then assembles a transfer channel, where relaxase nicks oriT to generate single-stranded T-DNA. This T-DNA translocates through the channel into recipient cells, reforms a double-stranded circular structure, and integrates into the recipient genome via integrase activity. In contrast, IMEs require auxiliary conjugation machinery for interbacterial resistance gene transfer. In seafood sold in China, tigecycline-resistant bacteria were identified with Shewanella spp. predominating (33/76 isolates, 43.4%). Genomic sequencing revealed that tet(X4) and tmexCD2-toprJ2 resistance genes in resistant Shewanella reside on SXT/R391-family ICEs. Thus, SXT/R391 ICEs mediate tigecycline resistance genes among aquatic bacteria, warranting further investigation into their contribution to clinical tigecycline resistance.128 Similarly, the ICE designated ICECiPOL15 was identified in the β-lactam-resistant Chryseobacterium indologenes strain POL15, harboring a class C β-lactamase gene (blaAQU) alongside genes for integrase, excisionase, relaxase, T4SS coupling proteins, and conjugative transposon proteins.129 Additionally, Guillem López de Egea demonstrated that the tetracycline resistance gene tet(M) in Streptococcus dysgalactiae subsp. equisimilis (SDSE) can reside within Tn916-family ICEs and a novel IME designated IME_SDSE_HUB-4529.130
Prophage
A prophage refers to the genome of a temperate bacteriophage that has integrated into the host bacterial chromosome. Typically dormant, it does not immediately lyse the host cell but replicates passively with the host during cell division. Prophages can persist long-term in bacterial genomes and may reactivate under specific conditions, entering the lytic cycle to produce new phage particles that rupture the host cell. The prophage genome contains integration and excision genes (mediating chromosomal integration and excision to initiate lysis), replication genes (encoding enzymes for accurate genome replication), transcription and translation genes (including promoters, operators, and regulatory proteins for gene expression), structural protein genes (encoding capsid and tail components for virion assembly), auxiliary metabolic genes (involved in bacterial metabolism and environmental adaptation), toxin/virulence factor genes (conferring pathogenicity), and antibiotic resistance genes (providing antibiotic resistance). These prophages play crucial roles in disseminating resistance genes within host bacteria. Tony et al observed that higher prophage abundance in synovial fluid samples from periprosthetic joint infection patients correlates with elevated antibiotic resistance gene counts.131 Furthermore, complex interactions exist between phages and plasmids in spreading ARGs.132
Niche-Specific Facilitators of HGT
Niche-specific facilitators of HGT encompass both host-related and environmental factors, where the genetic compatibility and ecological coexistence between recipient and donor bacteria critically determine the success of resistance gene transfer, while environmental elements such as antibiotic usage, biofilm formation, and plastic presence significantly influence the probability of horizontal resistance gene dissemination across bacterial populations. Delving into these mechanisms is essential for developing effective containment strategies.
Host Factors
Bacterial genetic compatibility refers to the ability of different bacteria to achieve compatible integration and stable transmission of genetic material during transfer processes.133 This characteristic not only determines the efficiency of antibiotic resistance gene transfer—for instance, conjugation efficiency between highly compatible E. coli and Salmonella strains can increase by 3- to 5-fold—but also governs post-transfer stability, where codon usage compatibility between donor and recipient bacteria serves as the critical determinant for successful integration.134,135 Ecological coexistence, manifested through physical proximity and metabolic interdependence among microorganisms in specific environments, significantly facilitates resistance gene exchange: mutualistic symbiosis between Enterococcus faecalis and Bacteroides in the human gut accelerates horizontal transfer of macrolide resistance gene ermB and tetracycline resistance gene tetM; biofilm alliances formed by Staphylococcus and Corynebacterium in skin microenvironments drive dissemination of β-lactamase gene blaCTX-M; whereas metabolic cooperation between Aeromonas and Chloroflexi in wastewater systems promotes prevalence of multidrug efflux pump gene mexAB-oprM.136–144 Concurrently, host-intrinsic factors regulate resistance gene acquisition through multiple mechanisms: carbon starvation activates the ComK pathway in B. subtilis to enhance transformation competence, while mitomycin C-induced DNA damage in Pseudomonas aeruginosa triggers SOS response (recA upregulation) increasing conjugation frequency 10-fold.145–148 Host restriction-modification systems constitute key barriers—E. coli Type I restriction enzymes reduce conjugative efficiency by >80% when methylation patterns are incompatible, and the CRISPR-Cas9 system in Streptococcus pneumoniae blocks stable integration of macrolide resistance gene mefA by PAM-guided DNA cleavage.149–151 Genomic architecture equally influences resistance maintenance, exemplified by Salmonella pathogenicity island SGI1 capturing cephalosporin resistance gene blaCTX-M through site-specific recombination.152 These bacterial factors, shown in Figure 1, synergize with environmental pressures to shape HGT “hotspots”, whose mechanistic elucidation provides critical theoretical foundations for targeted containment strategies.
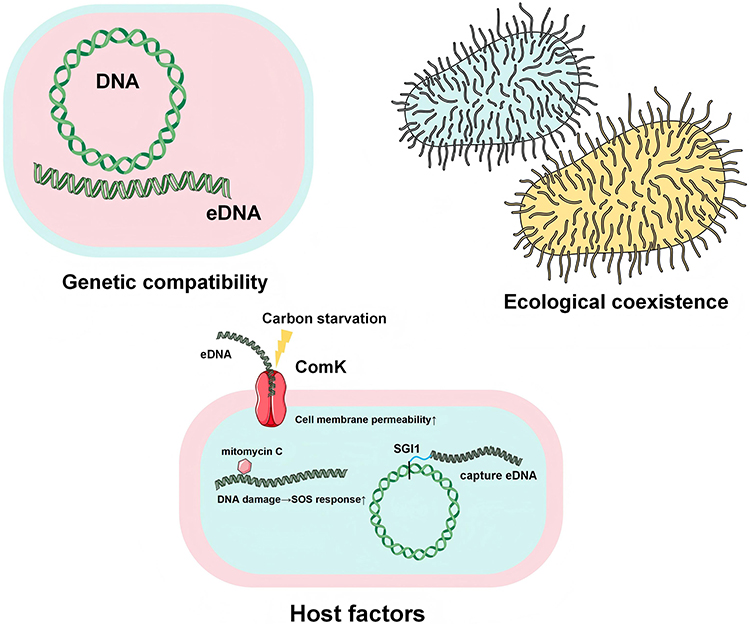 |
Figure 1 Bacterial Factors Contributing to Enhanced HGT. Genetic compatibility: the capacity for compatible integration between bacterial endogenous DNA and exogenous DNA (eDNA) during transfer processes; Ecological coexistence: mutualistic symbiosis achieved through physical proximity and biological communication between two or more bacterial species within specific environments; Host factors: 1) carbon starvation activates ComK on the cell membrane, enhancing membrane permeability to eDNA, 2) mitomycin C-induced host DNA damage upregulates the SOS response, facilitating integration of endogenous DNA with eDNA, 3) specialized genomic structures such as SGI1 enable eDNA capture. |
Environmental Factors
Environmental factors significantly accelerate the horizontal transfer and cross-media dissemination of antibiotic resistance genes via driving mechanisms such as antibiotic selection pressure, biofilm microenvironments, and plastics (shown in Figure 2). The widespread use of antibiotics directly drives horizontal resistance gene transfer, where environmental antibiotic residues (eg., subinhibitory concentrations of tetracycline) activate bacterial SOS stress responses (recA upregulation), increasing conjugative efficiency by up to 3.19-fold while promoting class 1 integrons to capture exogenous resistance genes.153,154 In aquaculture and medical wastewater, high-concentration antibiotics further select for strains carrying multidrug-resistant genes, accelerating resistance gene diffusion within environmental microbiota.155 Biofilms serve as “hotspot microenvironments”, where their three-dimensional structures enhance HGT through multiple mechanisms: extracellular polymeric substance (EPS) matrices encase bacteria to form physical barriers that block antibiotic penetration while prolonging plasmid conjugation duration; quorum sensing (QS) systems (eg., lasI/R in Pseudomonas aeruginosa) upregulate tra gene expression, elevating conjugation frequency within biofilms compared to free states; additionally, metabolic gradients in biofilms induce bacterial competence, augmenting uptake of free resistance genes such as ermB.156–160 For instance, S. aureus biofilms on hospital catheters exhibit transposon Tn916-mediated tetracycline resistance gene (tetM) transfer rates reaching 10−3 per cell.161 Plastic pollution provides persistent vectors for resistance genes: microplastics (<5 mm) adsorb pathogens and free DNA via high surface area, forming “plastic-biofilm” complexes that protect ARG (eg., mcr-1) from environmental degradation.162–166 Crucially, plastic surface hydrophobicity promotes tight adhesion between diverse bacterial species, elevating cross-species plasmid conjugative efficiency.167,168 Studies confirm that polyethylene microparticles in wastewater treatment plant effluents concentrate ARGs abundance at 102-103 times higher levels than ambient water, entering agricultural ecosystems through irrigation and forming invisible chains of resistance transmission.169,170
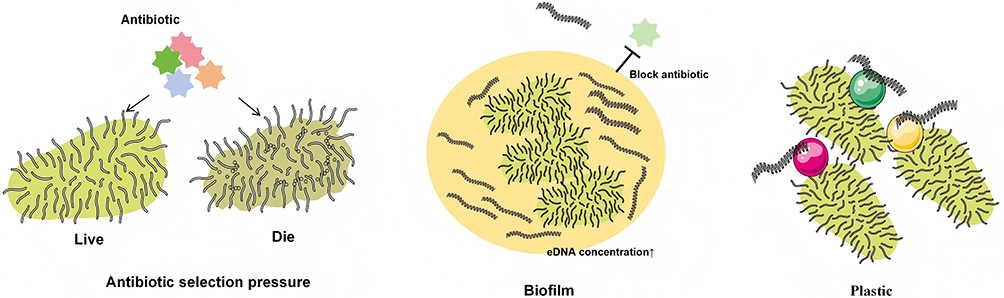 |
Figure 2 Environmental Factors Contributing to Enhanced HGT. Antibiotic selection pressure: the presence of antibiotics in the environment selects for the survival of antibiotic-resistant bacteria while eliminating non-resistant bacteria; Biofilm: biofilms in the environment enrich bacteria and exogenous DNA (eDNA), increasing the local concentration of eDNA, and additionally form physical barriers that impede antibiotic penetration; Plastics: the hydrophobic properties of plastic surfaces facilitate tight adhesion of bacteria and eDNA. |
Applications of Genomics in HGT
The advancement of high-throughput sequencing technologies and bioinformatics has enabled comprehensive genomic studies of bacteria. Current research utilizes these tools to investigate HGT.
High-Throughput Sequencing Technologies
Whole Genome Sequencing (WGS) precisely tracks horizontal transfer of resistance genes by resolving genome-wide variation sites. In clinical outbreak tracing, WGS identifies single-nucleotide polymorphism (SNP) differences between strains (threshold of ≤5 SNPs) and reconstructs mobile genetic elements carrying resistance genes using plasmid assembly tools (eg., PlasmidSPAdes). Applying this approach, Shravani et al revealed diverse resistance plasmids in carbapenemase-producing E. coli and K. pneumoniae isolated from Indian ICUs: blaNDM−1, blaNDM−5, blaNDM−7 (21/31), blaKPC−2 (1/31), and blaOXA−181, blaOXA−232 (8/31).171 Compared to individual-level WGS, metagenomic sequencing (mNGS) uncovers dynamic resistance gene exchange networks at the community scale. Through unbiased sequencing of total DNA combined with assembly and binning techniques, it reconstructs microbe-function correlation maps. Li et al employed population metagenomics in bovine fecal E. coli communities, detecting co-localization of multiple ARGs on the same MGE, which was linked to prevalent resistance phenotypes in isolates. Population metagenomics identified significantly distinct resistomes with higher ARG relative abundance and HGT risk compared to direct mNGS.172 Meanwhile, technical innovations enhance HGT monitoring resolution. Oxford Nanopore long-read sequencing (>10 kb) resolves repetitive regions, directly characterizing mobile genetic elements flanking resistance genes while avoiding short-read assembly errors. Francesca et al identified a 237-kb conjugative plasmid carrying acquired blaTEM−1 in a hypervirulent E. coli ST73 strain isolated from a clozapine-treated schizophrenia patient.173 EpicPCR (emulsion, paired isolation, and concatenation PCR)—a single-cell technology—links target genes to 16S fragments (spanning V4-V9, ~1000 bp) via optimized primer design, advancing host-specific ARG detection in microbial communities.174 These technologies demonstrate critical value in clinical surveillance. Kim et al analyzed carbapenemase-producing Enterobacteriaceae (CPE) and vancomycin-resistant Enterococcus (VRE) using WGS to investigate hospital-acquired transmission. Infectious disease inpatients showed higher CPE/VRE acquisition rates, with the first hospitalization week being the critical window. Dementia, central/urinary catheters, and carbapenem use were significant risk factors. Hospital-acquired CPE/VRE shared resistance genes with circulating strains, while community-acquired cases carried novel genes (eg., blaOXY−4-1, optrA). Microbiome analysis revealed significantly altered species abundance and reduced diversity at discharge.175
Bioinformatics Tools
The maturation of sequencing technologies has catalyzed explosive growth in bioinformatics, with extensive applications in clinically relevant contexts. Numerous bacterial genome databases—such as CARD and HGTree—have been established, providing researchers with rich genomic data resources (Table 1). Bioinformatics tools are being developed to efficiently process genomic data, construct resistance gene transmission networks, predict resistance trends, support precision medical decisions, optimize treatment strategies, and accelerate novel drug development. For instance, the Comprehensive Antibiotic Resistance Database (CARD) integrates multi-source data encompassing resistance genes, mutations, and resistance mechanisms, while also offering analytical tools and algorithms for antibiotic resistance research and surveillance.176 The HGTree database serves as a comprehensive HGT repository containing 2,472 prokaryotic genome HGT events,177 with its latest version HGTree v2.0 expanding to 20,536 non-redundant prokaryotic genomes and rigorously filtered HGT data. Additionally, HGTree v2.0 provides extended datasets including 6,361,199 putative horizontally transferred genes annotated with KEGG pathways, virulence factors, and antibiotic resistance features.178 The development of key bioinformatic tools focuses on mobile element identification, resistance gene detection, resistance phenotype prediction, and HGT network modeling (Table 2). PlasFlow distinguishes chromosomal from plasmid sequences in metagenomic data,179 while Plascad classifies bacterial plasmids, co-locates AMR genes, and visualizes plasmid structures.180 Prophage Hunter employs machine learning to assess prophage activity states and dissemination potential.181 ARG identification leverages tools like ResFinder and PointFinder (from the ResFinder database) alongside CARD’s Resistance Gene Identifier (RGI), both capable of detecting acquired resistance genes and point mutations.182 Breakthroughs in resistance transmission prediction include Zhou et al’s algorithm for constructing HGT networks in pathogens like Acinetobacter baumannii and E. coli to forecast antibiotic resistance emergence, achieving AUROC = 0.990 predictive performance.183 Pioneering live-tracing technologies integrate cross-disciplinary approaches, exemplified by Gong et al’s CRISPR-AMRtracker—a fluorescent tracing tool combining 16S rRNA sequencing, CRISPR/Cas9 tagging, fluorescence-activated cell sorting, and microbiome analysis. This system integrates fluorescent markers downstream of ARGs without compromising host antibiotic susceptibility, fitness, conjugation, or transposition, offering novel perspectives for studying ARG dissemination in complex microbiomes and elucidating resistance evolution mechanisms.184
 |
Table 1 The Classic Bacterial Genome Databases |
 |
Table 2 The Bioinformatic Tools for HGT Researches |
Strategies for Curbing HGT
Multi-omics strategies offer novel approaches to inhibit antibiotic resistance evolution by targeting multiple stages of HGT, restoring microbial ecological balance.
Genomic Strategies
Genomic strategies primarily rely on targeting MGEs like plasmids and transposons to combat HGT. The CRISPR-Cas system has emerged as a key tool to block this process through guide RNA (gRNA)-mediated precise cleavage. Within the same bacterial species, strains carrying the CRISPR-Cas system contain significantly fewer plasmids than those without it (0.86 vs. 1.86). Furthermore, CRISPR-Cas-positive strains show a 31% reduction in intact prophages (1.17 vs. 1.75). These data support the role of CRISPR-Cas in reducing the incidence of HGT.208 Using PCR technology, Tao et al detected the CRISPR-Cas system, antibiotic resistance genes, and virulence factors in clinical Enterococcus isolates. They found that 46 strains (46.0%) carried at least one CRISPR-Cas locus. Compared to CRISPR-Cas-negative isolates, CRISPR-Cas-positive isolates exhibited significantly lower resistance rates to ampicillin, erythromycin, levofloxacin, tetracycline, vancomycin, gentamicin, streptomycin, and rifampicin. Fewer CRISPR loci were identified in isolates carrying resistance genes such as vanA, tetM, ermB, aac6′-aph(2″), aadE, and ant(6). Additionally, the CRISPR-Cas locus was significantly negatively correlated with the enterococcal virulence factor enterococcal surface protein gene (esp).209 To target carbapenem-resistant E. coli carrying the blaKPC-2 gene, the same team constructed a specific prokaryotic CRISPR-Cas9 system plasmid designed to cleave blaKPC−2. Within 8 hours, this system achieved over 98.8% clearance of the resistance plasmid carrying blaKPC-2, restoring bacterial susceptibility to carbapenems.210 In Streptococcus pyogenes (S. pyogenes), two distinct CRISPR loci were identified. Strains lacking CRISPR contained significantly more prophages than those carrying CRISPR. Although S. pyogenes CRISPR arrays contained fewer spacers than those in other streptococci, a significant negative correlation was confirmed between validated spacer content and prophage abundance. Thus, the CRISPR system in S. pyogenes can restrict the acquisition of prophage-derived resistance determinants.211 Moreover, engineered phage technology can block transduction through a dual mechanism: first, by screening naturally lytic phages (such as JG004 targeting Pseudomonas aeruginosa) that specifically lyse strains carrying certain resistance genes (eg., aac(6′)-Ib-cr);212,213 second, by genetically modifying phages—enhancing their recognition and targeting capabilities—to achieve precise elimination of resistant bacteria.214–216 In patients with uncomplicated urinary tract infections caused by E. coli, a CRISPR-Cas3-enhanced phage cocktail, LBP-EC01, was evaluated in a Phase II clinical trial. When used in combination with trimethoprim-sulfamethoxazole (TMP-SMX), it resulted in rapid and sustained reduction of E. coli load in urine and corresponding resolution of clinical symptoms in evaluable patients.217
Proteomic and Metabolomic Strategies
The homeostasis of microbial communities serves as a natural barrier against the invasion of antibiotic resistance genes. Restriction-modification (R-M) systems, composed of DNA methyltransferases and restriction endonucleases, function as a form of “genomic immune defense” in bacteria by recognizing and degrading foreign unmethylated DNA, such as invading resistance plasmids, and are regarded as a barrier to horizontal gene exchange.218–220 Park et al performed comparative genomic analysis of S. aureus isolates associated with bovine mastitis and human infections, and found that the distribution of strain-specific genes, virulence genes, and AMR genes was closely linked to the presence of R-M systems. This suggests that R-M systems may promote clonal diversification by providing a genetic barrier against HGT. Moreover, lineage-specific R-M systems may restrict genetic exchange between strains from different lineages.221 Xu et al analyzed the distribution of AMR genes associated with MGEs in the context of a novel DNA phosphorothioate (PT) modification-based R-M system. They found that the presence of PT R-M effectively reduced the genomic distribution of horizontally acquired AMR genes. Furthermore, pangenome analysis and assessment of unique gene variants through MGE prediction demonstrated that PT R-M presence suppresses the frequency of HGT.222 Djordjevic et al developed a mathematical model reflecting the dynamics of R-M systems, linking changes in the methyltransferase-to-restriction enzyme (M/R) ratio to factors such as cell division time and plasmid replication rates, to predict the emergence of bacterial population heterogeneity resulting from horizontal gene transfer.223 QS is an interbacterial communication process that regulates numerous phenotypes and gene expression patterns, including ARGs and genes involved in AMR development. Quorum sensing inhibitors (QSIs) suppress conjugative plasmid transfer between bacteria by interfering with this process. For example, in wastewater, vanillin at a sub-minimum inhibitory concentration (sub-MIC, <0.1 g/L) reduced the conjugation transfer frequency of the resistance plasmid RP4 between E. coli and Pseudomonas aeruginosa by inhibiting biofilm formation, extracellular polymeric substance synthesis, and virulence factor secretion. This finding has important implications for controlling the environmental spread of bacterial resistance.224 Metabolic reprogramming strategies exploit nutrient competition to diminish the fitness advantage of resistant bacteria. Conjugation efficiency is influenced by the metabolic state of bacteria; high carbon concentration and an active physiological status can significantly enhance plasmid transfer kinetics. After transfer, energy demands related to plasmid recycling and host establishment—such as the metabolic burden associated with increased protein production—can limit the spread and trajectory of plasmids.225 However, in a ventricular assist device driveline infection model, Sun et al observed a strong negative correlation between bacterial metabolic activity and AMR when comparing different stages of biofilm development. The application of gentamicin—a weakly metabolism-dependent antibiotic—successfully killed staphylococcal biofilms, whereas vancomycin, which is strongly metabolism-dependent, did not. Thus, suppressed bacterial metabolism is a core mechanism of biofilm-associated AMR and a promising therapeutic target for biofilm-related driveline infections.226
Conclusion
Antibiotic resistance represents a critical global public health challenge, primarily driven by the dissemination of ARGs among bacteria via horizontal gene transfer mechanisms such as conjugation, transformation, and transduction. This review systematically elaborates on the key mechanisms of HGT and associated genetic elements, discusses environmental factors in different ecological niches that facilitate HGT, summarizes the applications of genomics and bioinformatics for monitoring HGT and predicting resistance phenotypes, and synthesizes the role of multi-omics strategies in restricting HGT. Conjugation and transduction are the most prominent pathways for HGT dissemination; among these, genomics-based targeting approaches—such as CRISPR-Cas systems—and phage therapy show promise as effective tools to suppress HGT. In the field of bacterial resistance prevention and control, the application of synthetic biology remains in its early stages. A future direction may involve the design of an engineered “nucleic acid sponge” probiotic capable of sequestering and degrading free plasmids and nucleic acid fragments in the environment, thereby reducing the incidence of HGT. Furthermore, artificial intelligence models powered by machine learning can be employed for early identification of hospitalized patients at high risk of resistant bacterial infections, monitoring the abundance dynamics of ARGs in the environment, and optimizing antibiotic use strategies in animal husbandry to prevent the spread of resistant bacteria. Despite these promising avenues, significant challenges remain in translating these strategies from bench to bedside. The precision of CRISPR-based editing is often compromised by off-target effects and the emergence of anti-CRISPR proteins; the narrow host range of phages limits their broad-spectrum applicability; and the high cost and lack of standardized protocols hinder the routine clinical implementation of metagenomic sequencing. Furthermore, interventions such as quorum sensing inhibitors and engineered probiotics have predominantly been validated in simplified systems, with their efficacy and safety in complex human microbiota or environmental ecosystems remaining largely unexplored. In conclusion, addressing antibiotic resistance, a pervasive global health threat, requires the integration of advanced computational models, mutation databases, and stringent regulatory frameworks. This interdisciplinary and international effort urgently demands close collaboration and concerted contributions from scientists worldwide.
Abbreviations
HGT, Horizontal Gene Transfer; ICEs, Integrative and Conjugative Elements; IMEs, Integrative and Mobilizable Elements; MDR, Multidrug-Resistant; CRAB, Carbapenem-resistant Acinetobacter baumannii; ICUs, Intensive Care Units; VRE, Vancomycin-resistant enterococci; AMR, Antimicrobial Resistance; GTAs, Gene Transfer Agents; OMVs, Outer Membrane Vesicles; ARGs, Antibiotic Resistance Genes; eDNA, Extracellular DNA; T6SS, Type VI Secretion System; dsDNA, Double-stranded DNA; ssDNA, Single-stranded DNA; DprA, DNA processing protein A; S. aureus, Staphylococcus aureus; B. subtilis, Bacillus subtilis; E. coli, Escherichia coli; cat, chloramphenicol acetyltransferase; erm, erythromycin resistance methylase; Inc group, Incompatibilitygroups; AMPs, Antimicrobial Peptides; IRs, Inverted Repeats; XDR, extensively drug-resistant; QS, Quorum Sensing; CPE, Carbapenemase-producing Enterobacteriaceae; VRE, Vancomycin-resistant Enterococcus; MGEs, Mobile Genetic Elements; CARD, Comprehensive Antibiotic Resistance Database; AMR, Antimicrobial Resistance; QSIs, Quorum Sensing Inhibitors; R-M system, Restriction-modification systems.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the National Key Clinical Specialties Construction Program.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hampton HG, Watson BNJ, Fineran PC. The arms race between bacteria and their phage foes. Nature. 2020;577(7790):327–19. doi:10.1038/s41586-019-1894-8
2. Han Z, Zhao L, Hu Q, et al. Gut microbiota-mediated modulation of host amino acid availability and metabolism. Gut Microbes. 2025;17(1):2552345. doi:10.1080/19490976.2025.2552345
3. Ikuta KS, Swetschinski LR, Robles Aguilar G. Global mortality associated with 33 bacterial pathogens in 2019: a systematic analysis for the Global Burden of Disease Study 2019. Lancet. 2022;400(10369):2221–2248. doi:10.1016/S0140-6736(22)02185-7
4. Nantongo M, Nguyen DC, Shin E, et al. Exploring β-lactam interactions with DacB1: unraveling optimal therapies for combating drug-resistant Mycobacterium tuberculosis. mBio. 2025;16(8):e0137225. doi:10.1128/mbio.01372-25
5. Al Amin M, Hossain MS, Habib MK, et al. Pharmacophore-guided computational modeling of quinolone-ATPase conjugate inhibitors targeting DNA GyrB subunit of Staphylococcus aureus. Comput Biol Med. 2025;186:109727. doi:10.1016/j.compbiomed.2025.109727
6. Kang MW, Ahn SY. Optimising personalised antibiotic treatment for methicillin-resistant Staphylococcus aureus bloodstream infections in ICU patients using a deep learning–based causal inference approach. J Glob Antimicrob Resist. 2025;45:70–76. doi:10.1016/j.jgar.2025.08.012
7. Muwonge A, Kakooza T, Johnson PCD, et al. Drivers of antimicrobial resistance in pig production systems of Uganda. Commun Earth Environ. 2025;6(1):517. doi:10.1038/s43247-025-02506-8
8. Schokker D, Stege PB, Duhamel M, et al. Rationally designed microbial communities in agri-food production systems: from research to market. ISME Commun. 2025;5(1):ycaf121. doi:10.1093/ismeco/ycaf121
9. Contarin R, Drapeau A, François P, et al. The interplay between mobilome and resistome in Staphylococcus aureus. mBio. 2024;15(10):e0242824. doi:10.1128/mbio.02428-24
10. Vesel N, Stare E, Štefanič P, et al. Naturally competent bacteria and their genetic parasites-a battle for control over horizontal gene transfer? FEMS Microbiol Rev. 2025;49.
11. Yan X, Liu W, Wen S, et al. Effect of sulfamethazine on the horizontal transfer of plasmid-mediated antibiotic resistance genes and its mechanism of action. J Environ Sci. 2023;127:399–409. doi:10.1016/j.jes.2022.06.014
12. Karakonstantis S, Kritsotakis EI, Tziolos RN, et al. Mortality due to carbapenem-resistant Acinetobacter baumannii bacteraemia: a 5-year cohort study in intensive care patients. Clin Microbiol Infect. 2025;31(6):1033–1039. doi:10.1016/j.cmi.2025.02.018
13. Ismail A, El-Hage-Sleiman AK, Majdalani M, et al. Device-associated infections in the pediatric intensive care unit at the American University of Beirut Medical Center. J Infect Dev Ctries. 2016;10(6):554–562. doi:10.3855/jidc.7303
14. Elshenawy RA, Umaru N, Aslanpour Z. An evaluation of the five rights antibiotic safety before and during COVID-19 at an NHS Foundation Trust in the United Kingdom. J Glob Antimicrob Resist. 2024;36:188–189. doi:10.1016/j.jgar.2023.12.019
15. Murray CJL, Ikuta KS, Sharara F. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629–655. doi:10.1016/S0140-6736(21)02724-0
16. Zhang X, Jiao P, Li B, et al. Phage lysis-mediated reduction of antibiotic-resistant bacteria alleviates micro/nanoplastic-driven antimicrobial resistance dissemination in anaerobic digestion. Water Res. 2025;285:124046. doi:10.1016/j.watres.2025.124046
17. Mom J, Valette O, Pieulle L, et al. Unraveling the molecular mechanisms of DNA capture by the Com pilus in naturally transformable monoderm bacteria. mBio. 2025;16(6):e0085125. doi:10.1128/mbio.00851-25
18. Xu Z, Li S, Ma Y, et al. Role of organophosphorus pesticides in facilitating plasmid-mediated conjugative transfer: efficiency and mechanisms. J Hazard Mater. 2025;487:137318. doi:10.1016/j.jhazmat.2025.137318
19. Kogay R, Zhaxybayeva O. Selection for translational efficiency in genes associated with alphaproteobacterial gene transfer agents. mSystems. 2022;7(6):e0089222. doi:10.1128/msystems.00892-22
20. Li P, Luo W, Xiang TX, et al. Horizontal gene transfer via OMVs co-carrying virulence and antimicrobial-resistant genes is a novel way for the dissemination of carbapenem-resistant hypervirulent Klebsiella pneumoniae. Front Microbiol. 2022;13:945972. doi:10.3389/fmicb.2022.945972
21. Sharma P, Kishore A, Singh M. Single-use polyethylene terephthalate bottle-derived nanoplastics propagate antibiotic resistance in bacteria via transformation and outer membrane vesicle secretion. Nanoscale. 2024;16(46):21360–21378. doi:10.1039/D4NR02613F
22. Li C, Zhu L, Wang D, et al. T6SS secretes an LPS-binding effector to recruit OMVs for exploitative competition and horizontal gene transfer. Isme J. 2022;16(2):500–510. doi:10.1038/s41396-021-01093-8
23. French KE, Zhou Z, Terry N. Horizontal ‘gene drives’ harness indigenous bacteria for bioremediation. Sci Rep. 2020;10(1):15091. doi:10.1038/s41598-020-72138-9
24. Morawska LP, Kuipers OP. Cell-to-cell non-conjugative plasmid transfer between Bacillus subtilis and lactic acid bacteria. Microb Biotechnol. 2023;16(4):784–798. doi:10.1111/1751-7915.14195
25. Silale A, Lea SM, Berks BC. The DNA transporter ComEC has metal-dependent nuclease activity that is important for natural transformation. Mol Microbiol. 2021;116(2):416–426. doi:10.1111/mmi.14720
26. Hua M, Dai D, Du P, et al. A chromosome-encoded T4SS independently contributes to horizontal gene transfer in Enterococcus faecalis. Cell Rep. 2022;41(6):111609. doi:10.1016/j.celrep.2022.111609
27. Furuya N, Komano T. NikAB- or NikB-dependent intracellular recombination between tandemly repeated oriT sequences of plasmid R64 in plasmid or single-stranded phage vectors. J Bacteriol. 2003;185(13):3871–3877. doi:10.1128/JB.185.13.3871-3877.2003
28. She T, Tan D, Balcazar JL, et al. Phage-mediated horizontal transfer of Salmonella enterica virulence genes with regulatory feedback from the host. Imeta. 2025;4(4):e70042. doi:10.1002/imt2.70042
29. Craske MW, Wilson JS, Fogg PCM. Gene transfer agents: structural and functional properties of domesticated viruses. Trends Microbiol. 2024;32(12):1200–1211. doi:10.1016/j.tim.2024.05.002
30. Johnston EL, Zavan L, Bitto NJ, et al. Planktonic and biofilm-derived Pseudomonas aeruginosa outer membrane vesicles facilitate horizontal gene transfer of plasmid DNA. Microbiol Spectr. 2023;11(2):e0517922. doi:10.1128/spectrum.05179-22
31. Velimirov B, Ranftler C. Unexpected aspects in the dynamics of horizontal gene transfer of prokaryotes: the impact of outer membrane vesicles. Wien Med Wochenschr. 2018;168(11–12):307–313. doi:10.1007/s10354-018-0642-2
32. Emamalipour M, Seidi K, Zununi Vahed S, et al. Horizontal gene transfer: from evolutionary flexibility to disease progression. Front Cell Dev Biol. 2020;8:229. doi:10.3389/fcell.2020.00229
33. Kumar G, Balakrishna K, Mukhopadhyay C, et al. Comparison of integron mediated antimicrobial resistance in clinical isolates of Escherichia coli from urinary and bacteremic sources. BMC Microbiol. 2024;24(1):102. doi:10.1186/s12866-024-03250-3
34. Wang YC, He LY, Wu HY, et al. High-risk plasmid-borne resistance genes from swine farm environments infiltrate deep soil and interact with the human gut microbiome via horizontal transfer. J Hazard Mater. 2025;496:139281. doi:10.1016/j.jhazmat.2025.139281
35. Domingues S, Lima T, Escobar C, et al. Large DNA fragment ISEc9-mediated transposition during natural transformation allows interspecies dissemination of antimicrobial resistance genes. Eur J Clin Microbiol Infect Dis. 2025;44(6):1417–1424. doi:10.1007/s10096-025-05113-9
36. Choufa C, Tidjani AR, Gauthier A, et al. Prevalence and mobility of integrative and conjugative elements within a Streptomyces natural population. Front Microbiol. 2022;13:970179. doi:10.3389/fmicb.2022.970179
37. Li M, Chen Q, Wu C, et al. A novel module promotes horizontal gene transfer in Azorhizobium caulinodans ORS571. Genes. 2022;13(10).
38. Botelho J, Cazares A, Schulenburg H. The ESKAPE mobilome contributes to the spread of antimicrobial resistance and CRISPR-mediated conflict between mobile genetic elements. Nucleic Acids Res. 2023;51(1):236–252. doi:10.1093/nar/gkac1220
39. Ku H, Kelk D, Bauer DC, et al. Phage-plasmid hybrids as vectors for antibiotic resistance in environmental Escherichia coli. Sci Total Environ. 2025;959:178157. doi:10.1016/j.scitotenv.2024.178157
40. Jonsdottir I, Given C, Penttinen R, et al. Preceding host history of conjugative resistance plasmids affects intra- and interspecific transfer potential from. Biofilm mSphere. 2023;8(3):e0010723. doi:10.1128/msphere.00107-23
41. Shaferman M, Gencel M, Alon N, et al. The fitness effects of codon composition of the horizontally transferred antibiotic resistance genes intensify at sub-lethal antibiotic levels. Mol Biol Evol. 2023;40(6). doi:10.1093/molbev/msad123
42. Pei Y, Hamar P, Pei DS. Deciphering multidrug-resistant Pseudomonas aeruginosa: mechanistic insights and environmental risks. Toxics. 2025;13(4):303. doi:10.3390/toxics13040303
43. Aggarwal R, Mahajan P, Pandiya S, et al. Antibiotic resistance: a global crisis, problems and solutions. Crit Rev Microbiol. 2024;50(5):896–921. doi:10.1080/1040841X.2024.2313024
44. Ding D, Wang B, Zhang X, et al. The spread of antibiotic resistance to humans and potential protection strategies. Ecotoxicol Environ Saf. 2023;254:114734. doi:10.1016/j.ecoenv.2023.114734
45. Blokesch M. Natural competence for transformation. Curr Biol. 2016;26(21):R1126–r1130. doi:10.1016/j.cub.2016.08.058
46. Carvalho G, Fouchet D, Danesh G, et al. Bacterial transformation buffers environmental fluctuations through the reversible integration of mobile genetic elements. mBio. 2020;11(2). doi:10.1128/mBio.02443-19
47. Crits-Christoph A, Hallowell HA, Koutouvalis K, et al. Good microbes, bad genes? The dissemination of antimicrobial resistance in the human microbiome. Gut Microbes. 2022;14(1):2055944. doi:10.1080/19490976.2022.2055944
48. Fukuda S, Tsuzuki K. Interspecific induction of transformation of bacteria with respect to phage-susceptibility. Nature. 1961;190:937–938. doi:10.1038/190937a0
49. Johnston C, Martin B, Fichant G, et al. Bacterial transformation: distribution, shared mechanisms and divergent control. Nat Rev Microbiol. 2014;12(3):181–196. doi:10.1038/nrmicro3199
50. Utnes AL, Sørum V, Hülter N, et al. Growth phase-specific evolutionary benefits of natural transformation in Acinetobacter baylyi. Isme J. 2015;9(10):2221–2231. doi:10.1038/ismej.2015.35
51. Borgeaud S, Metzger LC, Scrignari T, et al. The type VI secretion system of Vibrio cholerae fosters horizontal gene transfer. Science. 2015;347(6217):63–67. doi:10.1126/science.1260064
52. Coulthurst S. The Type VI secretion system: a versatile bacterial weapon. Microbiology. 2019;165(5):503–515. doi:10.1099/mic.0.000789
53. Peñil-Celis A, Garcillán-Barcia MP. Crosstalk between Type VI secretion system and mobile genetic elements. Front Mol Biosci. 2019;6:126. doi:10.3389/fmolb.2019.00126
54. Pimentel ZT, Zhang Y. Evolution of the natural transformation protein, ComEC, in bacteria. Front Microbiol. 2018;9:2980. doi:10.3389/fmicb.2018.02980
55. Facius D, Meyer TF. A novel determinant (comA) essential for natural transformation competence in Neisseria gonorrhoeae and the effect of a comA defect on pilin variation. Mol Microbiol. 1993;10(4):699–712. doi:10.1111/j.1365-2958.1993.tb00942.x
56. Graupner S, Wackernagel W. Identification and characterization of novel competence genes comA and exbB involved in natural genetic transformation of Pseudomonas stutzeri. Res Microbiol. 2001;152(5):451–460. doi:10.1016/S0923-2508(01)01218-9
57. Lerminiaux NA, Cameron ADS. Horizontal transfer of antibiotic resistance genes in clinical environments. Can J Microbiol. 2019;65(1):34–44. doi:10.1139/cjm-2018-0275
58. Kittredge HA, Dougherty KM, Evans SE. Dead but not forgotten: how extracellular DNA, moisture, and space modulate the horizontal transfer of extracellular antibiotic resistance genes in soil. Appl Environ Microbiol. 2022;88(7):e0228021. doi:10.1128/aem.02280-21
59. Sulek K. [Nobel prize for Joshua Lederberg in 1958 for discovery of genetic recombination and organization of the hereditary material in bacteria. Prize for George W. Beadle and Edward L. Tatum for discovery of the regulatory effect of genes on biochemical processes]. Wiad Lek. 1969;22(4):418–420.
60. Mergeay M, Gerits J. F’-plasmid transfer from Escherichia coli to Pseudomonas fluorescens. J Bacteriol. 1978;135(1):18–28. doi:10.1128/jb.135.1.18-28.1978
61. Lloyd RG, Buckman C. Conjugational recombination in Escherichia coli: genetic analysis of recombinant formation in Hfr x F- crosses. Genetics. 1995;139(3):1123–1148. doi:10.1093/genetics/139.3.1123
62. Lawley TD, Klimke WA, Gubbins MJ, et al. F factor conjugation is a true type IV secretion system. FEMS Microbiol Lett. 2003;224(1):1–15. doi:10.1016/S0378-1097(03)00430-0
63. Kennedy N, Beutin L, Achtman M, et al. Conjugation proteins encoded by the F sex factor. Nature. 1977;270(5638):580–585. doi:10.1038/270580a0
64. Wong JJ, Lu J, Glover JN. Relaxosome function and conjugation regulation in F-like plasmids - a structural biology perspective. Mol Microbiol. 2012;85(4):602–617. doi:10.1111/j.1365-2958.2012.08131.x
65. Mota-Bravo L, Camps M, Muñoz-Gutiérrez I, et al. Detection of horizontal gene transfer mediated by natural conjugative Plasmids in E. coli. J Vis Exp. 2023;193.
66. Yang X, Dong N, Chan EW, et al. Carbapenem resistance-encoding and virulence-encoding conjugative plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021;29(1):65–83. doi:10.1016/j.tim.2020.04.012
67. Zhang XW, Song JJ, Zeng SH, et al. Plasmid-mediated azithromycin resistance in non-typhoidal Salmonella recovered from human infections. J Antimicrob Chemother. 2024;79(10):2688–2697. doi:10.1093/jac/dkae281
68. Malaka De Silva P, Stenhouse GE, Blackwell GA, et al. A tale of two plasmids: contributions of plasmid associated phenotypes to epidemiological success among Shigella. Proc Biol Sci. 2022;289(1980):20220581. doi:10.1098/rspb.2022.0581
69. Wu W, Espedido B, Feng Y, et al. Citrobacter freundii carrying blaKPC-2 and blaNDM-1: characterization by whole genome sequencing. Sci Rep. 2016;6:30670. doi:10.1038/srep30670
70. Jiang X, Ellabaan MMH, Charusanti P, et al. Dissemination of antibiotic resistance genes from antibiotic producers to pathogens. Nat Commun. 2017;8:15784. doi:10.1038/ncomms15784
71. Fuentes S, Ding GC, Cárdenas F, et al. Assessing environmental drivers of microbial communities in estuarine soils of the Aconcagua River in Central Chile. FEMS Microbiol Ecol. 2015;91(10):fiv110. doi:10.1093/femsec/fiv110
72. Aquino de Muro M, Priest FG. Construction of chromosomal integrants of Bacillus sphaericus 2362 by conjugation with Escherichia coli. Res Microbiol. 2000;151(7):547–555. doi:10.1016/S0923-2508(00)00224-2
73. Leclerc QJ, Wildfire J, Gupta A, et al. Growth-dependent predation and generalized transduction of antimicrobial resistance by bacteriophage. mSystems. 2022;7(2):e0013522. doi:10.1128/msystems.00135-22
74. Chai Z, Zhang X, Dobbins AL, et al. Chimeric capsid proteins impact transduction efficiency of haploid adeno-associated virus vectors. Viruses. 2019;11(12):1138. doi:10.3390/v11121138
75. von Wintersdorff CJ, Penders J, van Niekerk JM, et al. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front Microbiol. 2016;7:173. doi:10.3389/fmicb.2016.00173
76. Watson BNJ, Staals RHJ, Fineran PC. CRISPR-Cas-mediated phage resistance enhances horizontal gene transfer by transduction. mBio. 2018;9(1). doi:10.1128/mBio.02406-17
77. Touchon M, Moura de Sousa JA, Rocha EP. Embracing the enemy: the diversification of microbial gene repertoires by phage-mediated horizontal gene transfer. Curr Opin Microbiol. 2017;38:66–73. doi:10.1016/j.mib.2017.04.010
78. Fillol-Salom A, Alsaadi A, Sousa JAM, et al. Bacteriophages benefit from generalized transduction. PLoS Pathog. 2019;15(7):e1007888. doi:10.1371/journal.ppat.1007888
79. Soler N, Forterre P. Vesiduction: the fourth way of HGT. Environ Microbiol. 2020;22(7):2457–2460. doi:10.1111/1462-2920.15056
80. Blesa A, Berenguer J. Methods to identify and analyze vesicle-protected DNA transfer. Methods Mol Biol. 2020;2075:209–221.
81. Esterman ES, Wolf YI, Kogay R, et al. Evolution of DNA packaging in gene transfer agents. Virus Evol. 2021;7(1):veab015. doi:10.1093/ve/veab015
82. Lang AS, Zhaxybayeva O, Beatty JT. Gene transfer agents: phage-like elements of genetic exchange. Nat Rev Microbiol. 2012;10(7):472–482. doi:10.1038/nrmicro2802
83. Lang AS, Westbye AB, Beatty JT. The distribution, evolution, and roles of gene transfer agents in prokaryotic genetic exchange. Annu Rev Virol. 2017;4(1):87–104. doi:10.1146/annurev-virology-101416-041624
84. Pallegar P, Peña-Castillo L, Langille E, et al. Cyclic di-GMP-mediated regulation of gene transfer and motility in Rhodobacter capsulatus. J Bacteriol. 2020;202(2). doi:10.1128/JB.00554-19
85. Shimizu T, Aritoshi T, Beatty JT, et al. Persulfide-responsive transcription factor SqrR regulates gene transfer and biofilm formation via the metabolic modulation of cyclic di-GMP in Rhodobacter capsulatus. Microorganisms. 2022;10(5).
86. Kogay R, Koppenhöfer S, Beatty JT, et al. Formal recognition and classification of gene transfer agents as viriforms. Virus Evol. 2022;8(2):veac100. doi:10.1093/ve/veac100
87. Sherlock D, Fogg PCM. Loss of the Rhodobacter capsulatus Serine Acetyl transferase gene, cysE1, impairs gene transfer by gene transfer agents and biofilm phenotypes. Appl Environ Microbiol. 2022;88(19):e0094422. doi:10.1128/aem.00944-22
88. Brimacombe CA, Ding H, Johnson JA, et al. Homologues of genetic transformation DNA import genes are required for Rhodobacter capsulatus gene transfer agent recipient capability regulated by the response regulator CtrA. J Bacteriol. 2015;197(16):2653–2663. doi:10.1128/JB.00332-15
89. Pérez J, Contreras-Moreno FJ, Marcos-Torres FJ, et al. The antibiotic crisis: how bacterial predators can help. Comput Struct Biotechnol J. 2020;18:2547–2555. doi:10.1016/j.csbj.2020.09.010
90. Moriyón I, Gamazo C, Díaz R. Properties of the outer membrane of Brucella. Ann Inst Pasteur Microbiol. 1987;138(1):89–91. doi:10.1016/0769-2609(87)90082-2
91. Briaud P, Carroll RK. Extracellular vesicle biogenesis and functions in gram-positive bacteria. Infect Immun. 2020;88(12). doi:10.1128/IAI.00433-20
92. Sartorio MG, Pardue EJ, Feldman MF, et al. Bacterial outer membrane vesicles: from discovery to applications. Annu Rev Microbiol. 2021;75:609–630. doi:10.1146/annurev-micro-052821-031444
93. Kulp A, Kuehn MJ. Biological functions and biogenesis of secreted bacterial outer membrane vesicles. Annu Rev Microbiol. 2010;64:163–184. doi:10.1146/annurev.micro.091208.073413
94. Ben-Hur S, Biton M, Regev-Rudzki N. Extracellular vesicles: a prevalent tool for microbial gene delivery? Proteomics. 2019;19(1–2):e1800170. doi:10.1002/pmic.201800170
95. Chatterjee S, Mondal A, Mitra S, et al. Acinetobacter baumannii transfers the blaNDM-1 gene via outer membrane vesicles. J Antimicrob Chemother. 2017;72(8):2201–2207. doi:10.1093/jac/dkx131
96. Marchant P, Carreño A, Vivanco E, et al. “One for all”: functional transfer of OMV-mediated polymyxin B resistance from Salmonella enterica sv. Typhi ΔtolR and ΔdegS to susceptible bacteria. Front Microbiol. 2021;12:672467. doi:10.3389/fmicb.2021.672467
97. Qiao W, Wang L, Luo Y, et al. Outer membrane vesicles mediated horizontal transfer of an aerobic denitrification gene between Escherichia coli. Biodegradation. 2021;32(4):435–448. doi:10.1007/s10532-021-09945-y
98. Dell’Annunziata F, Dell’Aversana C, Doti N, et al. Outer membrane vesicles derived from Klebsiella pneumoniae are a driving force for horizontal gene transfer. Int J Mol Sci. 2021;22(16). doi:10.3390/ijms22168732
99. Dubey GP, Ben-Yehuda S. Intercellular nanotubes mediate bacterial communication. Cell. 2011;144(4):590–600. doi:10.1016/j.cell.2011.01.015
100. Dubey GP, Malli Mohan GB, Dubrovsky A, et al. Architecture and characteristics of bacterial nanotubes. Dev Cell. 2016;36(4):453–461. doi:10.1016/j.devcel.2016.01.013
101. Gill S, Catchpole R, Forterre P. Extracellular membrane vesicles in the three domains of life and beyond. FEMS Microbiol Rev. 2019;43(3):273–303. doi:10.1093/femsre/fuy042
102. Boopathi S, Priya PS, Kesavan D, et al. Unveiling nanotubes-mediated communication: Enterococcus faecalis countering Salmonella ser. Typhi - in vitro and in vivo insights. Microb Pathog. 2023;184:106387. doi:10.1016/j.micpath.2023.106387
103. Fernández-Pato A, Sinha T, Garmaeva S, et al. Early-life development of the gut virome and plasmidome: a longitudinal study in cesarean-born infants. Cell Rep. 2025;44(6):115731. doi:10.1016/j.celrep.2025.115731
104. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/S1473-3099(15)00424-7
105. Liu JH, Liu YY, Shen YB, et al. Plasmid-mediated colistin-resistance genes: mcr. Trends Microbiol. 2024;32(4):365–378. doi:10.1016/j.tim.2023.10.006
106. Xavier BB, Lammens C, Ruhal R, et al. Identification of a novel plasmid-mediated colistin-resistance gene, mcr-2, in Escherichia coli, Belgium, June 2016. Euro Surveill. 2016;21(27). doi:10.2807/1560-7917.ES.2016.21.27.30280
107. Borowiak M, Fischer J, Hammerl JA, et al. Identification of a novel transposon-associated phosphoethanolamine transferase gene, mcr-5, conferring colistin resistance in d-tartrate fermenting Salmonella enterica subsp. enterica serovar Paratyphi B. J Antimicrob Chemother. 2017;72(12):3317–3324.
108. AbuOun M, Stubberfield EJ, Duggett NA, et al. mcr-1 and mcr-2 (mcr-6.1) variant genes identified in Moraxella species isolated from pigs in Great Britain from 2014 to 2015. J Antimicrob Chemother. 2018;73(10):2904. doi:10.1093/jac/dky272
109. Wang X, Wang Y, Zhou Y, et al. Emergence of a novel mobile colistin resistance gene, mcr-8, in NDM-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2018;7(1):122. doi:10.1038/s41426-018-0124-z
110. Wang C, Feng Y, Liu L, et al. Identification of novel mobile colistin resistance gene mcr-10. Emerg Microbes Infect. 2020;9(1):508–516. doi:10.1080/22221751.2020.1732231
111. Yang YQ, Li YX, Lei CW, et al. Novel plasmid-mediated colistin resistance gene mcr-7.1 in Klebsiella pneumoniae. J Antimicrob Chemother. 2018;73(7):1791–1795. doi:10.1093/jac/dky111
112. Yin W, Li H, Shen Y, et al. Novel plasmid-mediated colistin resistance gene mcr-3 in Escherichia coli. mBio. 2017;8(3).
113. Carattoli A, Villa L, Feudi C, et al. Novel plasmid-mediated colistin resistance mcr-4 gene in Salmonella and Escherichia coli, Italy 2013, Spain and Belgium, 2015 to 2016. Euro Surveill. 2017;22(31). doi:10.2807/1560-7917.ES.2017.22.31.30589
114. Carroll LM, Gaballa A, Guldimann C, et al. Identification of novel mobilized colistin resistance gene mcr-9 in a multidrug-resistant, colistin-susceptible Salmonella enterica serotype Typhimurium isolate. MBio. 2019;10(3):10–1128. doi:10.1128/mBio.00853-19
115. Liu Y, H LJ. Monitoring colistin resistance in food animals, an urgent threat. Expert Rev Anti Infect Ther. 2018;16(6):443–446. doi:10.1080/14787210.2018.1481749
116. Mmatli M, Mbelle NM, Osei Sekyere J. Global epidemiology, genetic environment, risk factors and therapeutic prospects of mcr genes: a current and emerging update. Front Cell Infect Microbiol. 2022;12:941358. doi:10.3389/fcimb.2022.941358
117. Xu Y, Lin J, Cui T, et al. Mechanistic insights into transferable polymyxin resistance among gut bacteria. J Biol Chem. 2018;293(12):4350–4365. doi:10.1074/jbc.RA117.000924
118. Dalmasso G, Beyrouthy R, Brugiroux S, et al. Genes mcr improve the intestinal fitness of pathogenic E. coli and balance their lifestyle to commensalism. Microbiome. 2023;11(1):12. doi:10.1186/s40168-022-01457-y
119. Santona A, Taviani E, Fiamma M, et al. Occult vancomycin-resistant Enterococcus faecium ST117 displaying a highly mutated vanB(2) Operon. Antibiotics. 2023;12(3):476. doi:10.3390/antibiotics12030476
120. Deng Y, Bao X, Ji L, et al. Resistance integrons: class 1, 2 and 3 integrons. Ann Clin Microbiol Antimicrob. 2015;14:45. doi:10.1186/s12941-015-0100-6
121. Kaushik M, Kumar S, Kapoor RK, et al. Integrons in Enterobacteriaceae: diversity, distribution and epidemiology. Int J Antimicrob Agents. 2018;51(2):167–176. doi:10.1016/j.ijantimicag.2017.10.004
122. Mondol SM, Islam MR, Mia ME, et al. Molecular and genomic insights into multidrug-resistant (MDR) and extensively drug-resistant (XDR) Pseudomonas aeruginosa causing burn wound infections in Bangladesh. Sci Rep. 2025;15(1):25445. doi:10.1038/s41598-025-11614-6
123. Biskri L, Mazel D. Erythromycin esterase gene ere(A) is located in a functional gene cassette in an unusual class 2 integron. Antimicrob Agents Chemother. 2003;47(10):3326–3331. doi:10.1128/AAC.47.10.3326-3331.2003
124. Ramírez MS, Quiroga C, Centrón D. Novel rearrangement of a class 2 integron in two non-epidemiologically related isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 2005;49(12):5179–5181. doi:10.1128/AAC.49.12.5179-5181.2005
125. Xu Z, Li L, Shirtliff ME, et al. Occurrence and characteristics of class 1 and 2 integrons in Pseudomonas aeruginosa isolates from patients in southern China. J Clin Microbiol. 2009;47(1):230–234. doi:10.1128/JCM.02027-08
126. Wang R, Li J, Kan B. Sequences of a co-existing SXT element, a chromosomal integron (CI) and an IncA/C plasmid and their roles in multidrug resistance in a Vibrio cholerae O1 El Tor strain. Int J Antimicrob Agents. 2016;48(3):305–309. doi:10.1016/j.ijantimicag.2016.05.020
127. Trudel MV, Vincent AT, Attéré SA, et al. Diversity of antibiotic-resistance genes in Canadian isolates of Aeromonas salmonicida subsp. salmonicida: dominance of pSN254b and discovery of pAsa8. Sci Rep. 2016;6:35617. doi:10.1038/srep35617
128. Zhang W, Peng K, Liu M, et al. Detection of SXT/R391 integrative conjugative elements carrying tigecycline resistance genes in Shewanella spp. isolated from retail seafood. Antimicrob Agents Chemother. 2025;69(8):e0174224. doi:10.1128/aac.01742-24
129. Zhu X, Wang X, Wang F, et al. The integrative and conjugative element ICECiPOL15 mediates horizontal transfer of β-lactam resistance gene in Chryseobacterium indoltheticum POL15. J Glob Antimicrob Resist. 2024;36:223–229. doi:10.1016/j.jgar.2023.12.028
130. López de Egea G, González-Díaz A, Guédon G, et al. A new integrative and mobilizable element is a major contributor to tetracycline resistance in Streptococcus dysgalactiae subsp. equisimilis. Antibiotics. 2023;12(3). doi:10.3390/antibiotics12030579
131. Chang TH, Manasherob R, Mrutyunjaya S, et al. High prophage count in Staphylococcus periprosthetic joint infection is associated with an increase in antibiotic resistance genes. J Arthroplasty. 2025;40:3254–3258. doi:10.1016/j.arth.2025.05.080
132. Gomathinayagam S, Kodiveri Muthukaliannan G. Dynamics of antibiotic resistance genes in plasmids and bacteriophages. Crit Rev Microbiol. 2025;51(2):219–228. doi:10.1080/1040841X.2024.2339262
133. Wiedenbeck J, Cohan FM. Origins of bacterial diversity through horizontal genetic transfer and adaptation to new ecological niches. FEMS Microbiol Rev. 2011;35(5):957–976. doi:10.1111/j.1574-6976.2011.00292.x
134. Medrano-Soto A, Moreno-Hagelsieb G, Vinuesa P, et al. Successful lateral transfer requires codon usage compatibility between foreign genes and recipient genomes. Mol Biol Evol. 2004;21(10):1884–1894. doi:10.1093/molbev/msh202
135. Li W, Bai X, Sheng H, et al. Conjugative transfer of mcr-1-bearing plasmid from Salmonella to Escherichia coli in vitro on chicken meat and in mouse gut. Food Res Int. 2022;157:111263. doi:10.1016/j.foodres.2022.111263
136. Zhu S, Hong J, Wang T. Horizontal gene transfer is predicted to overcome the diversity limit of competing microbial species. Nat Commun. 2024;15(1):800. doi:10.1038/s41467-024-45154-w
137. Babajanyan SG, Garushyants SK, Wolf YI, et al. Microbial diversity and ecological complexity emerging from environmental variation and horizontal gene transfer in a simple mathematical model. BMC Biol. 2024;22(1):148. doi:10.1186/s12915-024-01937-7
138. Herviou P, Balvay A, Bellet D, et al. Transfer of the integrative and conjugative element ICESt3 of Streptococcus thermophilus in physiological conditions mimicking the human digestive ecosystem. Microbiol Spectr. 2023;11(3):e0466722. doi:10.1128/spectrum.04667-22
139. Yin Z, Liang J, Zhang M, et al. Pan-genome insights into adaptive evolution of bacterial symbionts in mixed host-microbe symbioses represented by human gut microbiota Bacteroides cellulosilyticus. Sci Total Environ. 2024;927:172251. doi:10.1016/j.scitotenv.2024.172251
140. Aglomasa BC, Adu-Asiamah CK, Asiedu SO, et al. Multi-drug resistant bacteria isolates from lymphatic filariasis patients in the Ahanta West District, Ghana. BMC Microbiol. 2022;22(1):245. doi:10.1186/s12866-022-02624-9
141. ElRakaiby MT, Gamal-Eldin S, Amin MA, et al. Hospital microbiome variations as analyzed by high-throughput sequencing. Omics. 2019;23(9):426–438. doi:10.1089/omi.2019.0111
142. Skwor T, Stringer S, Haggerty J, et al. Prevalence of potentially pathogenic antibiotic-resistant Aeromonas spp. in treated urban wastewater effluents versus recipient riverine populations: a 3-year comparative study. Appl Environ Microbiol. 2020;86(3). doi:10.1128/AEM.02053-19
143. Shami AY, Abulfaraj AA, Refai MY, et al. Abundant antibiotic resistance genes in rhizobiome of the human edible Moringa oleifera medicinal plant. Front Microbiol. 2022;13:990169. doi:10.3389/fmicb.2022.990169
144. Zhu C, Wu L, Ning D, et al. Global diversity and distribution of antibiotic resistance genes in human wastewater treatment systems. Nat Commun. 2025;16(1):4006. doi:10.1038/s41467-025-59019-3
145. de Jong IG, Veening JW, Kuipers OP. Single cell analysis of gene expression patterns during carbon starvation in Bacillus subtilis reveals large phenotypic variation. Environ Microbiol. 2012;14(12):3110–3121. doi:10.1111/j.1462-2920.2012.02892.x
146. Meyer H, Weidmann H, Mäder U, et al. A time resolved metabolomics study: the influence of different carbon sources during growth and starvation of Bacillus subtilis. Mol Biosyst. 2014;10(7):1812–1823. doi:10.1039/C4MB00112E
147. Beaber JW, Hochhut B, Waldor MK. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature. 2004;427(6969):72–74. doi:10.1038/nature02241
148. Zhao R, Nawrocki A, Møller-Jensen J, et al. Mechanistic divergence between SOS response activation and antibiotic-induced plasmid conjugation in Escherichia coli. Microbiol Spectr. 2025;13(7):e0009025. doi:10.1128/spectrum.00090-25
149. Bower EKM, Cooper LP, Roberts GA, et al. A model for the evolution of prokaryotic DNA restriction-modification systems based upon the structural malleability of Type I restriction-modification enzymes. Nucleic Acids Res. 2018;46(17):9067–9080. doi:10.1093/nar/gky760
150. Skutel M, Yanovskaya D, Demkina A, et al. RecA-dependent or independent recombination of plasmid DNA generates a conflict with the host EcoKI immunity by launching restriction alleviation. Nucleic Acids Res. 2024;52(9):5195–5208. doi:10.1093/nar/gkae243
151. Chen W, Zhang H, Zhang Y, et al. Molecular basis for the PAM expansion and fidelity enhancement of an evolved Cas9 nuclease. PLoS Biol. 2019;17(10):e3000496. doi:10.1371/journal.pbio.3000496
152. de Curraize C, Neuwirth C, Bador J, et al. Two new Salmonella genomic islands 1 from Proteus mirabilis and description of blaCTX-M-15 on a variant (SGI1-K7). J Antimicrob Chemother. 2018;73(7):1804–1807. doi:10.1093/jac/dky108
153. Ding Y, Wang J, Chen Y, et al. Natural transformation of antibiotic resistance genes and the enhanced adaptability in bacterial biofilm under antibiotic and heavy metal stresses. J Hazard Mater. 2025;490:137740. doi:10.1016/j.jhazmat.2025.137740
154. Zhou H, Lu Z, Liu X, et al. Environmentally relevant concentrations of tetracycline promote horizontal transfer of antimicrobial resistance genes via plasmid-mediated conjugation. Foods. 2024;13(11).
155. Girijan SK, Paul R, Vj RK, et al. Investigating the impact of hospital antibiotic usage on aquatic environment and aquaculture systems: a molecular study of quinolone resistance in Escherichia coli. Sci Total Environ. 2020;748:141538. doi:10.1016/j.scitotenv.2020.141538
156. Li S, Duan G, Xi Y, et al. Insights into the role of extracellular polymeric substances (EPS) in the spread of antibiotic resistance genes. Environ Pollut. 2024;343:123285. doi:10.1016/j.envpol.2023.123285
157. Koraimann G, Wagner MA. Social behavior and decision making in bacterial conjugation. Front Cell Infect Microbiol. 2014;4:54. doi:10.3389/fcimb.2014.00054
158. Mukherjee S, Moustafa DA, Stergioula V, et al. The PqsE and RhlR proteins are an autoinducer synthase-receptor pair that control virulence and biofilm development in Pseudomonas aeruginosa. Proc Natl Acad Sci U S A. 2018;115(40):E9411–e9418. doi:10.1073/pnas.1814023115
159. Yang JX, Peng Y, Yu QY, et al. Gene horizontal transfers and functional diversity negatively correlated with bacterial taxonomic diversity along a nitrogen gradient. NPJ Biofilms Microbiomes. 2024;10(1):128. doi:10.1038/s41522-024-00588-4
160. Haenelt S, Richnow HH, Müller JA, et al. Antibiotic resistance indicator genes in biofilm and planktonic microbial communities after wastewater discharge. Front Microbiol. 2023;14:1252870. doi:10.3389/fmicb.2023.1252870
161. Jeric PE, Azpiroz A, Lopardo H, et al. Survey of molecular determinants in Gram-positive cocci isolated from hospital settings in Argentina. J Infect Dev Ctries. 2007;1(3):275–283. doi:10.3855/jidc.364
162. Qiu C, Zhou Y, Wang H, et al. Microplastics enrichment characteristics of antibiotic resistance genes and pathogens in landfill leachate. Chemosphere. 2023;341:140100. doi:10.1016/j.chemosphere.2023.140100
163. Ye G, Li M, Huang H, et al. Polystyrene microplastic exposure induces selective accumulation of antibiotic resistance genes in gut microbiota and its potential health risks. Int J Biol Macromol. 2025;309(Pt 3):142983. doi:10.1016/j.ijbiomac.2025.142983
164. Hongjin C, Ur Rahman S, Rehman A, et al. Microplastics and antibiotic resistance genes as rising threats: their interaction represents an urgent environmental concern. Curr Res Microb Sci. 2025;9:100447. doi:10.1016/j.crmicr.2025.100447
165. Ni N, Qiu J, Ge W, et al. Fibrous and fragmented microplastics discharged from sewage amplify health risks associated with antibiotic resistance genes in aquatic environments. Environ Sci Technol. 2025;59(30):15919–15930. doi:10.1021/acs.est.5c01335
166. Zhang D, Sun J, Peng S, et al. Biodegradable microplastics exacerbate the risk of antibiotic resistance genes pollution in agricultural soils. J Hazard Mater. 2025;496:139490. doi:10.1016/j.jhazmat.2025.139490
167. Zhang Q, Fan Y, Qian X, et al. Unraveling the role of microplastics in antibiotic resistance: insights from long-read metagenomics on ARG mobility and host dynamics. J Hazard Mater. 2025;490:137804. doi:10.1016/j.jhazmat.2025.137804
168. Luo Y, Xu T, Li B, et al. The effects of small plastic particles on antibiotic resistance gene transfer revealed by single cell and community level analysis. J Hazard Mater. 2024;480:136271. doi:10.1016/j.jhazmat.2024.136271
169. Miao S, Zhang Y, Wu L, et al. Resistance induction potency assessment of antibiotic production wastewater and associated resistome shaping mechanisms. Water Res. 2025;283:123811. doi:10.1016/j.watres.2025.123811
170. Nguyen AQ, Vu HP, Nguyen LN, et al. Monitoring antibiotic resistance genes in wastewater treatment: current strategies and future challenges. Sci Total Environ. 2021;783:146964. doi:10.1016/j.scitotenv.2021.146964
171. Mitra S, Naha S, Chakraborty J, et al. Diversity of mobile genetic elements in carbapenem-resistant Enterobacterales isolated from the intensive care units of a tertiary care hospital in Northeast India. Front Microbiol. 2025;16:1543427. doi:10.3389/fmicb.2025.1543427
172. Li B, Li X, Wang B, et al. A metagenomic approach for characterizing antibiotic resistance genes in specific bacterial populations: demonstration with Escherichia coli in cattle manure. Appl Environ Microbiol. 2022;88(7):e0255421. doi:10.1128/aem.02554-21
173. McDonagh F, Tumeo A, Murray EK, et al. Complete genome of Escherichia coli sequence type 73 with acquired blaTEM-1 and high genotypic virulence load identified in human saliva. J Glob Antimicrob Resist. 2024;39:149–152. doi:10.1016/j.jgar.2024.08.011
174. Liu S, Dai S, Deng Y, et al. Long-read epicPCR enhances species-level host identification of clinically relevant antibiotic resistance genes in environmental microbial communities. Environ Int. 2025;197:109337. doi:10.1016/j.envint.2025.109337
175. Kim JH, Seong JE, Lee YS, et al. Origin and transmission of carbapenemase-producing Enterobacterales and vancomycin-resistant Enterococci in hospitalized patients: a genomic and epidemiological analysis. J Hosp Infect. 2025;165:100–108. doi:10.1016/j.jhin.2025.07.017
176. Alcock BP, Huynh W, Chalil R, et al. CARD 2023: expanded curation, support for machine learning, and resistome prediction at the comprehensive antibiotic resistance database. Nucleic Acids Res. 2023;51(D1):D690–d699. doi:10.1093/nar/gkac920
177. Jeong H, Sung S, Kwon T, et al. HGTree: database of horizontally transferred genes determined by tree reconciliation. Nucleic Acids Res. 2016;44(D1):D610–9. doi:10.1093/nar/gkv1245
178. Choi Y, Ahn S, Park M, et al. HGTree v2.0: a comprehensive database update for horizontal gene transfer (HGT) events detected by the tree-reconciliation method. Nucleic Acids Res. 2023;51(D1):D1010–d1018. doi:10.1093/nar/gkac929
179. Krawczyk PS, Lipinski L, Dziembowski A. PlasFlow: predicting plasmid sequences in metagenomic data using genome signatures. Nucleic Acids Res. 2018;46(6):e35. doi:10.1093/nar/gkx1321
180. Che Y, Yang Y, Xu X, et al. Conjugative plasmids interact with insertion sequences to shape the horizontal transfer of antimicrobial resistance genes. Proc Natl Acad Sci U S A. 2021;118(6). doi:10.1073/pnas.2008731118
181. Song W, Sun HX, Zhang C, et al. Prophage Hunter: an integrative hunting tool for active prophages. Nucleic Acids Res. 2019;47(W1):W74–w80. doi:10.1093/nar/gkz380
182. Florensa AF, Kaas RS, Clausen P, et al. ResFinder - an open online resource for identification of antimicrobial resistance genes in next-generation sequencing data and prediction of phenotypes from genotypes. Microb Genom. 2022;8(1).
183. Zhou H, Beltrán JF, Brito IL. Functions predict horizontal gene transfer and the emergence of antibiotic resistance. Sci Adv. 2021;7(43):eabj5056. doi:10.1126/sciadv.abj5056
184. Li G, Long TF, Zhou SY, et al. CRISPR-AMRtracker: a novel toolkit to monitor the antimicrobial resistance gene transfer in fecal microbiota. Drug Resist Updat. 2024;77:101142. doi:10.1016/j.drup.2024.101142
185. Liu B, Pop M. ARDB--Antibiotic Resistance Genes Database. Nucleic Acids Res. 2009;37(Database issue):D443–7. doi:10.1093/nar/gkn656
186. Alcock BP, Raphenya AR, Lau TTY, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517–d525. doi:10.1093/nar/gkz935
187. Gibson MK, Forsberg KJ, Dantas G. Improved annotation of antibiotic resistance determinants reveals microbial resistomes cluster by ecology. Isme J. 2015;9(1):207–216. doi:10.1038/ismej.2014.106
188. Wallace JC, Port JA, Smith MN, et al. FARME DB: a functional antibiotic resistance element database. Database. 2017;2017:baw165. doi:10.1093/database/baw165
189. Scaria J, Chandramouli U, Verma SK. Antibiotic Resistance Genes Online (ARGO): a database on vancomycin and beta-lactam resistance genes. Bioinformation. 2005;1(1):5–7. doi:10.6026/97320630001005
190. Gupta SK, Padmanabhan BR, Diene SM, et al. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob Agents Chemother. 2014;58(1):212–220. doi:10.1128/AAC.01310-13
191. Arango-Argoty G, Garner E, Pruden A, et al. DeepARG: a deep learning approach for predicting antibiotic resistance genes from metagenomic data. Microbiome. 2018;6(1):23. doi:10.1186/s40168-018-0401-z
192. Yin X, Jiang XT, Chai B, et al. ARGs-OAP v2.0 with an expanded SARG database and Hidden Markov Models for enhancement characterization and quantification of antibiotic resistance genes in environmental metagenomes. Bioinformatics. 2018;34(13):2263–2270. doi:10.1093/bioinformatics/bty053
193. Zankari E, Hasman H, Cosentino S, et al. Identification of acquired antimicrobial resistance genes. J Antimicrob Chemother. 2012;67(11):2640–2644. doi:10.1093/jac/dks261
194. Doster E, Lakin SM, Dean CJ, et al. MEGARes 2.0: a database for classification of antimicrobial drug, biocide and metal resistance determinants in metagenomic sequence data. Nucleic Acids Res. 2020;48(D1):D561–d569. doi:10.1093/nar/gkz1010
195. Sandgren A, Strong M, Muthukrishnan P, et al. Tuberculosis drug resistance mutation database. PLoS Med. 2009;6(2):e2. doi:10.1371/journal.pmed.1000002
196. Ghosh A, N S, Saha S. Survey of drug resistance associated gene mutations in Mycobacterium tuberculosis, ESKAPE and other bacterial species. Sci Rep. 2020;10(1):8957. doi:10.1038/s41598-020-65766-8
197. Davis JJ, Wattam AR, Aziz RK, et al. The PATRIC Bioinformatics Resource Center: expanding data and analysis capabilities. Nucleic Acids Res. 2020;48(D1):D606–d612. doi:10.1093/nar/gkz943
198. Skoulakis A, Kostoula V, Daniilidis K, et al. AmrProfiler: a comprehensive tool for identifying antimicrobial resistance genes and mutations across species. Nucleic Acids Res. 2025;53(W1):W20–w31. doi:10.1093/nar/gkaf400
199. Ahmed S, Emon MI, Moumi NA, et al. ProtAlign-ARG: antibiotic resistance gene characterization integrating protein language models and alignment-based scoring. Sci Rep. 2025;15(1):30174. doi:10.1038/s41598-025-14545-4
200. Sherry NL, Horan KA, Ballard SA, et al. An ISO-certified genomics workflow for identification and surveillance of antimicrobial resistance. Nat Commun. 2023;14(1):60. doi:10.1038/s41467-022-35713-4
201. Dhariwal A, Junges R, Chen T, et al. ResistoXplorer: a web-based tool for visual, statistical and exploratory data analysis of resistome data. NAR Genom Bioinform. 2021;3(1):lqab018. doi:10.1093/nargab/lqab018
202. Lam MMC, Wick RR, Watts SC, et al. A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat Commun. 2021;12(1):4188. doi:10.1038/s41467-021-24448-3
203. De R, Mukhopadhyay AK, Dutta S. Metagenomic analysis of gut microbiome and resistome of diarrheal fecal samples from Kolkata, India, reveals the core and variable microbiota including signatures of microbial dark matter. Gut Pathog. 2020;12:32. doi:10.1186/s13099-020-00371-8
204. Cruz H, Pinheiro M, Borges V. ReporType: a flexible bioinformatics tool for targeted Loci screening and typing of infectious agents. Int J Mol Sci. 2024;25(6):3172. doi:10.3390/ijms25063172
205. Li X, Xie Y, Liu M, et al. oriTfinder: a web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements. Nucleic Acids Res. 2018;46(W1):W229–w234. doi:10.1093/nar/gky352
206. Leggett RM, Alcon-Giner C, Heavens D, et al. Rapid MinION profiling of preterm microbiota and antimicrobial-resistant pathogens. Nat Microbiol. 2020;5(3):430–442. doi:10.1038/s41564-019-0626-z
207. Muzondiwa D, Mutshembele A, Pierneef RE, et al. Resistance sniffer: an online tool for prediction of drug resistance patterns of Mycobacterium tuberculosis isolates using next generation sequencing data. Int J Med Microbiol. 2020;310(2):151399. doi:10.1016/j.ijmm.2020.151399
208. O’Meara D, Nunney L. A phylogenetic test of the role of CRISPR-Cas in limiting plasmid acquisition and prophage integration in bacteria. Plasmid. 2019;104:102418. doi:10.1016/j.plasmid.2019.102418
209. Tao S, Chen H, Li N, et al. Association of CRISPR-Cas system with the antibiotic resistance and virulence genes in nosocomial isolates of Enterococcus. Infect Drug Resist. 2022;15:6939–6949. doi:10.2147/IDR.S388354
210. Tao S, Chen H, Li N, et al. Elimination of bla(KPC-2)-mediated carbapenem resistance in Escherichia coli by CRISPR-Cas9 system. BMC Microbiol. 2023;23(1):310. doi:10.1186/s12866-023-03058-7
211. Nozawa T, Furukawa N, Aikawa C, et al. CRISPR inhibition of prophage acquisition in Streptococcus pyogenes. PLoS One. 2011;6(5):e19543. doi:10.1371/journal.pone.0019543
212. Uddin MJ, Kim B, Dawan J, et al. Assessment of antibiotic resistance in bacteriophage-insensitive Klebsiella pneumoniae. Microb Pathog. 2019;135:103625. doi:10.1016/j.micpath.2019.103625
213. Vázquez R, Seoane-Blanco M, Rivero-Buceta V, et al. Monomodular Pseudomonas aeruginosa phage JG004 lysozyme (Pae87) contains a bacterial surface-active antimicrobial peptide-like region and a possible substrate-binding subdomain. Acta Crystallogr D Struct Biol. 2022;78(Pt 4):435–454. doi:10.1107/S2059798322000936
214. Wang C, Wang S, Jing S, et al. Data-driven engineering of phages with tunable capsule tropism for Klebsiella pneumoniae. Adv Sci. 2024;11(33):e2309972. doi:10.1002/advs.202309972
215. Peng H, Chen IA, Qimron U. Engineering phages to fight multidrug-resistant bacteria. Chem Rev. 2025;125(2):933–971. doi:10.1021/acs.chemrev.4c00681
216. Zhu J, Liu Y, Chen T, et al. “Precision-guided killer” engineered phage for combating carbapenem-resistant klebsiella pneumoniae induced inflammatory bowel disease. Adv Funct Mater. 2025;35:e00754. doi:10.1002/adfm.202500754
217. Kim P, Sanchez AM, Penke TJR, et al. Safety, pharmacokinetics, and pharmacodynamics of LBP-EC01, a CRISPR-Cas3-enhanced bacteriophage cocktail, in uncomplicated urinary tract infections due to Escherichia coli (ELIMINATE): the randomised, open-label, first part of a two-part Phase 2 trial. Lancet Infect Dis. 2024;24(12):1319–1332. doi:10.1016/S1473-3099(24)00424-9
218. Huo W, Adams HM, Trejo C, et al. A Type I restriction-modification system associated with Enterococcus faecium subspecies separation. Appl Environ Microbiol. 2019;85(2). doi:10.1128/AEM.02174-18
219. Shmidov E, Villani A, Mendoza SD, et al. Multigenerational proteolytic inactivation of restriction upon subtle genomic hypomethylation in Pseudomonas aeruginosa. Nat Microbiol. 2025;10:2498–2510. doi:10.1038/s41564-025-02088-3
220. Qiu Y, Guo P, Tian H, et al. The restriction impacts of the Type III restriction-modification system on the transmission dynamics of antimicrobial resistance genes in Campylobacter jejuni. Front Microbiol. 2025;16:1496275. doi:10.3389/fmicb.2025.1496275
221. Park S, Jung D, O’Brien B, et al. Comparative genomic analysis of Staphylococcus aureus isolates associated with either bovine intramammary infections or human infections demonstrates the importance of restriction-modification systems in host adaptation. Microb Genom. 2022;8(2). doi:10.1099/mgen.0.000779
222. Xu C, Rao J, Xie Y, et al. The DNA phosphorothioation restriction-modification system influences the antimicrobial resistance of pathogenic bacteria. Microbiol Spectr. 2023;11(1):e0350922. doi:10.1128/spectrum.03509-22
223. Djordjevic M, Zivkovic L, Ou HY, et al. Nonlinear regulatory dynamics of bacterial restriction-modification systems modulates horizontal gene transfer susceptibility. Nucleic Acids Res. 2025;53(2). doi:10.1093/nar/gkae1322
224. Qiu X, Wang B, Ren S, et al. Regulation of quorum sensing for the manipulation of conjugative transfer of antibiotic resistance genes in wastewater treatment system. Water Res. 2024;253:121222. doi:10.1016/j.watres.2024.121222
225. Ahmad M, Aduru SV, Smith RP, et al. The role of bacterial metabolism in antimicrobial resistance. Nat Rev Microbiol. 2025;23(7):439–454. doi:10.1038/s41579-025-01155-0
226. Sun Y, McGiffin D, Ye L, et al. Low bacterial metabolism as a central mechanism of antimicrobial resistance and potential therapeutic target of staphylococcal biofilms in ventricular assist device driveline infections. J Antimicrob Chemother. 2025;80:2923–2933. doi:10.1093/jac/dkaf316
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.