Back to Journals » Cancer Management and Research » Volume 11
Suppressed OGT expression inhibits cell proliferation and modulates EGFR expression in renal cell carcinoma
Authors Wang L, Chen S, Zhang J, Mao S, Mao W, Zhang W, Guo Y, Wu Y, Wang R, Yan Y, Yao X ![]()
Received 12 October 2018
Accepted for publication 8 February 2019
Published 19 March 2019 Volume 2019:11 Pages 2215—2223
DOI https://doi.org/10.2147/CMAR.S190642
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rituraj Purohit
Longsheng Wang,1 Shaojun Chen,2 Junfeng Zhang,1 Shiyu Mao,1 Weipu Mao,1 Wentao Zhang,1 Yadong Guo,1 Yuan Wu,1 Ruiliang Wang,1 Yang Yan,1 Xudong Yao1
1Department of Urology, Shanghai Tenth People’s Hospital, Tongji University, School of Medicine, Shanghai 200072, China; 2Department of Urology, Shanghai General Hospital, Shanghai Jiaotong University, School of Medicine, Shanghai 200080, China
Purpose: O-linked N-acetylglucosamine (O-GlcNAc or O-GlcNAcylation) is a post-translational modification, which plays a vital role in the progression of various cancers. The purpose of the present study was to assess O-GlcNAcylation in human renal cell carcinoma (RCC).
Methods: O-GlcNAcylation levels and O-GlcNAc-transferase (OGT) expression in human RCC cell lines and 10 paired clinical tissues were detected by Western blot and Immunohistochemistry. Then, the effects of O-GlcNAcylation on RCC cell proliferation in vitro were investigated by Cell Counting Kit-8 assay. A xenograft assay was performed to assess the in vivo effects of OGT knockdown in RCC cells. Cell apoptosis and cell cycle assays were performed by flow cytometry. Co-immunoprecipitation assays were used to assess epidermal growth factor receptor (EGFR) O-GlcNAcylation and the interaction between OGT and EGFR.
Results: O-GlcNAcylation levels and OGT expression were increased in RCC, and the high amounts correlated with poor patient prognosis. OGT knockdown significantly suppressed RCC cell proliferation in vitro and in vivo. Notably, EGFR was modulated by O-GlcNAcylation and directly interacted with OGT.
Conclusion: These findings provide novel insights into the oncogenic roles of O-GlcNAcylation and OGT in the development of RCC, indicating that OGT might be used as a target for RCC therapy in the future.
Keywords: renal cancer, O-GlcNAcylation, OGT, proliferation, EGFR
Introduction
In 2018, it was expected that about 63,340 patients would be newly diagnosed with renal cell carcinoma (RCC) in the US, which is among the most lethal human genitourinary cancers.1,2 However, about 30% of RCC cases have metastases at initial diagnosis.3 Due to its resistance to chemotherapy and radiotherapy, targeted therapy remains the first-line treatment option for these advanced RCC patients.4 Most RCC cases show a notable clinical response; however, the treatment effects of targeted agents are limited due to the development of drug-resistant phenotypes.5 Hence, it is necessary to further assess the mechanisms involved in RCC and improve diagnosis and therapy strategies for this malignancy.
O-linked N-acetylglucosamine (O-GlcNAc or O-GlcNAcylation) is a post-translational modification, which is considered a new cancer hallmark based on multiple studies in the past decade.6 O-GlcNAc synthesis is catalyzed by O-GlcNAc-transferase (OGT) while the group is removed by O-GlcNAcase (OGA).7 As the substrate of O-GlcNAc, UDP-GlcNAc is synthesized from glycolytic metabolites through the hexosamine biosynthetic pathway (HBP) and add GlcNAc to serine or threonine residues of target proteins.8–10 Numerous biological processes are influenced by O-GlcNAcylation, such as cell cycle progression, signal transduction, and metabolism.11 Being dynamic and reversible, aberrant O-GlcNAc modulation is involved in the formation and progression of many diseases, especially carcinogenesis.8 The biological effects of O-GlcNAc in cancer development are mostly via O-GlcNAcylation of proteins such as p53 and β-catenin.12,13 For example, the oncogene c-Myc is O-GlcNAcylated, which could inhibit its phosphorylation and consequently suppress proteasome mediated degradation.14 Increased levels of O-GlcNAcylation have been reported in various cancers, including prostate cancer,15 colon cancer,16 liver carcinoma,17 breast cancer,14 and leukemia.18,19 However, the potential roles of O-GlcNAc in renal cancer remain undefined.
The number of proteins containing the O-GlcNAc modification has steadily increased over the past years. As a receptor tyrosine kinase (RTK), the epidermal growth factor receptor (EGFR) is always overexpressed in various cancers thus contributing to carcinogenesis by improving the invasive potential and metastatic behavior.20 EGFR is indeed overexpressed in RCC and is associated with cell cycle and tumorigenesis.21–24 Overexpression of EGFR in RCC could also lead to the upregulation of VEGF, which is involved in the angiogenic phenotype.25 Therefore, upregulated EGFR is considered an available molecular target for RCC therapy.26–28
The present study aimed to assess the expression and function of OGT as well as O-GlcNAcylation levels in RCC. Moreover, the potential mechanisms were explored.
Materials and methods
Antibodies and reagents
O-GlcNAc specific antibodies (RL2) were obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Antibodies against OGT and other primary antibodies were purchased from Abcam (Cambridge, MA, USA).
Cell culture and lentiviral shRNA infection
The human normal renal tubular epithelial HK-2 and RCC 786-O, A498, ACHN, and CAKI-2 cell lines were purchased from the Cell Bank of Type Culture Collection of Chinese Academy of Sciences (CCCAS, Shanghai, China). 786-O cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Thermo Fisher Scientific, Waltham, MA, USA). A498, ACHN, and CAKI-2 cells were cultured in RMPI 1640 (Thermo Fisher Scientific), while HK-2 cells were cultured in Keratinocyte Medium (KM, ScienCell, Carlsbad, CA, USA). All cell culture media were supplemented with 10% Fetal Bovine Serum (FBS, Hyclone, Logan, UT, USA), and cells were cultured at 37°C with 5% CO2.
The lentivirus expressing shRNA against OGT was produced by Jiman Co. (Shanghai, China). 786-O and A498 cells were infected with LV-sh-OGT or LV-sh-NC and selected by puromycin (Sigma–Aldrich, St. Louis, MO, USA). The expression levels of OGT and O-GlcNAc were examined at the RNA and protein levels.
Immunohistochemical staining
All tissues were formalin-fixed and embedded in paraffin blocks. Then, 4 µm sections were dewaxed in xylene and rehydrated in graded ethanol. Antigen retrieval was performed in pre-heated Tris-EDTA buffer for 20 minutes. Methanol containing 0.3% H2O2 was used to block endogenous peroxidase activity for 8 minutes. Next, bovine serum albumin (BSA) was used to block the sections for 30 minutes before incubation with the primary antibody overnight at 4°C. The sections were then incubated with horseradish peroxidase (HRP)-conjugated rabbit-anti-mouse IgG for 30 minutes. This was followed by treatment with the DAB+ EnVision System (Thermo Fisher Scientific) and counterstaining with Mayer/hematoxylin.
Cell proliferation in vitro
The shRNA sequence of OGT is shown in Table 1. The proliferation ability of renal cancer cells was evaluated by Cell Counting Kit-8 (CCK8) assay (Sigma-Aldrich) according to the manufacturer’s instructions. Briefly, 786-O and A498 cells were transfected with sh-NC or sh-OGT for 48 hours and seeded into 96-well plates at a density of 5×103 per well. After incubation for different times (0 hour, 24 hours, 48 hours, 72 hours, and 96 hours), 10 µL CCK-8 reagent was added to each well and further incubated for 4 hours. Finally, cell proliferation was determined by measuring absorbance at 450 nm on a plate reader.
 | Table 1 The shRNA sequence of OGT |
Xenograft assays in nude mice
Stably transfected 786-O (LV-sh-OGT and LV-sh-NC) cells (3×106 in 0.2 mL PBS) were implanted subcutaneously into the dorsal flank of male BALB/c nude mice (6 weeks old). The tumorigenic potential of cells was evaluated 3 weeks after inoculation. The mice were sacrificed by pentobarbital overdose (1%), and tumors were excised and weighed. Animal care and experiments were carried out under the National Institutes of Health (NIH) Guide for Care and Use of Laboratory Animals. All animal studies were approved by the Ethics Committees of Shanghai Tenth People’s Hospital.
Protein extraction and western blot
Total protein was extracted with RIPA lysis buffer containing protease and phosphatase inhibitors. The Bio-Rad protein assay system was used to measure total protein levels according to the manufacturer’s instructions. Equal amounts of protein were separated by SDS-PAGE and transferred onto nitrocellulose (NC) membranes (Bio-Rad). The blots were probed with primary antibodies overnight and incubated with secondary antibodies for 1 hour at room temperature, before signal visualization.
Co-immunoprecipitation
About 2 mg of cell lysate in 1 mL RIPA buffer was incubated with 2 µg of anti-EGFR antibody with shaking at 4°C overnight. Samples were then mixed with 30 µL of protein A/G-agarose (Santa Cruz) and incubated with shaking at 4°C for 2 hours. Agarose beads were washed five times in RIPA buffer. Immunoprecipitates were then eluted in 2 X SDS sample loading buffer and separated by SDS-PAGE.
Cell cycle distribution analysis
Cells were harvested and fixed with 70% ice-cold ethanol overnight. Then, cells were centrifuged and resuspended in PBS containing propidium iodide (PI; BD Biosciences, Franklin Lakes, NJ, USA), RNase (100 µg/mL) and Triton X-100 (0.2%) for 30 minutes. Finally, cell cycle distribution was analyzed by flow cytometry.
Statistical analysis
SPSS version 23.0 (IBM Corporation, Armonk, NY, USA) was used for statistical analysis. Data are mean ± SD from three independent experiments. The Wilcoxon signed-rank test, Student’s t-test, or chi-squared test was used to assess group differences. P<0.05 was considered statistically significant.
Ethics statement
The study was approved by the Ethics Committees of Shanghai Tenth People’s Hospital and written informed consent was obtained from all participating individuals in the study.
Results
Hyper-O-GlcNAcylation and OGT overexpression occur in renal cell carcinoma
First, we examined O-GlcNAc modification levels and OGT expression in human RCC (786-O, ACHN, CAKI-1, and A498) cell lines compared with the human normal renal tubular epithelial HK-2 cell line. Total O-GlcNAcylation and the expression of OGT was increased in RCC cells (Figure 1A). In addition, OGT expression was elevated in RCC tissues (Figure 1B). We also assessed the protein levels of OGT in RCC and adjacent normal tissues by immunohistochemistry (IHC). OGT signals were more intense in tumor tissues compared with adjacent normal samples (Figure 1C). The Oncomine database (https://www.oncomine.org/) was selected to analyze the levels of OGT in RCC and normal renal tissues. The results indicated that OGT mRNA expression levels were obviously elevated in RCC tissues compared with normal ones (Figure 1D).
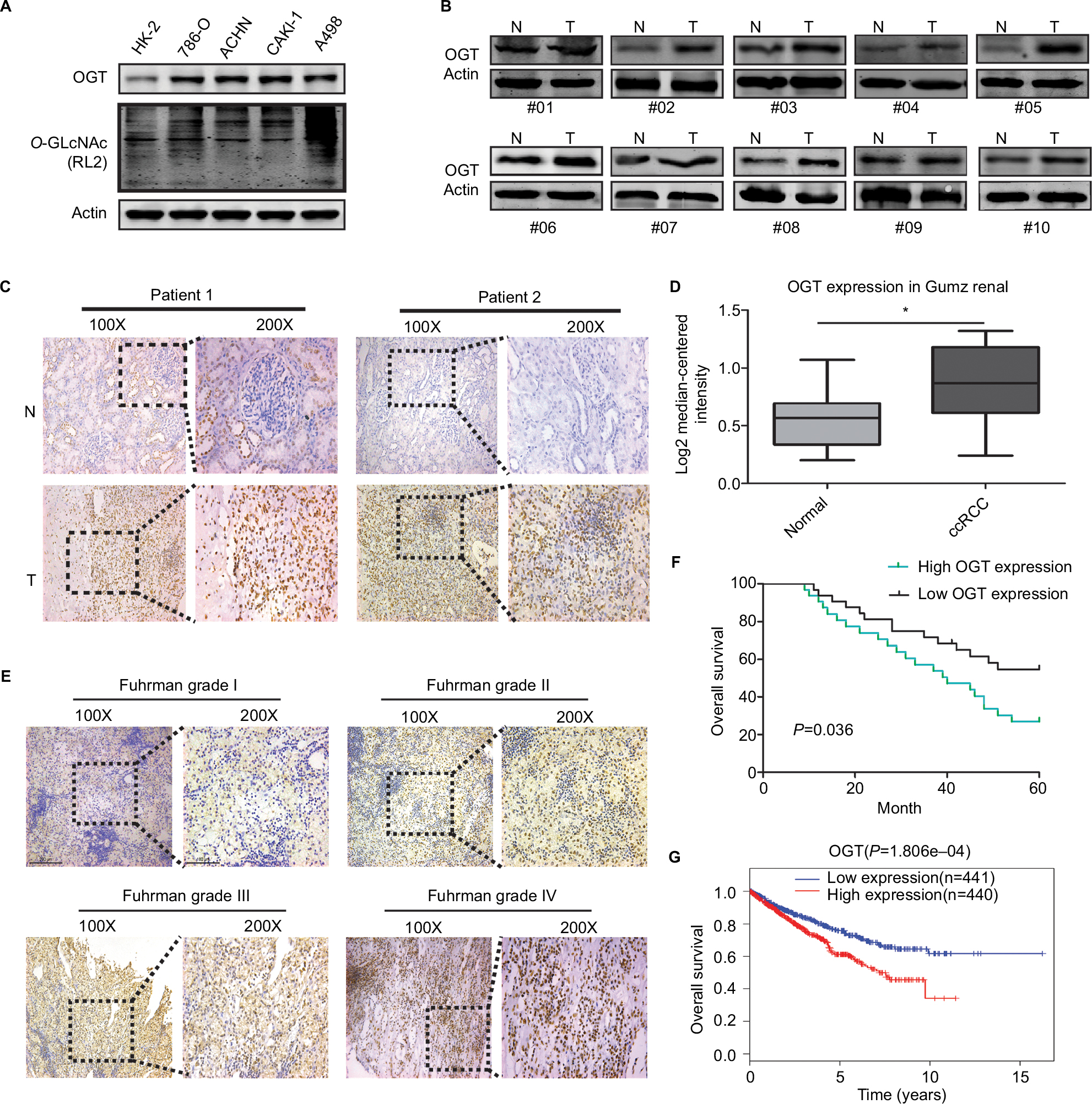 | Figure 1 Hyper-O-GlcNAcylation and OGT Overexpression Occur in renal cell carcinoma Notes: (A) Expression levels of O-GlcNAcylation and OGT in RCC (786-O, ACHN, CAKI-1, and A498) cell lines compared with the human normal renal tubular epithelial HK-2 cell line. (B) Expression levels of OGT in RCC and adjacent normal tissues were detected by Western blot. (C) Representative images of immunohistochemical staining for OGT expression in RCC and adjacent normal tissues. (D) Oncomine expression analysis of OGT in human RCC and normal renal tissues. (E) Comparison of OGT expression in Fuhrman grades I–IV of RCC tissue samples by IHC staining (left panel, magnification ×100; right panel, magnification ×200). (F) Kaplan–Meier analysis of the correlation between OGT expression and overall survival in RCC patients. (G) Overall survival in RCC patients with high or low expression of OGT from TCGA datasets. Abbreviations: ccRCC, clear cell RCC; O-GlcNAc , O-linked N-acetylglucosamine; OGT, O-GlcNAc-transferase; RCC, renal cell carcinoma; TCGA, The Cancer Genome Atlas. |
In addition, IHC data showed that OGT staining in RCC was positively correlated with the Fuhrman grade (Figure 1E). Next, we explored whether OGT expression was correlated with patient prognosis in RCC. Kaplan–Meier analysis showed that RCC patients with higher OGT expression levels had relatively poorer prognosis compared with those showing lower OGT expression (Figure 1F). Similarly, the TCGA datasets also supported the notion that high OGT levels predicted shorter survival time in RCC (Figure 1G).
Taken together, these findings suggested that hyper-O-GlcNAcylation and OGT overexpression occurred in renal cell carcinoma; moreover, OGT expression could serve as a prognostic marker of RCC progression.
Reduction of OGT level inhibits the growth of RCC in vitro and in vivo
O-GLcNAcylation plays an important role in the proliferation of various cancers. Meanwhile, suppressing OGT expression inhibits tumor formation and growth.6,29,30 Therefore, we assessed whether hyper-O-GlcNAcylation and OGT overexpression are associated with RCC cell proliferation. Stable OGT knockdown in 786-O and A498 cells was successfully performed (Figure 2A). First, we demonstrated that OGT knockdown effectively attenuated the proliferation of RCC cells as determined by CCK8 assay (Figure 2B, C).
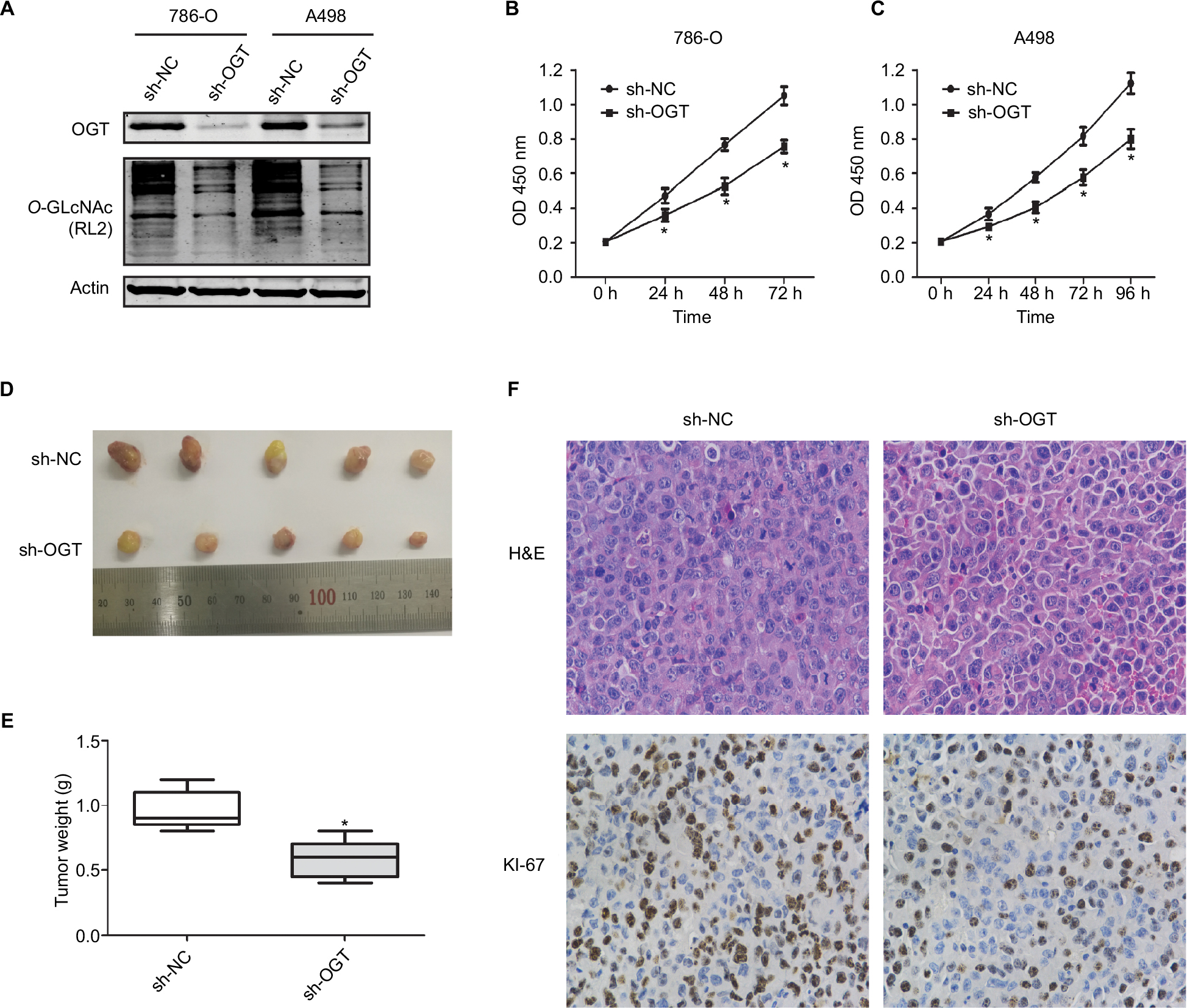 | Figure 2 OGT suppression inhibits RCC growth in vitro and in vivo. Notes: (A) Expression levels of OGT and O-GlcNAcylation after OGT knockdown in 786-O and A498 cells. (B, C) CCK8 assay was used to assess the proliferation of 786-O and A498 cells after OGT knockdown. (D) Representative images and animal weights. (E) Weights of the excised tumors derived from nude mice are shown. (F) Representative images of H&E staining and Ki-67 immunohistochemical detection in the excised tumors derived from nude mice. Data are mean ± SD from three independent experiments. *P<0.05. Magnification ×200. Abbreviations: CCK8, Cell Counting Kit-8; O-GlcNAc, O-linked N-acetylglucosamine; OGT, O-GlcNAc-transferase; NC, negative control; RCC, renal cell carcinoma. |
We next examined whether OGT suppression in RCC cells inhibits the tumorigenic phenotype in vivo. Stably transfected 786-O (LV-sh-OGT and LV-sh-NC) cells were injected into male BALB/c nude mice. A total of 3 weeks after implantation, the tumor-bearing mice were sacrificed, and tumors were harvested and weighted. Then, immunohistological examination of Ki-67 expression was performed. Significantly reduced tumor size and weight were observed in OGT knockdown 786-O cells (Figure 2D, E). As shown in Figure 2F, less Ki-67-positive cells were found in tumors from the OGT knockdown group compared with control samples, indicating a tumor-suppressing potential.
OGT suppression promotes RCC cell apoptosis and inhibits cell cycle progression
The effects of O-GLcNACylation on RCC cell apoptosis and cell cycle distribution were examined by flow cytometry. The results showed increased apoptosis of 786-O and A498 cells after OGT downregulation (Figure 3A–C). Moreover, cell cycle analysis indicated that the G0/G1 cell population was increased while the S and G2/M phases were decreased following OGT knockdown (Figure 3D, F and E, G). These data indicated that OGT downregulation inhibited RCC cell proliferation probably via apoptosis promotion and cell cycle inhibition.
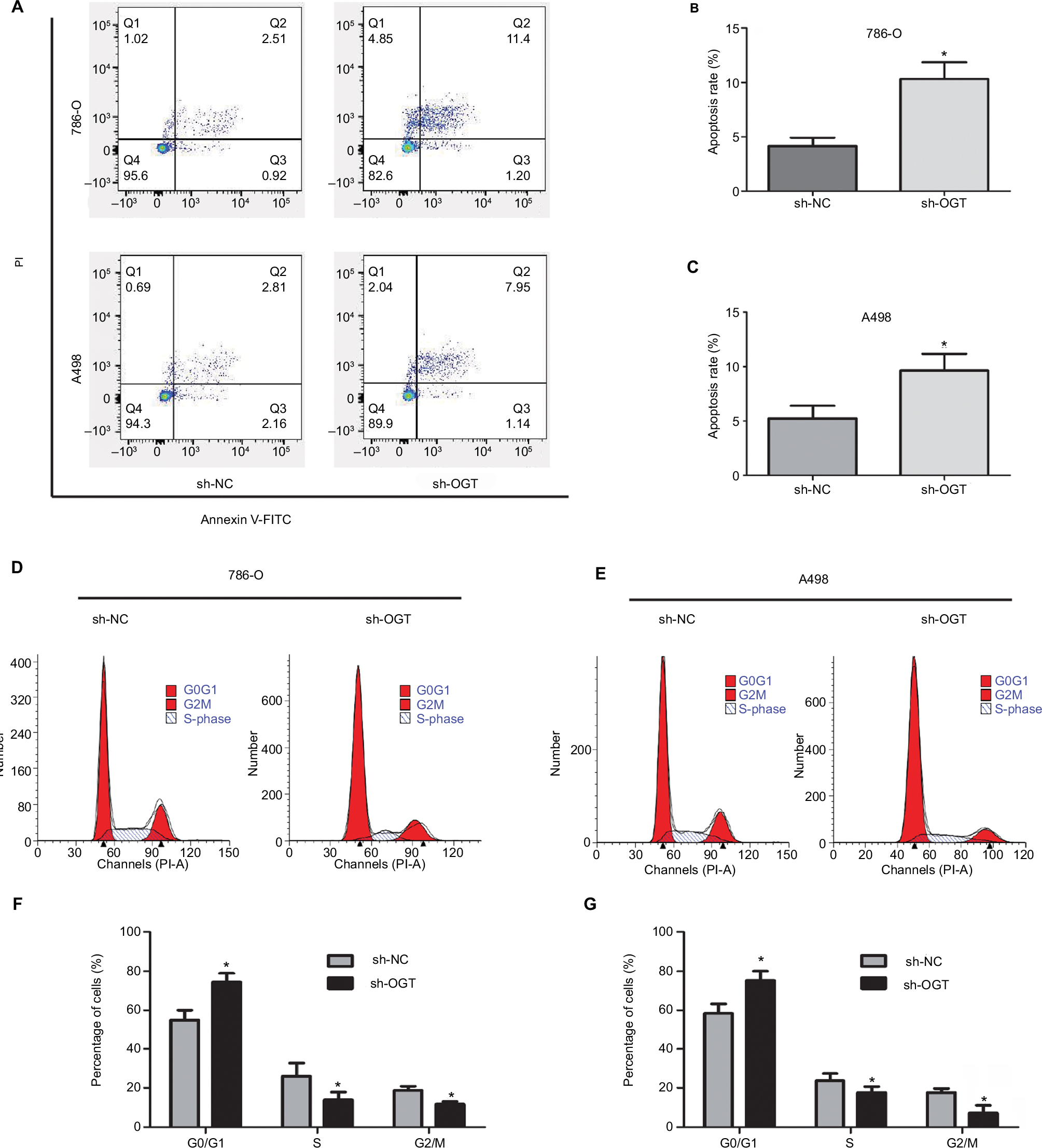 | Figure 3 OGT suppression promotes RCC cell apoptosis and inhibits the cell cycle. Notes: (A–C) Apoptotic rates in 786-O and A498 cells as measured by flow cytometry following OGT knockdown. (D–G) Effects of OGT knockdown on cell cycle distribution in T24 and UMUC-3 cells, as determined by flow cytometry. Data are mean ± SD from three independent experiments. *P<0.05. Abbreviations: FITC, Fluorescein isothiocyanate; OGT, O-GlcNAc-transferase;NC, negative control; PI, propidium iodide; RCC, renal cell carcinoma. |
OGT suppression reduces EGFR expression
It has been reported that OGT interacts with EGFR, which can be O-GlcNAcylated.31 Meanwhile, EGFR is over-expressed in RCC and shows correlation with poor prognosis in clear cell RCC.32,33 Thus, we first assessed whether EGFR is O-GlcNAcylated in RCC. Figure 4A reveals the correlation between OGT expression and EGFR in TCGA datasets. In this study, EGFR and its downstream PI3K/AKT pathway was downregulated in OGT knockdown 786-O cells (Figure 4B). We next found that EGFR and OGT could co-immunoprecipitate in 786-O cells (Figure 4C). Moreover, EGFR was O-GlcNAcylated in 786-O cells (Figure 4D). These results suggested EGFR O-GlcNAcylation in RCC.
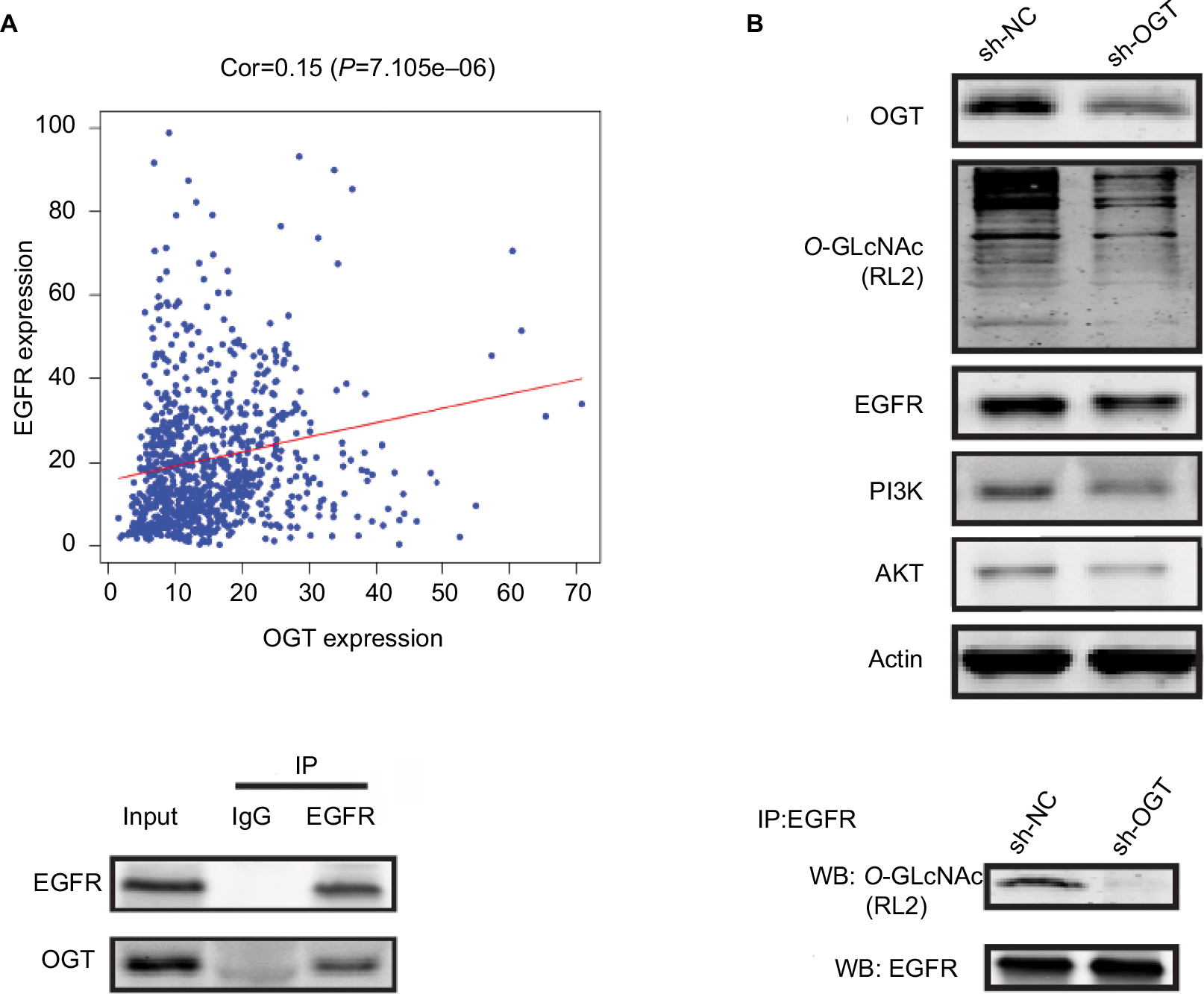 | Figure 4 OGT suppression downregulates EGFR. Notes: (A) Correlation between OGT expression and EGFR in TCGA datasets. (B) Effect of OGT knockdown on EGFR and PI3K/AKT expression in RCC cells. (C) Interaction between OGT and EGFR in 786-O cells, as determined by co-immunoprecipitation and immunoblot. (D) 786-O cells were treated with sh-NC or sh-OGT followed by EGFR immunoprecipitation with anti-EGFR antibodies; the resulting EGFR immunoprecipitates were assessed by WB for EGFR and O-GlcNAc (RL2) detection. Abbreviations: Cor, correlation; EGFR, epidermal growth factor receptor; IP, immunoprecipitation; OGT, O-GlcNAc-transferase; O-GlcNAc , O-linked N-acetylglucosamine; NC, negative control; RCC, renal cell carcinoma TCGA, The Cancer Genome Atlas; WB, Western blot. |
Discussion
In recent years, the role of O-GlcNAcylation in tumorigenesis has attracted increasing attention, with multiple related publications in various human malignancies such as prostate cancer,15 colon cancer,16 liver carcinoma,17 breast cancer,14 and leukemia.18,19 This study for the first time demonstrated that O-GlcNAcylation levels and OGT expression were markedly higher in RCC cell lines and tissue samples compared with respective normal controls. Meanwhile, high OGT expression was associated with the Fuhrman grade and poor patient prognosis in RCC. Functional assays revealed that silencing of OGT effectively inhibited RCC cell proliferation and cell cycle transition accompanied by increased apoptosis. These results suggest that hyper-O-GlcNAcylation and OGT upregulation function as an oncogenic factor in the development of RCC.
Excess food intake is considered an important cancer risk factor, and obesity was reported to be a risk factor for many cancer types, including breast, colon, liver, and endometrial cancers.34,35 A fraction (2%–3%) of glucose entering the cell is applied to the hexosamine biosynthetic pathway (HBP), which controls O-GlcNAcylation of cellular proteins.6 Similar to phosphorylation, O-GlcNAcylation is a reversible post-translational modification that influences almost all cellular processes.8,36 O-GlcNAcylation is mainly upregulated in cancer, and such upregulation is positively correlated with tumor grade or metastasis. However, discrepancies concerning the levels of O-GlcNAc and its cycling enzymes have emerged.14,16,17,37–39
RCC is considered one of the most vascularized human cancers;40 therefore, antiangiogenic therapies are well characterized in RCC.41,42 Upregulated EGFR has been reported to modulate cell functions in RCC, including cell cycle, cell migration, cell invasion and vascularization.21–25,43 In addition, previous study has demonstrated that EGFR is O-GLcNAcylated and directly interacts with OGT in A431 cells.31 The PI3K/AKT pathway was reported to be associated with RCC progression and was a promising drug target.44,45 In the present study, we for the first time reported that EGFR undergoes such post-translational modification and can be co-immunoprecipitated with OGT in human RCC 786-O cells. We also demonstrated that EGFR and its downstream PI3K/AKT pathway was downregulated in OGT knockdown 786-O cells.
Limitation
The present study did not assess whether EGFR is directly involved in OGT-associated effects on RCC cell functions, which deserves further investigation.
Conclusion
In summary, we for the first time reported that O-GlcNAcylation and OGT expression are upregulated and correlated with poor patient prognosis in RCC. In addition, OGT knockdown attenuates RCC cell proliferation in vitro and in vivo. We further found that EGFR is O-GlcNAcylated and interacts with OGT. Taken together, these findings provide novel insights into the oncogenic roles of O-GlcNAcylation and OGT in the development of RCC, and therefore OGT might be used as a target for RCC therapy in the future.
Acknowledgment
This study was supported by National Science Foundation of China (No. 81472389).
Disclosure
The authors report no conflicts of interest in this work.
References
Pruthi DK, Oomah S, Lu V, et al. Quality and quantity in kidney cancer surgery: the role of nonneoplastic kidney and kidney volumetrics in predicting postoperative renal function. Am J Clin Pathol. 2019;151(1):108–115. | ||
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2017. CA Cancer J Clin. 2017;67(1):7–30. | ||
Cindolo L, Patard J-J, Chiodini P, et al. Comparison of predictive accuracy of four prognostic models for nonmetastatic renal cell carcinoma after nephrectomy: a multicenter European study. Cancer. 2005;104(7):1362–1371. | ||
Calvo E, Schmidinger M, Heng DYC, Grünwald V, Escudier B. Improvement in survival end points of patients with metastatic renal cell carcinoma through sequential targeted therapy. Cancer Treat Rev. 2016;50:109–117. | ||
Heng DY, Mackenzie MJ, Vaishampayan UN, et al. Primary anti-vascular endothelial growth factor (VEGF)-refractory metastatic renal cell carcinoma: clinical characteristics, risk factors, and subsequent therapy. Ann Oncol. 2012;23(6):1549–1555. | ||
Fardini Y, Dehennaut V, Lefebvre T, Issad T. O-GlcNAcylation: a new cancer hallmark? Front Endocrinol. 2013;4:99. | ||
Trinca GM, Goodman ML, Papachristou EK, et al. O-GlcNAc-Dependent regulation of progesterone receptor function in breast cancer. Horm Canc. 2018;9(1):12–21. | ||
Ozcan S, Andrali SS, Cantrell JEL. Modulation of transcription factor function by O-GlcNAc modification. Biochim Biophys Acta. 2010;1799(5-6):353–364. | ||
Ma Z, Vosseller K. Cancer metabolism and elevated O-GlcNAc in oncogenic signaling. J Biol Chem. 2014;289(50):34457–34465. | ||
Ma Z, Vocadlo DJ, Vosseller K. Hyper-O-GlcNAcylation is anti-apoptotic and maintains constitutive NF-κB activity in pancreatic cancer cells. J Biol Chem. 2013;288(21):15121–15130. | ||
Hart GW, Housley MP, Slawson C. Cycling of O-linked beta-N-acetylglucosamine on nucleocytoplasmic proteins. Nature. 2007;446(7139):1017–1022. | ||
Yang WH, Kim JE, Nam HW, et al. Modification of p53 with O-linked N-acetylglucosamine regulates p53 activity and stability. Nat Cell Biol. 2006;8(10):1074–1083. | ||
Yang WH, Park SY, Nam HW, et al. NFkappaB activation is associated with its O-GlcNAcylation state under hyperglycemic conditions. Proc Natl Acad Sci U S A. 2008;105(45):17345–17350. | ||
Gu Y, Mi W, Ge Y, et al. GlcNAcylation plays an essential role in breast cancer metastasis. Cancer Res. 2010;70(15):6344–6351. | ||
Lynch TP, Ferrer CM, Jackson SR, Shahriari KS, Vosseller K, Reginato MJ. Critical role of O-Linked β-N-acetylglucosamine transferase in prostate cancer invasion, angiogenesis, and metastasis. J Biol Chem. 2012;287(14):11070–11081. | ||
Mi W, Gu Y, Han C, et al. O-GlcNAcylation is a novel regulator of lung and colon cancer malignancy. Biochim Biophys Acta. 2011;1812(4):514–519. | ||
Zhu Q, Zhou L, Yang Z, et al. O-GlcNAcylation plays a role in tumor recurrence of hepatocellular carcinoma following liver transplantation. Med Oncol. 2012;29(2):985–993. | ||
Ding X, Jiang W, Zhou P, et al. Mixed lineage leukemia 5 (MLL5) protein stability is cooperatively regulated by O-GlcNAc transferase (OGT) and ubiquitin specific protease 7 (USP7). PLoS One. 2015;10(12): e0145023. | ||
Shi Y, Tomic J, Wen F, et al. Aberrant O-GlcNAcylation characterizes chronic lymphocytic leukemia. Leukemia. 2010;24(9):1588–1598. | ||
Mendelsohn J. Antibody-mediated EGF receptor blockade as an anticancer therapy: from the laboratory to the clinic. Cancer Immunol Immunother. 2003;52(5):342–346. | ||
Uhlman DL, Nguyen P, Manivel JC, et al. Epidermal growth factor receptor and transforming growth factor alpha expression in papillary and nonpapillary renal cell carcinoma: correlation with metastatic behavior and prognosis. Clin Cancer Res. 1995;1(8):913–920. | ||
Hofmockel G, Riess S, Bassukas ID, Dämmrich J. Epidermal growth factor family and renal cell carcinoma: expression and prognostic impact. Eur Urol. 1997;31(4):478–484. | ||
Moch H, Sauter G, Buchholz N, et al. Epidermal growth factor receptor expression is associated with rapid tumor cell proliferation in renal cell carcinoma. Hum Pathol. 1997;28(11):1255–1259. | ||
Kallio JP, Hirvikoski P, Helin H, et al. Membranous location of EGFR immunostaining is associated with good prognosis in renal cell carcinoma. Br J Cancer. 2003;89(7):1266–1269. | ||
Le Q-T, Denko NC, Giaccia AJ. Hypoxic gene expression and metastasis. Cancer Metastasis Rev. 2004;23(3-4):293–310. | ||
Yang JC. Bevacizumab for patients with metastatic renal cancer: an update. Clin Cancer Res. 2004;10(18 Pt 2):6367S–6370S. | ||
Motzer RJ, Michaelson MD, Redman BG, et al. Activity of SU11248, a multitargeted inhibitor of vascular endothelial growth factor receptor and platelet-derived growth factor receptor, in patients with metastatic renal cell carcinoma. J Clin Oncol. 2006;24(1):16–24. | ||
Ratain MJ, Eisen T, Stadler WM, et al. Phase II placebo-controlled randomized discontinuation trial of sorafenib in patients with metastatic renal cell carcinoma. J Clin Oncol. 2006;24(16):2505–2512. | ||
Itkonen HM, Minner S, Guldvik IJ, et al. O-GlcNAc transferase integrates metabolic pathways to regulate the stability of c-myc in human prostate cancer cells. Cancer Res. 2013;73(16):5277–5287. | ||
Itkonen HM, Gorad SS, Duveau DY, et al. Inhibition of O-GlcNAc transferase activity reprograms prostate cancer cell metabolism. Oncotarget. 2016;7(11):12464–12476. | ||
Stateva SR, Villalobo A. O-GlcNAcylation of the human epidermal growth factor receptor. Org Biomol Chem. 2015;13(30):8196–8204. | ||
Dorđević G, Matušan Ilijaš K, Hadžisejdić I, Maričić A, Grahovac B, Jonjić N. EGFR protein overexpression correlates with chromosome 7 polysomy and poor prognostic parameters in clear cell renal cell carcinoma. J Biomed Sci. 2012;19(1):40. | ||
Merseburger AS, Hennenlotter J, Simon P, et al. Membranous expression and prognostic implications of epidermal growth factor receptor protein in human renal cell cancer. Anticancer Res. 2005;25(3B):1901–1907. | ||
Dossus L, Kaaks R. Nutrition, metabolic factors and cancer risk. Best Pract Res Clin Endocrinol Metab. 2008;22(4):551–571. | ||
Roberts DL, Dive C, Renehan AG. Biological mechanisms linking obesity and cancer risk: new perspectives. Annu Rev Med. 2010;61(1):301–316. | ||
Hart GW, Copeland RJ. Glycomics hits the big time. Cell. 2010;143(5):672–676. | ||
Slawson C, Pidala J, Potter R. Increased N-acetyl-beta-glucosaminidase activity in primary breast carcinomas corresponds to a decrease in N-acetylglucosamine containing proteins. Biochim Biophys Acta. 2001;1537(2):147–157. | ||
Krześlak A, Forma E, Bernaciak M, Romanowicz H, Bryś M. Gene expression of O-GlcNAc cycling enzymes in human breast cancers. Clin Exp Med. 2012;12(1):61–65. | ||
Krześlak A, Wójcik-Krowiranda K, Forma E, Bieńkiewicz A, Bryś M. Expression of genes encoding for enzymes associated with O-GlcNAcylation in endometrial carcinomas: clinicopathologic correlations. Ginekol Pol. 2012;83(1):22–26. | ||
Lôrincz T, Tímár J, Szendrôi M. Alterations of microvascular density in bone metastases of adenocarcinomas. Pathol Oncol Res. 2004;10(3):149–153. | ||
Pantuck AJ, Zeng G, Belldegrun AS, Figlin RA. Pathobiology, prognosis, and targeted therapy for renal cell carcinoma: exploiting the hypoxia-induced pathway. Clin Cancer Res. 2003;9(13):4641–4652. | ||
Rini BI, Small EJ. Biology and clinical development of vascular endothelial growth factor-targeted therapy in renal cell carcinoma. J Clin Oncol. 2005;23(5):1028–1043. | ||
Lu Z, Jiang G, Blume-Jensen P, Hunter T. Epidermal growth factor-induced tumor cell invasion and metastasis initiated by dephosphorylation and downregulation of focal adhesion kinase. Mol Cell Biol. 2001;21(12):4016–4031. | ||
Di JH, Gao KY, Qu DB, Yang J, Zheng JN. Rap2B promotes angiogenesis via PI3K/AKT/VEGF signaling pathway in human renal cell carcinoma. Tumor Biol. 2017;39(7):1010428317701653. | ||
Liu F, Shangli Z, Hu Z. CAV2 promotes the growth of renal cell carcinoma through the EGFR/PI3K/Akt pathway. Oncotargets Ther. 2018;11:6209–6216. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.