Back to Journals » Neuropsychiatric Disease and Treatment » Volume 22
Stem Cells from Human Exfoliated Deciduous Teeth Attenuate Depression-Like Behaviors in CUMS Rats by Suppressing Neuroinflammation and Enhancing Synaptic Plasticity
Authors Yu F, Gao Y, Du X, Liu S ![]() , Xu Y
, Xu Y ![]()
Received 4 December 2025
Accepted for publication 16 February 2026
Published 24 February 2026 Volume 2026:22 586668
DOI https://doi.org/10.2147/NDT.S586668
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Fei Yu,1,2 Yao Gao,1 Xinzhe Du,1 Sha Liu,1 Yong Xu3
1Department of Psychiatry, First Hospital/First Clinical Medical College of Shanxi Medical University, Taiyuan, People’s Republic of China; 2Department of Psychiatry, Heping Hospital Affiliated to Changzhi Medical College, Changzhi, People’s Republic of China; 3Department of Clinical Psychology, The Eighth Affiliated Hospital of Sunyat-Sen University, Shenzhen, People’s Republic of China
Correspondence: Yong Xu; Sha Liu, Department of Psychiatry, First Hospital/First Clinical Medical College of Shanxi Medical University, Taiyuan, SX030001, People’s Republic of China, Email [email protected]; [email protected]
Purpose: This study aimed to evaluate the therapeutic potential of stem cells from human exfoliated deciduous teeth (SHED) against depression and to elucidate the underlying mechanisms involving neuroinflammation and synaptic plasticity in a rat model of chronic unpredictable mild stress (CUMS).
Methods: A robust rat model of depression was established using chronic unpredictable mild stress (CUMS) paradigm. CUMS-exposed rats received intracerebroventricular transplantation of SHED at three doses (0.5× 106, 1× 106, or 2× 106 cells/rat); a fluoxetine group served as positive control. Depressive-like behaviors were assessed through sucrose preference, forced swimming, open field, and Y-maze tests. Post-behavioral molecular analyses in the hippocampus and prefrontal cortex included qRT-PCR and ELISA for key inflammatory cytokines (IL-1β, IL-6, TNF-α, IL-10, iNOS, Arg1, NLRP3, Caspase-1) and neuroplasticity markers (BDNF, TrkB, PSD95). Microglial activation and polarization were examined by immunofluorescence. Hippocampal transcriptomic profiling was conducted via RNA sequencing.
Results: SHED transplantation rapidly ameliorated CUMS-induced behavioral deficits, showing efficacy comparable to fluoxetine but with a notably faster onset. Mechanistically, SHED potently attenuated neuroinflammation by reducing hippocampal and cortical levels of pro inflammatory cytokines and by promoting a phenotypic shift in microglia from the M1 to the M2 state, as evidenced by morphology and marker expression. Transcriptomic analysis revealed that SHED treatment upregulated gene sets related to postsynaptic density, while downregulating the NOD like receptor (NLRP3 inflammasome) signaling pathway. At the molecular level, SHED enhanced the expression of key synaptic protein (PSD95) and restored the impaired BDNF/TrkB signaling axis in stress-vulnerable brain regions.
Conclusion: SHED exerts rapid and potent antidepressant effects in the CUMS model through a convergent dual mechanism: suppressing neuroinflammation via microglial reprogramming and inflammasome inhibition, and enhancing structural and functional synaptic plasticity. These robust preclinical findings strongly support SHED as a novel, mechanism-based, cell therapeutic strategy for major depressive disorder.
Keywords: major depressive disorder, stem cells from human exfoliated deciduous teeth, BDNF/TrkB signaling, microglial polarization, postsynaptic density, NLRP3 inflammasome
Introduction
Major depressive disorder (MDD) is a prevalent and debilitating psychiatric condition characterized by persistent low mood, anhedonia, and cognitive deficits, leading to severe functional impairment and reduced quality of life.1 MDD affects over 280 million people globally and is a leading cause of disability worldwide.2 Alarmingly, suicide linked to depression accounts for more than 700,000 deaths annually.2 Projections suggest that by 2030, MDD will represent the largest global disease burden among mental disorders.3
The pathophysiology of MDD is multifactorial. While the traditional monoamine hypothesis has guided drug development, current first-line antidepressants—primarily selective serotonin reuptake inhibitors (SSRIs)—are limited by slow onset, variable efficacy, significant side effects, and high relapse rates.4 Consequently, there is an urgent need for novel therapeutic strategies that act more rapidly and through distinct mechanisms.
Accumulating evidence highlights the central roles of neuroinflammation and impaired neuroplasticity in MDD.5–7 Chronic stress, a major risk factor for depression, can trigger systemic inflammation and elevate peripheral pro-inflammatory cytokines such as interleukin-6 (IL-6), IL-1β, and tumor necrosis factor-alpha (TNF-α), which are consistently observed in MDD patients.8,9 These inflammatory mediators can cross a compromised blood–brain barrier, activate microglia—the resident immune cells of the central nervous system—toward a pro-inflammatory (M1-like) phenotype, and disrupt neurotrophic signaling, particularly the brain-derived neurotrophic factor (BDNF) pathway.10,11 This cascade leads to synaptic dysfunction, reduced neuroplasticity, and neuronal damage, especially in stress-sensitive regions such as the hippocampus and prefrontal cortex.12,13 Therefore, interventions that simultaneously dampen neuroinflammation and enhance synaptic integrity represent a promising therapeutic direction.
Mesenchymal stem cells (MSCs) have emerged as a potential regenerative therapy due to their immunomodulatory, anti-inflammatory, and neurotrophic properties.14,15 Among various MSC sources, stem cells from human exfoliated deciduous teeth (SHED) offer distinct advantages: they are derived from the ectoderm-derived neural crest (sharing a common lineage with neural cells), exhibit strong neurogenic potential, express high levels of neurotrophic factors, and can be obtained non-invasively with minimal ethical concerns.16,17 Recent studies have demonstrated the therapeutic potential of SHED in neurodegenerative models;16 however, their efficacy and mechanisms in depression remain underexplored.
Translating findings from animal models to human therapeutics requires robust preclinical evidence. The chronic unpredictable mild stress (CUMS) model reliably recapitulates key behavioral and neurobiological features of human depression, including anhedonia, despair, and anxiety-like behaviors, along with neuroinflammatory and synaptic deficits.18,19 Thus, it serves as a valid platform for evaluating novel antidepressant interventions.
This study aimed to investigate whether intracerebroventricular transplantation of SHED could ameliorate depressive-like behaviors in CUMS-exposed rats. We hypothesized that SHED would exert antidepressant effects through a dual mechanism: (1) attenuating neuroinflammation by modulating microglial polarization and suppressing pro-inflammatory signaling pathways, and (2) enhancing neuroplasticity by restoring BDNF/TrkB signaling and synaptic protein expression. By integrating behavioral, molecular, and transcriptomic analyses, we sought to provide a comprehensive mechanistic basis for SHED-based therapy, supporting its future translation as a novel cell-based strategy for MDD.
Materials and Methods
Study Design and Timeline
This experimental study was conducted between February and December 2024. The overall design included CUMS modeling, SHED transplantation, behavioral testing, and tissue collection/analysis.
Culture and Characterization of SHED
SHED (Batch No. RDP000660403) were purchased from Sailiai Stem Cell Technology Co., Ltd. (Guangzhou, China). Cells were cultured in DMEM/F12 medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) and maintained at 37°C with 5% CO2. Upon reaching 80–90% confluence, cells were passaged. Only cells at passages 5–6 were used for transplantation.
Multilineage differentiation potential was assessed using commercial induction media (Pricella™). Osteogenic differentiation was induced for 16 days and confirmed by Alizarin Red staining for calcium deposition. Adipogenic differentiation was induced for 21 days and confirmed by Oil Red O staining for lipid droplets. Stained cells were imaged using a fluorescence microscope (Fluo-View-FV1000, Olympus, Japan).
Animals
Male 7-week-old Wistar rats (n=70) were purchased from Charles River Experimental Animal Technology Co., Ltd. (Beijing, China). Animals were housed under a 12 h light/dark cycle at 22 ± 1°C with ad libitum access to food and water. After one week of acclimation, 8-week-old rats were randomly assigned to experimental groups. All procedures were approved by the Experimental Animal Ethics Committee of Shanxi Medical University (Approval No. DWLL-2024-034) and complied with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Chronic Unpredictable Mild Stress (CUMS) Procedure
The CUMS protocol was adapted from established methods.18,20 Over six weeks, rats were exposed to 1–2 different mild stressors daily in an unpredictable order. The stressors included: empty water bottles (12 h); food deprivation (12 h); tail clipping (10 min); restraint (2 h); light cycle disruption (3–24 h); cage shaking (1 h); cage tilting (45°, 24 h); strobe lighting (12 h); damp bedding (24 h); soiled cage (24 h); social stress (24 h); plantar electrostimulation (10 × 10s pulses at 1 mA); and noise exposure (85 dB, 3 h). Control rats were housed undisturbed in a separate room.
Experimental Groups and SHED Transplantation
Rats were divided into seven groups (n=10 per group): Control (CON): No stress, no treatment. CUMS: Stress only. CUMS + Fake surgery (FS): Stress + intracerebroventricular injection of 0.9% NaCl. CUMS + Fluoxetine (Flu): Stress + oral fluoxetine (10 mg/kg/day) for the final three weeks.21 CUMS + SHED-Low (SHED-L): Stress + ICV transplantation of 0.5×106 SHED. CUMS + SHED-Medium (SHED-M): Stress + ICV transplantation of 1×106 SHED. CUMS + SHED-High (SHED-H): Stress + ICV transplantation of 2×106 SHED.
After three weeks of CUMS, SHED or vehicle were transplanted under isoflurane anesthesia using a stereotactic apparatus (RWD Life Science, Shenzhen, China). A 25 μL Hamilton syringe was used to inject cells into the lateral ventricle (coordinates: −1.0 mm posterior to bregma, ±1.5 mm lateral to midline, and −4.5 mm ventral to the dura mater) at 0.5 μL/min. The syringe remained in place for 8 min before slow withdrawal.
Behavioral Tests
To validate the depression animal model and the treatment effect, we conducted behavioral tests including the measurement of body weight, Sucrose preference,22 Open field,23 Forced swim and Y-maze.24 All behavioral tests were performed by an observer blinded to group assignment. Detailed experimental methods are provided in the Supplementary Information.
Tissue Preparation
After the completion of the behavioral tests, rats were anesthetized using inhaled isoflurane to collect brain tissue samples. Rats were perfused with 0.9% NaCl, then the whole brain was removed. The hippocampus and prefrontal cortex were identified and collected by dissection from removing the cerebral cortex of each hemisphere, and placed into separate enzyme-free 2-mL microcentrifuge tubes, then immediately transferred into a pre- chilled frozen vial and snap- frozen in liquid nitrogen and then stored at −80°C for subsequent RNA high-throughput sequencing, qRT- PCR and ELISA.
Quantitative Real-Time Polymerase Chain Reaction (qRT- PCR)
qRT-PCR analysis was performed based on a previously described method with modifications.25 Total RNA was extracted using TransZol Up Plus RNA reagent (TransGen Biotech). Complementary DNA (cDNA) was then synthesized from the extracted RNA using the TransScript Uni All-in-One First-Strand cDNA Synthesis SuperMix (TransGen Biotech). Quantitative PCR was performed on a CFX 96 Real-Time PCR system (Bio-Rad) using PerfectStart Green qPCR SuperMix (TransGen Biotech). The thermal cycling protocol consisted of an initial denaturation at 95 °C for 30 seconds, followed by 39 cycles of 95 °C for 5 seconds and an appropriate primer-specific annealing temperature for 34 seconds. All samples were assayed in triplicate. Relative gene expression levels were calculated using the 2−ΔΔCt method, with β-actin serving as the endogenous control. The sequences of all primers used are provided in the Supplementary Information.
Enzyme-Linked Immunosorbent Assay (ELISA)
The ELISA procedure was modified from established methods.25 Briefly, hippocampus and prefrontal cortex tissues from rats were homogenized individually in ice-cold PBS (5 mL/g tissue). Following centrifugation at 13,000 rpm for 20 min, the total protein concentration in the supernatant was determined using a BCA kit (Abbkine, China). All supernatants were diluted to a uniform protein concentration and subsequently assayed for IL-1β, IL-6, IL-10, TNF-α, and BDNF using respective commercial ELISA kits (ChemicalBook, China) as per the manufacturer’s protocols.
Histochemistry and Immunofluorescence
Histochemical staining was performed using the Nissl staining method to visualize neuronal cell bodies and assess cytoarchitecture.26 Hippocampal tissue sections were dehydrated, rehydrated, and stained with 0.1% cresyl violet solution according to standard protocols. After differentiation, sections were dehydrated, cleared, and mounted for microscopic examination.
The immunofluorescence staining was performed as previously described with modifications.27 Brain sections with prefrontal cortex were incubated overnight at 4°C with primary antibodies against Iba1 (Abcam, ab178847), followed by incubation with corresponding DyLight 594 conjugated secondary antibodies (Abbkine) for 1 h. Nuclei were counterstained with DAPI, and images of prefrontal cortex were captured using a fluorescence microscope (OLYMPUS BX53 F2) and analyzed with ImageJ software (v1.45J).
RNA High-Throughput Sequencing
RNA-seq was conducted as described previously with modifications.28 Total RNA was extracted with Trizol and quality-assessed (Agilent Bioanalyzer 2100); samples with RIN >7.0 were used for library preparation. mRNA was enriched using Dynabeads Oligo(dT), fragmented at 94 °C, and reverse-transcribed. Second-strand cDNA was synthesized incorporating dUTP, followed by A-tailing, adapter ligation, and size selection. After UDG treatment, libraries were amplified by PCR (8 cycles) and sequenced on an Illumina Novaseq™ 6000 (PE150). Gene set enrichment analysis (GSEA v4.1.0)29 was performed using MSigDB. GO terms with |NES|>1, NOM p-val<0.05, and FDR q-val<0.25 were considered significantly enriched.
Statistical Analysis
Data are presented as mean ± SEM. Statistical analysis was performed using SPSS 22.0 and GraphPad Prism 9.3.0. One-way ANOVA followed by appropriate post-hoc tests was used for group comparisons. Statistical significance was set at P < 0.05.
Results
Culture and Characterization of SHED
SHEDs displayed a fibroblast-like morphology, characterized by abundant cytoplasm, large nuclei with fine chromatin and prominent nucleoli, and were often arranged in a whorl-like pattern (Figure 1A and B). To evaluate their multipotent differentiation capacity, SHEDs were subjected to osteogenic and adipogenic induction. Following 16 days of culture in osteogenic medium, the cells differentiated into osteocytes, as evidenced by Alizarin Red staining of calcium mineral deposits. In contrast, after 21 days in adipogenic medium, SHEDs underwent adipogenic differentiation, indicated by the formation of lipid vacuoles stained with Oil Red O (Figure 1C and D).
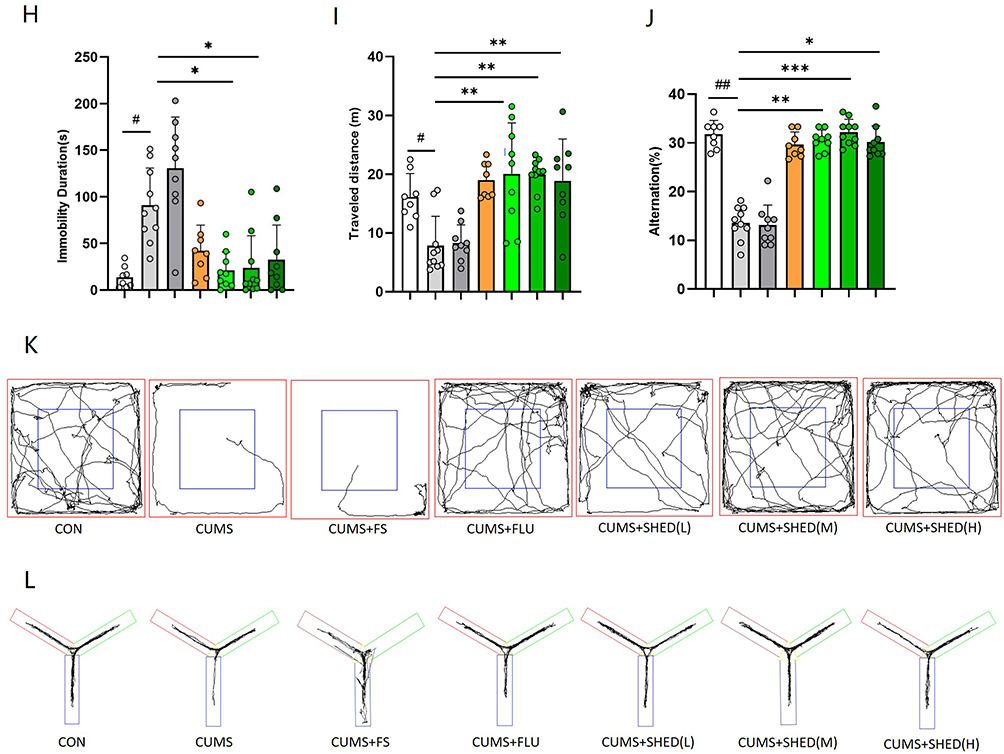
Figure 1 continued. Figure 1 Culture and characterization of SHEDs (A–D). SHEDs exhibited a fibroblast-like morphology and arranged in a whorl-like pattern (A with scale bar: 200 μm, B with scale bar: 100 μm) Bright- field images for cell morphology and staining of osteogenic derivatives with calcium deposition, which was stained Orange- red by Alizarin Red (C with scale bar: 100 μm) and adipogenic derivatives with formation of lipid droplets, which was stained red in Oil Red O (D with scale bar: 100 μm). SHEDs ameliorated depressive-like behaviors induced by CUMS (E–L). The schematic of the experimental time line (E). Body weight was measured during the experimental period (F). The sucrose preference test (G), the forced swimming test (H), the open field test (OFT)(I), the Y maze test (J), the graphic representation of OFT (K), the graphic representation of Y maze test (L). Data were presented as mean ± SD, n = 8–10 rats per group. ns, P > 0.05. #P < 0.05, ##P < 0.01, ###P < 0.001 versus the CON group. *P < 0.05, **P < 0.01, ***P < 0.001 versus the CUMS group.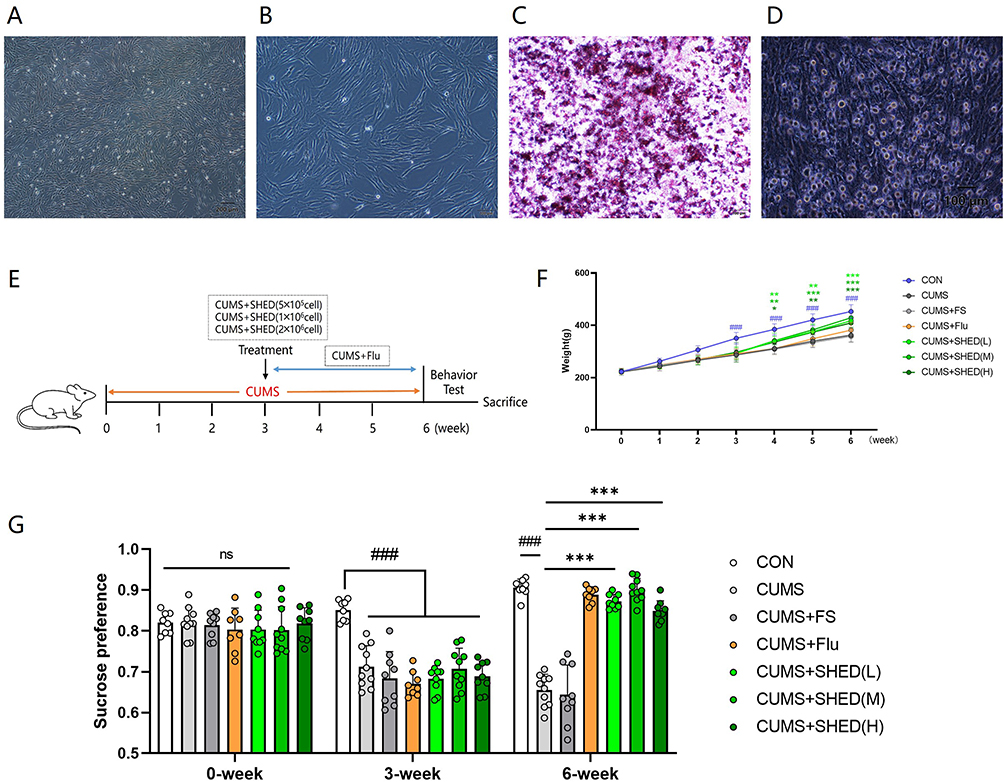
SHED Transplantation Rapidly Ameliorates CUMS-Induced Depressive-Like Behaviors
To evaluate the therapeutic potential of SHED, CUMS-exposed rats received intracerebroventricular transplantation of SHED at three doses (0.5×106, 1×106, or 2×106 cells/rat). As illustrated in the experimental timeline (Figure 1E), behavioral tests were conducted following treatment.
CUMS induced a significant reduction in body weight compared to unstressed controls (Figure 1F). While fluoxetine treatment did not reverse this weight loss, all three SHED doses effectively restored body weight to near-normal levels, with no significant differences among dosage groups.
In the sucrose preference test (SPT), CUMS rats exhibited marked anhedonia, shown by reduced sucrose preference. Both SHED transplantation and fluoxetine treatment significantly increased sucrose preference, with SHED groups showing a rapid response (Figure 1G). In the forced swim test (FST), CUMS prolonged immobility time, indicating behavioral despair. SHED treatment significantly shortened immobility time, to an extent comparable to fluoxetine (Figure 1H). Anxiety-like behavior was assessed in the open field test (OFT). CUMS reduced total distance traveled and center zone activity, both of which were restored by SHED and fluoxetine (Figure 1I). Spatial working memory, evaluated using the Y-maze spontaneous alternation test, was impaired in CUMS rats and significantly improved by SHED treatment (Figure 1J). Figure 1K and L vividly represent the results of OFT and Y-maze. Notably, the antidepressant-like effects of SHED were not dose-dependent within the tested range, suggesting a possible ceiling effect due to direct intracerebroventricular delivery.
SHED Attenuates Neuroinflammation and Modulates Microglial Polarization
Given the established link between depression and neuroinflammation, we examined inflammatory cytokine expression in the hippocampus and prefrontal cortex. CUMS significantly upregulated mRNA levels of pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) and SHED transplantation effectively reversed these changes, meanwhile SHED upregulated the anti-inflammatory cytokine IL-10. (Figure 2A–D). Consistent results were observed at the protein level via ELISA in hippocampus tissue (Figure 2E–H). Microglial activation and phenotype switching play a key role in stress-induced neuroinflammation. At the molecular level, SHED downregulated M1 marker iNOS and upregulated the M2 marker Arg1, indicating a shift toward an anti-inflammatory phenotype (Figure 2I and J). Neuroinflammation can damage the neurons. Nissl staining of the rat hippocampal CA3 region revealed impaired neuronal cytoarchitecture in CUMS model rats, which was ameliorated by SHED treatment. Normal controls exhibited densely packed, orderly arranged neurons with large, clear nuclei and abundant Nissl substance. In contrast, the CUMS group showed neuronal disorganization, cell shrinkage, loss of Nissl bodies, and increased interstitial vacuolation. SHED treatment markedly restored neuronal density and Nissl body integrity (Figure 2K). Immunofluorescence analysis of the prefrontal cortex revealed that CUMS increased the number and soma size of Iba1⁺ microglia, accompanied by shortened processes and reduced branching—features of a pro-inflammatory (M1-like) state. SHED treatment reversed these morphological changes, promoting a more ramified, surveillant morphology (Figure 2L–O). Figure 2 continued. Figure 2 SHEDs modulated neuroinflammation in the central nervous system (CNS). Expressions of neural protective factors and inflammatory factors in rat hippocampus and prefrontal cortex were quantitated using qRT- PCR for pro- inflammatory factors IL- 1β (A), IL- 6 (B), TNF- α (C), and anti- inflammatory factors IL- 10 (D). The protein levels of inflammatory factors in rat hippocampus were measured using ELISA for IL- 1β (E), IL- 6 (F), TNF- α (G), IL-10 (H). Expressions of markers of microglial polarization in rat prefrontal cortex were quantitated using qRT- PCR for Arg1 (I), and iNOS (J) (n = 4 rats per group). Representative photomicrographs of the hippocampal CA3 region stained with Nissl staining (scale bar: 200 μm) (K). Representative fluorescence micrographs of microglia in prefrontal cortex. Microglia were stained with Iba1 (red) by Immunofluorescence (L), area of Iba1+ cells (%) (M), Iba1+ cells /mm2 (N), the number of branches (O) were measured. n = 5 rats per group. 5–8 slices containing hippocampus or prefrontal cortex were collected from each sample. Data were presented as mean ± SD. ns, P > 0.05. #P < 0.05, ##P < 0.01, ###P < 0.001 versus the CON group. *P < 0.05, **P < 0.01, ***P < 0.001 versus the CUMS group.

Transcriptomic Profiling Reveals SHED-Mediated Synaptic Restoration and Suppression of Inflammatory Signaling
To gain an unbiased view of SHED’s actions, RNA sequencing was performed on hippocampal tissue. Differential expression analysis identified significant changes between the CUMS+SHED and CUMS groups (Figure 3A–C). Gene set enrichment analysis (GSEA) revealed that SHED upregulated genes associated with postsynaptic density, glutamatergic synapse, and long-term memory, while downregulating genes involved in the NOD-like receptor (NLR) signaling pathway (Figure 3D). Enrichment score (ES) plots demonstrated a distinct positive peak for the gene set “postsynaptic density” (NES = 1.94, P = 0), indicating widespread upregulation in the SHED-treated group and suggesting synaptic functional recovery (Figure 3E). Conversely, the “nod-like receptor signaling pathway” displayed a negative peak (NES = −1.46, P = 0.004), reflecting significant upregulation in the CUMS group and subsequent normalization after SHED treatment (Figure 3F). Figure 3 continued. Figure 3 Transcriptomic profile of rats’ hippocampus (A–F). The barplot (A), the volcano (B) and the heatmap (C) for DEGs derived from the CUMS+SHED group and the CUMS group were compared. The top 30 GSEA enrichment analysis showed the downregulated and upregulated pathways between the CUMS+SHED group and the CUMS group (D). The GSEA Enrichment Score (ES) line plot for postsynaptic density (E) and nod like receptor signaling pathway (F). Expressions of synaptic marker, the markers of nod like receptor signaling pathway, and neural protective factors in rat hippocampus and prefrontal cortex were quantitated using qRT- PCR for PSD95 (G), NLRP3 (H), Caspase-1 (I), BDNF (J) and TrkB (K). The protein level of BDNF (L) in rat hippocampus was measured using ELISA. Data were presented as mean ± SD, n = 5 rats per group. ns, P > 0.05. #P < 0.05, ##P < 0.01, ###P < 0.001 versus the CON group. *P < 0.05, **P < 0.01, ***P < 0.001 versus the CUMS group.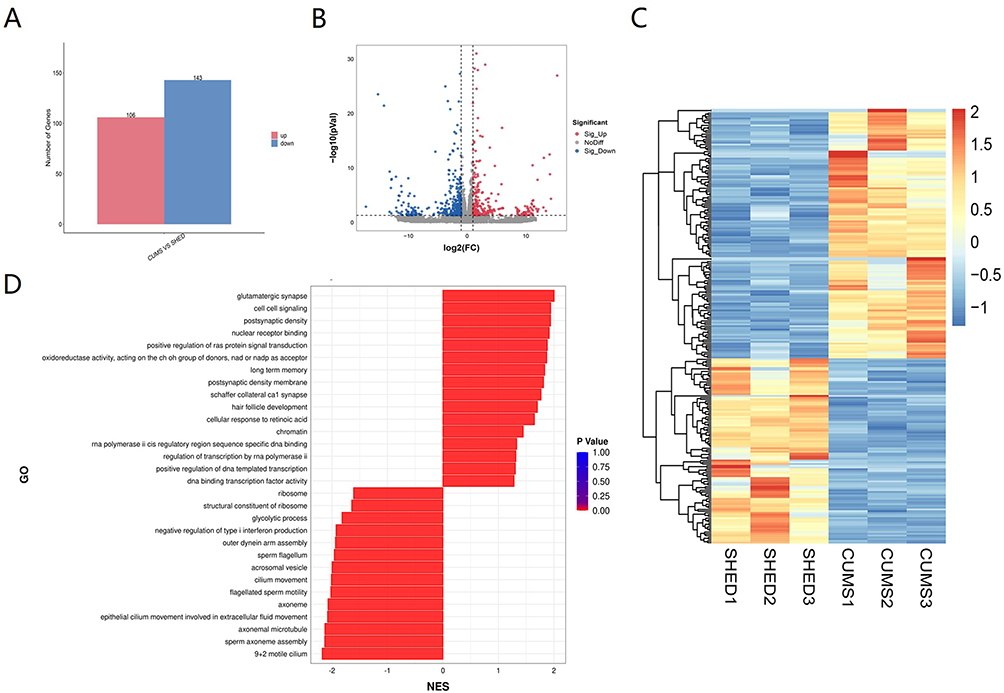

Consistent with transcriptomic findings, qRT-PCR confirmed that SHED increased mRNA level of synaptic marker PSD95, which were reduced by CUMS (Figure 3G). Concurrently, SHED downregulated key components of the NLRP3 inflammasome pathway—NLRP3 and caspase-1—that were elevated in CUMS rats (Figure 3H and I).
SHED Enhances BDNF/TrkB Signaling, a Key Pathway for Neuroplasticity
The BDNF/TrkB pathway is central to synaptic plasticity and is often impaired in depression. qRT-PCR analysis showed that CUMS downregulated BDNF and TrkB mRNA expression in both the hippocampus and prefrontal cortex. SHED treatment significantly restored their expression to levels comparable to those in fluoxetine-treated rats (Figure 3J and K). ELISA further confirmed these trends at the protein level in hippocampal tissue (Figure 3L). These results suggest that SHED promotes neuroplasticity, at least in part, through modulation of the BDNF/TrkB signaling axis.
Discussion
The present study demonstrates that intracerebroventricular transplantation of stem cells from human exfoliated deciduous teeth (SHED) significantly alleviates depressive-like behaviors in the CUMS rat model. Our findings support the initial hypothesis that SHED exerts its antidepressant effects through a dual mechanism: attenuating neuroinflammation and enhancing neuroplasticity. Notably, SHED produced a rapid behavioral improvement, with efficacy comparable to fluoxetine, yet without the dose-dependent response within the tested range—an observation that underscores its distinct mode of action.
Rapid Behavioral Recovery and Possible Mechanisms
Unlike conventional antidepressants, which often require weeks to take effect, SHED transplantation led to a swift reversal of CUMS-induced deficits in sucrose preference, locomotor activity, and behavioral despair. This rapid onset aligns with emerging evidence that certain stem cell therapies can modulate neuroimmune responses more directly than pharmacotherapies acting solely on monoamine systems. The lack of dose dependence may be attributable to the intracerebroventricular delivery route, which likely maximizes local bioavailability of SHED-secreted factors in peri ventricular brain regions involved in mood regulation.
Suppression of Neuroinflammation via Microglial Reprogramming
Chronic stress triggers a neuroinflammatory cascade characterized by elevated pro inflammatory cytokines and microglial activation, both of which are implicated in the pathophysiology of depression.8,10 Our results confirm that SHED effectively reduces levels of IL 1β, IL 6, and TNF α while increasing the anti-inflammatory cytokine IL 10. More importantly, SHED promoted a phenotypic shift in microglia from a pro inflammatory M1 like state (marked by elevated IL 1β, IL 6, TNF α, and iNOS) toward an anti-inflammatory, tissue reparative M2 like state (indicated by increased IL-10 and Arg1). These changes were accompanied by a morphological reversion from an amoeboid, activated shape to a ramified, surveillant morphology. This reprogramming of microglial function represents a key mechanism through which SHED mitigates neuroinflammation, consistent with recent studies highlighting the role of mesenchymal stem cells in modulating glial responses in psychiatric models.30
Transcriptomic Insights into Synaptic Restoration and Inflammasome Suppression
RNA sequencing analysis provided an unbiased systems level view of SHED’s actions. GSEA revealed that SHED upregulated gene sets related to postsynaptic density and glutamatergic synapse—functional categories tightly linked to synaptic plasticity and cognitive function.31 Conversely, SHED downregulated the NOD like receptor (NLR) signaling pathway, a key intracellular inflammatory cascade activated by stress associated damage-associated molecular patterns (DAMPs). In particular, SHED reduced hippocampal expression of NLRP3 and caspase 1, core components of the NLRP3 inflammasome. This pathway has been identified as a critical mediator linking chronic stress to neuroinflammation and depressive like behaviors.32 Our data thus position SHED as a regulator of both synaptic integrity and innate immune signaling in the stressed hippocampus.
Enhancement of Neuroplasticity via BDNF/TrkB Signaling
The BDNF/TrkB pathway is a well-established molecular substrate of neuroplasticity and is frequently impaired in depression.11 Our results show that SHED restored CUMS induced reductions in BDNF and TrkB expression in both the hippocampus and prefrontal cortex. This restoration was associated with increased expression of synaptic proteins PSD95, suggesting that SHED not only elevates trophic support but also promotes structural synaptic remodeling. Nissl staining further confirmed that CUMS induced significant neuronal loss, shrinkage, and loss of Nissl substance in the hippocampal CA3 region, all of which were attenuated by SHED treatment. This structural preservation aligns with the observed functional recovery in synaptic and behavioral endpoints. The coordinated upregulation of BDNF/TrkB signaling and postsynaptic scaffolding proteins indicates that SHED targets a central plasticity related axis that is often resistant to conventional antidepressants in treatment resistant cases.33
Contribution to the Literature and Translational Perspective
While previous studies have demonstrated the potential of SHED in neurodegenerative,16 neuroinflammatory,34 and injury contexts—such as alleviating Alzheimer’s pathology,35 ameliorating autistic-like behaviors,36 promoting spinal cord recovery,37 and mitigating high-altitude cerebral edema,38—evidence for its efficacy in depression remains scarce. Our study extends this literature by providing integrated behavioral, molecular, and transcriptomic evidence that SHED acts through a convergent dual mechanism in a validated depression model. This dual action—simultaneously suppressing neuroinflammation (via microglial repolarization and NLRP3 inflammasome inhibition) and enhancing synaptic plasticity (via BDNF/TrkB signaling and postsynaptic protein restoration)—aligns with the growing view that effective antidepressant interventions should address both pathological axes. Notably, our work highlights the rapid behavioral onset of SHED compared to fluoxetine and reveals previously unreported transcriptomic shifts involving postsynaptic density and NLR pathways. Compared to other MSC sources, SHED offers practical advantages such as noninvasive harvesting and neural crest origin, which may enhance its neurogenic and immunomodulatory potency.39 These robust preclinical findings strongly support further investigation of SHED as a novel, mechanism‑based therapeutic strategy for MDD.
Limitations and Future Directions
Several limitations warrant consideration. First, while the CUMS model is well validated, it does not fully capture the etiological complexity of human MDD. Second, the long-term durability of SHED’s effects remains to be evaluated. Third, although we identified key pathways affected by SHED, the specific paracrine factors (eg, exosomes, cytokines, or metabolites) responsible for these changes require further isolation and characterization. Future studies should focus on identifying these effector molecules to develop potentially cell free, targeted therapies based on SHED’s mechanism.
Conclusion
In summary, this study demonstrates that SHED transplantation produces rapid and potent antidepressant-like effects in CUMS-exposed rats. These benefits are mediated through a convergent dual mechanism: suppressing neuroinflammation via microglial reprogramming and NLRP3 inflammasome inhibition, and enhancing neuroplasticity via BDNF/TrkB signaling and synaptic protein restoration. These findings support further investigation of SHED as a novel, mechanism-based therapeutic strategy for major depressive disorder.
Abbreviations
MDD, major depressive disorder; CUMS, Chronic Unpredictable Mild Stress; Flu, fluoxetine; ICV, intracerebroventricular; RT-qPCR, real-time quantitative reverse transcription polymerase chain reaction; ELISA, Enzyme-Linked Immunosorbent Assay; DEGs, differentially expressed genes; GSEA, Gene set enrichment analysis; BDNF, brain-derived neurotrophic factor; TrkB, tropomyosin receptor kinase B; Arg1, Arginase 1; iNOS, Inducible nitric oxide synthase; PSD95, synaptic scaffolding protein 95; NLRP3, NLR Family, PYRIN domain-containing protein 3; Caspase-1, Cysteine-requiring aspartate protease-1.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
All procedures were approved by the Experimental Animal Ethics Committee of Shanxi Medical University (Approval No. DWLL-2024-034) and complied with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Acknowledgments
The authors would like to express their gratitude to everyone who contributed to this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82371511, 82271546, 82301725); National Key Research and Development Program of China (2023YFC2506201); Key Project of Science and Technology Innovation 2030 of China (2021ZD0201805); Fundamental Research Program of Shanxi Province (202203021212028).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Malhi GS, Mann JJ. Depression. Lancet. 2018;392(10161):2299–14. doi:10.1016/s0140-6736(18)31948-2
2. GBD 2019 Mental Disorders Collaborators. Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990–2019: a systematic analysis for the global burden of disease study 2019. Lancet Psychiatry. 2022;9(2):137–150. doi:10.1016/s2215-0366(21)00395-3
3. Liu Q, He H, Yang J, Feng X, Zhao F, Lyu J. Changes in the global burden of depression from 1990 to 2017: findings from the global burden of disease study. J Psychiatr Res. 2020;126:134–140. doi:10.1016/j.jpsychires.2019.08.002
4. Marx W, Penninx B, Solmi M, et al. Major depressive disorder. Nat Rev Dis Primers. 2023;9(1):44. doi:10.1038/s41572-023-00454-1
5. Troubat R, Barone P, Leman S, et al. Neuroinflammation and depression: a review. Eur J Neurosci. 2021;53(1):151–171. doi:10.1111/ejn.14720
6. Beurel E, Toups M, Nemeroff CB. The bidirectional relationship of depression and inflammation: double trouble. Neuron. 2020;107(2):234–256. doi:10.1016/j.neuron.2020.06.002
7. Duman RS, Sanacora G, Krystal JH. Altered connectivity in depression: GABA and glutamate neurotransmitter deficits and reversal by novel treatments. Neuron. 2019;102(1):75–90. doi:10.1016/j.neuron.2019.03.013
8. Köhler CA, Freitas TH, Maes M, et al. Peripheral cytokine and chemokine alterations in depression: a meta-analysis of 82 studies. Acta Psychiatr Scand. 2017;135(5):373–387. doi:10.1111/acps.12698
9. Osimo EF, Pillinger T, Rodriguez IM, Khandaker GM, Pariante CM, Howes OD. Inflammatory markers in depression: a meta-analysis of mean differences and variability in 5166 patients and 5083 controls. Brain Behav Immun. 2020;87:901–909. doi:10.1016/j.bbi.2020.02.010
10. Wang H, He Y, Sun Z, et al. Microglia in depression: an overview of microglia in the pathogenesis and treatment of depression. J Neuroinflammation. 2022;19(1):132. doi:10.1186/s12974-022-02492-0
11. Zhang JC, Yao W, Hashimoto K. Brain-derived Neurotrophic Factor (BDNF)-TrkB signaling in inflammation-related depression and potential therapeutic targets. Curr Neuropharmacol. 2016;14(7):721–731. doi:10.2174/1570159x14666160119094646
12. McEwen BS, Nasca C, Gray JD. Stress effects on neuronal structure: hippocampus, amygdala, and prefrontal cortex. Neuropsychopharmacology. 2016;41(1):3–23. doi:10.1038/npp.2015.171
13. Duman RS, Aghajanian GK, Sanacora G, Krystal JH. Synaptic plasticity and depression: new insights from stress and rapid-acting antidepressants. Nat Med. 2016;22(3):238–249. doi:10.1038/nm.4050
14. Song N, Scholtemeijer M, Shah K. Mesenchymal stem cell immunomodulation: mechanisms and therapeutic potential. Trends Pharmacol Sci. 2020;41(9):653–664. doi:10.1016/j.tips.2020.06.009
15. Andrzejewska A, Dabrowska S, Lukomska B, Janowski M. Mesenchymal stem cells for neurological disorders. Adv Sci. 2021;8(7):2002944. doi:10.1002/advs.202002944
16. Candelise N, Santilli F, Fabrizi J, et al. The importance of stem cells isolated from human dental pulp and exfoliated deciduous teeth as therapeutic approach in nervous system pathologies. Cells. 2023;12(13):1686. doi:10.3390/cells12131686
17. Hochuli AHD, Senegaglia AC, Selenko AH, Fracaro L, Brofman PRS. Dental pulp from human exfoliated deciduous teeth-derived stromal cells demonstrated neuronal potential: in vivo and in vitro studies. Curr Stem Cell Res Ther. 2021;16(5):495–506. doi:10.2174/1574888x16666210215160402
18. Antoniuk S, Bijata M, Ponimaskin E, Wlodarczyk J. Chronic unpredictable mild stress for modeling depression in rodents: meta-analysis of model reliability. Neurosci Biobehav Rev. 2019;99:101–116. doi:10.1016/j.neubiorev.2018.12.002
19. Planchez B, Surget A, Belzung C. Animal models of major depression: drawbacks and challenges. J Neural Transm. 2019;126(11):1383–1408. doi:10.1007/s00702-019-02084-y
20. Wang P, Li Y, Song Y, et al. Human umbilical cord mesenchymal stem cells reverse depression in rats induced by chronic unpredictable mild stress combined with lipopolysaccharide. CNS Neurosci Ther. 2024;30(3):e14644. doi:10.1111/cns.14644
21. Taguchi R, Shikata K, Furuya Y, Ino M, Shin K, Shibata H. Selective corticotropin-releasing factor 1 receptor antagonistE2508has potent antidepressant-like and anxiolytic-like properties in rodent models. Behav Brain Res. 2016;312:138–147. doi:10.1016/j.bbr.2016.06.017
22. Liu MY, Yin CY, Zhu LJ, et al. Sucrose preference test for measurement of stress-induced anhedonia in mice. Nat Protoc. 2018;13(7):1686–1698. doi:10.1038/s41596-018-0011-z
23. Kraeuter AK, Guest PC, Sarnyai Z. The open field test for measuring locomotor activity and anxiety-like behavior. Methods Mol Biol. 2019;1916:99–103. doi:10.1007/978-1-4939-8994-2_9
24. Kraeuter AK, Guest PC, Sarnyai Z. The Y-maze for assessment of spatial working and reference memory in mice. Methods Mol Biol. 2019;1916:105–111. doi:10.1007/978-1-4939-8994-2_10
25. Jiang X, Yi S, Liu Q, et al. Asperosaponin VI ameliorates the CMS-induced depressive-like behaviors by inducing a neuroprotective microglial phenotype in hippocampus via PPAR-γ pathway. J Neuroinflammation. 2022;19(1):115. doi:10.1186/s12974-022-02478-y
26. Dai S, Wei J, Zhang H, et al. Intermittent fasting reduces neuroinflammation in intracerebral hemorrhage through the Sirt3/Nrf2/HO-1 pathway. J Neuroinflammation. 2022;19(1):122. doi:10.1186/s12974-022-02474-2
27. Liu G, Miao L, Niu H, et al. Human umbilical cord mesenchymal stem cells ameliorated chronic unpredictable mild stress-induced depression and anxiety by alleviating neuroinflammation. J Neuroimmune Pharmacol. 2025;20(1):45. doi:10.1007/s11481-025-10198-2
28. Li B, Li Y, Zhou H, et al. Multiomics identifies metabolic subtypes based on fatty acid degradation allocating personalized treatment in hepatocellular carcinoma. Hepatology. 2024;79(2):289–306. doi:10.1097/hep.0000000000000553
29. Subramanian A, Tamayo P, Mootha VK, et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A. 2005;102(43):15545–15550. doi:10.1073/pnas.0506580102
30. Sánchez-Castillo AI, Sepúlveda MR, Marín-Teva JL, et al. Switching roles: beneficial effects of adipose tissue-derived mesenchymal stem cells on microglia and their implication in neurodegenerative diseases. Biomolecules. 2022;12(2):219. doi:10.3390/biom12020219
31. Yang X, Huang YA, Marshall J. Targeting TrkB-PSD-95 coupling to mitigate neurological disorders. Neural Regen Res. 2025;20(3):715–724. doi:10.4103/nrr.Nrr-d-23-02000
32. Feng X, Zhao Y, Yang T, et al. Glucocorticoid-driven NLRP3 inflammasome activation in hippocampal microglia mediates chronic stress-induced depressive-like behaviors. Front Mol Neurosci. 2019;12:210. doi:10.3389/fnmol.2019.00210
33. Shi X, Zhou XZ, Chen G, et al. Targeting the postsynaptic scaffolding protein PSD-95 enhances BDNF signaling to mitigate depression-like behaviors in mice. Sci Signal. 2024;17(834):eadn4556. doi:10.1126/scisignal.adn4556
34. Su J, Ge X, Jiang N, Zhang Z, Wu X. Efficacy of mesenchymal stem cells from human exfoliated deciduous teeth and their derivatives in inflammatory diseases therapy. Curr Stem Cell Res Ther. 2022;17(4):302–316. doi:10.2174/1574888x17666220417153309
35. Zhang X, Lei T, Wang D, et al. Stem cells from human exfoliated deciduous teeth relieves Alzheimer’s disease symptoms in SAMP8 mice by up-regulating the PPARγ pathway. Biomed Pharmacother. 2022;152:113169. doi:10.1016/j.biopha.2022.113169
36. Zhao L, Li Y, Kou X, et al. Stem cells from human exfoliated deciduous teeth ameliorate autistic-like behaviors of SHANK3 mutant beagle dogs. Stem Cells Transl Med. 2022;11(7):778–789. doi:10.1093/stcltm/szac028
37. Nishii T, Osuka K, Nishimura Y, et al. Protective mechanism of stem cells from human exfoliated deciduous teeth in treating spinal cord injury. J Neurotrauma. 2024;41(9–10):1196–1210. doi:10.1089/neu.2023.0251
38. Wang YM, Zhou YK, Han CS, et al. Stem cells from human exfoliated deciduous teeth alleviate high-altitude cerebral oedema by shifting microglial M1/M2 polarisation. Chin J Dent Res. 2023;26(3):153–162. doi:10.3290/j.cjdr.b4330807
39. Huang L, Zheng Z, Bai D, Han X. Stem cells from human exfoliated deciduous teeth and their promise as preventive and therapeutic strategies for neurological diseases and injuries. Curr Stem Cell Res Ther. 2022;17(6):527–536. doi:10.2174/1574888x17666211229155533
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Altered Neuroplasticity in Epilepsy is Associated with Neuroinflammation and Oxidative Stress: In vivo Evidence of Brain-Derived Extracellular Vesicles
Wang S, Zhang H, Li R, Liu Z, Xiang D
International Journal of Nanomedicine 2025, 20:7185-7197
Published Date: 4 June 2025
Sodium-Glucose Cotransporter-2 Inhibitors in Mood Disorders: A Narrative Review of Mechanisms, Evidence, and Challenges
Guo X, Yang T, Fang Y, Chen J
Neuropsychiatric Disease and Treatment 2026, 22:599108
Published Date: 8 April 2026