Back to Journals » Cancer Management and Research » Volume 12
SPOCK1 Involvement in Epithelial-to-Mesenchymal Transition: A New Target in Cancer Therapy?
Authors Sun L ![]() , Li S, Guo Q, Zhou W, Zhang H
, Li S, Guo Q, Zhou W, Zhang H
Received 14 February 2020
Accepted for publication 22 April 2020
Published 18 May 2020 Volume 2020:12 Pages 3561—3569
DOI https://doi.org/10.2147/CMAR.S249754
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Yong Teng
Li-rui Sun,1,* Si-yu Li,2,* Qiu-shi Guo,1 Wei Zhou,1 Hong-mei Zhang1
1Department of Pharmacy, The First Hospital of Jilin University, Changchun, Jilin, People’s Republic of China; 2Department of Pathology, Hangzhou Third Hospital, Hangzhou, Zhejiang, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Wei Zhou; Hong-mei Zhang
Department of Pharmacy, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China
Email [email protected]; [email protected]
Background: Cancer metastasis is the main obstacle to increasing the lifespan of cancer patients. Epithelial-to-mesenchymal transition (EMT) plays a significant role in oncogenic processes, including tumor invasion, intravasation, and micrometastasis formation, and is especially critical for cancer invasion and metastasis. The extracellular matrix (ECM) plays a crucial role in the occurrence of EMT corresponding to the change in adhesion between cells and matrices.
Conclusion: SPOCK1 is a critical regulator of the ECM and mediates EMT in cancer cells. This suggests an important role for SPOCK1 in tumorigenesis, migration and invasion. SPOCK1 is a critical regulator of some processes involved in cancer progression, including cancer cell proliferation, apoptosis and migration. Herein, the functions of SPOCK1 in cancer progression are expounded, revealing the association between SPOCK1 and EMT in cancer metastasis. SPOCK1 is a positive downstream regulator of transforming growth factor-β, and SPOCK1-mediated EMT regulates invasion and metastasis through the Wnt/β-catenin pathway and PI3K/Akt signaling pathway. It is of significance that SPOCK1 may be an attractive prognostic biomarker and therapeutic target in cancer treatment.
Keywords: SPOCK1, epithelial-to-mesenchymal transition, extracellular matrix, cancer, metastasis
Introduction
The number of deaths caused by cancer was estimated to be 9.6 million in 2018, and it is anticipated that cancer will be the top cause of death worldwide in the 21st century.1 Increasing the life expectancy of patients who have cancer with a poor survival rate is a major obstacle, mainly because of early spread of tumors via metastasis or direct invasion.2
Metastasis is the spread of tumors from the primary site to distant sites of the body. The spread of cancer cells is considered to be a vital step in the progression of tumors. Epithelial-to-mesenchymal transition (EMT) is an embryonic program that relaxes cell-cell binding complexes and gives cells enhanced migration and invasion characteristics that can be exploited by cancer cells during metastatic progression. Cancer cells undergoing EMT are more aggressive, showing more invasive, stem cell-like characteristics and resistance to apoptosis.3 As a critical element of the cancer microenvironment, the extracellular matrix (ECM) provides mechanical support for tissues and mediates the interaction between cells and the microenvironment, which is critical to the invasion of cancer cells. The ECM is increasingly taken as a key regulator in cancer.4 The ECM plays a crucial role in the occurrence of EMT according to the change in adhesion between cells and matrices. To maintain tissue differentiation, the ECM is highly dynamic and continually remodeled.5 This process is mediated by a variety of types of peptidases that can degrade the macromolecules of the ECM.6 In addition, peptidases play a role in the development of cancer and are involved in physiological functions such as angiogenesis, receptor activation, morphogenesis and apoptosis.7 During tumorigenesis, dramatic changes in ECM homeostasis are driven by alterations in peptidases and interruptions in the balance between ECM synthesis and degradation.8
SPARC/osteonectin, cwcv and kazal-like domains proteoglycan 1 (SPOCK1), a glycoprotein isolated from human testes, is one of the secreted protein, acidic, cysteine-rich (SPARC) family members and was originally called “testican-1”. SPOCK1 is highly expressed in the brain, cartilage, vascular endothelium, myoblasts, fibroblasts, lymphocytes, and neuromuscular junctions (NMJs), which suggests that the proteoglycan is involved in several physiological functions.9–14 It is worthwhile to investigate the effects of SPOCK1 on cancer progression based on the structural similarities between SPOCK1 and SPARC and the role of SPARC in cancer development.15 SPOCK1 is a critical regulator of the ECM and mediates EMT in cancer cells.16 This suggests an important role for SPOCK1 in tumorigenesis, migration and invasion.
In recent years, an increasing number of publications have paid close attention to the role of SPOCK1 in cancer diagnosis and treatment. Herein, the functions of SPOCK1 in cancer progression are expounded, revealing the association of SPOCK1 with ECM and EMT in cancer metastasis.
The Structure of SPOCK1
The SPARC family, comprising SPARC, SPOCK1, SPOCK2 and SPOCK3, secretes follistatin-like protein 1, Hevin and modular calcium binding protein 1 and 2.17 SPARC family members share a basic structure, which is composed of an N-terminus, follistatin-like domain and C-terminus.16 In contrast to SPOCK2 and SPOCK3, which are pure heparan sulfate proteoglycans, SPOCK1 carries both heparan and chondroitin sulfate chains, SPOCK2 is N-glycosylated and SPOCK3 is composed of several mucin-type O-glycans, suggesting that there are specific physiological roles for each SPOCK.18–20
The 439-amino acid of SPOCK1 is encoded by SPOCK1, which is located at 5q31. SPOCK1 consists of 11 exons. The first, second and fourth osteonectin domains are encoded by the overlap of exons 2, 3, 4 and exon 8. The third osteonectin domain and the Kazal-like domains are encoded by exons 5 and 6. The CWCV domain is encoded by exon 10.21 Thyropin domain homology occurs between residues 310 and 379, comprised of exons 9 and 10.22,23
SPOCK1 is a multidomain protein that is homologous to different protease inhibitors.24 The proteoglycan is composed of the N-terminus, follistatin-like domain, extracellular calcium binding domain and C-terminus containing a thyropin domain and domain V, including two potential glycosaminoglycan attachment (GAG) sites (Figure 1).23
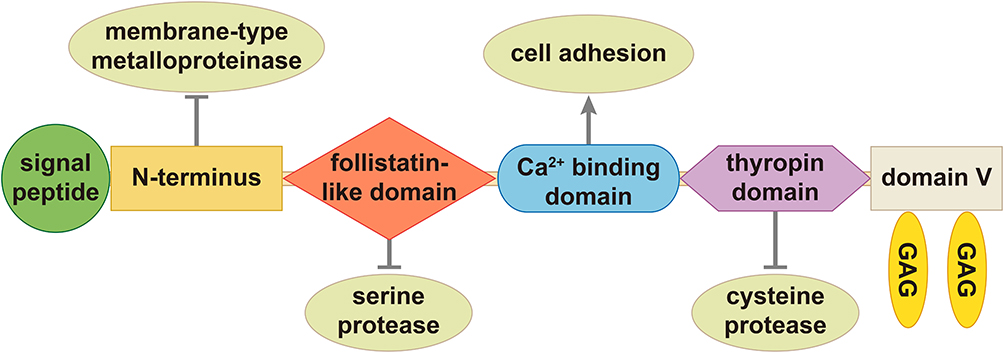 |
Figure 1 The structure of SPOCK1. SPOCK1 is constitutive of N-terminus, follistatin-like domain, extracellular calcium binding domain and C-terminus containing a thyropin domain and domain V including two potential glycosaminoglycan attachment (GAG) sites. Notes: Reproduced with permission from Bocock JP, Edgell CJ, Marr HS, Erickson AH. Human proteoglycan 435 testican-1 inhibits the lysosomal cysteine protease cathepsin L. Eur J Biochem. 2003;270(19):4008–4015. John Wiley and Sons copyright 2003.23 |
The unique N-terminus is responsible for the inhibition of a membrane-type metalloproteinase.26 There is similarity between the six-cysteine-follistatin-like domain and the Kazal domains in serine protease inhibitors.27–29 The extracellular calcium binding domain is an independent domain that belongs to the SPARC family and is involved in cell adhesion.30,31 The thyropin domain is constitutive of 64 amino acids and homologous to protein sequences that inhibit cysteine proteases cathepsin-L.22,23 SPOCK1 can be classified in a subset of the osteonectin domains and is a highly conservative chimeric proteoglycan that carries both heparan sulfate and chondroitin sulfate chains.9 Due to its modular structure, SPOCK1 is classified as a regulator of the ECM that can interact with cell surfaces.
Physiological Functions of SPOCK1
SPOCK1 was first isolated in human testicular seminal plasma.23,32,33 SPOCK1 has different expression levels in tissues, and its expression is highest in specific regions of the brain, including the axons and neuromuscular junctions.9,34,35 It has been indicated that there are systemic effects of SPOCK1 because it can also be found in several other tissues and organs, such as lymphocytes and blood (Table 1).2,13,14,18,36,37
The highest expression of human SPOCK1 is in the central nervous system (CNS), including neurons, endothelial cells, and activated astroglia, predominantly in the postsynaptic region of pyramidal cells in the hippocampus CA3 region.34,35 Neuronal cell growth and substrate attachment are regulated by SPOCK1, which indicates the role of SPOCK1 in brain development.25 SPOCK1 may participate in diverse steps of neurogenesis and participate in various neuronal processes in the CNS via involvement in brain cell adhesion.26,32,38 SPOCK1 plays a role in the behavior of the Rett syndrome phenotype, modulation of dopamine neuron population size and necrosis after brain injury.13,39,40
In the peripheral nervous system, SPOCK1 is expressed in axons, Schwann cells and the periphery of the ganglion, suggesting that SPOCK1 is crucial to NMJ immunoreactivity, neurogenesis, tactile and proprioceptive functions and mutations relevant to developmental delay and microcephaly.36,41,42
In addition to its role in the neuronal system, SPOCK1 contributes other physiological functions that are consistent with its tissue distribution. SPOCK1 is a component of the ECM and a regulator of MMP-2. According to the regulation of activity and expression of MMP-2, SPOCK1 has a function of menarche, chronic inflammation and infection, and matrix turnover in cartilage.9,14,43,44 In addition, SPOCK1 can participate in other physiological and disease processes, such as embryonic development correlated with MafB, tissue remodeling under spaceflight conditions and coronary artery disease through the low-density lipoprotein-cholesterol pathway.31,45,46
The Role of SPOCK1 in EMT
EMT is a significant process associated with the weakness of cell-cell junctions, loss of cell polarity and epithelial characteristics, resulting in increased motor ability and invasion.47 EMT plays a significant role in oncogenic processes, including tumor invasion, intravasation, and micrometastasis formation.3,48-50 The EMT process promotes the acquisition of mesenchymal properties of epithelial cells and demonstrates an increase in cohesive intercellular adhesion and motor motile characteristics to migrate towards surrounding tissues, which facilitates subsequent metastases.51–53
In the EMT process, there are a range of ECM factors and membrane receptors involved in alternative signaling pathways that cooperate and compete to promote the connection between cancer cells and different extracellular clues. EMT is associated with not only loss of cell adhesion and elongation but also latent ECM invasion.47 The specific cell ligands in the microenvironment of tumors depend on ECM components that are correlated with cancer cell processes and promote the expression of survival pathways so that cells can escape drug therapy.54,55 The development of tumors is often related to the upregulation of different ECM proteins.56 The major molecular components of the ECM are proteoglycans and fibrillary proteins.37,45 Members of the SPARK family are important regulators of ECM assembly and are also involved in protease activities and cytokine signaling pathways.45 Similar to other members of the SPARK family, SPOCK1 is a regulator of the ECM and is crucial for the degradation and remodeling of the ECM, which is critical to ECM homeostasis as well as tumorigenesis.8 SPOCK1 has been indicated to regulate the activity and expression of MMPs, such as pro-MMP-2, membrane type (MT)1-MMP or MT3-MMP, MMP3 and MMP9, as a means to modulate ECM degradation and restoration.44,47
The progression of EMT is complicated by the regulation of mediators, such as growth factors and hormones.57 EMT is initiated by EMT-inducing transcriptional factors (EMT-TFs), including zinc finger proteins Snail, Slug (Snail2), Zeb1, Zeb2, and Twist.58 Stimulated by transforming growth factor-β (TGF-β), Snail controls transcriptional ZEB1 expression by cooperating with Twist and Ets1.58 The network of EMT-inducing transcriptional factors is activated and leads to temporal and spatial coordination between EMT biomarkers.57 In addition, multiple transcriptional factors and signaling pathways are positively correlated with EMT, including epithelial markers such as E-cadherin and mesenchymal markers such as vimentin.46,52,58 Among the signaling pathways, the signaling pathways of TGF-β, Wnt and Notch are considered to be key to the induction of the EMT phenotype.56
TGF-β, the primary enhancer of EMT, plays a crucial role in cancer viability and metastasis. At the transcription level, a range of transcription factors, such as zinc finger proteins Snail (SNAI1) and Slug (SNAI2), can be activated by TGF-β, subsequently leading to EMT.59 In addition, TGF-β can be connected to the Wnt/β-catenin signaling pathway and Notch signaling pathway to promote EMT.52 Wnt integrates with other signaling pathways, including RTKs and their downstream regulators MAPK, PI3K and PKB, to promote the progression of EMT.51 The expression of GSK3β, Snail, cyclin D1 and c-MYC is associated with Wnt/β-catenin.60 The Notch pathway is a supportive factor that must synergize with other carcinogenic pathways to lead to cell transformation and induce the EMT process. Notch target genes include Akt, mTOR, NF-κB, cyclin D1, c-myc, VEGF.61–63
Cells are changed to more scattered mesenchymal-like cells with the overexpression of SPOCK1. Several kinds of cancer cells undergo EMT changes according to the induction of N-cadherin and vimentin and the inhibition of ZO-1 and E-cadherin; these cell types include lung cancer, gallbladder cancer, gastric cancer, prostate cancer, esophageal cancer and glioma cells.2,16,50,59,60,64
It is suggested that several signaling pathways are associated with SPOCK1-mediated EMT. In addition, in the EMT process induced by SPOCK1, the interaction and cooperation between the expression of regulators and signaling pathways have been further discussed. There is a positive correlation between the expression of SPOCK1 and TGF-β in lung and breast cancer, which demonstrates that SPOCK1 is a positive downstream regulator of TGF-β.24,59 In addition, SPOCK1 expression is regulated through the Wnt/β-catenin pathway in NSCLC.65 SPOCK1-mediated EMT regulates proliferation and invasion via activation of the phosphoinositide 3-kinase (PI3K)/Akt signaling pathway in gallbladder cancer, colorectal cancer, pancreatic ductal adenocarcinoma and glioma.2,60,66,67 It is worth noting that invasion and metastasis are regulated by SPOCK1 through Slug-mediated EMT in gastric cancer (Figure 2).16
 |
Figure 2 The signaling pathways of SPOCK1 in EMT. SPOCK1 is a downstream regulator of TGF-β. EMT-inducing transcriptional factors Snail and Slug are regulated by TGF-β/SPOCK1 through PI3K/Akt and Wnt/β-catenin signaling pathways which cause regulation of epithelial markers E-cadherin, ZO-1 and mesenchymal markers vimentin, N-cadherin. Abbreviations: TGF-β, transforming growth factor-β; PI3K, phosphoinositide 3-kinase; Akt, v-akt murine thymoma viral oncogene homolog; Wnt, wingless/integrated. |
Functions of SPOCK1 in Cancer Progression
In addition to its physiological functions, SPOCK1 is a critical regulator in cancer progression. Accumulating evidence has indicated a role for SPOCK1 in many kinds of cancers, such as gallbladder cancer, prostate cancer, breast cancer, urothelial carcinomas, colorectal cancer, glioblastomas, ovarian cancer, esophageal squamous cell carcinoma, lung cancer and hepatocellular carcinoma (HCC) (Table 2).2,8,24,27,30,51,54,55,59,64,68-70 The importance of SPOCK1 in cancer proliferation, apoptosis, migration and drug resistance has been reported.15
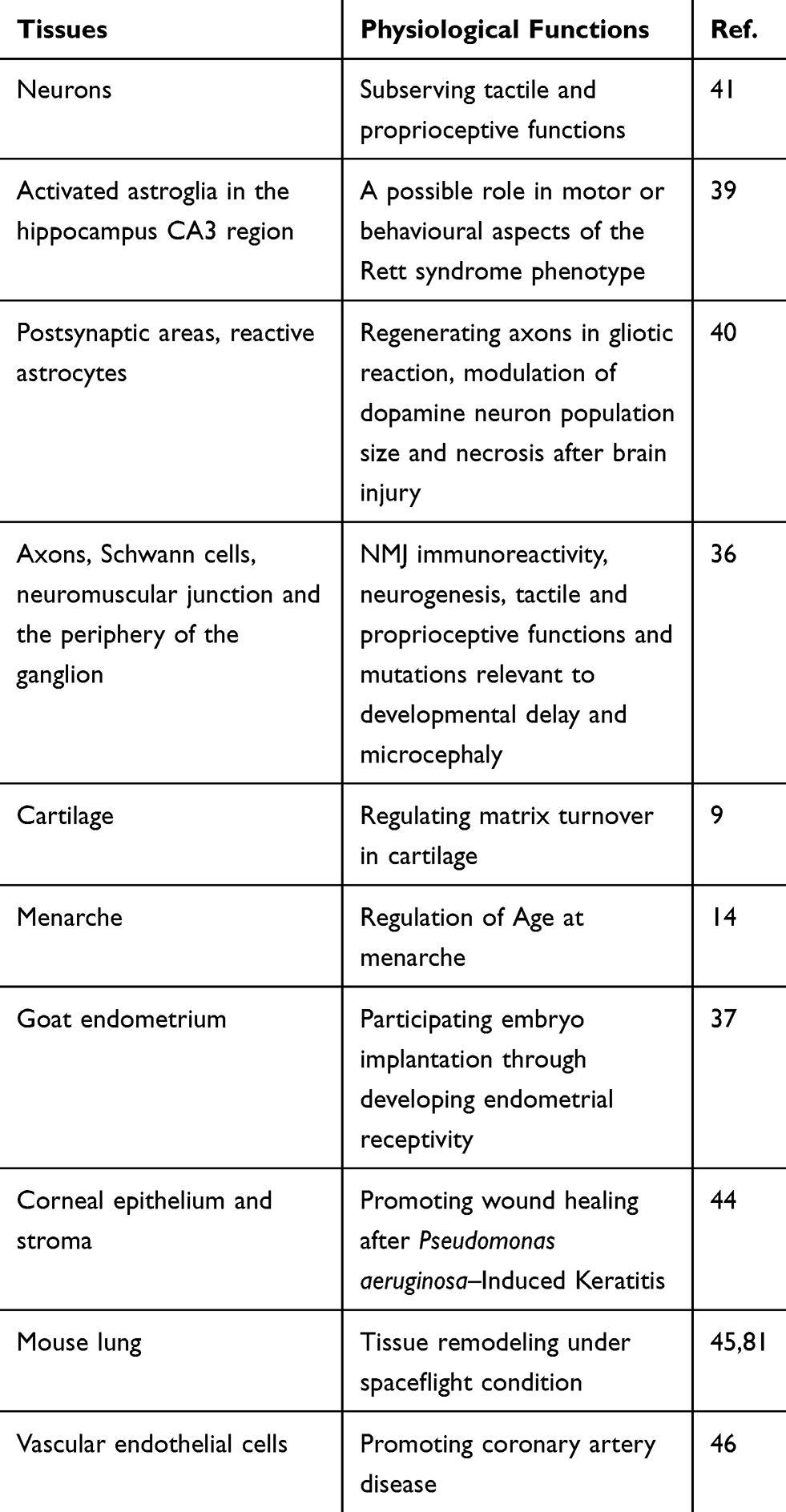 |
Table 1 The Physiological Functions |
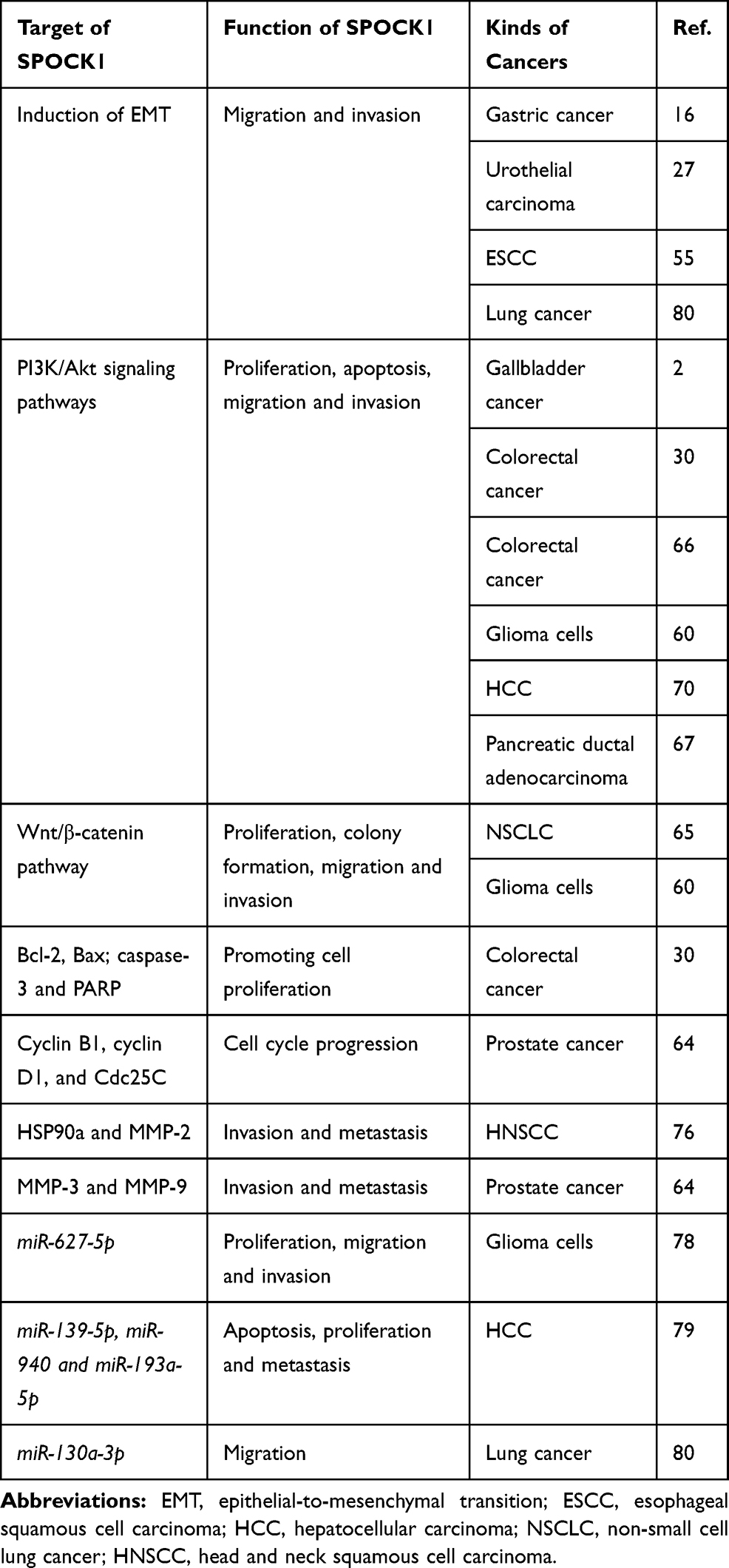 |
Table 2 The Mechanisms of SPOCK1 in Cancer Progression |
It is disputed whether SPOCK1 is a regulator of cancer cell proliferation. Some studies demonstrated that SPOCK1 had no effect on gastric cancer cell growth in vivo and in vitro.16 Other findings showed that SPOCK1 was essential in non-small-cell lung cancer (NSCLC) and osteosarcoma via the Wnt/β-catenin and mTOR-S6K signaling pathways, respectively.65,71 In addition, the enhancement of tumor cell survival by SPOCK1 may be attributed to its inhibition of apoptosis and regulation of cell cycle progression in cancers, such as colorectal cancer, prostate cancer and gallbladder cancer.2,30,64,70 The cell cycle regulators Cdc25C, cyclin B1 and cyclin D1 are regulated by SPOCK1 in colorectal cancer cell lines SW480 and HT29.30 SPOCK1 regulates G0/G1-phase cell cycle arrest and apoptosis in prostate cancer.64,72
SPOCK1 inhibits apoptosis through activation of the PI3K/Akt signaling pathway and subsequently inhibits the cyt c/caspase-9/caspase-3 pathway, which leads to upregulation of Bax and downregulation of Bcl-2.51,70,72
It is noteworthy that SPOCK1 also has effects on drug resistance to chemotherapeutics in cancer cells. SPOCK1 mediates temozolomide (TMZ) resistance in glioblastoma multiforme (GBM) cells and lapatinib resistance in HER2-positive gastric cancer via the Akt and Wnt/β-catenin signaling pathways, respectively.47,54 SPOCK1 is associated with osimertinib resistance in lung cancer, which is correlated with TGF-β1 and MMP11.73
SPOCK1 plays a critical role in invasion in many cancer types, such as gallbladder cancer, esophageal squamous cell carcinoma and colorectal cancer.2,51,66 SPOCK1 interferes with cancer cells by affecting extracellular collagen, subsequently promoting the expansion of cancer cells.15 SPOCK1 is involved in prostate cancer metastasis via the SPOCK1-Akt-Snail/Slug signaling pathway.74
Endothelial protein C receptor (EPCR), plakophilin 1 (PKP1) and chromodomain helicase/adenosine triphosphatase DNA binding protein 1-like (CHD1L) are upstream factors of SPOCK1.8,29,70 The high level of EPCR-SPOCK1 expression is associated with 3D growth in vitro, which is consistent with poor clinical outcome in breast cancer patients.29 CHD1L binds directly to SPOCK1 and activates transcription, blocks apoptosis and induces invasion of HCC cells through the Akt signaling pathway.70 SPOCK1 is also a target of miRNAs, including miR-129-5p, miR-150-3p, and miR-150-5p, in gastric cancer (GC) cells and head and neck squamous cell carcinoma (HNSCC). The miRNA/SPOCK1 axis is crucial for cancer proliferation, migration and invasion.75,76
Potential Applications of SPOCK1
Because of its high expression in cancer patients, SPOCK1 might be a prognostic marker in cancers. The expression level of SPOCK1 is associated with overall survival in many cancers, such as urothelial carcinoma, pancreatic ductal adenocarcinoma, prostate cancer, gastric cancer, breast cancer, lung adenocarcinoma and lung squamous carcinoma.16,24,27,67,73,74 High expression of SPOCK1 is consistent with shorter overall survival time, which indicates that SPOCK1 is associated with poor prognosis in univariate Log rank tests and in the multivariate analysis. There is a link between cancer stage and the expression of SPOCK1. SPOCK1 expression is higher in advanced stages than earlier stages in prostate cancer.69 Patients with upper tract urothelial carcinoma and poor disease-specific survival (DSS) had high SPOCK1 expression.27 HNSCC patients with poor prognoses had high SPOCK1 expression in TCGA database analyses.76 The expression of SPOCK1 in gallbladder cancer tissues is higher than that in their nontumor counterparts.2 SPOCK1 in lung cancer is significantly upregulated in metastatic tissues compared to nonmetastatic tissues.59
Several inhibitors have been identified, indicating that SPOCK1 may be a potential therapeutic target. Apigenin (20–80 μM), 4ʹ,5,7-trihydroxyflavone, can inhibit prostate cancer cell proliferation, migration, and invasion in vivo and in vitro.74 The expression of SPOCK1 is downregulated by 100 μM nitroglycerin in human monocytic leukemia cells to alter atherosclerotic plaque stability.77 These results indicated that nitroglycerin could be a candidate SPOCK1 inhibitor. Several endogenous inhibitors have also been identified. The microRNAs mentioned above (miR-130a-3p, miR-129-5p, miR-150-3p, miR-150-5p, miR-627-5p, miR-139-5p, miR-940 and miR-193a-5p) can regulate cancer progression by downregulating the expression of SPOCK1.75,76,78-80
Taken together, these results indicate that SPOCK1 could be a potential therapeutic and prognostic target.
Conclusion
SPOCK1 is a proteoglycan with systemic physiological functions. The anticancer effects have been investigated widely. SPOCK1 plays an important role in cancer proliferation and chemotherapy resistance and especially participates in cancer cell invasion and metastasis via EMT regulation. Patients with low SPOCK1 expression have increased life expectancy compared to those with high expression.59 The connection between the expression of SPOCK1 and poor prognosis makes SPOCK1 an effective prognostic factor in cancer. It is of clinical significance that the expression of SPOCK1 in metastatic tumor tissues is significantly higher than that in nonmetastatic tumor tissues.59 It is worth noting that multiple signaling pathways of SPOCK1 are involved in tumorigenesis and development. SPOCK1 may be a critical therapeutic target in cancer. Above all, SPOCK1 may be an attractive prognostic biomarker and therapeutic target in cancer treatment.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
2. Shu YJ, Weng H, Ye YY, et al. SPOCK1 as a potential cancer prognostic marker promotes the proliferation and metastasis of gallbladder cancer cells by activating the PI3K/AKT pathway. Mol Cancer. 2015;14:12. doi:10.1186/s12943-014-0276-y
3. Suarez-Carmona M, Lesage J, Cataldo D, Gilles C. EMT and inflammation: inseparable actors of cancer progression. Mol Oncol. 2017;11(7):805–823. doi:10.1002/1878-0261.12095
4. Insua-Rodríguez J, Oskarsson T. The extracellular matrix in breast cancer. Adv Drug Deliv Rev. 2016;97:41–55. doi:10.1016/j.addr.2015.12.017
5. Egeblad M, Rasch MG, Weaver VM. Dynamic interplay between the collagen scaffold and tumor evolution. Curr Opin Cell Biol. 2010;22(5):697–706. doi:10.1016/j.ceb.2010.08.015
6. Mott JD, Werb Z. Regulation of matrix biology by matrix metalloproteinases. Curr Opin Cell Biol. 2004;16(5):558–564. doi:10.1016/j.ceb.2004.07.010
7. Bode W, Huber R. Proteinase-protein inhibitor interaction. Biomed Biochim Acta. 1991;50(4–6):437–446.
8. Yang C, Fischer-Kešo R, Schlechter T, Ströbel P, Marx A, Hofmann HI. Plakophilin 1-deficient cells upregulate SPOCK1: implications for prostate cancer progression. Tumor Biol. 2015;36(12):9567–9577. doi:10.1007/s13277-015-3628-3
9. Hausser HJ, Decking R, Brenner RE. Testican-1, an inhibitor of pro-MMP-2 activation, is expressed in cartilage. Osteoarthritis Cartilage. 2004;12(11):870–877. doi:10.1016/j.joca.2004.07.008
10. Marr HS, BaSalamah MA, Edgell CJ. Endothelial cell expression of testican mRNA. Endothelium. 1997;5(3):209–219. doi:10.3109/10623329709053399
11. Genini M, Schwalbe P, Scholl FA, Schafer BW. Isolation of genes differentially expressed in human primary myoblasts and embryonal rhabdomyosarcoma. Int J Cancer. 1996;66(4):571–577. doi:10.1002/(SICI)1097-0215(19960516)66:4<571::AID-IJC24>3.0.CO;2-9
12. Nitsche EM, Moquin A, Adams PS, et al. Differential display RT PCR of total RNA from human foreskin fibroblasts for investigation of androgen-dependent gene expression. Am J Med Genet. 1996;63(1):231–238. doi:10.1002/(SICI)1096-8628(19960503)63:1<231::AID-AJMG40>3.0.CO;2-M
13. Delgado IJ, Kim DS, Thatcher KN, LaSalle JM, Van Den Veyver IB. Expression profiling of clonal lymphocyte cell cultures from Rett syndrome patients. BMC Med Genet. 2006;7:61. doi:10.1186/1471-2350-7-61
14. Liu YZ, Guo YF, Wang L, et al. Genome-wide association analyses identify SPOCK as a key novel gene underlying age at menarche. PLoS Genet. 2009;5(3):e1000420. doi:10.1371/journal.pgen.1000420
15. Veenstra VL, Damhofer H, Waasdorp C, et al. Stromal SPOCK1 supports invasive pancreatic cancer growth. Mol Oncol. 2017;11(8):1050–1064. doi:10.1002/1878-0261.12073
16. Chen D, Zhou H, Liu G, Zhao Y, Cao G, Liu Q. SPOCK1 promotes the invasion and metastasis of gastric cancer through Slug-induced epithelial-mesenchymal transition. J Cell Mol Med. 2018;22(2):797–807. doi:10.1111/jcmm.13602
17. Bradshaw AD. Diverse biological functions of the SPARC family of proteins. Int J Biochem Cell Biol. 2012;44(3):480–488. doi:10.1016/j.biocel.2011.12.021
18. Hartmann U, Hülsmann H, Seul J, et al. Testican-3: a brain-specific proteoglycan member of the BM-40/SPARC/osteonectin family. J Neurochem. 2013;125(3):399–409. doi:10.1111/jnc.12212
19. Schnepp A, Komp Lindgren P, Hülsmann H, Kröger S, Paulsson M, Hartmann U. Mouse testican-2. Expression, glycosylation, and effects on neurite outgrowth. J Biol Chem. 2005;280(12):11274–11280. doi:10.1074/jbc.M414276200
20. Yamamoto A, Uchiyama K, Nara T, et al. Structural abnormalities of corpus callosum and cortical axonal tracts accompanied by decreased anxiety-like behavior and lowered sociability in spock3-mutant mice. Dev Neurosci. 2014;36(5):381–395. doi:10.1159/000363101
21. Charbonnier F, Périn JP, Mattei MG, et al. Genomic organization of the human SPOCK gene and its chromosomal localization to 5q31. Genomics. 1998;48(3):377–380. doi:10.1006/geno.1997.5199
22. Lenarcic B, Bevec T. Thyropins – new structurally related proteinase inhibitors. Biol Chem. 1998;379(2):105–111.
23. Bocock JP, Edgell CJ, Marr HS, Erickson AH. Human proteoglycan testican-1 inhibits the lysosomal cysteine protease cathepsin L. Eur J Biochem. 2003;270(19):4008–4015. doi:10.1046/j.1432-1033.2003.03789.x
24. Fan LC, Jeng YM, Lu YT, Lien HC. SPOCK1 is a novel transforming growth factor-β-induced myoepithelial marker that enhances invasion and correlates with poor prognosis in breast cancer. PLoS One. 2016;11(9):e0162933. doi:10.1371/journal.pone.0162933
25. Röll S, Seul J, Paulsson M, Hartmann U. Testican-1 is dispensable for mouse development. Matrix Biol. 2006;25(6):373–381. doi:10.1016/j.matbio.2006.05.004
26. Nakada M, Yamada A, Takino T, et al. Suppression of membrane type 1 matrix metalloproteinase (MMP) -mediated MMP-2 activation and tumor invasion by testican 3 and its splicing variant gene product, N-Tes. Cancer Res. 2001;61(24):8896–8902.
27. Ma LJ, Wu WJ, Wang YH, et al. SPOCK1 overexpression confers a poor prognosis in urothelial carcinoma. J Cancer. 2016;7(4):467–476. doi:10.7150/jca.13625
28. Greene LJ, DiCarol JJ, Sussman AJ, Bartelt DC. Two trypsin inhibitors from porcine pancreatic juice. J Biol Chem. 1968;243:1804–1815.
29. Perurena N, Zandueta C, Martínez-Canarias S, et al. EPCR promotes breast cancer progression by altering SPOCK1/testican 1-mediated 3D growth. J Hematol Oncol. 2017;10(1):23. doi:10.1186/s13045-017-0399-x
30. Zhang J, Zhi X, Shi S, et al. SPOCK1 is up-regulated and promotes tumor growth via the PI3K/AKT signaling pathway in colorectal cancer. Biochem Biophys Res Commun. 2017;482(4):870–876. doi:10.1016/j.bbrc.2016.11.126
31. Zhang Y, Chen Q, Ross AC. Retinoic acid and tumor necrosis factor-α induced monocytic cell gene expression is regulated in part by induction of transcription factor MafB. Exp Cell Res. 2012;318(18):2407–2416. doi:10.1016/j.yexcr.2012.07.011
32. Bonnet F, Perin JP, Maillet P, Jolles P, Alliel PM. Characterization of a human seminal plasma glycosaminoglycanbearing polypeptide. Biochem J. 1992;288(Pt 2):565–569. doi:10.1042/bj2880565
33. Alliel PM, Pedrin J-P, Jolles P, Bonnet FJ. Testican, a multidomain testicular proteoglycan resemblingmodulators of cell social behaviour. Eur J Biochem. 1993;214(1):347–350. doi:10.1111/j.1432-1033.1993.tb17930.x
34. Bonnet F, Périn JP, Charbonnier F, et al. Structure and cellular distribution of mouse brain testican. J Biol Chem. 1996;271(8):4373–4380. doi:10.1074/jbc.271.8.4373
35. Marr HS, Basalamah MA, Bouldin TW, Duncan AW, Edgell CJ. Distribution of testican expression in human brain. Cell Tissue Res. 2000;302:139–144. doi:10.1007/s004410000277
36. Cifuentes-Diaz C, Alliel PM, Charbonnier F, et al. Regulated expression of the proteoglycan SPOCK in the neuromuscular system. Mech Dev. 2000;94(1–2):277–282. doi:10.1016/S0925-4773(00)00285-9
37. Zhang L, An XP, Liu XR, et al. Characterization of the transcriptional complexity of the receptive and pre-receptive endometria of dairy goats. Sci Rep. 2015;5:14244. doi:10.1038/srep14244
38. Colin C, Baeza N, Bartoli C, et al. Identification of genes differentially expressed in glioblastoma versus pilocytic astrocytoma using suppression subtractive hybridization. Oncogene. 2006;25(19):2818–2826. doi:10.1038/sj.onc.1209305
39. Vadasz C, Smiley JF, Figarsky K, et al. Mesencephalic dopamine neuron number and tyrosine hydroxylase content: genetic control and candidate genes. Neuroscience. 2007;149(3):561–572. doi:10.1016/j.neuroscience.2007.06.049
40. Iseki K, Hagino S, Zhang Y, et al. Altered expression pattern of testican-1 mRNA after brain injury. Biomed Res. 2011;32(6):373–378. doi:10.2220/biomedres.32.373
41. Charbonnier F, Chanoine C, Cifuentes-Diaz C, et al. Expression of the proteoglycan SPOCK during mouse embryo development. Mech Dev. 2000;90(2):317–321. doi:10.1016/S0925-4773(99)00255-5
42. Dhamija R, Graham JM, Smaoui N, Thorland E, Kirmani S. Novel de novo SPOCK1 mutation in a proband with developmental delay, microcephaly and agenesis of corpus callosum. Eur J Med Genet. 2014;57(4):181–184. doi:10.1016/j.ejmg.2014.02.009
43. Wlazlinski A, Engers R, Hoffmann MJ, et al. Downregulation of several fibulin genes in prostate cancer. Prostate. 2007;67(16):1770–1780. doi:10.1002/pros.20667
44. Berger EA, McClellan SA, Barrett RP, Hazlett LD. Testican-1 promotes resistance against pseudomonas aeruginosa–induced keratitis through regulation of MMP-2 expression and activation. Invest Ophthalmol Vis Sci. 2011;52(8):5339–5346. doi:10.1167/iovs.10-6920
45. Gridley DS, Mao XW, Tian J, et al. Genetic and apoptotic changes in lungs of mice flown on the STS-135 mission in space. In Vivo. 2015;29(4):423–433.
46. Chen F, Zhao X, Peng J, Bo L, Fan B, Ma D. Integrated microRNA-mRNA analysis of coronary artery disease. Mol Biol Rep. 2014;41(8):5505–5511. doi:10.1007/s11033-014-3426-9
47. Kim HP, Han SW, Song SH, et al. Testican-1-mediated epithelial–mesenchymal transition signaling confers acquired resistance to lapatinib in HER2-positive gastric cancer. Oncogene. 2014;33(25):3334–3341. doi:10.1038/onc.2013.285
48. Thiery JP, Acloque H, Huang RY, Nieto MA. Epithelial-mesenchymal transitions in development and disease. Cell. 2009;139(5):871–890. doi:10.1016/j.cell.2009.11.007
49. Thompson EW, Newgreen DF, Tarin D. Carcinoma invasion and metastasis: a role for epithelial-mesenchymal transition? Cancer Res. 2005;65(14):5991–5995. doi:10.1158/0008-5472.CAN-05-0616
50. Radisky DC. Epithelial-mesenchymal transition. J Cell Sci. 2005;118(Pt 19):4325–4326. doi:10.1242/jcs.02552
51. Song X, Han P, Liu J, et al. Up-regulation of SPOCK1 induces epithelial–mesenchymal transition and promotes migration and invasion in esophageal squamous cell carcinoma. J Mol Histol. 2015;46(4–5):347–356. doi:10.1007/s10735-015-9627-2
52. Basu S, Cheriyamundath S, Ben-Ze’ev A. Cell-cell adhesion: linking Wnt/β-catenin signaling with partial EMT and stemness traits in tumorigenesis. F1000Res. 2018;7:
53. Das V, Bhattacharya S, Chikkaputtaiah C, Hazra S, Pal M. The basics of epithelial-mesenchymal transition (EMT): a study from a structure, dynamics, and functional perspective. J Cell Physiol. 2019 ();234(9):14535–14555. doi:10.1002/jcp.28160
54. Yu F, Li G, Gao J, et al. SPOCK1 is upregulated in recurrent glioblastoma and contributes to metastasis and Temozolomide resistance. Cell Prolif. 2016;49(2):195–206. doi:10.1111/cpr.12241
55. Zhang LQ, Wang Y, Zhang L. Effects of shRNA mediated knockdown of SPOCK1 on ovarian cancer growth and metastasis. Cell Mol Biol (Noisy-Le-Grand). 2015;61(7):102–109.
56. Holle AW, Young JL, Spatz JP. In vitro cancer cell–ECM interactions inform in vivo cancer treatment. Adv Drug Deliv Rev. 2016;97:270–279. doi:10.1016/j.addr.2015.10.007
57. Stefania D, Vergara D. The many-faced program of epithelial-mesenchymal transition: a system biology-based view. Front Oncol. 2017;7:274. doi:10.3389/fonc.2017.00274
58. Simeone P, Trerotola M, Franck J, et al. The multiverse nature of epithelial to mesenchymal transition. Semin Cancer Biol. 2019;58:1–10. doi:10.1016/j.semcancer.2018.11.004
59. Miao L, Wang Y, Xia H, Yao C, Cai H, Song Y. SPOCK1 is a novel transforming growth factor-β target gene that regulates lung cancer cell epithelial-mesenchymal transition. Biochem Biophys Res Commun. 2013;440(4):792–797. doi:10.1016/j.bbrc.2013.10.024
60. Yang J, Yang Q, Yu J, Li X, Yu S, Zhang X. SPOCK1 promotes the proliferation, migration and invasion of glioma cells through PI3K/AKT and Wnt/β-catenin signaling pathways. Oncol Rep. 2016;35(6):3566–3576. doi:10.3892/or.2016.4757
61. Miele L, Osborne B. Arbiter of differentiation and death: notch signaling meets apoptosis. J Cell Physiol. 1999;181(3):393–409. doi:10.1002/(SICI)1097-4652(199912)181:3<393::AID-JCP3>3.0.CO;2-6
62. Miele L, Miao H, Nickoloff BJ. Notch signaling as a novel cancer therapeutic target. Curr Cancer Drug Targets. 2006;6(4):313–323. doi:10.2174/156800906777441771
63. Miele L. Notch signaling. Clin Cancer Res. 2006;12(4):1074–1079. doi:10.1158/1078-0432.CCR-05-2570
64. Chen Q, Yao YT, Xu H, et al. SPOCK1 promotes tumor growth and metastasis in human prostate cancer. Drug Des Devel Ther. 2016;10:2311–2321. doi:10.2147/DDDT.S91321
65. Wang T, Liu X, Tian Q, Liang T, Chang P. Reduced SPOCK1 expression inhibits non-small cell lung cancer cell proliferation and migration through Wnt/β-catenin signaling. Eur Rev Med Pharmacol Sci. 2018;22(3):637–644. doi:10.26355/eurrev_201802_14288
66. Zhao P, Guan HT, Dai ZJ, Ma YG, Liu XX, Wang XJ. Knockdown of SPOCK1 inhibits the proliferation and invasion in colorectal cancer cells by suppressing the PI3K/Akt pathway. Oncol Res. 2016;24(6):437–445. doi:10.3727/096504016X14685034103554
67. Li J, Ke J, Fang J, Chen JP. A potential prognostic marker and therapeutic target: SPOCK1 promotes the proliferation, metastasis, and apoptosis of pancreatic ductal adenocarcinoma cells. J Cell Biochem. 2020;121(1):743–754. doi:10.1002/jcb.29320
68 Kong L, Wu Q, Zhao L, Ye J, Li N, Yang H. Identification of messenger and long noncoding RNAs associated with gallbladder cancer via gene expression profile analysis. J Cell Biochem. 2019;120(12):19377–19387. doi:10.1002/jcb.28953
69. Chen ML, Ho CJ, Yeh CM, et al. High SPOCK1 Expression Is Associated with Advanced Stage, T Value, and Gleason Grade in Prostate Cancer. Medicina (Kaunas). 2019;55(7):
70. Li Y, Chen L, Chan TH, et al. SPOCK1 is regulated by CHD1L and blocks apoptosis and promotes HCC cell invasiveness and metastasis in mice. Gastroenterology. 2013;144(1):179–191.e4. doi:10.1053/j.gastro.2012.09.042
71. Wang Y, Wang W, Qiu E. SPOCK1 promotes the growth of Osteosarcoma cells through mTOR-S6K signaling pathway. Biomed Pharmacother. 2017;95:564–570. doi:10.1016/j.biopha.2017.08.116
72. Yan L, Sun K, Liu Y, Liang J, Cai K, Gui J. MiR-129-5p influences the progression of gastric cancer cells through interacting with SPOCK1. Tumour Biol. 2017;39(6):1010428317706916. doi:10.1177/1010428317706916
73. Gao Y, Yu M, Ma M, et al. SPOCK1 contributes to the third-generation EGFR tyrosine kinase inhibitors resistance in lung cancer. J Cell Biochem. 2019;120(8):12566–12573. doi:10.1002/jcb.28523
74. Chien MH, Lin YW, Wen YC, et al. Targeting the SPOCK1-snail/slug axis-mediated epithelial-to-mesenchymal transition by apigenin contributes to repression of prostate cancer metastasis. J Exp Clin Cancer Res. 2019;38(1):246. doi:10.1186/s13046-019-1247-3
75. Osako Y, Seki N, Koshizuka K, et al. Regulation of SPOCK1 by dual strands of pre-miR-150 inhibit cancer cell migration and invasion in esophageal squamous cell carcinoma. J Hum Genet. 2017;62(11):935–944. doi:10.1038/jhg.2017.69
76. Koshizuka K, Hanazawa T, Kikkawa N, et al. Antitumor miR-150-5p and miR-150-3p inhibit cancer cell aggressiveness by targeting SPOCK1 in head and neck squamous cell carcinoma. Auris Nasus Larynx. 2018;45(4):854–865. doi:10.1016/j.anl.2017.11.019
77. Krishnatry AS, Brazeau DA, Fung HL. Broad regulation of matrix and adhesion molecules in THP-1 human macrophages by nitroglycerin. Nitric Oxide. 2010;22(1):11–17. doi:10.1016/j.niox.2009.10.004
78. Fan Z, Zheng J, Xue Y, et al. NR2C2-uORF targeting UCA1-miR-627-5p- NR2C2 feedback loop to regulate the malignant behaviors of glioma cells. Cell Death Dis. 2018;9(12):1165. doi:10.1038/s41419-018-1149-x
79. Li P, Xiao Z, Luo J, Zhang Y, Lin L. MiR‐139‐5p, miR‐940 and miR‐193a‐5p inhibit the growth of hepatocellular carcinoma by targeting SPOCK1. J Cell Mol Med. 2019;23(4):2475–2488. doi:10.1111/jcmm.14121
80. Yu XF, Wang J, Ouyang N, et al. The role of miR-130a-3p and SPOCK1 in tobacco exposed bronchial epithelial BEAS-2B transformed cells: comparison to A549 and H1299 lung cancer cell lines. J Toxicol Environ Health A. 2019;82(15):862–869. doi:10.1080/15287394.2019.1664479
81. Tian J, Pecaut MJ, Slater JM, Gridley DS. Spaceflight modulates expression of extracellular matrix, adhesion, and profibrotic molecules in mouse lung. J Appl Physiol (1985). 2010;108(1):162–171. doi:10.1152/japplphysiol.00730.2009
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.