Back to Journals » Journal of Inflammation Research » Volume 16
Sparcl1 and Atherosclerosis
Authors Cheng X ![]() , Chen X, Zhang M, Wan Y, Ge S
, Chen X, Zhang M, Wan Y, Ge S ![]() , Cheng X
, Cheng X ![]()
Received 3 February 2023
Accepted for publication 3 May 2023
Published 17 May 2023 Volume 2023:16 Pages 2121—2127
DOI https://doi.org/10.2147/JIR.S406907
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tara Strutt
Xu Cheng,1,2 Xinyan Chen,2,3 Min Zhang,3 Yufeng Wan,4 Shenglin Ge,1 Xiaowen Cheng2,3,5
1Department of Cardiovascular Surgery, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, 230022, People’s Republic of China; 2Laboratory of Molecular Biology and Department of Biochemistry, Anhui Medical University, Hefei, Anhui, 230022, People’s Republic of China; 3Department of Clinical Laboratory, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, 230022, People’s Republic of China; 4Department of Otolaryngology-Head Neck Surgery, the Affiliated Chaohu Hospital of Anhui Medical University, Hefei, Anhui, 238001, People’s Republic of China; 5Anhui Province Key Laboratory of Translational Cancer Research, Bengbu Medical College, Bengbu, Anhui, 233030, People’s Republic of China
Correspondence: Xiaowen Cheng, Department of Clinical Laboratory, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, 230022, People’s Republic of China, Email [email protected] Shenglin Ge, Department of Cardiovascular Surgery, the First Affiliated Hospital of Anhui Medical University, Hefei, Anhui, 230022, People’s Republic of China, Email [email protected]
Abstract: Atherosclerosis and its complications constitute some of the major diseases affecting humans worldwide. A core component of atherogenesis is endothelial cell damage and dysfunction, which also includes factors such as adhesion and proliferation of various cells. Multiple studies have shown that atherosclerosis and cancer share a common pathophysiological process and exhibit a degree of similarity. Sparcl-1 is a cysteine-rich secretory stromal cell protein present in the extracellular matrix and belongs to the Sparc family of proteins. Its role in tumor development has been widely investigated; however, its role in cardiovascular diseases has rarely been studied. Sparcl-1 is considered an oncogene correlated with the regulation of cell adhesion, migration, and proliferation and is also related to blood vessel integrity. In this review, the potential link between Sparcl-1 and atherosclerosis development is investigated, and recommendations on future research on the role of Sparcl-1 in atherogenesis are provided.
Keywords: Sparcl-1, atherogenesis, cancer
Introduction
Atherosclerosis and its complications constitute some of the major causes of cardiovascular diseases and account for a few of the most important diseases threatening the health of humans.1 Atherosclerosis is a chronic inflammatory state caused by the uncontrolled proliferation of vascular smooth muscle cells, endothelial cells, macrophages, and other cells.2,3 Endothelial dysfunction is thought to be an initiating and aggravating factor in atherosclerosis.4 Endothelial cell dysfunction and hemodynamics are closely related to the activation of various inflammatory pathways and the reduced nitric oxide (NO) production.5 When pro-inflammatory factors activate the endothelium, it expresses some adhesion molecules that attract lymphocytes and monocytes, such as intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1), and makes them infiltrate into the intima of the arterial wall.6 Subsequently, monocytes transform into macrophages in the endothelium, phagocytose oxidized low-density lipoprotein (ox-LDL), impede the clearance of cholesterol and ox-LDL, and transform into foam cells.7 Finally, migration and proliferation of vascular smooth muscle cells (VSMC) promote atheromatous plaque formation and are involved in fibrous cap formation.8 In summary, the initiation of atherosclerosis can be roughly divided into the following steps: (1) endothelial dysfunction and accumulation of low-density lipoproteins (LDL),9,10 (2) activation of endothelial cells to induce monocyte recruitment and foam cell formation,11 and (3) focal accumulation of smooth muscle cells.12 Therefore, endothelial cell dysfunction contributes to the development of atherosclerosis. In recent years, much of the atherosclerosis research has focused on the inflammatory response. Targeting inflammatory pathways could represent an attractive new route for the treatment of atherosclerotic diseases.13,14
Some researchers have suggested that the pathophysiological processes of atherosclerosis and cancer are similar and have viewed atherosclerosis as a cancer of the blood vessel wall.1,15–17 Sparcl1 has been a major player in cancer development studies in recent years and has been mentioned in studies related to anti-angiogenicity.18 Similarly, it is also mentioned in non-alcoholic inflammatory diseases of the liver.19 However, few studies have shown a direct relationship between Sparcl1 and atherosclerosis. It has been demonstrated that Sparcl1 is indeed involved in atherogenesis and is expressed in atherosclerotic tissues,20,21 however, the specific process of Sparcl1 involvement in atherosclerosis and its particular role has not been investigated. However, as mentioned above, endothelial dysfunction is an important initiating factor of atherosclerosis,22 and Sparcl1 has a role in stabilizing the vascular endothelium. Therefore, Sparcl1 may have an undiscovered role in atherosclerosis development. Sparcl1, also known as Hevin, SC1, and ECM2, belongs to the SPARC family, an extracellular matrix protein family involved in the regulation of cell adhesion, migration, and proliferation.23 It was first isolated from a rat CNS expression library by Jonhtson et al in 1990 and was named SC1.24 Subsequently, Schraml et al25 and Girard et al26 cloned the gene named MAST9 and hevin from the endothelial cells of non-small cell lung cancer and from high endothelial small veins of human tonsillar lymphoid tissue, respectively. Sparcl1 is localized in chromosome 4q22-25 and spans approximately 47 kb of the genome.27 It is highly homologous to the Sparc family and composed of three structural domains: (1) N-terminal acidic structural domain, (2) follicle inhibitor-like structural domains, and (3) extracellular Ca2+ binding domain.26 It is widely expressed in various tissues in the human body, but its expression is downregulated in some cancers, such as prostate cancer,28 non-small cell carcinoma,29 ovarian cancer,30 and colon cancer,31 suggesting that Sparcl1 acts as a tumor-suppressor gene. This review aims to explore the potential link between Sparcl1 and atherosclerosis and to provide possible future research directions. This study aims to explore the potential association between Sparcl1 and atherosclerosis and the possibility of Sparcl1 as a new therapeutic target for atherosclerosis in the future.
Structure and Signaling Pathway of Sparcl-1
Gene Localization and Structure of Sparcl-1
As mentioned earlier, Sparcl1 is an extracellular matrix protein involved in the regulation of cell adhesion, proliferation and migration. Sparcl1 is localized to human chromosome 4q22-25, which is also the region missing in many cancers. Its full gene length is approximately 47 kb and consists of 11 exons and 10 introns. The length of these exons varies from 65 to 1017 kb, whereas the length of introns varies from about 400bp to 21.5kb.27 In terms of structure, it has high homology with the SPARC family, and they can all be divided into three parts: (1) the N-terminal acidic domain; (2) the follicle repressor-like domain (FS domain, aa52-132); and (3) the extracellular calcium binding domain (EC domain, aa133-285). Furthermore, a characteristic segment of the EC structural domain is present in each member of the SPARC family. This segment consists of an amphipathic α-helix and two EF-hand motifs.32 In addition, Brekken & Sage reported in 2000 that the EC domain can stabilize calcium binding through the interaction of several specific bases with the FS domain, and that some of these peptides can inhibit cell proliferation and reverse adhesion formation.33 This sets the stage for the next prediction of the role of Sparcl-1. Sparcl-1 is one of the four types of SPARC family members based on the homology of the EC structural domain. Sparcl1 shares 56% homology with the FS region of SPARC and 63% homology with EC (Figure 1).34
 |
Figure 1 The homology of SPARC with Sparc-1 is clearly seen by the overlap of the relevant colors in the figure. The gene schematic of SPARC is shown at the top of the figure, and the gene schematic of Sparcl1 is shown at the bottom. The number of corresponding amino acids is shown below the structural domain. |
Sparcl-1-Associated Signaling Pathways
Currently, there are studies on Sparcl1 that are still predominantly oncology-based. For instance, Hui Ye and his colleagues found, in 2017, that Sparcl-1 overexpression would considerably reduce the phosphorylation of JNK, ERK and p38 proteins in kidney cancer cells. This suggests that Sparcl1 has the potential to inhibit the proliferation and migration of kidney cancer cells by inactivating the JNK/ERK/p38 pathway.35 In a 2018 study on ovarian cancer, by overexpressing Sparcl1 as well as suppressing the expression, researchers found that overexpression of Sparcl1 extensively inhibited the mitogen-activated protein kinase (MEK)/extracellular signal-related kinase (ERK) signaling pathway. It is also hypothesized that Sparcl1 may inhibit the proliferation and migration of ovarian cancer cells by downregulating the MEK/ERK signaling pathway.30 In addition, Yuxin Wang demonstrated, in 2020, that Sparcl1 can also regulate ITGB1-mediated signaling pathways by affecting integrin β1 (ITGB1), thereby affecting cell migration and differentiation.36 Furthermore, it has been shown that Sparcl1 can also affect cell differentiation through the BMP/TGF-βpathway (Figure 2).37 These results suggest that the biological actions of Sparcl-1 are mediated through a wide variety of signaling pathways.
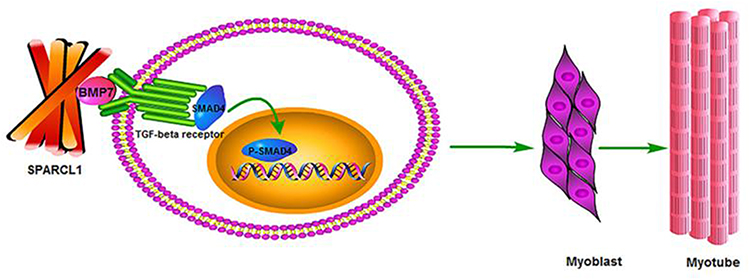 |
Figure 2 Sparcl1 can affect cell differentiation through the BMP/TGF-β pathway. Notes: Reproduced from Wang Y, Liu S, Yan Y, Li S, Tong H. SPARCL1 promotes C2C12 cell differentiation via BMP7-mediated BMP/TGF-β cell signaling pathway. Cell Death Dis. 2019;10:852. Creative Commons.37 |
Role of Sparcl1 in Cancer Development in Relation to Atherogenesis
Effects of Sparcl-1 on Blood Vessels
Regensburger and colleagues studied the role of Sparcl1 related to colorectal disease and found that Sparcl1 has an anti-angiogenic effect.18 They compared the vascular integrity of Sparcl1 knockout mice and wild-type mice and revealed that the vascular integrity of Sparcl1 knockout mice is markedly lower than that of wild-type mice. These results suggested that Sparcl1 expression is positively correlated with vascular integrity. Their follow-up experiment compared the permeability of WT and Sparcl1 knockout mice and concluded that vascular permeability considerably increases in the absence of Sparcl1.18 These results have constructive implications for the involvement of Sparcl1 in tumor metastasis as well as lipid infiltration in atherosclerosis.
Sparcl1 is considered a marker of tumor vascular endothelial cell quiescence, ie, Sparcl1 is highly expressed in quiescent vascular endothelial cells and low in activated vascular endothelial cells,22 suggesting that Sparcl1 may be associated with maintaining vessel wall stability. Subsequent experiments have suggested that a high Sparcl1 expression is associated with blood vessel maturation and contributes to mature vessel stability; it inhibits endothelial cell proliferation and migration.
Thus, the metastatic process of tumors and the lipid infiltration process of atherosclerosis involve changes in vascular permeability. Therefore, Sparcl1 is potentially linked to atherosclerosis.
Role of Sparcl-1 in Cell Proliferation and Migration
Funk and colleagues provided evidence, in as early as 1991 that Sparcl1 may negatively regulate the proliferative effects of bovine aortic endothelial cells by inhibiting the progression of the S phase.38 Subsequently, Claeskens et al constructed a Hela 3S tet-off cell model and used FACS to detect different phases of the cell cycle.39 They demonstrated that stably expressed Sparcl1 can reduce the proliferation rate of cells by inhibiting the transition from G1 to S phase or by prolonging the G1 phase.
A recent study on ovarian cancer has shown a marked increase in the proliferation rate of SKOV-3 cells after Sparcl-1 knockdown; follow-up experiments have demonstrated that Sparcl11 inhibits SKOV-3 cell migration. In addition, Sparcl1 inhibits the proliferation and migration of SKOV-3 at least in part by inhibiting the MEK/ERK pathway.30 Zhang et al also found that Hela cells overexpressing Sparcl-1 have a weaker proliferation and migration ability.40 Sparcl1 can impede the migration and invasion of trophoblast cells by altering the expression of related molecules in embryonic trophoblast cells.41 As mentioned above, Sparcl1 has an inhibitory effect on cell proliferation; however, there have been no studies on the effect of Sparcl1 on vascular smooth muscle cell (VSMC) proliferation. The role of smooth muscle cells in the development of atherosclerosis has changed over the years; however, with the proliferation of smooth muscle cells previously thought to be a driver of atherosclerosis, the most recent studies suggest that the proliferation of vascular smooth muscle appears to be a reparative factor.42 Therefore, the effect of Sparcl1 interaction with vascular smooth muscle on atherosclerosis needs further investigated.
Other Related Mechanisms
Girard et al isolated Sparcl1 from tonsillar lymphoid tissue in 1995 and named it Hevin,42 they suggested that Sparcl1 has 62% homology with the anti-adhesive extracellular matrix protein SPARC.26 Therefore, Sparcl1 likely has an anti-adhesive function. It also exhibits inhibitory effects on cell motility and cell adhesion in osteosarcoma.43 It mediates the activation of WNT/β protein signaling to increase chemokine ligand 5 (CCL-5) production in human osteosarcoma cells, thereby promoting macrophage recruitment. These functions of Sparcl1 have also been further confirmed in atherosclerosis. In addition, Sparcl1 inhibits adipogenesis by reducing the lipogenic capacity of preadipocytes.44 Moreover, Cheng Xiao and Hai Guojin et al found that overexpression of Sparcl1 significantly inhibited lipid droplet accumulation and triglyceride content by increasing Wnt10b, Fzd8, IL6, and β-linked protein and suppressing PPARγ, C/EBPα, LPL, and IGF1 gene expression during the preadipocyte differentiation stage. Disruptive expression of Sparcl1 increases apoptosis and viability of sheep preadipocytes during the proliferative phase.45 As early as 2009, a study confirmed that the SPARC family is associated with the amount of body fat and may be linked to metabolic dysregulation of obesity.46 In a study by Bin Liu et al in 2021, Sparcl1 was found to be upregulated in white adipose tissue markers in nonalcoholic steatohepatitis (NASH) mice and indicated that Sparcl1 might be a causative factor in NASH.19 However, the specific role of Sparcl1 in human adipose tissue requires further investigation.
Link Between Atherosclerosis and Cancer
Atherosclerosis and cancer are two chronic diseases and leading causes of human death worldwide.16 Studies have speculated the existence of a link between atherosclerosis and cancer, and some researchers have even suggested that atherosclerosis is a cancer of the blood vessel wall.1,16 The common predisposing factors for atherosclerosis and cancer are age, smoking, and unhealthy lifestyles.16 Atherosclerosis and cancer also share similar pathophysiological processes. For example, (1) atherosclerosis and cancer both have uncontrolled cell proliferation.17 (2) Both diseases have changes in cell adhesion molecules associated with the formation of atheromatous plaques and the migration and invasion of tumor cells, respectively.16 Both conditions (3) have corresponding inflammatory manifestations1, (4) are closely related to oxidative stress1, and (5) have angiogenesis and cell apoptosis.
The link between atherosclerosis and cancer suggests that certain molecules that participate in tumorigenesis may also play a role in the development of atherosclerosis. Therefore, further studies should focus on the relationship between Sparcl1 and atherosclerosis.
Conclusion and Future Research Perspectives
Sparcl1 was identified as a new candidate gene associated with atherosclerosis in the analysis of the peripheral blood leukocyte transcriptome in human aortic atherosclerotic lesions as early as 2009.20 Recently, in a study of the single-cell transcriptome of human atherosclerotic plaques, Sparcl1 was found to be expressed in the ninth cluster of endothelial cells, in addition to COL4A1, COL4A2, and PLVA.21 This is constructive for the relationship between Sparcl1 and atherosclerosis, and this is the first time that Sparcl1 expression has been clarified in atherosclerotic tissue. All of these studies suggest that Sparcl1 is indeed associated with atherosclerosis. Combining the role of Sparcl1 in some physiological processes and the development of atherosclerosis, it is not difficult to speculate that Sparcl1 and atherosclerosis are closely related. However, the direct link between Sparcl1 and atherogenesis remains to be investigated, such as whether Sparcl1 promotes or inhibits the development of atherosclerosis, how and through what signaling pathways Sparcl1 affects the progression of atherosclerosis, and whether inhibition or enhancement of Sparcl1 expression makes a substantial difference to the development of atherosclerosis. The results of these forthcoming studies will provide insights into the development of atherosclerosis. These upcoming studies will have a very favorable impact on the study of the progression of atherosclerosis and may offer a new therapeutic modality.
Abbreviations
NO, nitric oxide; ICAM-1, intercellular adhesion molecule-1; VCAM-1, vascular cell adhesion molecule-1; ox-LDL, oxidized low-density lipoprotein; VSMC, vascular smooth muscle cells; LDL, low-density lipoproteins; Sparcl-1, Secreted protein acidic and rich in cysteine-like 1; EC domain, extracellular calcium binding domain; MEK, mitogen-activated protein kinase kinase; ERK, extracellular signal-related kinase; ITGB1, integrin β1.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by Anhui Medical University Scientific Research Fund (Grant No.2020xkj220 and 2020xkj165), the Natural Science Foundation of the Anhui Higher Education Institutions of China (Grant No.KJ2019A0260), the Key Scientific Research Platform Open Project Fund of Anhui Province (Grant No.KFZZ202204) and Postgraduate Innovation Research and Practice Program of Anhui Medical University (Grant No.YJS20230072).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Li -J-J, Gao R-L. Should atherosclerosis be considered a cancer of the vascular wall? Medical Hypotheses. 2005;64(4):694–698. doi:10.1016/j.mehy.2004.11.043
2. Tapia-Vieyra JV, Delgado-Coello B, Mas-Oliva J. Atherosclerosis and cancer; a resemblance with far-reaching implications. Arch Med Res. 2017;48:12–26.
3. Ouimet M. Autophagy in obesity and atherosclerosis: interrelationships between cholesterol homeostasis, lipoprotein metabolism and autophagy in macrophages and other systems. Biochim Biophys Acta. 2013;1831:1124–1133.
4. Ross R. Atherosclerosis--an inflammatory disease. N Engl J Med. 1999;340:115–126.
5. Zhang B, Gu J, Qian M, Niu L, Zhou H, Ghista D. Correlation between quantitative analysis of wall shear stress and intima-media thickness in atherosclerosis development in carotid arteries. Biomed Eng OnLine. 2017;16:137.
6. Chistiakov DA, Melnichenko AA, Grechko AV, Myasoedova VA, Orekhov AN. Potential of anti-inflammatory agents for treatment of atherosclerosis. Exp Mol Pathol. 2018;104(2):114–124. doi:10.1016/j.yexmp.2018.01.008
7. Pirillo A, Norata GD, Catapano AL. LOX-1, OxLDL, and atherosclerosis. Mediators Inflamm. 2013;2013:152786. doi:10.1155/2013/152786
8. Lim S, Park S. Role of vascular smooth muscle cell in the inflammation of atherosclerosis. BMB Rep. 2014;47(1):1–7. doi:10.5483/bmbrep.2014.47.1.285
9. Hermida N, Balligand JL. Low-density lipoprotein-cholesterol-induced endothelial dysfunction and oxidative stress: the role of statins. Antioxid Redox Signal. 2014;20:1216–1237.
10. Mundi S, Massaro M, Scoditti E, et al. Endothelial permeability, LDL deposition, and cardiovascular risk factors-a review. Cardiovasc Res. 2018;114:35–52.
11. Jebari-Benslaiman S, Galicia-García U, Larrea-Sebal A, et al. Pathophysiology of atherosclerosis. Int J Mol Sci. 2022;23:3346.
12. Ross R, Glomset JA. Atherosclerosis and the arterial smooth muscle cell: proliferation of smooth muscle is a key event in the genesis of the lesions of atherosclerosis. Science. 1973;180:1332–1339.
13. Ridker PM, Everett BM, Thuren T, et al. Antiinflammatory therapy with canakinumab for atherosclerotic disease. N Engl J Med. 2017;377:1119–1131.
14. Ait-Oufella H, Salomon BL, Potteaux S, et al. Natural regulatory T cells control the development of atherosclerosis in mice. Nat Med. 2006;12:178–180.
15. Ross JS, Stagliano NE, Donovan MJ, Breitbart RE, Ginsburg GS. Atherosclerosis: a cancer of the blood vessels? Am J Clin Pathol. 2001;116(Suppl):S97–S107.
16. Cordero A, Fácila L, García-Carrilero M, Gunturiz C, Montagud V, Núñez J. Breakfast habits in patients hospitalized for acute coronary syndrome. Rev Esp Cardiol. 2015;68:814–815.
17. de Nigris F, Sica V, Herrmann J, et al. c-Myc oncoprotein: cell cycle-related events and new therapeutic challenges in cancer and cardiovascular diseases. Cell Cycle. 2003;2:325–328.
18. Regensburger D, Tenkerian C, Pürzer V, et al. Matricellular protein SPARCL1 regulates blood vessel integrity and antagonizes inflammatory bowel disease. Inflam Bowel Dis. 2021;27:1491–1502.
19. Liu B, Xiang L, Ji J, et al. Sparcl1 promotes nonalcoholic steatohepatitis progression in mice through upregulation of CCL2. J Clin Invest. 2021;131(20):e144801. doi:10.1172/JCI144801
20. Timofeeva AV, Goriunova LE, Khaspekov GL. [Comparative transcriptome analysis of human aorta atherosclerotic lesions and peripheral blood leukocytes from essential hypertension patients]. Kardiologiia. 2009;49(9):27–38. Russian.
21. Depuydt MAC, Prange KHM, Slenders L, et al. Microanatomy of the human atherosclerotic plaque by single-cell transcriptomics. Circ Res. 2020;127(11):1437–1455. doi:10.1161/CIRCRESAHA.120.316770
22. Naschberger E, Liebl A, Schellerer VS, et al. Matricellular protein SPARCL1 regulates tumor microenvironment–dependent endothelial cell heterogeneity in colorectal carcinoma. J Clin Invest. 2016;126:4187–4204.
23. Bradshaw AD. Diverse biological functions of the SPARC family of proteins. Int J Biochem Cell Biol. 2012;44:480–488.
24. Johnston IG, Paladino T, Gurd JW, Brown IR. Molecular cloning of SC1: a putative brain extracellular matrix glycoprotein showing partial similarity to osteonectin/BM40/SPARC. Neuron. 1990;4:165–176.
25. Schraml P, Shipman R, Stulz P, Ludwig CU. cDNA subtraction library construction using a magnet-assisted subtraction technique (MAST). Trends Genet. 1993;9:70–71.
26. Girard JP, Springer TA. Cloning from purified high endothelial venule cells of hevin, a close relative of the antiadhesive extracellular matrix protein SPARC. Immunity. 1995;2:113–123.
27. Isler SG, Schenk S, Bendik I, et al. Genomic organization and chromosomal mapping of SPARC-like 1, a gene down regulated in cancers. Int J Oncol. 2001;18:521–526.
28. Nelson PS, Plymate SR, Wang K, et al. Hevin, an antiadhesive extracellular matrix protein, is down-regulated in metastatic prostate adenocarcinoma. Cancer Res. 1998;58:232–236.
29. Bendik I, Schraml P, Ludwig CU. Characterization of MAST9/Hevin, a SPARC-like protein, that is down-regulated in non-small cell lung cancer. Cancer Res. 1998;58:626–629.
30. Ma Y, Xu Y, Li L. SPARCL1 suppresses the proliferation and migration of human ovarian cancer cells via the MEK/ERK signaling. Exp Ther Med. 2018;16:3195–3201.
31. Han W, Cao F, Ding W, et al. Prognostic value of SPARCL1 in patients with colorectal cancer. Oncol Lett. 2018;15:1429–1434.
32. Maurer P, Hohenadl C, Hohenester E, Göhring W, Timpl R, Engel J. The C-terminal portion of BM-40 (SPARC/osteonectin) is an autonomously folding and crystallisable domain that binds calcium and collagen IV. J Mol Biol. 1995;253:347–357.
33. Brekken RA, Sage EH. SPARC, a matricellular protein: at the crossroads of cell-matrix communication. Matrix Biol. 2001;19(8):816–827.
34. Vannahme C, Gösling S, Paulsson M, Maurer P, Hartmann U. Characterization of SMOC-2, a modular extracellular calcium-binding protein. Biochem J. 2003;373:805–814.
35. Ye H, Wang WG, Cao J, Hu XC. SPARCL1 suppresses cell migration and invasion in renal cell carcinoma. Mol Med Rep. 2017;16:7784–7790.
36. Wang Y, Liu S, Yan Y, Li S, Tong H. SPARCL1 Influences Bovine Skeletal Muscle-Derived Satellite Cell Migration and Differentiation through an ITGB1-Mediated Signaling Pathway. Animals. 2020;10:1361.
37. Wang Y, Liu S, Yan Y, Li S, Tong H. SPARCL1 promotes C2C12 cell differentiation via BMP7-mediated BMP/TGF-β cell signaling pathway. Cell Death Dis. 2019;10:852.
38. Funk SE, Sage EH. The Ca2(+)-binding glycoprotein SPARC modulates cell cycle progression in bovine aortic endothelial cells. Proc Natl Acad Sci U S A. 1991;88:2648–2652. doi:10.1073/pnas.88.7.2648
39. Claeskens A, Ongenae N, Neefs JM, et al. Hevin is down-regulated in many cancers and is a negative regulator of cell growth and proliferation. Br J Cancer. 2000;82:1123–1130. doi:10.1054/bjoc.1999.1051
40. Zhang S, Zhang F, Feng L. The inhibition of HeLa cells proliferation through SPARCL1 mediated by SPP1. Cytotechnology. 2021;73:71–78. doi:10.1007/s10616-020-00443-2
41. Liu X, Zhao J, Luan X, et al. SPARCL1 impedes trophoblast migration and invasion by down-regulating ERK phosphorylation and AP-1 production and altering EMT-related molecule expression. Placenta. 2020;89:33–41. doi:10.1016/j.placenta.2019.10.007
42. Girard JP, Springer TA. Modulation of endothelial cell adhesion by hevin, an acidic protein associated with high endothelial venules. J Biol Chem. 1996;271(8):4511–4517. doi:10.1074/jbc.271.8.4511
43. Zhao SJ, Jiang YQ, Xu NW, et al. SPARCL1 suppresses osteosarcoma metastasis and recruits macrophages by activation of canonical WNT/beta-catenin signaling through stabilization of the WNT-receptor complex. Oncogene. 2018;37:1049–1061. doi:10.1038/onc.2017.403
44. Meissburger B, Perdikari A, Moest H, Müller S, Geiger M, Wolfrum C. Regulation of adipogenesis by paracrine factors from adipose stromal-vascular fraction - a link to fat depot-specific differences, Biochim. Biophys Acta. 1861;2016:1121–1131. doi:10.1016/j.bbalip.2016.06.010
45. Xiao C, Jin HG, Zhang LC, et al. Effects of SPARCL1 on the proliferation and differentiation of sheep preadipocytes. Adipocyte. 2021;10(1):658–669. doi:10.1080/21623945.2021.2010901
46. Kos K, Wong S, Tan B. Regulation of the fibrosis and angiogenesis promoter SPARC/osteonectin in human adipose tissue by weight change, leptin, insulin, and glucose. Diabetes. 2009;58(8):1780–1788. doi:10.2337/db09-0211
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.