Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Skin Ageing: A Progressive, Multi-Factorial Condition Demanding an Integrated, Multilayer-Targeted Remedy
Authors Liang Y ![]() , Su W, Wang F
, Su W, Wang F
Received 14 March 2023
Accepted for publication 1 May 2023
Published 9 May 2023 Volume 2023:16 Pages 1215—1229
DOI https://doi.org/10.2147/CCID.S408765
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Yihuai Liang,1,2 Wenrou Su,1,2 Feifei Wang1– 3
1Research and Development Center, Yunnan Botanee Bio-Technology Group Co. Ltd, Shanghai, People’s Republic of China; 2Yunnan Yunke Characteristic Plant Extraction Laboratory Co. Ltd, Kunming, Yunnan, People’s Republic of China; 3Medaesthee (Shanghai) Biotechnology Co., Ltd, Shanghai, People’s Republic of China
Correspondence: Feifei Wang, Research and Development Center, Yunnan Botanee Bio-Technology Group Co. Ltd, No. 999 Huaxu Road, Shanghai, 201702, People’s Republic of China, Tel +86 21 39880807, Email [email protected]
Abstract: Of the human organs, skin is the most visible one that displays the manifestations of ageing. It has a very intricate microanatomical structure and performs several key physiological functions. The pathophysiology of cutaneous ageing is characterized by deterioration of structural stability and functional integrity, implying a continuous reduction in maximal function and reserve capacity, as a result of the accumulating damage due to both intrinsic and extrinsic factors. Elimination of unfavorable expressions associated with facial and cutaneous ageing is the key patient demand in aesthetic dermatology. Even though the progress has been made in nonsurgical therapies like fillers and lasers, non-invasive interventions by using skin care products designed for rejuvenation at an early stage are the most popular and accessible solution among people. In this review, we have scrutinized the ageing-associated cutaneous changes at molecular, cellular and tissue levels. To optimize the ageing process towards a healthy skin, we propose an integrated, multilayer-targeted intervention, which involves both topical application of anti-ageing formulations from outside and oral supplementation from inside. Additionally, several promising naturally derived ingredients are reviewed from an anti-aging perspective. Most of them possess various bioactivities and may contribute to the development of the mentioned anti-ageing remedy.
Keywords: skin barrier, skin ageing, causative factors, anti-ageing remedy, naturally derived ingredients
Introduction
Ageing occurs as a gradual breakdown of homeostasis—the capability of the body system to maintain an internal balanced state while adapting to changing conditions. This failure ultimately give rise to the damage accumulation and cause the progressive dysfunction of all tissues and cells. The ageing problem can become even more tricky when the protective function of skin has been lost. “Dermatoporosis”, a chronic cutaneous disorder, occurs as high as one-third among in-patient elders. This syndrome is featured by “atrophic skin with solar purpura”, “white pseudoscars on the extremities”, “skin lacerations” and “delayed healing”.1,2
As the most visible organ, the skin can display the most apparent indication of human ageing. However, people desire to look younger than others at the same chronological age, by maintaining a well-functioned body including skin. Perceived age (self-perceived or perceived by others, relevant to ageing characteristics at functional and molecular levels) is used as a robust indicator of ageing by clinicians to evaluate physical health condition among patients over 70s.3
Since ageing itself cannot be physically avoided, skin anti-ageing means to defer the progress of skin senescence as late as possible, or to mitigate severity of age-dependent alterations. It is essential to reveal the underlying mechanism of skin ageing and develop anti-ageing remedies to combat and suppress the symptoms brought about by ageing. Correction of unfavorable expressions associated with facial ageing is the first reason for patients consulting in aesthetic dermatology. The remarkable progress has been made not only in cosmetic surgery but also in nonsurgical therapies like fillers and lasers. However, non-invasive interventions by using skin care products designed for rejuvenation at an early stage are still the most popular and accessible solution for most people. In this review, we try to go through the consensus about ageing skin and highlight published discoveries of “senescence” at levels from the molecule to the system, and discuss an integrated, multilayer-targeted remedy.
We further focus on naturally derived bioactive ingredients with potential anti-aging properties, and take some examples from China and the rest of the world. Some of them are antioxidants that scavenge free radicals, and some have an ability of anti-inflammation to mitigate skin “inflammaging”. There are also some botanical extracts that can boost collagen synthesis in the skin matrix or protect the collagen from enzymatic degradation, and so on. Admittedly, scientific evidence gathered in this review is mostly from in vitro studies. We cannot emphasize enough the importance of in vivo evidence on their efficacy and safety, effect concentrations in formulations, and duration of the anticipated effect.
Skin Architecture and Physiological Functions
Skin, measuring around 2m2 in area and a full one-sixth of the total weight for an adult, is the largest human organ. It has a very intricate microanatomical structure with several important cellular types, ie keratinocyte, fibroblast, melanocyte, Merkel cell and various immune cells (mast cell, migrating mononuclear cell and Langerhans). In addition to skin cells, there are sweat glands, hair follicles, plus a complex capillary and nerve network. Connective tissues rich in extracellular matrix (ECM) are the foundation of skin’s flexibility.
Basically, epidermis, dermis, and hypodermis (also known as the subcutaneous layer) are the main layers of skin. The stratified squamous epidermis is able to self-renew. The primary epidermal cells are keratinocytes which origin from the ectoderm in early embryonic development. The dermis, a layer much thicker than the epidermis, is composed of papillary dermis (the upper one) and reticular dermis (the lower one), containing fibroblasts and complex ECM. Its elastic property is supplied by elastin fibers, while the structural elements are provided by collagen fibers. Between the epidermis and the dermis is the dermal-epidermal junction (DEJ), which holds the two layers together and functions as a barrier to chemicals and cells. The deepest skin layer, hypodermis, lies below above the underlying fascia and muscle and the dermis. It contains connective tissues, blood vessels, and fat.
Skin performs a series of key functions. It is the front line for protecting bodies against disruption from the outside and self-replenish constantly in normal homeostasis. The epidermal barrier plays a well-known protective role at five functional levels (chemical, physical, immune, microbial and neuronal).4 Besides, the skin helps keep body temperature constant through the moisture modulation. It collects sensory signals via nerve endings, secretes sweat, sebum, hormones, and enzymes, and excretes harmful metabolic products from inner organs. In addition to the anatomic basis of cutaneous system,5–8 emerging proof has made it clear that skin can produce the same biomolecules (eg neurotrophins, neuropeptides, hormones and cytokines) like the other part of body and interact with the system via them,9–11 identifying the skin as a “neuro–endocrine–immune” organ.12
Pathophysiology of Skin-Ageing
Cutaneous ageing pertaining to a steady deterioration of structural stability and functional integrity,13 manifests at the molecular level as a continuous decline in reserve capacity and maximum function. From some perspectives, ageing is a programmed genetically, inevitable process, ie intrinsic, or chronological ageing. Additionally, signs of the premature senescence are considered partially an outcome of exogenous insults in the short term or long term, ie extrinsic ageing.
For intrinsically aged skin, the typical clues are fine wrinkles, dryness, pale in color, and less elasticity.14 Histologically, the epidermis becomes thinner and the DEJ increasingly flattened out, which accounts for increased skin fragility and decreased nutrient deliver through the DEJ interface. The loss in interface area between two layers may pose a risk to the separation of epidermis from dermis, possibly a process in which wrinkles develop. Slower wound healing and less effective desquamation also take place because of decline in epidermal cell reproductive activities. The epidermal turnover rate is found as slower as at least 30% in adults at 80s than that in adults at 30s.15
Reduction of dermis thickness occurs along with a decrement in vascularity and cellularity as well as the quantity of fibroblasts and mast cells. The content levels of hyaluronic acid and glycosaminoglycans in this layer declines with the ageing process, and depletion of collagen and elastin leads to disorganization of connective tissue and causes the rise of wrinkles.
For extrinsically aged skin, photoaging is the most common form induced by environmental factors. As much as eighty percent of premature facial ageing is attributable to solar radiation in a long term.16 Clinical signs of photoaging include roughness, dryness, deep wrinkle, lack of elasticity, uneven pigmentation, mainly superimposed on face, neck, and dorsal forearms. Histologically, “elastosis”, a phenomenon of amorphous elastin material accumulated just below the DEJ, is the main characteristic of the pathognomonic of photoaged skin. Atrophic epidermis, fragmented elastic fibers and collagen are also found to develop with constant sun exposure.15
Ageing occurs at any layer of the skin, and hypodermis is not an exception. Site-specific changes, including atrophy and hypertrophy, occur in this layer, ie fat losses in the shins, hands and face, and fat gains in the area between the hips and rib cage.
It is a natural inclination to focus on epidermal and dermal changes when evaluating skin ageing. The skin of face and neck unlike other anatomical locations of human body, however, has retained the cutaneous musculature, namely “panniculus carnosus”, a thin layer of striated muscles affixed to the fascia and skin and providing skin twitching and contraction functions.17 The cutaneous muscles with their aponeuroses (superficial musculoaponeurotic system, SMAS), as the laxity of which increases with age, have a significant impact on facial ageing.18 Facial movements, through contraction of facial muscles, are correlated with facial wrinkles, both dynamic (ie occurring along with contraction) and static (ie appearing constantly and more noticeable with contraction). The formation of dynamic wrinkle is attribute to the persistent hypercontraction that compensates the age-related cutaneous atrophy.19
Mechanisms of Skin-Ageing and the Causative Factors
The systemic ageing is suggested the synergistic or cumulative consequences of nine interconnected hallmarks: genomic instability, telomere attrition, epigenetic alterations, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication.20 Mechanisms such as transcriptional deregulation, stem cell exhaustion, advanced glycation end products (AGEs), ECM degradation and etc., are considered to be involved in the cutaneous senescence process.21–24
The senescence gradually proceeds at a relatively mutual rate at different levels of an individual organism. To present a patchwork of skin ageing, much effort has been devoted to scrutinizing age-related changes in skin at the molecular level (carbohydrates, lipids, nucleic acids, and proteins, etc.), at the cellular (keratinocytes, melanocytes, fibroblasts, etc.) and tissue (the different cutaneous layers) levels, and further, from the perspective of an individual.
Ageing of Biomolecules
During the ageing process, disruptions of both intracellular and extracellular components occur at the molecular level through direct UV damage or catalysis by oxidative enzymes, especially overwhelming chemical oxidation caused by reactive oxygen species (ROS).
UVA (320–400 nm) can penetrate deep into the dermal matrix. The absorbed UV energy causes structural change of biomolecules and often generates ROS. UV or endogenous ROS induce a variety of oxidative impairment to DNA, which are primarily restored by nucleotide or base excision repair (NER or BER) systems.25,26 Generally, the repair system or apoptosis (programmed cell death) is initiated rapidly and deal with this malfunctioned DNA. However, nucleotides that is not repaired or repaired in a wrong way, ie mutations, progressively build up in cells with ageing due to the decline in DNA repair capability by NER or BER systems, and then senescent-like cells accumulate in the skin.
Lipids, rich in organisms, can be an easy target of ROS. It should be considered how lipid peroxidation play a role in the ageing-associated alterations. A typical example is lipofuscin, autofluorescent materials of oxidized lipids and proteins in lysosomes. It has been described as “age pigment” due to its accumulation increased age, leading to reduction in the proteasome activity and homeostasis.27 Dermal photodamage may be induced as a result of UVA-induced photo-oxidative stress by failure in lysosomal removal of lipofuscin.28
Collagen, a large family of ECM proteins in the dermis, is susceptible to the aged cross-linked proteins, such as AGEs (products from a non-enzymatic glycation reaction between proteins and sugars) and HHL (histidinohydroxylysinonorleucine, one cross-link of type I collagen). Disorganized collagen becomes fragile and brittle, and the skin structural support get weakened.29 The physiological function that its surrounding keratinocytes and fibroblasts migrate or proliferate can be impaired by AGEs as well. The effect of UV on pentosidine (a biomarker of AGEs) generates ROS in the ECM leading to additional adverse consequences. Furthermore, AGEs provoke an enzyme activation of matrix degradation (metalloproteinases), and then MMP-1 and MMP-3 from the matrix metalloproteinase (MMP) family can break down collagen fibers causing loss to functional ECM.29
Among the degenerative changes of cutaneous ageing, the elasticity loss induced by alterations in production or degradation of elastin is concerned. “Solar elastosis” is one of the major histologic indicators of photoaged skin, characterized by deposition of thickened and tangled amorphous elastotic materials. This phenomenon may be explained by the alternative splicing of the elastin gene led by chronic sun exposure, and then triggering the wrong assembly of elastic fibers.30 Similarly, overproduction of ROS is involved significantly in elastolysis. Both elastin and collagen can be broken up by MMPs,31 which is probably associated with ROS-induced activation of activator protein 1 (AP-1).32
Ageing of Cutaneous Cells and Tissues
Generally, cell viability tends to decline with ageing because its capacity to repair various forms of injury gets lost. As dysfunctional molecules are accumulated in cells and tissues, senescent phenotypes occur.
Although cellular senescence is proposed as one of the ageing hallmarks,20 its origin in the human skin is still under investigated.33 Senescent cells share common features with alteration of gene expression and cell cycle, reorganized chromatin, and changes in morphology and metabolism.34 Cell cycle, a series of events like cell division, differentiation and regeneration is dominated by genomic stability. This stability is mainly attributed to the modulation of the telomeric ends that protect chromosomes by a nucleoprotein cap from exposure to the DNA damage reaction.35,36
For most types of normal cells, telomeres get shortened with each run of cell cycle owing to incomplete lagging-strand DNA synthesis, and when too short, the cell will be unable to divide, and the outcome of cellular senescence arises. The telomerase enzyme complex with the ability to maintain telomere length can be found in the basal epidermis (keratinocytes) but is almost undetectable in the dermis (fibroblasts). A balance between telomerase activity and telomere length is essential to keep skin cells normal.37 For patients suffering from a disease called dyskeratosis congenita, a number of epidermal abnormalities are found related to in the mutated telomerase component genes.38
Telomeres are likely to be influenced by rapid shortening caused by DNA-damaging agents such as ROS. The cutaneous effects of ROS vary with cell types, ie keratinocytes appear more resistant to UVB-induced injury, and are likely subjected to more apoptosis and less senescence in comparison with fibroblasts in the same condition.39 Therefore, while the defective keratinocytes are removed by apoptosis, fibroblasts with telomere shortening may get degenerative, remaining in the dermis layer to adversely affect epidermal growth via cellular communication (eg paracrine signaling) and ECM deposition.37
Mitochondrial dysfunction is also linked to phenotypic signs of skin ageing: wrinkle formation, delayed or decreased wound healing, and irregular pigmentation.40 The constant renewal capacity of skin depends on the rapid proliferation of its progenitor cells, the energy of which is supplied through mitochondrial respiration. Mitochondria can be both targets and producers of oxidative stress: during mitochondrial respiration ROS is produced and then disrupt macromolecular and cellular structures if not suppressed by the antioxidant system; and additionally, mitochondria themselves are the organelles susceptible to the process of UV-induced and chronological skin ageing. Higher rates of mutations and deletions in mitochondrial DNA (mtDNA) are detected in photoaged skin and skin cancer lesions.41
Senescence-associated secretory phenotype (SASP), a phenomenon that senescent cells release pro-inflammatory chemokines, cytokines, proteases and growth factors, may promote chronic inflammation and frailty in aged skin.42 The SASP seems closely linked to the immune system. Macrophage chemokines, eg monocyte chemoattractant protein-1, are prominent elements of the SASP, while innate immunity including infiltration of macrophage is involved in removing senescent cells. A decline of tissue macrophage responses potentially contributing to a cumulating burden of senescent cells in elderly, which in turn disrupts the immune function.43 This phenomenon is referred to as “inflammaging”, a continuous, low-grade inflammation associated with ageing.44
From a perspective of tissues, cutaneous manifestations such as fine wrinkles, senile pigmentation,45 and senile xerosis,46 and are common among the elderly. As mentioned above, the epidermal proliferation and turnover slow down significantly in aged skin. Xerosis (dry skin) displays aberrations in the keratin expression in the epidermis. Pigmentation homeostasis is altered in the ageing process. In addition to “pigment-producing” melanocytes damaged directly, the physiological interaction between melanocytes and other surrounding senescent cells such as keratinocytes, fibroblasts, endothelial cells take part in the development of senile lentigo.47–49
Systemic Ageing of an Individual
Ageing is a pathophysiological process than an individual suffers a gradual impaired homeostasis of the whole system including skin, skeleton, internal organs, brain and so on. Different body sites show various signs of senescence over time, and particularly, the skin ageing is the most important element that determine how people estimate the degree of individual senescence.
In essence, the causative factors of skin ageing are broadly categorized as “intrinsic” and “extrinsic”.16,50 A changeable interaction between intrinsic and extrinsic modes may exist, which make it very difficult to discriminate the levels of involvement in the ageing processes between the two types of factors.
From an intrinsic standpoint, an intrinsic defect, such as gene mutation that impairs genome stability and DNA repair, can cause a genetic disorder characterized by premature ageing. Werner Syndrome,51 xeroderma pigmentosa,52 and Cockayne Syndrome,53 are well-known progeria syndromes with distinct features of individual senescence. However, among the middle-aged at the identical chronological age, disparities in paces of skin/facial ageing and varying degree of senescence may be shown, the reason of which should not be explained only by the governance of innate genetic factors but also by the susceptibility to external causes, eg solar radiation,54 diet,55 pollution,56,57 and even lack of sleep.58,59
To elaborate how important each external cause (extrinsic ageing) is, the twin studies have been designed, which allow a unique opportunity to control for the majority of the genetic factors (intrinsic ageing). Interestingly, a Japanese monozygotic twin study demonstrates that facial texture and wrinkle scores are likely to be influenced by environmental factors, suggesting tobacco smoking accelerates the visible ageing of skin, and whereas use of skin protectant decelerates this process.60
Skin Anti-Ageing Regimen
More than a mere aesthetic problem, skin degeneration in structural stability and functional integrity with the passage of time is defined as a disease by Russell-Goldman and Murphy.61 Skin ageing can negatively and significantly affect the “quality of life” (QOL) of the elderly, especially for those with dermatological disorders worthy of medical attentions.
Since skin serves as “a neuro–endocrine–immune organ”,12 any locally disturbed cutaneous homeostatic status can influence the steady conditions of the whole system. According to the concept of “inflammatory skin march” in atopic dermatitis, once pro-inflammatory cytokines like interleukin (IL) −1 and tumor necrosis factor-α (TNF-α) from skin lesions are released beyond normal limits, chronic low-grade systemic inflammation may be caused, which predisposes the patient to suffer from cardiovascular disease and type 2 diabetes.62 In other words, the attempt to prevent skin senescence should have “implications that transcend the integument” and will benefit those vital internal organs that “youthful skin was designed to envelop and safeguard”61 in terms of systemic ageing and age-related dysfunction.
A constant pursuit of human is to arrest, or postpone the pace of skin ageing, and even get rejuvenated. Hence, a number of anti-ageing approaches pertaining to the skin have been developed to minimize the cutaneous signs such as wrinkle, laxity, roughness, pigmentation, dryness and chronic inflammation.63 It is not realistic to get rid of all these signs with one single anti-ageing approach. Consequently, we will discuss in this chapter an integrated regimen of skin anti-ageing, a combination of various approaches to skin revitalization and restoration of each layer (see Figure 1).
 |
Figure 1 A schematic illustration of an integrated, multilayer-targeted intervention, which involves both topical application of anti-ageing formulations from outside and oral supplementation from inside. |
Multilayered Targets at Various Depths
Restoring Impaired Epidermal Barrier
Primarily, the topical use of moisturizers is the cornerstone of skin barrier regulation and repair.64,65 Direct supplements of the triple lipid mixture “ceramides, cholesterol and free fatty acids” are beneficial to maintain the cutaneous barrier by filling the space of the outermost layers of keratinocytes (stratum corneum).66 Furthermore, the effort to improve epidermal barrier is worth via increasing expressions of “cornified envelope”-related proteins such as keratin,67 filaggrin,68 and loricrin.69
Attenuating Skin Inflammaging
Since tissue homeostatic status can be influenced by SASP significantly, the possibility of suppressing SASP factors from senescent cells and mitigating the negative impacts of “inflammaging” is of particular interest.70–72 The JAK/STAT pathway,73 the transcription factor NF-κB,74 as well as senescence-associated β-galactosidase (SA-β-gal),75 represent favorable targets to control SASP.
Upregulating the Cellular and ECM Turnover
Epidermal turnover can be promoted via stimulating proliferation of keratinocytes and accelerating the peeling process of stratum corneum (desquamation). The cosmetic use of retinoids,76 or hydroxy acids such as alpha hydroxy acids (AHAs) and polyhydroxy acids (PHAs) may exhibit this benefit. Modulation of the dermal fibroblast–ECM interactions, by breaking the self-perpetuating cycle of collagen fibril fragmentation, is also an important procedure to skin anti-ageing.77,78 This involves introducing agents that can stimulate fibroblast proliferation,79 suppress expression of MMPs,80 boost collagenesis,81 elastin production,82 and glycosaminoglycan synthesis,83 etc.
Lessening Oxidative Stress
Oxidative stress initiated by excessive ROS generation causes cell injury. Antioxidants are conventionally adopted to relieve skin ageing by neutralizing formed ROS. There are many popular antioxidants, eg vitamin C, vitamin E, glutathione, coenzyme Q10, as well as some botanical extracts.
Reducing AGEs Deposition
AGEs are viewed as “key players in skin ageing”.84 Various AGEs affect skin at different layers: weakening epidermis barrier function, destroying structure of keratinocytes, promoting melanin production in melanocytes, obstructing skin wound healing, stimulating fibroblast apoptosis, and damaging fiber contracture in the dermis.85 Preventative agents that inhibit AGEs formation, break the generated AGEs, or block receptors of AGEs, are of particular interest in counteracting AGEs induced skin ageing.
Rescuing Telomere Shortening
Attention has been paid to telomerase and telomere length for anti-ageing interventions.37,72,86 The shield of telomeric DNA, known as “shelterin”, is a specialized six-protein complex, and has fundamental actions on modulating telomere length and telomere capping.87 Therefore, potential shelterin enhancer, telomerase activator88 and telomerase reverse transcriptase (TERT) expression activator89 have been sought for anti-ageing interventions.86,90
Relaxing Contraction of Facial Musculature
To postpone the progress of “expression wrinkles” with the elapsing time, non-invasive cosmetic approaches have been developed to suppress repetitive facial muscle movement. Herein, neurotransmitters such as acetylcholine released from nerve endings bind to their corresponding receptors on the surface of myocytes, which depolarizes/activates Na+ channels, and accordingly, stimulates Ca2+ channels, and eventually, triggers contraction.91 Based on the above signaling pathway, novel compounds that inhibit nerve endings from releasing acetylcholine, competitively bind acetylcholine receptors, or deactivate Na+ channels can be promising for the cosmetic use.
Crosstalk Between the Skin and the System
Skin functioning relies on the homeostasis of the whole body, and conversely, cutaneous functions in an intact and steady state contribute a lot to everyone’s well-being. Apparently, skin ageing is part of the systemic ageing of the whole body. It is rational to take care of skin from both outside and inside to combat ageing.
From the Outside in: Topical Application
The traditional approach to anti-ageing is topical use of skin care products, directly applied to a body surface. Although the cosmetic industry has succeeded in discovering and incorporating several active ingredients in their products, the epidermal barrier remains an obstacle in the way that these actives penetrate the skin. In order to overcome the barrier for the expected efficacy, an effective transdermal delivery of actives from the anti-ageing formulations should be designed and optimized, by using chemical permeation enhancers, as well as appropriate vehicles.92
In addition to positive impacts on physical appearance and emotional experience, an appropriate use of skin care products may bring imperceptible benefits to one’s health through preventing systemic comorbidity in cutaneous degenerative alterations. For instance, if a shift toward cellular senescence in skin (inflammaging) is reversed at an early stage, ideally, circulating pro-inflammatory cytokines will be reduced to some extent, and the occurrence of age-related disorders such as arthritis, type 2 diabetes, and dementia, may be postponed.
From the Inside Out: Oral Supplement
Good nutrition is the foundation of healthy skin. Consuming a healthy diet throughout the life-course assists in prolonging lifespan and delaying ageing.55 Rational nutrient intakes are found associated with a more pleasant skin appearance,93,94 while a positive relation between dietary AGEs intake and AGEs accumulation in skin suggests potential detrimental effects on health outcomes.95 Oral anti-ageing preparations acting from the interior of human body, therefore, can be anticipated to manifest cutaneous benefits.
In the near decade, oral supplementation of collagen has come to be popular, and been an option of the anti-ageing remedy.96 A series of randomized control trials reveals that oral supplementation of collagen may boost skin hydration and elasticity, and lessen skin roughness and wrinkles.97 When orally administered, ceramides and hyaluronan, are as effective for skin moisturization as their topical application.98,99
Foodborne antioxidants (eg phytosterols, polyphenols, minerals and vitamins) can scavenge ROS obtained from diet and mitigate the related oxidative damage. Such an example is Rosa gallica extract taken by an oral route, which prevent UVB-induced photoaging in vivo.100
The communication between the skin and gut microbiome in a bidirectional manner, namely the gut-skin axis, works through the immune system, mediators of inflammation and so on.101 Oral prebiotics and probiotics may aid in regulating cutaneous conditions.102 The fermented dietary fibers can be metabolized by colonic bacteria into products called short-chain fatty acids (SCFA), which help to maintain an integral skin barrier by upregulating the turnover of keratinocytes.103
Apart from nutrition, sleep of good quality is another necessity for skin health. Sleep problems are common among adults, and likely to increase. Chronic sleep deficiency along with the disrupted circadian rhythm is relevant to cutaneous symptoms of damaged barrier integrality and accelerated ageing.59 Oral supplements derived from plant extracts that ameliorate sleep quality via neurotransmitter-mediated mechanisms have been extensively reviewed,104 which provide evidence for oral skin care and further, an auxiliary to skin anti-ageing.
Promising Naturally Derived Ingredients from an Anti-Ageing Perspective
With increased knowledge of cutaneous physiology and senescence, a great number of bioactives aiming the anti-ageing segment have emerged in the cosmetic market. The following section will dig deep to understand the underlying mechanisms by which some promising ingredients currently available take effect, while focusing on different anti-ageing targets mentioned above, as summarized in Table 1.
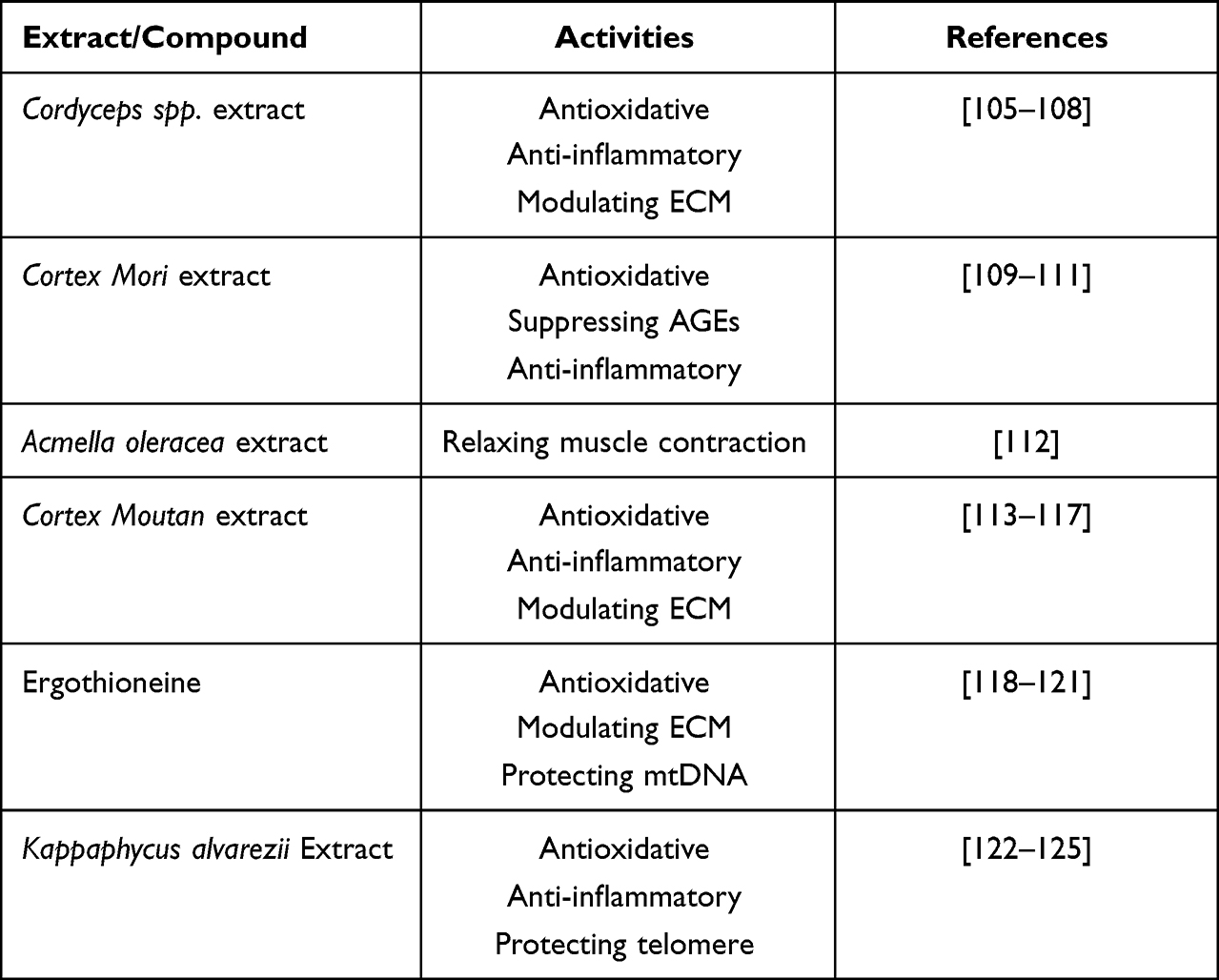 |
Table 1 Some Naturally Derived Ingredients of Interest in the Skin Anti-Ageing Regimen |
Cordyceps Spp. Extract
Cordyceps is a wide-ranging term for a fungi genus parasitic mainly on insects and other arthropods. The genus, comprising more than 400 species, has a worldwide distribution, especially in east and southeast Asia. More than sixty species have been discovered in China, two of which Cordyceps sinensis and Cordyceps militaris are the most valued species in the traditional medicine. Diverse bioactive components present in Cordyceps, including cordycepic acid, polysaccharides, ergosterol, peptides, nucleosides, especially cordycepin and its derivatives, form the basis of their broad range of biological potentials.126
How Cordyceps interacts with UVB has been studied. The aqueous extract of C. sinensis is found to protect keratinocytes against UV-induced damage, via reducing ROS production and intracellular hydrogen peroxide content. In the network pharmacology analysis, bioactive compounds in C. sinensis were suggested to act on PPAR (peroxisome proliferator-activated receptors) signaling pathway, cholesterol metabolism, and ovarian steroidogenesis against low dose UVB-induced alterations.105
Hirsutella sinensis, C. sinensis at an early stage, can suppress inflammasomes through inhibiting activated macrophages from secreting IL-1β and IL-18, supported by the finding than its ethanol extract reduces the production of ROS, the activation of caspase-4, and the inflammasome components (P2X7R and NLRP1).106
C. militaris can also serve as an ROS scavenger. After being pretreated by its aqueous extract, human dermal fibroblasts in vitro resist H2O2-induced apoptotic cell death, and show less activity of SA-β-gal, a biomarker of premature ageing at the cellular level.107 Furthermore, C. militaris extract can protect skin ECM against degradation. Based on its chemical profiles, phenolic compounds in the extract are able to inhibit hyaluronidase, MMP-1, and elastase, while flavonoids show an inhibitory impact on hyaluronidase.108
Cortex Mori Extract
Morus alba is a medium-sized mulberry tree, originally from China and now widely cultivated worldwide. Its root bark, Cortex Mori, is bioactives including stilbenes, flavonoids, terpenes, benzofurans, and alkaloids have been isolated from Cortex Mori.127 Cortex Mori extract exhibits a protective effect on diet-induced neuronal damage in mice by reducing AGEs deposit, RAGE (receptors for AGEs) expression and pro-inflammatory cytokines, and inhibiting mitogen-activated protein kinases (MAPKs),109 suggesting that this extract could be valuable for skin anti-ageing via modulation of the AGE-RAGE/MAPK signaling pathway. With respect to anti-inflammation, Cortex Mori shows the ability to down-regulate cyclooxygenase-2 mRNA, and suppress the production of prostaglandin E2 and nitric oxide in macrophages.110
Hydroxystilbenes, polyphenols naturally occurring, are also ROS scavengers. Of them, resveratrol (one polyhydroxy trans-stilbene compound) and its derivatives are found in mulberry root bark.128,129 Aside from its inhibition toward cellular tyrosinase activity and melanin formation,130 the anti-ageing potentials of resveratrol have been reviewed.131 In vitro, resveratrol can delay cellular senescence through enhancing telomerase activity of endothelial progenitor cells.132 Beneficial impacts of the resveratrol treatment on human fibroblasts in relation to ageing changes are also reported, eg promoting cell duplications, reducing the acetylation levels of p53 protein and histones, and restricting elevations in DNA content and nuclear size.133 Moreover, resveratrol may lower mRNA expressions of MMP-1 and MMP-3 in dermal fibroblasts treated by TNF-α via down-regulating p300/NF-κB acetylation,111 or improve mitochondrial functions, possibly correcting defects of mitochondrial fatty acid oxidation in cells.134 Nevertheless, clinical trials are required to clarify whether human will benefit from these findings.
Acmella oleracea Extract
“Neuro-relaxing” botanical ingredients have drawn attention from the cosmetic industry. Among those, Acmella oleracea extract is considered as a safe alternative to Botox, a drug that relaxes or paralyzes muscles. Acmella oleracea, is an annual, flowering herb species, native to South American. Owing to its anti-inflammatory and local anesthetic effects, Acmella, with a common name “toothache plant”, has been used to relieve oral pain for hundreds of years.135
Spilanthol (the main representative of N-alkylamides) is the key bioactive constituent abstracted from Acmella oleracea, flowers in particular.136 Using a nerve–muscle coculture model, the influence of spilanthol on muscle contraction frequency has been investigated, as well as the recuperation of contractile activity after spilanthol blocks muscle contractions. This compound manifests a Botox-like action, thus inhibiting contractions in subcutaneous muscles.112 The fact that spilanthol penetrates the epidermal barrier and reaches the dermis and even deeper sites has been demonstrated in Franz-diffusion cells using human skin,137 making it feasible to reduce facial expression wrinkles by topical application.
Interestingly, Acmella oleracea may also favor tendon repair, via increasing the molecular organization and content of collagen in partially transected tendons.138 Whether this effect help restore the impaired ECM in aged skin needs further studies.
Cortex Moutan (Peony) Extract
Cortex Moutan, a plant crude drug in traditional Chinese medicine, is the root bark of Paeonia suffruticosa. Phytochemical profiles indicate that Cortex Moutan contains many compounds, eg phenols and monoterpene glycosides. Paeoniflorin and paeonol are two of the characteristic and major components in Cortex Moutan.139 In addition to the pharmaceutical use, its efficacy against skin ageing has been concerned.
Cortex Moutan extracts can stimulate dermal keratinocytes and fibroblasts into proliferation at low concentrations.113 Paeoniflorin exerts a cytoprotective effect, when cultured keratinocytes from both hairless mouse and human are subjected to UVB-induced DNA damage. And reduced facial wrinkles were observed among volunteers using a paeoniflorin-containing formulation for eight weeks.114 These findings may be correlated with paeoniflorin’s inhibitory effect on the ROS-p38-p53 pathway.115
In a study of the senescence model of MRC-5 cells exposed to H2O2, paeonol can weaken the cellular ageing condition by attenuating the activity of SA-β-gal and expression of other senescence biomarkers. This anti-ageing effect is in part due to promotion of the transcription factor, “nuclear factor-erythroid 2–related factor 2” (Nrf2). Meanwhile, premalignant human keratinocytes display a decline in levels of SASP factors (IL-6 and -8) after the paeonol treatment.116
Paeonol extracted from Paeonia suffruticosa seems promising in protection skin from UVB-induced photoaging. The UVB exposure to keratinocytes causes the phosphorylation of MAPKs. MAPKs are the promoters of AP-1, and the latter is an important transcription activator of MMP-1 which degrades the ECM structural protein, procollagen type I. In vitro, paeonol disrupts this pathway in keratinocytes and attenuate potential cutaneous damage, further proved by the finding of an in vivo study, ie topical application of the extract on UVB-exposed hairless mice can decrease expression of MMP-1 and increase production of procollagen type I.117
Ergothioneine
Ergothioneine, a unique amino acid, is a natural antioxidant discovered from the rye ergot fungus Claviceps purpurea by Charles Tanret over one century ago.140 In UVA-irradiated human dermal fibroblasts, ergothioneine decrease expression of TNF-α and MMP-1.118 In a keratinocyte system co-cultured with fibroblasts, ergothioneine activates HSP70 as well as the Nrf2/HO-1 pathway in keratinocytes, and protect those skin cells from UVB damage by inhibiting ROS production, cytokines mediating inflammation and apoptosis, and thereby alleviates the subsequent senescence of fibroblasts via maintaining the steady-state of collagen.119
A specific transporter for ergothioneine (ETT) is concentrated in mitochondria. Cells lack of ETT suffer more mitochondrial DNA (mtDNA) damage caused by oxidative stress than the normal ones, suggesting the role of ergothioneine in protecting mtDNA from the adverse effects of oxidative stress related to superoxide generation in mitochondria.120 Another in vitro study shows that ergothioneine enhances the level of glutathione in a reduced state and prevents against photoaging-associated mtDNA mutations in cells.121
Kappaphycus alvarezii Extract
Seaweed or marine macroalgae refers to thousands of species of macroscopic multicellular organisms. It is a favorable source abundant in bioactive chemicals, contributing to the beneficial cutaneous activities, such as moisturized, antioxidative, photoprotective, anti-ageing, and anti-inflammatory.141 Among various seaweeds, Kappaphycus alvarezii (formerly Eucheuma cottonii) is an edible red seaweed predominating in the coastal and continental shelf areas. It contains high amounts of carbohydrates, proteins, phenolic compounds and polyunsaturated fatty acids, and thereby has been cultivated widely, and developed for carrageenans and even functional food.142
As a rich source of antioxidants, K. alvarezii extracts protect mice against liver damage caused by lead acetate.143 Oral supplementation with K. alvarezii modulates the balance of gut microbiota and reverses metabolic syndrome in obese rats,122 or decreases plasma H2O2 level and renal NOX4 (NADPH oxidase 4) gene expression in diabetic rats.123 In an asthma rat study, the seaweed consumption reduces mucin synthesis, suppresses eosinophil infiltration, and downregulates expression of MMP-9 and inflammatory mediators (NF-κB, IL-4 and TNF-α).124
K. alvarezii extract promotes migration in human keratinocytes in vitro through the p38 MAPK/CREB (cAMP response element‑binding protein) pathway.144 Hydrolysis product of K. alvarezii containing linear galactans stimulates the expression of POT1 and TPP1 (two of the components of “shelterin”), thus limiting the telomere shortening and decelerating the senescence process of dermal fibroblasts.125
Concluding Remarks
The visible signs of ageing, health, and disease can be easily displayed by skin. Skin ageing is a progressive condition as a consequence of the accumulating damage. The age-associated cutaneous changes, induced by both intrinsic and extrinsic factors, gradually emerge at different levels, ie biomolecules, cells, and tissues, and ultimately, leads to develop malfunctions of different cutaneous layers and even psychosomatic disorders. In this review, to optimize the ageing process towards a healthy skin, we propose an integrated, multilayer-targeted intervention, which involves introducing agents that can restore impaired epidermal barrier, attenuate inflammaging, fight against oxidative stress and AGEs deposition, inhibit telomere shortening, modulate abnormal ECM, and relax contracted facial muscles, with the aid of oral supplementation of anti-ageing preparations from inside. Moreover, naturally derived ingredients have played an important role in present means of treating ageing skin. Many of them possess various bioactivities, contributing to different skin benefits. However, further clinical trials are required to corroborate the safety and efficacy for the application of naturally derived ingredients.
Acknowledgments
The authors gratefully acknowledge the funding “Science and Technology Project” (2017IB007) from the Key Research and Development Plan of Yunnan Province and the fund of Yunnan Characteristic Plant Extraction Laboratory (2022YKZY006).
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Wollina U, Lotti T, Vojvodic A, Nowak A. Dermatoporosis – the chronic cutaneous fragility syndrome. Open Access Maced J Med Sci. 2019;7(18):3046. doi:10.3889/oamjms.2019.766
2. Kaya G, Kaya A, Sorg O, Saurat JH. Dermatoporosis, a prevalent skin condition affecting the elderly: current situation and potential treatments. Clin Dermatol. 2019;37(4):346–350. doi:10.1016/j.clindermatol.2019.04.006
3. Christensen K, Thinggaard M, McGue M, et al. Perceived age as clinically useful biomarker of ageing: cohort study. BMJ. 2009;339:b5262. doi:10.1136/bmj.b5262
4. Lefèvre-Utile A, Braun C, Haftek M, Aubin F. Five functional aspects of the epidermal barrier. Int J Mol Sci. 2021;22(21):11676. doi:10.3390/ijms222111676
5. Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nat Rev Immunol. 2009;9(10):679–691. doi:10.1038/nri2622
6. Sevilla LM, Pérez P. Roles of the glucocorticoid and mineralocorticoid receptors in skin pathophysiology. Int J Mol Sci. 2018;19(7):1906. doi:10.3390/ijms19071906
7. Glatte P, Buchmann SJ, Hijazi MM, Illigens BMW, Siepmann T. Architecture of the cutaneous autonomic nervous system. Front Neurol. 2019;10:970. doi:10.3389/fneur.2019.00970
8. Mancino G, Miro C, Di Cicco E, Dentice M. Thyroid hormone action in epidermal development and homeostasis and its implications in the pathophysiology of the skin. J Endocrinol Invest. 2021;44(8):1571–1579. doi:10.1007/s40618-020-01492-2
9. Russo AF. Overview of neuropeptides: awakening the senses? Headache. 2017;57(Suppl2):37–46. doi:10.1111/head.13084
10. Slominski AT, Manna PR, Tuckey RC. Cutaneous glucocorticosteroidogenesis: securing local homeostasis and the skin integrity. Exp Dermatol. 2014;23(6):369–374. doi:10.1111/exd.12376
11. Hannen RF, Michael AE, Jaulim A, Bhogal R, Burrin JM, Philpott MP. Steroid synthesis by primary human keratinocytes; implications for skin disease. Biochem Biophys Res Commun. 2011;404(1):62–67. doi:10.1016/j.bbrc.2010.11.059
12. Jin R, Luo L, Zheng J. The trinity of skin: skin homeostasis as a neuro-endocrine-immune organ. Life Basel Switz. 2022;12(5):725. doi:10.3390/life12050725
13. Farage MA, Miller KW, Elsner P, Maibach HI. Characteristics of the Aging Skin. Adv Wound Care. 2013;2(1):5–10. doi:10.1089/wound.2011.0356
14. Chaudhary M, Khan A, Gupta M. Skin ageing: pathophysiology and current market treatment approaches. Curr Aging Sci. 2020;13(1):22–30. doi:10.2174/1567205016666190809161115
15. Baumann L. Skin ageing and its treatment. J Pathol. 2007;211(2):241–251. doi:10.1002/path.2098
16. Uitto J. Understanding premature skin aging. N Engl J Med. 1997;337(20):1463–1465. doi:10.1056/NEJM199711133372011
17. Naldaiz‐Gastesi N, Bahri OA. The panniculus carnosus muscle: an evolutionary enigma at the intersection of distinct research fields. J Anat. 2018;233(3):275–288. doi:10.1111/joa.12840
18. Okuda I, Yoshioka N, Shirakabe Y, Akita K. Basic analysis of facial ageing: the relationship between the superficial musculoaponeurotic system and age. Exp Dermatol. 2019;28(Suppl 1):38–42. doi:10.1111/exd.13827
19. Carroll JM, Hanna S, Guenther LC, Boucher N. Comparison of topical antiaging creams in the management of lateral canthal lines. J Cosmet Dermatol. 2020;19(3):694–704. doi:10.1111/jocd.13062
20. López-Otín C, Blasco MA, Partridge L, Serrano M, Kroemer G. The hallmarks of aging. Cell. 2013;153(6):1194–1217. doi:10.1016/j.cell.2013.05.039
21. Gragnani A, Cornick SM, Chominski V. Review of major theories of skin aging. Adv Aging Res. 2014;3(4):265–284. doi:10.4236/aar.2014.34036
22. Haydont V, Bernard BA, Fortunel NO. Age-related evolutions of the dermis: clinical signs, fibroblast and extracellular matrix dynamics. Mech Ageing Dev. 2019;177:150–156. doi:10.1016/j.mad.2018.03.006
23. Nguyen HP, Katta R. Sugar sag: glycation and the role of diet in aging skin. Skin Ther Lett. 2015;20(6):1–5.
24. Cai Y, Song W, Li J, et al. The landscape of aging. Sci China Life Sci. 2022;65(12):2354–2454. doi:10.1007/s11427-022-2161-3
25. Krokan HE, Bjørås M. Base excision repair. Cold Spring Harb Perspect Biol. 2013;5(4):a012583. doi:10.1101/cshperspect.a012583
26. Marteijn JA, Lans H, Vermeulen W, Hoeijmakers JHJ. Understanding nucleotide excision repair and its roles in cancer and ageing. Nat Rev Mol Cell Biol. 2014;15(7):465–481. doi:10.1038/nrm3822
27. Skoczyńska A, Budzisz E, Trznadel-Grodzka E, Rotsztejn H. Melanin and lipofuscin as hallmarks of skin aging. Adv Dermatol Allergol Dermatol Alergol. 2017;34(2):97–103. doi:10.5114/ada.2017.67070
28. Lamore SD, Qiao S, Horn D, Wondrak GT. Proteomic Identification of Cathepsin B and Nucleophosmin as Novel UVA-Targets in Human Skin Fibroblasts. Photochem Photobiol. 2010;86(6):1307–1317. doi:10.1111/j.1751-1097.2010.00818.x
29. Reilly DM, Lozano J. Skin collagen through the lifestages: importance for skin health and beauty. Plast Aesthetic Res. 2021;8:2. doi:10.20517/2347-9264.2020.153
30. Weihermann AC, Lorencini M, Brohem CA, de Carvalho CM. Elastin structure and its involvement in skin photoageing. Int J Cosmet Sci. 2017;39(3):241–247. doi:10.1111/ics.12372
31. Van Doren SR. Matrix metalloproteinase interactions with collagen and elastin. Matrix Biol J Int Soc Matrix Biol. 2015;1:224–231. doi:10.1016/j.matbio.2015.01.005
32. Chiang HM, Chen HC, Chiu HH, Chen CW, Wang SM, Wen KC. Neonauclea reticulata (Havil) Merr Stimulates Skin Regeneration after UVB Exposure via ROS Scavenging and Modulation of the MAPK/MMPs/Collagen Pathway. Evid-Based Complement Altern Med ECAM. 2013;2013:324864. doi:10.1155/2013/324864
33. Toutfaire M, Bauwens E, Debacq-Chainiaux F. The impact of cellular senescence in skin ageing: a notion of mosaic and therapeutic strategies. Biochem Pharmacol. 2017;142:1–12. doi:10.1016/j.bcp.2017.04.011
34. Rübe CE, Bäumert C, Schuler N, et al. Human skin aging is associated with increased expression of the histone variant H2A.J in the epidermis. Npj Aging Mech Dis. 2021;7(1):1–11. doi:10.1038/s41514-021-00060-z
35. O’Sullivan RJ, Karlseder J. Telomeres: protecting chromosomes against genome instability. Nat Rev Mol Cell Biol. 2010;11(3):171–181. doi:10.1038/nrm2848
36. Jacczak B, Rubiś B, Totoń E. Potential of naturally derived compounds in telomerase and telomere modulation in skin senescence and aging. Int J Mol Sci. 2021;22(12):6381. doi:10.3390/ijms22126381
37. Buckingham EM, Klingelhutz AJ. The role of telomeres in the ageing of human skin. Exp Dermatol. 2011;20(4):297–302. doi:10.1111/j.1600-0625.2010.01242.x
38. Kirwan M, Dokal I. Dyskeratosis congenita: a genetic disorder of many faces. Clin Genet. 2008;73(2):103–112. doi:10.1111/j.1399-0004.2007.00923.x
39. D’Errico M, Lemma T, Calcagnile A. Cell type and DNA damage specific response of human skin cells to environmental agents. Mutat Res. 2007;614(1–2):37–47. doi:10.1016/j.mrfmmm.2006.06.009
40. Stout R, Birch-Machin M. Mitochondria’s role in skin ageing. Biology. 2019;8(2):29. doi:10.3390/biology8020029
41. Sreedhar A, Aguilera-Aguirre L, Singh KK. Mitochondria in skin health, aging, and disease. Cell Death Dis. 2020;11(6):444. doi:10.1038/s41419-020-2649-z
42. Velarde MC, Demaria M. Targeting senescent cells: possible implications for delaying skin aging: a mini-review. Gerontology. 2016;62(5):513–518. doi:10.1159/000444877
43. Tchkonia T, Zhu Y, van Deursen J, Campisi J, Kirkland JL. Cellular senescence and the senescent secretory phenotype: therapeutic opportunities. J Clin Invest. 2013;123(3):966–972. doi:10.1172/JCI64098
44. Zhuang Y, Lyga J. Inflammaging in skin and other tissues - The roles of complement system and macrophage. Inflamm Allergy Drug Targets. 2014;13(3):153–161. doi:10.2174/1871528113666140522112003
45. Kim JC, Park TJ, Kang HY. Skin-aging pigmentation: who is the real enemy? Cells. 2022;11(16):2541. doi:10.3390/cells11162541
46. Görög A, Bánvölgyi A, Holló P. Characteristics of the ageing skin, xerosis cutis and its complications. Dev Health Sci. 2022;4(4):77–80. doi:10.1556/2066.2021.00051
47. Yoon JE, Kim Y, Kwon S, et al. Senescent fibroblasts drive ageing pigmentation: A potential therapeutic target for senile lentigo. Theranostics. 2018;8(17):4620–4632. doi:10.7150/thno.26975
48. Kang HY, Lee JW, Papaccio F, Bellei B, Picardo M. Alterations of the pigmentation system in the aging process. Pigment Cell Melanoma Res. 2021;34(4):800–813. doi:10.1111/pcmr.12994
49. Fulop T, Larbi A, Pawelec G, et al. Immunology of aging: the birth of inflammaging. Clin Rev Allergy Immunol. 2021. doi:10.1007/s12016-021-08899-6
50. Farage MA, Miller KW, Elsner P, Maibach HI. Intrinsic and extrinsic factors in skin ageing: a review. Int J Cosmet Sci. 2008;30(2):87–95. doi:10.1111/j.1468-2494.2007.00415.x
51. Lebel M. Werner syndrome: genetic and molecular basis of a premature aging disorder. Cell Mol Life Sci CMLS. 2001;58(7):857–867. doi:10.1007/s00018-001-8398-y
52. Rizza ERH, DiGiovanna JJ, Khan SG, Tamura D, Jeskey JD, Kraemer KH. Xeroderma pigmentosum: a model for human premature aging. J Invest Dermatol. 2021;141(4):976–984. doi:10.1016/j.jid.2020.11.012
53. Hussain M, Krishnamurthy S, Patel J, et al. Skin abnormalities in disorders with DNA repair defects, premature aging, and mitochondrial dysfunction. J Invest Dermatol. 2021;141(4S):968–975. doi:10.1016/j.jid.2020.10.019
54. Han A, Chien AL, Kang S. Photoaging. Dermatol Clin. 2014;32(3):291–299, vii. doi:10.1016/j.det.2014.03.015
55. Cao C, Xiao Z, Wu Y, Ge C. Diet and skin aging—from the perspective of food nutrition. Nutrients. 2020;12(3):870. doi:10.3390/nu12030870
56. Schikowski T, Hüls A. Air pollution and skin aging. Curr Environ Health Rep. 2020;7(1):58–64. doi:10.1007/s40572-020-00262-9
57. Martic I, Jansen-Dürr P, Cavinato M. Effects of air pollution on cellular senescence and skin aging. Cells. 2022;11:14. doi:10.3390/cells11142220
58. Sundelin T, Lekander M, Kecklund G, Van Someren EJW, Olsson A, Axelsson J. Cues of fatigue: effects of sleep deprivation on facial appearance. Sleep. 2013;36(9):1355–1360. doi:10.5665/sleep.2964
59. Oyetakin-White P, Suggs A, Koo B, et al. Does poor sleep quality affect skin ageing? Clin Exp Dermatol. 2015;40(1):17–22. doi:10.1111/ced.12455
60. Ichibori R, Fujiwara T, Tanigawa T, et al. Objective assessment of facial skin aging and the associated environmental factors in Japanese monozygotic twins. J Cosmet Dermatol. 2014;13(2):158–163. doi:10.1111/jocd.12081
61. Russell-Goldman E, Murphy GF. The pathobiology of skin aging: new insights into an old dilemma. Am J Pathol. 2020;190(7):1356–1369. doi:10.1016/j.ajpath.2020.03.007
62. Furue M, Kadono T. “Inflammatory skin march” in atopic dermatitis and psoriasis. Inflamm Res off J Eur Histamine Res Soc Al. 2017;66(10):833–842. doi:10.1007/s00011-017-1065-z
63. Zouboulis CC, Ganceviciene R, Liakou AI, Theodoridis A, Elewa R, Makrantonaki E. Aesthetic aspects of skin aging, prevention, and local treatment. Clin Dermatol. 2019;37(4):365–372. doi:10.1016/j.clindermatol.2019.04.002
64. Schwartz J, Friedman AJ. Exogenous factors in skin barrier repair. J Drugs Dermatol JDD. 2016;15(11):1289–1294.
65. Rosso JD, Zeichner J, Alexis A, Cohen D, Berson D. Understanding the epidermal barrier in healthy and compromised skin: clinically relevant information for the dermatology practitioner. J Clin Aesthetic Dermatol. 2016;9(4 Suppl 1):S2–S8.
66. Feingold KR, Elias PM. Role of lipids in the formation and maintenance of the cutaneous permeability barrier. Biochim Biophys Acta. 2014;1841(3):280–294. doi:10.1016/j.bbalip.2013.11.007
67. Kumar V, Bouameur JE, Bär J, et al. A keratin scaffold regulates epidermal barrier formation, mitochondrial lipid composition, and activity. J Cell Biol. 2015;211(5):1057–1075. doi:10.1083/jcb.201404147
68. Kim Y, Lim KM. Skin barrier dysfunction and filaggrin. Arch Pharm Res. 2021;44(1):36–48. doi:10.1007/s12272-021-01305-x
69. Ishitsuka Y, Roop DR. Loricrin at the boundary between inside and outside. Biomolecules. 2022;12(5):673. doi:10.3390/biom12050673
70. Cuollo L, Antonangeli F, Santoni A, Soriani A. The Senescence-Associated Secretory Phenotype (SASP) in the Challenging Future of Cancer Therapy and Age-Related Diseases. Biology. 2020;9(12):485. doi:10.3390/biology9120485
71. Ghosh K, Capell BC. The senescence-associated secretory phenotype: critical effector in skin cancer and aging. J Invest Dermatol. 2016;136(11):2133–2139. doi:10.1016/j.jid.2016.06.621
72. Zhang S, Duan E. Fighting against skin aging: the way from bench to bedside. Cell Transplant. 2018;27(5):729–738. doi:10.1177/0963689717725755
73. Xu M, Tchkonia T, Ding H, et al. JAK inhibition alleviates the cellular senescence-associated secretory phenotype and frailty in old age. Proc Natl Acad Sci. 2015;112(46):E6301–E6310. doi:10.1073/pnas.1515386112
74. Laberge RM, Zhou L, Sarantos MR, et al. Glucocorticoids suppress selected components of the senescence-associated secretory phenotype. Aging Cell. 2012;11(4):569–578. doi:10.1111/j.1474-9726.2012.00818.x
75. Jeon S, Choi M. Anti-inflammatory and anti-aging effects of hydroxytyrosol on human dermal fibroblasts (HDFs). Biomed Dermatol. 2018;2(1):21. doi:10.1186/s41702-018-0031-x
76. Milosheska D, Roškar R. Use of retinoids in topical antiaging treatments: a focused review of clinical evidence for conventional and nanoformulations. Adv Ther. 2022;39(12):5351–5375. doi:10.1007/s12325-022-02319-7
77. Widgerow AD, Fabi SG, Palestine RF, et al. Extracellular matrix modulation: optimizing skin care and rejuvenation procedures. J Drugs Dermatol JDD. 2016;15(4 Suppl):s63–71.
78. Cole MA, Quan T, Voorhees JJ, Fisher GJ. Extracellular matrix regulation of fibroblast function: redefining our perspective on skin aging. J Cell Commun Signal. 2018;12(1):35–43. doi:10.1007/s12079-018-0459-1
79. Lee JH, Park J, Shin DW. The molecular mechanism of polyphenols with anti-aging activity in aged human dermal fibroblasts. Mol Basel Switz. 2022;27(14):4351. doi:10.3390/molecules27144351
80. Sárdy M. Role of matrix metalloproteinases in skin ageing. Connect Tissue Res. 2009;50(2):132–138. doi:10.1080/03008200802585622
81. Enescu CD, Bedford LM, Potts G, Fahs F. A review of topical vitamin C derivatives and their efficacy. J Cosmet Dermatol. 2022;21(6):2349–2359. doi:10.1111/jocd.14465
82. Rossetti D, Kielmanowicz MG, Vigodman S, et al. A novel anti-ageing mechanism for retinol: induction of dermal elastin synthesis and elastin fibre formation. Int J Cosmet Sci. 2011;33(1):62–69. doi:10.1111/j.1468-2494.2010.00588.x
83. Wang ST, Neo BH, Betts RJ. Glycosaminoglycans: sweet as sugar targets for topical skin anti-aging. Clin Cosmet Investig Dermatol. 2021;14:1227–1246. doi:10.2147/CCID.S328671
84. Gkogkolou P, Böhm M. Advanced glycation end products: key players in skin aging? Dermatoendocrinol. 2012;4(3):259–270. doi:10.4161/derm.22028
85. Chen C, Zhang JQ, Li L, et al. Advanced Glycation End Products in the Skin: molecular Mechanisms, Methods of Measurement, and Inhibitory Pathways. Front Med. 2022;9:e46.
86. Mishra SK, Balendra V, Esposto J, et al. Therapeutic Antiaging Strategies. Biomedicines. 2022;10(10):2515. doi:10.3390/biomedicines10102515
87. Martínez P, Blasco MA. Role of shelterin in cancer and aging. Aging Cell. 2010;9(5):653–666. doi:10.1111/j.1474-9726.2010.00596.x
88. Tsoukalas D, Fragkiadaki P, Docea AO, et al. Discovery of potent telomerase activators: unfolding new therapeutic and anti-aging perspectives. Mol Med Rep. 2019;20(4):3701–3708. doi:10.3892/mmr.2019.10614
89. Bernardes de Jesus B, Vera E, Schneeberger K, et al. Telomerase gene therapy in adult and old mice delays aging and increases longevity without increasing cancer. EMBO Mol Med. 2012;4(8):691–704. doi:10.1002/emmm.201200245
90. Aydin Y. Antiaging Strategies Based on Telomerase Activity. In: Rizvi SI, Çakatay U editors. Molecular Basis and Emerging Strategies for Anti-Aging Interventions. Springer; 2018:97–109. doi:10.1007/978-981-13-1699-9_7
91. Kuo IY, Ehrlich BE. Signaling in Muscle Contraction. Cold Spring Harb Perspect Biol. 2015;7(2):a006023. doi:10.1101/cshperspect.a006023
92. Kim B, Cho HE, Moon SH, et al. Transdermal delivery systems in cosmetics. Biomed Dermatol. 2020;4(1):10. doi:10.1186/s41702-020-0058-7
93. Cosgrove MC, Franco OH, Granger SP, Murray PG, Mayes AE. Dietary nutrient intakes and skin-aging appearance among middle-aged American women. Am J Clin Nutr. 2007;86(4):1225–1231. doi:10.1093/ajcn/86.4.1225
94. Draelos ZD, Oral A. Supplement and the nutrition–skin connection. J Clin Aesthetic Dermatol. 2019;12(7):13–16.
95. Chen J, Waqas K, Tan RC, et al. The association between dietary and skin advanced glycation end products: the Rotterdam Study. Am J Clin Nutr. 2020;112(1):129–137. doi:10.1093/ajcn/nqaa117
96. Jhawar N, Wang JV, Saedi N. Oral collagen supplementation for skin aging: a fad or the future? J Cosmet Dermatol. 2020;19(4):910–912. doi:10.1111/jocd.13096
97. Al-Atif H. Collagen supplements for aging and wrinkles: a paradigm shift in the fields of dermatology and cosmetics. Dermatol Pract Concept. 2022;12(1):e2022018. doi:10.5826/dpc.1201a18
98. Sun Q, Wu J, Qian G, Cheng H. Effectiveness of dietary supplement for skin moisturizing in healthy adults: a systematic review and meta-analysis of randomized controlled trials. Front Nutr. 2022;9:895192. doi:10.3389/fnut.2022.895192
99. Hsu TF, Su ZR, Hsieh YH, et al. Oral Hyaluronan Relieves Wrinkles and Improves Dry Skin: a 12-Week Double-Blinded, Placebo-Controlled Study. Nutrients. 2021;13(7):2220. doi:10.3390/nu13072220
100. Jo S, Jung Y-S, Cho Y-R, et al. Oral Administration of Rosa gallica Prevents UVB−Induced Skin Aging through Targeting the c−Raf Signaling Axis. Antioxidants. 2021;10(11):1663. doi:10.3390/antiox10111663
101. Thye AY-K, Bah Y-R, Law JW-F, et al. Gut–skin axis: unravelling the connection between the gut microbiome and psoriasis. Biomedicines. 2022;10(5):1037. doi:10.3390/biomedicines10051037
102. Sinha S, Lin G, Ferenczi K. The skin microbiome and the gut-skin axis. Clin Dermatol. 2021;39(5):829–839. doi:10.1016/j.clindermatol.2021.08.021
103. Trompette A, Pernot J, Perdijk O, et al. Gut-derived short-chain fatty acids modulate skin barrier integrity by promoting keratinocyte metabolism and differentiation. Mucosal Immunol. 2022;15(5):908–926. doi:10.1038/s41385-022-00524-9
104. He B, Chen Y, Yu S, Hao Y, Wang F, Qu L. Food plant extracts for sleep-related skin health: mechanisms and prospects. Food Biosci. 2022;49:101951. doi:10.1016/j.fbio.2022.101951
105. He H, Tang J, Ru D, et al. Protective effects of Cordyceps extract against UVB‑induced damage and prediction of application prospects in the topical administration: an experimental validation and network pharmacology study. Biomed Pharmacother. 2020;121:109600. doi:10.1016/j.biopha.2019.109600
106. Huang TT, Chong KY, Ojcius DM, et al. Hirsutella sinensis mycelium suppresses interleukin-1β and interleukin-18 secretion by inhibiting both canonical and non-canonical inflammasomes. Sci Rep. 2013;3:1374. doi:10.1038/srep01374
107. Park JM, Lee JS, Lee KR, Ha SJ, Hong EK. Cordyceps militaris extract protects human dermal fibroblasts against oxidative stress-induced apoptosis and premature senescence. Nutrients. 2014;6(9):3711–3726. doi:10.3390/nu6093711
108. Prommaban A, Sriyab S, Marsup P, et al. Comparison of chemical profiles, antioxidation, inhibition of skin extracellular matrix degradation, and anti-tyrosinase activity between mycelium and fruiting body of Cordyceps militaris and Isaria tenuipes. Pharm Biol. 2022;60(1):225. doi:10.1080/13880209.2021.2025255
109. You S, Jang M, Kim GH. Mori Cortex radicis extract protected against diet-induced neuronal damage by suppressing the AGE-RAGE/MAPK signaling pathway in C. elegans and mouse model. J Funct Foods. 2022;91:104996. doi:10.1016/j.jff.2022.104996
110. Seo CS, Lim HS, Jeong SJ, Ha H, Shin HK. HPLC-PDA analysis and anti-inflammatory effects of mori cortex radicis. Nat Prod Commun. 2013;8(10):1934578X1300801027. doi:10.1177/1934578X1300801027
111. Lu YE, Chen YJ. Resveratrol inhibits matrix metalloproteinase-1 and −3 expression by suppressing of p300/NFκB acetylation in TNF-α-treated human dermal fibroblasts. Chem Biol Interact. 2021;337:109395. doi:10.1016/j.cbi.2021.109395
112. Demarne F, Passaro G Use of an Acmella oleracea extract for the botulinum toxin-like effect thereof in an anti-wrinkle cosmetic composition. Available from: https://patents.google.com/patent/US7531193B2/en.
113. Wang R, Lechtenberg M, Sendker J, Petereit F, Deters A, Hensel A. Wound-healing plants from TCM: in vitro investigations on selected TCM plants and their influence on human dermal fibroblasts and keratinocytes. Fitoterapia. 2013;84:308–317. doi:10.1016/j.fitote.2012.12.020
114. Lee S, Lim JM, Jin MH, et al. Partially purified paeoniflorin exerts protective effects on UV-induced DNA damage and reduces facial wrinkles in human skin. J Cosmet Sci. 2006;57(1):57–64.
115. Kong L, Wang S, Wu X, Zuo F, Qin H, Wu J. Paeoniflorin attenuates ultraviolet B-induced apoptosis in human keratinocytes by inhibiting the ROS-p38-p53 pathway. Mol Med Rep. 2016;13(4):3553–3558. doi:10.3892/mmr.2016.4953
116. Yang L, Xing S, Wang K, Yi H, Du B. Paeonol attenuates aging MRC-5 cells and inhibits epithelial-mesenchymal transition of premalignant HaCaT cells induced by aging MRC-5 cell-conditioned medium. Mol Cell Biochem. 2018;439(1–2):117–129. doi:10.1007/s11010-017-3141-7
117. Sun Z, Du J, Hwang E, Yi TH. Paeonol extracted from Paeonia suffruticosa Andr. ameliorated UVB-induced skin photoaging via DLD/Nrf2/ARE and MAPK/AP-1 pathway. Phytother Res PTR. 2018;32(9):1741–1749. doi:10.1002/ptr.6100
118. Obayashi K, Kurihara K, Okano Y, Masaki H, Yarosh DB. L-Ergothioneine scavenges superoxide and singlet oxygen and suppresses TNF-alpha and MMP-1 expression in UV-irradiated human dermal fibroblasts. J Cosmet Sci. 2005;56(1):17–27.
119. Ko HJ, Kim J, Ahn M, Kim JH, Lee GS, Shin T. Ergothioneine alleviates senescence of fibroblasts induced by UVB damage of keratinocytes via activation of the Nrf2/HO-1 pathway and HSP70 in keratinocytes. Exp Cell Res. 2021;400(1):112516. doi:10.1016/j.yexcr.2021.112516
120. Paul BD, Snyder SH. The unusual amino acid L-ergothioneine is a physiologic cytoprotectant. Cell Death Differ. 2010;17(7):1134–1140. doi:10.1038/cdd.2009.163
121. Bazela K, Solyga-Zurek A, Debowska R, Rogiewicz K, Bartnik E, Eris I. l-Ergothioneine Protects Skin Cells against UV-Induced Damage—A Preliminary Study. Cosmetics. 2014;1(1):51–60. doi:10.3390/cosmetics1010051
122. Wanyonyi S. Kappaphycus alvarezii as a Food Supplement Prevents Diet-Induced Metabolic Syndrome in Rats. Nutrients. 2017;9(11):1261. doi:10.3390/nu9111261
123. Yulianti E. The effect of Kappaphycus alvarezii active fraction on oxidative stress and inflammation in streptozotocin and nicotinamide-induced diabetic rats. BMC Complement Med Ther. 2022;22(1):15. doi:10.1186/s12906-021-03496-8
124. Abu Bakar N, Anyanji VU, Mustapha NM, Lim SL, Mohamed S. Seaweed (Eucheuma cottonii) reduced inflammation, mucin synthesis, eosinophil infiltration and MMP-9 expressions in asthma-induced rats compared to Loratadine. J Funct Foods. 2015;19:710–722. doi:10.1016/j.jff.2015.10.011
125. Paufique J Use of an active principle derived from eucheuma cottonii and rich in linear galactans for controlling skin cell aging. 2014. Available from: https://patents.google.com/patent/EP2811974A2/en.
126. Yue K, Ye M, Zhou Z, Sun W, Lin X. The genus Cordyceps: a chemical and pharmacological review. J Pharm Pharmacol. 2013;65(4):474–493. doi:10.1111/j.2042-7158.2012.01601.x
127. Bayazid AB, Kim JG, Park SH, Lim BO. Antioxidant, anti-inflammatory, and antiproliferative activity of mori cortex radicis extracts. Nat Prod Commun. 2020;15(1):1934578X19899765. doi:10.1177/1934578X19899765
128. Chan EWC, Lye PY, Wong SK. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin J Nat Med. 2016;14(1):17–30. doi:10.3724/SP.J.1009.2016.00017
129. Li Z, Chen X, Liu G, et al. Antioxidant Activity and Mechanism of Resveratrol and Polydatin Isolated from Mulberry (Morus alba L). Mol Basel Switz. 2021;26(24):7574. doi:10.3390/molecules26247574
130. Zeng HJ, Li QY, Ma J, Yang R, Qu LB. A comparative study on the effects of resveratrol and oxyresveratrol against tyrosinase activity and their inhibitory mechanism. Spectrochim Acta A Mol Biomol Spectrosc. 2021;251:119405. doi:10.1016/j.saa.2020.119405
131. Hecker A, Schellnegger M, Hofmann E, et al. The impact of resveratrol on skin wound healing, scarring, and aging. Int Wound J. 2022;19(1):9–28. doi:10.1111/iwj.13601
132. Wang XB, Zhu L, Huang J, et al. Resveratrol-induced augmentation of telomerase activity delays senescence of endothelial progenitor cells. Chin Med J. 2011;124(24):4310–4315.
133. Giovannelli L, Pitozzi V, Jacomelli M, et al. Protective effects of resveratrol against senescence-associated changes in cultured human fibroblasts. J Gerontol Ser A. 2011;66A(1):9–18. doi:10.1093/gerona/glq161
134. Aires V, Delmas D, Le Bachelier C, et al. Stilbenes and resveratrol metabolites improve mitochondrial fatty acid oxidation defects in human fibroblasts. Orphanet J Rare Dis. 2014;9(1):79. doi:10.1186/1750-1172-9-79
135. Rondanelli M, Fossari F, Vecchio V, et al. Acmella oleracea for pain management. Fitoterapia. 2020;140:104419. doi:10.1016/j.fitote.2019.104419
136. Barbosa AF, de Carvalho MG, Smith RE, Sabaa-Srur AUO. Spilanthol: occurrence, extraction, chemistry and biological activities. Rev Bras Farmacogn. 2016;26(1):128–133. doi:10.1016/j.bjp.2015.07.024
137. Boonen J, Baert B, Roche N, Burvenich C, De Spiegeleer B. Transdermal behaviour of the N-alkylamide spilanthol (affinin) from Spilanthes acmella (Compositae) extracts. J Ethnopharmacol. 2010;127(1):77–84. doi:10.1016/j.jep.2009.09.046
138. Moro SD, de Oliveira Fujii L, Teodoro LFR, et al. Acmella oleracea extract increases collagen content and organization in partially transected tendons. Microsc Res Tech. 2021;84(11):2588–2597. doi:10.1002/jemt.23809
139. Wang Z, He C, Peng Y, Chen F, Xiao P. Origins, Phytochemistry, Pharmacology, Analytical Methods and Safety of Cortex Moutan (Paeonia suffruticosa Andrew): a Systematic Review. Mol J Synth Chem Nat Prod Chem. 2017;22(6):946. doi:10.3390/molecules22060946
140. Borodina I, Kenny LC, McCarthy CM, et al. The biology of ergothioneine, an antioxidant nutraceutical. Nutr Res Rev. 2020;33(2):190–217. doi:10.1017/S0954422419000301
141. Kalasariya HS, Yadav VK, Yadav KK, et al. Seaweed-based molecules and their potential biological activities: an eco-sustainable cosmetics. Molecules. 2021;26(17):5313. doi:10.3390/molecules26175313
142. Shah MD, Venmathi Maran BA, Shaleh SRM, Zuldin WH, Gnanaraj C, Yong YS. Therapeutic potential and nutraceutical profiling of north Bornean seaweeds: a review. Mar Drugs. 2022;20(2):101. doi:10.3390/md20020101
143. Wardani G, Farida N, Andayani R, Kuntoro M, Sudjarwo SA. The Potency of Red Seaweed (Eucheuma cottonii) Extracts as Hepatoprotector on Lead Acetate-induced Hepatotoxicity in Mice. Pharmacogn Res. 2017;9(3):282–286. doi:10.4103/pr.pr_69_16
144. Kim I, Kim S, Manggau M, et al. Red seaweed (Eucheuma cottonii) extract promotes human keratinocyte migration via p38 mitogen-activated protein kinase phosphorylation. Pharmacogn Mag. 2020;16:192. doi:10.4103/pm.pm_203_19
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.