Back to Journals » Journal of Pain Research » Volume 18
Shared Genetic Architecture of Obstructive Sleep Apnea and Neck Pain: A Large-Scale Genome-Wide Association Study
Authors Yang F ![]() , Zhu D, Yu R, Zhang Z, He J
, Zhu D, Yu R, Zhang Z, He J
Received 30 April 2025
Accepted for publication 5 November 2025
Published 1 December 2025 Volume 2025:18 Pages 6423—6436
DOI https://doi.org/10.2147/JPR.S534380
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor King Hei Stanley Lam
Fangjie Yang,1,* Dan Zhu,2,* Renyu Yu,3 Zhiyi Zhang,4 Jian He2,5
1Fujian Vocational College of Bioengineering, Fuzhou, Fujian Province, 350007,People’s Republic of China; 2Fujian University of Traditional Chinese Medicine, Fuzhou, Fujian Province, 350122, People’s Republic of China; 3The Affiliated People’s Hospital of Fujian University of Traditional Chinese Medicine, Fuzhou, Fujian Province, 350004, People’s Republic of China; 4Department of Massage, Quanzhou Orthopedic-Traumatological Hospital, Quanzhou, Fujian Province, 362000, People’s Republic of China; 5Zhangzhou Health Vocational College, Zhangzhou, Fujian Province, 363000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jian He, Email [email protected]
Background: This study aims to explore the role of genetic factors in the co-occurrence of neck pain and obstructive sleep apnea (OSA), thereby providing a deeper understanding of their intrinsic relationship.
Methods: We analyzed GWAS data for OSA from the FinnGen consortium (38,998 cases and 336,659 controls), and neck pain from the UK Biobank (106,521 cases and 355,336 controls), all of European ancestry. Genetic correlation between neck pain and OSA using the Linkage Disequilibrium Score Regression (LDSC) method and High-definition Likelihood (HDL) method. Local correlation was examined across 2,795 loci with Local Analysis of Variant Association (LAVA). Mendelian randomization (MR) was conducted to infer causality. Cross-Phenotype Association (CPASSOC) identified pleiotropic SNPs, which were annotated with FUMA and validated by MAGMA. Transcriptome-wide association study (TWAS) and tissue enrichment analyses were further applied to explore biological mechanisms.
Results: MR analysis indicated that NP increases the risk of OSA. CPASSOC identified 150 pleiotropic SNPs, with 40 loci annotated by FUMA, yielding 85 pleiotropic genes consistent with MAGMA. TWAS highlighted seven genes with significant pleiotropic effects on both NP and OSA. Tissue enrichment revealed that pleiotropic gene expression was predominantly localized in brain tissues, particularly the cerebral cortex (P=2.17E-6). These genes suggested involvement of neurodevelopmental and metabolic pathways in neck pain-OSA comorbidity. In MR,we found that individuals with neck pain had an increased risk of developing OSA.
Conclusion: Our findings underscore the pivotal role of genetic factors in neck pain–OSA comorbidity and provide theoretical support for considering pain management as part of clinical strategies for OSA.
Keywords: GWAS, neck pain, OSA, genetic correlation
Introduction
Neck pain refers to discomfort or pain localized in the cervical region, typically involving the cervical vertebrae, muscles, ligaments, nerves, and surrounding tissues.1 With the advent of modern lifestyles, including prolonged sitting at work and extensive use of electronic devices, the prevalence of neck pain has increased, with an average rate as high as 23.1%.2 Obstructive Sleep Apnea (OSA) is a common sleep disorder characterized by the repeated collapse of the upper airway during sleep, leading to episodes of apnea and hypopnea. This condition is not only associated with various health issues such as cardiovascular diseases, hypertension, and type 2 diabetes, but it also negatively impacts the patient’s quality of life.
Previous research has identified that sleep disturbances serve as an independent risk factor for long-term activity-limiting neck pain.3 Poor sleep quality or insufficient sleep has been strongly associated with the incidence of neck pain, with this relationship particularly pronounced among women.4,5 Moreover, sleep deprivation has been shown to heighten pain sensitivity,6 while chronic pain may lead to psychological consequences such as anxiety, stress, and depression, which in turn further disrupt sleep patterns.7 OSA is one of the most common sleep disorders, yet research on its association with neck pain is still limited.8 Individuals with OSA often unconsciously adjust their head and neck position during sleep to reduce airway resistance. These adjustments can impose excessive mechanical strain on cervical muscles and joints, potentially contributing to neck pain.9 Additionally, the sympathetic nervous system overactivation caused by OSA can further increase muscle tension and pain in the neck and shoulder areas.10 Beyond mechanical and autonomic factors, other shared biological mechanisms may also underlie the relationship between OSA and neck pain. Systemic inflammation appears to be a key link between OSA and neck pain.11 Studies have shown that OSA can induce a chronic low-grade inflammatory state, characterized by elevated circulating levels of pro-inflammatory cytokines such as IL-6, TNF-α, and CRP. This inflammatory response may increase nociceptive sensitivity and, in turn, exacerbate musculoskeletal pain.12 Furthermore, abnormalities in central pain processing represent another potential mechanism.13 Neuroimaging studies further reveal structural brain changes, especially reduced gray matter volume in pain-related regions such as the thalamus, prefrontal cortex, and insula.14 These neural alterations may impair pain modulation and increase susceptibility to chronic pain conditions, including neck pain.15 Despite growing evidence suggesting a potential association between OSA and neck pain, the physiological and pathophysiological mechanisms connecting the two remain inadequately explored. Future research should prioritize a deeper exploration of the underlying mechanisms of this association.
Compared to the limited clinical research on the co-occurrence of OSA and neck pain, their genetic underpinnings have received growing attention with the advancement of genome-wide association studies (GWAS).16,17 Shi et al identified new genetic and immune-related biomarkers linked to OSA, which indicate potential involvement of inflammatory processes in disease development.18 Similarly, Xu et al conducted a GWAS in a Han Chinese population and discovered new risk loci associated with sleep traits, highlighting the potential for population-specific genetic factors.19 Environmental and lifestyle factors such as obesity, physical inactivity, and poor sleep hygiene may influence gene expression and interact with genetic susceptibility loci, thereby jointly increasing the risk of both OSA and neck pain. Accordingly, investigating the genetic overlap between these conditions may help uncover shared biological mechanisms. However, research on the common genetic basis of OSA and neck pain remains limited. Such analyses could not only enhance our understanding of their comorbidity but also inform integrated strategies for prevention and treatment.20,21
This study aims to investigate the genetic correlation between OSA and neck pain. We performed cross-trait analysis to identify pleiotropic single nucleotide polymorphisms (SNPs) and shared genes associated with both conditions. We further conducted tissue-specific enrichment analysis to explore the potential biological relevance of these genes. Lastly, Mendelian randomization (MR) was applied to assess the potential causal relationship between OSA and neck pain, thereby providing stronger evidence for a genetic link between the two traits.
Method
The Source of GWAS
The GWAS data for both traits were derived from European samples. Summary GWAS data for OSA were sourced from the FinnGen consortium R9 (https://www.finngen.fi/en/access_results), encompassing a large cohort of 38,998 cases and 336,659 controls. Neck pain GWAS data were obtained from the UK Biobank, with the case group consisting of 106,521 individuals and the control group comprising 355,336 healthy participants (https://gwas.mrcieu.ac.uk/datasets/ukb-b-18596/). Then, the data underwent rigorous cleaning, including standardization of major effect alleles, removal of duplicate SNPs, and supplementation of missing genetic information, such as rsID numbers and chromosome positions. The cleaned GWAS data were then aligned with the European 1000 Genomes Project to ensure consistency and accuracy across shared regions. These steps were undertaken to ensure data reliability and provide a robust foundation for subsequent genetic analyses (Figure 1).
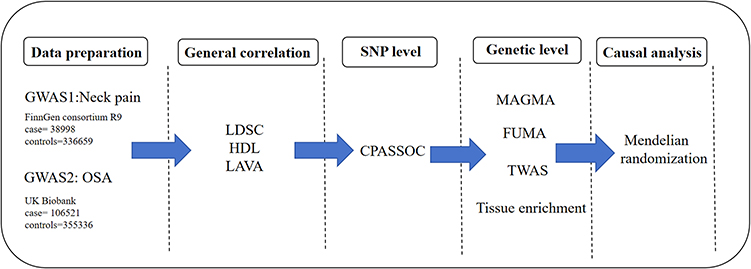 |
Figure 1 Flowchart of the Genetic Architecture Analysis of Neck Pain and OSA. |
General Genetic Correlation
Linkage Disequilibrium Score Regression22 (LDSC) is a statistical method used in genomic studies to analyze data from GWAS. This method leverages linkage disequilibrium (LD) scores across the genome to help researchers differentiate between genetic and environmental contributions to phenotypic variation. Through regression analysis, LDSC can estimate the heritability of specific phenotypes and reveal the widespread distribution of genetic effects. This approach is particularly valuable in studies of the genetic underpinnings of complex traits, offering insights into the overall impact of genetic factors.In addition, we employed the High-Definition Likelihood (HDL) method to validate the genetic correlation results between Neck pain and OSA that were obtained through LDSC. By modeling LD structure more precisely, HDL efficiently utilizes information from GWAS summary data, particularly in cases with smaller sample sizes or weaker genetic signals. Compared to LDSC, HDL reduces the variance of genetic correlation estimates by approximately 60%, which is equivalent to increasing the sample size by 150 when using LDSC. We conducted the HDL analysis using the R package HDL-v1.4.0 (https://github.com/zhenin/HDL), utilizing 1,029,876 well-imputed HapMap3 SNPs as the reference panel (https://github.com/zhenin/HDL/wiki/Reference-panels).
Local Genetic Correlation
Genetic correlation analysis is conducted across the entire genome, considering only the average of shared signals within the genome. However, this approach may not effectively identify genetic correlations when they are restricted to specific genomic regions or when opposing directions are present at different loci.23 Local Analysis of Variance Approach (LAVA) is an integrated framework for local genetic correlation analysis that tests standard bivariate local genetic correlations and is capable of analyzing both binary and continuous phenotypes, even with varying degrees of sample overlap. We performed pairwise local rgs analysis across 2,495 genomic loci. The Bonferroni-corrected P threshold is 0.05/2495 = 2.00E-05.
CPASSOC
Next, we analyzed the genetic overlap between OSA and neck pain at the SNP level. Cross-Phenotype Association analysis (CPASSOC) is a novel statistical method designed to identify genetic variants associated with multiple phenotypes.24 By evaluating the genetic associations of multiple phenotypes within the same analytical framework, CPASSOC can reveal the shared genetic mechanisms underlying complex traits based on the meta-analysis. It utilizes two key statistics (SHom and SHet) to assess shared and distinct genetic effects across phenotypes, allowing for a comprehensive understanding of the genetic architecture underlying multiple traits.
MAGMA
Gene-level analysis offers the advantage of capturing broader genetic effects and interactions, providing a more comprehensive understanding of genetic influences compared to SNP-level analysis. We explored the genetic architecture of OSA and neck pain at the genetic level. Multi-marker Analysis of GenoMic Annotation (MAGMA)25 is a method that can assess the association between genes and traits using multiple linear regression models based on principal component analysis. Initially, we used Functional Mapping and Annotation (FUMA)24 to annotate Pleiotropic SNPs identified by CPASSOC located in close proximity within genomic regions, linking them to the corresponding genes. Following this, we conducted a gene- and trait-based association analysis with MAGMA, utilizing principal component regression models and the 1000 Genomes European reference sample to enhance the power of detection. We selected the overlapping genes from the two methods and performed corrections to derive the final results. We conducted a tissue enrichment analysis utilizing the FUMA platform, incorporating data from 54 tissue types provided by GTEx (v.8).
TWAS
Based on GWAS results, we can identify associations between SNPs and traits; however, these findings only point to candidate chromosomal regions and cannot precisely pinpoint the causative genes. To address this limitation, we performed a transcriptome-wide association study (TWAS) analysis26 to investigate the shared gene expression structure between OSA and neck pain. TWAS employs transcriptional regulation (expression) as a mediator between genetic variation (genotype) and phenotype, thereby transforming the SNP-phenotype association into a gene/transcript-phenotype association.
First, we conducted TWAS separately for each trait: 1) Constructing the relationship between SNPs and gene expression using a reference panel, 2) Predicting the gene expression levels for OSA and neck pain based on this relationship, and 3) Analyzing the association between genes and traits using the imputed gene expression levels. We then selected genes that were significantly expressed in both traits (PFDR < 0.05) as candidate pleiotropic genes for OSA and neck pain at the gene expression level. For the reference tissue, we utilized the GTEx V8 dataset, which provides extensive gene expression data from 49 different tissues, collected from 838 post-mortem donors (https://ftp.ebi.ac.uk/pub/databases/spot/eQTL/imported/GTEx_V8).
MR
We explored the causal link between OSA and neck pain by employing a bidirectional MR27 approach, leveraging genetic variants as instrumental variables (IV) to determine causality. A valid IV requires a strong association with the exposure while remaining independent of the outcome, typically meeting a significance threshold of P < 5E-8, with clumping parameters set at R2 < 0.001 (clumping window size = 1000kb). IVs with F-values under 10 are deemed weak. The inverse variance weighting (IVW) method was employed as our primary MR analysis, complemented by MR-Egger and weighted median methods to validate results under less stringent assumptions. To ensure robustness and reliability, we also performed pleiotropy and heterogeneity analyses. Additionally, the sensitivity of the results was tested using the leave-one-out method, where each SNP was sequentially removed to observe its impact on the overall meta-effects.
Enrichment
Enrichment analyses were conducted to characterize the functions of shared genes. GO analysis classified them into biological processes, molecular functions, and cellular components, while KEGG mapping revealed their involvement in signaling and metabolic pathways. These results provided mechanistic insights and identified biologically relevant networks linked to the traits of interest.
Results
Genetic Correlation
Using European populations from different cohorts, LDSC revealed a heritability of 0.0324 for neck pain and 0.0438 for OSA. Additionally, a significant positive genetic correlation was found between OSA and neck pain (rg = 0.068, P = 3.0E-04).Consistent with this, the HDL analysis revealed a heritability of 0.0306 for neck pain and 0.027 for OSA, with a significant positive genetic correlation between the two traits (rg = 0.355, P = 1.51E-22).
The LAVA analysis revealed significant positive genetic correlations between neck pain and OSA at three regions: chromosome (CHR): position (BP) = 5:177,659,903~178595252 (P=1.29e-05, rho=1); CHR:BP=11:121,208,187~122589224 (P=2.55e-05, rho=0.79), and CHR:BP=3:139,831,268~141034637 (P=2.58e-05, rho=0.92) (Figure 2). Although these loci showing significant local rgs have not been annotated to specific genes, their strong positive associations suggest that they may harbor previously unrecognized regulatory elements that influence the expression of genes involved in neck muscle tone, neural signaling, or inflammatory pathways.
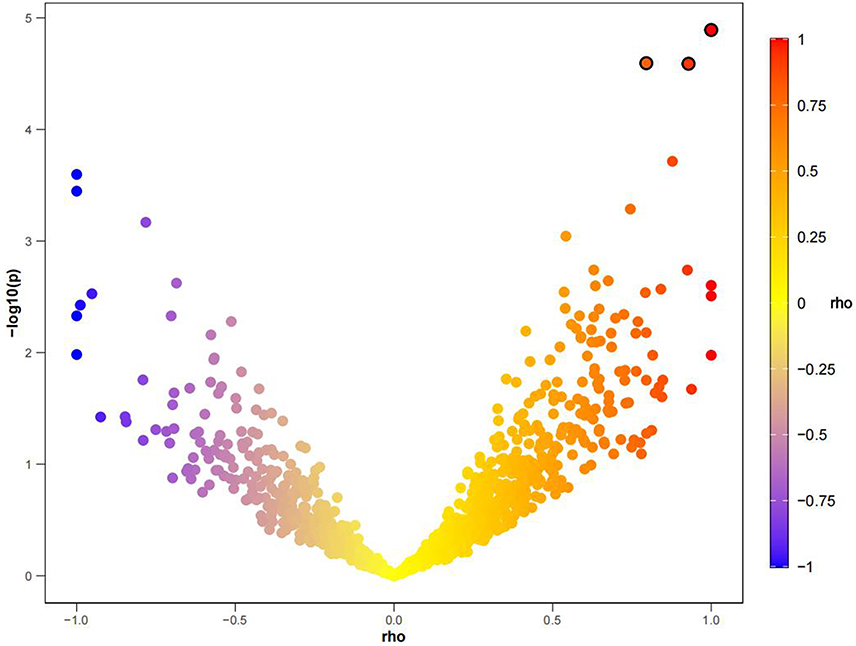 |
Figure 2 The scatter plot of the bivariate LAVA analysis results for OSA and neck pain. |
CPASSOC
We identified a total of 150 pleiotropic SNPs that met the criteria of Pmeta < 5E-08, with individual trait significance thresholds of P < 0.001 for both OSA and neck pain. The most significant SNP was rs8084619 (POSA = 3.50E-07, Pneck pain = 4.11E-05, PCPASSOC = 9.68E-11), located at CHR:BP = 18:31208660. The second most significant SNP was rs6507051 (POSA = 3.60E-07, Pneck pain = 4.18E-05, PCPASSOC = 1.02E-10), located at CHR:BP= 18:31208453. The third most significant SNP was rs12957987 (POSA = 3.60E-07, Pneck pain = 4.39E-05, PCPASSOC = 1.02E-10), located at CHR:BP= 18:31212209 (Supplementary Table 01).
Pleiotropic Genes
Using CPASSOC analysis, we identified pleiotropic SNPs, which were then annotated on the FUMA platform, resulting in 40 loci (Table 1). Subsequently, MAGMA analysis identified 85 overlapping genes (Figure 3,4, Supplementary Table 02), of which 36, consistent with the FUMA annotation results (Supplementary Table 03), remained significant after Bonferroni correction (P < 5.81E-04). Among these, the gene most significantly associated with both OSA and neck pain is FTO (PMAGMA = 1.12E-11), located at CHR:BP = 16:53,687,875~54205853. The FTO gene encodes the enzyme alpha-ketoglutarate-dependent dioxygenase, which plays a role in energy homeostasis and metabolism, and is linked to an increased risk of obesity.28 The second most significant gene is AUTS2 (PMAGMA = 2.68E-10), located at CHR:BP = 7:69,013,905~70308054. AUTS2, also known as Autism Susceptibility Candidate 2, encodes a protein involved in modulating synaptic plasticity, which is crucial for normal brain development and function.29 Alterations in AUTS2 are associated with a range of neurodevelopmental and psychiatric conditions.30 The third most significant gene is PRUNE2 (PMAGMA = 5.06E-09), located at CHR:BP = 9:79,176,292~79571003. PRUNE2 is similarly involved in neuronal differentiation and may influence neural plasticity, which is essential for learning and memory.31
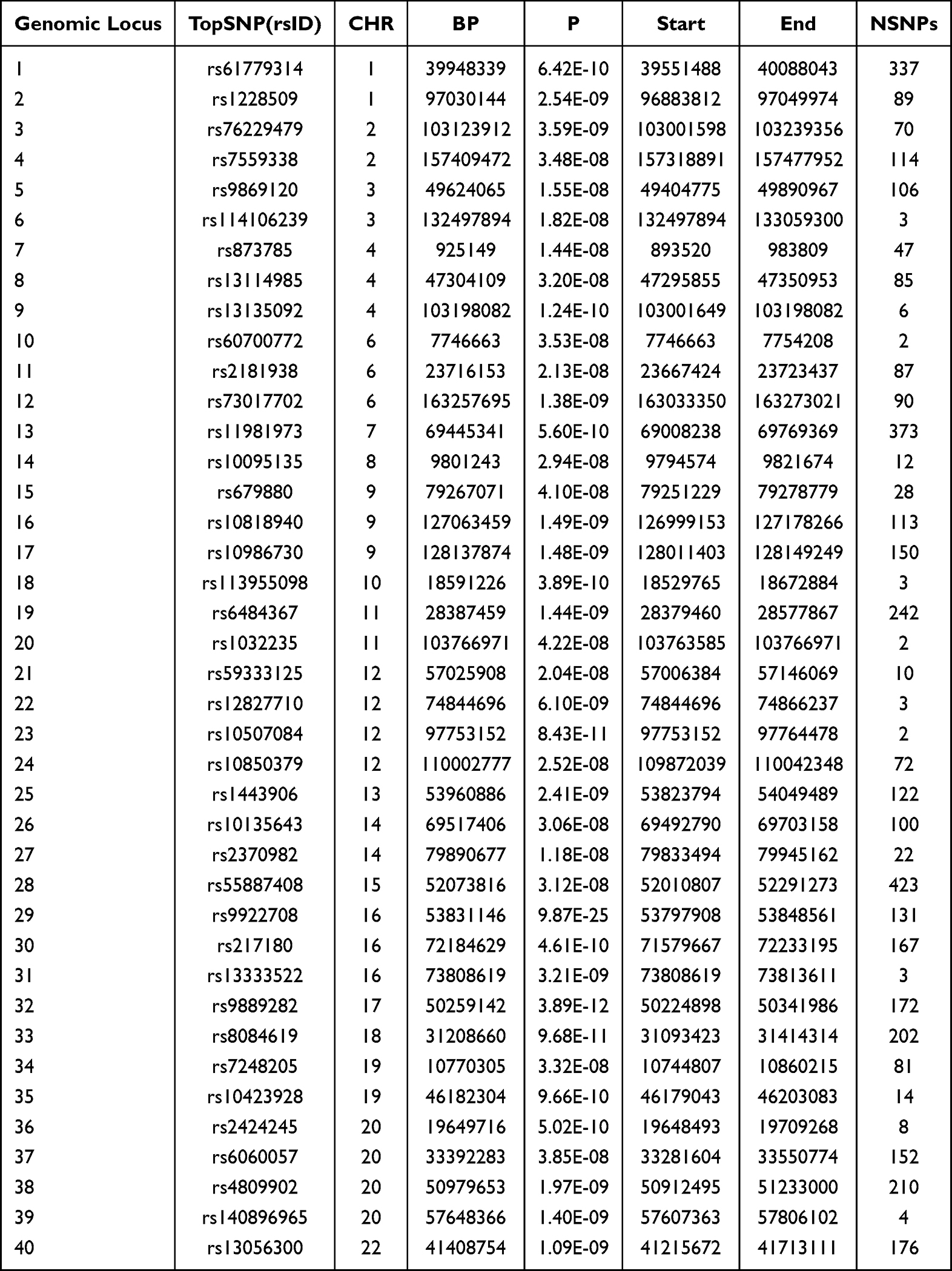 |
Table 1 Annotation Results of CPASSOC Outcomes Using the FUMA Platform |
 |
Figure 3 Manhattan Plot of the Shared Genes of OSA and Neck pain by MAGMA. |
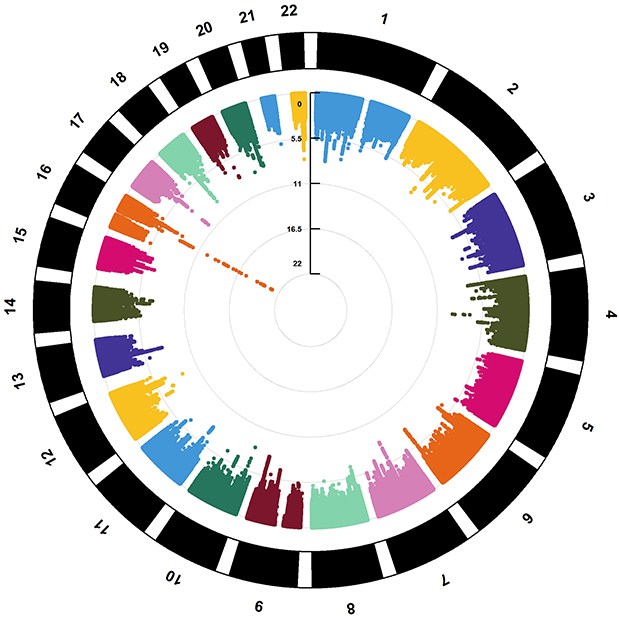 |
Figure 4 Circle_Manhattan Plot of the Shared Genes of OSA and Neck pain by MAGMA. |
Tissue Enrichment
We conducted tissue enrichment analysis using MAGMA to identify tissues with high expression levels of the associated genes. The analysis revealed significant enrichment in 13 tissues (Figure 5). Notably, the pituitary gland showed pronounced enrichment (P = 1.33E-03), while the remaining significantly enriched tissues were all in the brain. Among these, the Brain cortex exhibited the strongest enrichment (P = 2.17E-06), followed by the Brain frontal cortex BA9 and Brain anterior cingulate cortex BA24 (Supplementary Table 04). These results suggest that the associated genes may play crucial roles in these specific tissues, offering insights into the potential mechanisms underlying the studied trait.
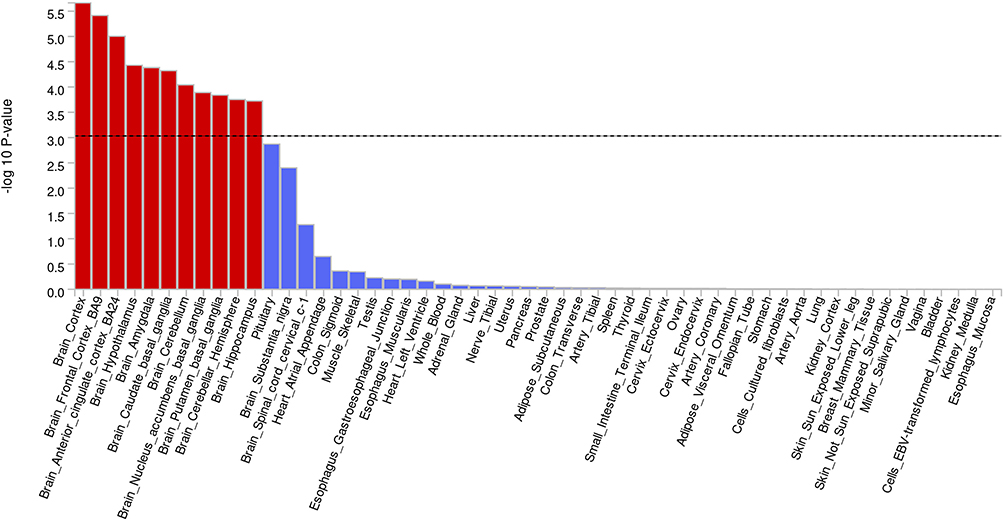 |
Figure 5 The Result of Tissue Enrichment based on GTEx 8 platform. |
TWAS
In the TWAS analysis, a total of seven genes were identified to exhibit significant pleiotropic effects on both OSA and neck pain. Among these, GPD2 demonstrated significant expression across the greatest number of tissues, including Esophagus_Muscularis, Whole_Blood, and Artery_Tibial, which may be attributed to its role in mitochondrial energy metabolism; its upregulation could influence oxidative stress levels or local hypoxia responses, thereby exacerbating hypoxic injury in OSA, or alternatively, modulate metabolic byproducts to affect neuroinflammatory pathways involved in neck pain. The second most highly expressed gene across tissues was MMAB, which showed significant associations with both OSA and neck pain in Colon_Transverse and Nerve_Tibial tissues. MMAB encodes an enzyme essential for mitochondrial cobalamin metabolism and energy homeostasis, suggesting a possible role of metabolic dysfunction in the shared pathogenesis of these two traits.32,33 Other genes located in coding regions, such as NCK1-AS1, were significantly associated with neck pain in Esophagus_Muscularis (P=4.32e-05, PFDR=2.42e-02) and OSA (P=9.22e-08, PFDR=4.29e-04). AP1G1 expression in Brain_Nucleus_accumbens_basal_ganglia showed significant associations with neck pain (P=1.06e-06, PFDR=4.77e-03) and OSA (P=5.99e-05, PFDR=0.027). AP1G1 is involved in clathrin-mediated vesicle trafficking, which may influence neuronal signaling pathways relevant to both pain perception and sleep regulation.34 Additionally, FOXN4 expression in Adipose_Visceral_Omentum was significantly associated with neck pain (P=3.51e-05, PFDR=2.59e-02) and OSA (P=7.9e-05, PFDR=0.030), indicating a potential contribution of adipose tissue regulation and systemic metabolic processes to the comorbidity of these conditions35 (Supplementary Table 05).
MR
In our analysis with neck pain as the exposure and OSA as the outcome, we selected 51 IVs with a significance threshold of P < 5E-08, ensuring all F-values exceeded 10 (Supplementary Table 06). Using the IVW method, we found that individuals with neck pain had an increased risk of developing OSA (b = 1.19, P = 4.38E-05) (Table 2, Figure 6A and B). The results were consistent when using the weighted median method (b = 0.96, P = 0.009). Heterogeneity testing showed a Cochran’s Q statistic p-value of 0.07, which is not significant at the 0.05 level, and pleiotropy testing yielded a p-value of 0.20, also above 0.05. However, when OSA was treated as the exposure and neck pain as the outcome, no significant causal relationship was detected (b = −0.002, P = 0.437) (Supplementary Table 07).
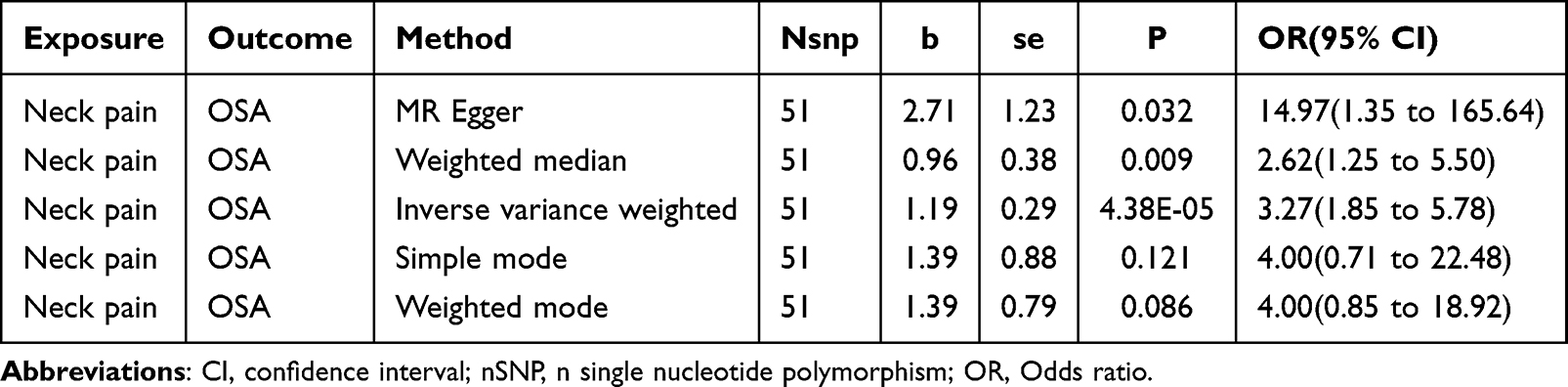 |
Table 2 MR Results of Causal Effects Between Neck Pain and OSA |
 |
Figure 6 (A) A Scatter Plots for the Causal Association between Neck pain and OSA. (B) A Leave-One-Out for the Causal Association between Neck Pain and OSA. |
Enrichment
The GO enrichment analysis of pleiotropic genes shared between OSA and neck pain highlighted several significantly enriched biological processes. Neural mechanisms accounted for the most prominent enrichment, with synaptic vesicle endocytosis (GO:0048488, DGKQ, DNM2, GAK; P = 2.34E-04), presynaptic endocytosis (GO:0140238, DGKQ, DNM2, GAK; P = 2.44E-04), and synaptic vesicle recycling (GO:0036465, DGKQ, DNM2, GAK; P = 3.58E-04) emerging as key processes, suggesting that alterations in neurotransmitter release and vesicle trafficking may underlie shared neural mechanisms. In addition, metabolic and endocrine regulation, as well as epigenetic modification, were also involved (Supplementary Table 08). Similarly, KEGG pathway analysis provided complementary insights, identifying significant enrichment in the Polycomb repressive complex (hsa03083; P = 8.95e-04), supporting an epigenetic regulatory component. Additional pathways included choline metabolism in cancer (hsa05231; P = 0.0219), TGF-βsignaling (hsa04350; P = 0.0267), and phospholipase D signaling (hsa04072; P = 0.0465), which together implicate metabolic reprogramming, extracellular matrix remodeling, and inflammatory signal transduction in the shared pathophysiology (Supplementary Table 09).
Discussion
This study not only utilized genetic correlation analysis and cross-trait analysis to investigate the genetic sharing structure between OSA and neck pain, but also employed Mendelian randomization to reveal a causal relationship between the two conditions.
This study employed LDSC analysis to identify a significant positive genetic correlation between OSA and neck pain, a finding consistent with clinical observations. The co-occurrence of OSA and neck pain may be attributed to shared genetic factors, involving mechanisms such as inflammation, neuromuscular regulation, or structural abnormalities. In OSA patients, elevated inflammatory markers such as C-reactive protein and interleukins (eg, IL-6) may lead to increased neck circumference and insulin resistance, thereby exacerbating neck pain. Degenerative changes, such as the loss of cervical lordosis or segmental spinal kyphosis, are also risk factors for OSA in neck pain patients. However, in clinical practice, symptoms of OSA, such as daytime sleepiness, snoring, or apnea, leading to delays in diagnosis and treatment of neck pain. This study identifies shared genetic loci between OSA and neck pain at multiple levels, providing new insights into the biological mechanisms underlying their co-occurrence. It offer valuable guidance for the comprehensive management of patients suffering from both conditions in clinical practice.
Through a cross-trait analysis, we identified several pleiotropic genes, among which the FTO gene showed the most significant shared genetic correlation with both OSA and neck pain. The FTO gene encodes a protein associated with fat mass and obesity, which can influence energy intake and expenditure, thereby regulating appetite and metabolic rate. Previous studies have found that the FTO gene variant rs8050136 is significantly associated with OSA risk.36 Strausz ect also pointed out that FTO gene variants are associated with an increased risk of COVID-19 infection in OSA patients.37 Moreover, FTO gene silencing, via a YTHDF2-dependent mechanism, enhances Grp177 m6A methylation, reducing oxidative stress and alleviating neuropathic pain.38 Functional studies using CRISPR/Cas9-mediated FTO knockout in porcine endothelial cells revealed significant transcriptomic and metabolic alterations, particularly in pathways related to insulin signaling, oxidative stress, and inflammation—which may link FTO to OSA and pain regulation.39 Similarly, FTO-deficient human embryonic stem cells maintained normal pluripotency and differentiation capacity, providing a stable and reliable model for investigating the mechanistic role of FTO in development and disease.40 This study confirms the role of the FTO gene in the co-occurrence of OSA and neck pain, consistent with previous findings. Obesity is recognized as a risk factor for pain.41 Furthermore, we identified AUTS2 and PRUNE2 as additional pleiotropic genes influencing the co-occurrence of OSA and neck pain. The AUTS2 gene plays a crucial role in neurodevelopment and is associated with conditions such as autism and intellectual disabilities.42 Zhang et al found that the PRUNE2 gene is linked to arteriovenous malformations.43 The proteins encoded by PRUNE2 and AUTS2 are involved in synaptic development and plasticity in neurons. While no direct association between these genes and OSA or pain has been demonstrated, future studies may explore whether neurodevelopmental and vascular mechanisms contribute to the co-occurrence of these two conditions.
At the gene expression level, we conducted TWAS analysis and found that the expression of the mitochondrial glycerophosphate dehydrogenase (GPD2) gene in Whole_Blood, Esophagus_Muscularis, and Artery_Tibial tissues was significantly associated with the occurrence of OSA and neck pain. The enzyme encoded by the GPD2 gene transfers electrons to the mitochondria via the glycerol-phosphate shuttle system, helping cells generate energy.44 Li et al conducted a cross-tissue transcriptome-wide study and identified GPD2 as a novel susceptibility gene for OSA, which regulates energy metabolism to reduce OSA risk.45 Furthermore, another study found that GPD1, a gene within the same family as GPD2, was upregulated in the cortex and six other regions after 24 or 96 hours of sleep deprivation.46 GPD2 is considered to be one of the key components linking carbohydrate and lipid metabolism. High triglyceride and low-density lipoprotein levels are associated with more severe local and systemic pain progression.47 Additionally, lipid molecules, such as prostaglandins and leukotrienes, enhance pain perception and promote inflammation by activating receptors in the nervous system.48,49 Therefore, we hypothesize that the expression of GPD2 may be negatively correlated with the shared risk of OSA and neck pain, likely due to its regulatory role in energy metabolism. MMAB (methylmalonic acid dehydrogenase subunit B) has been identified as the second pleiotropic gene associated with the co-occurrence of OSA and neck pain. This gene influences homocysteine metabolism, a pathway implicated in vascular and neurological regulation. Notably, Li et al observed that lower homocysteine levels correlated with greater reductions in diastolic blood pressure in non-obese OSA patients with normal body weight.50 Elevated homocysteine levels have been identified can attenuate vascular pain responses.50 Whether homocysteine directly contributes to the shared pathophysiology of OSA and neck pain remains speculative and warrants further investigation. Other gene expressions have not been found to be associated with the occurrence of OSA and neck pain in previous studies, warranting further exploration in the future.
Our bidirectional MR study indicates that neck pain has a direct detrimental effect on OSA, whereas no causal relationship was found between OSA and neck pain in the reverse analysis. The management of OSA traditionally focuses on physiological factors related to airway obstruction. For instance, Schwartz et al demonstrated that fat deposition around the pharynx due to obesity increases the collapsibility of the pharyngeal airway.51 Previous studies have demonstrated that the minimal cross-sectional area of the upper airway is significantly smaller in OSA patients compared to non-OSA individuals.52 Craniofacial abnormalities, such as retrognathia and micrognathia, are also common features among OSA patients.53 However, this evidence does not directly clarify the role of neck pain. Our results indicate that neck pain may act as a risk factor for OSA rather than a mere coexisting symptom, suggesting a potential causal contribution to its development. Pathophysiological mechanisms such as sympathetic overactivation and low-grade systemic inflammation associated with chronic pain may underlie this relationship.54 In addition, metabolic dysregulation, including altered energy homeostasis and lipid metabolism, may further contribute to the development of OSA in patients with chronic pain.55 Consistent with this, the enrichment results also revealed that these pleiotropic genes are associated with metabolic mechanisms. Recognizing and addressing neck pain may enhance treatment outcomes for patients with OSA and should be part of routine clinical care.
While MR reduces the influence of confounding by using the random allocation of genetic variants, residual confounders and methodological limitations may still affect the interpretation of the unidirectional causality observed. In this study, we systematically examined the causal relationship and genetic overlap between OSA and neck pain at both the SNP and gene levels, but our analysis was limited to European populations. Future research should extend to trans-ethnic GWAS or meta-analytic frameworks to increase diversity and strengthen the generalizability of the findings.;
Conclusion
This study not only identified a causal relationship between neck pain and OSA, but also revealed the shared genetic architecture at the SNP level between the two traits and the underlying physiological tissues involved. These findings provide novel insights into the common pathogenic mechanisms of pain and sleep disorders, offering a foundation for future efforts aimed at precision risk stratification and targeted interventions.
Data Sharing Statement
No datasets were generated or analysed during the current study.
Ethics Approval
In accordance with item 1 and 2 of Article 32 of the Measures for Ethical Review of Life Science and Medical Research Involving Human Subjects dated February 18, 2023, China, the current study, which utilizes publicly available and legally obtained anonymized data, is exempt from ethical review.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The National Key Research and Development Program of China (Grant Number: 2022YFC2009700); Fujian Provincial Young and Middle-aged Teacher Education Research Project (Science and Technology)(No.JAT241293). The funding organizations played no further role in study design, data collection, analysis and interpretation, and paper writing.
Disclosure
The authors declare no competing interests in this work.
References
1. Southall WJI. Neck pain. InnovAiT. 2021;14(506):511.
2. Hoy DG, Protani M, De R, Buchbinder R. The epidemiology of neck pain. Best Pract Res. 2010;24(6):783–792. doi:10.1016/j.berh.2011.01.019
3. Holm LW, Bohman T, Lekander M, Magnusson C, Skillgate E. Risk of transition from occasional neck/back pain to long-duration activity limiting neck/back pain: a cohort study on the influence of poor work ability and sleep disturbances in the working population in Stockholm County. BMJ open. 2020;10(6):e033946. doi:10.1136/bmjopen-2019-033946
4. Auvinen JP, Tammelin TH, Taimela SP, et al. Is insufficient quantity and quality of sleep a risk factor for neck, shoulder and low back pain? A longitudinal study among adolescents. Eur Spine J. 2010;19(4):641–649. doi:10.1007/s00586-009-1215-2
5. Moldofsky H. Sleep and pain. Sleep Med Rev. 2001;5(5):385–396. doi:10.1053/smrv.2001.0179
6. Lautenbacher S, Kundermann B, Krieg JC. Sleep deprivation and pain perception. Sleep Med Rev. 2006;10(5):357–369. doi:10.1016/j.smrv.2005.08.001
7. Irwin MR, Olmstead R, Carrillo C, et al. Sleep loss exacerbates fatigue, depression, and pain in rheumatoid arthritis. Sleep. 2012;35(4):537–543. doi:10.5665/sleep.1742
8. Pham T, Lin CK, Leek D, Chandrashekhar R, Annaswamy TM. Obstructive sleep Apnea’s association with the cervical spine abnormalities, posture, and pain: a systematic review. Sleep Med. 2020;75:468–476. doi:10.1016/j.sleep.2020.09.008
9. Araújo Lda S, Fernandes JF, Klein MR, Sanjuliani AF. Obstructive sleep apnea is independently associated with inflammation and insulin resistance, but not with blood pressure, plasma catecholamines, and endothelial function in obese subjects. Nutrition. 2015;31(11–12):1351–1357. doi:10.1016/j.nut.2015.05.017
10. Kohler M, Stradling JR. CrossTalk proposal: most of the cardiovascular consequences of OSA are due to increased sympathetic activity. J Physiol. 2012;590(12):2813–5;discussion2823. doi:10.1113/jphysiol.2012.229633
11. Xie X, Pan L, Ren D, Du C, Guo Y. Effects of continuous positive airway pressure therapy on systemic inflammation in obstructive sleep apnea: a meta-analysis. Sleep Med. 2013;14(11):1139–1150. doi:10.1016/j.sleep.2013.07.006
12. Stannus OP, Jones G, Blizzard L, Cicuttini FM, Ding C. Associations between serum levels of inflammatory markers and change in knee pain over 5 years in older adults: a prospective cohort study. Ann Rheum Dis. 2013;72(4):535–540. doi:10.1136/annrheumdis-2011-201047
13. Joo EY, Tae WS, Lee MJ, et al. Reduced brain gray matter concentration in patients with obstructive sleep apnea syndrome. Sleep. 2010;33(2):235–241. doi:10.1093/sleep/33.2.235
14. Taylor KS, Millar PJ, Murai H, et al. Cortical autonomic network gray matter and sympathetic nerve activity in obstructive sleep apnea. Sleep. 2018;41(2). doi:10.1093/sleep/zsx208
15. Jensen KB, Regenbogen C, Ohse MC, Frasnelli J, Freiherr J, Lundström JN. Brain activations during pain: a neuroimaging meta-analysis of patients with pain and healthy controls. Pain. 2016;157(6):1279–1286. doi:10.1097/j.pain.0000000000000517
16. Meng W, Chan BW, Harris C, et al. A genome-wide association study finds genetic variants associated with neck or shoulder pain in UK Biobank. Human Mol Genetics. 2020;29(8):1396–1404. doi:10.1093/hmg/ddaa058
17. Sofer T, Kurniansyah N, Murray M, et al. Genome-wide association study of obstructive sleep apnoea in the Million Veteran Program uncovers genetic heterogeneity by sex. EBioMedicine. 2023;90:104536. doi:10.1016/j.ebiom.2023.104536
18. Liu Q, Ou Y, Liu T, et al. Preliminary evidence of immune infiltration and neutrophil degranulation in peripheral blood of non-obese OSA patients related to cognitive decline. Sci Rep. 2025;15(1):3481. doi:10.1038/s41598-025-88034-z
19. Cade BE, Gharib SA. Breath of fresh air: toward unraveling the molecular underpinnings of sleep apnea. Am J Respir Crit Care Med. 2022;206(12):1450–1451. doi:10.1164/rccm.202207-1410ED
20. Gui H, Kwan JS, Sham PC, Cherny SS, Li M. Sharing of genes and pathways across complex phenotypes: a multilevel genome-wide analysis. Genetics. 2017;206(3):1601–1609. doi:10.1534/genetics.116.198150
21. Zhu Z, Hasegawa K, Camargo CA Jr, Liang L. Investigating asthma heterogeneity through shared and distinct genetics: insights from genome-wide cross-trait analysis. J Allergy Clin Immunol. 2021;147(3):796–807. doi:10.1016/j.jaci.2020.07.004
22. Song S, Jiang W, Zhang Y, Hou L, Zhao H. Leveraging LD eigenvalue regression to improve the estimation of SNP heritability and confounding inflation. Am J Hum Genet. 2022;109(5):802–811. doi:10.1016/j.ajhg.2022.03.013
23. Werme J, van der Sluis S, Posthuma D, de Leeuw CA. An integrated framework for local genetic correlation analysis. Nat Genet. 2022;54(3):274–282. doi:10.1038/s41588-022-01017-y
24. Zhu X, Feng T, Tayo BO, et al. Meta-analysis of correlated traits via summary statistics from GWASs with an application in hypertension. Am J Hum Genet. 2015;96(1):21–36. doi:10.1016/j.ajhg.2014.11.011
25. de Leeuw CA, Mooij JM, Heskes T, Posthuma D. MAGMA: generalized gene-set analysis of GWAS data. PLoS Comput Biol. 2015;11(4):e1004219. doi:10.1371/journal.pcbi.1004219
26. Gusev A, Ko A, Shi H, et al. Integrative approaches for large-scale transcriptome-wide association studies. Nat Genet. 2016;48(3):245–252. doi:10.1038/ng.3506
27. Sekula P, Del Greco MF, Pattaro C, Köttgen A. Mendelian randomization as an approach to assess causality using observational data. J Am Soc Nephrol. 2016;27(11):3253–3265. doi:10.1681/asn.2016010098
28. Zhao X, Yang Y, Sun BF, Zhao YL, Yang YG. FTO and obesity: mechanisms of association. Curr Diabetes Rep. 2014;14(5):486. doi:10.1007/s11892-014-0486-0
29. Jha U, Kondrychyn I, Korzh V, Thirumalai V. High behavioral variability mediated by altered neuronal excitability in auts2 mutant zebrafish. eNeuro. 2021;8(5):ENEURO.0493–20.2021. doi:10.1523/eneuro.0493-20.2021
30. Oksenberg N, Haliburton GD, Eckalbar WL, et al. Genome-wide distribution of Auts2 binding localizes with active neurodevelopmental genes. Transl Psychiatry. 2014;4(9):e431. doi:10.1038/tp.2014.78
31. Gradari S, Herrera A, Tezanos P, Fontán-Lozano Á, Pons S, Trejo JL. The role of Smad2 in adult neuroplasticity as seen through hippocampal-dependent spatial learning/memory and neurogenesis. J Neurosci. 2021;41(32):6836–6849. doi:10.1523/jneurosci.2619-20.2021
32. Brasil S, Richard E, Jorge-Finnigan A, et al. Methylmalonic aciduria cblB type: characterization of two novel mutations and mitochondrial dysfunction studies. Clin Genet. 2015;87(6):576–581. doi:10.1111/cge.12426
33. Keller SA, Luciani A. Mitochondrial distress in methylmalonic acidemia: novel pathogenic insights and therapeutic perspectives. Cells. 2022;11(19):3179. doi:10.3390/cells11193179
34. Usmani MA, Ahmed ZM, Magini P, et al. De novo and bi-allelic variants in AP1G1 cause neurodevelopmental disorder with developmental delay, intellectual disability, and epilepsy. Am J Hum Genet. 2021;108(7):1330–1341. doi:10.1016/j.ajhg.2021.05.007
35. Gharib SA, Hayes AL, Rosen MJ, Patel SR. A pathway-based analysis on the effects of obstructive sleep apnea in modulating visceral fat transcriptome. Sleep. 2013;36(1):23–30. doi:10.5665/sleep.2294
36. Cronin RM, Field JR, Bradford Y, et al. Phenome-wide association studies demonstrating pleiotropy of genetic variants within FTO with and without adjustment for body mass index. Front Genetics. 2014;5:250. doi:10.3389/fgene.2014.00250
37. Johnson R, Shi H, Pasaniuc B, Sankararaman S. A unifying framework for joint trait analysis under a non-infinitesimal model. Bioinformatics. 2018;34(13):i195–i201. doi:10.1093/bioinformatics/bty254
38. Wu SF, Wang Y, Zhao QC. Demethylase FTO promotes neuropathic pain development via regulating the m6A methylation levels of CXCR3. Acta Biochim Pol. 2022;69(4):819–824. doi:10.18388/abp.2020_6185
39. Xie L, Fan N, Ding X, et al. Comparative transcriptomic and metabolomic analysis of FTO knockout and wild-type porcine iliac artery endothelial cells. Gene. 2025;936:149094. doi:10.1016/j.gene.2024.149094
40. Wei C, Luo Q, Wang B, et al. Generation of a FTO gene knockout human embryonic stem cell line using CRISPR/Cas9 editing. Stem Cell Res. 2021;53:102362. doi:10.1016/j.scr.2021.102362
41. Vincent HK, Heywood K, Connelly J, Hurley RW. Obesity and weight loss in the treatment and prevention of osteoarthritis. PMR. 2012;4(5 Suppl):S59–67. doi:10.1016/j.pmrj.2012.01.005
42. Biel A, Castanza AS, Rutherford R, et al. AUTS2 syndrome: molecular mechanisms and model systems. Front Mol Neurosci. 2022;15:858582. doi:10.3389/fnmol.2022.858582
43. Zhang M, Ding X, Zhang Q, et al. Exome sequencing of 112 trios identifies recessive genetic variants in brain arteriovenous malformations. J Neurointervent Surg. 2021;13(6):568–573. doi:10.1136/neurintsurg-2020-016469
44. Daoud H, Gruchy N, Constans JM, et al. Haploinsufficiency of the GPD2 gene in a patient with nonsyndromic mental retardation. Hum Genet. 2009;124(6):649–658. doi:10.1007/s00439-008-0588-3
45. Meng L, Gui J, Han Z, et al. Novel susceptibility genes for sleep apnea revealed by a cross-tissue transcriptome-wide association study. Int J Biol Macromol. 2025;297:139841. doi:10.1016/j.ijbiomac.2025.139841
46. Conti B, Maier R, Barr AM, et al. Region-specific transcriptional changes following the three antidepressant treatments electro convulsive therapy, sleep deprivation and fluoxetine. Mol Psychiatry. 2007;12(2):167–189. doi:10.1038/sj.mp.4001897
47. Pan F, Tian J, Cicuttini F, Jones G. THU0420 Metabolic syndrome and trajectories of localised pain and generalised pain. Ann Rheumatic Dis. 2019;78(Suppl 2):498. doi:10.1136/annrheumdis-2019-eular.1245
48. Noguchi K, Okubo M. Leukotrienes in nociceptive pathway and neuropathic/inflammatory pain. Biol Pharm Bull. 2011;34(8):1163–1169. doi:10.1248/bpb.34.1163
49. Wagner KM, Gomes A, McReynolds CB, Hammock BD. Soluble epoxide hydrolase regulation of lipid mediators limits pain. Neurotherapeutics. 2020;17(3):900–916. doi:10.1007/s13311-020-00916-4
50. Joseph EK, Green PG, Ferrari LF, Levine JD. Homocysteine-induced attenuation of vascular endothelium-dependent hyperalgesia in the rat. Neuroscience. 2015;284:678–684. doi:10.1016/j.neuroscience.2014.10.056
51. Schwartz AR, Patil SP, Squier S, Schneider H, Kirkness JP, Smith PL. Obesity and upper airway control during sleep. J Appl Physiol. 2010;108(2):430–435. doi:10.1152/japplphysiol.00919.2009
52. Chen H, Aarab G, de Ruiter MH, de Lange J, Lobbezoo F, van der Stelt PF. Three-dimensional imaging of the upper airway anatomy in obstructive sleep apnea: a systematic review. Sleep Med. 2016;21:19–27. doi:10.1016/j.sleep.2016.01.022
53. Tsuiki S, Isono S, Ishikawa T, Yamashiro Y, Tatsumi K, Nishino T. Anatomical balance of the upper airway and obstructive sleep apnea. Anesthesiology. 2008;108(6):1009–1015. doi:10.1097/ALN.0b013e318173f103
54. Reynolds CA, Minic Z. Chronic pain-associated cardiovascular disease: the role of sympathetic nerve activity. Int J Mol Sci. 2023;24(6):5378. doi:10.3390/ijms24065378
55. Ito A, Suganami T. Lipid metabolism in myeloid cell function and chronic inflammatory diseases. Front Immunol. 2024;15:1495853. doi:10.3389/fimmu.2024.1495853
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.