Back to Journals » Neuropsychiatric Disease and Treatment » Volume 14
Serum levels of GPER -1 in euthymic bipolar patients
Authors Orhan FÖ ![]() , Kurutaş EB
, Kurutaş EB ![]() , Doğaner A, Türker E, Taner Özcü SŞ, Güngör M, Çakmak S
, Doğaner A, Türker E, Taner Özcü SŞ, Güngör M, Çakmak S
Received 5 December 2017
Accepted for publication 19 February 2018
Published 26 March 2018 Volume 2018:14 Pages 855—862
DOI https://doi.org/10.2147/NDT.S158822
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Roger Pinder
Fatma Özlem Orhan,1 Ergül Belge Kurutaş,2 Adem Doğaner,3 Ebru Türker,1 Safiye Şeyma Taner Özcü,4 Meltem Güngör,5 Seyfettin Çakmak1
1Department of Psychiatry, Faculty of Medicine, Kahramanmaraş Sütçüimam University, Kahramanmaras, Turkey; 2Department of Biochemistry, Faculty of Medicine, Kahramanmaraş Sütçüimam University, Kahramanmaras, Turkey; 3Department of Biostatistics and Medical Informatics, Faculty of Medicine, Kahramanmaraş Sütçüimam University, Kahramanmaras, Turkey; 4Department of Bioengineering, Institute of Science, Kahramanmaraş Sütçüimam University, Kahramanmaras, Turkey; 5Vocational High School, Toros University, Mersin, Turkey
Introduction: Estrogen and its receptors have been suggested as playing a role in the pathogenesis of bipolar disorder (BD). Estrogen functions through the estrogen receptors alpha and beta and the recently discovered G-protein–coupled estrogen receptor-1 (GPER-1). The aim of this study was to evaluate serum GPER-1 levels in euthymic BD patients.
Patients and methods: The study population consisted of 38 euthymic outpatients meeting the criteria for BD in the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition and 35 age- and gender-matched healthy controls. Medical histories were obtained and physical examinations and laboratory tests conducted.
Results: Serum GPER-1 levels were measured in both patients and controls and found to be significantly higher in the BD patients than in controls. These results were not influenced by the medications in use.
Conclusion: The results of this study demonstrated that GPER-1 may play a role in BD pathophysiology.
Keywords: estrogen receptor, sex hormones, GPER-1, bipolar, euthymic
Introduction
Bipolar disorder (BD) is a severe, recurrent mood disorder that affects millions of people worldwide, yet the etiology of the illness remains unclear.1,2 It has been found that some women with BD are more vulnerable to developing mood instability during periods of hormone fluctuation. It appears that reproductive events and hormonal treatments may affect the course of BD in women.3 There is a robust relationship between gonadal hormones, including estrogens, and mood disorders in women.4 There are some gender differences in patients with BD, even though the results are conflicting. The gender differences support further investigation of the possible interaction between BD and reproduction-related hormonal changes, particularly changes in the main female sex hormone estrogen.3 Although the role of estrogen in cycle-dependent mood disorders such as premenstrual syndrome or postpartum and menopausal depressive disorders is not entirely clear,5,6 a broad range of interactions with neuronal signaling has been demonstrated.7 However, the role of estrogen in BD patients is not fully understood.
Estrogens encompass steroid hormones that not only display physiologic roles in the female reproductive system, but also affect several other systems in the body, including the cardiovascular, musculoskeletal, immune, and central nervous system (CNS).8,9 Estrogens are synthesized in the brain10,11 and are known to exhibit widespread neurologic effects ranging from cognition to emotional status to sensory processing.12 Mood changes occurring due to the effects of estrogen on the CNS have been extensively investigated in studies on rats.13–18 A correlation has been observed between decreased estrogen levels (eg, premenstrually, during the postpartum period, and perimenopausally) and increased anxiety and depressive symptoms.19–21 Estrogen appears to function as an agonist on the serotonergic system, by decreasing the monoamine oxidase activity (the enzyme catabolizing serotonin and dopamine)5 and affecting the interneuronal serotonin transport – both processes increasing the level of serotonin in the synapse, leading to possible mood enhancement.22 Additionally, estrogen is involved in regulating serotonin receptor number and function,23–25 thus controlling the activity of serotonergic neurons. Also, estrogen has been shown to affect many neurotransmitter systems in addition to the serotonergic system. For example, estrogen generally increases the activity of noradrenaline, acts as a cholinergic agonist in particular areas of the brain, decreases dopamine D2 receptor sensitivity, and appears to act as an adjunct agonist of gamma-aminobutyric acid.26 While estrogens can modulate serotonin and noradrenaline neurotransmission, they appear to play an important role in neuroprotection27 and anti-inflammation in the brain.28
Estrogen mediates its effects mostly by binding to estrogen receptors (ERs). There are two different types of ERs: one is a member of the nuclear hormone family of intracellular receptors and the other one is a membrane-bound G-protein–coupled estrogen receptor (GPER).29 The membrane-bound G-protein–coupled estrogen receptor-1 (GPER-1), or GPR-30, was identified by several groups in the late 1990s.30 GPER has been found to be responsible for the rapid actions of estrogen.31 Revankar et al reported that estradiol binds to and signals through GPER-1 with high affinity and potency in vitro, leading to the classification of GPER-1 as a membrane-bound ER.32 GPER-1 was shown to act independently of ERα and Erβ, but likely stimulates the same second-messenger pathways and has genomic actions.33 Thus, the effects of estrogens in the brain likely depend on the relative expression and cellular locations of multiple ERs. Immunohistochemical studies have revealed that GPER is expressed in both the CNS and peripheral nervous system; throughout the nervous system, the GPER regulates its own expression in regionally distinct ways, and it has increasingly been shown to mediate estrogen’s physiologic and pathologic functions, with expression found in the brain of both adult male and nonpregnant female rats.34 ERs are expressed in a variety of areas in the brain, where they predominate in limbic-related areas known to be important for emotion, cognition, and behavior.35,36 In animal model experiments, GPER has been found in male and female rodents throughout the CNS and peripheral nervous system, including the hypothalamus, hippocampus, midbrain, spinal cord, and dorsal root ganglia.34,37–39 GPER-1 is expressed in neurons and microglia.28,40 In addition, ERs have been identified in the leukocytes isolated from peripheral blood and the blood vessel endothelia of both men and women.41,42 GPER-1 in macrophages is associated with the anti-inflammatory action of estrogens.43,44 Till now, the physiologic functions for GPER have been described in almost every organ system.45
Several studies have investigated the possible involvement of ERs in the etiology of psychiatric disorders.29 To the best of our knowledge, there has been no study to date examining serum GPER-1 levels in BD patients. The aim of this study was to evaluate the potential role of GPER-1 in euthymic BD patients and compare their GPER-1 levels with those of healthy controls.
Patients and methods
Patients
The study included 38 BD patients and 35 age- and gender-matched healthy controls. The patients included in the study had already received the diagnosis of BD according to the Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition.46 The study protocol was approved by the Scientific Research Ethics Committee of Kahramanmaraş Sütçüimam University, and detailed written informed consent was obtained from each patient and control subject.
The study excluded those who were pregnant or postmenopausal, having irregular menstrual cycles (anovulatory cycle), receiving hormone replacement therapy (eg, oral contraceptives), having endocrine disorders (diabetes mellitus or impaired levels of thyroid-stimulating hormone), addicted to alcohol and/or other substances, and having a psychiatric disorder other than BD or a concomitant physical disease.
The demographic and clinical characteristics, including gender, age, and comorbid conditions, were recorded for all participants, and physical examinations and all required laboratory tests were performed.
Procedure
Blood samples were collected from all patients in the BD and the control groups between 9:00 am and 11.00 am Samples were centrifuged and the serum samples stored at −20°C until analysis. The GPER-1 levels were analyzed in the Biochemistry Laboratory at Kahramanmaraş Sütçüimam University School of Medicine using a quantitative sandwich enzyme-linked immunosorbent assay technique and a commercial kit (SEG045Hu; Cloud-Clone Corp.; Houston, TX, USA) according to the manufacturer’s instructions.
Statistical analysis
The normal distribution fitness of the data was examined using the Shapiro–Wilk test. An independent samples t-test was used to compare the groups for variables that had normal distributions. Descriptive statistics were specified with mean ± SD. Mann–Whitney U test was used to compare the groups for variables that did not have normal distribution. Descriptive statistics were expressed in median (Quartile 1–Quartile 3). Chi-square test and Fisher’s exact test were applied to categorical variables. The relationship between variables was examined using Pearson’s correlation test. Statistical significance was accepted as p<0.05. Analysis of the data was conducted using IBM SPSS version 22 (IBM SPSS for Windows version 22; IBM Corporation; Armonk, NY, USA). A receiver operating characteristic (ROC) curve was plotted to obtain a cut-off value of GPER-1 for predicting the presence of BD.
Results
This study included 38 patients with euthymic BD and 35 age- and gender-matched healthy control subjects. The groups did not differ significantly in age (patients: 37.61±12.16, controls: 33.51±9.59 [mean ± SD], p=0.117) or gender (patients male/female: 15/23, controls male/female: 17/18, p=0.434). Table 1 shows the sociodemographic characteristics of the sample.
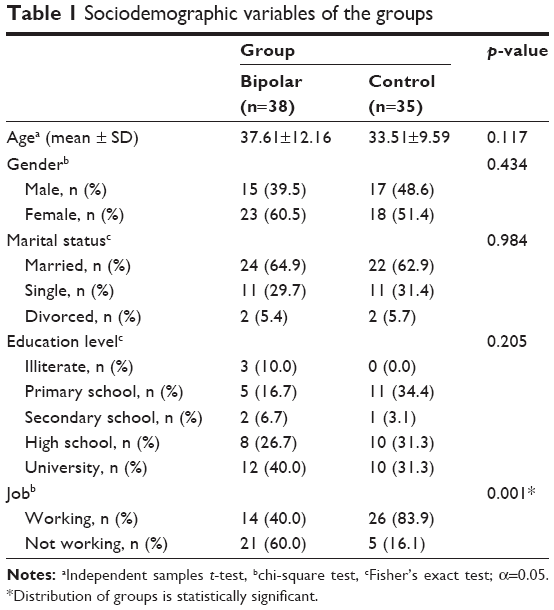 | Table 1 Sociodemographic variables of the groups |
Table 2 shows the BD patients’ duration of illness and the medication data. The levels of estradiol were found to be similar in both groups (patients: 41.27 pg/mL, controls: 38.17 pg/mL, [mean ± SD], p=0.774). However, the serum GPER-1 level was significantly higher in patients with euthymic BD compared to healthy controls (0.55±0.14 and 0.23±0.09 ng/mL [mean ± SD], respectively, p<0.001), as shown in Table 3 and Figure 1.
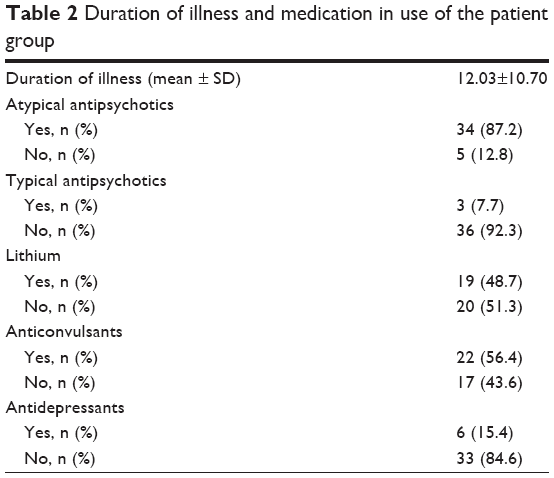 | Table 2 Duration of illness and medication in use of the patient group |
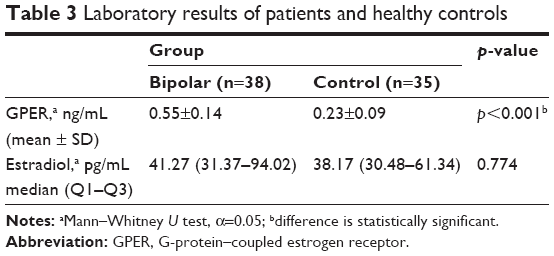 | Table 3 Laboratory results of patients and healthy controls |
 | Figure 1 GPER-1 levels in the euthymic BD and control groups. |
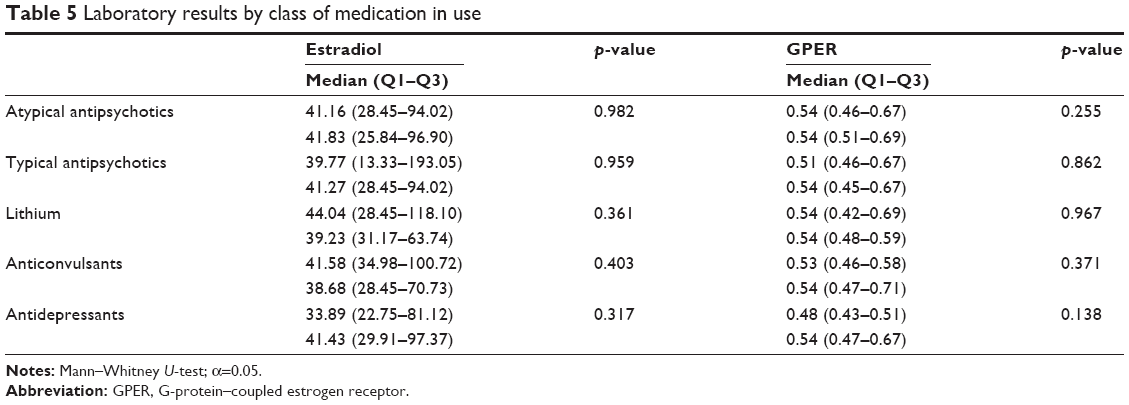
Subgroup analysis found significantly higher GPER-1 levels in all patients, regardless of gender (see Table 4). This result indicates the possible diagnostic potential of GPER-1 in BD, independent of gender. No significant difference was found between the BD and control groups in the levels of estradiol in males and females (Table 4). Pearson’s correlation analysis showed no significant correlation of the levels of estradiol and GPER with the duration of illness (p>0.05). Additionally, the laboratory results were not influenced by the medications in use (Table 5). The ROC analysis revealed that GPER-1 had a predictive value for the presence of BD (area under the curve, 0.977; 95% CI, 0.948–1.000; p<0.001). When the GPER-1 value was ≥0.3850, the sensitivity and specificity for the presence of BD were 92.1 and 97.1%, respectively (Figure 2). The GPER-1 level was under the cut-off point in all subjects in the control group.
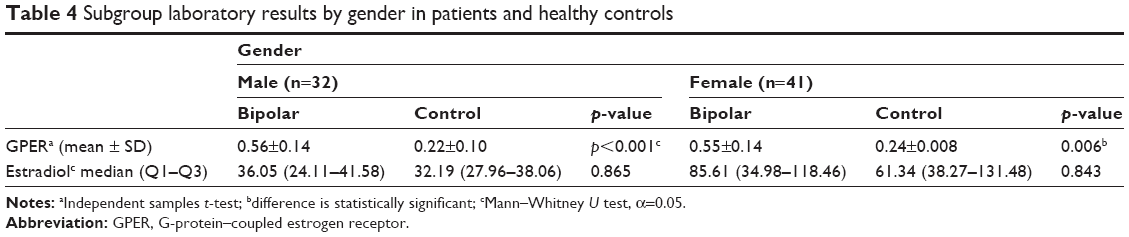 | Table 4 Subgroup laboratory results by gender in patients and healthy controls |
 | Table 5 Laboratory results by class of medication in use |
 | Figure 2 ROC curve for GPER-1. |
Discussion
The main finding of this study was that serum GPER-1 levels were higher in euthymic BD patients than in healthy control subjects. In addition, the results showed that these increased GPER-1 levels were independent of gender. Additionally, in ROC analysis, it was found that the diagnostic value of GPER-1 levels for BD was significantly high. This finding indicates that GPER-1 could play a potential role in the pathogenesis of BD. To the best of our knowledge, this is the first study to evaluate serum GPER-1 levels in BD patients.
A significant proportion of women with BD face emotional challenges during periods of normal hormonal fluctuation.47 In addition, numerous studies have reported that periods of hormonal fluctuation (eg, premenstrually, postpartum, and postmenopausally) are associated with increased risk of affective dysregulation and vulnerability to developing depression.5,48–50 Several studies have shown that periods of depression often correlate with hormonal fluctuations in women with BD or major depression.5,26,48–52 It has been suggested that estrogen participates in coordinating synaptic plasticity in the nervous system,53 and it is thought to play a significant role in neuropsychiatric disorders. Recent data suggest estrogen can serve as a short-term adjuvant to selective serotonin reuptake inhibitors.54 In most humans, high and constant levels of estrogen are described as anxiolytic and “emotionally positive”. In contrast, low and/or fluctuating levels correlate with dysphoric emotional states and increased anxiety.16
There has been extensive research into the role of estradiol and the activation of ERs in the neuroprotective and neurodegenerative processes, using various animal models of neurodegeneration, cognitive impairment, and affective disorders.19,55 Two studies of possible involvement of the ERα and ERβ genes in BD found no association with the disease.56,57 However, Weickert et al58 reported that differences in the ERα gene and its mRNA expression contributed to the risk of developing schizophrenia. Another study found that persons committing suicide had decreased levels of ERβ mRNA.36 The detection of both ERα and ERβ mRNAs in the human hippocampus, amygdala, and cerebral cortex59–63 raises the possibility that ERs may directly mediate the known effects of estrogen on mood.64,65 It has been reported that favorable effect of estrogen on mood is associated with ERβ signaling, and that estrogen leads to a decrease in glucocorticoid activity via ERβ and has antidepressant activity.66
Although the cellular activities of GPER have been examined in a number of systems, the physiologic functions of GPER are just beginning to be investigated. Based on the extensive effects of estrogen in a vast array of physiologic systems, the contributions of GPER to estrogen-mediated effects could be substantial.12 Recently, GPER stimulation was shown to attenuate serotonin receptor signaling in the paraventricular nucleus, as demonstrated by reduced oxytocin and adrenocorticotropic hormone responses, suggesting that GPER may play a role in mood disorders.67 GPER-1 is expressed in the hypothalamus,67 pituitary gland,68 hippocampal formation, and amygdala69 in both male and female rodents, which indicates a role for GPER-1 in controlling emotions and regulating endocrine responses. In addition, a role of GPER-1 in mood disorders has been reported.12,69 Experimental studies on animals have suggested that antidepressant-like effect of estradiol is mediated by ERβ and GPER-1, and that GPER-1 might be a new target in the treatment of depression.70,71 Furthermore, the GPER-1 level was increased in the serum of patients with generalized anxiety disorder, significantly and positively correlating with the severity of the anxiety, and it has been suggested that increased serum GPER-1 levels may play a role in the etiology of generalized anxiety disorder.72 GPER-1 level has been examined in tissue in rat models,37,73 and recently, serum has been studied to correlate GPER-1 level with various diseases.72
According to our hypothesis, it is expected that GPER-1 is a more selective biomarker for BD in females than in males. But both male and female BD group had higher GPER-1 levels than that of each control group. In rat studies, there is a controversial effect of GPER-1 on mood according to the gender. Some studies show equal effect in both genders,13–15 and others show gender differences.16
There is evidence that neuroinflammation underlies BD.74 Dysfunction still occurs in the brain of euthymic patients, while no BD symptoms appears in euthymic state,75 implying that neuroinflammation persists in BD patients with euthymic state. Serum GPER-1 level is higher in euthymic patients compared with controls, while there is no difference in serum estradiol level between the two groups. Accordingly, it is possible that GPER-1 signal is increased to attenuate neuroinflammation in BD patients with euthymic state. Improved GPER-1 signal may also occur in the brain.
GPR30 mediates estrogen-induced G-protein activation of several signaling cascades, such as protein kinase cascades-phosphatidylinositol 3 kinase and calcium signaling,32 at its locations in the plasma membrane, Golgi apparatus, and endoplasmic reticulum.76 Additionally, estrogen treatment increases the expression of protein kinase C (PKC), an important intracellular messenger.77 Biochemical data support the potential involvement of PKC and its substrates in BD patients,2 and mood stabilizers such as lithium and valproate have been shown to inhibit PKC.78 In this context, two small, randomized controlled trials have revealed robust anti-manic effects with tamoxifen, which is an ER and PKC antagonist and a selective estrogen receptor modulator.77,79 However, in this study, the laboratory results were not influenced by the medications in use (Table 5).
The limitations of this study include its small sample size, its cross-sectional design, and the ongoing medication of the BD patients. Because it is unethical to stop patients’ treatments, medications were not suspended during the study. In addition, the study measured serum levels, which offer insight into global alterations of GPER-1 levels in BD, but may not reflect the levels in the brain. In contrast, the study’s strength is that it is the first to investigate blood GPER-1 levels in BP patients, as far as we know.
Conclusion
This study demonstrated that serum GPER-1 levels were significantly higher in BP patients than in control subjects. Moreover, it was observed that GPER-1 level had a good diagnostic value for the presence of BD. Therefore, GPER-1 receptor activity may be a candidate biomarker for BP. These results should be considered preliminary and need to be confirmed by future studies.
Acknowledgments
The authors are grateful to Mehmet Karadöl for data collection.
Disclosure
The authors report no conflicts of interest in this work.
References
Kulkarni J, Garland KA, Scaffi di A, et al. A pilot study of hormone modulation as a new treatment for mania in women with bipolar affective disorder. Psychoneuroendocrinology. 2006;31:543–547. | ||
Zarate CA Jr, Singh JB, Carlson PJ, et al. Efficacy of a protein kinase C inhibitor (tamoxifen) in the treatment of acute mania: a pilot study. Bipolar Disord. 2007;9:561–570. | ||
Meinhard N, Kessing LV, Vinberg M. The role of estrogen in bipolar disorder, a review. Nord J Psychiatry. 2014;68:81–87. | ||
Borrow AP, Cameron NM. Estrogenic mediation of serotonergic and neurotrophic systems: implications for female mood disorders. Prog Neuropsychopharmacol Biol Psychiatry. 2014;54:13–25. | ||
Payne JL. The role of estrogen in mood disorders in women. Int Rev Psychiatry. 2003;15:280–290. | ||
Westberg L, Eriksson E. Sex steroid-related candidate genes in psychiatric disorders. J Psychiatry Neurosci. 2008;33:319–330. | ||
Wittmann W, Schunk E, Rosskothen I, et al. Prodynorphin-derived peptides are critical modulators of anxiety and regulate neurochemistry and corticosterone. Neuropsychopharmacology. 2009;34(3):775–785. | ||
Heldring N, Pike A, Andersson S, et al. Estrogen receptors: how do they signal and what are their targets. Physiol Rev. 2007;87:905–931. | ||
van Nas A, Guhathakurta D, Wang SS, et al. Elucidating the role of gonadal hormones in sexually dimorphic gene coexpression networks. Endocrinology. 2009;150:1235–1249. | ||
Azcoitia I, Yague JG, Garcia-Segura LM. Estradiol synthesis within the human brain. Neuroscience. 2011;191:139–147. | ||
Jie C, Shen Y, Li R. Estrogen synthesis and signaling pathways during aging: from periphery to brain. Trends Mol Med. 2013;19:197–209. | ||
Prossnitz ER, Barton M. Signaling, physiological functions and clinical relevance of the G protein-coupled estrogen receptor GPER. Prostaglandins Other Lipid Mediat. 2009;89:89–97. | ||
Anchan D, Clark S, Pollard K, Vasudevan N. GPR30 activation decreases anxiety in the open field test but not in the elevated plus maze test in female mice. Brain Behav. 2014;4:51–59. | ||
Hart D, Nilges M, Pollard K, et al. Activation of the G-protein coupled receptor 30 (GPR30) has different effects on anxiety in male and female mice. Steroids. 2014;81:49–56. | ||
Kastenberger I, Lutsch C, Schwarzer C. Activation of the G-protein-coupled receptor GPR30 induces anxiogenic effects in mice, similar to oestradiol. Psychopharmacology. 2012;221:527–535. | ||
Kastenberger I, Schwarzer C. GPER1 (GPR30) knockout mice display reduced anxiety and altered stress response in a sex and paradigm dependent manner. Horm Behav. 2014;66:628–636. | ||
Liu S, Tian Z, Guo Y, Zhang N, Feng B, Zhao M. Activation of GPR30attenuates chronic pain-related anxiety in ovariectomized mice. Psychoneuroendocrinology. 2015;53:94–107. | ||
Tian Z, Wang Y, Zhang N, et al. Estrogen receptor GPR30 exerts anxiolytic effects by maintaining the balance between GABAergic and glutamatergic transmission in the basolateral amygdala of ovariectomized mice after stress. Psychoneuroendocrinology. 2013;38:2218–2233. | ||
Osterlund MK, Witt MR, Gustafsson JA. Estrogen action in mood and neurodegenerative disorders: estrogenic compounds with selective properties-the next generation of therapeutics. Endocrine. 2005;28:235–242. | ||
Ryan J, Ancelin ML. Polymorphisms of estrogen receptors and risk of depression: therapeutic implications. Drugs. 2012;72:1725–1738. | ||
Watson CS, Alyea RA, Cunningham KA, Jeng YJ. Estrogens of multiple classes and their role in mental health disease mechanisms. Int J Womens Health. 2010;2:153–166. | ||
Douma SL, Husband C, O’Donnell ME, Barwin BN, Woodend AK. Estrogen-related mood disorders: reproductive life cycle factors. ANS Adv Nurs Sci. 2005;28:364–375. | ||
Andrade TG, Nakamuta JS, Avanzi V, Graeff FG. Anxiolytic effect of estradiol in the median raphe nucleus mediated by 5-HTlA receptors. Behav Brain Res. 2005;163:18–25. | ||
Matsuda T, Nakano Y, Kanda T, Iwata H, Baba A. Gonadal hormones affect the hypothermia induced by serotoninl A (5-HTlA) receptor activation. Life Sci. 1991;48:1627–1632. | ||
Summer BE, Fink G. Estrogen increases the density of 5-hydroxytryptamine(2A) receptors in cerebral cortex and nucleus accumbens in the female rat. J Steroid Biochem Mol Biol. 1995;54:15–20. | ||
Halbreich U, Kahn LS. Role of estrogen in the aetiology and treatment of mood disorders. CNS Drugs. 2001;15:797–817. | ||
Lebesgue D, Traub M, De Butte-Smith M, et al. Acute administration of non-classical estrogen receptor agonists attenuates ischemia-induced hippocampal neuron loss in middle-aged female rats. PLoS One. 2010;5(1):e8642. | ||
Zhao TZ, Ding Q, Hu J, He SM, Shi F, Ma LT. GPER expressed on microglia mediates the anti-inflammatory effect of estradiol in ischemic stroke. Brain Behav. 2016;6(4):e00449. | ||
Graae L, Karlsson R, Paddock S. Significant association of estrogen receptor binding site variation with bipolar disorder in females. PLoS One. 2012;7:e32304. | ||
Owman C, Blay P, Nilsson C, Lolait SJ. Cloning of human cDNA encoding a novel heptahelix receptor expressed in Burkitt’s lymphoma and widely distributed in brain and peripheral tissues. Biochem Biophys Res Commun. 1996;228:285–292. | ||
Prossnitz ER, Barton M. The G-protein-coupled estrogen receptor GPER in health and disease. Nat Rev Endocrinol. 2011;7:715–726. | ||
Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science. 2005;307:1625–1630. | ||
Maggiolini M, Vivacqua A, Fasanella G, et al. The G protein-coupled receptor GPR30 mediates c-fos up-regulation by 17beta-estradiol and phytoestrogens in breast cancer cells. J Biol Chem. 2004;279:27008–27016. | ||
Brailoiu E, Dun SL, Brailoiu GC, et al. Distribution and characterization of estrogen receptor G protein-coupled receptor 30 in the rat central nervous system. J Endocrinol. 2007;193:311–321. | ||
Osterlund MK, Hurd YL. Estrogen receptors in the human forebrain and the relation to neuropsychiatrie disorders. Prog Neurobiol. 2001;64:251–267. | ||
Ostlund H, Keller E, Hurd YL. Estrogen receptor gene expression in relation to neuropsychiatrie disorders. Ann NY Acad Sci. 2003;1007:54–63. | ||
Altun I, Kurutas EB. G protein-coupled estrogen receptor levels after peripheral nerve injury in an experimental rat model. World Neurosurg. 2015;84:1903–1906. | ||
Cheng D, Liang B, Hao Y, Zhou W. Estrogen receptor alpha gene polymorphisms and risk of Alzheimer’s disease: evidence from a meta-analysis. Clin Interv Aging. 2014;9:1031–1038. | ||
Dun SL, Brailoiu GC, Gao X, et al. Expression of estrogen receptor GPR30 in the rat spinal cord and in autonomic and sensory ganglia. J Neurosci Res. 2009;87:1610–1619. | ||
Filardo EJ, Thomas P. Minireview: G protein-coupled estrogen receptor-1, GPER-1: its mechanism of action and role in female reproductive cancer, renal and vascular physiology. Endocrinology. 2012;153:2953–2962. | ||
Stygar D, Westlund P, Eriksson H, Sahlin L. Identification of wild type and variants of oestrogen receptors in polymorphonuclear and mononuclear leucocytes. Clin Endocrinol. 2006;64:74–81. | ||
Han G, Li F, Yu X, White RE. GPER: a novel target for non-genomic estrogen action in the cardiovascular system. Pharmacol Res. 2013;71:53–60. | ||
Pelekanou V, Kampa M, Kiagiadaki F, et al. Estrogen anti-inflammatory activity on human monocytes is mediated through cross-talk between estrogen receptor ERα36 and GPR30/GPER1. J Leukoc Biol. 2016;99:333–347. | ||
Rettew JA, McCall SH, Marriott I. GPR30/GPER-1 mediates rapid decreases in TLR4 expression on murine macrophages. Mol Cell Endocrinol. 2010;328:87–92. | ||
Prossnitz ER, Oprea TI, Sklar LA, Arterburn JB. The ins and outs of GPR30: a transmembrane estrogen receptor. J Steroid Biochem Mol Biol. 2008;109(3–5):350–353. | ||
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM5). 5th ed. Washington, DC: American Psychiatric Press; 2013. | ||
Frey BN, Macritchie KA, Soares CN, Steiner M. Bipolar disorder in women. In: Yatham LN, Maj M, editors. Bipolar Disorder: Clinical and Neurobiological Foundations. West Sussex: John Wiley & Sons; 2010:463–476. | ||
Freeman MP, Smith KW, Freeman SA, et al. The impact of reproductive events on the course of bipolar disorder in women. J Clin Psychiatry. 2002;63:284–287. | ||
Grigoriadis S, Kennedy SH. Role of estrogen in the treatment of depression. Am J Ther. 2002;9:503–509. | ||
Leibenluft E. Women with bipolar illness: clinical and research issues. Am J Psychiatry. 1996;153:163–173. | ||
Mahon PB, Payne JL, MacKinnon DF, et al. Genome-wide linkage and follow-up association study of postpartum mood symptoms. Am J Psychiatry. 2009;166:1229–1237. | ||
Viguera AC, Tondo L, Koukopoulos AE, Reginaldi D, Lepri B, Baldessarini RJ. Episodes of mood disorders in 2,252 pregnancies and postpartum periods. Am J Psychiatry. 2011;168:1179–1185. | ||
Suzuki S, Brown CM, Wise PM. Mechanisms of neuroprotection by estrogen. Endocrine. 2006;29:209–215. | ||
Lu CL, Herndon C. New roles for neuronal estrogen receptors. Neurogastroenterol Motil. 2017;29:1–7. | ||
Garcia-Segura LM, Balthazart J. Steroids and neuroprotection: new advances. Front Neuroendocrinol. 2009;30:v–ix. | ||
Kealey C, Reynolds A, Mynett-Johnson L, Claffey E, McKeon P. No evidence to support an association between the oestrogen receptor beta gene and bipolar disorder. Psychiatr Genet. 2001;11:223–226. | ||
Middle F, Jones I, Robertson E, Morey J, Lendon C, Craddock N. Variation in the coding sequence and flanking splice junctions of the estrogen receptor alpha (ERalpha) gene does not play an important role in genetic susceptibility to bipolar disorder or bipolar affective puerperal psychosis. Am J Med Genet B Neuropsychiatr Genet. 2003;118B:72–75. | ||
Weickert CS, Miranda-Angulo AL, Wong J, Perlman WR, Ward SE, Radhakrishna V. Variants in the estrogen receptor alpha gene and its mRNA contribute to risk for schizophrenia. Hum Mol Genet. 2008;17:2293–2309. | ||
Osterlund MK, Grandien K, Keller E, Hurd YL. The human brain has distinct regional expression patterns of estrogen receptor alpha mRNA isoforms derived from alternative promoters. J Neurochem. 2000;75:1390–1397. | ||
Osterlund MK, Gustafsson JA, Keller E, Hurd YL. Estrogen receptor beta (ERbeta) messenger ribonucleic acid (mRNA) expression within the human forebrain: distinct distribution pattern to ER mRNA. J Clin Endocrinol Metab. 2000;85:3840–3846. | ||
Osterlund MK, Keller E, Hurd YL. The human forebrain has discrete estrogen receptor alpha messenger RNA expression: high levels in the amygdaloid complex. Neuroscience. 2000;95:333–342. | ||
Perlman WR, Matsumoto M, Beltaifa S, et al. Expression of estrogen receptor alpha exon-deleted mRNA variants in the human and non-human primate frontal cortex. Neuroscience. 2005;134:81–95. | ||
Perlman WR, Webster MJ, Kleinman JE, Weickert CS. Reduced glucocorticoid and estrogen receptor alpha messenger ribonucleic acid levels in the amygdala of patients with major mental illness. Biol Psychiatry. 2004;56:844–852. | ||
Fink G, Sumner BE, McQueen JK, Wilson H, Rosie R. Sex steroid control of mood, mental state and memory. Clin Exp Pharmacol Physiol. 1998;25:764–775. | ||
Miller KJ, Conney JC, Rasgon NL, Fairbanks LA, Small GW. Mood symptoms and cognitive performance in women estrogen users and nonusers and men. J Am Geriatr Soc. 2002;50:1826–1830. | ||
Solomon MB, Herman JP. Sex differences in psychopathology: of gonads, adrenals and mental illness. Physiol Behav. 2009;97:250–258. | ||
Xu H, Qin S, Carrasco GA, et al. Extra-nuclear estrogen receptor GPR30 regulates serotonin function in rat hypothalamus. Neuroscience. 2009;158:1599–1607. | ||
Hazell GG, Yao ST, Roper JA, Prossnitz ER, O’Carroll AM, Lolait SJ. Localisation of GPR30, a novel G protein-coupled oestrogen receptor, suggests multiple functions in rodent brain and peripheral tissues. J Endocrinol. 2009;202:223–236. | ||
Prossnitz ER, Arterburn JB, Smith HO, Oprea TI, Sklar LA, Hathaway HJ. Estrogen signaling through the transmembrane G protein-coupled receptor GPR30. Annu Rev Physiol. 2008;70:165–190. | ||
Benmansour S, Weaver RS, Barton AK, Adeniji OS, Frazer A. Comparison of the effects of estradiol and progesterone on serotonergic function. Biol Psychiatry. 2012;71:633–641. | ||
McAllister CE, Mi Z, Mure M, Li Q, Muma NA. GPER1 stimulation alters posttranslational modification of RGSz1 and induces desensitization of 5-HT1A receptor signaling in the rat hypothalamus. Neuroendocrinology. 2014;100:228–239. | ||
Findikli E, Camkurt MA, Karaaslan MF, et al. Serum levels of G protein-coupled estrogen receptor 1 (GPER1) in drug-naive patients with generalized anxiety disorder. Psychiatry Res. 2016;244:312–316. | ||
Kurt AH, Buyukafsar K. Vasoconstriction induced by G1, a G-protein-coupled oestrogen receptor1 (GPER-1) agonist, in the isolated perfused rat kidney. Eur J Pharmacol. 2013;702:71–78. | ||
Muneer A. Bipolar disorder: role of inflammation and the development of disease biomarkers. Psychiatry Investig. 2016;13(1):18–33. | ||
Van der Schot A, Kahn R, Ramsey N, Nolen W, Vink M. Trait and state dependent functional impairments in bipolar disorder. Psychiatry Res. 2010;184(3):135–142. | ||
Matsuda K, Sakamoto H, Mori H, et al. (2008) Expression and intracellular distribution of the G protein-coupled receptor 30 in rat hippocampal formation. Neurosci Lett. 2008;441:94–99. | ||
Yildiz A, Guleryuz S, Ankerst DP, Ongür D, Renshaw PF. Protein kinase C inhibition in the treatment of mania: a double-blind, placebo-controlled trial of tamoxifen. Arch Gen Psychiatry. 2008;65:255–263. | ||
Kawa I, Carter JD, Joyce PR, et al. Gender differences in bipolar disorder: age of onset, course, comorbidity, and symptom presentation. Bipolar Disord. 2005;7:119–125. | ||
Amrollahi Z, Rezaei F, Salehi B, et al. Double-blind, randomized, placebo-controlled 6-week study on the efficacy and safety of the tamoxifen adjunctive to lithium in acute bipolar mania. J Affect Disord. 2011;129:327–331. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.