Back to Journals » Drug Design, Development and Therapy » Volume 18
Semaglutide Reduces Cardiomyocyte Damage Caused by High-Fat Through HSDL2
Authors Yang L ![]() , Pan X
, Pan X ![]() , Pan Z, Gao H, Ban J, Chen S
, Pan Z, Gao H, Ban J, Chen S
Received 15 October 2024
Accepted for publication 21 November 2024
Published 30 November 2024 Volume 2024:18 Pages 5501—5515
DOI https://doi.org/10.2147/DDDT.S495659
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Anastasios Lymperopoulos
Lin Yang,1,2 Xiaoyu Pan,2 Zhenyu Pan,3 Haina Gao,4 Jiangli Ban,4 Shuchun Chen2,5,6
1Department of Internal Medicine, Hebei North University, Zhangjiakou, Hebei, People’s Republic of China; 2Department of Endocrinology, Hebei General Hospital, Shijiazhuang, Hebei, People’s Republic of China; 3Department of Internal Medicine, Shandong Medical College, Linyi, Shandong, People’s Republic of China; 4Department of Endocrinology, The Second Hospital of Shijiazhuang, Shijiazhuang, Hebei, People’s Republic of China; 5Department of Internal Medicine, Hebei Medical University, Shijiazhuang, Hebei, People’s Republic of China; 6Hebei Key Laboratory of Metabolic Diseases, Hebei General Hospital, Shijiazhuang, Hebei, People’s Republic of China
Correspondence: Shuchun Chen, Department of Internal Medicine, Hebei Medical University, People’s Republic of China, Tel +86 31185988406, Email [email protected]
Introduction: Obesity-induced inflammation and oxidative stress can cause damage to cardiomyocytes. Semaglutide has the potential to reduce glucose levels and weight, while hydroxysteroid dehydrogenase-like protein 2 (HSDL2) also plays a role in regulating lipid metabolism. This study aimed to investigate the expression of oxidative stress markers and HSDL2 in myocardium and serum under high-fat conditions, in order to elucidate the mechanism of obesity-induced myocardial injury and evaluate the impact of semaglutide on myocardial injury through HSDL2.
Methods: Mouse models of obesity were established with semaglutide treatment. Palmitic acid-cultured mouse cardiomyocytes with HSDL2 knockout were used, as well as palmitic acid-induced high-fat environment models followed by semaglutide treatment. The levels of inflammatory and oxidative stress markers in serum and cardiomyocytes were measured. Additionally, the expression of HSDL2 and autophagy levels in different cell groups were assessed to evaluate the effect of semaglutide on high-fat diet-induced cardiomyocyte injury mediated by HSDL2.
Results: Obesity increased oxidative stress, which was alleviated by intervention with semaglutide. Furthermore, semaglutide down-regulated HSDL2 expression in obese individuals. Moreover, palmitic acid-induced oxidative stress and autophagy were reduced when using cells with knocked out HSDL2 gene.
Conclusion: These findings suggest that semaglutide may mitigate cardiomyocyte injury caused by a high-fat diet through regulation of HDLSDSLEP-1 expression. These discoveries are expected to unveil novel molecular mechanisms and provide new targets for clinical treatment.
Keywords: obesity, inflammation, oxidative stress, semaglutide, HSDL2
Introduction
Obesity is a chronic multifactorial condition characterized by the accumulation of multiple adipose tissues, which exerts detrimental effects on human health. With the advancement of the global economy, the prevalence of obesity continues to escalate in the global population, showing no apparent signs of abatement. Obesity directly contributes to the onset and progression of metabolic disorders, cardiomyopathy, ischemic coronary heart disease, peripheral arterial disease, heart failure, and arterial hypertension; thereby significantly impacting worldwide morbidity and mortality rates. Consequently, obesity represents a syndromic ailment that warrants sustained attention.
The presence of obesity induces a range of hemodynamic alterations that can result in modifications to cardiac morphology, ventricular function, and vascular endothelial cells, ultimately leading to the development of diverse cardiovascular disorders such as cardiac hypertrophy, heart failure, and atherosclerosis,1,2 the metabolic dysregulation induced by obesity plays a pivotal role in the pathogenesis of cardiovascular diseases. The findings of various studies have demonstrated that the consumption of a high-fat diet (HFD) leads to obesity in mice, accompanied by impaired endothelium-dependent vasodilation and increased arterial wall thickness.3 Additionally, obesity-induced lipid metabolism disorders significantly elevate oxidative stress levels, thereby potentially impairing various target organs and primarily affecting cardiovascular cell function, consequently initiating a cascade of pathological alterations. Concurrently, heightened oxidative stress also promotes the accumulation of lipid peroxidation products, leading to myocardial cell damage.4 The mechanism underlying obesity-induced myocardial injury, however, requires further investigation.
Glucagon-like peptide-1 receptor agonists (GLP-1RA) stimulate insulin secretion and lower blood glucose levels by activating the GLP-1 receptor. Semaglutide, a long-acting glucagon-like peptide-1 receptor agonist, also demonstrates glucose-lowering and weight-reducing properties. In a clinical trial, we observed a significant decrease in the risk of cardiac disease and an improved prognosis for patients with type 2 diabetes who were treated with semaglutide. However, most studies on GLP-1R agonists have primarily focused on the mechanism of diabetes-induced atherosclerosis, while limited research exists regarding their protective effects against early myocardial injury caused by obesity. Furthermore, there is a need for further investigation into the molecular mechanisms of these drugs and identification of new targets due to current knowledge gaps. It should be noted that obesity and diabetes share common pathogenic mechanisms that can contribute to cardiovascular complications.
The Hydroxysteroid dehydrogenase-like protein 2 (HSDL2) belongs to the short-chain dehydrogenase/reductase (SDR) family,5,6 is expressed in adipose tissue, cardiac muscle, and other various tissues. It exhibits catalytic activity for the oxidation and reduction of diverse substrates including steroids, sugars, retinoids, and fatty acids.7 Numerous studies have demonstrated that HSDL2 plays a pivotal role in the regulation of fatty acids within lipid metabolism and also actively participates in cholesterol synthesis.8–10 However, Shang et alobserved that the long non-coding RNA LNMICC could enhance lymph node metastasis in cervical cancer through reprogramming of fatty acid metabolism, and they also demonstrated the pivotal role of HSDL2 in regulating lipid metabolism.11 Recent studies have revealed that aberrant lipid metabolism can expedite the development of cardiomyopathy.12 The involvement of HSDL2 in cholesterol synthesis is facilitated through its binding to the coenzyme NADPH. It has been observed that overexpression of HSDL2 in vivo leads to a significant acceleration of intracellular cholesterol synthesis and a notable increase in total cholesterol levels within the organism.9 Meanwhile, our previous proteomic investigations on cardiovascular protein expression in mice subjected to a high-fat diet have revealed that Hsd17b4, HSDL2, and Acadvl are proteins primarily implicated in lipid metabolism. These proteins, identified as common differentially expressed proteins (Co-DEPs) in both the cardiac tissue and aorta, hold significant potential as novel targets for the diagnosis and treatment of obesity-related cardiovascular disease. Therefore, we postulate that both Hsd17b4 and HSDL2 exert regulatory roles in lipid metabolism and oxidative stress due to their association with catalase activity. Furthermore, peroxisomes play an indispensable role in maintaining cellular REDOX homeostasis.13 The dysregulation of HSDL2 has been observed in various cancer types and is closely associated with tumor progression.14,15 The protein HSDL2 localizes to peroxisomes and exerts a profound influence on tumor progression through its intricate regulation of lipid metabolism.7,8 The cell death phenomenon known as ferroptosis is characterized by an iron-dependent mechanism and the accumulation of lipid peroxidation. This mode of cellular demise exhibits distinct features compared to necrosis, apoptosis, and autophagy.16 The potential involvement of ferroptosis extends to neurogenesis, oncogenesis, and organ dysfunction.17,18 The scientific research has provided evidence for the association between HSDL2 and the initiation and progression of ferroptosis.19 However, it remains unclear whether abnormalities in HSDL2 are associated with obesity-induced myocardial injury.
In summary, we performed a comprehensive screening of cardiac and aortic Co-DEP based on proteomic results, followed by functional analysis of key proteins.13 The HSDL2 gene was selected for further investigation to elucidate the molecular mechanism underlying semaglutide’s protective effect on myocardial injury in obese mice and identify potential novel clinical therapeutic targets for obesity-related cardiomyopathy. To validate this mechanism, we established a high-fat cardiomyocyte model to observe changes in cardiomyocyte injury. Western blot (Wb) and polymerase chain reaction(PCR) techniques were employed to assess alterations in protein and mRNA levels in the cardiomyocytes of high-fat mice before and after semaglutide intervention as well as HSDL2 knockout. The present study aimed to explore the molecular mechanism through which semaglutide exerts its protective effect against high-fat diet-induced cardiomyocyte injury by regulating HSDL2.
Methods
Animals and Experimental Design
In this study, 24 male C57BL/6 mice at the age of 6 weeks were obtained from Hebei Shengwu Technology Co., LTD. The company possesses a license (Certificate number: SYXK)(20150004 June) for the sale and distribution of laboratory animals. Subsequently, the mice were housed at the Laboratory Animal Center of Hebei General Hospital, where they were maintained under standardized laboratory conditions including a temperature range of 22±2°C, humidity levels between 55±10%, a light-dark cycle lasting for 12 hours each, and ad libitum access to water. These standardized conditions ensured that the mice had an appropriate environment throughout the duration of the study.
After a one-week acclimatization period, the mice were randomly allocated into three groups: the normal diet group (NC), high-fat diet group (HF), and high-fat diet plus semaglutide group (Sema). It is important to note that all experimental procedures were approved by the Animal Ethics Committee of the General Hospital of Hebei Province and conducted in accordance with the Regulations on the Management of Experimental Animals of Hebei Province. The NC group was fed a normal diet (D1035, Beijing Huafukang Biotechnology Co., LTD., China) containing 4% fat, 20% protein, and 20% carbohydrate with a total energy content of 34.8 kcal/100 g. The HF group was fed a high-fat diet (H10060, Beijing Huafukang Biotechnology Co., LTD., China) containing 60% fat, 20% protein, and 20% carbohydrate with a total energy content of 524 kcal/100 g. The Sema group followed a similar high-fat diet for fourteen weeks while receiving daily subcutaneous injections of semaglutide at a dose of thirty nmol per kilogram body weight per day (Novo Nordisk, Bagsvaerd, Denmark). This dosage was selected based on previously published mouse studies. After eight weeks of treatment, glucose tolerance tests as well as measurements for body weight and serum levels were performed after fasting for twelve hours before euthanasia via intraperitoneal injection using one percent sodium pentobarbital at sixty mg/kg. Blood samples were collected from retroorbital sinuses in sterile tubes containing one millimeter ethylenediamine tetraacetic acid (EDTA). Finally, heart tissues from each mouse were collected then weighed before being stained with hematoxylin and eosin or frozen in liquid nitrogen then stored at −80°C for further analysis.
Weight Assessment
The body weights of mice were measured on a weekly basis throughout the duration of the experiment. These measurements can provide valuable data for assessing the impact of drug interventions on mouse physiology and metabolism.
Enzyme Linked Immunosorbent Assay (ELISA)
The measurement of plasma insulin (INS) levels was conducted using the Elabscience Mouse insulin ELISA Kit, manufactured by Wuhan Elarite Biotechnology Co., LTD. This kit is specifically designed for quantifying insulin levels in mouse plasma samples.
The levels of low-density lipoprotein cholesterol (LDL-C), high-density lipoprotein cholesterol (HDL-C), serum total cholesterol (TC), triglyceride (TG), inflammatory cytokines interleukin-1β (IL-1β) and tumor necrosis factor-alpha (TNF-α), as well as the oxidative stress markers malondialdehyde (MDA) and superoxide dismutase (SOD), were quantified using commercially available kits from Nanjing Jianchang. All measurements were conducted on a VERSAmax, a fully automated ELISA reader manufactured in the United States. Data obtained with the ELISA reader were analyzed using GraphPad Prism software.
IHC
Formalin-fixed paraffin sections (3µm thick) of mouse cardiac tissue were subjected to dewaxing and rehydration. Endogenous peroxidase activity was blocked using a 3% hydrogen peroxide solution. Slides were incubated overnight with a primary antibody against HSDL2 (1:200), followed by incubation with horseradish peroxidase (HRP)-conjugated immunoglobulin G (IgG) for 50 minutes at room temperature. The sections were stained using freshly prepared DAB chromogenic solution, and the color development time was meticulously controlled under microscopic observation to achieve a brown-yellow positive signal. Nuclei were counterstained with hematoxylin for 3 minutes. Finally, the sections were examined and interpreted under a white light microscope (Nikon Instruments Co., LTD., China).
Cell Culture
The mouse cardiomyocytes (HL-1) were cultured in DMEM medium (Gibco, USA, 22400089) supplemented with 10% fetal bovine serum(FBS) (Gibco, USA, 16140071) and 1% dual antibiotics (100µg/mL penicillin and 100 U/mL streptomycin). The cells were incubated at a temperature of 37 °C in a humidified incubator enriched with 5% CO2.
Study Subgroups
High-fat cell additives (sodium palmitate concentration is 6mmol/L) and complete medium at a ratio of 1:30, and finally palmitic acid (PA) 0.25mmol/L. The cell suspension is inoculated into the 6-hole plate, and the prepared palmitine solution is added after the cell is attached to the wall. It is cultured at 37°C for 24 hours to build a high-fat cardiomyocyte model. In the same way, the prepared semaglutide (concentration of 100nmol/L) will intervene in the high-fat cardiomyocyte model for 24 hours for the following experiments.
HSDL2 Transfection
The siHSDL2 (HSDL2 knockdown RNA) was processed by GenePharma (Shanghai, China). The cell lines were transfected with Lipofectamine 3000 transfection assay from Genechem (Shanghai, China). HSDL2 small interfering RNA (siRNA) (si-HSDL2) and siRNA control were obtained from GenePharma.
The HL-1 cells were transfected with siRNA using Lipofectamine 3000 according to the experimental protocol. The transfection efficiency was assessed by Western blotting after 24 hours, and the stably transfected cells were utilized in subsequent experiments. In the transfection sequence, the forward and reverse sequences of siRNA-HSDL2 were as follows: GGGAGGACCUGGUAUCGAATT and UUCGAUACCAGGUCCUCCCTT, respectively. The cells were divided into four groups: palmitic acid (PA) group, palmitic acid plus semaglutide (Sema) group, HSDL2 knockout plus palmitic acid (Si-HSDL2) group, and normal control (NC) group.
Western Blotting
The protein extraction was performed using cell lysis buffer (Biosharp, China, BL509A) added with phosphatase inhibitors (Biosharp, China, BL507A). The protein concentrations were measured by the bicinchoninic acid protein assay kit (Servicebio, China,G2026). The proteins were subjected to sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and transferred to a polyvinylidene difluoride (PVDF) membrane (Immobilon-P, USA). After being blocked with 5% skim milk powder, appropriate dilutions of rabbit-derived HSDL2 antibody (Proteintech, USA), P62 antibody (Servicebio, China), LC3 antibody (Servicebio, China), and mouse-derived GAPDH antibody (ABclonal, USA) primary antibodies were incubated overnight at 4 °C. Then the cells were incubated with goat anti-rabbit IgG secondary antibody (ABclonal, USA) or anti-mouse IgG secondary antibody (ABclonal, USA) for 1.5 hours at room temperature. Finally, the film was exposed to obtain images using an Amersham Imager 600 ultrasensitive Multifunction imager (General Electric, American).
Quantitative Reverse Transcription PCR (qRT-PCR)
The total RNA was extracted using TRIzol reagent, followed by reverse transcription into complementary DNA (cDNA) using SweScript All-in-one RT SuperMix for qPCR (Servicebio, China, G3337). The quantification of the cDNA samples was performed using SuperReal PreMix Plus (SYBR Green) (TIANGEN, China, FP205) on a StepOnePlus real-time PCR system (Applied Biosystem, USA, Cat#4376600). The results were analyzed and calculated using the 2-ΔΔCT method. Table 1 provides the details of the primers used for qRT-PCR.
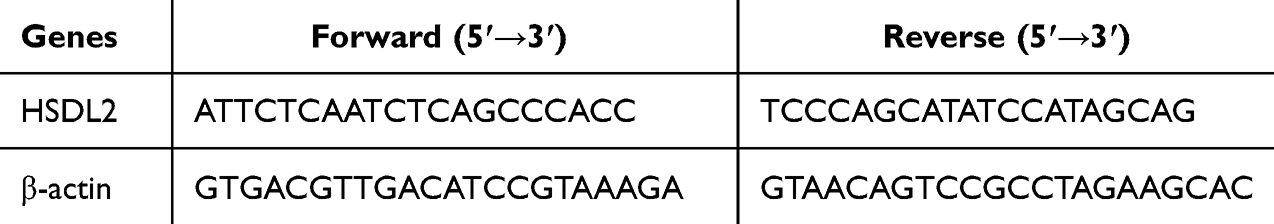 |
Table 1 Sequences of Primers Used for qPCR |
Cell Viability Detection
The cell viability was assessed using the CCK-8 assay (Biosharp, China, BS350A). Cells were seeded at a density of 1×103 cells per well in 96-well plates and cultured in DEME medium supplemented with 10% fetal bovine serum (FBS) for 24 hours at 37 °C and 5% CO2 in an incubator. After cell adhesion, the PA group was treated with a concentration of 20% PA for 24 hours, while the Sema group was also treated with a concentration of 20% PA for the same duration followed by treatment with semaglutide at a concentration of 200 nM/L for another period of 24 hours. Subsequently, the cells were incubated with CCK-8 solution for two hours. The absorbance at a wavelength of 450 nm was measured using a Synergy HT multimode microwell plate reader (BioTek, USA).
IF
To detect the expression of HSDL2, cells were seeded in confocal small dishes and cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum for 24 hours at a temperature of 37 °C and a CO2 concentration of 5%. The Si group was treated with 20% palmitic acid for 24 hours, the PA group was treated with the same duration of exposure to 20% palmitic acid, while the Sema group received treatment with both 20% palmitic acid followed by an additional period of time lasting another day with a concentration of semaglutide equal to 200nmmol/L. Following these treatments, cells were fixed using a solution containing paraformaldehyde at a concentration of 4%, permeabilized using permeabilizing solution, and blocked by incubation in Bovine Serum Protein (BSA) at a concentration of 1%. Subsequently, samples were incubated overnight with primary antibody against HSDL2 (dilution ratio:1:500), followed by incubation with fluorescently labeled anti-rabbit IgG conjugate specific to antibody isotype IgG subtype γ bound to Alexa Fluor® dye number-486 for one and half hours at room temperature. Nuclei were stained using Hoechst dye and visualized under laser confocal microscope (Observer Z1, Zeiss, Germany).
Enzyme-Linked Immunosorbent Assay
The levels of interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) in mice were quantified using corresponding ELISA kits (Esebio, Shanghai, China). The cell supernatant from each group was added to the wells and incubated with horseradish peroxidase (HRP) labeled detection antibody at 37°C for 60 minutes. After removing the liquid, the well plate was cleaned. Chromogens A and B were added and incubated at 37 °C in the dark for 10 minutes. Subsequently, 50µL of reaction termination solution was added to each well, and the absorbance (OD) value of each well was measured at a wavelength of 450nm.
Measurement of ROS Production
The levels of reactive oxygen species (ROS) were quantified using a fluorescent probe, dichlorodihydrofluorescein diacetate (DCFH-DA, Zomanbio, China). Following treatment, 10μM DCFHDA was added to each experimental group as per the manufacturer’s instructions. Subsequently, cells were incubated for 30 minutes, washed three times with DMEM, and the fluorescence intensity was measured using laser confocal microscopy (BD Biosciences, USA).
Assessment of MDA
The treated cells were incubated with 300μL of a mixed MDA working solution containing thiobarbituric acid (TBA) for 60 minutes to facilitate the detection of malondialdehyde levels. Subsequently, centrifugation was performed to remove the supernatant, and analysis was conducted using a Gen5 multifunctional microplate analyzer (Gene, USA).
Cell Apoptosis Assay
The Annexin V-FITC/propidium iodide (PI) Apoptosis assay kit (Elabscience, China) was utilized to perform apoptosis assays in accordance with the reagent manufacturer’s protocol. A personal-type flow cytometer (BD Biosciences, USA) was employed to analyze at least 10,000 cells within the gated area for apoptosis. The results were expressed as a percentage of total cells.
Statistical Processing
The Student’s t-test was employed to compare the means of the two groups of data. One-way analysis of variance (ANOVA) was utilized to compare data means among three or more groups. A statistically significant difference between groups was defined as P < 0.05. All statistical analyses were conducted using GraphPad 8.0 software.
Results
Changes in Weight in Three Groups of Mice
Initially, there were no discernible differences in body weight among the three groups of mice. However, following a 14-week high-fat diet, the HF group exhibited a significant increase in body weight compared to the NC group (Supplementary Figure 1, P<0.01), indicating successful establishment of the obesity model. Subsequently, after 8 weeks of semaglutide treatment, the Sema group demonstrated significantly lower body weight than the HF group (Supplementary Figure 2, P<0.01). Nevertheless, from week 18 onwards (Supplementary Figure 2, P > 0.05), there was no notable disparity in body weight between the Sema and NC groups. The alterations in body weight due to high-fat diet and semaglutide treatment are depicted through line plots in Supplementary Figure 3.
Morphological Alterations of Mice Cardiomyocytes
Compared to the NC group, the PA group exhibited reduced cardiomyocyte volume, slight disarray, increased extracellular matrix deposition, varying nuclear sizes, and evident lipid droplet formation. Semaglutide intervention demonstrated a mitigating effect on the pathological alterations of cardiomyocytes induced by palmitic acid (Supplementary Figures10–13).
Changes in Blood Sugar and Blood Lipids in Three Groups of Mice
The levels of total cholesterol(TC), high-density lipoprotein cholesterol(HDL-C), low-density lipoprotein cholesterol(LDL-C), fasting blood glucose(FBG), and oral glucose tolerance test(OGTT) were measured to assess changes in blood lipid and glucose levels in obese mice and the effects of semaglutide on regulating glucose and lipid disorders. Serum levels of TC, HDL-C, LDL-C and FBG were significantly higher in the high-fat group compared to the normal control group (Supplementary Figures 4–7; P <0.01). In contrast, the Sema group showed significant reductions in serum lipid and FBG levels (Supplementary Figures 4–7; P <0.01). Additionally, OGTT levels were significantly higher in the HF group compared to other groups (Supplementary Figure 8; P <0.01). Interestingly, the Sema group exhibited an earlier increase in oral glucose tolerance test at 15 minutes compared to the NC group’s increase at 30 minutes; thereafter their levels remained similar (Supplementary Figure 8; P >0 0.05). Furthermore, the serum insulin level was significantly lower in both Sema groups when compared with that of NC groups (Supplementary Figure 9, P<0 0.01).
The Effect of Smeglupeptide on the Level of Inflammation and Oxidative Stress Caused by Obesity
The levels of TNF-α, MDA, IL-1β, T-SOD, and adenosine in the myocardial tissue of obese mice exhibited a significant elevation compared to those in normal mice (Figure 1, P <0.05). Conversely, there was a notable reduction in the levels of TNF-α, IL-6, MDA, ROS, IL-1β, T-SOD and adenosine in myocardial tissue (Figure 1, P <0.05).
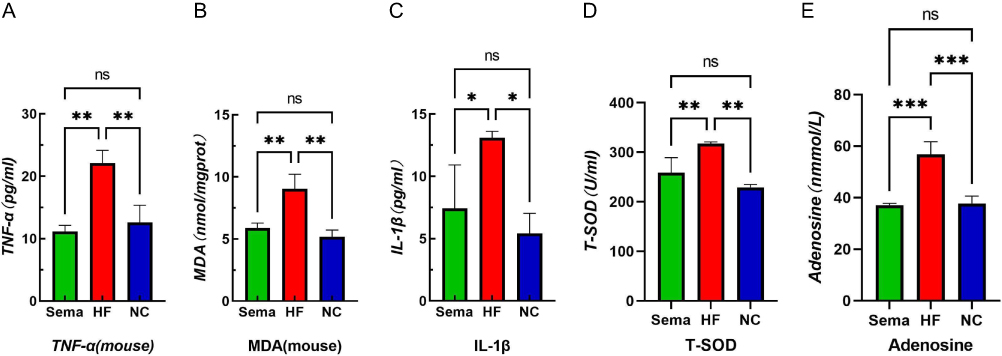 |
Figure 1 Effects of obesity and semaglutide on myocardial inflammation and oxidative stress levels in mice. (A) TNF-α, (B)MDA, (C) IL-1β, (D) T-SOD, (E) Adenosine. Notes: ***P<0.01, **P<0.05, *P<0.05, ns P>0.05. |
Effect of HSDL2 on Myocardial Cells in Obese Mice
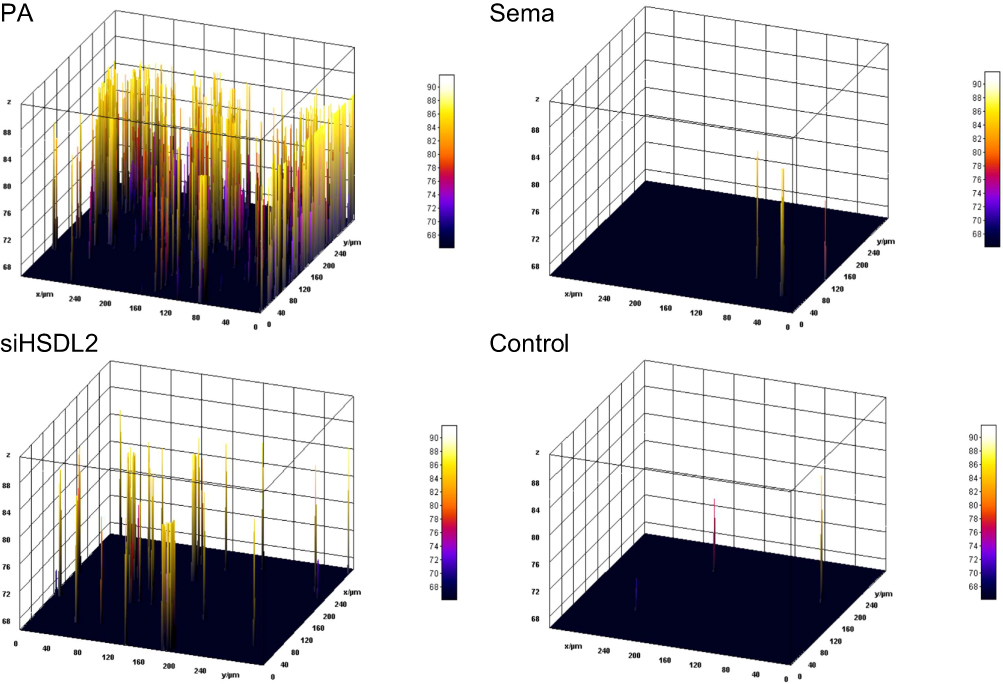
Exposure to a high-fat environment induced by PA resulted in increased production of ROS) in cardiomyocytes, which was attenuated by knockdown of HSDL2 (Figure 2, P < 0.05). And after transfection of HSDL2-siRNA, there was a corresponding trend in the expression of HSDL2 in Wb, and an increase in the expression of HSDL2 in the PA group. The results obtained from PCR and immunofluorescence (IF) were consistent with those observed in Wb analysis (Figure 3, P < 0.05). Additionally, PA promoted autophagy in cardiomyocytes, and this effect was partially alleviated by downregulation of HSDL2 expression (Figure 3, P < 0.05). Figure 2 Continued. Figure 2 Effect of semaglutide on cardiomyocyte function in mice (A) TNF–α (B) IL–6 (C) MDA (D) ROS (E) ROS fiuorescence intensity bands. Notes: ****P<0.001, ***P<0.01, **P<0.05, *P<0.05, ns P>0.05. Figure 3 Continued. Figure 3 Effect of HSDL2 on cardiomyocyte function in mice. (A) HSDL2 protein expression bands, (B) Relative HSDL2 protein expression, (C) Relative LC3 protein expression, (D) Relative P62 protein expression, (E) mean fluorescence intensity of HSDL2, (F) mean fluorescence intensity of P62, (G) HSDL2 fluorescence intensity bands, (H) P62 fluorescence intensity bands. Notes: ****P<0.001, ***P<0.01, **P<0.05, *P<0.05, ns P>0.05.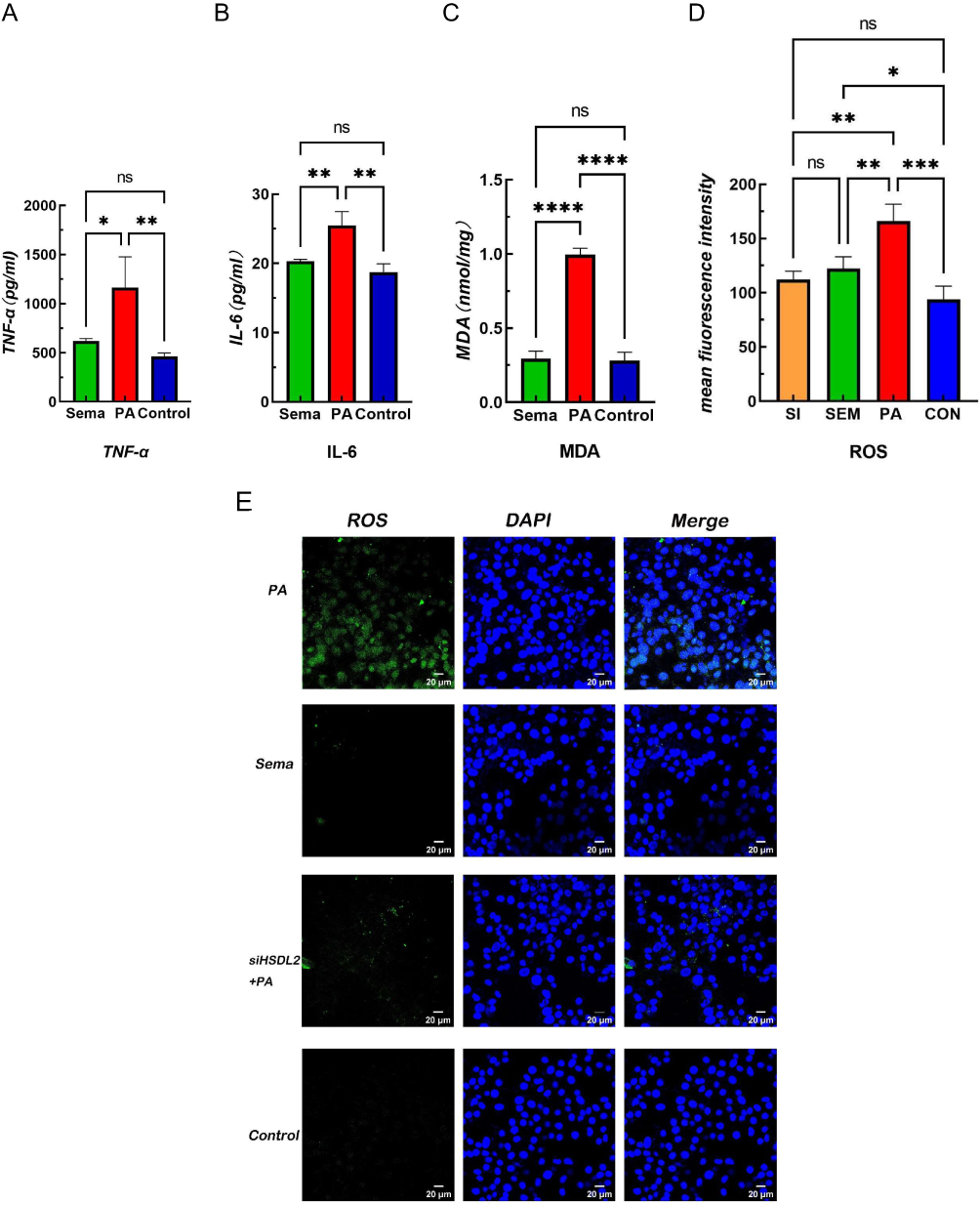
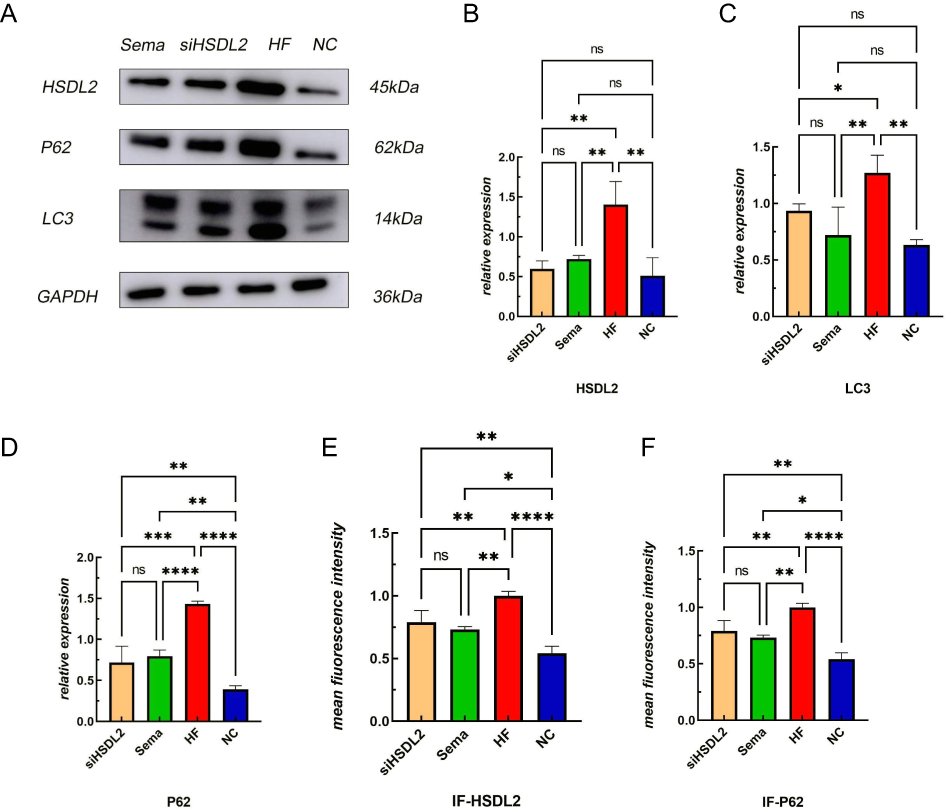
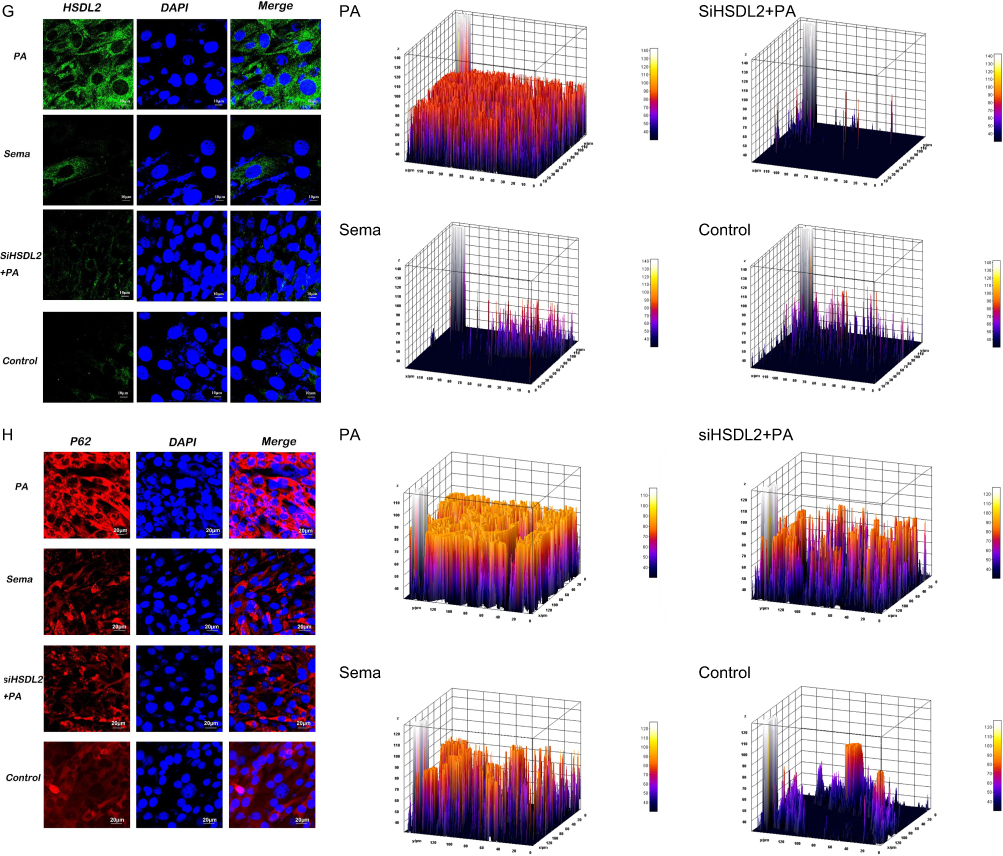
Statistical Processing
The levels of TNF-α, IL-6, MDA, and ROS in the Sema group exhibited a significant decrease compared to those in the control group (Figure 2, P <0.05). Moreover, Sema treatment significantly enhanced cell viability and mitigated PA-induced apoptosis (Figure 4, P <0.001). Furthermore, both Wb and IF results demonstrated that Sema attenuated autophagy induced by PA (Figure 3, P<0.05). In terms of HSDL2 expression, Sema treatment led to a reduction in protein expression levels of HSDL2 in cardiomyocytes when compared with the PA group; this finding was consistent with PCR results (Figure 5, P<0.05). Additionally, immunohistochemistry analysis revealed similar trends for HSDL2 expression between the Sema and PA groups as observed through Wb analysis (Figure 5, P<0.05).
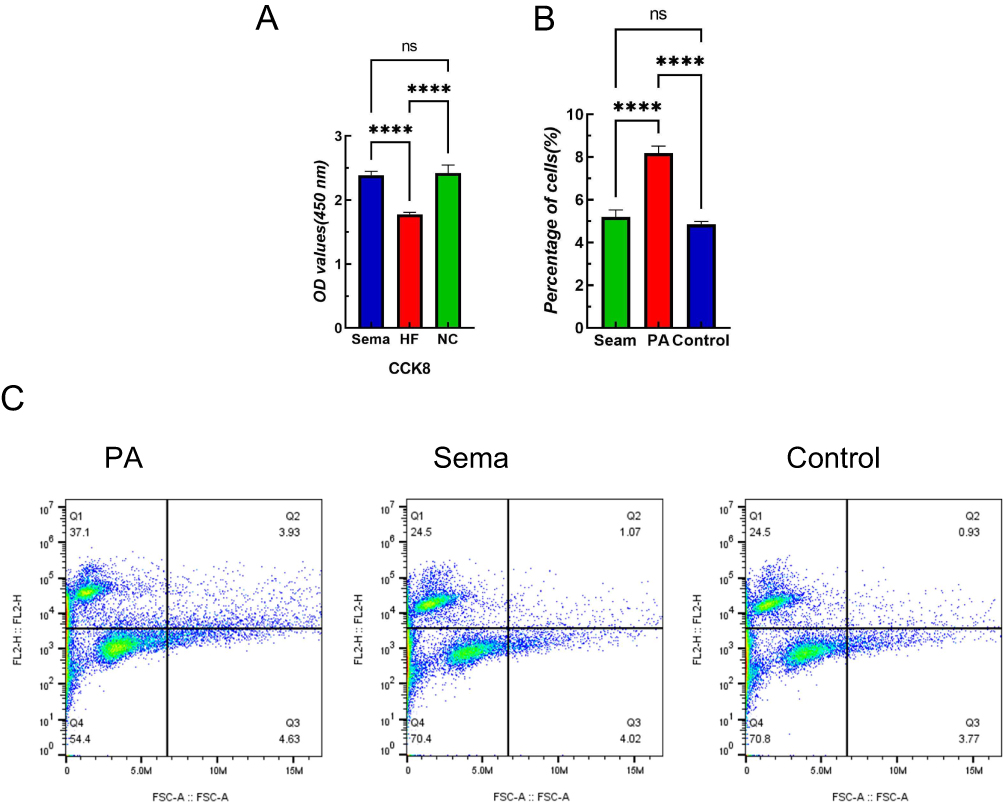 |
Figure 4 Semaglutide reduces apoptosis and increases cell viability in mouse cardiomyocytes induced by high-fat diet. (A) Cardiomyocyte viability assay, (B) Cardiomyocyte apoptosis rate. (C) Flow cytometry. Notes: ****P<0.001, ns P>0.05. |
 |
Figure 5 Effect of semaglutide on cardiomyocyte function in mice (A) HSDL2 IOD ration expression bands. (B) Relative mRNA expression of HSDL2. Notes: ****P<0.001, ***P<0.01, **P<0.05, ns P>0.05. |
Discussion
In recent years, the prevalence of obesity has witnessed a remarkable upsurge owing to the enhancement of individuals’ living standards. This surge in adiposity has posed significant threats to people’s health by inflicting substantial damage on vital organs such as the heart, pancreas, and kidneys.20 According to a multitude of clinical and fundamental experiments, obesity exerts its deleterious effects on the human body by significantly elevating lipid and glucose levels, thereby precipitating metabolic disorders.21 The present study has demonstrated that diet-induced obesity in a murine model leads to an elevation in blood levels of triglycerides (TG), TC, LDL-C, and fasting plasma glucose. This concomitant increase in circulating lipid and glucose concentrations, along with the inflammatory milieu associated with obesity, accelerates lipid oxidation, thereby promoting the development of atherosclerosis and impairing cardiac function.22 The simultaneous occurrence of obesity-induced elevation in LDL-C and reduction in HDL-C is associated with oxidative stress, which subsequently triggers extensive lipid oxidation and further amplifies the production of lipid peroxidation products.23 The activation of multiple oxidoreductases, in conjunction with a decrease in the activity of cellular antioxidant enzymes such as Cu-Zn SOD, catalase, and GPx, can initiate lipid peroxidation and result in an elevation of ROS levels that cause detrimental effects on cells and tissues.24,25 Additionally, obesity also led to significantly elevated levels of TNF-α and IL-6 in both serum and cardiac tissue, indicating the presence of inflammation in vivo. However, obesity-induced increase in ROS within the cardiac tissue may result in the generation of toxic MDA. The excessive production of MDA through lipid peroxidation is attributed to enhanced lipid synthesis, inflammation, and oxidative stress. Levels of MDA in cardiac tissue are associated with mitochondrial activity, cell membrane damage, and the extent of peroxidation injury. Alterations in lipid profiles and indicators of oxidative stress can impact myocardial cell metabolism, thereby influencing the progression of obesity-related cardiomyopathy.26,27 The presence of myocardial hypertrophy and ventricular wall thickening can still provide sufficient compensatory function in pre-obesity; however, decompensation may lead to irreversible and severe impairment of cardiac function.28,29 The echocardiography analysis conducted in our previous study on obese mice revealed concentric hypertrophy, characterized by ventricular wall thickening and reduced ventricular volume.30 In this study, we established mouse models of HF group and Sema group, as well as cell models of PA group and Sema group, to gain a comprehensive understanding of the mechanisms underlying obesity-induced myocardial injury and investigate the potential protective effects of semaglutide. Compared to the Sema group, the HF group exhibited increased body weight and elevated levels of TC, TG, and LDL-C. Moreover, serum levels of TNF-α, IL-6, ROS, and MDA were significantly higher in the HF group. The PA group demonstrated similar patterns of oxidative stress expression compared to the HF group. Additionally, cardiomyocyte morphology in the PA group showed alterations characterized by lipid droplet formation, cellular atrophy, and extensive cell debris resulting from cell death. The cck8 assay revealed a significant decrease in cell viability within the PA group. Autophagy and apoptosis were also markedly increased in this experimental cohort. These findings suggest that obesity leads to elevated lipid levels and oxidative stress within the body, resulting in enhanced production of lipid peroxidation products which subsequently induce myocardial damage and trigger myocardial diseases.
The HSDL2 gene is located on the 9Q32 region of human chromosome 9, spanning a length of 92 kilodaltons. It consists of 11 exons and encodes a protein comprising 418 amino acids. Belonging to the SDR family, HSDL2 is classified as one among several proteins that facilitate oxidation and reduction reactions involving diverse substrates such as steroid hormones, oxysterols, bile acids (BAs), prostaglandins, retinoids, fatty acids, amino acids, and sugars.31 The HSDL2 protein consists of an N-terminal SDR domain and a C-terminal SCP2 domain, both of which play pivotal roles in nicotinamide adenine dinucleotide (oxidized form) (NAD)/NAD phosphate (NADP)-dependent REDOX reactions and the binding of sterols and other lipids, respectively.32,33 The capacity of HSDL2 to interact with NADP/H, sterol derivatives, and fatty acids can be attributed to its possession of a well-preserved NADP/H binding site and SCP2 domain. These molecular features serve as crucial structural components that facilitate the transportation of sterols and various lipids across cellular membranes.34,35 In previous studies, the impact of HSDL2 on lipid metabolism was only proposed without further investigation into its association with high-fat myocardium. This study reveals a significantly higher abundance of brown particles in the HF group compared to the Sema group, indicating upregulation of HSDL2 expression in the former and downregulation in the latter. The results obtained from Wb, PCR, and IF analyses in both PA and Sema groups were consistent. Therefore, it can be concluded that a high-fat diet indeed enhances myocardial HSDL2 expression while semaglutide suppresses it. To determine whether HSDL2 is one of the mechanisms through which high-fat induces changes in myocardial function, we conducted knockdown experiments targeting HSDL2 expression. The inhibition of HSDL2 expression demonstrated amelioration of autophagy and oxidative stress induced by a high-fat diet in cardiomyocytes as observed through IF analysis for autophagy markers and ROS. Based on these findings, it can be inferred that HSDL2 may serve as one mechanism contributing to myocardial injury. Additionally, considering the aforementioned association between HSDL2 and ferroptosis, it is plausible to speculate that semaglutide may potentiate cellular ferroptosis through its interaction with HSDL2, thereby establishing a robust foundation for further clinical investigations.
Semaglutide, a glucagon-like peptide-1 receptor agonist, has been extensively studied in numerous trials and consistently demonstrated its efficacy in reducing blood glucose levels and promoting weight loss.36 The weight loss efficacy of semaglutide has been extensively investigated in recent clinical practice, demonstrating a reduction in basal body weight by 20 to 30% with minimal adverse effects.37,38 The effectiveness of semaglutide in mitigating cardiovascular events has been established among individuals diagnosed with type 2 diabetes, while also offering cardioprotective benefits to overweight or obese patients suffering from preexisting cardiovascular disease but not afflicted by diabetes.39,40 The results of our previous study suggest that semaglutide possesses the potential to ameliorate the detrimental cardiac effects associated with obesity and modulate the expression of numerous proteins involved in lipid metabolism within cardiac tissue.30 The results of this study demonstrate the effective reduction of body weight and blood lipid levels in mice with HF by semaglutide. Additionally, semaglutide decreased serum lipids, TNF-α, IL-6, ROS, and MDA levels in both the HF group and PA-exposed myocardial cells. Although semaglutide did not alter PA-induced cell morphology, microscopic examination revealed a significant increase in cell death. Moreover, the Sema group exhibited higher cell viability compared to the PA group. Furthermore, the Sema group displayed significantly lower levels of autophagy and apoptosis compared to the PA group. The findings suggest that semaglutide may mitigate myocardial inflammatory injury induced by a high-fat diet and restore myocardial viability. Semaglutide significantly reduces the expression of HSDL2 in the high-fat myocardium, as mentioned above. Based on these results, HSDL2 might be one of the mechanisms contributing to myocardial injury, while semaglutide appears to protect the myocardium by modulating HSDL2 expression.
Conclusions
Our study provides evidence that obesity triggers oxidative stress, inflammatory response, autophagy, and apoptosis in cardiomyocytes. Downregulating HSDL2 expression significantly attenuates autophagy levels and oxidative stress in these cells. Moreover, semaglutide not only alleviates cellular damage induced by obesity but also suppresses the upregulation of HSDL2 resulting from this condition. These findings offer a fresh insight into the use of GLP-1 receptor agonists as targeted interventions to regulate myocardial injury associated with obesity while identifying a potential therapeutic target for managing cardiac damage related to excessive weight gain.
Data Sharing Statement
Data available on request from the authors.
Ethics Approval
All experimental procedures and animal management procedures were approved by the Ethics Committee of Hebei General Hospital (Approval Number: 2023-32; Date of approval: Jun 14, 2023).
Acknowledgment
This paper has been uploaded to ResearchGate as a preprint: https://www.researchgate.net/publication/385060147_Semaglutide_reduces_cardiomyocyte_damage_caused_by_high-fat_through_HSDL2.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Hebei Province Natural Science Foundation (H2022307026). The funding bodies played no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pan X, Chen S, Chen X, et al. Effect of high-fat diet and empagliflozin on cardiac proteins in mice. Nutr Metab. 2022;19(1):69. doi:10.1186/s12986-022-00705-0
2. Rosengren A. Obesity and cardiovascular health: the size of the problem. Eur Heart J. 2021;42(34):3404–3406. doi:10.1093/eurheartj/ehab518
3. Ma L, Ma S, He H, et al. Perivascular fat-mediated vascular dysfunction and remodeling through the AMPK/mTOR pathway in high-fat diet-induced obese rats. Hypertens Res. 2010;33(5):446–453. doi:10.1038/hr.2010.11
4. Ionică LN, Gaiță L, Bînă AM, et al. Metformin alleviates monoamine oxidase-related vascular oxidative stress and endothelial dysfunction in rats with diet-induced obesity. Mol Cell Biochem. 2021;476(11):4019–4029. doi:10.1007/s11010-021-04194-2
5. Kowalik D, Haller F, Adamski J, Moeller G. In search for function of two human orphan SDR enzymes: hydroxysteroid dehydrogenase like 2 (HSDL2) and short-chain dehydrogenase/reductase-orphan (SDR-O). J Steroid Biochem Mol Biol. 2009;117(4–5):117–124. doi:10.1016/j.jsbmb.2009.08.001
6. Dai J, Xie Y, Wu Q, et al. Molecular cloning and characterization of a novel human hydroxysteroid dehydrogenase-like 2 (HSDL2) cDNA from fetal brain. Biochem Genet. 2003;41(5–6):165–174. doi:10.1023/A:1023377627138
7. Yang Y, Han A, Wang X, Yin X, Cui M, Lin Z. Lipid metabolism regulator human hydroxysteroid dehydrogenase-like 2 (HSDL2) modulates cervical cancer cell proliferation and metastasis. J Cell Mol Med. 2021;25(10):4846–4859. doi:10.1111/jcmm.16461
8. Han A, Xu R, Liu Y, Yin X, Lin Z, Yang W. HSDL2 Acts as a Promoter in Pancreatic Cancer by Regulating Cell Proliferation and Lipid Metabolism. Onco Targets Ther. 2021;14:435–444. doi:10.2147/OTT.S287722
9. Cheng Z, Li Y, Sui C, Sun X, Xie Y. Synthesis, purification and crystallographic studies of the C-terminal sterol carrier protein type 2 (SCP-2) domain of human hydroxysteroid dehydrogenase-like protein 2. Acta Crystallogr F Struct Biol Commun. 2015;71(Pt 7):901–905. doi:10.1107/S2053230X15008559
10. Gronemeyer T, Wiese S, Ofman R, et al. The proteome of human liver peroxisomes: identification of five new peroxisomal constituents by a label-free quantitative proteomics survey. PLoS One. 2013;8(2):e57395. doi:10.1371/journal.pone.0057395
11. Shang C, Wang W, Liao Y, et al. LNMICC Promotes Nodal Metastasis of Cervical Cancer by Reprogramming Fatty Acid Metabolism. Cancer Res. 2018;78(4):877–890. doi:10.1158/0008-5472.CAN-17-2356
12. Kim MS, Kim WJ, Khera AV, et al. Association between adiposity and cardiovascular outcomes: an umbrella review and meta-analysis of observational and Mendelian randomization studies. Eur Heart J. 2021;42(34):3388–3403. doi:10.1093/eurheartj/ehab454
13. Pan X, Zhang X, Ban J, Yue L, Ren L, Chen S. Effects of High-Fat Diet on Cardiovascular Protein Expression in Mice Based on Proteomics. Diabetes Metab Syndr Obes. 2023;16:873–882. doi:10.2147/DMSO.S405327
14. Zhang DY, Liu Z, Lu Z, et al. Lentivirus-mediated overexpression of HSDL2 suppresses cell proliferation and induces apoptosis in cholangiocarcinoma. Onco Targets Ther. 2018;11:7133–7142. doi:10.2147/OTT.S176410
15. Sun Q, Zhang Y, Su J, Li T, Jiang Y. Role of Hydroxysteroid Dehydrogenase-Like 2 (HSDL2) in Human Ovarian Cancer. Med Sci Monit. 2018;24:3997–4008. doi:10.12659/MSM.909418
16. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
17. Stockwell BR, Friedmann Angeli JP, Bayir H, et al. Ferroptosis: a Regulated Cell Death Nexus Linking Metabolism, Redox Biology, and Disease. Cell. 2017;171(2):273–285. doi:10.1016/j.cell.2017.09.021
18. Tang D, Chen X, Kang R, Kroemer G. Ferroptosis: molecular mechanisms and health implications. Cell Res. 2021;31(2):107–125. doi:10.1038/s41422-020-00441-1
19. Ma S, Ma Y, Qi F, et al. HSDL2 knockdown promotes the progression of cholangiocarcinoma by inhibiting ferroptosis through the P53/SLC7A11 axis. World J Surg Oncol. 2023;21(1):293. doi:10.1186/s12957-023-03176-6
20. Bunbupha S, Apaijit K, Maneesai P, Prasarttong P, Pakdeechote P. Nobiletin ameliorates high-fat diet-induced vascular and renal changes by reducing inflammation with modulating AdipoR1 and TGF-β1 expression in rats. Life Sci. 2020;260:118398. doi:10.1016/j.lfs.2020.118398
21. O’Neil PM, Birkenfeld AL, McGowan B, et al. Efficacy and safety of semaglutide compared with liraglutide and placebo for weight loss in patients with obesity: a randomised, double-blind, placebo and active controlled, dose-ranging, Phase 2 trial. Lancet. 2018;392(10148):637–649. doi:10.1016/S0140-6736(18)31773-2
22. Piché ME, Tchernof A, Després JP. Obesity Phenotypes, Diabetes, and Cardiovascular Diseases. Circ Res. 2020;126(11):1477–1500. doi:10.1161/CIRCRESAHA.120.316101
23. Cafe SL, Nixon B, Dun MD, Roman SD, Bernstein IR, Bromfield EG. Oxidative Stress Dysregulates Protein Homeostasis Within the Male Germ Line. Antioxid Redox Signal. 2020;32(8):487–503. doi:10.1089/ars.2019.7832
24. Rani V, Deep G, Singh RK, Palle K, Yadav UC. Oxidative stress and metabolic disorders: pathogenesis and therapeutic strategies. Life Sci. 2016;148:183–193. doi:10.1016/j.lfs.2016.02.002
25. Holvoet P, Kritchevsky SB, Tracy RP, et al. The metabolic syndrome, circulating oxidized LDL, and risk of myocardial infarction in well-functioning elderly people in the health, aging, and body composition cohort. Diabetes. 2004;53(4):1068–1073. doi:10.2337/diabetes.53.4.1068
26. Kolijn D, Pabel S, Tian Y, et al. Empagliflozin improves endothelial and cardiomyocyte function in human heart failure with preserved ejection fraction via reduced pro-inflammatory-oxidative pathways and protein kinase Gα oxidation. Cardiovasc Res. 2021;117(2):495–507. doi:10.1093/cvr/cvaa123
27. Watanabe K, Nagao M, Toh R, et al. Critical role of glutamine metabolism in cardiomyocytes under oxidative stress. Biochem Biophys Res Commun. 2021;534:687–693. doi:10.1016/j.bbrc.2020.11.018
28. Alpert MA, Omran J, Bostick BP. Effects of Obesity on Cardiovascular Hemodynamics, Cardiac Morphology, and Ventricular Function. Curr Obes Rep. 2016;5(4):424–434. doi:10.1007/s13679-016-0235-6
29. Harada T, Obokata M. Obesity-Related Heart Failure with Preserved Ejection Fraction: pathophysiology, Diagnosis, and Potential Therapies. Heart Fail Clin. 2020;16(3):357–368. doi:10.1016/j.hfc.2020.02.004
30. Pan X, Yue L, Ban J, Ren L, Chen S. Effects of Semaglutide on Cardiac Protein Expression and Cardiac Function of Obese Mice. J Inflamm Res. 2022;15:6409–6425. doi:10.2147/JIR.S391859
31. Kavanagh KL, Jörnvall H, Persson B, Oppermann U. Medium- and short-chain dehydrogenase/reductase gene and protein families: the SDR superfamily: functional and structural diversity within a family of metabolic and regulatory enzymes. Cell Mol Life Sci. 2008;65(24):3895–3906. doi:10.1007/s00018-008-8588-y
32. Persson B, Kallberg Y, Bray JE, et al. The SDR (short-chain dehydrogenase/reductase and related enzymes) nomenclature initiative. Chem Biol Interact. 2009;178(1–3):94–98. doi:10.1016/j.cbi.2008.10.040
33. Chiapparino A, Maeda K, Turei D, Saez-Rodriguez J, Gavin AC. The orchestra of lipid-transfer proteins at the crossroads between metabolism and signaling. Prog Lipid Res. 2016;61:30–39. doi:10.1016/j.plipres.2015.10.004
34. Bhatia C, Oerum S, Bray J, et al. Towards a systematic analysis of human short-chain dehydrogenases/reductases (SDR): ligand identification and structure-activity relationships. Chem Biol Interact. 2015;234:114–125. doi:10.1016/j.cbi.2014.12.013
35. Burgardt NI, Gianotti AR, Ferreyra RG, Ermácora MR. A structural appraisal of sterol carrier protein 2. Biochim Biophys Acta Proteins Proteom. 2017;1865(5):565–577. doi:10.1016/j.bbapap.2017.03.002
36. Ryan DH, Lingvay I, Colhoun HM, et al. Semaglutide Effects on Cardiovascular Outcomes in People With Overweight or Obesity (SELECT) rationale and design. Am Heart J. 2020;229:61–69. doi:10.1016/j.ahj.2020.07.008
37. Aroda VR, Ahmann A, Cariou B, et al. Comparative efficacy, safety, and cardiovascular outcomes with once-weekly subcutaneous semaglutide in the treatment of type 2 diabetes: insights from the SUSTAIN 1–7 trials. Diabetes Metab. 2019;45(5):409–418. doi:10.1016/j.diabet.2018.12.001
38. Thethi TK, Pratley R, Meier JJ. Efficacy, safety and cardiovascular outcomes of once-daily oral semaglutide in patients with type 2 diabetes: the PIONEER programme. Diabetes Obes Metab. 2020;22(8):1263–1277. doi:10.1111/dom.14054
39. Husain M, Birkenfeld AL, Donsmark M, et al. Oral Semaglutide and Cardiovascular Outcomes in Patients with Type 2 Diabetes. N Engl J Med. 2019;381(9):841–851. doi:10.1056/NEJMoa1901118
40. Bain SC, Mosenzon O, Arechavaleta R, et al. Cardiovascular safety of oral semaglutide in patients with type 2 diabetes: rationale, design and patient baseline characteristics for the PIONEER 6 trial. Diabetes Obes Metab. 2019;21(3):499–508. doi:10.1111/dom.13553
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.