Back to Journals » ImmunoTargets and Therapy » Volume 14
Selective Deletion of HLA-B, and -C Class I Genes Promotes Immunocompatibility of Humanized Skin Graft Model
Authors Cobos-Figueroa L ![]() , Notario L
, Notario L ![]() , Mir C, Molpeceres C, Lauzurica S, López D, Lorente E, Lauzurica P
, Mir C, Molpeceres C, Lauzurica S, López D, Lorente E, Lauzurica P
Received 23 November 2024
Accepted for publication 13 March 2025
Published 9 April 2025 Volume 2025:14 Pages 451—463
DOI https://doi.org/10.2147/ITT.S506352
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sarah Wheeler
Laura Cobos-Figueroa,1,2,* Laura Notario,1,* Carmen Mir,1 Carlos Molpeceres,2 Sara Lauzurica,2 Daniel López,1 Elena Lorente,1,* Pilar Lauzurica1,*
1Centro Nacional de Microbiología, Insituto de Salud Carlos III, Majadahonda, Madrid, Spain; 2Centro Láser, Universidad Politécnica de Madrid, Madrid, Madrid, Spain
*These authors contributed equally to this work
Correspondence: Laura Notario, Centro Nacional de Microbiología, Insituto de Salud Carlos III, Ctra. de Pozuelo, 28, Majadahonda, Madrid, 28222, Spain, Tel +34 918223718, Email [email protected]
Background: The treatment of extensive burns requires rapid allogeneic skin transplantation, but HLA diversity poses a significant challenge in finding histocompatible donor-recipient matches.
Methods: In this study, we developed a humanized skin graft model using HLA class I transgenic mice to closely examine the HLA-mediated immune response in skin transplantation. Additionally, this model was used to analyse the response against a human lymphoblastoid cell line, JY, with HLA-B and -C genes knocked out by a single-step CRISPR-Cas9 strategy, retaining the most common HLA class I allele, HLA-A*02:01.
Results: Mice expressing the HLA-A02:01 allele alone or in combination with HLA-B07:02 do not reject the skin of animals expressing only HLA-A02:01. However, skin from HLA-A02:01/B07:02 mice transplanted into HLA-A02:01 mice is rejected, triggering a strong specific CD8 T cell response mediated by the HLA-B*07:02 molecule. In these latter mice, unlike the parental JY cell line, the edited cells did not induce a CD8 T cell response in vitro, suggesting that the selective deletion of HLA-B and -C may contribute to improve skin graft compatibility.
Conclusion: This genetic engineering approach, repeated without modification for the five HLA-A class I most common alleles known to be associated with HLA-B7 and -C7 in the same haplotype, would cover 83.4% of the world population. Our findings offer a scalable HLA-compatible skin graft model, potentially improving practices in burn units worldwide.
Plain Language Summary: This study addresses a key challenge in treating severe burns: finding suitable skin donors. When someone needs a skin transplant, it is important to match certain proteins on their cells, called HLA proteins, to avoid rejection by the immune system. However, there is a lot of variety in HLA types among people, making it hard to find good matches. Here, we developed a mouse model that mimics human skin transplants. We used mice that carry human HLA proteins, allowing us to study how the immune system reacts to different skin grafts. We also used a gene-editing tool (CRISPR-Cas9) to modify cells and remove some HLA proteins, keeping only the most common one, HLA-A*02:01. The results shows that when mice lack some HLA proteins but retain compatible ones, the skin is not rejected. However when there are differences, the immune system attacks the graft. Importantly, removing certain HLA proteins (like HLA-B and -C), in cell lines, helps avoiding their rejection. This technique could potentially apply to most people worldwide (about 83.4%), offering a new way to make skin transplants more successful and easier to match. This could greatly benefit burn patients by speeding up treatment and reducing the risk of rejection.
Keywords: transplant, allorejection, CRISPR, genetic engineering, histocompatibility
Introduction
Burns are among the most severe forms of trauma. According to the World Health Organization, approximately 11 million people suffer burns worldwide each year, with 180,000 of these cases resulting in death. [https://www.who.int/news-room/fact-sheets/detail/burns]. Although the vast majority of burns occur in low- and middle-income countries, burn injuries are also a significant issue in high-income countries. For example, in the United States, 1.4 million burn injuries occur each year, making it the fourth leading cause of unintentional injury death.1 Extensive burns dramatically reduce survival rates, with the mortality rate reaching 97.8% in patients with burns covering more than 70% of the total body surface area.2 Rapid allogeneic skin transplantation is the only way to avoid certain death in these severe cases. Thus, one of the greatest challenges in transplantation is identifying donors compatible with the patient. This is crucial to avoiding immune rejection, which is primarily mediated by cytotoxic T lymphocytes (CTLs) and natural killer (NK) cells. CTLs recognize allogeneic cells through the major histocompatibility complex (MHC) class I—known as HLA class I in humans—triggering the elimination of “non-self” cells. In contrast, NK cells eliminate cells with insufficient HLA class I expression through “missing-self” recognition. HLA class I molecules can be divided into classical (HLA-A, HLA-B, and HLA-C), which are highly polymorphic and primarily involved in antigen presentation to CD8+ T cells, and non-classical (HLA-E, HLA-F, and HLA-G), which exhibit limited polymorphism and play immunoregulatory roles, particularly in immune tolerance and NK cell modulation. As a result, while HLA mismatch induces CTL-mediated rejection, complete HLA loss enhances NK cell-mediated killing, creating a major barrier to allogeneic graft survival.
For more than a decade, efforts have been made to solve the problem of immune rejection by depleting HLA class I genes to generate hypoimmunogenic cells to serve as universal donors. Initially, HLA expression was depleted by beta-2-microglobulin knock-down3,4 or knock-out5,6 but soon it was realized that complete HLA removal could reduce T cell activity but exacerbate NK cells killing. To reduce NK cell activity in these mutant cells, overexpression of HLA-E, HLA-G or CD47 which interact with NK inhibitory receptors seemed to be partially effective.7–9 However, the cutting edge approach to reduce T cell immunogenicity keeping NK cells at check, it is selective targeting of all but one HLA-Class I proteins expressed in a cell. This strategy allows to present HLA-restricted peptides to a certain extent and reduce NK “missing self” activity through interaction with inhibitory receptors. Therefore, some HLA-edited cells have been designed retaining different HLA genes, on the basis of being the most prevalent of a certain population or being more prompt to interact with KIR receptors.10–12
There are transgenic mouse models which lack expression of mouse MHC and instead express individual HLA class I genes.13–15 HLA-educated T cells are present in these transgenic mice, and have been widely used for identifying T cell epitopes, studying CTLs responses generated by those epitopes in the context of viral infections and tumors and exploring its use as potential T-cell based vaccines.16–18 However, these HLA transgenic mice have never been utilized in studies analyzing allogeneic responses to skin transplants in the absence of mouse histocompatibility complex.
The main goal in rapid management of severe burn patients is to ensure the availability of compatible skin for transplantation when needed. One approach to achieve this, is the generation of skin grafts from cells where several HLA molecules, excluding the most common allele, are deleted to ensure broad compatibility. HLA-A*02:01 (HLA-A2) is the predominant allele in the Caucasian population, making cells retaining this allele potentially accepted by approximately 50% of Caucasians.19 In this study, we introduce the development of a skin graft model using HLA class I transgenic mice and characterize its immune response. Additionally, a strategy for generating human cell clones that retain HLA-A2 expression while lacking HLA-B and HLA-C using a single guide RNA, and subsequently assess their histocompatibility in this murine skin allograft model was proposed. Furthermore, we explore the potential of this strategy in advancing towards a universal HLA-compatible skin graft model.
Material and Methods
Mice
All HLA transgenic mice used13–15 were males between 8 and 12 weeks of age that were bred and housed under specific-pathogen-free conditions in the Instituto de Salud Carlos III (ISCIII) animal facilities (Madrid, Spain).
Cell Culture
JY cell line, kindly provided by Dr. JA López de Castro, is a B cell lymphoblastoid cell line, part of the HLA Typed Collection maintained by the European Collection of Cell Cultures (ECACC). JY cells express HLA-A*02:01, HLA-B*07:02 and HLA-C*07:02 and were cultured in RPMI-1640 supplemented with 10% fetal bovine serum (FBS), 1% Penicillin (10,000 UI)-Streptomycin (10 mg/mL) and 1% L-Glutamine (200mM). For cell cloning, FBS was incremented to 20%. Cells were cultured under standard conditions (37°C, 5% CO2).
Skin Grafts
The donor mice were anesthetized with isoflurane using 4–5% for induction and 2–3% for maintenance with a mouse cone throughout the procedure. Back skin was shaved, disinfected with chlorhexidine and harvested from the hip to the neck using blunt dissection. Donor mice were euthanized by cervical dislocation after harvesting the skin graft. Under a microscope, the connective tissue, fat tissue, and panniculus carnosus was separated from the back skin using fine tenotomy scissors. Approximately 10-mm x 10-mm grafts were cut out and stored on gauzes soaked with sterile phosphate-buffered saline (PBS) in a petri dish on ice. Recipient mice were treated with the anti-inflammatory Metacam (0.2 mg/kg) and the antibiotic Enrofloxacin (5 mg/kg) prior to the start of the procedure. They were then anesthetized, shaved and disinfected as previously described. Using scissors, 10-mm x 10-mm square of skin was cut preserving the panniculus carnosus and vessels, and the graft was positioned on the graft bed, avoiding folds along the edges. Eight sutures were placed at the corners and in the middle of each edge. Recipient mice were wrapped in an adhesive bandage with folded gauze over the graft and anaesthesia was removed. Mice were closely monitored during recovery.
Bone Marrow Dendritic Cell Generation
Mice were euthanized in accordance with ethical and animal welfare guidelines. Long bones, such as the femurs and tibias were extracted and the muscles and soft tissues were removed. The bones were washed with PBS to eliminate any remaining blood and tissue. Next, the ends of the bones were cut and centrifugated. For dendritic cell obtention, bone marrow cells were resuspended in RPMI-1640 medium supplemented with 10% FBS, 0.5% Penicillin (10,000 UI)-Streptomycin (10 mg/mL), 0.1% Kanamycin (10 mg/mL), 1% L-Glutamine (200mM), 2-Mercaptoethanol (5x10−5 M) and 10% of J558L supernatant containing GM-CSF for DC differentiation. Fresh medium was added every 3–4 days to maintain differentiation towards dendritic cells.
Allograft Immune Response Assay
Recipient mouse spleens were disaggregated in 100 μm nylon cell strainers, lysed in ACK buffer for 5 min at room temperature, washed in PBS and cell counted. The antibodies used against mouse intracellular and surface antigens were purchased from BioLegend: anti-CD8 (clone 53–6.7); anti-CD4 (clone GK1.5), anti-NKp46 (clone 29A1.4), anti-IFN-γ (clone XMG 1.2) and anti-TNF-α (clone MP6-XT22).
To assess the intracellular production of IFN-γ and TNF-α, 2×106 recipient splenocytes were challenged for 24h with either 106 DC or 105 JY edited cells, in presence of Brefeldin A (2.5 ug/mL). Following incubation, the cells were fixed with 4% paraformaldehyde (Electron Microscopy Sciences) for 12 min at room temperature in the dark and permeabilized with 1% saponin (Sigma-Aldrich) in 1× PBS and 3% FBS for 20 min at 4°C. The immune response was analyzed by flow cytometry with a FACSCanto flow cytometer (Becton, Dickinson, Franklin Lakes, NJ, USA) using BD FACSDiva software (Becton, Dickinson), and data were analysed with FlowJo (TreeStar Inc., Ashland, OR, USA).
CRISPR/Cas9 HLA Disruption
Selected HLA genes were disrupted by Integrated DNA Technologies (IDT) Alt-R CRISPR-cas9 system (IDT, Coralville, IA, USA). The crRNA 615 (5’-CGACGCCGCGAGTCCGAGAG-3’), named after its position in HLA-B*07:02 gene, was designed to target exon 2, one of the most variable exons in HLA genes, which correspond to α1, subunit comprised in the peptide groove. Ribonucleoprotein (RNP) complexes were delivered to the JY cell line by nucleofection using the Neon transfection system (Thermo Fisher Scientific). For each reaction 2×105 cells were resuspended in 14 uL buffer R with crRNA/tracrRNA (2.68µM) and Cas9 (2.57uM) and electroporated at 1450 V/10 ms/3 pulses. After nucleofection, cells were seeded in 96 well plates. Edited cells were cloned to 1 cell per well in a rounded bottom 96 well plate.
HLA Clones Characterization
For HLA protein characterization, clones were analysed by flow cytometry using supernatant from PA2.1 (anti-HLA-A*02) or ME-1 (anti-HLA-B*07) hybridomas, as well as a goat anti-mouse IgG polyclonal antibody (Thermo Fisher Scientific). Further genomic characterization was performed by PCR, and sequencing. Primers used were: HLA-A*02_Fwd (5’CCTCTGTGGGGAGAAGCAAC3’), HLA-A*02_Rv (5’TCCCGGGGCAAGGGTCTC3’), HLA-B*07:02_Fwd (5’GGAGGAGCGAGGGGACCGCA3’), HLA-B*07:02_Rv (5’GGGTCCCGCGGCCTCAGGGA3’), HLA-C*07:02_Fwd (5’GAGAGGAGCGAGGGGC3’) and HLA-C*07:02_Rv (5’CGGGTTCCGCAGATCCACC3’). For sequencing, PCR products were cloned with TOPO-TA cloning kit (Thermo Fisher, K4575J10) according to manufacturer’s instructions.
Statistical Analysis
All data were plotted and statistically analysed using GraphPad Prism software. Graphs show means ±SEM. Statistical significance was determined using an Two-way ANOVA. P value of <0.01 was considered significant.
Results
HLA-Matched Skin Grafting: Murine Model Validation
The development of a model to study the effect of the absence of certain human MHC molecules on the immune response to transplants, aimed at facilitating HLA matching, would significantly advance the field of skin transplantation. Using humanized HLA mice (Table 1), we established a skin graft model involving the transplantation of small fragments of dorsal skin from an HLA-A*02:01 (HLA-A2) mouse onto the back of HLA-A*02:01/B*07:02 (HLA-A2B7) double transgenic mice. To ensure compatibility and monitor rejection, HLA-A2B7 and HLA-B*27:05 (HLA-B27) skin were also transplanted into HLA-A2B7 mice (Figure 1A).
 |
Table 1 Humanized Transgenic Mouse Models Used in the Study |
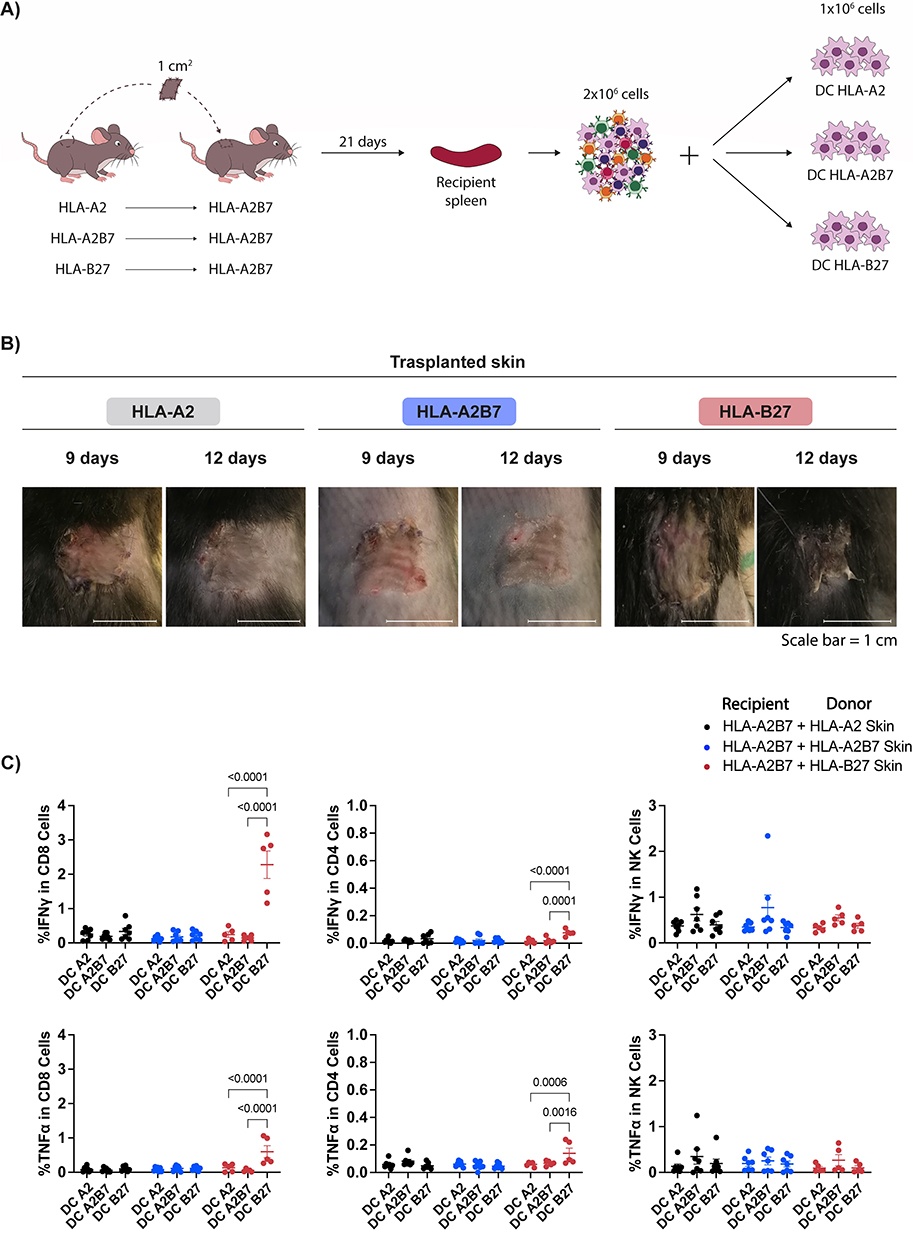 |
Figure 1 HLA-A*02:01 mouse skin does not trigger a CD8 mediated allorejection in HLA-A*02:01/B*07:02 recipient mice. (A) Schematic of skin graft model and strategy to evaluate the immune response to the grafts. (B) Representative images of skin grafts from HLA-A2, HLA-A2B7 and HLA-B27 donor mice in HLA-A2B7 recipient mice 9 and 12 days post-transplantation. (C) Percentage of activated CD8 T cells, CD4 T cells, or NK cells, as measured by IFN-γ and TNF-α production through intracellular flow cytometry of recipient mouse splenocytes exposed to the different DC in the presence of Brefeldin-A for 24 hours. Data represent the mean ± SEM of two experiment with n = 5 to 7 mice per group. Data were analysed by two-way ANOVA test. |
By day 12 post-transplantation, differences between the grafts were visible. HLA-A2 and HLA-A2B7 skin grafts appeared healthy, while HLA-B27 graft showed necrosis (Figure 1B). By day 21, the incompatible HLA-B27 graft was completely rejected. At this point, we assessed the immune response developed against the graft. Recipient spleens were harvested, processed, and challenged with dendritic cells (DC) expressing HLA-A2 (DC A2), HLA-A2B7 (DC A2B7), or HLA-B27 (DC B27) molecules. Activation was analysed by IFN-γ and TNF-α production in CD8 and CD4 T cells and NK cells (Figure 1C). Only CD8 T cells from HLA-A2B7 mice transplanted with HLA-B27 incompatible skin showed significant secretion of both cytokines. This response is exclusively triggered when challenged with DC expressing HLA-B27, but not with HLA-A2 or HLA-A2B7 DC, demonstrating HLA-restricted specificity. A weak CD4 T cell response limited to TNF-α production was also observed. No significant differences in NK response were detected.
These results validate the feasibility of the proposed HLA transgenic mouse model to recapitulate the cutaneous allogenic response and suggest potential acceptance of simplified HLA skin grafts, characterized by a reduced number of HLA molecules, by recipients with broader HLA profiles.
Development and Immune Response Characterization of a Skin Allograft Model Using HLA Class I Transgenic Mice
To further explore immune-mediated allorejection mechanisms, we constructed a converse scenario model. HLA-A2B7 mouse skin was grafted into HLA-A2 mice to promote a specific HLA-B7 immune response. Following the same procedure as in the first experiment, we analysed the immune response by measuring IFN-γ and TNF-α production in various immune cells. Splenocytes from recipient mice were harvested at 3 weeks and 2 months post-transplantation and challenged with DC expressing HLA-A2, HLA-B7, or both HLA class I molecules (Figure 2).
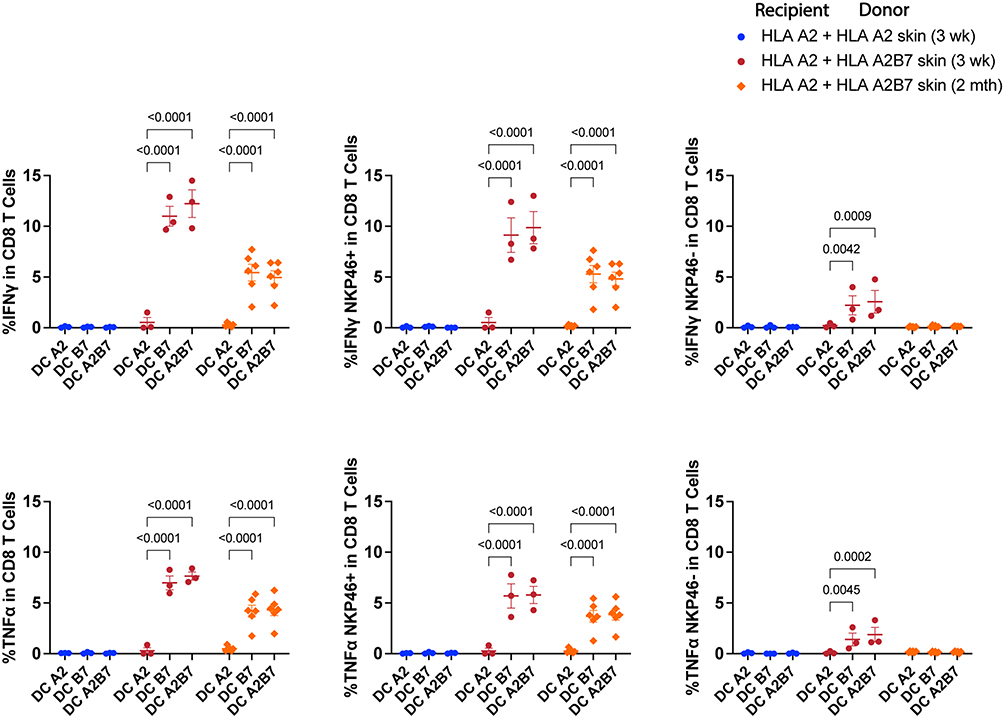 |
Figure 2 HLA-A*02:01/B*07:02 mouse skin induces a CD8 mediated allograft rejection in HLA-A*02:01 recipient mice. Following similar experimental strategy as in Figure 1, skin from HLA-A2 or HLA-A2B7 mice was grafted into HLA-A2 recipient mouse. Three weeks or 2 months post-transplantation, recipient spleens were harvested and co-culture with the different DC in presence of Brefeldin-A for 24 h, followed by immune response evaluation by flow cytometry. The plots show activation of total (left), NKP46+ (middle) and NKP46- (right) CD8 T cells measured by IFN-γ and TNF-α production through intracytoplasmic flow cytometry. Data represent the mean ± SEM of one experiment with n = 3 to 6 mice per group. Data were analysed by two-way ANOVA test. |
Significant IFN-γ and TNF-α secretion was observed in CD8 T cells exposed to DC expressing HLA-B7 or HLA-A2B7, but not HLA-A2 alone (Figure 2, left). The activation of CD8 T cells by B7 and A2B7 DC suggests that T cell activation is specifically triggered by the HLA-B7 molecule. The response was reduced by half at 2 months, indicating an acute response at 3 weeks rather than solely immunological memory. Moreover, it is known that T cells when activated can gain expression of NK receptors such as NKp46.20 In our skin model the CD8 NKp46+ T cells were the major contributors to the allograft response, while CD8 NKp46- T cell activation was minor and only detected at 3 weeks post-engraftment (Figure 2, middle and right).
To confirm the specificity of activated CD8 T cells in HLA-A2 mice grafted with HLA-A2B7 skin, cytokine levels were measured in the presence of specific antibodies against HLA class I molecules during co-culture (Figure 3). Inhibition of IFN-γ and TNF-α activation was observed in the presence of the specific antibody against HLA-B7 (ME1) but not with HLA-A2 specific antibody (PA2.1). These results evidence that allorecognition is mediated by the HLA-B*0702 molecule.
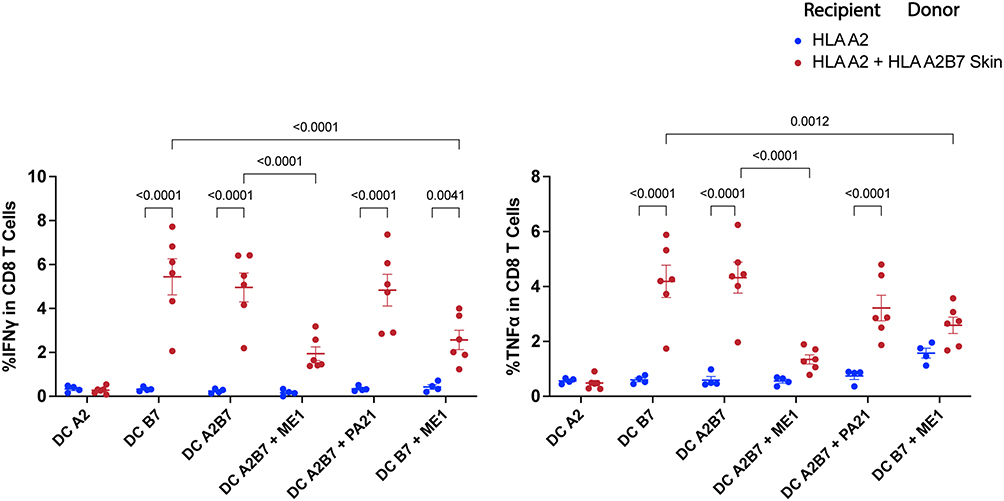 |
Figure 3 CD8 T cell mediated HLA-A2B7 skin allograft rejection in HLA-A*02:01 recipient mice is HLA-B*07:02 specific. Two months post-transplantation recipient splenocytes were challenged with DC in presence of HLA monoclonal antibodies: anti-HLA-A*02:01 (PA2.1) and anti-HLA-B*07:02 (ME1) to evaluate immune response specificity. IFN-γ and TNF-α producing CD8 T cells were analysed by intracellular flow cytometry. Data represent the mean ± SEM of one experiment with n = 4 to 6 mice per group. Data were analysed by two-way ANOVA test. |
Therefore, we established a mouse model to investigate the adaptive alloimmune response mediated by CD8 T cells and specific HLA class I molecules, which play a pivotal role in allograft rejection.
Generation of Human Cells Deficient in HLA-B and -C Genes
To address the complex issue of human transplant rejection, we employed genetic engineering to selectively remove all but one HLA class I molecule from a human cell line. Our initial approach involved retaining HLA-A2, as it represents the most prevalent allele in the human population, in JY cell line which expresses HLA-A2, HLA-B*07:02 (HLA-B7), and HLA-C*07:02 (HLA-C7) class I molecules in a homozygous manner. We designed a one-step strategy, targeting the simultaneous deletion of both HLA-B7 and HLA-C7 class I molecules. The requirements for finding the appropriate guide were to limit the search to exons 2 and 3, which are the most variable regions coding for the antigenic peptide groove. We selected the 615 guide (g615), which sequence upstream of the PAM was conserved in HLA-B7 and -C7 genes, but not in HLA-A2 (Figure 4A).
 |
Figure 4 HLA disruption of JY cell line for the generation of immune-compatible cells. (A) Sequence of the crRNA used for the CRISPR/Cas9 deletion strategy and correspondent sequences in the interest HLA genes and expected HLA disruption scheme. (B) Flow cytometry analysis of HLA-A*02:01 and HLA-B*07:02 expression in JY edited CRISPR pool and selected clones. (C) Schematic representation of genetic characterization of selected HLA-A2ΔBC and unedited clones. |
Flow cytometry analysis showed 60% depletion of HLA-B7 in the edited cell pool (HLA-A2ΔBC), while HLA-A2 expression remained unchanged (Figure 4B, left). After single-cell cloning, we successfully isolated 10 out of 14 clones expressing HLA-A2 but lacking HLA-B7 protein (Figure 4B, right). We further characterized these clones through a genomic analysis by PCR and sequencing of the HLA-B7 and -C7 genes. Since we are using a guide that induces two cuts, one in HLA-B7 and one in HLA-C7 gene, in our genomic analysis we observe two main outcomes: clones that either locally delete a small number of bases in each molecule (clone 13) or that fuse B7 and C7 genes (clone 15) (Figure 4C). In both types of KO clones, the detected modifications either shift the open reading frame by introducing a premature stop codon or delete a significant number of bases, leading to protein truncation.
Assessing Histocompatibility of HLA-A2ΔBC Clones in the Mouse Skin Allograft Model
We evaluated the role of HLA-B and -C double knock-out cells in a hybrid mouse-human allograft model by co-culturing splenocytes from skin-transplanted mice with either unedited or HLA-A2ΔBC human clones (Figure 5A). A specific HLA-B7 immune response with IFN-γ and TNF-α secretion in CD8 T cells was detected with unedited HLA-A2B7 clones. In contrast, no specific activation of CD8 or CD4 T cells was observed with HLA-A2ΔBC KO clones (Figure 5B, left). As expected, no CD4 T cell activation occurred with unedited HLA-A2B7 cells (Figure 5B, middle). Increased unspecific TNF-α production by NK effector cells was observed due to the xenogeneic nature of the co-cultures, regardless of the number of HLA alleles expressed by human target cells (Figure 5B, right).
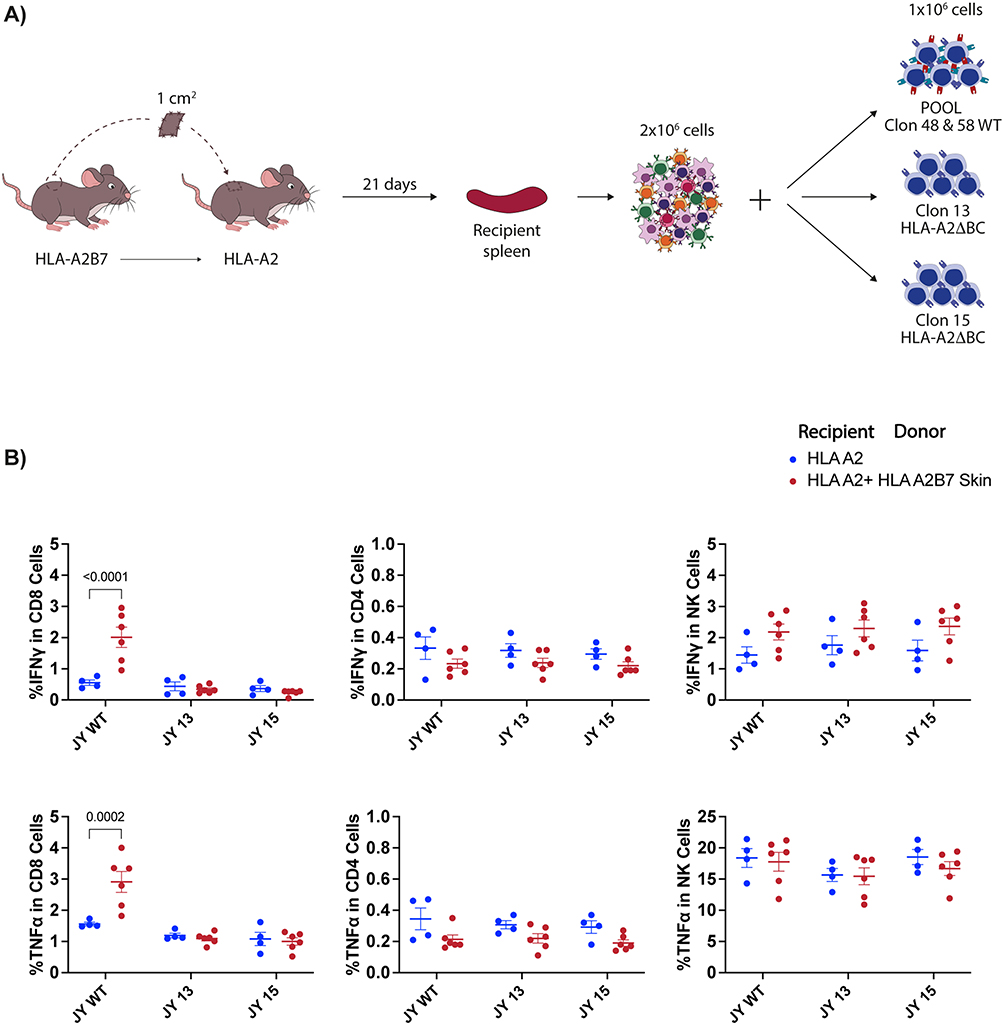 |
Figure 5 JY HLA-A2ΔBC clones escape HLA-B*07:02 specific CD8 T cell response against skin allografts. (A) Schematic of skin graft model and strategy to evaluate the immune response against edited and unedited JY clones. (B) Analysis of IFN-γ and TNF-α producing CD8 T cells, CD4 T cells and NK cells from recipient mouse splenocytes when exposed to JY cell clones in presence of Brefeldin-A for 24h. Data represent the mean ± SEM of one experiment with n = 4 to 6 mice per group. Data were analysed by two-way ANOVA test. |
These findings demonstrate that HLA-A2ΔBC clones can evade recognition by CD8 T cells, suggesting their potential suitability for transplantation.
Towards a Universal HLA-Compatible Skin Graft Model
In this study, we showed that human HLA-A2 expressing cells lacking HLA-B and -C genes did not elicit an allograft response in a humanized skin allograft model. Given HLA-A2’s prevalence in 39.1% of the human population, generating HLA-A2ΔBC skin and storing it in skin banks could be a fast and effective medical technique to treat a significant, but minor, percentage of patients with major burns who cannot wait for an bioengineered autograft. Extending this strategy to other common HLA-A alleles could benefit a majority of the human population.
According to world population coverage data from the Immune Epitope Database (IEDB), using the five most abundant HLA-A alleles (A*01:01, A*02:01, A*03:01, A*11:01, and A*24:02) could potentially provide coverage for 83.4% of the human population. B7 and C7 alleles are frequently associated with these HLA-A alleles on the same haplotype, particularly in the Caucasoid population (Table 2). This approach could be repeated without modification for the five HLA-A class I alleles mentioned.
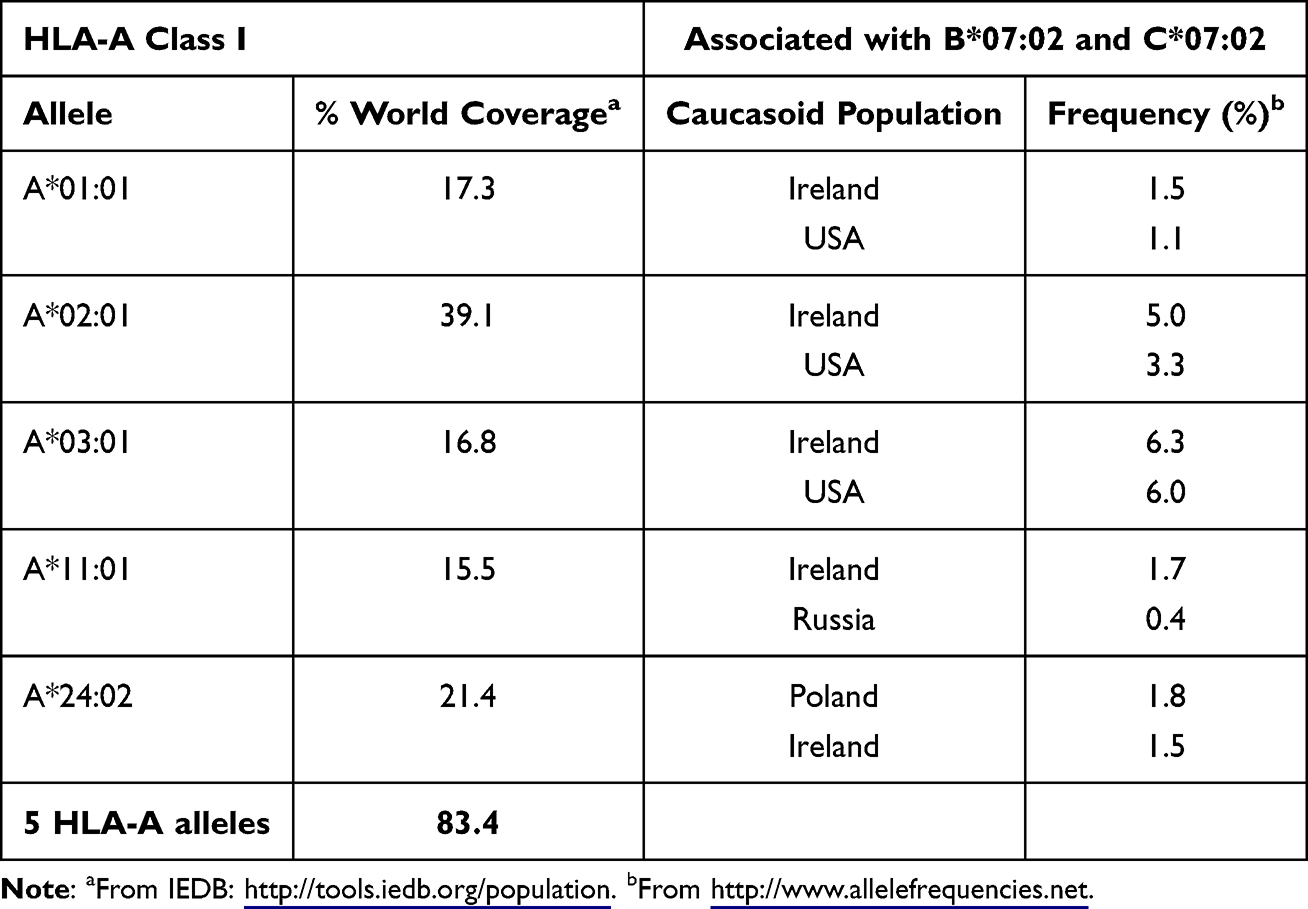 |
Table 2 The Five Most Common HLA-A Alleles Cover the Majority of the World Population and are Associated With HLA-B*07:02 and HLA-C*07:02 in the Caucasoid Population. HLA-A most common alleles world frequency obtained from IEDB Database and HLA-AX/B7/C7 haplotype frequencies in the caucasoid population retrieved from the Allele Frequency Net Database |
Discussion
The primary challenge in treating burn patients is achieving HLA compatibility for available skin grafts, a task complicated by the extensive variation in HLA class I molecules. In this study, we developed a humanized HLA-transgenic skin graft mouse model that faithfully mimics allogenic immune responses. This model enables accurate examination of immunogenicity and graft rejection in the presence and absence of HLA class I molecules. Additionally, we established a one-step strategy for generating human HLA-A2 expressing cell lines devoid of HLA-B and -C genes. These engineered cells exhibit excellent immune tolerance, as demonstrated by their successful integration into our hybrid mouse-human allograft model without eliciting signs of rejection. This strategy, applied to the five most frequent HLA-A alleles, advances us towards a universal HLA-compatible skin graft model.
The experimental subjects in this study were HLA humanized mice that lack murine MHC class I molecules but are transgenic for human MHC class I. Previous studies have shown that these models are versatile preclinical models for tumour processes and cardiovascular diseases, such as atherosclerosis,21 HLA-restricted T cell epitope identification,18,22–24 study of human autoimmune diseases associated with specific HLA alleles,25 and the evaluation of human T-cell-based vaccines.16,17 Our study proposes a new use of HLA class I transgenic mice as a model to examine the immune response in HLA-restricted grafts, specifically skin transplantation. This approach allows for a comprehensive analysis of immune populations impacting allografts. We observed a predominant CD8 T cell-driven allogeneic response, consistent with allogenic responses in humans.26 Previous findings from an HLA transgenic model by Borenstein et al suggest that CD8 T cells may not be indispensable for allograft rejection.27 However, this discrepancy could be attributed to the presence in their model of mouse MHC molecules. Our experimental approach accurately reproduces skin graft rejection when HLA class I-unmatched skin is transplanted, resulting in total rejection at three weeks post-implantation. Additionally, our methodology includes re-stimulating recipient splenocytes with DC expressing unmatched HLA molecules, allowing detailed evaluation of the immune response, including cytokine production, by CD4 and CD8 T lymphocytes. And, the use of HLA-specific antibodies which indicated that T cell activation in our model is likely through direct DC presentation, rather than an indirect mechanism as proposed in several other models.28,29
Moreover, our HLA transgenic model demonstrated that skin grafts from donors expressing only a single HLA molecule could be transplanted into recipients expressing that HLA molecule, among others, without causing rejection. This approach can be extended to as many models as types of HLA transgenic mice are available.
The experimental HLA disruption strategy developed here can be extrapolated to manufacture skin equivalents expressing only one HLA class I allele, a plausible clinical strategy to avoid allogenic response. Such equivalents may be well-tolerated by patients sharing that specific HLA class I molecule, regardless of other HLA-A, -B, and -C alleles expressed in their cells. To achieve this goal, the appropriate strategy involves deleting all genes encoding HLA class I molecules, except for one. Xu et al pioneered this approach by targeting HLA-A and -B genes, while retaining HLA-C in induced pluripotent stem cells using specific crRNA for each targeted gene. However, HLA-C’s protective capacity is limited due to its lower surface expression and less ligand diversity compared to HLA-A and -B molecules,30 which is crucial for burn patients highly susceptible to infections. Therefore, preserving HLA-A class I molecules may offer a broader range of protection and influence survival.
Two recent studies focused on selective deletion of HLA-B and -C genes while preserving the most common HLA-A alleles in specific populations as is the HLA-A*11:01 allele in the Chinese population,12 or the HLA-A2 allele in the world population.11 Our approach differs by using a single CRISPR-Cas9 guide to target both HLA-B7 and -C7 genes, and by developing models for immunogenicity testing using humanized transgenic mice expressing HLA-A2, HLA-B7, and HLA-A2B7.
We outlined a one-step approach using crRNA 615, which enables to generate skin tissue characterized by depleted HLA-B7 and -C7 molecules, and the presence of a single prevalent HLA-A class I allele (A*01:01, A*02:01, A*03:01, A*11:01, and A*24:02), known to be associated with HLA-B7 and -C7 in the same haplotype. These skins could be preserved in skin banks, addressing the demand for skin grafts across a significant proportion of the global population. Modifying only five cell types could achieve 83.4% world coverage. Given the association of the partial haplotype B7/C7 with other multiple HLA-A class I molecules, our experimental paradigm could augment the diversity of samples in skin banks, increasing coverage of the human population.
Conclusions
In this study we have established a genetic engineering strategy to deplete HLA-B07:02 and HLA-C07:02 genes while preserving the most common HLA-A allele, HLA-A*02:01, and a skin graft model using HLA transgenic mice that allows precise study of the immune response to grafts and demonstrates the histocompatibility of the edited cells. The extension of this strategy to any cell type and to the five most common HLA-A molecules represents a significant advance in the field of skin substitutes and burn treatment, potentially providing ready-to-use skins compatible with 83.4% of the world population. This work advances our understanding of immune responses in skin grafts and may pave the way for more effective treatments for burn patients.
Abbreviations
DC, dendritic cells; FBS, fetal bovine serum; MHC, major histocompatibility complex; PBS, phosphate-buffered saline; RNP, ribonucleoprotein.
Data Sharing Statement
Source data are provided with this paper. The raw data generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Ethics Approval
All experiments involving mice were done with the approval and in accordance with the Instituto de Salud Carlos III (ISCIII) and Comunidad de Madrid Ethics Committees (PROEX 355.1/21) and followed current Spanish legislation (Real Decreto 53/2013), which is in compliance with European laws (Directive 2010/63/EU). The use of JY cell line in this study was reviewed by the Instituto de Salud Carlos III Ethics Committee and deemed exempt from specific ethical approval.
Author Contributions
All authors contributed significantly to the conception and design, data acquisition, or data analysis and interpretation, participated in the drafting of the article or critically revising it for important intellectual content, agreed to submit to the current journal, gave final approval for the version to be published, and agreed to be accountable for all aspects of the work.
Funding
This work was supported by grants from the Comunidad de Madrid (Programa de I+D en tecnologías; S2018/BAA-4480), Instituto de Salud Carlos III (ISCIII-AESI; PI19CIII/0033) and the Agencia Estatal de Investigación (AEI; PID2021-125757OB-100). LC-F holds a FPU fellowship (FPU20/03864) from Ministerio de Ciencia, Innovación y Universidades (Spain).
Disclosure
The authors state no competing interests in this work.
References
1. Kraft R, Herndon DN, Al-Mousawi AM, Williams FN, Finnerty CC, Jeschke MG. Burn size and survival probability in paediatric patients in modern burn care: a prospective observational cohort study. Lancet. 2012;379(9820):1013–1021. doi:10.1016/S0140-6736(11)61345-7
2. Iqbal T, Saaiq M, Ali Z. Epidemiology and outcome of burns: early experience at the country’s first national burns Centre. Burns. 2013;39(2):358–362. doi:10.1016/J.BURNS.2012.07.011
3. Börger AK, Eicke D, Wolf C, et al. Generation of hla-universal ipsc-derived megakaryocytes and platelets for survival under refractoriness conditions. mol Med. 2016;22(1):274–285. doi:10.2119/MOLMED.2015.00235/FIGURES/8
4. Figueiredo C, Seltsam A, Blasczyk R. Class-, gene-, and group-specific HLA silencing by lentiviral shRNA delivery. J mol Med. 2006;84(5):425–437. doi:10.1007/S00109-005-0024-2/FIGURES/8
5. Lu P, Chen J, He L, et al. Generating hypoimmunogenic human embryonic stem cells by the disruption of beta 2-microglobulin. Stem Cell Rev Rep. 2013;9(6):806–813. doi:10.1007/S12015-013-9457-0/FIGURES/4
6. Mattapally S, Pawlik KM, Fast VG, et al. Human leukocyte antigen class I and II knockout human induced pluripotent stem cell–derived cells: universal donor for cell therapy. J Am Heart Assoc. 2018;7(23). doi:10.1161/JAHA.118.010239
7. Han X, Wang M, Duan S, et al. Generation of hypoimmunogenic human pluripotent stem cells. Proc Natl Acad Sci U S A. 2019;116(21):10441–10446. doi:10.1073/PNAS.1902566116/SUPPL_FILE/PNAS.1902566116.SD01.XLSX
8. Gornalusse GG, Hirata RK, Funk SE, et al. HLA-E-expressing pluripotent stem cells escape allogeneic responses and lysis by NK cells. Nature Biotechnol. 2017;35(8):765–772. doi:10.1038/nbt.3860
9. Deuse T, Hu X, Gravina A, et al. Hypoimmunogenic derivatives of induced pluripotent stem cells evade immune rejection in fully immunocompetent allogeneic recipients. Nature Biotechnol. 2019;37(3):252–258. doi:10.1038/s41587-019-0016-3
10. Xu H, Wang B, Ono M, et al. Targeted disruption of HLA genes via CRISPR-Cas9 generates iPSCs with enhanced immune compatibility. Cell Stem Cell. 2019;24(4):566–578.e7. doi:10.1016/j.stem.2019.02.005
11. Parent AV, Faleo G, Chavez J, et al. Selective deletion of human leukocyte antigens protects stem cell-derived islets from immune rejection. Cell Rep. 2021;36(7):109538. doi:10.1016/j.celrep.2021.109538
12. Ji TT, Niu SS, Fang MH, et al. Genome editing HLA alleles for a pilot immunocompatible hESC line in a Chinese hESC bank for cell therapies. Cell Prolif. 2023;56(5):e13471. doi:10.1111/CPR.13471
13. Firat H, Garcia-Pons F, Tourdot S, et al. H-2 class I knockout, HLA-A2.1-transgenic mice: a versatile animal model for preclinical evaluation of antitumor immunotherapeutic strategies. Eur J Immunol. 1999;29(10):3112–3121. doi:10.1002/(sici)1521-4141(199910)29:10<3112::aid-immu3112>3.0.co;2-q
14. Rohrlich PS, Cardinaud S, Firat H, et al. HLA-B*0702 transgenic, H-2KbDb double-knockout mice: phenotypical and functional characterization in response to influenza virus. Int Immunol. 2003;15(6):765–772. doi:10.1093/INTIMM/DXG073
15. Khare SD, Luthra HS, David CS. Spontaneous inflammatory arthritis in HLA-B27 transgenic mice lacking beta 2-microglobulin: a model of human spondyloarthropathies. J Exp Med. 1995;182(4):1153–1158. doi:10.1084/JEM.182.4.1153
16. Akazawa Y, Saito Y, Yoshikawa T, et al. Efficacy of immunotherapy targeting the neoantigen derived from epidermal growth factor receptor T790M/C797S mutation in non-small cell lung cancer. Cancer Sci. 2020;111(8):2736–2746. doi:10.1111/CAS.14451
17. Chiuppesi F, d’Alincourt SM, Contreras H, et al. Development of a multi-antigenic SARS-CoV-2 vaccine candidate using a synthetic poxvirus platform. Nat Commun. 2020;11(1):1–16. doi:10.1038/s41467-020-19819-1
18. Takagi A, Matsui M. Identification of HLA-A*02:01-restricted candidate epitopes derived from the non-structural polyprotein 1a of SARS-CoV-2 that may be natural targets of CD8+ T cell recognition in vivo. J Virol. 2021;95(5). doi:10.1128/JVI.01837-20
19. Gonzalez-Galarza FF, Christmas S, Middleton D, Jones AR. Allele frequency net: a database and online repository for immune gene frequencies in worldwide populations. Nucleic Acids Res. 2011;39(suppl_1):D913–D919. doi:10.1093/NAR/GKQ1128
20. Hudspeth K, Silva-Santos B, Mavilio D. Natural cytotoxicity receptors: broader expression patterns and functions in innate and adaptive immune cells. Front Immunol. 2013;4(MAR). doi:10.3389/FIMMU.2013.00069
21. Schaftenaar FH, Amersfoort J, Douna H, et al. Induction of HLA-A2 restricted CD8 T cell responses against ApoB100 peptides does not affect atherosclerosis in a humanized mouse model. Sci Rep. 2019;9(1). doi:10.1038/S41598-019-53642-Z
22. Cardinaud S, Moris A, Février M, et al. Identification of cryptic MHC I-restricted epitopes encoded by HIV-1 alternative reading frames. J Exp Med. 2004;199(8):1053–1063. doi:10.1084/JEM.20031869
23. Blanc E, Holtgrewe M, Dhamodaran A, et al. Identification and ranking of recurrent neo-epitopes in cancer. BMC Med Genomics. 2019;12(1):1–14. doi:10.1186/S12920-019-0611-7/FIGURES/4
24. Scheikl-Gatard T, Tosch C, Lemonnier F, Rooke R. Identification of new MUC1 epitopes using HLA-transgenic animals: implication for immunomonitoring. J Transl Med. 2017;15(1):154. doi:10.1186/s12967-017-1254-0
25. Taneja V, David CS. Lessons from animal models for human autoimmune diseases. Nat Immunol. 2001;2(9):781–784. doi:10.1038/ni0901-781
26. Benichou G, Gonzalez B, Marino J, Ayasoufi K, Valujskikh A. Role of memory T cells in allograft rejection and tolerance. Front Immunol. 2017;8:170. doi:10.3389/FIMMU.2017.00170
27. Borenstein SH, Graham J, Zhang XL, Chamberlain JW. CD8+ T cells are necessary for recognition of allelic, but not locus-mismatched or xeno-, HLA class I transplantation antigens. J Immunol. 2000;165(5):2341–2353. doi:10.4049/JIMMUNOL.165.5.2341
28. Benichou G, Yamada Y, Yun SH, Lin C, Fray M, Tocco G. Immune recognition and rejection of allogeneic skin grafts. Immunotherapy. 2011;3(6):757–770. doi:10.2217/IMT.11.2
29. Jurcevic S, Chandler P, Sacks SH, Simpson E. Rapid rejection of HLA-A2 transgenic skin graft due to indirect allorecognition. Transplantation. 2001;72(6):994–997. doi:10.1097/00007890-200109270-00003
30. Crux NB, Elahi S. Human Leukocyte Antigen (HLA) and immune regulation: how do classical and non-classical HLA alleles modulate immune response to human immunodeficiency virus and hepatitis C virus infections? Front Immunol. 2017;8(JUL):281746. doi:10.3389/FIMMU.2017.00832/BIBTEX
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.