Back to Journals » Journal of Hepatocellular Carcinoma » Volume 13
ScRNA-Seq Deciphers an Autocrine EFNA1-EPHA1 Loop That Reprograms the Microenvironment in Hepatocellular Carcinoma
Authors Chen Y, Tang Y, Ning Y, Yang Y, Tian R, Mao Y, Feng Z ![]() , Lin W
, Lin W ![]() , Wang D, Feng X
, Wang D, Feng X
Received 8 November 2025
Accepted for publication 24 February 2026
Published 10 March 2026 Volume 2026:13 577864
DOI https://doi.org/10.2147/JHC.S577864
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr David Gerber
Yuanhong Chen,1,2,* Yulian Tang,3,* Yufan Ning,4 Yang Yang,4 Rensheng Tian,4 Yongjiao Mao,4 Zhiquan Feng,4 Wenxian Lin,5 Decai Wang,6 Xueping Feng1
1Department of Pathogenic Biology and Immunology, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China; 2Key Laboratory of Basic Research and Transformation of Tumor Immunity and Infectious Diseases, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China; 3School of Medical Technology and Artificial Intelligence, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China; 4School of Basic Medical Sciences, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China; 5Institute of Cardiovascular Sciences, Guangxi Academy of Medical Sciences & the People’s Hospital of Guangxi Zhuang Autonomous Region, Nanning, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China; 6Library, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xueping Feng, Department of Pathogenic Biology and Immunology, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China, Email [email protected] Decai Wang, Library, Youjiang Medical University for Nationalities, Baise, Guangxi Zhuang Autonomous Region, 533000, People’s Republic of China, Email [email protected]
Background: Hepatocytes demonstrate significant heterogeneity between normal liver tissue and hepatocellular carcinoma (HCC), with malignant hepatocytes playing a crucial role in remodeling the tumor microenvironment through specific ligand-receptor interactions. However, the mechanisms by which hepatocytes drive HCC progression at the single-cell level remain poorly understood.
Methods: We analyzed single-cell RNA sequencing datasets (GSE174748 and GSE166635) from the GEO database using CellChat to decode intercellular communication networks, which specifically revealed enhanced EPHA signaling in HCC hepatocytes and identified EFNA1-EPHA1 as the most prominent ligand-receptor pair. This key finding was validated through immunofluorescence analysis in both clinical HCC tissues and HepG2/LO2 cell lines, and its clinical relevance was assessed using the TCGA-LIHC dataset via UALCAN.
Results: Single-cell analysis revealed that HCC hepatocytes act as both senders and receivers of pro-tumorigenic signals, with upregulated expression of malignancy-related genes (AFP, ACSL4, and SERPINA1). CellChat inference demonstrated significantly strengthened outgoing interaction signals from hepatocytes in the HCC microenvironment. The EFNA1-EPHA1 axis was identified as a key mediator of hepatocyte-microenvironment crosstalk, showing marked activation in HCC tissues and high co-expression in HepG2 cells. TCGA analysis confirmed EFNA1 upregulation in HCC, correlating with advanced clinical stage, higher tumor grade, and metastatic events.
Conclusion: Our study provides single-cell resolution evidence that malignant hepatocytes promote HCC progression through autocrine and paracrine signaling via the EFNA1-EPHA1 axis, reshaping the tumor microenvironment. These findings delineate a key autocrine-paracrine mechanism in HCC progression and nominate the EFNA1-EPHA1 axis as a promising candidate for therapeutic development.
Keywords: heterogeneity, hepatocyte, HCC, EFNA1, EPHA1, scRNA-seq, TME
Introduction
Hepatocellular carcinoma (HCC), the sixth most common malignancy and the third leading cause of cancer-related deaths globally,1,2 is a highly aggressive disease whose progression is critically driven by its complex tumor microenvironment (TME).1,2 Despite advances in treatment, including molecular-targeted agents such as tyrosine kinase inhibitors (TKIs), immune checkpoint inhibitors (ICIs), and combination regimens (eg, atezolizumab plus bevacizumab), which have become standard therapies for advanced HCC,3,4 drug resistance remains a major clinical challenge. This resistance is closely linked to the high heterogeneity and dynamic remodeling of the TME.5,6 Hepatocytes, the primary functional cells of the liver, exhibit significant phenotypic and functional alterations during HCC development, contributing to tumor initiation, immune evasion, and metastasis.7 However, the precise mechanism by which cancer-associated hepatocytes influence HCC progression remains unclear.
Notably, these studies have identified dysregulated intercellular signaling pathways that may drive HCC progression, including the ephrin/Eph receptor (Eph/EFN), which is a critical mediator of cell-cell communication in development and cancer.8–10 Among the numerous interactions within this axis, EFNA1–EPHA1 stands out. Bioinformatic analyses of HCC cohorts have consistently identified EFNA1 as one of the most significantly upregulated ephrin family members, and its high expression is strongly correlated with advanced tumor stage and poor patient prognosis.8 This establishes EFNA1 as a clinically relevant target within the HCC landscape itself. Notably, the functional significance of EphA1, a primary receptor for EFNA1, has been validated in HCC, where a specific lncRNA-miRNA axis regulates its expression to promote tumor progression.11 However, the precise mechanistic role of the EFNA1–EPHA1 interaction—specifically how it reprograms the tumor microenvironment to fuel invasion, immune evasion, or metastasis—remains a critical knowledge gap, which is driven by the frequent dysregulation of EFNA1 in tumors.12,13 This gap is especially striking despite its documented oncogenic roles in other cancers;14–16 the functional significance and clinical relevance of EFNA1–EPHA1 signaling in driving HCC progression remain underexplored.
In this study, we integrated scRNA-seq (single-cell RNA sequencing) analysis of HCC datasets17,18 (GSE174748, GSE166635) with functional validation to investigate the role of hepatocyte-derived EFNA1–EPHA1 signaling in tumor progression. Our comprehensive analysis revealed the EFNA1–EPHA1 axis as a critical driver of HCC, a finding validated in clinical cohorts, which offers new mechanistic insights and nominates it as a potential therapeutic target.
Materials and Methods
Bioinformatic Analysis of scRNA-Seq Data
Single-cell RNA sequencing (scRNA-seq) data were acquired from NCBI GEO under accession numbers GSE166635 (HCC cohort, n=2) and GSE174748 (non-tumor controls, n=2). Data processing and analysis were performed using the Seurat package (v5.1.0). Low-quality cells (expressing <200 genes) and genes detected in fewer than three cells were filtered out, followed by normalization and variance stabilization using the SCTransform to correct for technical variability. Highly variable features (n = 3000) were selected for downstream analysis, and the cells were clustered at a resolution of 0.6. Cell type-specific marker genes were identified using thresholds of |log2FC| > 0.25 and a minimum expression fraction (min.pct) of 0.25. Cell-cell communication networks were systematically inferred using CellChat (v1.5.0), a computational framework integrating ligand-receptor interaction databases and pattern recognition algorithms. To ensure robustness, only genes expressed in ≥10 cells (min. cells =10), a threshold informed by the high prevalence of our key signaling molecules, were included, with interactions further filtered by empirical P-values < 0.05, and significance thresholds (|log2FC| > 0.25) to identify biologically relevant signaling pathways.
Biopsies Of Liver Cancer
Treatment-naïve HCC specimens were collected during surgical resection and processed for histopathological analysis with ethical approval from the Youjiang Medical University for Nationalities (#2023082802). All participants provided written informed consent prior to enrollment in the study, in accordance with the ethical principles of the Declaration of Helsinki. Following diagnostic evaluation, residual tissues were embedded in OCT compound, snap-frozen in liquid nitrogen, and stored at −80°C until sectioning. Serial cryosections were obtained at 8-μm thickness for H&E (Solarbio, Beijing, China, Cat# G1120) and Masson’s trichrome staining (Solarbio, Beijing, China, Cat# G1340) to assess tissue morphology and fibrosis, and consecutive sections of 12-μm and 20-μm thicknesses were prepared for multiplex immunofluorescence studies to enable three-dimensional protein localization mapping and intercellular interaction analysis (Figure 1A).
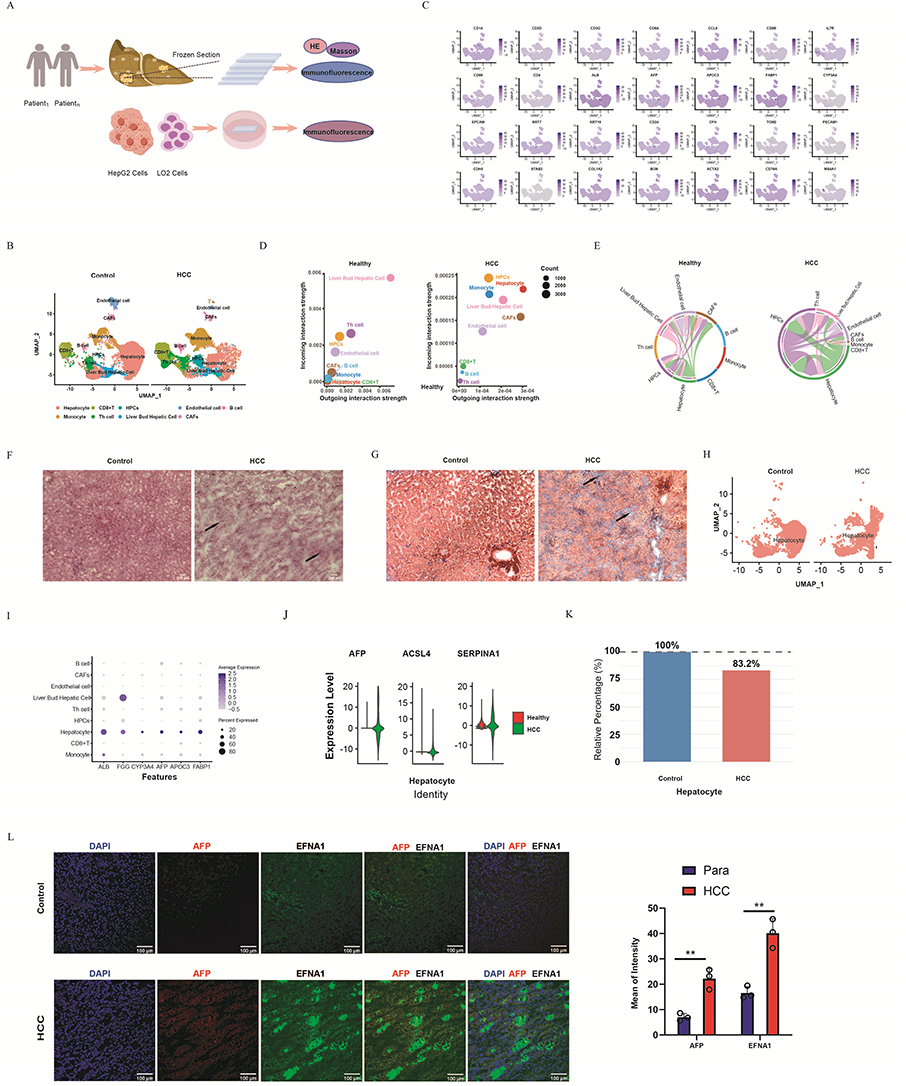 |
Figure 1 Distinct hepatocyte alterations in HCC by single-cell analysis. (A) Experimental Flowchart: Clinical Samples and Cell Cultures. (B) UMAP plots revealed nine cell types in healthy livers and HCC biopsy tissues. Hepatocytes were the cell types displayed, with a decrease in hepatocytes in HCC. (C) Cell populations were annotated using marker gene expression visualized in FeaturePlots. (D) Dot plots showed inferred interaction strengths (outgoing/incoming) between infiltrating immune cells differed between the control and HCC groups, with hepatocytes showing enhanced interactions. (E) Chord diagram comparison of EPHA-mediated hepatocyte-microenvironment crosstalk between Healthy and HCC. The width of the chords corresponds to the communication probability. (F) Histological comparison of adjacent normal liver tissue and hepatocellular carcinoma (HCC) (H&E staining). The black arrow highlights the characteristic disorganized architecture and high cellular density of the malignant region. (G) Masson’s trichrome staining of a serial section from the tissues shown in (F), visualizing collagen deposition (blue). The black arrow identifies a corresponding area of prominent intratumoral fibrosis within the HCC. (H) Hepatocytes were extracted for comparison using UMAP. (I) Dotplot displayed the expression levels of hepatocyte-specific markers (ALB,AFP, APOC3, FABP1, and CYP3A4) in hepatocyte populations. (J) Violin plots revealed upregulated expression of malignancy-associated genes (AFP, ACSL4, and SERPINA1) in HCC compared to normal liver tissue. (K) Stacked bar graph demonstrating a decreased proportion of hepatocytes in HCC, reflecting tumor-driven parenchymal loss. (L) Tumors showed concurrent upregulation of AFP/EFNA1 genes and their protein co-localization (**P<0.01). |
Cell Culture and Assays
The hepatocellular carcinoma cell line HepG2 and normal hepatocyte line LO2 were acquired from the Cell Resource Center of the Chinese Academy of Sciences (Shanghai, China). Cells were propagated in Dulbecco’s modified Eagle’s medium (DMEM; Gibco, USA) supplemented with 10% heat-inactivated fetal bovine serum and 1% penicillin-streptomycin (100 U/mL penicillin and 100 μg/mL streptomycin). All the cell cultures were maintained at 37°C in a humidified atmosphere containing 5% CO2. For the experimental procedures, only cells in the active proliferation phase (logarithmic growth) were used to ensure consistency.
Immunofluorescence Staining and Colocalization
Following the preparation of 12–20μm frozen sections and cell cultures grown to 60–80% confluency on 12–15 mm coverslips, samples were 15-minute air-drying at RT, fixed in −20°C methanol (5–10 min), and washed twice with PBS. The sections were blocked in PBS containing 0.5% BSA and Fcγ blocker at 4°C for one hour, followed by overnight incubation with primary antibodies at the same temperature. After three washes with PBS, samples were treated with fluorophore-conjugated secondary antibodies for one hour at room temperature. Following two additional PBS washes, the sections were finally sealed with coverslips using Fluoromount-G mounting medium (SouthernBiotech, #0100-01). Fluorescence imaging was conducted using a Leica DMI3000B inverted microscope. The primary antibodies for immunofluorescence assays were goat-anti-human/mouse EPHA1 (Invitrogen,#PA5-47837), rabbit-anti-human/mouse EFNA1 (Invitrogen, #34-3300), and mouse anti-human AFP (Invitrogen, #14- 9760-82).
EFNA1 and EPHA1 in HCC Prognosis
Survival analysis revealed that elevated EFNA1 expression was significantly associated with poorer overall survival, whereas EPHA1 showed no significant prognostic correlation. Analysis via UALCAN using TCGA data further illustrated the expression patterns of both genes across clinical stages, tumor grades, and lymph node metastasis status, with the results presented in the respective figures (*P<0.05, **P<0.01, ***P<0.001).
Statistical Analyses
All statistical analyses were performed using GraphPad Prism 12.0. Paired t-tests were applied for comparisons between tumor and matched adjacent non-tumor tissues from the same patients, whereas two-sample t-tests were used for comparisons between HepG2 and control cell groups. Immunofluorescence intensities of EFNA1 and EPHA1 were quantified and normalized to the reference standards. Statistical significance was defined as P<0.05 (two-tailed), with data presented as the mean ± SD from three biological replicates.
Results
A Single-Cell Atlas of the Hepatocellular Carcinoma and Human Liver
The scRNA-seq datasets, GSE174748 (53,986 cells) and GSE166635 (24,832 cells), were processed using Seurat (v4.0) in R (v4.1.0). Initial quality control excluded cells with either <200 detected genes or >10% mitochondrial gene expression (Supplementary Figure 1A and B), with additional outlier removal performed using FeatureScatter. Data normalization was performed using the SCTransform wrapper. Subsequent integration of normal liver and hepatocellular carcinoma (HCC) datasets was achieved and then integrated using RPCA (dims=1:30) for comparison of normal liver and HCC. UMAP clusters (resolution=0.6; Figure 1B) were definitively annotated through a multi-source consensus approach. Cluster marker genes were first identified via differential expression analysis (FindMarkers; |log2FC| > 0.25, adj. p < 0.01), and then used to cross-validate CellMatch v2.1 predictions against the CellMarker 2.0 database and HCC literature, requiring concordance from at least two sources for definitive annotation. Distinct cell populations were identified using characteristic marker expression: hepatocytes (ALB/AFP/APOC3/FABP1/CYP3A4), fibroblasts (COL3A1/BGN/COL1A2), CD8+ T cells (CD3D/CD3G/CCL5/CD8A/CD8B), monocytes (CD14), B cells (MS4A1/CD79A), endothelial cells (PECAM1/STAB2/CDH5) and CD4+ T cells (CD3D/CD4/CD69/IL7R),Liver progenitor cells (LPCs, EPCAM/KRT7/KRT19/CD24) and liver bud hepatocytes (CFH/TGM2) (Figure 1C and Supplementary Table 1).
CellChat Reveals Key HCC Hepatocyte Functions and Oncogenic Pathways
CellChat analysis revealed significant remodeling of cell-cell communication networks in hepatocellular carcinoma (HCC) compared to adjacent non-tumor tissue, with hepatocytes demonstrating drastically enhanced interactions with infiltrating immune cells, as evidenced by increased outgoing and incoming signaling strengths (Figure 1D and Supplementary Data 1). Further investigation identified a prominent reorganization of EPHA-mediated crosstalk in HCC in contrast to healthy tissue (Supplementary Figure 2). This was characterized by significantly strengthened autocrine signaling among hepatocytes (Figure 1E), suggesting potential self-reinforcing mechanisms driving tumor progression. Histological analysis confirmed the malignancy of HCC tissue compared to adjacent normal liver (Figure 1F). Similarly, compared to the normal architecture, Masson staining revealed prominent tumor fibrosis (arrows, Figure 1G), consistent with the observed intercellular communication alterations. Hepatocytes from HCC and normal tissues were isolated for comparative analyses (Figure 1H). Marker expression (ALB/AFP/APOC3/FABP1/CYP3A4) confirmed hepatocyte identity, while revealing HCC-specific heterogeneity (Figure 1I). Malignant transformation was evidenced by the upregulation of oncogenes (AFP/ACSL4/SERPINA1; Figure 1J) and significant parenchymal loss (16.8%; Figure 1K). Notably, tumors exhibited concurrent upregulation and co-localization of AFP and EFNA1 (p < 0.01, Figure 1L). This co-expression pattern associates the key EPHA pathway ligand EFNA1 with the canonical HCC marker AFP.
Elevated EFNA1/EPHA1 Expression in Malignant Tissues and in vitro Models
Immunofluorescence revealed significantly elevated co-expression of EFNA1 and EPHA1 in tumor tissues compared to that in adjacent normal controls (Figure 2A–C). A key finding was the observation of EFNA1 deposition gradients extending beyond producer cell boundaries, which colocalized with EPHA1 in the tumor microenvironment, supporting the functional role of soluble ligand effects in vivo. Notably, these soluble EFNA1 gradients were not observed in HepG2/LO2 cocultures. Instead, the in vitro models recapitulated membrane-localized EFNA1 (via GPI anchoring) in direct contact with EPHA1 on adjacent cells, confirming elevated co-expression at the protein level (Figure 2D–F).
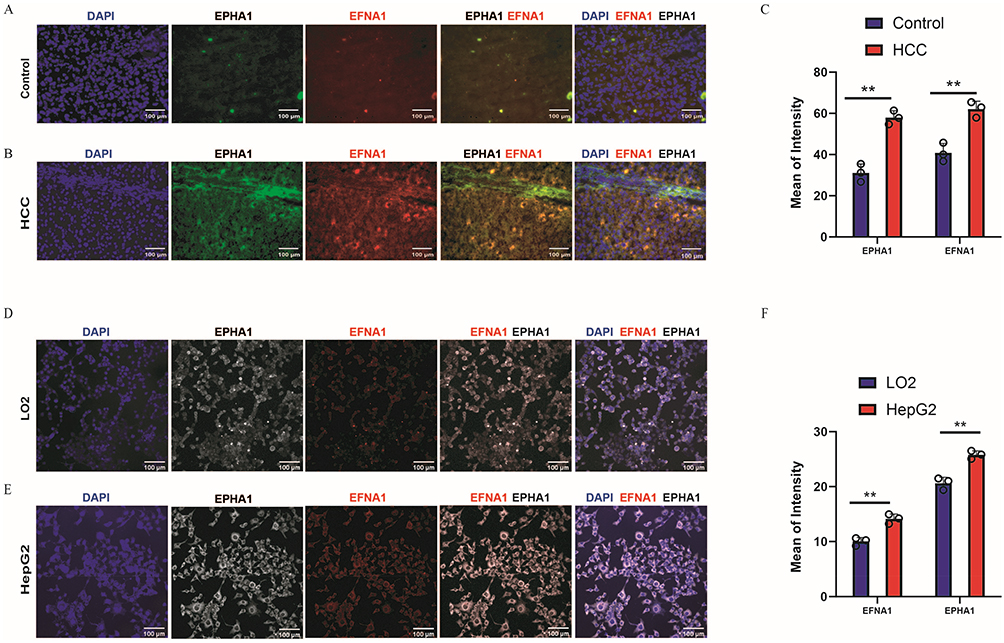 |
Figure 2 Oncogenic Upregulation of EFNA1-EPHA1 Signaling in HCC. (A) Immunofluorescence showing low EFNA1/EPHA1 co-expression in adjacent normal liver tissues (20×), Scale bar: 100 μm(B) Immunofluorescence showing high EFNA1/EPHA1 co-expression in hepatocellular carcinoma (HCC) tissues (20×), Scale bar: 100 μm. (C) Quantification of fluorescence intensity from (A and B) confirming significant overexpression in tumor tissues (n=3; mean ± SEM; **P< 0.01; t-test). (D) Immunofluorescence showing low EFNA1/EPHA1 co-expression in LO2 (normal hepatocytes) cells (20×), Scale bar: 100 μm. (E) Immunofluorescence showing high EFNA1/EPHA1 co-expression in HepG2 (HCC) cells (20×), Scale bar: 100 μm. (F) Quantification of fluorescence intensity from (D and E) confirmed significant overexpression in HepG2 cells (n=3; mean ± SEM; **P < 0.01; t-test). |
The Mechanism of EFNA1–EPHA1 Signaling Pathway in HCC
The consistent colocalization of EFNA1–EPHA1 in HCC specimens and cellular models establishes its pathogenic relevance, although the mechanistic contributions of hepatocyte-autonomous EFNA1–EPHA1 signaling to HCC progression remain to be elucidated. Comparative scRNA-seq analysis of HCC versus normal livers (GSE174748/GSE166635) revealed enhanced EPHA signaling-mediated interactions, indicating that HCC hepatocytes actively engage in both autocrine and paracrine signaling with LPCs, liver bud hepatocytes, and endothelial cells (Figure 3A). CellChat analysis demonstrated that these hepatocyte-derived signals were predominantly received by the same cell population, establishing autocrine signaling loops and intercellular crosstalk within the HCC microenvironment (Figure 3B). Notably, we identified a fundamental rewiring of EPHA signaling networks in HCC and mapped the general underlying mechanism (Figure 3C and D). While normal hepatocytes primarily utilized EFNA1–EPHA3, EFNA1–EPHA2, and EFNA2–EPHA3 pairs, HCC tissues exhibited a dramatic shift toward EFNA1–EPHA1, EFNA1–EPHA5, and EFNA1–EPHA7 interactions. Of these, EFNA1-EPHA1 was examined by single-cell RNA sequencing, which revealed fundamental differences in its signaling between normal and HCC tissues. In the normal liver, signaling originates exclusively from liver progenitor cells (LPCs), whereas HCC tissues exhibit both LPC-derived signaling and robust autocrine activation specifically within malignant hepatocytes. This cell-autonomous EFNA1–EPHA1 engagement suggests a pathological rewiring of ephrin signaling in transformed hepatocytes (Figure 3E–H).
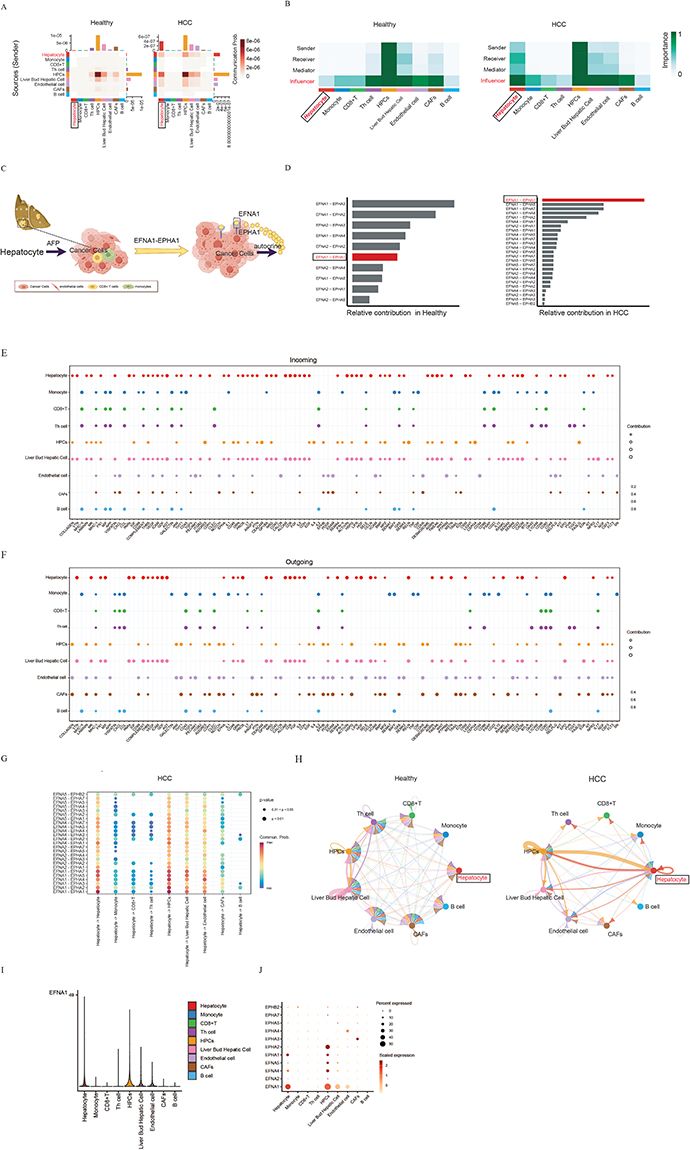 |
Figure 3 Mechanism of EPHA Signaling Pathway in HCC. (A) Communication Probability: Both the sending and receiving probabilities of the EPHA signaling pathway are elevated in HCC hepatocytes compared to healthy hepatocytes. (B) Importance Heatmap: Color intensity maps to the degree of interaction importance, demonstrating the enhanced sender and receiver functions of HCC hepatocytes relative to healthy ones. (C) Molecular Signaling in the EPHA pathway. (D) EFNA1–EPHA1 was identified as the most prominent ligand–receptor pair in HCC. (E) Screening of altered incoming ligand-receptor interactions identifies hepatocyte-derived EPHA receptors as key nodes undergoing significant change in HCC, visualized across cell types (y-axis) and a broad gene set (x-axis). (F) Screening of altered outgoing ligand-receptor interactions identifies hepatocyte-derived EPHA receptors as key nodes undergoing significant change in HCC, visualized across cell types (y-axis) and a broad gene set (x-axis). (G) Hepatocyte-derived EPHA receptors and their cognate ligand partnerships in hepatocellular carcinoma. (H). The interaction patterns of EFNA1-EPHA1 differed significantly between the healthy liver and HCC tissues. Nodes represent cell types; edge width is proportional to communication probability. (I) Violin plots showing EFNA1 expression across distinct cell populations in HCC. (J) Dot-plot analysis of hepatocyte-derived EFNA1 secretion and EPHA1 receptor interactions in HCC. |
Further comparative analysis of EFNA1–EPHA1 binding dynamics revealed distinct intercellular signaling networks in healthy and HCC tissues. In healthy liver, EFNA1–EPHA1 primarily mediated autocrine and paracrine communication between liver progenitor cells (LPCs) and liver bud hepatocytes. In contrast, HCC tissues exhibited significantly enhanced interactions between LPCs and malignant hepatocytes, concurrent with attenuated autocrine signaling in liver bud hepatocytes, but reinforced autocrine activity in both LPCs and malignant hepatocytes (Figure 3H). These findings demonstrate pathological rewiring of EFNA1–EPHA1-mediated crosstalk, wherein the ligand-receptor pair selectively amplifies “co-opt EFNA1–EPHA1 for self-sustaining proliferation”. This shift was driven by autonomous EFNA1 secretion from HCC hepatocytes and compensatory EPHA1 upregulation (Figure 3I–J).
EFNA1/EPHA1 Associates with Advanced HCC
EFNA1 and EPHA1 in HCC prognosis and clinicopathological associations. Survival analysis identified EFNA1 as a strong prognostic marker, with high expression significantly correlating with worse overall survival (P<0.001), whereas EPHA1 showed no significant survival association. However, UALCAN-TCGA analysis revealed that both genes exhibited progressively increasing expression patterns with advancing clinical stage, higher tumor grade, and lymphatic metastasis, demonstrating their consistent clinicopathological relevance (Figure 4A and B).
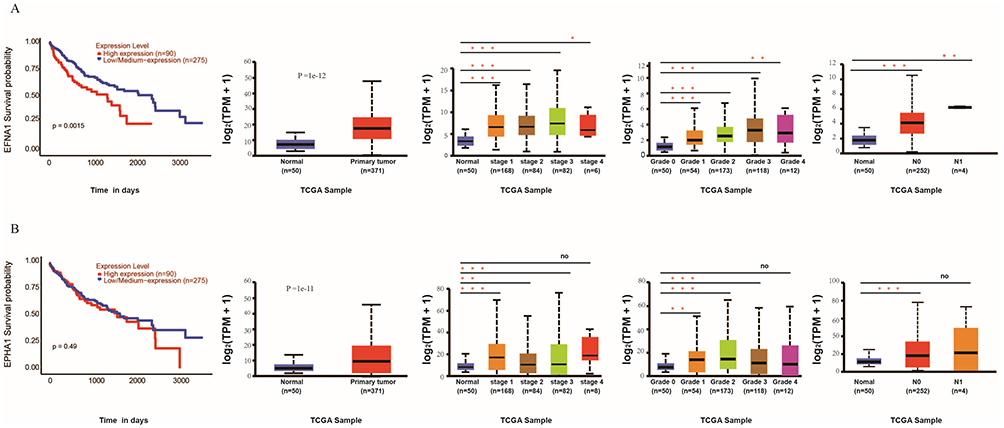 |
Figure 4 EFNA1 overexpression correlates with poor prognosis in hepatocellular carcinoma (LIHC). (A) EFNA1 and (B) EPHA1 Survival analysis demonstrated that high EFNA1 expression was significantly associated with poor OS, whereas EPHA1 showed no significant prognostic correlation. UALCAN analysis of TCGA data demonstrated the expression profiles of both genes stratified by clinical stage, tumor grade, and lymphatic metastasis, visualized in corresponding plots (noP>0.05, *P<0.05, **P<0.01, ***P<0.001). |
Discussion
The tumor microenvironment (TME) of hepatocellular carcinoma (HCC) is a complex ecosystem, in which cellular heterogeneity and dysregulated intercellular communication drive malignant progression.2 This finding is consistent with growing evidence that aberrant cell–cell communication within the TME contributes to HCC aggressiveness.16
EFNA1–EPHA1 Signaling as a Central Driver in HCC
Ligand–receptor analysis at single-cell resolution revealed a pronounced reorganization of EFNA1-centered signaling in HCC. A striking shift toward EFNA1–EPHA1 signaling emerges as the dominant characteristic of the rewired interactome in HCC, displacing the conventional EFNA1–EPHA2/EPHA3 interactions observed in normal hepatocytes (Figure 3D). In contrast, we observed consistent upregulation of EFNA1 in HCC tissues and cell lines, with pronounced co-localization with EPHA1, supporting its functional involvement in vivo. Strikingly, EFNA1 formed spatial gradients in vivo, indicative of proteolytic shedding and paracrine signaling, a phenomenon absent in 2D co-cultures. (Figure 2A–F). This divergence highlights that the full spectrum of EFNA1 signaling is likely regulated by tumor microenvironment-specific factors, such as hypoxia-induced expression and matrix metalloproteinase (MMP) activity, which are challenging to recapitulate in vitro.19,20 This aligns with emerging evidence that both EPHA1 and EFNA1 are frequently overexpressed in various malignancies, including gastric cancer, cervical cancer, and esophageal squamous cell carcinoma.13,15,16,21,22 Although EPHA1 affects hepatocellular carcinoma cell prognosis,11,23 the mechanistic role of EFNA1–EPHA1 signaling in HCC progression remains poorly characterized.
EFNA1-EPHA1 Rewires the HCC Microenvironment
Building on this rewired interaction, CellChat analysis delineated stark contrasts in EFNA1–EPHA1-mediated crosstalk between normal and malignant liver tissues. In a healthy liver, EFNA1–EPHA1 signaling is largely confined to liver progenitor cells (LPCs). In HCC, we detected expansive autocrine activation within malignant hepatocytes, along with strengthened paracrine signaling between LPCs and endothelial cells (Figure 3A and H). This rewiring may enhance tumor–stromal communication, potentially facilitating angiogenesis and immune evasion, hallmarks of HCC progression.24
The concurrent upregulation of EFNA1 and AFP in HCC tissues (Figure 1K) suggests a potential link between hepatocyte dedifferentiation and EFNA1–EPHA1 activation. Given AFP’s role as an HCC marker and its association with aggressive phenotypes,24,25 further investigation is warranted to determine whether EFNA1–EPHA1 signaling contributes to stem-like properties in HCC cells.
Clinical and Prognostic Relevance
The strong association between high EFNA1 expression and poor HCC prognosis underscores its potential as a biomarker of disease progression. Although EPHA1 expression alone was not predictive of survival, its co-upregulation with EFNA1 in advanced tumors suggested possible synergistic oncogenic effects. The progressive increase in EFNA1 and EPHA1 expression with tumor grade and metastatic status further underscores their roles in HCC aggression (Figure 4A and B), corroborating earlier studies that have implicated Eph/ephrin families in invasion, angiogenesis, immune modulation, and therapy resistance.26,27 Notably, while EPHA1 expression alone was not a standalone prognostic marker in our cohort, its functional role as the cognate receptor within the dominant, rewired autocrine loop is what defines the EFNA1–EPHA1 pair as a central signaling axis in HCC.
Therapeutic Potential of Targeting EFNA1-EPHA1
The autocrine EFNA1-EPHA1 activation loop identified in this study suggests that targeting this axis may offer a novel therapeutic strategy for HCC. Preclinical studies support the therapeutic potential of targeting this axis: EPHA1 blockade has been shown to suppress tumor growth and metastasis in Colorectal cancer and breast cancer,23,28 while EFNA1 inhibition has demonstrated efficacy in other malignancies, such as counteracting AKT/ERK1/2–driven progression in HNSCC13 and suppressing lung metastasis in ESCC via nanoparticle-mediated delivery of siEFNA1 (EA1–EYLNs–PTX/siEFNA1).29
Our findings suggest that similar approaches may be effective for HCC, particularly given the limited efficacy of current targeted therapies such as sorafenib in advanced disease.30 Future studies should evaluate the potential synergy of EFNA1–EPHA1 inhibition with immune checkpoint blockade, as this combination represents a promising strategy in light of emerging evidence that ephrin signaling modulates the TME.27,31
Limitations and Future Directions
While our study provides compelling evidence for EFNA1–EPHA1’s role in HCC, some limitations remain. First, the scRNA-seq datasets were derived from a limited number of patients, thus necessitating validation in larger cohorts. Second, the precise intracellular signaling cascades it engages (eg, potential activation of MAPK/ERK or PI3K/AKT pathways) represent a crucial next step. Future work employing phospho-specific flow cytometry, Western blot analysis in EFNA1-knockdown models, or pathway-focused reporter assays will be essential to map the direct downstream effectors, thereby transitioning our correlative discovery into a definitive mechanistic understanding. Finally, in vivo studies using hepatocyte-specific EFNA1 knockout models will help determine whether this signaling axis is necessary for tumor initiation, maintenance, or both.
Conclusion
By applying single-cell transcriptomics to decode intercellular communication, this study moves beyond correlation to provide a precise, functional definition of a pathogenic circuit in HCC: the autocrine EFNA1-EPHA1 signaling loop within malignant hepatocytes. This mechanism explains how these cells reprogram their microenvironment to fuel progression. Our work thus translates high-resolution molecular data into a directly actionable therapeutic target, with EFNA1 serving as a co-incident prognostic biomarker.
Abbreviations
HCC, Hepatocellular Carcinoma; scRAN-seq, Single-cell RNA sequencing; TME, tumor microenvironment; TKIs, kinase inhibitors; ICIs, immune checkpoint inhibitors; LPCs, liver progenitor cells.
Data Sharing Statement
The data that support the findings of this study are available from the corresponding author Xueping Feng upon reasonable request.
Funding
The Youjiang Medical University for Nationalities’ High-level Talent Research Project (#RZ2300001266), the Baise Science & Technology Research Program on Development (#Baike20234214), and the 2024 Guangxi Autonomous Region College Students Innovation and Entrepreneurship Training Program (#33) provided funding for this work.
Disclosure
The authors declare no commercial conflicts of interest or financial relationships related to this study.
References
1. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–11. doi:10.3322/caac.21660
2. Liu P, Kong L, Liu Y, Li G, Xie J, Lu X. A key driver to promote HCC: cellular crosstalk in tumor microenvironment. Front Oncol. 2023;13:1135122. doi:10.3389/fonc.2023.1135122
3. Sensi B, Angelico R, Toti L, et al. Mechanism, Potential, and Concerns of Immunotherapy for Hepatocellular Carcinoma and Liver Transplantation. Curr Mol Pharmacol. 2024;17:e18761429310703.
4. Childs A, Aidoo-Micah G, Maini MK, Meyer T. Immunotherapy for hepatocellular carcinoma. JHEP Rep. 2024;6(10):101130. doi:10.2174/0118761429310703240823045808
5. Yin Y, Feng W, Chen J, et al. Immunosuppressive tumor microenvironment in the progression, metastasis, and therapy of hepatocellular carcinoma: from bench to bedside. Exp Hematol Oncol. 2024;13(1):72. doi:10.1186/s40164-024-00539-x
6. Sneller L, Mathur K, Kottilil S, Mathur P. Hot and Cold HCC: uncoupling Viral Oncogenesis and Therapy. Viruses. 2025;17(9):1255. doi:10.3390/v17091255
7. Lu Y, Yang A, Quan C, et al. A single-cell atlas of the multicellular ecosystem of primary and metastatic hepatocellular carcinoma. Nat Commun. 2022;13(1):4594. doi:10.1038/s41467-022-32283-3
8. Huang S, Dong C, Zhang J, Fu S, Lv Y, Wu J. A comprehensive prognostic and immunological analysis of ephrin family genes in hepatocellular carcinoma. Front Mol Biosci. 2022;9:943384. doi:10.3389/fmolb.2022.943384
9. Wang P, Ma Y, Zhao Y, et al. Single-cell RNA sequencing unveils tumor heterogeneity and immune microenvironment between subungual and plantar melanoma. Sci Rep. 2024;14(1):7039. doi:10.1038/s41598-024-57640-8
10. Li P, Wang L, Li P, et al. Retraction Note: silencing of long non-coding RNA XIST represses gastric cancer progression through blocking NFκB pathway via inhibiting HNF4A-mediated transcription of EPHA1. Cancer Gene Ther. 2023;30(10):1441. doi:10.1038/s41417-023-00667-2
11. Li M, Cheng Y, Qiu F, et al. LncRNA EBLN3P/microRNA-323a-3p/EphA1 axis affects the biological function of hepatocellular carcinoma cells. Discov Oncol. 2025;16(1):1145.
12. Li S, Sun M, Cui Y, et al. Ephrin A1 functions as a ligand of EGFR to promote EMT and metastasis in gastric cancer. EMBO J. 2025;44(5):1464–1487. doi:10.1038/s44318-025-00363-x
13. Chu LY, Huang BL, Huang XC, Peng YH, Xie JJ, Xu YW. EFNA1 in gastrointestinal cancer: expression, regulation and clinical significance. World J Gastrointest Oncol. 2022;14(5):973–988.
14. Cui Y, Chang Y, Ma X, et al. Ephrin A1 stimulates CCL2 secretion to facilitate premetastatic niche formation and promote gastric cancer liver metastasis. Cancer Res. 2025;85(2):263–276. doi:10.1158/0008-5472.CAN-24-1254
15. Chu LY, Wu FC, Fang WK, et al. Secreted proteins encoded by super enhancer-driven genes could be promising biomarkers for early detection of esophageal squamous cell carcinoma. Biomed J. 2024;47(4):100662. doi:10.1016/j.bj.2023.100662
16. Li P, Wang L, Li P, et al. Silencing of long non-coding RNA XIST represses gastric cancer progression through blocking NFκB pathway via inhibiting HNF4A-mediated transcription of EPHA1. Cancer Gene Ther. 2021;28(3–4):307–320. doi:10.1038/s41417-020-00220-5
17. Meng Y, Zhao Q, An L, et al. A TNFR2-hnRNPK axis promotes primary liver cancer development via activation of YAP signaling in hepatic progenitor cells. Cancer Res. 2021;81(11):3036–3050. doi:10.1158/0008-5472.CAN-20-3175
18. Filliol A, Saito Y, Nair A, et al. Opposing roles of hepatic stellate cell subpopulations in hepatocarcinogenesis. Nature. 2022;610(7931):356–365. doi:10.1038/s41586-022-05289-6
19. Ning W, Qiao N, Zhou L, et al. Integrative multiscale analysis reveals EFNA1-driven immune remodeling promotes colorectal cancer lymph node metastasis. Hum Mutat. 2026;2026(1):8553028. doi:10.1155/humu/8553028
20. Gonzalez-Avila G, Sommer B, Flores-Soto E, Aquino-Galvez A. Hypoxic effects on matrix metalloproteinases’ expression in the tumor microenvironment and therapeutic perspectives. Int J Mol Sci. 2023;24(23):16887. doi:10.3390/ijms242316887
21. Jiang H, Wang S, Liu Y, et al. Targeting EFNA1 suppresses tumor progression via the cMYC-modulated cell cycle and autophagy in esophageal squamous cell carcinoma. Discov Oncol. 2023;14(1):64. doi:10.1007/s12672-023-00664-9
22. Liu SQ, Cheng XX, He S, et al. Super-enhancer-driven EFNA1 fuels tumor progression in cervical cancer via the FOSL2-Src/AKT/STAT3 axis. J Clin Invest. 2025;135(8):e177599. doi:10.1172/JCI177599
23. Wu Y, Du Z, Mou J, et al. The functions of EphA1 receptor tyrosine kinase in several tumors. Curr Med Chem. 2023;30(20):2340–2353. doi:10.2174/0929867329666220820125638
24. Lu X, Deng S, Xu J, et al. Combination of AFP vaccine and immune checkpoint inhibitors slows hepatocellular carcinoma progression in preclinical models. J Clin Invest. 2023;133(11):e163291. doi:10.1172/JCI163291
25. Tan XP, Zhou K, Zeng QL, Yuan YF, Chen W. Influence of AFP on surgical outcomes in non-B non-C patients with curative resection for hepatocellular carcinoma. Clin Exp Med. 2023;23(1):107–115. doi:10.1007/s10238-022-00813-4
26. Giannopoulos K, Karikis I, Byrd C, et al. Eph/ephrin-mediated immune modulation: a potential therapeutic target. Front Immunol. 2025;16:1539567. doi:10.3389/fimmu.2025.1539567
27. Pasquale EB. Eph receptors and ephrins in cancer progression. Nat Rev Cancer. 2024;24(1):5–27. doi:10.1038/s41568-023-00634-x
28. Zhang Y, Lai X, Yue Q, et al. Bone marrow mesenchymal stem cells-derived exosomal microRNA-16-5p restrains epithelial-mesenchymal transition in breast cancer cells via EPHA1/NF-κB signaling axis. Genomics. 2022;114(3):110341. doi:10.1016/j.ygeno.2022
29. Xie F, Qiu J, Sun C, et al. Development of a specific aptamer-modified nano-system to treat esophageal squamous cell carcinoma. Adv Sci. 2024;11(28):e2309084. doi:10.1002/advs.202309084
30. Li Q, Chen K, Zhang T, et al. Understanding sorafenib-induced ferroptosis and resistance mechanisms: implications for cancer therapy. Eur J Pharmacol. 2023;955:175913. doi:10.1016/j.ejphar.2023.175913
31. Xie R, Yuan M, Jiang Y. The pan-cancer crosstalk between the EFNA family and tumor microenvironment for prognosis and immunotherapy of gastric cancer. Front Cell Dev Biol. 2022;10:790947. doi:10.3389/fcell.2022.790947
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.