Back to Journals » Journal of Inflammation Research » Volume 19
S100A12 as a Shared Inflammatory Driver of Intervertebral Disc Degeneration and Osteoporosis: A Mendelian Randomization and Experimental Study
Authors Wang Z, Chen B ![]() , Liu Z, Zhang Y, Han L
, Liu Z, Zhang Y, Han L
Received 8 September 2025
Accepted for publication 7 March 2026
Published 9 April 2026 Volume 2026:19 562027
DOI https://doi.org/10.2147/JIR.S562027
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ujjwol Risal
Zexin Wang,1,* Bin Chen,1,* Zhenchuan Liu,1 Yuanqiang Zhang,1 Leixiang Han2
1Department of Orthopedic Surgery, Qilu Hospital, Cheeloo College of Medicine, Shandong University, Jinan, Shandong, 250012, People’s Republic of China; 2Department of Spine Surgery, Qilu Hospital (Qingdao), Cheeloo College of Medicine, Shandong University, Qingdao, Shandong, 266035, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Leixiang Han, Email [email protected]
Objective: S100A12 as an indispensable molecular components inflammatory cytokines in human physiology. Notably, similar pathological mechanisms involving chronic low-grade inflammation are also observed in intervertebral disc degeneration (IVDD) and osteoporosis (OP), making the interplay between these diseases and inflammatory cytokines indisputable. Nevertheless, the precise identity of cytokines that concurrently fuel IVDD and OP remains elusive, hindering targeted therapeutic strategies.
Methods: We employed genome-wide significant SNPs as instrumental variables (IVs) to assess causality between circulating inflammatory cytokines, IVDD, and OP using Mendelian randomization (MR). Data on cytokines came from Zhao et al, while IVDD and OP data were from the FinnGen database, predominantly European participants. The inverse-variance weighted (IVW) method was employed, with validation from MR-Egger, weighted median, weighted mode, and simple mode. In later experimental studies, one-way ANOVA was used to analyze differences among multiple groups, followed by Dunnett’s t-test for post-hoc comparisons. Finally, cell and human samples were utilized to validate our findings.
Results: In our study, positive correlations were found between CXCL10, CXCL6, S100A12, NT-3, uPA, IL7, FLT3L, Caspase 8, TNF, and IL18 levels with IVDD/OP. While, negative correlations were observed with DNER, IL10, SIRT2, OPG, MCP-3, CCL19, SCF, GDNF, MCP-2, CCL25, and CD40L receptor levels. Reverse MR analysis showed no significant associations, indicating unidirectional causal relationships. Notably, S100A12 was identified as a shared positive risk factor influencing both IVDD and OP pathogenesis. Also, the S100A12 was significantly increased in IVDD and OP patients/cells than normal samples.
Conclusion: Our study indicates a potential causal relationship that S100A12 exhibiting positive correlations with both IVDD and OP. The results strengthen the understanding of the pathogenesis of IVDD and OP, as well as, provide new treatment strategies for IVDD and OP.
Keywords: intervertebral disc degeneration, osteoporosis, inflammatory cytokines, mendelian randomization, S100A12
Introduction
Inflammatory cytokines are indispensable molecular components in human physiology, functioning as critical mediators that orchestrate immune surveillance, modulate inflammatory cascades, and maintain homeostasis.1 Their dual role as regulators and biomarkers has solidified their prominence in the pathogenesis of diverse diseases, ranging from degenerative disease to metabolic disease.2 Emerging evidence highlights their central role in these diseases under specific pathological conditions.3 For instance, during COVID-19, dynamic shifts in cytokine profiles—notably elevated IL-6 and IL-8—have been identified as prognostic indicators, correlating with disease severity and clinical outcomes.4,5 S100A12, a ubiquitously expressed factor in humans, has been implicated in diverse pathological conditions including malignancies, cardiovascular diseases and inflammatory disorders. However, there are currently no research reports indicating that S100A12 is associated with intervertebral disc degeneration (IVDD) and osteoporosis (OP). Notably, similar pathological mechanisms involving chronic low-grade inflammation are also observed in IVDD and OP, making the interplay between these diseases and inflammatory cytokines indisputable.6,7
IVDD, a pervasive degenerative spinal disorder, clinically manifests as chronic low back pain (LBP), disc herniation, and spinal stenosis, significantly impairing quality of life.8 Mechanistic studies reveal that IVDD progression is intricately linked to a pro-inflammatory microenvironment within degenerated discs.9 Key cytokines exhibit stage-dependent upregulation, mirroring the severity of disc degeneration and serving as quantifiable biomarkers for disease staging and therapeutic monitoring.10,11
While traditionally viewed as a metabolic imbalance, emerging paradigms implicate inflammatory cytokines in OP pathogenesis.12 Chronic inflammation accelerates bone resorption via osteoclast activation, mediated by cytokines such as IL-1β, IL-6, and TNF-α, which disrupt the bone remodeling equilibrium.13 Epidemiological studies further highlight a clinical overlap: OP patients exhibit a heightened predisposition to IVDD, suggesting shared inflammatory pathways.14 Paradoxically, despite this comorbidity, investigations focusing on bone mineral density (BMD)—a cornerstone of OP diagnosis—have failed to establish a direct causal or correlative link between OP and IVDD.15,16 This discordance underscores the necessity to pivot toward inflammatory mechanisms as a unifying framework for understanding their coexistence.
Nevertheless, the precise identity of cytokines that concurrently fuel IVDD and OP remains elusive, hindering targeted therapeutic strategies. Resolving this knowledge gap is imperative for developing dual-purpose interventions, stratifying high-risk cohorts, and implementing preemptive measures.
To address this, Mendelian randomization (MR) analysis offers a robust methodological approach. By leveraging genetic variants associated with inflammatory cytokines as instrumental variables, MR mitigates confounding biases inherent in observational studies, enabling causal inference between exposures and outcomes.17 This method circumvents limitations posed by reverse causation and unmeasured confounders, providing higher evidentiary validity.18 Our study employs a bidirectional MR framework to dissect the causal relationships among IVDD, OP, and inflammatory cytokines. The objectives are twofold: (1) to identify cytokines with pleiotropic effects on both conditions and (2) to delineate actionable targets for preventive and therapeutic modalities.
Materials and Methods
Genome-Wide Association Study (GWAS) Data Source
In this study, the GWAS summary statistics for circulating inflammatory cytokines were collected from the Zhao etal The GWAS summary statistics for IVDD and OP were derived from the FinnGen database, which including 368,519 participants (Ncase = 46,205, Ncontrol = 322,314) and 438,772 participants (Ncase = 9046, Ncontrol = 429,826), respectively.19
Study Design of Mendelian Randomization (MR)
We aimed to investigate the reciprocal causal relationships among circulating inflammatory cytokines, IVDD and OP by performing a two-sample MR analysis. Based on the random allocation of genetic variation, an MR analysis is a research method that identifies the direct causal relationship between an exposure and an outcome. In observational studies, MR has the benefit of avoiding reverse causality and minimizing confounding effects. The following three key assumptions must be satisfied by effective instrumental variables when constructing causal inferences: (1) significant association between the single-nucleotide polymorphisms (SNPs) and the exposure; (2) no confounding factors associated with the SNP as a result of its association with the outcome; (3) independent association between the SNP and the outcome. The experimental design is shown in Figure 1. Several previous studies collected GWAS data from large-scale human participants for our analysis that were publically available. The institutional review committees in each study approved the data after receiving informed consent from participants.
 |
Figure 1 Mendelian randomization study design schematic diagram. |
Selection of Instrumental Variables (IVs) and Data Harmonization
In our study, SNPs were defined as instrumental variables (IVs).20 Initially, we included SNPs associated with the circulating inflammatory cytokines exposure at a genome-wide significance threshold P < 5×10−6. Subsequently, a pairwise linkage disequilibrium analysis was then performed on the selected SNPs to exclude those in linkage disequilibrium (R2 > 0.001, window distance < 10000 kb). The R2-statistic were calculated as. For each SNP, the F-statistic was computed, and the strength of each instrumental variable was evaluated based on the variance the SNP explained for each exposure, calculated as:
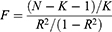
where K is the number of genetic variants and N is the sample size. We excluded SNPs with an F less than 10 and harmonized the data so that the effect of each SNP on exposure and outcome corresponds to the same allele.
Cell Culture
Human nucleus pulposus cells (hNPCs) and human umbilical cord mesenchymal stem cells (hUCMSCs) were purchased from the Immocell Biotechnology Company (Xiamen, China). hNPCs were grown in α-Minimum Essential Medium (α-MEM, Gibco, USA) with the addition of 10% fetal bovine serum (FBS, Gibco, USA) and 1% penicillin-streptomycin. Cells were used between passages 3 and 5 for all experiments and were seeded at a density of 8 × 103 cells/cm2 for intervention studies. hUCMSCs were cultured in Dulbecco’s Modified Eagle Medium (DMEM, Gibco, USA) with 10% FBS and 1% penicillin-streptomycin, and used between passages 3 and 5 at a seeding density of 5000 cells per cm2. All cells were incubated in a humidified atmosphere containing 5% CO2 at 37°C and were routinely tested to ensure the absence of mycoplasma contamination.
Cell Intervention
The hUCMSCs were divided into control and model groups. The control group was cultured in osteogenic induction medium (OIM), while the model group was treated with 50 nM dexamethasone (Dex) added to the OIM for 7 days to inhibit osteogenic differentiation and simulate the in vitro phenotype of OP. Similarly, hNPCs were divided into control and model groups. The model group was stimulated with interleukin-1β (IL-1β) (10 ng/mL) for 48 hours to establish a chronic inflammation-induced degeneration model, whereas the control group received no cytokine treatment.
RNA Isolation and qRT-PCR
Using Trizol Reagent (Invitrogen, Carlsbad, CA, USA), total RNA was extracted from cultured cells according to the provided instructions. Then, cDNA was synthesized with a reverse transcription kit (Takara, Kohoku-Cho, Kusatsu, Japan). The qRT-PCR was performed with FastStart Essential DNA Green Master on a LightCycler96 device from Roche, Basel, Switzerland. The mRNA expression relative to the internal control (Arppo RNA) was determined using the 2-ΔΔCt method. The target gene primer sequences were see in Table 1.
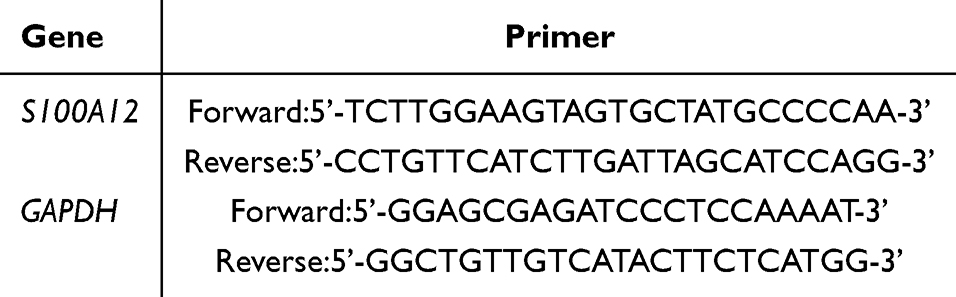 |
Table 1 Primer Sequences |
Western Blot (WB)
Cells from various groups were rinsed three times with PBS, and total proteins were extracted using RIPA lysis buffer with the addition of 10% phosphatase inhibitors and 1% PMSF. Proteins were denatured by boiling, and the supernatant was separated by centrifugation to prepare samples for analysis. Total protein content was quantified using the BCA Protein Assay Kit and then normalized. The samples were then processed using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and moved onto a nitrocellulose membrane. The membrane was blocked with skim milk at room temperature for 1 hour, followed by incubation with various primary antibodies S100A12 (diluted 1:500, Affinity Biosciences, DF7277, CN) and β-actin (diluted 1:5000, proteintech, 51067-2-AP, CN) at 4°C overnight, and then incubated with HRP-conjugated secondary antibodies (1:10000) for 1 hour. Chemiluminescence detection was performed using ECL developer, and semi-quantitative analysis was conducted using ImageJ software. To guarantee reproducibility, each experiment was conducted three times.
Collection of Human Specimens
After obtaining informed consent, human nucleus pulposus tissues were collected from 15 patients undergoing lumbar discectomy for symptomatic lumbar disc herniation (LDH) with imaging-confirmed Pfirrmann grade IV–V disc degeneration. Bone tissues were obtained from 15 patients with OP undergoing total knee arthroplasty, diagnosed based on a femoral neck or lumbar spine T-score ≤ −2.5. Control nucleus pulposus samples (n=10) were obtained from patients with traumatic lumbar fractures without prior disc degeneration (Pfirrmann grade I–II). Control bone samples (n=10) were collected from age- and sex- matched patients undergoing joint replacement due to trauma, with normal bone mineral density (T-score > −1.0). All donors were aged 45–70 years and were excluded if they had malignancies, metabolic bone diseases other than OP, chronic inflammatory disorders, or previous treatment with bone-modifying agents. All human samples were obtained from Qilu Hospital. This study was approved by the Ethics Committee of Qilu Hospital (KYLL-2022(ZM)-136).
Immunohistochemistry (IHC)
For IHC analysis, the tissue sections were dewaxed and then treated with 0.3% hydrogen peroxide to inhibit endogenous peroxidase activity. Subsequently, they were blocked with normal goat serum at room temperature for 20 minutes. The sections were then incubated with primary antibody against S100A12 (diluted 1:50, Affinity Biosciences, DF7277, China) overnight at 4 °C. Non‑specific IgG antibody was used as the negative control. The next day, sections were treated with suitable HRP-conjugated secondary antibodies for one hour at room temperature. Positive staining was visualized using diaminobenzidine (DAB) solution (Invitrogen, MD). Two seasoned pathologists independently assessed all slides without knowledge of the clinical and experimental group assignments. Staining intensity and the proportion of positively stained cells were assessed using a semi‑quantitative H‑score method. In short, the staining intensity was classified as 0 for negative, 1 for weak, 2 for moderate, and 3 for strong. The H‑score was calculated as: H‑score = Σ (intensity × percentage of cells at that intensity), resulting in a range of 0–300. For each case, five representative high‑power fields (×400) were analyzed. The mean H‑score of the two observers was used for statistical analysis; discrepancies were resolved by joint re‑evaluation.
Statistical Analysis
We performed a two-sample bidirectional MR to evaluate the mutual causal links between circulating inflammatory cytokines, IVDD, and OP. According to a random-effects model, the IVW was the primary statistical analysis approach.21 Additionally, a further validation of IVW results was conducted using four complementary methods (MREgger,22 weighted median,23 weighted mode,23 and simple mode). All SNPs are assumed to satisfy the three MR assumptions in the IVW approach. The intercept was used to identify horizontal pleiotropy in MR-Egger analysis. A causal relationship was assessed by calculating a weighted median and weighted mode.
An analysis of SNP heterogeneity was conducted using Cochran’s Q statistic and funnel plots.24 The MR-Egger intercept method21 and the MR-PRESSO method25 were used to detect horizontal pleiotropy. If outliers were detected, they were removed, and the MR causal estimates were re-assessed. In cases where heterogeneity persisted after removing outliers, a random effects model was tested for stability, which is less prone to weaker SNP-exposure associations. Ultimately, a leave-one-out analysis was performed to evaluate the influence of each SNP on the overall causal estimates. P > 0.05 means that there is no heterogeneity and multiple effects.
All statistical analyses in 2.6/2.7/2.9 subject were performed using SPSS 25.0 software.26 Data from separate experiments are shown as the mean ± SEM, and comparisons between two groups were conducted using Dunnett’s t-test. A one-way ANOVA was used to analyze data across multiple groups, followed by an LSD test for post hoc analysis, with statistical significance determined at P<0.05.
Results
Identification of Genetic Instruments
Following the removal of palindromic/ambiguous SNPs, SNPs without proxy variants, and SNPs with incorrect causal direction as identified by MR Steiger filtering, genome-wide significant SNPs were selected as instrumental variables. The number of SNPs used for each pQTL and eQTL analysis ranged from 30 to 9682. All SNPs possessed an F-statistic greater than 10, indicating no weak instrumental variants were included. The SNPs from the exposure datasets that showed a causal association with the outcome measures are detailed in Supplementary Table 1.
Association of Circulating Inflammatory Cytokines with IVDD
MR-PRESSO analysis detected no SNP outliers, and the MR-Egger regression showed no evidence of pleiotropy (P > 0.05; Supplementary Table 2). The causal relationships between 91 circulating inflammatory cytokines and IVDD, as analyzed by the methods above, are summarized in Figure 2. Eight of these cytokines do not have multiple effects. The IVW analysis revealed a positive correlation between C-X-C motif chemokine 10 (CXCL10) levels (P < 0.001, OR = 1.031, 95% CI: 1.023–1.04), C-X-C motif chemokine 6 (CXCL6) levels (P < 0.001, OR = 1.011, 95% CI: 1.008–1.015), Protein S100-A12 (S100A12) levels (P < 0.001, OR = 1.09, 95% CI: 1.076–1.105), Neurotrophin-3 (NT-3) levels (P < 0.001, OR = 1.108, 95% CI: 1.082–1.135), Urokinase-type plasminogen activator (uPA) levels (P < 0.001, OR = 1.016, 95% CI: 1.01–1.023) and IVDD (Figure 2 and Supplementary Table 2). The IVW analysis revealed a negative correlation between Delta and Notch-like epidermal growth factor-related receptor (DNER) levels (P < 0.001, OR = 0.982, 95% CI: 0.977–0.987), Interleukin-10 (IL10) levels (P < 0.001, OR = 0.782, 95% CI: 0.776–0.788), SIR2-like protein 2 (SIRT2) levels (P = 0.014, OR = 0.967, 95% CI: 0.942–0.993) and IVDD (Figures 3 and 4, Supplementary Table 3). The P value of all IVW and MR-Egger regression analyses of CXCL10, IL10, uPA yielded Cochran’s Q statistics were less than 0.05, indicating a possible heterogeneity among the SNPs (Supplementary Table 4).
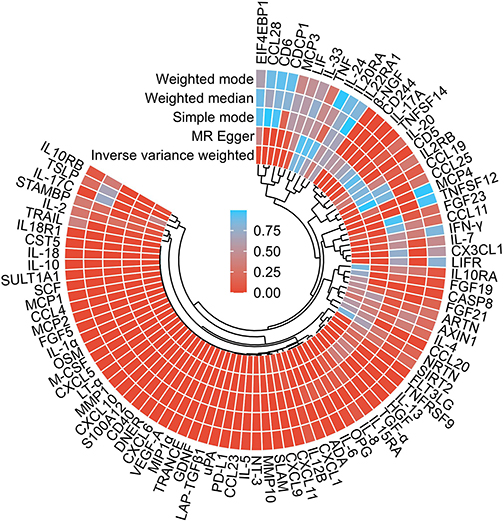 |
Figure 2 A panorama of all the 91 circulating inflammatory cytokines and their causal estimate on the risk of IVDD. |
 |
Figure 3 Positive MR estimates of the causal association between circulating inflammatory cytokines and the risk of IVDD. |
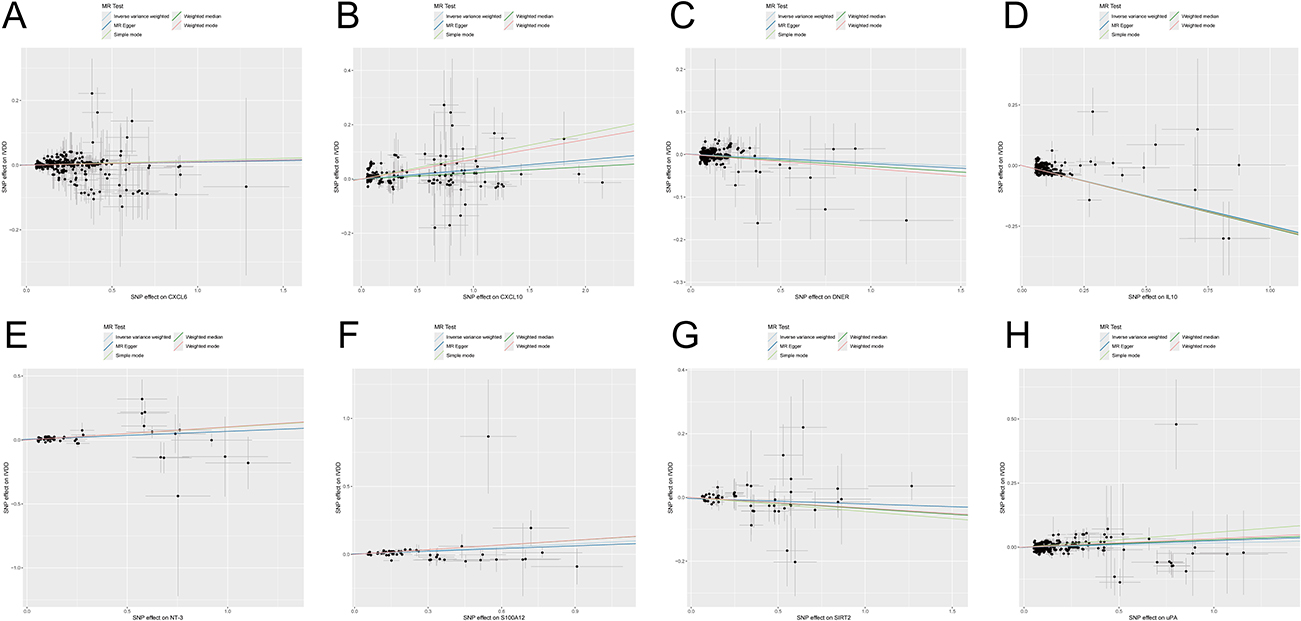 |
Figure 4 Scatter plot of (A) CXCL6, (B) CXCL10, (C) DNER, (D) IL10, (E) NT-3, (F) S100A12, (G) SIRT2, and (H) uPA with IVDD. |
Association of Circulating Inflammatory Cytokines with OP
MR-PRESSO analysis detected no SNP outliers, and the MR-Egger regression showed no evidence of pleiotropy (P > 0.05; Supplementary Table 5). The causal relationships between 91 circulating inflammatory cytokines and OP, as analyzed by the methods above, are summarized in Figure 5. Fourteen of these cytokines do not have multiple effects. The IVW analysis revealed a positive correlation between Interleukin-7 levels (IL-7) (P < 0.001, OR = 1.082, 95% CI: 1.057–1.108), S100A12 levels (P < 0.001, OR = 1.099, 95% CI: 1.068–1.132), Fms-related tyrosine kinase 3 ligand (FLT3L) levels (P < 0.001, OR = 1.108, 95% CI: 1.093–1.123), Caspase 8 levels (P < 0.001, OR = 1.131, 95% CI: 1.098–1.165), Tumor necrosis factor (TNF) levels (P < 0.001, OR = 1.166, 95% CI: 1.105–1.231), Interleukin-18 (IL-18) levels (P < 0.001, OR = 1.171, 95% CI: 1.161–1.181) and OP (Figure 5 and Supplementary Table 5). The IVW analysis revealed a negative correlation between Osteoprotegerin (OPG) levels (P < 0.001, OR = 0.934, 95% CI: 0.921–0.946), Monocyte chemoattractant protein-3 (MCP-3) levels (P < 0.001, OR = 0.943, 95% CI: 0.926–0.961), C-C motif chemokine 19 levels (CXCL19) (P < 0.001, OR = 0.943, 95% CI: 0.933–0.954), Stem cell factor (SCF) levels (P < 0.001, OR = 0.945, 95% CI: 0.936–0.954), Glial cell line-derived neurotrophic factor (GDNF) levels (P < 0.001, OR = 0.95, 95% CI: 0.939–0.961), Monocyte chemoattractant protein 2 (MCP-2) levels (P < 0.001, OR = 0.968, 95% CI: 0.964–0.973), C-C motif chemokine 25 levels (CXCL25) (P < 0.001, OR = 0.972, 95% CI: 0.967–0.977), CD40L receptor levels (P < 0.001, OR = 0.978, 95% CI: 0.967–0.989) and OP (Figures 6 and 7, Supplementary Table 6). Neither IVW nor MR-Egger regression showed significant heterogeneity (Cochran’s Q P > 0.05; Supplementary Table 7). In contrast, analyses of CXCL19, CD40L receptor, OPG, and SCF indicated potential heterogeneity (Cochran’s Q P < 0.05; Supplementary Table 7).
 |
Figure 5 A panorama of all the 91 circulating inflammatory cytokines and their causal estimate on the risk of OP. |
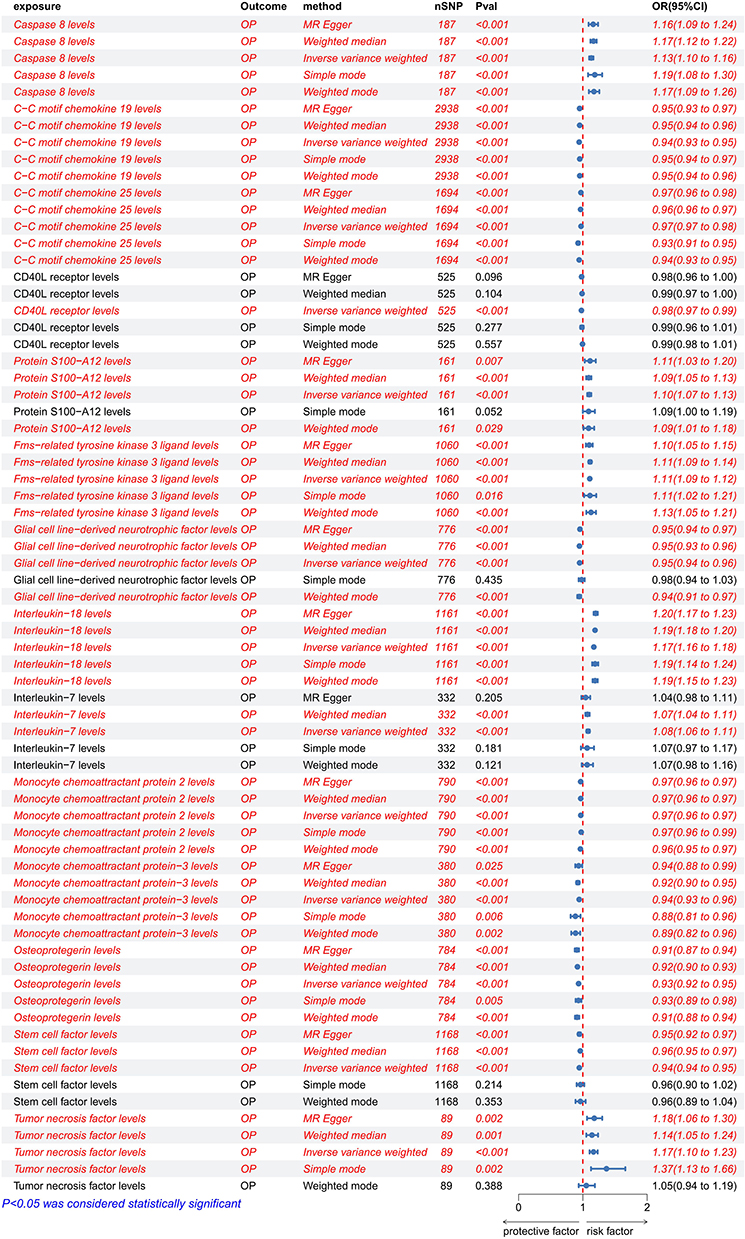 |
Figure 6 Positive MR estimates of the causal association between circulating inflammatory cytokines and the risk of OP. |
 |
Figure 7 Scatter plot of (A) Caspase 8, (B) CXCL19, (C) CXCL25, (D) CD40L receptor, (E) S100A12, (F) FLT3L, (G) GDNF, (H) IL18, (I) IL7, (J) MCP-2, (K) MCP-3, (L) OPG, (M) SCF, and (N) TNF with IVDD. |
Reverse MR Analysis
Following filtration for palindromic/ambiguous SNPs, SNPs without proxies, and reverse causality via MR Steiger, genome-wide significant SNPs were used as instrumental variables. MR-PRESSO detected no outliers, and MR-Egger regression indicated no significant pleiotropy (P > 0.05). Reverse MR analysis using IVW showed no associations, and no heterogeneity was observed in IVW or MR-Egger models (all Cochran’s Q P > 0.05). See Supplementary Tables 8–10 for details.
Experiments Analysis
We next examined the S100A12 expression in vitro and in vivo (Figure 8A). We examined the expression of S100A12 in hUCMSCs and hNPCs under different intervention conditions using WB. The results showed that the expression of S100A12 was significantly upregulated in osteogenesis-inhibited hUCMSCs, and a similar trend was observed in degenerated hNPCs (Figure 8B). Further qRT-PCR analysis was conducted to determine whether S100A12 was regulated at the pre-translational level. The results indicated that the expression of S100A12 in osteogenesis-inhibited hUCMSCs was approximately 5.2 times that of the control group, and it was increased by 9.9 times in IL-1β group than control (Figure 8C). To further validate the association between S100A12 and OP as well as IVDD, we performed IHC on human-derived nucleus pulposus and bone tissues. By comparing samples from patients with varying degrees of the conditions, the images and quantitative results indicated that the expression of S100A12 was significantly increased in the bone tissues of OP patients compared to the normal group, and there was also a certain degree of upregulation in the nucleus pulposus tissues of IVDD patients (Figure 8D).
 |
Figure 8 S100A12 is pathologically upregulated in OP and IVDD. (A) Schematic diagram of S100A12 expression examined in vitro and in vivo. (B) Western blot bands and semi-quantitative analysis of S100A12 after stimulation with IL-1β or dexamethasone. (C) Relative mRNA expression levels of S100A12 after stimulation with IL-1β or dexamethasone. (D) Representative images and quantification of S100A12 expression in tissues with different degrees of degeneration as examined by IHC. *: p < 0.05, ***: p < 0.001. |
Discussion
IVDD and OP are common musculoskeletal disorders that severely affect patients’ quality of life and impose a significant socioeconomic burden worldwide.27 Despite the high incidence rates of these conditions, their shared molecular mechanisms remain poorly understood. Previous studies explored IVDD and OP from the perspectives of pyroptosis and autophagy. The pyroptosis pathway represents a major mechanism influencing both IVDD and OP.28 Experiments clearly demonstrated that the slow loss of bone mass is associated with the disrupted microenvironment following cell apoptosis, while increased apoptosis of osteoblasts and elevated expression of inflammatory cytokines further limit compensatory increases in bone formation. Reduced bone strength in aging is also closely linked to osteocyte death. Additionally, apoptosis amplifies the inflammatory environment intracellularly, thereby affecting IVDD and OP. Cellular autophagy is a key factor in maintaining bone homeostasis; its dysregulation disrupts the stable environment, leading to excessive bone loss and reduced bone formation, ultimately triggering degenerative bone diseases like IVDD and OP.29 Furthermore, studies indicate that stem cell-derived extracellular vesicles (EVs) show significant therapeutic potential for such diseases.30 EVs can regulate osteoclast activity and bone resorption, playing a vital role in restoring bone homeostasis.31 EVs also alleviate further disc degeneration by exerting anti-inflammatory effects, modulating inflammatory responses in intervertebral discs, and inhibiting apoptosis of disc cells.32 However, previous studies mostly reported relationships between individual diseases and inflammatory factors, and common inflammatory factors affecting both IVDD and OP have not yet been reported. Research indicates that inflammatory factors play a pivotal role in bone tissue metabolism, significantly influencing BMD and thereby driving the onset and progression of OP.33 Concurrently, OP induces structural compromise of vertebral bodies,34 triggering the release of substantial inflammatory factors from bone cells,15 which subsequently exacerbates the progression of IVDD.28 Therefore, discussing IVDD and OP together is necessary, aiding in the prevention and early detection of these diseases. Our study were the first one using MR and experimental validation linking S100A12 to both IVDD and OP.
Shared Inflammatory Pathways in IVDD and OP
Previous studies have demonstrated that the pathogenesis of IVDD is mechanistically linked to inflammatory cytokines. For example, the upregulated expression of IL-1β and TNF-α in IVDD drive disease progression and initiation through modulating inflammatory responses, cellular proliferation.11 Integrated MR and immunomics analyses revealed a robust causal association between the C-type lectin domain family 11, member A (CLEC11A) causative gene and the pathogenesis and progression of IVDD. Notably, MR analyses demonstrated that CLEC11A elevates IVDD risk by mediating the upregulation of artemin (ARTN), a potent inflammatory mediator involved in nociceptive signaling pathways.35 MR analyses demonstrated that elevated levels of stromal cell-derived factor 1 alpha (SDF-1α) were positively associated with an increased risk of IVDD. ELISA experiments demonstrated a significant positive correlation between prostaglandin E2 (PGE2) levels and the pathogenesis of IVDD, with elevated PGE2 concentrations mechanistically linked to cyclooxygenase-2 (COX-2) -mediated inflammatory cascades in degenerated disc tissues.36 Conversely, higher concentrations of IFN-γ, IL-1β, IL-4, IL-18, and G-CSF were inversely associated with susceptibility to IVDD.10,36
OP, a prevalent systemic disorder, has been mechanistically associated with dysregulated inflammatory mediators as demonstrated by accumulating evidence.37 MR analyses revealed a protective association where elevated circulating levels of interleukin-8 (IL-8), macrophage inflammatory protein-1 beta (MIP-1β), and cutaneous T-cell attracting chemokine (CTACK) were inversely correlated with OP risk. Conversely, reduced concentrations of hepatocyte growth factor (HGF), interleukin-1 receptor antagonist (IL-1RA), interleukin-10 (IL-10), macrophage colony-stimulating factor (MCSF), and Macrophage Inflammatory Protein-1α (MIP-1α) exhibited negative correlations with OP pathogenesis, suggesting bidirectional immunomodulatory effects on bone remodeling homeostasis.13 MR analyses demonstrated that inflammatory bowel disease (IBD) causally contributes to genetic variations in interleukin-6 (IL-6) and interleukin-1 receptor antagonist (IL-1RA), with dysregulation of these cytokines significantly correlated with OP pathogenesis. Furthermore, elevated interleukin-1 beta (IL-1β) secretion levels were identified as a predictive biomarker for OP risk.38 Previous MR studies have demonstrated that Interferon-inducible T-cell α-chemoattractant (CXCL11) elevates the risk of OP.37 MR analyses demonstrated a causal association between inflammatory cytokines and BMD.39
Our MR analysis revealed that elevated secretion levels of CXCL6, S100A12, and neurotrophin-3 (NT3) increase the risk of IVDD, while decreased levels of DNER and sirtuin-2 (SIRT2) also elevate IVDD susceptibility. For OP, heightened levels of IL-7, S100A12, FLT3L, caspase-8, TNF, and IL-18 were associated with increased disease risk, whereas reduced levels of MCP-3, GDNF, MCP-2, and CXCL25 similarly contributed to higher OP incidence. Notably, S100A12 was identified as a shared positive risk factor influencing both IVDD and OP pathogenesis.
The Mechanistic Role of S100A12
S100A12, a ubiquitously expressed factor in humans, has been implicated in diverse pathological conditions including malignancies, cardiovascular diseases and inflammatory disorders. Previous studies employing immunohistochemical analyses have demonstrated that S100A12 serves as a diagnostic biomarker in papillary thyroid cancer,40 breast cancer41 and bladder cancer.42 Studies have demonstrated that S100A12 serves as a biomarker in atherosclerosis.43 In the context of inflammatory diseases, S100A12 exhibits ubiquitous expression across diverse cell types and modulates critical biological processes including cellular proliferation, apoptosis, and inflammatory responses through RAGE-dependent signaling cascades.44 Its expression in IBD further establishes S100A12 as a key biomarker for this disorder.45 Furthermore, research has found that S100A12 also plays a significant role in osteoarthritis. It is significantly upregulated in the cartilage, meniscus, and synovium of osteoarthritis patients.46 Simultaneously, S100A12 promotes macrophage M1 polarization through the NF-κB signaling pathway, thereby enhancing the secretion of inflammatory factors such as IL-1β. These secreted inflammatory factors damage chondrocytes, leading to increased S100A12 production and forming a vicious cycle that exacerbates osteoarthritis progression. S100A12 serves as a critical biomarker in juvenile idiopathic arthritis.47 However, the relevant effects of S100A12 have not been reported in IVDD and OP. We speculate whether S100A12 also affects the intracellular NF-κB signaling pathway, thereby causing alterations in IL-1β and TNF-α.
Based on above results, we used qRT-PCR assay and WB to detect the expression of S100A12 in human cell lines and human samples from patients. The observed increase in S100A12 levels in patients/cells with IVDD and OP, compared to normal samples, supports our conclusion.
However, while we have revealed the correlation between S100A12 and IVDD and OP, its underlying pathogenic mechanisms in these diseases remain unclear. As well, due to the limitations of the available databases, the population we discussed mainly revolved around Europeans, lacking samples of Asian people. Furthermore, supporting evidence from clinical studies is currently lacking. Additional investigations like mechanistic in vivo validation are warranted to elucidate the specific causal relationships and explore its intrinsic pathogenic role.
Conclusion
Based on our integrated research employing MR analysis to investigate inflammation factors exhibiting positive and negative correlations with both IVDD and OP, S100A12 was identified through screening as a pro-inflammatory factor positively associated with the pathogenesis of both conditions. Further investigations are needed to verify our results, these findings provide a new idea for the treatment of both IVDD and OP.
Data Sharing Statement
The GWAS summary statistics for IVDD and OP were derived from data released by FinnGen database. The GWAS summary statistics for circulating inflammatory cytokines were derived from data released by Zhao etal. The data used to provide support for the results of this study can be obtained from the corresponding authors.
Ethics Declaration
This study was approved by the Ethics Committee of Qilu Hospital (KYLL-2022(ZM)-136). Our study complies with the Declaration of Helsinki.
Acknowledgments
We acknowledge FinnGen database and Zhao etal for providing their platforms and contributors for uploading their meaningful datasets, and all the patients who gave consent to disclose their medical records.
Author Contributions
Zexin Wang: Software, Formal analysis, Methodology, Writing- original draft; Bin Chen: Software, Methodology, Investigation, Writing- review & editing; Zhenchuan Liu: Formal analysis, Methodology, Writing- original draft; Yuanqiang Zhang: Methodology, Project administration, Resources, Writing -review & editing and Writing- original draft; Leixiang Han: Formal analysis, Methodology, Supervision, Writing- original draft, Writing -review & editing. All authors have agreed on the final version for publication, agreed on the journal to which the paper was submitted, and have agree to be accountable for all aspects of the work. Zexin Wang and Bin Chen have contributed equally to this work.
Funding
This work was supported by grants from National Natural Science Foundation of China (82402758).
Disclosure
Zexin Wang, Bin Chen, Zhenchuan Liu, Yuanqiang Zhang, Leixiang Han declare that they have no conflict of interest.
References
1. Opal SM, DePalo VA. Anti-inflammatory cytokines. Chest. 2000;117:1162–15. doi:10.1378/chest.117.4.1162
2. Liu C, Chu D, Kalantar‐Zadeh K, et al. Cytokines: from clinical significance to quantification. Adv Sci. 2021;8:e2004433. doi:10.1002/advs.202004433
3. Wojdasiewicz P, Poniatowski LA, Szukiewicz D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediators Inflamm. 2014;2014:561459. doi:10.1155/2014/561459
4. Long JP, Prakash R, Edelkamp P, et al. Cytokine storms in COVID-19, hemophagocytic lymphohistiocytosis, and CAR-T therapy. JAMA Network Open. 2025;8:e253455. doi:10.1001/jamanetworkopen.2025.3455
5. Liu PL, Diao J-Y, Wang Q, et al. Cartilage damage pathological characteristics of diabetic neuropathic osteoarthropathy. Anal Cell Pathol. 2023;2023:7573165. doi:10.1155/2023/7573165
6. Nie J, Zhou L, Tian W, et al. Deep insight into cytokine storm: from pathogenesis to treatment. Signal Transduct Target Ther. 2025;10:112. doi:10.1038/s41392-025-02178-y
7. Loi F, Córdova LA, Pajarinen J, et al. Inflammation, fracture and bone repair. Bone. 2016;86:119–130. doi:10.1016/j.bone.2016.02.020
8. Xin J, Wang Y, Zheng Z, et al. Treatment of intervertebral disc degeneration. Orthop Surg. 2022;14:1271–1280. doi:10.1111/os.13254
9. Zhou L, Cai F, Zhu H, et al. Immune-defensive microspheres promote regeneration of the nucleus pulposus by targeted entrapment of the inflammatory cascade during intervertebral disc degeneration. Bioact Mater. 2024;37:132–152. doi:10.1016/j.bioactmat.2024.03.020
10. Xu T, Chen G, Li J, Zhang Y. Exploring causal correlations between inflammatory cytokines and intervertebral disc degeneration: a Mendelian randomization. JOR Spine. 2024;7:e1349. doi:10.1002/jsp2.1349
11. Wang Y, Che M, Xin J, et al. The role of IL-1beta and TNF-alpha in intervertebral disc degeneration. Biomed Pharmacother. 2020;131:110660. doi:10.1016/j.biopha.2020.110660
12. Anam AK, Insogna K. Update on Osteoporosis Screening and Management. Med Clin North Am. 2021;105:1117–1134. doi:10.1016/j.mcna.2021.05.016
13. Yi KJ, Kang RM, Zhang YY, Li Q. Causal relationship between circulating inflammatory factors and osteoporosis: a bidirectional Mendelian randomization study. Eur Rev Med Pharmacol Sci. 2024;28:2237–2249.
14. Mei J, Hu H, Ding H, et al. Investigating the causal relationship between ankylosing spondylitis and osteoporosis in the European population: a bidirectional Mendelian randomization study. Front Immunol. 2023;14:1163258. doi:10.3389/fimmu.2023.1163258
15. Liu G, Zhang H, Chen M, Chen W. Causal relationship between intervertebral disc degeneration and osteoporosis: a bidirectional two-sample Mendelian randomization study. Front Endocrinol. 2024;15:1298531. doi:10.3389/fendo.2024.1298531
16. Zhao J, Wang J, Xu H, et al. Intervertebral disk degeneration and bone mineral density: a bidirectional mendelian randomization study. Calcif Tissue Int. 2024;114:228–236. doi:10.1007/s00223-023-01165-1
17. Chong M, Sjaarda J, Pigeyre M, et al. Novel drug targets for ischemic stroke identified through mendelian randomization analysis of the blood proteome. Circulation. 2019;140:819–830. doi:10.1161/CIRCULATIONAHA.119.040180
18. Emdin CA, Khera AV, Kathiresan S. Mendelian Randomization. JAMA. 2017;318:1925–1926. doi:10.1001/jama.2017.17219
19. Kurki MI, Karjalainen J, Palta P, et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature. 2023;613:508–518. doi:10.1038/s41586-022-05473-8
20. Hemani G, Zheng J, Elsworth B, et al. The MR-Base platform supports systematic causal inference across the human phenome. Elife. 2018;7. doi:10.7554/eLife.34408
21. Burgess S, Thompson SG. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur J Epidemiol. 2017;32:377–389. doi:10.1007/s10654-017-0255-x
22. Bowden J, Davey Smith G, Burgess S. Mendelian randomization with invalid instruments: effect estimation and bias detection through Egger regression. Int J Epidemiol. 2015;44:512–525. doi:10.1093/ije/dyv080
23. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40:304–314. doi:10.1002/gepi.21965
24. Tan JS, Ren J-M, Fan L, et al. Genetic Predisposition of Anti-Cytomegalovirus Immunoglobulin G Levels and the Risk of 9 Cardiovascular Diseases. Front Cell Infect Microbiol. 2022;12:884298. doi:10.3389/fcimb.2022.884298
25. Verbanck M, Chen CY, Neale B, Do R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat Genet. 2018;50:693–698. doi:10.1038/s41588-018-0099-7
26. Kuzu S, Canli M, Valamur İ, et al. Effects of aerobic exercise in addition to core stabilization exercises on functional capacity, physical performance and fall risk in geriatric individuals with chronic non-specific low back pain. BMC Sports Sci Med Rehabil. 2025;17:218. doi:10.1186/s13102-025-01271-7
27. Li HZ, Zhang J-L, Yuan D-L, et al. Role of signaling pathways in age-related orthopedic diseases: focus on the fibroblast growth factor family. Mil Med Res. 2024;11:40. doi:10.1186/s40779-024-00544-5
28. Li Z, Cheng W, Gao K, et al. Pyroptosis: a spoiler of peaceful coexistence between cells in degenerative bone and joint diseases. J Adv Res. 2025;71:227–262. doi:10.1016/j.jare.2024.06.010
29. Wang J, Zhang Y, Cao J, et al. The role of autophagy in bone metabolism and clinical significance. Autophagy. 2023;19:2409–2427. doi:10.1080/15548627.2023.2186112
30. Jia Z, Zhang S, Li W. Harnessing stem cell-derived extracellular vesicles for the regeneration of degenerative bone conditions. Int J Nanomed. 2023;18:5561–5578. doi:10.2147/IJN.S424731
31. Lee KS, Lee J, Kim HK, et al. Extracellular vesicles from adipose tissue-derived stem cells alleviate osteoporosis through osteoprotegerin and miR-21-5p. J Extracell Vesicles. 2021;10:e12152. doi:10.1002/jev2.12152
32. Liu C, Fan L, Guan M, et al. A redox homeostasis modulatory hydrogel with GLRX3 + extracellular vesicles attenuates disc degeneration by suppressing nucleus pulposus cell senescence. ACS Nano. 2023;17:13441–13460. doi:10.1021/acsnano.3c01713
33. Charles JF, Nakamura MC. Bone and the innate immune system. Curr Osteoporos Rep. 2014;12:1–8. doi:10.1007/s11914-014-0195-2
34. Armas LA, Recker RR. Pathophysiology of osteoporosis: new mechanistic insights. Endocrinol Metab Clin North Am. 2012;41:475–486. doi:10.1016/j.ecl.2012.04.006
35. Jiang N, Wang Q, Hu Z, Tian X. CLEC11A-driven molecular mechanisms in intervertebral disc degeneration: a comprehensive multi-omics study. J Inflamm Res. 2025;18:1353–1375. doi:10.2147/JIR.S505296
36. Gong Y, Qiu J, Jiang T, et al. Maltol ameliorates intervertebral disc degeneration through inhibiting PI3K/AKT/NF-kappaB pathway and regulating NLRP3 inflammasome-mediated pyroptosis. Inflammopharmacology. 2023;31:369–384. doi:10.1007/s10787-022-01098-5
37. Zheng Q, Wang D, Lin R, et al. Effects of circulating inflammatory proteins on osteoporosis and fractures: evidence from genetic correlation and Mendelian randomization study. Front Endocrinol. 2024;15:1386556. doi:10.3389/fendo.2024.1386556
38. Xu D, Chen Y, Gao X, et al. The genetically predicted causal relationship of inflammatory bowel disease with bone mineral density and osteoporosis: evidence from two-sample Mendelian randomization. Front Immunol. 2023;14:1148107. doi:10.3389/fimmu.2023.1148107
39. Wu F, Huang Y, Hu J, Shao Z. Mendelian randomization study of inflammatory bowel disease and bone mineral density. BMC Med. 2020;18:312. doi:10.1186/s12916-020-01778-5
40. Wang X, Sun Z, Tian W, et al. S100A12 is a promising biomarker in papillary thyroid cancer. Sci Rep. 2020;10:1724. doi:10.1038/s41598-020-58534-1
41. Gunaldi M, Okuturlar Y, Gedikbasi A, et al. Diagnostic importance of S100A9 and S100A12 in breast cancer. Biomed Pharmacother. 2015;76:52–56. doi:10.1016/j.biopha.2015.10.029
42. Liu Z, Pu X. Orosomucoid 1 interacts with S100A12 and activates ERK signalling to expedite the advancement of bladder cancer. Cell Adh Migr. 2025;19:1–11. doi:10.1080/19336918.2024.2434209
43. Nazari A, Khorramdelazad H, Hassanshahi G, et al. S100A12 in renal and cardiovascular diseases. Life Sci. 2017;191:253–258. doi:10.1016/j.lfs.2017.10.036
44. Xia P, Ji X, Yan L, et al. Roles of S100A8, S100A9 and S100A12 in infection, inflammation and immunity. Immunology. 2024;171:365–376. doi:10.1111/imm.13722
45. van de Logt F, Day AS. S100A12: a noninvasive marker of inflammation in inflammatory bowel disease. J Dig Dis. 2013;14:62–67. doi:10.1111/1751-2980.12012
46. Zhang Y, Li Z, Chen C, et al. S100A12 is involved in the pathology of osteoarthritis by promoting M1 macrophage polarization via the NF-kappaB pathway. Connect Tissue Res. 2024;65:133–145. doi:10.1080/03008207.2024.2310852
47. Sharma TK, Saxena S, Arora P, Sharma AK. S100A12 protein levels in Juvenile Idiopathic Arthritis (JIA): a systematic review and meta-analysis. Clin Lab. 2025;71. doi:10.7754/Clin.Lab.2024.241041
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.