Back to Journals » OncoTargets and Therapy » Volume 13
Role of miRNA-424 in Cancers
Authors Li S ![]() , Wu Y
, Wu Y ![]() , Zhang J, Sun H, Wang X
, Zhang J, Sun H, Wang X
Received 18 June 2020
Accepted for publication 4 September 2020
Published 28 September 2020 Volume 2020:13 Pages 9611—9622
DOI https://doi.org/10.2147/OTT.S266541
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr William C. Cho
Shulin Li,1 Yuqi Wu,1 Jiawei Zhang,1 Hao Sun,2 Xiangwei Wang1
1Department of Urology & Carson International Cancer Center, Shenzhen University General Hospital & Shenzhen University Clinical Medical Academy Center, Shenzhen University, Shenzhen 518000, People’s Republic of China; 2Department of Urology, Shenzhen Second People’s Hospital & the First Affiliated Hospital of Shenzhen University, Shenzhen 518000, People’s Republic of China
Correspondence: Xiangwei Wang
Department of Urology & Carson International Cancer Center, Shenzhen University General Hospital & Shenzhen University Clinical Medical Academy Center, Shenzhen University, No. 1098, Xueyuan Road, Shenzhen University City, Nanshan District, Shenzhen 518000, People’s Republic of China
Tel +86 17796349157
Fax +86 075521839000
Email [email protected]
Abstract: microRNA (miRNA) is an important part of non-coding RNA that regulates gene expression at a posttranscriptional level. miRNA has gained increasing interest in recent years, both in research and clinical fields. miRNAs have been found to play an important role in various diseases, particularly cancer. Aberrant miR-424 expression is found in several tumors where they can function as either oncogenes or tumor-suppressor genes. Meanwhile, miR-424 is also affected by the reorganization of many other non-coding RNAs such as lncRNA and cirRNA. Several studies have found that miR-424 participates in proliferation, differentiation, apoptosis, invasion, angiogenesis, and drug resistance, and plays an important role in the tumorigenesis and progression of tumors. This review will focus on the recent progress of research on miR-424 in tumors.
Keywords: miRNAs, miR-424, tumor, regulation, carcinogenesis, cancer progression
Introduction
MicroRNA (miRNA) has been the subject of many studies since its discovery in 1993.1 miRNA is an important class of non-coding RNA that function as post-transcriptional regulators of gene expression in cells by repressing and fine-tuning protein output. Although miRNA accounts for only 1% of the human genome, it regulates one-third of the gene expression in humans.2–4 miRNAs are ubiquitous in eukaryotes, and are highly conservative, time-series, and tissue-cell specific. miRNAs are composed of a large group of non-coding small RNAs (approximately 21–23nt).5 miRNA formation comprises three main steps: transcription, maturation, and functional complex assembly. The gene encoding miRNA in the nucleus is first transcribed under the catalysis of RNA polymerase II to form a primary transcript with a cap structure (5MGpppG) and a poly-A tail (PolyA) of several hundred nucleotides in length.6 The primary transcript is further processed into a single precursor pre-miRNA, with a stem-loop structure containing only 70–100 nt under the action of the RNase family enzyme Drosha. The pre-miRNA is then transported to the cytoplasm by HASTY (HST), a homolog of the export protein Exportin-5.7–9 In conjunction with the RNaseIII family enzyme Dicer, the pre-miRNA is processed to form double-stranded miRNA, which subsequently forms double strand mature miRNA.10 Mature miRNAs combine with a ribonucleoprotein complex similar to RISC (RNA-induced silencing complex) to form miRNP, which recognizes target genes.9,11 After miRNA is loaded into the RISC complex, it functions in the regulation of gene expression. miRNA recognizes binding sites on the mRNA 3ʹUTR through its seed sequence (nucleotides 2–8 at the 5 ‘end), carries RISC, and inhibits protein expression. The three main functions of miRNA are transcriptional repression, and mRNA cleavage or degradation.12–14 Existing studies have found that approximately 30% of human protein expression is regulated by miRNAs.15 By interacting with the 3ʹUTR, miRNA can simultaneously regulate the expression of many genes, thereby regulating signaling pathways at multiple levels.16,17
Evidence increasingly suggests that miR-424 plays an important role in different types of diseases, including sperm damage,18 psoriasis,19 amyotrophic lateral sclerosis (ALS),20 insulin resistance,21 and acute respiratory distress syndrome (ARDS).22 However, the research on miR-424 is more abundant in cases of tumors involving early diagnosis, development, treatment, prognosis. This review focuses on the role of miR-424 in tumor progression.
Dual Regulatory Function of miRNA-424
The expression level of miR-424 in tumors can be either high or low, highlighting its multiple roles. Due to differences in expression, it is possible to diagnose disease and predict prognosis by detecting the amount of miRNA-424 in the blood or other body fluids.
Multilateral statistical evaluations have repeatedly confirmed the combination of highly specific biomarkers composed of four urine microRNAs (miR-424, miR-423, miR-660, and let7-i) as indicators to distinguish breast cancer patients from healthy controls, with sensitivity of 98.6% and specificity of 100%. This is of great significance for the non-invasive auxiliary diagnosis of breast cancer.23 Meanwhile, currently basing on the PSA to diagnosis prostate cancer is limited. A previous study identified that a low expression level of miR-424-3p was associated with the invasive phenotype and recurrence of prostate cancer.24,25 It can be detected by miR-424 combined with PSA value and prostate volume to assist in the diagnosis of prostate cancer in patients with elevated PSA.26
In addition to studies on endocrine organs, the association between miRNAs and digestive system tumors has also been explored. The overall survival rate of pancreatic adenocarcinoma (PAAD) is associated with miRNAs including miR-424, which may have the potential to become a prognostic biomarker for PAAD.27 Several studies have found that miR-424-5p is a tumor suppressor miRNA in hepatocellular carcinoma (HCC), and miR-424-5p is closely related to cell proliferation and invasion.28–30 Furthermore, it has been shown that miR-424 can predict the recurrence of liver cancer after liver transplantation, and that low expression of miR-424 is an independent prognostic factor of relapse, primarily through promoting the migration, invasion, and proliferation of HCC cells.31 In the respiratory system, it has been shown that high expression of miR-424 indicates a worse prognosis for patients with non-small cell lung cancer.32
In summary, a large number of studies have found that the expression level of miRNA-424 in different tumors has significance in the diagnosis and prognosis of tumors.
Function of miR-424 and Roles in Tumor Progression
MicroRNA regulates gene expression at the post-transcription level and is involved in regulating several key processes including cell proliferation, apoptosis, stem cell differentiation, invasion and metastasis, and other physiological and pathological processes. As mentioned above, the expression of miR-424 in tumors is closely related to the occurrence and development of disease. At the same time, a large number of studies have found that by changing the expression of miR-424 in tumors to regulate different target genes, cell function can also be altered.
Multiplication, Invasion, and Migration
Tumor proliferation, invasion and migration are complex processes with multiple factors and steps that involve oncogenes, tumor suppressor genes, signal transduction genes, and numerous cytokines and regulatory factors. Studies have found that miRNAs play different functions in different diseases. For instance, in the nervous system, glioma is one of the most deadly malignant tumors, in which miR-424 plays a role in suppressing cancer in glioma cells and is down-regulated by DNA methylation.33 Further research found that miR-424 can inhibit cell migration and epithelial-mesenchymal transition (EMT) by targeting KIF23 in glioma.34 Moreover, in the reproductive system, based on the exact relationship between HPV infection and cervical cancer, studies have found that in HPV (+) cervical cancer cells, CHK1 and Wee1, the two targets of miR-424, are suppressed. These factors have important roles in DNA damage repair,35 and further research found that regulating CHK1 can affect cervical cancer cell migration and invasion, apoptosis, and cell cycle changes. Furthermore, miR424-5p can target KDM5B through the Notch signaling pathway to influence cervical cancer cell proliferation.36,37 In endometrial cancer, miR-424 can inhibit cell proliferation by targeting E2F7, increase cell apoptosis, and cause cell cycle arrest38 Another study found that miR-424 can inhibit cell invasion and sphere formation by inhibiting MMSET, as well as inhibiting cell migration, invasion, and EMT by downregulating doublecortin-like kinase 1 (DCLK1). miR-424 can also target G protein-coupled estrogen receptor −1 (GPER-1), which in turn affects the PI3K/AKT signaling pathway to regulate cell proliferation.39–42
In summary, a large number of studies on the female reproductive system have indicated that miR-424 is involved in regulating the malignant biological behavior of tumors by regulating the expression of target genes, which may become an important marker for tumor diagnosis and prognosis, and may even have potential as an anti-tumor adjuvant therapy.
In nasopharyngeal carcinoma, miR-424-5p acts an oncogene that promotes the aggressive progression of laryngosquamous cell carcinoma (LSCC), and a high miR-424-5p level is significantly associated with poor differentiation, advanced tumors, and cervical lymph node metastasis.43 In oral squamous cell carcinoma, it has been found that miR-424-5p is involved in cell migration and invasion by negatively regulating the expression of SOCS2.44 Meanwhile, studies in a variety of digestive system tumors have found that miR-424 can either promote or suppress cancer. miR-424-5p is a carcinogen in colorectal cancer (CRC) that directly promotes proliferation and metastasis by targeting SCN4B, CCNE1, AKT3, and Rictor.45–48 In the field of hepatobiliary carcinoma, it has been found that miR-424-5p can inhibit the metastasis and invasion of intrahepatic cholangiocarcinoma by targeting ARK5. Moreover, in liver cancer, miR-424 can inhibit the proliferation and invasion of tumors by targeting TRIM29, YAP1, and c-Myb, while miR-424-5p can also target ICAT and inhibit tumor progression by reversing the EMT process of primary liver cancer.28,49–52 Another group has found that miR-424 inhibits tumor growth by inhibiting the Akt3/E2F3 axis in liver cancer tissues.53 Other studies have found that in hepatoma cells, the transcription of miR-424 and miR-503 increases after thyroid hormone binds to its receptor, further confirming its role in promoting tumor proliferation and invasion.54 miR-424 targets TGFBR3, thereby promoting EMT and migration of tongue squamous cell carcinoma (TSCC).55 miR-424-5p is involved in the invasion and metastasis of esophageal squamous cell carcinoma (ESCC) through the SMAD7 pathway; therefore, the upregulation of miR-424-5p may reduce the invasion and metastasis of the tumor.56 miR-424-5p affects the ERK1/2 signaling pathway by negatively regulating SOCS6 in pancreatic cancer, thereby increasing proliferation, migration, and invasion, and inhibiting apoptosis.57
There are relatively few studies on miR-424 in the urinary system. In prostate cancer, a study has found that low expression of miR-424-3p is closely related to increased invasiveness.24 Moreover, the expression level of miR-424 was shown to significantly correlate with aggressive tumor growth, advanced clinical stage, and poor prognosis of bladder cancer.58 In kidney cancer, miR-424 and miR-381 jointly target WEE1, up-regulate the activity of Cdc2 to synergistically inhibit the proliferation and survival of renal cancer cells, and participate in tumor-related processes.59 In basal-like breast cancer, a study has found that miR-424-5p regulates tumor cell proliferation, migration, and invasion by targeting doublecortin like kinase 1 (DCLK1).60 In the respiratory system, non-small cell lung cancer accounts for approximately 80% of all lung cancer cases. In the past few decades, the average survival time of patients diagnosed with non-small cell lung cancer (NSCLC) is less than 6 months.61 Another study found that miR-424 promotes the progression and metastasis of non-small cell lung cancer by regulating the tumor suppressor gene TNFAIP1.62 miR-424 can inhibit the proliferation and invasion of NSCLC by inhibiting the expression of E2F6.63 In the motor system, a study group demonstrated that miR-424 inhibits osteosarcoma cell migration and invasion by targeting fatty acid synthase.64
Drug Resistance and Radio-Sensitivity of Radiotherapy
Drug resistance is the main obstacle to the long-term effectiveness of cancer treatment. Drug resistance is mainly due to the generation of drug-resistant cells in the tumor tissue, which in turn leads to chemotherapeutic drugs being unable to exert a full anti-cancer effect. However, even if most of the tumor cells are killed, this small population of drug-resistant tumor cells will continue to proliferate, ultimately causing cancer relapse. In summary, following the emergence of drug resistance, the efficacy of chemotherapy will gradually decline, and eventually malignant tumors will no longer be treatable. In the reproductive system, studies have found that high expression of miR-424 and miR-503 is closely related to chemoresistance and tumor progression of ovarian cancer.65 miR-424-3p increases the sensitivity of the ovarian cancer cells SK-OV-3 and TOV-21G to cisplatin by reducing the expression of galectin-3 (anti-apoptotic protein); these results indicate that miR-424-3p may be an appropriate target to recover the sensitivity to chemotherapy drugs.66
Studies have found that the sensitivity of the commonly used chemotherapy drug, docetaxel, is affected by miR-424.67 Further research confirmed that miR-424 is involved in the regulation of PD-L1/PD-1 and CD80/CTLA-4 in chemotherapy-resistant ovarian cancer.68 In the endocrine system, miR-424-5p inactivates Hippo signaling, thereby promoting anoikis drug resistance and lung metastasis. Therefore, miR-424-5p has the potential to become a therapeutic target for lung metastases from thyroid cancer.69 A study in hormone receptor-positive metastatic breast cancer found that miR-424-5p can be used as a potential indicator of the sensitivity response to dovitinib.70 Furthermore, the deletion of miR-424 (322)/503 can promote breast cancer resistance by up-regulating its two targets: BCL-2 and insulin-like growth factor-1 receptor (IGF1R).71 In the digestive system, preliminary verification based on the GEO database and human gastric cancer tissues showed that miR-424 regulates gastric cancer cisplatin resistance by targeting SMURF1.72 In non-small cell lung cancer, studies have confirmed that the deletion of miR-424-3p is involved in chemotherapy resistance by targeting YAP1.73 In addition to chemotherapy resistance, the decline of tumor sensitivity to radiotherapy after long-term treatment has also attracted increasing attention. Studies have found that miR-424 can promote IR-induced DNA damage, apoptosis, and G2/M cell cycle arrest through APTX, thereby enhancing the radiation sensitivity of radiation-resistant cervical cancer cells.74
Cell Cycle
The cell cycle is the time required from the end of cell division to the end of the next cell division. The cell cycle is divided into four stages: G1 phase (gap1), which refers to the gap time before DNA replication is due to complete from mitosis; S phase (synthesis phase), which refers to the period of DNA replication; G2 phase (gap2), which refers to the completion of DNA replication but before the onset of mitosis; M phase, also known as D phase (mitosis or division), begins to end cell division. Dysregulation of the cell cycle is one of the main mechanisms of tumorigenesis.
A study identified that miR-424 in the digestive system regulates the cell cycle of esophageal squamous cell carcinoma (ESCC) to promote proliferation, and can be used as a new prognostic marker and an effective therapeutic target.75 In gastric cancer, miR-424-5p regulates the TGF-β signaling pathway and cyclin (c-Myc, CDK2, CDK4, CDK6) through SMAD3, where it has roles in regulating the cell cycle.76 Meanwhile, in the motion system, miR-424 suppressed the development of osteosarcoma by targeting the expression of cyclin cell A2.77 In epithelial ovarian cancer, miR-424-5p suppresses the expression of CCNE1, and blocks cells in the G1/G0 phase through the E2F1-pRb pathway.41 In conclusion, some studies have shown that miR-424 plays an important role in regulating the cell cycle, and can regulate the expression of miR-424 to inhibit the occurrence and development of tumors.
Stem Cell Properties
Cancer stem cells (CSCs) represent a subset of the cancer cell population that possess self-renewal ability and account for cancer initiation, progression, metastasis, and recurrence. It has been shown that in breast cancer cells, hyperglycemia can excessively activate tumor stem cells through the miR-424/cdc42/prdm14 axis, and enhance tumor invasion.78 Similarly, in another common endocrine-related tumor, prostate cancer, miR-424 targets E3 ubiquitin ligase COP1 and recognizes STAT3 as a key substrate of COP1, promoting prostate epithelial cell tumorigenicity and cancer stem cell-like properties.79
Relationship with Viruses
Various viruses have been shown to be closely related to the occurrence of tumors. EBV was the first human virus to be shown to be directly related to canceration, which has been shown to be closely related to various malignant tumors.80 The causal relationship between human papillomavirus (HPV) and cervical cancer is well established in biology and epidemiology. HPV-16 and −18 infections account for approximately 70% of cervical cancer cases.81 A previous study reported that miR-424 plays a key role in the regulation of HPV replication. Furthermore, HPV E6 and E7 proteins inhibit miR-424 levels, control CHK1, and have been shown to play a central role in viral replication.37 Furthermore, the interaction between HPV16 and E7CUL2 recruits CUL2 and activates the regulatory loops between CUL2, E2F1, and miR-424, which results in persistent CUL2 overexpression, and subsequent promotion of HPV16-induced cervical cancer development.82
In conclusion, existing evidence indicates that miR-424 plays an important role in various stages of tumorigenesis. Moreover, analysis of miR-424 expression can help to diagnose diseases, and has the potential to become an important molecule in tumor diagnosis and prognosis. In addition, in the future, the expression of miR-424 may be adjusted to regulate the occurrence and development of tumors and limit tumor progression. Taken together, these studies have suggested that miR-424 performs different functions in various cancers (Table 1).
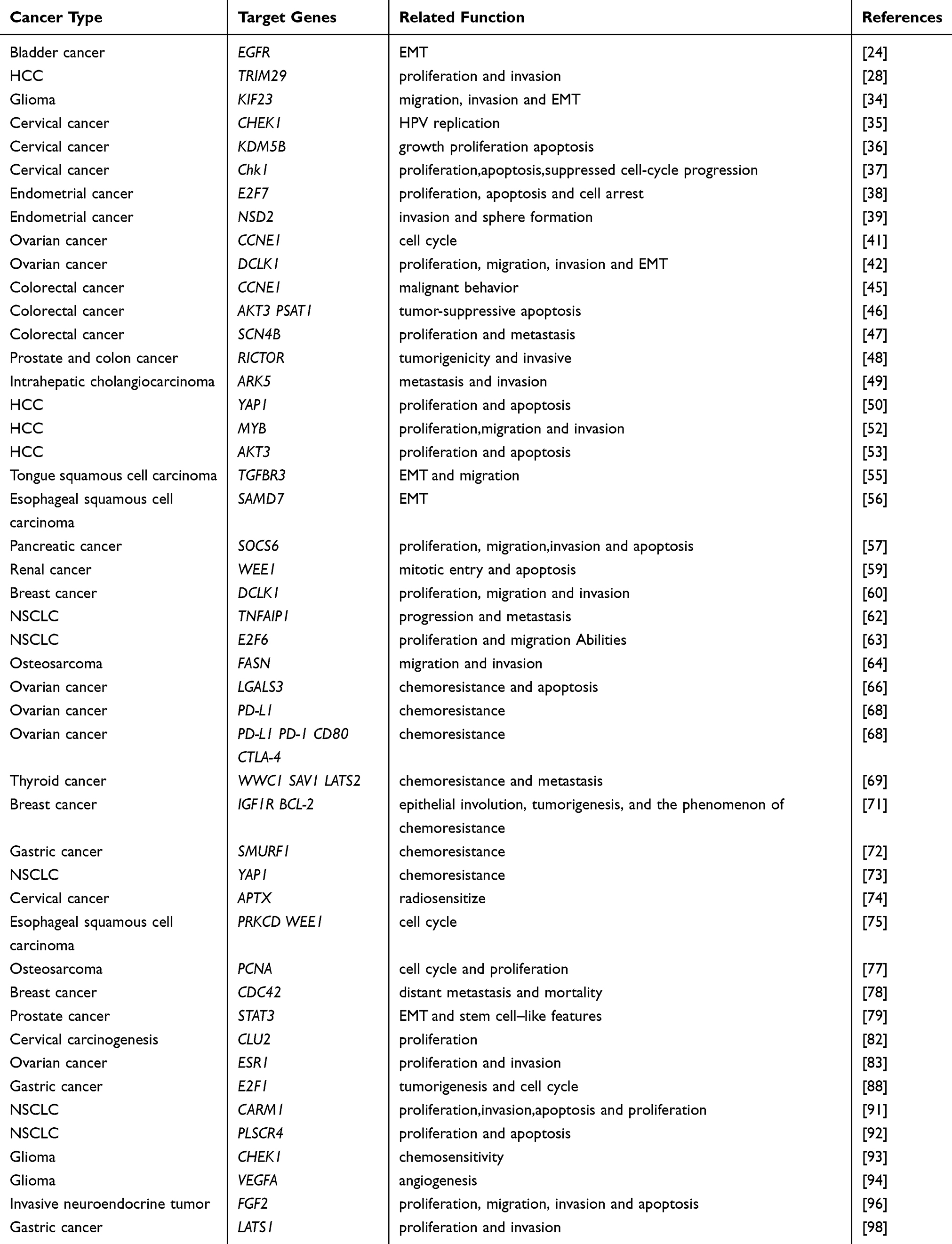 |
Table 1 The Function of the Target Gene of miR-424 in Difference Cancers |
Molecules That Regulate the Expression of miR-424 in Tumors
LncRNA
Accumulating evidence has shown that a variety of lncRNAs and cirRNAs are involved in the occurrence and development of tumors by regulating mir-424. For example, in the reproductive system, estrogen receptor 1 (ESR1) plays an important role in the pathological process of ovarian cancer. ESR1-induced upregulation of LINC00511 may be caused by sponged miR-424-5p and miR-370- 5p, which in turn promotes the proliferation and invasion of ovarian cancer CAOV3 cells.83 Moreover, it has been shown that Lnc-SNHG12 negatively regulates miR-424-5p to promote cancer.84 Another study found that Lnc-CCAT2 negatively targets miR-424 as a competitive endogenous RNA (ceRNA) or sponge for ovarian cancer, and represents a new diagnostic marker and therapeutic target.85 Tumor promoting effects of Lnc-PVT1 were found in cervical cancer, as competitive endogenous RNA (ceRNA) or molecular sponge negatively regulates miR-424.86 Furthermore, LncRNA BLACAT1 expression is increased in cervical cancer (CSCC), while BLACAT1 overexpression may promote tumor progression by down-regulating miR-424.87 Another study found that the tumor-promoting Small nucleolar RNA host gene 12 (SNHG12) acts as a molecular sponge of miR-424-5p and has a negative regulatory effect.84 With regards to the digestive system, Lnc-RNA NNT-AS1 can affect the tumorigenesis and cell cycle of gastric cancer cells by binding to miR-424/E2F1.88 Moreover, in hepatocellular carcinoma, LncRNA-LINC00511 plays an important role in the malignant progression of HCC by regulating miR-424.89 Another study has found that Lnc-RNA DLX6-AS1 increases the expression of WEE1 by targeting miR-424-5p, which in turn promotes hepatocellular carcinoma.90 In the respiratory system, non-small cell lung cancer accounts for approximately 80% of lung cancer subclasses, and is the disease with the highest cancer-related mortality in both men and women. A research team found that the miR-424-5p/lncRNA PVT1/CARM1 axis can influence non-small cell lung cancer. Furthermore, radiotherapy sensitivity of small cell lung carcinoma, overexpression of miR-424-5p, or inhibition of lncRNA PVT1 can increase the radiotherapy sensitivity of NSCLC.91 Lnc-RNA LINC00641 was found to inhibit the progression of non-small cell lung cancer by sponging miR-424-5p and up-regulating PLSCR4. Further rescue experiments showed that miR-424-5p and PLSCR4 can reverse the cell process mediated by LINC00641. In conclusion, LINC00641 exerts a tumor suppressive effect in NSCLC through miR-424-5p.92
In the central nervous system, a previous study demonstrated that lncRNA-CCAT2 enhances the chemoresistance of glioma cells through the lncRNA-CCAT2/miR-424/Chk1 axis, and promotes the migration of glioma cells. As such, the curative effect of chemotherapy for glioma can be improved by inhibiting this axis.93 Recent studies have found that lncRNA-CCAT2 also can down-regulate miR-424 to promote VEGFA expression, induce PI3K/AKT signaling, and promote cell proliferation and endothelial angiogenesis.94 It was also found that down-regulation of lncRNA-PVT1 inhibits the progression of glioma by up-regulating the expression of miR-424.95 In addition, in invasive pituitary neuroendocrine tumors, LncRNA-XIST competitively combines with miR-424-5p to increase the expression of bFGF to promote cell proliferation, migration, invasion, and inhibit cell cycle arrest and apoptosis.96 In the circulatory system, a research group used tumor xenograft experiments to show that LncRNA-MALAT1 inhibits miR-424 and activates the MEKK3-mediated IKK/NF-kappaB pathway to promote the development of hemangioma. This suggests that LncRNA-MALAT1, miR-424, and MEKK3 can be used as potential targets for the improvement of IH treatment efficiency.97
CircRNA
In addition to LncRNA, part of CircRNA is also involved in the regulation of microRNA. Circ-LARP4 inhibits the proliferation and invasion of gastric cancer cells by sponging miR-424-5p and regulating the expression of LATS1. The increased expression of miR-424 or the decreased expression of LATS1, is closely related to the pathological stage and poor prognosis of GC patients.98 Studies have also found that the high expression of Circ-LARP4 is related to a shorted perioperative period and prolonged survival of osteosarcoma, as well as increased chemical sensitivity to cisplatin and doxorubicin through cavernous miR-424.99 In short, an increasing number of circ-RNAs have been shown to be involved in the regulation of miR-424, and their function needs to be explored further.
Taken together, these studies have suggested that miR-424 regulated by different non-coding RNA in various cancers (Table 2).
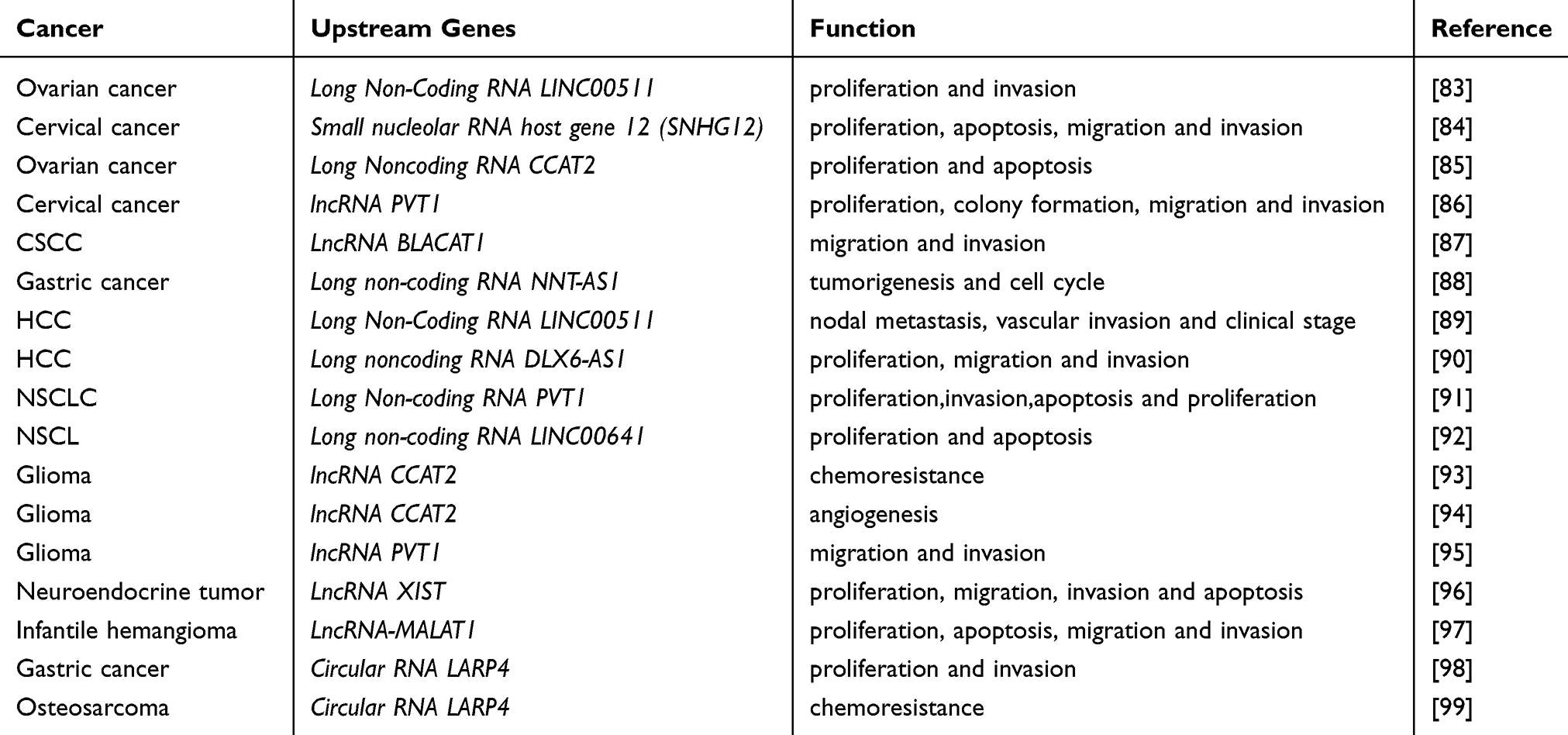 |
Table 2 The Function of the Upstream Genes of miR-424 in Difference Cancers |
Bioinformatics Analysis of miR-424
We performed KEGG pathway analysis of miR-424 using Webgestalt (http://bioinfo.vanderbilt.edu/webgestalt/) to predict the possible downstream pathway of miR-424 target genes, screening is performed with P < 0.05 as the critical value. After KEGG analysis, the 20 most relevant routes were selected, 15 of which were found to have significant P-values. The 20 KEGG pathways with the highest concentration are shown in Figure 1A. miR-424 target genes were mainly enriched in the Hippo signaling pathway, pancreatic cancer, EGFR tyrosine kinase inhibitor resistance, AGE-RAGE signaling pathways in diabetic complications, Hepatitis B, proteoglycans in cancer, cell cycle, and several cancer-related pathways. These data indicated that miR-424-5p might play important roles in the malignant behavior of cancer. Among them, Hippo signaling pathway affected the genesis and development of tumors by regulating cell proliferation, apoptosis and stem cell self-renewal ability.100,101 In addition, miR-424 regulates tumor proliferation and invasion processes by regulating EGFR. A research has revealed a wide range of role of EGFR via adding EGF to cultured HeLa cells to activate EGFR resulting in phosphorylation of 6600 sites in 2244 proteins.102 After ligand binding, the intracellular tyrosine sites of EGFR are phosphorylated, which consequently match to different signal transduction proteins to initiate different signal pathways.103 The intricate signal transduction network play roles in tumor cell proliferation, metastasis and anti-apoptosis.104 Therefore, overexpression or mutation of EGFR is closely related to the pathogenesis of cancer and the poor prognosis of patients.
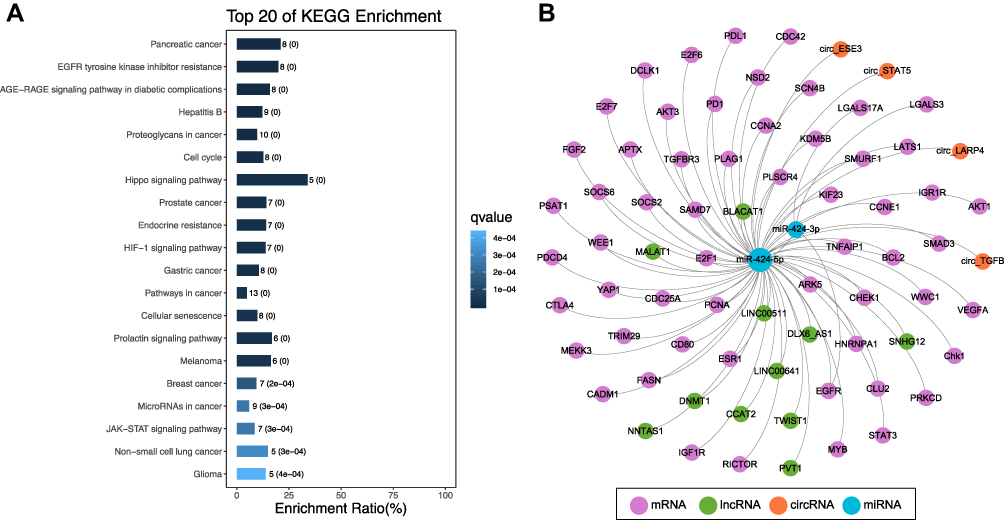 |
Figure 1 (A) KEGG analysis showing the number of transcripts mapped to the function involved in the target genes of miR-424. The 20 KEGG pathways most strongly enriched by functional-related gene groups. (B) The competing endogenous RNA network of cirRNA/lncRNA-microRNA-mRNA. The pink dots represent target genes of miR-424. The green dots represents lncRNA, while the orange dots diamonds represent cirRNA. The blue dots represents the miR-424-5p or miR-424-3p. |
In sum, miR-424 participates in downstream target genes through regulation, and then plays an important role in the malignant behavior of cancer. If we can further clarify the relationship between miR-424 and downstream pathways and the mechanism of action between pathways, we will clarify the potential molecular mechanism of miR-424 involved in regulating the occurrence and development of malignant tumors. It will be beneficial to tumor diagnosis and treatment. Next, we carried out the construction of ce-network (Figure 1B), combined with the above review and network diagram, we clearly found that miRNAs in organisms not only play a regulatory role on downstream target genes at the post-transcriptional level, but also their expression and function are affected by multiple signal transduction pathways and transcription factors. The regulatory relationship between them is related to each other through a complex linear relationship of point-to-multipoint divergence or multipoint-to-point convergence. The body constitutes a network-type deployment control to play its corresponding physiological or pathological role.
Discussion
In recent years, the role of miRNA in the occurrence, development, and diagnosis of diseases has attracted considerable attention, and evidence for the association between miRNA and the occurrence and development of tumors is only increasing. In line with this, miR-424 has been a hotspot in the field of cancer research, and evidence shows that it is closely related to the biological processes of tumor cell proliferation, migration, invasion, and apoptosis, as well as tumor occurrence, development, diagnosis and treatment, chemosensitivity and recurrence monitoring, prognosis assessment, and viral infections. We employed the use of bioinformatics to show the KEGG pathway of miR-424 based on the reported mRNA reported in the literature, as well as the competing endogenous RNA network of cirRNA/lncRNA-microRNA-mRNA. miR-424 can act as either a cancer-promoting factor or a tumor suppressor. miR-424 regulates multiple mRNAs and affects multiple signal transduction pathways, while the target genes regulated by miRNA in different microenvironments are also different. The multiple target gene pathways are related to each other, and the relationship is extremely complex. Therefore, further studies are needed to clarify the precise function of miR-424 in tumors. In order to reduce the side effects of miRNA binding to other targets during cancer treatment, it is necessary to develop highly specific miRNA. In addition, to ensure that the drug can function safely and effectively after reaching the body, the design of the drug carrier needs to be continuously improved and perfected. In view of the potential value of miR-424 in the clinical application of cancer, future applied research needs to further explore the application value of miR-424 in the prevention, diagnosis, prognosis, and targeted therapy of malignant tumors. This will ultimately lead to more promising treatment opportunities for accurate diagnosis and safe and effective options for the long-term treatment of patients with tumors.
Acknowledgments
The study was sponsored by the Science and Technology Innovation Commission Foundation of Shenzhen (Grant No. JCYJ20190808141013454 and JCYJ20180305124827261)
Disclosure
The authors report no conflicts of interest in this work.
References
1. Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854.
2. Pillai RS, Bhattacharyya SN, Filipowicz WJT. Repression of protein synthesis by miRNAs: how many mechanisms? Trend Cell Biol. 2007;17:118–126.
3. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136:215–233.
4. Garzon R, Calin GA, Croce CMJ. MicroRNAs in cancer. Annu Rev Med. 2009;60:167–179.
5. Carthew RW, Sontheimer EJ. Origins and mechanisms of miRNAs and siRNAs. Cell. 2009;136:642–655. doi:10.1016/j.cell.2009.01.035
6. Lee Y, Kim M, Han J, et al. MicroRNA genes are transcribed by RNA polymerase II. EMBO J. 2004;23:4051–4060. doi:10.1038/sj.emboj.7600385
7. Lee Y, Ahn C, Han J, et al. The nuclear RNase III Drosha initiates microRNA processing. Nature. 2003;425:415–419. doi:10.1038/nature01957
8. Carmell MA, Hannon GJ. RNase III enzymes and the initiation of gene silencing. Nat Struct Mol Biol. 2004;11:214–218. doi:10.1038/nsmb729
9. Gregory RI, Yan K-P, Amuthan G, et al. The Microprocessor complex mediates the genesis of microRNAs. Nature. 2004;432:235–240. doi:10.1038/nature03120
10. Bernstein E, Caudy AA, Hammond SM, Hannon GJ. Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature. 2001;409:363–366. doi:10.1038/35053110
11. Khvorova A, Reynolds A, Jayasena SD. Functional siRNAs and miRNAs exhibit strand bias. Cell. 2003;115:209–216. doi:10.1016/S0092-8674(03)00801-8
12. John B, Enright AJ, Aravin A, Tuschl T, Sander C, Marks DS. Human microRNA targets. PLoS Biol. 2004;2:e363.
13. Krek A, Grun D, Poy MN, et al. Combinatorial microRNA target predictions. Nat Genet. 2005;37:495–500. doi:10.1038/ng1536
14. Mogilyansky E, Rigoutsos I. The miR-17/92 cluster: a comprehensive update on its genomics, genetics, functions and increasingly important and numerous roles in health and disease. Cell Death Differ. 2013;20:1603–1614. doi:10.1038/cdd.2013.125
15. Lewis BP, Burge CB, Bartel DP. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets. Cell. 2005;120:15–20.
16. Baek D, Villén J, Shin C, Camargo FD, Gygi SP, Bartel DPJN. The impact of microRNAs on protein output. Nature. 2008;455:64–71.
17. Selbach M, Schwanhäusser B, Thierfelder N, Fang Z, Khanin R, Rajewsky N. Widespread changes in protein synthesis induced by microRNAs. Nature. 2008;455:58–63.
18. Zhao K, Chen Y, Yang R, et al. miR-424/322 is downregulated in the semen of patients with severe DNA damage and may regulate sperm DNA damage. Reprod Fertil Dev. 2016;28:1598–1607. doi:10.1071/RD15052
19. Tsuru Y, Jinnin M, Ichihara A, et al. miR‐424 levels in hair shaft are increased in psoriatic patients. J Dermatol. 2014;41:382–385.
20. de Andrade HM, de Albuquerque M, Avansini SH, et al. MicroRNAs-424 and 206 are potential prognostic markers in spinal onset amyotrophic lateral sclerosis. J Neurol Sci. 2016;368:19–24. doi:10.1016/j.jns.2016.06.046
21. Min K-H, Yang W-M, Lee WJB. Saturated fatty acids-induced miR-424–5p aggravates insulin resistance via targeting insulin receptor in hepatocytes. Biochem Biophys Res Commun. 2018;503:1587–1593. doi:10.1016/j.bbrc.2018.07.084
22. Cheng D, Zhu C, Liang Y, Xing Y, Shi C. MiR-424 overexpression protects alveolar epithelial cells from LPS-induced apoptosis and inflammation by targeting FGF2 via the NF-κB pathway. Life Sci. 2020;242:117213. doi:10.1016/j.lfs.2019.117213
23. Hirschfeld M, Rucker G, Weiss D, et al. Urinary exosomal microRNAs as potential non-invasive biomarkers in breast cancer detection. Mol Diagn Ther. 2020;24:215–232. doi:10.1007/s40291-020-00453-y
24. Richardsen E, Andersen S, Al-Saad S, et al. Low expression of miR-424-3p is highly correlated with clinical failure in prostate cancer. Sci Rep. 2019;9:10662. doi:10.1038/s41598-019-47234-0
25. Suer I, Guzel E, Karatas OF, Creighton CJ, Ittmann M, Ozen M. MicroRNAs as prognostic markers in prostate cancer. Prostate. 2019;79:265–271. doi:10.1002/pros.23731
26. Lyu J, Zhao L, Wang F, et al. Discovery and validation of serum microRNAs as early diagnostic biomarkers for prostate cancer in Chinese population. Biomed Res Int. 2019;2019:9306803. doi:10.1155/2019/9306803
27. Wang ZX, Deng TX, Ma Z. Identification of a 4-miRNA signature as a potential prognostic biomarker for pancreatic adenocarcinoma. J Cell Biochem. 2019;120:16416–16426. doi:10.1002/jcb.28601
28. Du H, Xu Q, Xiao S, et al. MicroRNA-424-5p acts as a potential biomarker and inhibits proliferation and invasion in hepatocellular carcinoma by targeting TRIM29. Life Sci. 2019;224:1–11. doi:10.1016/j.lfs.2019.03.028
29. Lu M, Kong X, Wang H, Huang G, Ye C, He Z. A novel microRNAs expression signature for hepatocellular carcinoma diagnosis and prognosis. Oncotarget. 2017;8:8775–8784. doi:10.18632/oncotarget.14452
30. Yao H, Liu X, Chen S, Xia W, Chen X. Decreased expression of serum miR-424 correlates with poor prognosis of patients with hepatocellular carcinoma. Int J Clin Exp Pathol. 2015;8:14830–14835.
31. Wu L, Yang F, Lin B, et al. MicroRNA-424 expression predicts tumor recurrence in patients with hepatocellular carcinoma following liver transplantation. Oncol Lett. 2018;15:9126–9132.
32. Wang Y, Lv Z, Fu J, Wang Z, Fan Z, Lei T. Endogenous microRNA-424 predicts clinical outcome and its inhibition acts as cancer suppressor in human non-small cell lung cancer. Biomed Pharmacother. 2017;89:208–214. doi:10.1016/j.biopha.2017.01.163
33. Jin C, Li M, Ouyang Y, Tan Z, Jiang Y. MiR-424 functions as a tumor suppressor in glioma cells and is down-regulated by DNA methylation. J Neurooncol. 2017;133:247–255. doi:10.1007/s11060-017-2438-4
34. Zhao C, Wang XB, Zhang YH, Zhou YM, Yin Q, Yao WC. MicroRNA-424 inhibits cell migration, invasion and epithelial-mesenchymal transition in human glioma by targeting KIF23 and functions as a novel prognostic predictor. Eur Rev Med Pharmacol Sci. 2018;22:6369–6378.
35. Hong S, Cheng S, Songock W, Bodily J, Laimins LA. Suppression of microRNA 424 levels by human papillomaviruses is necessary for differentiation-dependent genome amplification. J Virol. 2017;91.
36. Zhou Y, An Q, Guo RX, et al. miR424-5p functions as an anti-oncogene in cervical cancer cell growth by targeting KDM5B via the Notch signaling pathway. Life Sci. 2017;171:9–15. doi:10.1016/j.lfs.2017.01.006
37. Xu J, Li Y, Wang F, et al. Suppressed miR-424 expression via upregulation of target gene Chk1 contributes to the progression of cervical cancer. Oncogene. 2013;32:976–987. doi:10.1038/onc.2012.121
38. Li Q, Qiu XM, Li QH, et al. MicroRNA-424 may function as a tumor suppressor in endometrial carcinoma cells by targeting E2F7. Oncol Rep. 2015;33:2354–2360. doi:10.3892/or.2015.3812
39. Dong P, Xiong Y, Yue J, Hanley SJB, Watari H. miR-34a, miR-424 and miR-513 inhibit MMSET expression to repress endometrial cancer cell invasion and sphere formation. Oncotarget. 2018;9:23253–23263. doi:10.18632/oncotarget.25298
40. Lu Z, Nian Z, Jingjing Z, Tao L, Quan L. MicroRNA-424/E2F6 feedback loop modulates cell invasion, migration and EMT in endometrial carcinoma. Oncotarget. 2017;8:114281–114291. doi:10.18632/oncotarget.23218
41. Liu J, Gu Z, Tang Y, Hao J, Zhang C, Yang X. Tumour-suppressive microRNA-424-5p directly targets CCNE1 as potential prognostic markers in epithelial ovarian cancer. Cell Cycle. 2018;17:309–318. doi:10.1080/15384101.2017.1407894
42. Wu X, Ruan Y, Jiang H, Xu C. MicroRNA-424 inhibits cell migration, invasion, and epithelial mesenchymal transition by downregulating doublecortin-like kinase 1 in ovarian clear cell carcinoma. Int J Biochem Cell Biol. 2017;85:66–74. doi:10.1016/j.biocel.2017.01.020
43. Li Y, Liu J, Hu W, et al. miR-424-5p promotes proliferation, migration and invasion of laryngeal squamous cell carcinoma. Onco Targets Ther. 2019;12:10441–10453. doi:10.2147/OTT.S224325
44. Peng H-Y, Jiang -S-S, Hsiao J-R, et al. IL-8 induces miR-424-5p expression and modulates SOCS2/STAT5 signaling pathway in oral squamous cell carcinoma. Mol Oncol. 2016;10:895–909. doi:10.1016/j.molonc.2016.03.001
45. Jiang B, Wu D, Huang L, Fang H. miR-424-5p inhibited malignant behavior of colorectal cancer cells by targeting CCNE1. Panminerva Med. 2019.
46. Fang Y, Liang X, Xu J, Cai X. miR-424 targets AKT3 and PSAT1 and has a tumor-suppressive role in human colorectal cancer. Cancer Manag Res. 2018;10:6537–6547. doi:10.2147/CMAR.S185789
47. Dai W, Zhou J, Wang H, Zhang M, Yang X, Song W. miR-424-5p promotes the proliferation and metastasis of colorectal cancer by directly targeting SCN4B. Pathol Res Pract. 2020;216:152731. doi:10.1016/j.prp.2019.152731
48. Oneyama C, Kito Y, Asai R, et al. MiR-424/503-mediated Rictor upregulation promotes tumor progression. PLoS One. 2013;8:e80300. doi:10.1371/journal.pone.0080300
49. Wu J, Yang B, Zhang Y, et al. miR-424-5p represses the metastasis and invasion of intrahepatic cholangiocarcinoma by targeting ARK5. Int J Biol Sci. 2019;15:1591–1599. doi:10.7150/ijbs.34113
50. Piao L, Wang F, Wang Y, et al. miR-424-5p regulates hepatoma cell proliferation and apoptosis. Cancer Biother Radiopharm. 2019;34:196–202. doi:10.1089/cbr.2018.2625
51. Zhang Y, Li T, Guo P, et al. MiR-424-5p reversed epithelial-mesenchymal transition of anchorage-independent HCC cells by directly targeting ICAT and suppressed HCC progression. Sci Rep. 2014;4:6248. doi:10.1038/srep06248
52. Yu L, Ding GF, He C, Sun L, Jiang Y, Zhu L. MicroRNA-424 is down-regulated in hepatocellular carcinoma and suppresses cell migration and invasion through c-Myb. PLoS One. 2014;9:e91661. doi:10.1371/journal.pone.0091661
53. Yang H, Zheng W, Shuai X, et al. MicroRNA-424 inhibits Akt3/E2F3 axis and tumor growth in hepatocellular carcinoma. Oncotarget. 2015;6:27736–27750. doi:10.18632/oncotarget.4811
54. Ruiz-Llorente L, Ardila-Gonzalez S, Fanjul LF, Martinez-Iglesias O, Aranda A. microRNAs 424 and 503 are mediators of the anti-proliferative and anti-invasive action of the thyroid hormone receptor beta. Oncotarget. 2014;5:2918–2933. doi:10.18632/oncotarget.1577
55. Li D, Liu K, Li Z, Wang J, Wang X. miR-19a and miR-424 target TGFBR3 to promote epithelial-to-mesenchymal transition and migration of tongue squamous cell carcinoma cells. Cell Adh Migr. 2018;12:236–246. doi:10.1080/19336918.2017.1365992
56. Wang F, Wang J, Yang X, Chen D, Wang L. MiR-424-5p participates in esophageal squamous cell carcinoma invasion and metastasis via SMAD7 pathway mediated EMT. Diagn Pathol. 2016;11:88. doi:10.1186/s13000-016-0536-9
57. Wu K, Hu G, He X, et al. MicroRNA-424-5p suppresses the expression of SOCS6 in pancreatic cancer. Pathol Oncol Res. 2013;19:739–748. doi:10.1007/s12253-013-9637-x
58. Wu CT, Lin WY, Chang YH, Lin PY, Chen WC, Chen MF. DNMT1-dependent suppression of microRNA424 regulates tumor progression in human bladder cancer. Oncotarget. 2015;6:24119–24131. doi:10.18632/oncotarget.4431
59. Chen B, Duan L, Yin G, Tan J, Jiang X. Simultaneously expressed miR-424 and miR-381 synergistically suppress the proliferation and survival of renal cancer cells—Cdc2 activity is up-regulated by targeting WEE1. Clinics (Sao Paulo). 2013;68:825–833. doi:10.6061/clinics/2013(06)17
60. Wang J, Wang S, Zhou J, Qian Q. miR-424-5p regulates cell proliferation, migration and invasion by targeting doublecortin-like kinase 1 in basal-like breast cancer. Biomed Pharmacother. 2018;102:147–152.
61. Devesa SS, Bray F, Vizcaino AP, Parkin DM. International lung cancer trends by histologic type: male: female differences diminishing and adenocarcinoma rates rising. Int J Cancer. 2005;117:294–299. doi:10.1002/ijc.21183
62. Zhang M, Gao C, Yang Y, et al. MiR-424 promotes non-small cell lung cancer progression and metastasis through regulating the tumor suppressor gene TNFAIP1. Cell Physiol Biochem. 2017;42:211–221. doi:10.1159/000477314
63. Li H, Lan H, Zhang M, et al. [Effects of miR-424 on proliferation and migration abilities in non-small cell lung cancer A549 cells and its molecular mechanism]. Zhongguo Fei Ai Za Zhi. 2016;19:571–576. Chinese.
64. Long XH, Mao JH, Peng AF, Zhou Y, Huang SH, Liu ZL. Tumor suppressive microRNA-424 inhibits osteosarcoma cell migration and invasion via targeting fatty acid synthase. Exp Ther Med. 2013;5:1048–1052. doi:10.3892/etm.2013.959
65. Park YT, Jeong JY, Lee MJ, et al. MicroRNAs overexpressed in ovarian ALDH1-positive cells are associated with chemoresistance. J Ovarian Res. 2013;6:18. doi:10.1186/1757-2215-6-18
66. Bieg D, Sypniewski D, Nowak E, Bednarek I. MiR-424-3p suppresses galectin-3 expression and sensitizes ovarian cancer cells to cisplatin. Arch Gynecol Obstet. 2019;299:1077–1087. doi:10.1007/s00404-018-4999-7
67. Geretto M, Pulliero A, Rosano C, Zhabayeva D, Bersimbaev R, Izzotti A. Resistance to cancer chemotherapeutic drugs is determined by pivotal microRNA regulators. Am J Cancer Res. 2017;7:1350–1371.
68. Xu S, Tao Z, Hai B, et al. miR-424(322) reverses chemoresistance via T-cell immune response activation by blocking the PD-L1 immune checkpoint. Nat Commun. 2016;7:11406. doi:10.1038/ncomms11406
69. Liu X, Fu Y, Zhang G, et al. miR-424-5p promotes anoikis resistance and lung metastasis by inactivating hippo signaling in thyroid cancer. Mol Ther Oncolytics. 2019;15:248–260. doi:10.1016/j.omto.2019.10.008
70. Shivapurkar N, Vietsch EE, Carney E, Isaacs C, Wellstein A. Circulating microRNAs in patients with hormone receptor-positive, metastatic breast cancer treated with dovitinib. Clin Transl Med. 2017;6:37. doi:10.1186/s40169-017-0169-y
71. Rodriguez-Barrueco R, Nekritz EA, Bertucci F, et al. miR-424(322)/503 is a breast cancer tumor suppressor whose loss promotes resistance to chemotherapy. Genes Dev. 2017;31:553–566. doi:10.1101/gad.292318.116
72. Lu L, Wu M, Lu Y, et al. MicroRNA-424 regulates cisplatin resistance of gastric cancer by targeting SMURF1 based on GEO database and primary validation in human gastric cancer tissues. Onco Targets Ther. 2019;12:7623–7636. doi:10.2147/OTT.S208275
73. Zhang M, Zeng J, Zhao Z, Liu Z. Loss of MiR-424-3p, not miR-424-5p, confers chemoresistance through targeting YAP1 in non-small cell lung cancer. Mol Carcinog. 2017;56:821–832. doi:10.1002/mc.22536
74. Wang X, Li Q, Jin H, et al. miR-424 acts as a tumor radiosensitizer by targeting aprataxin in cervical cancer. Oncotarget. 2016;7:77508–77515. doi:10.18632/oncotarget.12716
75. Wen J, Hu Y, Liu Q, et al. miR-424 coordinates multilayered regulation of cell cycle progression to promote esophageal squamous cell carcinoma cell proliferation. EBioMedicine. 2018;37:110–124. doi:10.1016/j.ebiom.2018.10.043
76. Wei S, Li Q, Li Z, Wang L, Zhang L, Xu Z. miR-424-5p promotes proliferation of gastric cancer by targeting Smad3 through TGF-β; signaling pathway . Oncotarget. 2016;7:75185.
77. Shekhar R, Priyanka P, Kumar P, et al. The microRNAs miR-449a and miR-424 suppress osteosarcoma by targeting cyclin A2 expression. J Biol Chem. 2019;294:4381–4400. doi:10.1074/jbc.RA118.005778
78. Nandy SB, Orozco A, Lopez-Valdez R, et al. Glucose insult elicits hyperactivation of cancer stem cells through miR-424-cdc42-prdm14 signalling axis. Br J Cancer. 2017;117:1665–1675. doi:10.1038/bjc.2017.335
79. Dallavalle C, Albino D, Civenni G, et al. MicroRNA-424 impairs ubiquitination to activate STAT3 and promote prostate tumor progression. J Clin Invest. 2016;126:4585–4602. doi:10.1172/JCI86505
80. Thompson MP, Kurzrock R. Epstein-Barr virus and cancer. Clin Cancer Res. 2004;10:803–821. doi:10.1158/1078-0432.CCR-0670-3
81. Munoz N, Castellsagué X, de González AB, Gissmann L. HPV in the etiology of human cancer. Vaccine. 2006;24:S1–S10. doi:10.1016/j.vaccine.2006.05.115
82. Xu J, Fang Y, Wang X, et al. CUL2 overexpression driven by CUL2/E2F1/miR-424 regulatory loop promotes HPV16 E7 induced cervical carcinogenesis. Oncotarget. 2016;7:31520–31533. doi:10.18632/oncotarget.9127
83. Wang K, Zhu G, Bao S, Chen S. Long non-coding RNA LINC00511 mediates the effects of ESR1 on proliferation and invasion of ovarian cancer through miR-424-5p and miR-370-5p. Cancer Manag Res. 2019;11:10807–10819. doi:10.2147/CMAR.S232140
84. Dong J, Wang Q, Li L, Xiao-Jin Z. Upregulation of long non-coding RNA small nucleolar RNA host gene 12 contributes to cell growth and invasion in cervical cancer by acting as a sponge for MiR-424-5p. Cell Physiol Biochem. 2018;45:2086–2094. doi:10.1159/000488045
85. Hua F, Li CH, Chen XG, Liu XP. Long noncoding RNA CCAT2 knockdown suppresses tumorous progression by sponging miR-424 in epithelial ovarian cancer. Oncol Res. 2018;26:241–247. doi:10.3727/096504017X14953948675412
86. Gao YL, Zhao ZS, Zhang MY, Han LJ, Dong YJ, Xu B. Long noncoding RNA PVT1 facilitates cervical cancer progression via negative regulating of miR-424. Oncol Res. 2017;25:1391–1398. doi:10.3727/096504017X14881559833562
87. Cheng H, Tian J, Wang C, Ren L, Wang N. LncRNA BLACAT1 is upregulated in cervical squamous cell carcinoma (CSCC) and predicts poor survival. Reprod Sci. 2020;27:585–591. doi:10.1007/s43032-019-00058-9
88. Chen B, Zhao Q, Guan L, et al. Long non-coding RNA NNT-AS1 sponges miR-424/E2F1 to promote the tumorigenesis and cell cycle progression of gastric cancer. J Cell Mol Med. 2018;22:4751–4759.
89. Wang RP, Jiang J, Jiang T, Wang Y, Chen LX. Increased long noncoding RNA LINC00511 is correlated with poor prognosis and contributes to cell proliferation and metastasis by modulating miR-424 in hepatocellular carcinoma. Eur Rev Med Pharmacol Sci. 2019;23:3291–3301.
90. Li D, Tang X, Li M, Zheng Y. Long noncoding RNA DLX6-AS1 promotes liver cancer by increasing the expression of WEE1 via targeting miR-424-5p. J Cell Biochem. 2019;120:12290–12299. doi:10.1002/jcb.28493
91. Wang D, Hu Y. Long non-coding RNA PVT1 competitively binds microRNA-424-5p to regulate CARM1 in radiosensitivity of non-small-cell lung cancer. Mol Ther Nucleic Acids. 2019;16:130–140. doi:10.1016/j.omtn.2018.12.006
92. Li Y, Zhao L, Zhao P, Liu Z. Long non-coding RNA LINC00641 suppresses non-small-cell lung cancer by sponging miR-424-5p to upregulate PLSCR4. Cancer Biomark. 2019;26:79–91. doi:10.3233/CBM-190142
93. Ding J, Zhang L, Chen S, Cao H, Xu C, Wang X. lncRNA CCAT2 enhanced resistance of glioma cells against chemodrugs by disturbing the normal function of miR-424. Onco Targets Ther. 2020;13:1431–1445. doi:10.2147/OTT.S227831
94. Sun S-L, Shu Y-G, Tao M-Y. LncRNA CCAT2 promotes angiogenesis in glioma through activation of VEGFA signalling by sponging miR-424. Mol Cell Biochem. 2020;468:69–82. doi:10.1007/s11010-020-03712-y
95. Han Y, Li X, He F, et al. Knockdown of lncRNA PVT1 inhibits glioma progression by regulating miR-424 expression. Oncol Res. 2019;27:681–690. doi:10.3727/096504018X15424939990246
96. Zhou K, Li S, Du G, et al. LncRNA XIST depletion prevents cancer progression in invasive pituitary neuroendocrine tumor by inhibiting bFGF via upregulation of microRNA-424-5p. Onco Targets Ther. 2019;12:7095–7109. doi:10.2147/OTT.S208329
97. Li MM, Dong CX, Sun B, et al. LncRNA-MALAT1 promotes tumorogenesis of infantile hemangioma by competitively binding miR-424 to stimulate MEKK3/NF-kappaB pathway. Life Sci.. 2019;239:116946. doi:10.1016/j.lfs.2019.116946
98. Zhang J, Liu h, Hou L, et al. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR-424-5p and regulating LATS1 expression. Mol Cancer. 2017;16:151.
99. Hu Y, Gu J, Shen H, et al. Circular RNA LARP4 correlates with decreased Enneking stage, better histological response, and prolonged survival profiles, and it elevates chemosensitivity to cisplatin and doxorubicin via sponging microRNA-424 in osteosarcoma. J Clin Lab Anal. 2020;34:e23045. doi:10.1002/jcla.23045
100. Zhao B, Tumaneng K, Guan KL. The Hippo pathway in organ size control, tissue regeneration and stem cell self-renewal. Nat Cell Biol. 2011;13(8):877–883.
101. Zheng Y, Pan d. The Hippo Signaling Pathway in Development and Disease. Dev Cell. J Clin Lab Anal. 2019;50(3):264–282.
102. Olsen JV, Global, in vivo, and site-specific phosphorylation dynamics in signaling networks. Cell. 2006;127(3):635–648.
103. Wang X, Zhang S, MacLennan GT, et al. Epidermal growth factor receptor protein expression and gene amplification in small cell carcinoma of the urinary bladder. Clinical Cancer Research. 2007;13(3):953–957.
104. Wang X, Zhang S, MacLennan GT, et al. Epidermal growth factor receptor protein expression and gene amplification in the chemorefractory metastatic embryonal carcinoma. Modern Pathology. 2009;22(1):7–12.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.