")
Back to Journals » Journal of Blood Medicine » Volume 14
Role of Hemin in the Immune Response of T Follicular Helper Lymphocytes Expressing T-Cell Immunoreceptor with Immunoglobulin and Immunoreceptor Tyrosine-Based Inhibitory Domains, Programmed Cell Death-1, and Interleukin-21 in Allo-Auto Positive and Negative Thalassemia
Authors Tambunan BA , Ugrasena IDG, Aryati
Received 20 October 2022
Accepted for publication 6 January 2023
Published 13 January 2023 Volume 2023:14 Pages 7—17
DOI https://doi.org/10.2147/JBM.S393134
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Martin H Bluth
Betty Agustina Tambunan,1– 3 I Dewa Gede Ugrasena,3,4 Aryati2,3
1Doctoral Program of Medical Science, Faculty of Medicine, Universitas Airlangga, Surabaya, Indonesia; 2Department of Clinical Pathology, Faculty of Medicine, Universitas Airlangga, Surabaya, Indonesia; 3Dr. Soetomo General Academic Hospital, Surabaya, Indonesia; 4Department of Pediatrics, Faculty of Medicine, Universitas Airlangga, Surabaya, Indonesia
Correspondence: Aryati, Department of Clinical Pathology, Faculty of Medicine, Universitas Airlangga, St. Mayjen. Prof. Dr. Moestopo 47, Surabaya, East Java, 60132, Indonesia, Tel +6281230570493 ; +62-031-5020251, Email [email protected]
Introduction: Repeated transfusions in thalassemia patients can cause several complications, including alloimmunization and autoimmunization.
Purpose: This study compares the immune response of T follicular helper (Tfh) lymphocytes expressing T-cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory domains (TIGIT), programmed cell death-1 (PD-1), and interleukin-21 (IL-21) between patients with allo-auto positive and negative thalassemia before and after hemin administration.
Materials and Methods: This study used a quasi-experimental pre- and post-test design and was performed between April and November 2021 at the Dr. Soetomo General Academic Hospital in Surabaya, Indonesia. It enrolled 29 patients with allo-auto positive thalassemia and 28 with allo-auto negative, and 9 mL of whole blood (WB) was drawn from each patient. Hemin solution (20 μM) was added to 5 mL of WB, incubated for two hours, processed into peripheral blood mononuclear cells (PBMCs) in RPMI media, and cultured with 5% CO2 for three days. The 4 mL WB sample was also processed into PBMCs. PBMC cells cultured and without cultured were examined by flow cytometry using a BD FACSCalibur after surface and intracellular staining. Differences in Tfh cells expressing TIGIT, PD-1, and IL-21 between thalassemia groups before and after hemin administration were compared using independent t-tests or Mann–Whitney U-tests (p < 0.05).
Results: Tfh cell expression did not differ between groups before hemin administration and increased after hemin administration. The increase in Tfh cell expression was higher in the allo-auto positive group. TIGIT and PD-1 expression in Tfh cells did not differ between groups, but TIGIT decreased after hemin administration in contrast to PD-1 result. IL-21 expression in Tfh cells did not differ between groups and did not change after hemin administration.
Conclusion: Hemin affected the expression of Tfh cells in both group thalassemia, but there was no difference of Tfh cell expression between the groups.
Keywords: thalassemia disease, T follicular helper, TIGIT, PD-1, interleukin-21
Introduction
Thalassemia is an autosomal recessive hereditary disease. A partial or complete deficiency of globin chain tetramers in thalassemia causes chronic hemolytic anemia. The Thalassemia International Federation classifies thalassemia into transfusion-independent and transfusion-dependent based on its need for transfusion.1 As technology advances, disease-causing mutations have become treatment targets. Several approaches have been explored as nucleic acid-based therapies. However, several limitations challenge their implementation, such as stability, delivery method, suitable vector, and optimal nucleic acid dose.2
Transfusion remains the primary treatment recommended for thalassemia. However, repeated transfusions can cause iron overload, characterized by increased serum ferritin levels that interfere with 25(OH)D synthesis in the liver.3 High excess iron also causes impaired glucose metabolism.4 Other complications are common, such as mild depression, especially in patients with high serum cortisol levels,5 transfusion reactions, infection, alloimmunization, and autoimmunization.6 Alloimmunization rates are reported to be 5–30% in thalassemia major in Mediterranean and Southeast Asian countries. A Thai study found an alloimmunization rate of 17.5% in transfusion-dependent thalassemia patients.7 Research on patients with homozygous beta-thalassemia who received repeated transfusions found that 87 of 385 patients (22.6%) experienced autoimmunization.8 The reported autoimmunization rates in thalassemia patients in several countries include 28.8% in Egypt,9 23% in Hong Kong,10 22.8% in Albania,11 and 6.5% in the US.12
Alloimmunization and autoimmunization can have many disadvantages, including acute and delayed hemolytic transfusion reactions, difficulty in obtaining compatible blood leading to delayed transfusions, additional pre-transfusion tests, and increased costs for laboratory tests and patient management.6 Therefore, the role of the immune response in alloimmunization and autoimmunization incidence in thalassemia patients must be further studied.13
Several studies on the immune response in chronic hemolytic conditions have been performed in sickle cell disease (SCD) patients. Cell culture studies of SCD patients who received repeated transfusions showed an increased response of CD4+ T cells to the hemin administration in alloimmunized patients compared to those without alloimmunization.14 Hemin is a marker of red blood cell degradation products that induce changes in TCD4+ cell subsets through activation of heme oxygenase-1 (HO-1).15 Another study on SCD patients showed that T follicular helper (Tfh) cells expressing programmed cell death-1 (PD-1) are more numerous in patients without alloimmunization compared to those with alloimmunization, while interleukin-21 (IL-21) expressing cells were more numerous in patients with alloimmunization.16 Cell culture studies of SCD patients with repeated transfusions showed an increase in Tfh cells expressing T-cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory domains (TIGIT) in alloimmunized versus non-alloimmunized subjects, while Tfh cells expressing PD-1 were not different.17
Whether the immune response in thalassemia is similar to SCD patients remains unknown.18 Since chronic hemolysis occurs in both SCD and thalassemia patients, this study compares the immune response of Tfh lymphocytes expressing TIGIT, PD-1, and IL-21 between transfusion-dependent thalassemia patients with (allo-auto positive) and without (allo-auto negative) alloimmunization and autoimmunization before and after hemin administration. The study’s results are expected to provide information for therapy development in thalassemia patients through modification of the immune response of Tfh cells, especially in patients with alloimmunization.
Materials and Methods
This study was quasi-experimental with a non-equivalent control group and pre- and post-test design. It was performed between April and November of 2021 at the Dr. Soetomo General Academic Hospital in Surabaya, Indonesia. This study was approved by Local Institutional Human Research Ethics Committee at the Dr. Soetomo General Academic Hospital (no. 0527/LOE/301.4.2/VII/2021) in accordance with the standards laid down in the 1964 Declaration of Helsinki.
We enrolled 29 patients with transfusion-dependent allo-auto positive thalassemia and 28 with allo-auto negative thalassemia. Sample size calculated by determining the sample size for experimental research with two groups (1-β=0.80). Patients underwent standard set blood sample drawing after obtaining informed consent. Informed consent was obtained from parents or legal guardians for research participants under 18 years old. Heparin blood samples were taken from each patient. In particular, 9 mL of whole blood was drawn from each patient and divided into two heparinized tubes; one contained 5 mL for culture and the other contained 4 mL for non-culture. A 20 µM hemin solution was added to culture samples, incubated for two hours, and prepared to manufacture peripheral blood mononuclear cells (PBMCs).
PBMC cells were diluted to a density of 106 cells/mL and divided into three 1 mL tubes. Fetal bovine serum, streptomycin, and penicillin were added to each tube and mixed. Next, they were placed in an incubator with 5% CO2 for three days. PBMC cultures were harvested after three days and centrifuged at 1500 rpm for 10 minutes to separate PBMCs from the supernatant. PBMCs were washed twice and prepared for flow cytometry. The 4 mL blood samples were added to 4 mL of PBS, mixed, divided into two tubes, and prepared for PBMC manufacturing.
Tfh lymphocytes expressing TIGIT, PD-1, and IL-21 were identified using flow cytometry with a BD FACS Calibur instrument. The Tfh cells were identified by surface staining by adding mouse anti-human CD4 labeled with fluorescein isothiocyanate (BD Pharmingen) and rat anti-human C-X-C motif chemokine receptor 5 (CXCR5/CD185) labeled with PerCP-Cy5.5 (BD Pharmingen). Monoclonal anti-TIGIT labeled with allophycocyanin (eBioscience) as the TIGIT surface marker. Monoclonal mouse anti-human PD-1 (CD279) antibody (BD Pharmingen) labeled with phycoerythrin was added as the PD-1 surface marker. Intracellular staining was performed by adding mouse anti-human IL-21 labeled with Alexa Fluor 647 (BD Pharmingen).
Statistical Analysis
The differences in Tfh cells expressing TIGIT, PD-1, and IL-21 between allo-auto positive and negative thalassemia groups before and after hemin administration were assessed with independent t-tests when normally distributed and with Mann–Whitney U-tests when nonnormally distributed. All results with p < 0.05 were considered statistically significant and analyzed using IBM SPSS Statistics Version 21.
Results
Participants’ demographic data (gender, age, and blood type) were collected when they first received blood transfusions. The allo-auto negative thalassemia group was dominated by the O blood type (50%), while the allo-auto positive thalassemia group was dominated by the B blood type (44.8%). All patients were rhesus positive (Table 1).
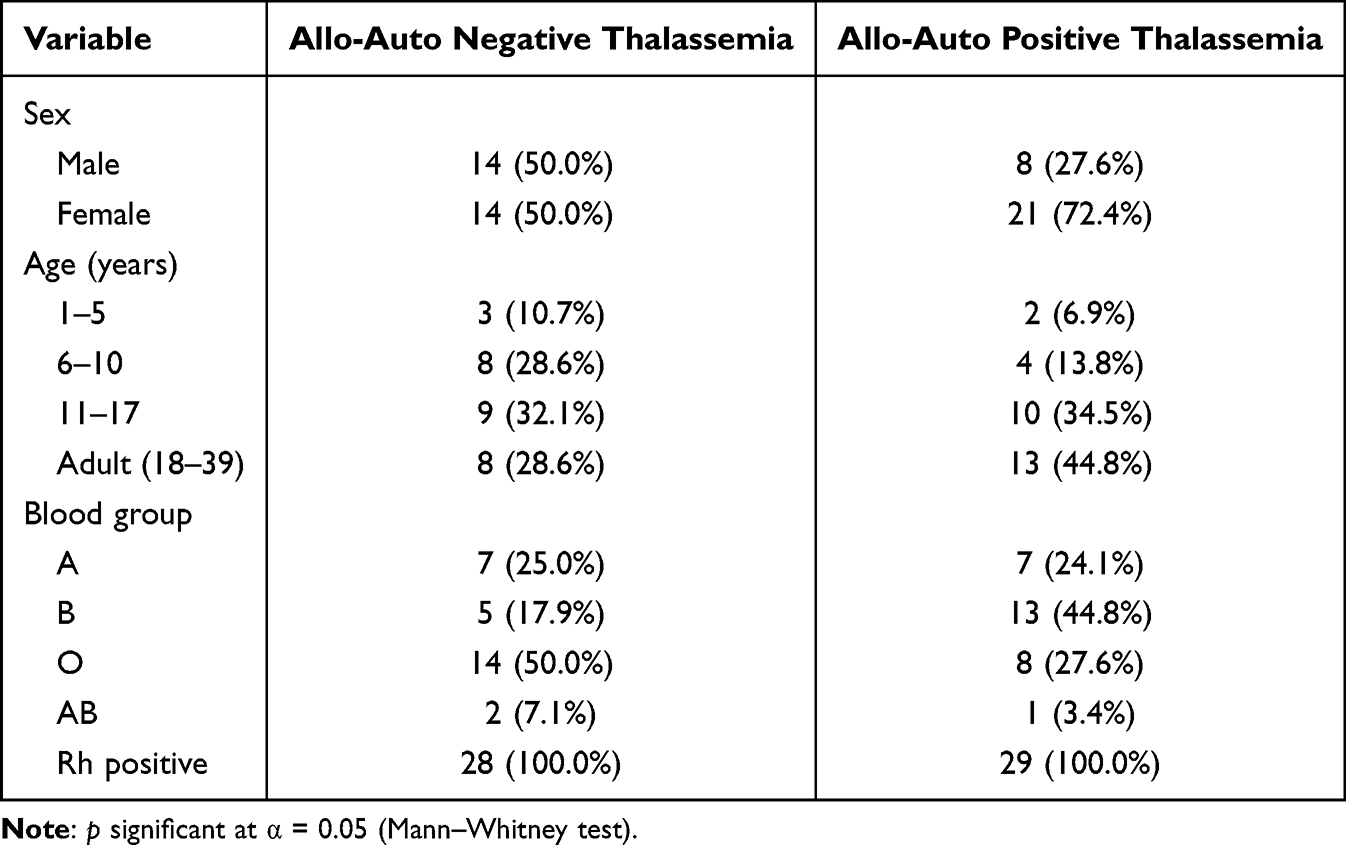 |
Table 1 Participant Characteristics |
Of the 29 allo-auto positive thalassemia patients, 13 showed positive autocontrol and negative antibody screening (44.8%), termed thalassemia with autoimmunization. Negative autocontrol and positive antibody screening were found in four patients (13.8%), termed alloimmunization thalassemia. Positive autocontrol and positive antibody screening were found in 12 patients (41.4%), termed alloimmunization and autoimmunization thalassemia (Table 2).
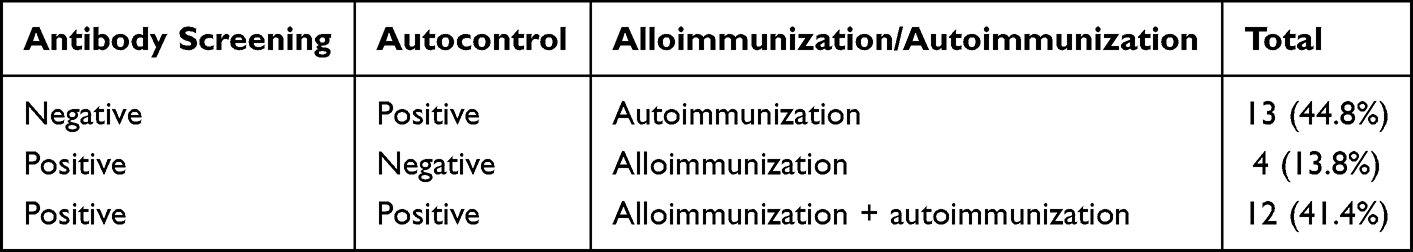 |
Table 2 Antibody and Autocontrol Screening Results |
T Follicular Helper Cells
Tfh cells in allo-auto negative and positive thalassemia significantly increased after hemin administration (p = 0.024 and p = 0.000, respectively). In addition, the increase in Tfh cells differed significantly between groups (p = 0.030). Tfh cell numbers in allo-auto negative thalassemia were not significantly different from allo-auto positive thalassemia before hemin administration (p = 0.267) but were significantly higher after hemin administration (p = 0.024) (Figure 1).
 |
Figure 1 T follicular helper (Tfh) cells. |
Activation of Tfh Cells Expressing TIGIT
Tfh cells expressing TIGIT in allo-auto negative and positive thalassemia significantly decreased after hemin administration (p = 0.000 and p = 0.001, respectively). The change in Tfh cells expressing TIGIT did not differ significantly between groups (p = 0.326). Tfh cells expressing TIGIT did not differ significantly between groups before (p = 0.083) or after (p = 0.199) hemin administration (Figure 2).
 |
Figure 2 Tfh cells expressing TIGIT. |
Tfh Cells Expressing PD-1
Tfh cells expressing PD-1 did not differ significantly between allo-auto negative and positive thalassemia before (p = 0.835) and after (p = 0.133) hemin administration. However, Tfh cells expressing PD-1 significantly increased after hemin administration in allo-auto negative (p = 0.000) and allo-auto positive (p = 0.001) thalassemia. However, the change of Tfh cells expressing PD-1 did not differ significantly between groups (p = 0.238) (Figure 3).
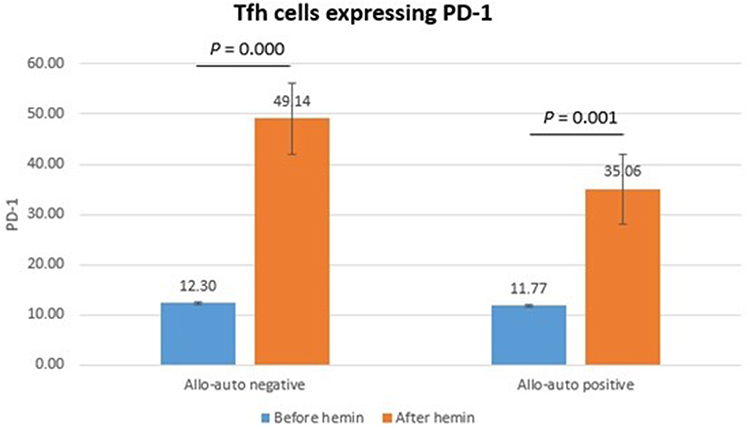 |
Figure 3 Tfh cells expressing PD-1. |
Tfh Cells Expressing IL-21
Tfh cells expressing IL-21 did not differ significantly between allo-auto negative and positive thalassemia before (p = 0.914) and after (p = 0.195) hemin administration. Tfh cells expressing IL-21 did not differ significantly after hemin administration in allo-auto negative (p = 0.995) and positive (p = 0.063) thalassemia. The change in Tfh cells expressing IL-21 did not differ significantly between groups (p = 0.513) (Figure 4).
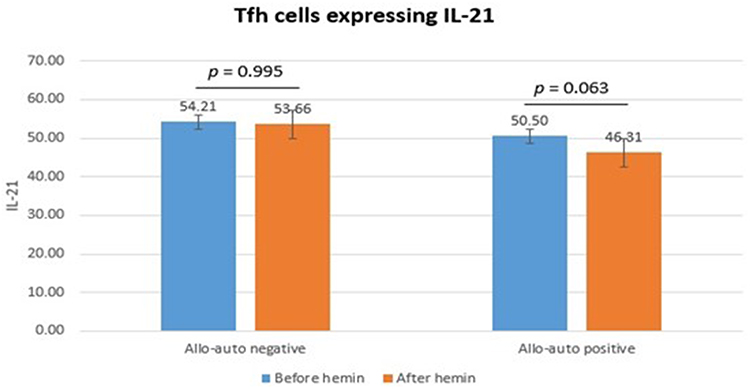 |
Figure 4 Tfh cells expressing IL-21. |
Discussion
Fifty-seven transfusion-dependent thalassemia patients were included in this study, 28 with allo-auto negative and 29 with allo-auto positive thalassemia. Our results suggest that the allo-auto positive thalassemia group was dominated by 13 autoimmunization patients (44.8%), followed by 12 alloimmunization and autoimmunization patients (41.4%), and four alloimmunized patients (13.8%). An Albanian prospective cohort study followed 118 thalassemia patients receiving regular transfusions for five years, screened for antibodies, and given blood with matched ABO, rhesus, and Kell blood group systems. Of these 118 patients, 14 (11.8%) were alloimmunized and 27 (22.8%) were autoimmunization. They found that of the 14 alloimmunized patients, 11 also had autoimmunization.11
An Egyptian study found that of 200 thalassemia patients receiving repeated transfusions, those with alloimmunization were more alloimmunized (36; 18%) than autoimmunized (33; 16.5%).18 A Chinese study reported that of their 382 patients, 294 (77.0%) were antibody negative and 88 (23.0%) were antibody positive. Of the antibody-positive patients, 70 (79.5%) showed alloimmunization and 18 (20.5%) showed autoimmunization.10 Our study is consistent with the Albanian study, suggesting that many patients with alloimmunization also have autoimmunization. The frequency of patients with autoimmunization is higher than with alloimmunization, which is inconsistent with the Egyptian and Chinese studies. The influencing factors in the frequency of alloimmunization and autoimmunization are unclear. Overall, our study is consistent with all three studies because thalassemia patients experienced unexplainable alloimmunization or autoimmunization.
Our study found that allo-auto negative thalassemia is dominated by blood type O, while allo-auto positive thalassemia is dominated by blood type B. A Greek study of 65 thalassemia patients receiving repeated transfusions found that 36.9% had blood group B and were rhesus positive.18 An Indian study of 500 thalassemia patients receiving repeated transfusions found that 36.8% had blood group B and were rhesus positive.19 Unfortunately, these two studies did not classify thalassemia as with or without alloimmunization. Whether there is a relationship between the incidence of alloimmunization and autoimmunization with the ABO blood group remains unclear.
Tfh cells in allo-auto negative thalassemia were not significantly different from allo-auto positive thalassemia before hemin administration. Tfh cells are a subset of CD4+ T cells that stimulate B cells to produce antibodies in the effective humoral immune response.20 Tfh cells were characterized by their expression of the transcription factor nuclear transcriptional repressor B cell lymphoma 6 (BCL6), the CXCR5, inducible co-stimulator (ICOS), PD-1, and IL-21.21
Since 2011, CD4+ CXCR5+ T cells in peripheral blood have been known as circulating Tfh cells, which have been shown to promote B cell differentiation into plasma cells and produce immunoglobulin G (IgG) in vitro.22 Elevated Tfh cells have been found in patients with systemic lupus erythematosus (SLE), myasthenia gravis, and juvenile dermatomyositis.23 Excessive activation of Tfh cells can also lead to autoimmunization.24 Other studies have also shown an increase in circulating Tfh cells in patients with active autoimmune disease.25
Circulating Tfh cells account for about 6% of the total CD4+ T cells in peripheral blood in healthy individuals.22 Tfh cells have antigen-specific memory and regulate memory B cells.26 Our study found that the median percentage of Tfh cells from the CD4+ T cells in allo-auto negative thalassemia patients was lower (5.03%) than in allo-auto positive thalassemia patients (6.29%). However, the percentage of Tfh cells did not differ significantly between groups. Another study found that the percentage of Tfh cells in SCD patients was not significantly different between those without alloimmunization and those with alloimmunization.17
A study in mice modeled autoimmune hemolytic anemia (AIHA) and showed an increase in the number of Tfh cells and the ratio of Tfh to T follicular regulatory T cells (Tfr) in mice with positive autoantibodies. They also found that Tfh cells injected into AIHA-modeled mice stimulated the formation of autoantibodies, indicating the important role of Tfh cells in controlling the induction of B cells to produce antibodies in AIHA. Increased Tfh cell activity is associated with a response to exposure to a given red cell antigen. Weekly intraperitoneal injections of red blood cells in rats led to the formation of autoantibodies against erythrocytes five to six weeks after immunization. Peak erythrocyte autoantibody levels were formed within 10–12 weeks post-injection. They correlated with an increase in reticulocytes and significantly reduced the lifespan of red blood cells. The increase in Tfh cells correlated positively with anti-RBC IgG levels.21
Hemin administration increased the number of Tfh cells in both groups. Tfh cells were higher in allo-auto positive thalassemia patients after hemin administration. Hemoglobin and heme resulting from the destruction of red blood cells in thalassemia hemolytic anemia can act as erythrocytic danger-associated molecular pattern molecules that activate the innate (natural) immune response.27 Heme induces T regulatory (Treg) cell proliferation and inhibits Th1 through HO-1 activation in SCD patients without alloimmunization but not in patients with alloimmunization.28 Hemin administration was believed to induce Tfh cell proliferation in both groups due to the Tfh response to heme. Hemin induced a stronger Tfh cell response in allo-auto positive thalassemia patients, indicated by a larger increase in the percentage of Tfh cells. We hypothesize that the increase in Tfh cells in allo-auto positive thalassemia patients might be related to Tfh cells, which are more sensitive to hemin exposure due to their memory of specific red blood cell antigens contacted.
Tfh cells expressing TIGIT in allo-auto negative thalassemia patients were not significantly different from allo-auto positive thalassemia patients. Another study of healthy individuals showed that the percentage of Tfh cells expressing TIGIT in peripheral blood was 47% ± 3.2%.17 Our study showed that the median percentage of Tfh cells expressing TIGIT in peripheral blood of allo-auto negative thalassemia patients was lower than in healthy individuals in Godefroy et al but with a wider range.17 The highest percentage was almost the same as normal individuals (20.88%; 4.93–47.29%) and allo-auto positive thalassemia patients (29.20%; 4.23–51.55%). Our study showed that the percentage of Tfh cells expressing TIGIT did not differ significantly between groups, consistent with the SCD study that showed the percentage of Tfh cells expressing TIGIT did not differ significantly between patients with allo-auto positive and negative thalassemia.
A significant decrease in Tfh cells expressing TIGIT in both groups after hemin administration was showed in our study. Studies of autoimmune SLE patients showed that they have more Tfh cells expressing TIGIT than healthy individuals, correlating with severity as assessed by the SLE disease activity index criteria.29 Cancer research shows that TIGIT inhibition in Treg cells (CD4+ FOXP3+ TIGIT+) increases the activity of TCD8+ and NK cells, inhibiting tumor growth.30 The role of Tfh cells in thalassemia remains unclear. Our study showed that hemin administration to PBMCs from thalassemia patients decreased Tfh cells expressing TIGIT. The decrease in the percentage of Tfh cells expressing TIGIT after hemin administration might be an anti-inflammatory response to HO-1 activation. This hypothesis is consistent with the effect of hemin on cytokine regulation through the activation of HO-1, which has an anti-inflammatory effect by inhibiting macrophages secreting proinflammatory cytokines (interleukin-1β and tumor necrosis factor α) and inducing the release of interleukin-10.31
Another study of SLE patients showed a decrease in Tfh cells expressing TIGIT after stimulation with mitogen phytohemagglutinin. They also used recombinant CD155 (TIGIT receptors on dendritic cells), which decreased CD4+ T cell proliferation. This finding suggests that TIGIT may act as a negative regulator of TCD4+ cell activity.29 HO-1 acts as an immunosuppressant, inhibiting dendritic cell maturation.32 This study proposed that hemin might activate HO-1, inhibiting the response of dendritic cells to TIGIT and decreasing the percentage of Tfh cells expressing TIGIT after hemin administration.
Tfh cells expressing PD-1 did not differ significantly between groups before and after hemin administration. The median percentage of Tfh cells expressing PD-1 in our study for allo-auto negative and positive thalassemia patients was lower but broader than in healthy individuals in another study.33 Our study also found that Tfh cells expressing PD-1 increased significantly in both groups after hemin administration. The median percentage of Tfh cells expressing PD-1 result was lower than in lymphoma patients from another study.33
PD-1 is a co-inhibitor of cytokine production and effector T cell proliferation. This mechanism maintains tolerance to autoantigens and prevents autoimmunity.34 Inhibition of T-cell receptor signaling and costimulatory CD28 is considered the main pathway of PD-1 action.35 Programmed death-ligand 1 (PD-L1) is expressed by hematopoietic and nonhematopoietic cells. Cells expressing PD-1 will bind to PD-L1, inducing signals that inhibit the function and survival of Tfh cells expressing PD-1.36 Another study found that PD-1 inhibits the function of Tfr. PD-1 deficiency in Tfr cells in the peripheral blood increases suppressive capacity.37 In-vitro studies of T cells in the lymph nodes of patients infected with the human immunodeficiency virus (HIV) showed an increase in Tfh cells expressing PD-1 followed by a decrease in ICOS expression and IL-21 secretion, suggesting that PD-1 inhibits the increased production of HIV-specific immunoglobulins.20
Experimental animal studies have shown that dendritic cells are required to initiate the transformation of naive TCD4 cells into Tfh cells expressing PD-1. The culture of naive TCD4 cells with naive B cells alone did not produce Tfh cells. However, when dendritic cells were added, Tfh cells expressing PD-1 increased significantly.38 Hemin is responsible for activating HO-1, which is responsible for inducing dendritic cell responses.39
Tfh cells expressing PD-1 increased in both groups after hemin administration in this study, possibly due to hemin inducing HO-1. This process activates dendritic cell responses, increasing Tfh cells expressing PD-1. The increase in Tfh cells expressing PD-1 inhibits Tfh cell activation against specific antigen stimulation, making Tfh cells unable to induce B cells to form antibodies. This inability might have a protective effect against transfusion-dependent thalassemia after repeated exposure to red cell antigens that are different from the self-antigens.
Our study showed that Tfh cells expressing IL-21 did not differ significantly between groups before and after hemin administration. Tfh cells produce IL-21 in response to antigens presented by dendritic cells, inducing B cell differentiation in germinal centers. One study reported that circulating Tfh cells also had memory capabilities.40 Tfh cells expressing IL-21 did not differ between groups, possibly due to repeated transfusions resulting in exposure to different red cell antigens under the same conditions in both groups that were not affected by hemin.
The results showed a higher median percentage of Tfh cells expressing IL-21 in both groups but a wide interval compared to osteoarthritis patients in another study.41 Studies on lymphocyte cells from mouse lymph nodes have shown that IL-21 mediates the growth and survival of Tfh cells. The IL-21 receptor is expressed on Tfh cells and B cells, suggesting a central role of IL-21 in Tfh cell formation and antibody production. Tfh cells are known to express ICOS and IL-21. ICOS expression has an impact on IL-21 production.42
Another study that cocultured circulating Tfh cells with dendritic cells showed that dendritic cells led to increased Tfh cell activation and cytokine production, including IL-21, but IL-21 also inhibited dendritic cell activation and maturation.43 Studies in patients with Sjogren’s syndrome with positive antibodies have reported circulating Tfh cells expressing higher IL-21 and positively correlated with IgG levels.44 We suspect that hemin administration did not increase Tfh cells expressing IL-21 because hemin is not a specific antigen presented by dendritic cells to Tfh cells, leading to a weaker effect on increasing IL-21 expression. In addition, the number of Tfh cells expressing IL-21 was already high, which might be sufficient to maintain Tfh cell activation and survival.
We recognize that this study has limitations, so it cannot comprehensively describe the immune response of the two thalassemia groups. Some factors that affected the study results cannot be completely ruled out. The level of heme in the blood was not measured, so the hemolysis level of individual subjects was not evaluated. This study did not measure the transcription factor nuclear transcriptional repressor B cell lymphoma 6 (BCL6), the CXCR5, an inducible co-stimulator (ICOS) that affects the immune response of T cells, including Tfh, to stimulate B cells forming antibodies. This study was limited to the Tfh immune response, while the hemolysis-related immune response involved the innate immune response. We also did not examine T regulators, which control T effector, including Tfh cells.
Conclusions
Tfh cells did not differ significantly between thalassemia groups before hemin administration, but a significant increase was observed after hemin administration. The increase in Tfh cells was higher in allo-auto positive thalassemia patients. Tfh cells expressing TIGIT did not differ significantly between thalassemia groups, but a significant decrease was observed after hemin administration. Tfh cells expressing PD-1 did not differ significantly between thalassemia groups, but hemin administration increased Tfh cells expressing PD-1 in both groups. Tfh cells expressing IL-21 did not differ significantly between thalassemia groups before and after hemin administration. The immune response of Tfh cells expression did not differ between allo-auto positive and negative thalassemia. Hemin administration affected Tfh cell expression in both thalassemia groups.
Data Sharing Statement
The statistical data used to support the findings of this study are available from the corresponding author upon request.
Ethical Approval
The study was approved by the Health Research Ethics Committee of the Dr. Soetomo General Academic Hospital (no. 0527/LOE/301.4.2/VII/2021).
Acknowledgments
We acknowledge all participants for supporting and cooperating in this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declare no competing financial interests or personal relationships that could have influenced the work reported in this article.
References
1. Viprakasit V, Ekwattanakit S. Clinical classification, screening and diagnosis for thalassemia. Hematol Oncol Clin North Am. 2018;32:193–211. doi:10.1016/j.hoc.2017.11.006
2. D’Arqom A. Nucleic acid therapy for β-thalassemia. Biologics. 2020;14:95–105.
3. Jazuli MI, Bintoro SUY, Mudjanarko SW. The association between serum ferritin levels and 25(OH)D levels in adult patients with transfusion dependent thalassemia. J Med Chem Sci. 2022;5:35–41.
4. Fonda T, Novida H. Management of gestasional diabetes mellitus in a beta major thalassemia patient. Curr Intern Med Res Pract Surabaya J. 2021;2:45. doi:10.20473/cimrj.v2i2.26255
5. Suryawan A, Ningtiar HW, Irwanto I, et al. Determinant factors of depression in beta major thalassemia children. Folia Medica Indones. 2021;57:46. doi:10.20473/fmi.v57i1.13664
6. Meulenbroek EM, Wouters D, Zeerleder SS. Lyse or not to lyse: clinical significance of red blood cell autoantibodies. Blood Rev. 2015;29:369–376. doi:10.1016/j.blre.2015.05.001
7. Tangvarasittichai S. Impact of alloimmunization on transfusion-dependent patient. Ann Adv Chem. 2017;1:70–82. doi:10.29328/journal.aac.1001009
8. Ben KM, Ouederni M, Sahli N, et al. Predictors of autoimmune hemolytic anemia in beta-thalassemia patients with underlying red blood cells autoantibodies. Blood Cells, Mol Dis. 2019;79. doi:10.1016/j.bcmd.2019.102342
9. Al-Riyami AZ, Al-Muqbali A, Al-Sudiri S, et al. Risks of red blood cell alloimmunization in transfusion-dependent β-thalassemia in Oman: a 25-year experience of a university tertiary care reference center and a literature review. Transfusion. 2018;58:871–878. doi:10.1111/trf.14508
10. Cheng CK, Lee CK, Lin CK. Clinically significant red blood cell antibodies in chronically transfused patients: a survey of Chinese thalassemia major patients and literature review. Transfusion. 2012;52:2220–2224. doi:10.1111/j.1537-2995.2012.03570.x
11. Seferi I, Xhetani M, Face M, et al. Frequency and specificity of red cell antibodies in thalassemia patients in Albania. Int J Lab Hematol. 2015;37:569–574. doi:10.1111/ijlh.12362
12. Vichinsky E, Neumayr L, Trimble S, et al. Transfusion complications in thalassemia patients: a report from the CDC. Transfusion. 2015;54:1–20.
13. Ryter SW, Alam J, Choi AMK. Heme oxygenase-1/carbon monoxide: from basic science to therapeutic applications. Physiol Rev. 2006;86:583–650. doi:10.1152/physrev.00011.2005
14. Zhong H, Bao W, Friedman D, et al. Hemin controls T cell polarization in sickle cell alloimmunization. J Immunol. 2014;193:102–110. doi:10.4049/jimmunol.1400105
15. Martins R, Knapp S. Heme and hemolysis in innate immunity: adding insult to injury. Curr Opin Immunol. 2018;50:14–20. doi:10.1016/j.coi.2017.10.005
16. Vingert B, Tamagne M, Habibi A, et al. Phenotypic differences of CD4+ T cells in response to red blood cell immunization in transfused sickle cell disease patients. Eur J Immunol. 2015;45:1868–1879. doi:10.1002/eji.201445187
17. Godefroy E, Zhong H, Pham P, et al. TIGIT-positive circulating follicular helper T cells display robust B-cell help functions: potential role in sickle cell alloimmunization. Haematologica. 2015;100:1415–1425. doi:10.3324/haematol.2015.132738
18. Saifeldeen E, Awad M, El-Tonbary Y, et al. Risk for red cell immunization among thalassemic patients. Egypt J Haematol. 2017;42:58. doi:10.4103/1110-1067.216113
19. Datta SS, Mukherjee S, Talukder B, et al. Frequency of red cell alloimmunization and autoimmunization in thalassemia patients: a report from Eastern India. Adv Hematol. 2015;2015:1–6. doi:10.1155/2015/610931
20. Raul Badell I, Ford ML, Follicular Helper T. Cells in the generation of alloantibody and graft rejection. Physiol Behav. 2016;176:139–148.
21. Gao Y, Jin H, Nan D, et al. The Role of T follicular helper cells and T follicular regulatory cells in the pathogenesis of autoimmune hemolytic anemia. Sci Rep. 2019;9:1–11. doi:10.1038/s41598-019-56365-3
22. Brenna E, Davydov AN, Ladell K, et al. CD4+ T follicular helper cells in human tonsils and blood are clonally convergent but divergent from non-Tfh CD4+ cells. Cell Rep. 2020;30:137–152.e5. doi:10.1016/j.celrep.2019.12.016
23. Crotty S. T follicular helper cell differentiation, function and roles in disease. Immunity. 2015;41:529–542.
24. Laguna-Goya R, Suàrez-Fernández P, Paz-Artal E. Follicular helper T cells and humoral response in organ transplantation. Transplant Rev. 2019;33:183–190. doi:10.1016/j.trre.2019.06.003
25. Schey KL, Luther JM, Rose KL. Pathophysiology of T follicular helper cells in humans and mice. Nat Immunol. 2016;16:1–21.
26. Pauff SM, Miller SC. Follicular helper T cells: lineage and location. Bone. 2012;78:711–716.
27. Kato GJ, Steinberg MH, Gladwin MT. Intravascular hemolysis and the pathophysiology of sickle cell disease. J Clin Invest. 2017;127:750–760. doi:10.1172/JCI89741
28. Zhong H, Yazdanbakhsh K. Hemolysis and immune regulation. Curr Opin Hematol. 2018;25:177–182. doi:10.1097/MOH.0000000000000423
29. Luo Q, Ye J, Zeng L, et al. Elevated expression of TIGIT on CD3+CD4+ T cells correlates with disease activity in systemic lupus erythematosus. Allergy Asthma Clin Immunol. 2017;13:1–12. doi:10.1186/s13223-017-0188-7
30. Kurtulus S, Sakuishi K, Ngiow SF, et al. TIGIT predominantly regulates the immune response via regulatory T cells. J Clin Invest. 2015;125:4053–4062. doi:10.1172/JCI81187
31. Ma JL, Yang PY, Rui YC, et al. Hemin modulates cytokine expressions in macrophage-derived foam cells via heme oxygenase-1 induction. J Pharmacol Sci. 2007;103:261–266. doi:10.1254/jphs.FP0060270
32. Chauveau C, Rémy S, Royer PJ, et al. Heme oxygenase-1 expression inhibits dendritic cell maturation and proinflammatory function but conserves IL-10 expression. Blood. 2005;106:1694–1702. doi:10.1182/blood-2005-02-0494
33. Byford ET, Carr M, Ladikou E, et al. Circulating Tfh1 (cTfh1) cell numbers and PD1 expression are elevated in low-grade B-cell non-Hodgkin’s lymphoma and cTfh gene expression is perturbed in marginal zone lymphoma. PLoS One. 2018;13:1–18. doi:10.1371/journal.pone.0190468
34. Bardhan K, Anagnostou T, Boussiotis VA. The PD1: PD-L1/2 pathway from discovery to clinical implementation. Front Immunol. 2016;7. doi:10.3389/fimmu.2016.00550
35. Shi J, Hou S, Fang Q, et al. PD-1 controls follicular T helper cell positioning and function. Immunity. 2018;49:264–274.e4. doi:10.1016/j.immuni.2018.06.012
36. Jogdand GM, Mohanty S, Devadas S. Regulators of Tfh cell differentiation. Front Immunol. 2016;7:1–14. doi:10.3389/fimmu.2016.00520
37. Sage PT, Francisco LM, Carman CV, et al. PD-1 controls lymph node and blood T follicular regulatory cells. Nat Immunol. 2013;14:152–161. doi:10.1038/ni.2496
38. Martin-Gayo E, Cronin J, Hickman T, et al. Circulating CXCR5+CXCR3+PD-1lo Tfh-like cells in HIV-1 controllers with neutralizing antibody breadth. JCI Insight. 2017;2. doi:10.1172/jci.insight.89574
39. Thomas RAB, Czopek A, Bellamy COC, et al. Hemin preconditioning upregulates heme oxygenase-1 in deceased donor renal transplant recipients: a randomized, controlled, phase IIB Trial. Transplantation. 2016;100:176–183. doi:10.1097/TP.0000000000000770
40. Sage PT, Alvarez D, Godec J, et al. Circulating T follicular regulatory and helper cells have memory-like properties. J Clin Invest. 2014;124:5191–5204. doi:10.1172/JCI76861
41. Shan Y, Qi C, Liu Y, et al. Increased frequency of peripheral blood follicular helper T cells and elevated serum IL-21 levels in patients with knee osteoarthritis. Mol Med Rep. 2017;15:1095–1102. doi:10.3892/mmr.2017.6132
42. Vogelzang A, McGuire HM, Yu D, et al. A fundamental role for interleukin-21 in the generation of T follicular helper cells. Immunity. 2008;29:127–137. doi:10.1016/j.immuni.2008.06.001
43. Patente TA, Pinho MP, Oliveira AA, et al. Human dendritic cells: their heterogeneity and clinical application potential in cancer immunotherapy. Front Immunol. 2019;10:1–18. doi:10.3389/fimmu.2019.00001
44. Pontarini E, Murray-Brown WJ, Croia C, et al. Unique expansion of IL-21+ Tfh and Tph cells under control of ICOS identifies Sjögren’s syndrome with ectopic germinal centres and MALT lymphoma. Ann Rheum Dis. 2020;79:1588–1599. doi:10.1136/annrheumdis-2020-217646
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.