Back to Journals » International Journal of General Medicine » Volume 18
RNA Binding Motif Protein 15 (RBM15): Structure, Function and Its Research Progress in Tumors
Authors Long X, Hu B, Zhu S, Wu Y, Wang T
Received 18 February 2025
Accepted for publication 23 June 2025
Published 4 July 2025 Volume 2025:18 Pages 3635—3649
DOI https://doi.org/10.2147/IJGM.S519741
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Dana Kristjansson
Xiangyu Long,* Bintao Hu,* Shiqing Zhu, Yue Wu, Tao Wang
Department of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Tao Wang, Department of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, Hubei, People’s Republic of China, Email [email protected]
Abstract: N6-methyladenosine (m6A) is the most common modification on mammalian mRNA and has gained considerable attention in recent years as a key regulator of gene expression. RBM15 is an important component of the methyltransferase complex. Although its functions are increasingly being recognized, a comprehensive overview of its critical roles in cancer regulation and potential therapeutic relevance is still lacking. This review aims to summarize recent advances in RBM15 by outlining its physiological functions in normal organisms, detailing its biological roles in tumor regulation and discussing its potential clinical applications. This work highlights its potential as a therapeutic target in oncology.
Keywords: RBM15, m6A modification, tumor, therapeutic targets
Background
Previous studies have elucidated the hierarchical nature of epigenetic regulation mediated by DNA and protein modifications. However, the epigenetic regulatory role of RNA modifications is not yet been fully investigated. RNA methylation, which accounts for over 60% of all RNA modifications, is the most prevalent form of RNA modification, with N6-methyladenosine (m6A) being the most common modification in mammalian messenger RNA (mRNA).1 Initially identified in the 1970s, the m6A modification did not attract considerable attention at that time. Recently, it has emerged as a crucial regulator of gene expression,2 sparking renewed interest in both cancer and non-cancer research. The m6A modification has been shown to be critically involved in cancer biology, modulating essential biological processes such as RNA maturation, localization, and translation.3–5
M6A RNA modification is a dynamical and reversible biological process—orchestrated by three classes of enzymes: m6A methyltransferases (“Writers”), m6A demethylases (“Erasers”), and m6A binding proteins (“Readers”). These enzymes collaboratively regulate m6A modifications, thus facilitating RNA processing, degradation, nuclear export, and translation, which controls RNA expression and function.6,7 The m6A methyltransferase complex, a multi-component entity, mediates m6A modifications on mRNA. METTL3 is the most widely studied methyltransferase and forms a core catalytic complex with METTL14, which primarily enhances its enzymatic activity for m6A deposition.8 WTAP, acts as a regulatory subunit, guiding the complex to target RNAs and stabilizing its catalytic function,9 RBM15, an adaptor protein, directs the complex to uracil-rich mRNA regions.10 Well-known m6A demethylases include ALKBH511 and FTO,12 which remove m6A from RNA. M6A binding proteins, known as “readers”, are classified into nuclear and cytoplasmic proteins based on their subcellular localization. These proteins play a pivotal role in recognizing and binding methylated RNA, thereby regulating key post-transcriptional processes, including mRNA splicing, translation, stability, and degradation.13–15
As an m6A methyltransferase, RBM15 exerts a variety of physiological functions, including a critical role in hematopoiesis. In hematopoiesis, RBM15 plays a vital role by regulating hematopoietic cell differentiation, maintaining the quiescent state of hematopoietic stem cells (HSCs), and influencing cell fate decisions. It promotes B lymphocyte development while inhibiting myeloid differentiation. In addition to its role in hematopoiesis, RBM15 is also involved in the regulation of tumorigenesis, studies on the relationship between RBM15 and cancer have shown that dysregulated expression of RBM15 closely regulates tumor initiation and progression through various molecular mechanisms, including in hepatocellular carcinoma, lung cancer, cervical cancer, and acute leukemia, and is associated with patient clinical outcomes. RBM15 is implicated in critical aspects of tumorigenesis, such as angiogenesis within tumors, immune cells infiltrations, tumor cell proliferation, invasion, migration, and modulation of stemness.16 RBM15 was identified later than other m6A “writers” and serves as a critical component of the methyltransferase complex (MTC), it plays an essential role in directing the complex to uracil-rich regions of target mRNAs,10 Due to its relatively recent discovery, the precise role of RBM15 within the m6A modification landscape remains to be fully elucidated and warrants further investigation. Understanding its function is crucial for elucidating the impact of m6A regulation in cancer and for advancing our knowledge of tumor biology. Numerous studies utilizing bioinformatics analyses have uncovered that RBM15 exhibits copy number variations and abnormal expression across multiple cancers, which are significantly associated with overall survival rates. Risk models based on the expression profiles of m6A-modulating genes, including RBM15, can be effectively utilized for early diagnosis of cancer and patient prognosis.17 Consequently, a comprehensive and systematic review of the association between RBM15 and cancer is imperative. This review synthesizes the current understanding of RBM15’s structure, function, and biological implications in cancer, and to explore its influence on cancer progression, therapeutic strategies, and prognostic outcomes. This study aims to elucidate the molecular mechanisms influenced by RBM15, thereby providing new insights into targeted cancer therapy and prognostic improvement.
Protein Structure of RBM15
The gene encoding RBM15 protein is also known as OTT (ovary testis transcribed), OTT1 (ovary testis transcribed 1), and SPEN (Split end) in humans, and KIAA1438 in mice. It encodes an RNA-binding protein with a molecular weight of 105.7 kDa. RBM15, along with its homolog RBM15B, serves as a crucial component of the m6A methyltransferase complex. Although RBM15 itself lacks catalytic activity, both RBM15 and RBM15B can interact with WTAP and the catalytic subunit METTL3 within the m6A methyltransferase complex, guiding these proteins to specific RNA sites for m6A modification.18–20
RNA-binding proteins (RBPs) typically possess one or more RNA-binding domains, including RNA recognition domains (RBDs, also known as RNP domains), K homology (KH) domains (types I and II), RGG (Arg-Gly-Gly) boxes, Sm domains, DEAD/DEAH boxes, zinc fingers (ZnFs, primarily C-x8-X5-X-x3-H), double-stranded RNA-binding domains (dsRBDs), and Pumilio/FBF (PuF or Pum-HD) domains.21,22 Cooperative interactions among these RNA-binding domains determine protein specificity for RNA sequences. RBPs are highly diverse, comprising over 200 different types, including heterogeneous nuclear ribonucleoproteins (hnRNPs), alternative splicing regulators (such as members of the SPEN family), and components of small nuclear ribonucleoproteins (snRNPs). A common feature of these proteins is the presence of RNA recognition motifs (RRMs). RBM15, a member of the SPEN (Split end) family, is involved in regulating alternative splicing. The SPEN (Split end) family, also known as the RRM Spen family, is characterized by multiple RRM domains and a highly conserved SPOC domain. Despite their structural similarities, different members of the family have distinct functions. For example, SHARP (SMART/HDAC1-associated repressor protein) binds to corepressors SMRT/NCoR to inhibit transcription, while OTT3 (RBM15B) and OTT1 (RBM15) interact with spliceosomes and the nuclear export factor NXF1 to regulate splicing and export.23 RBM15 plays a crucial role in normal development in animals. Both RBM15 and RBM15B contain three RNA recognition motifs (RRMs) and are notably enriched at m6A methylation sites in adjacent regions. Despite their similar sequence and domain organization, suggesting that RBM15B might compensate for RBM15, their biological functions differ.24 Notably, RBM15 knockout in mouse embryos is lethal, and RBM15B cannot compensate for the loss of RBM15.25
Physiological Function of RBM15
RBM15 was first identified and named in the context of acute megakaryoblastic leukemia with the t (1;22) translocation,26 Subsequent research has explored its critical role in hematopoietic development. Evidence suggests that RBM15 may inhibit myeloid differentiation in hematopoietic cells by stimulating Notch signaling through RBPJ. Its function is cell-type specific: it promotes Notch-induced HES1 transcription in hematopoietic cell lines but suppresses this activity in non-hematopoietic cells;27 Consistent with this, further studies have shown that RBM15-deficient mice exhibit peripheral B cell loss due to impaired pre-B cell differentiation, alongside an expansion of myeloid and megakaryocytic cells in the spleen and bone marrow. This results in a shift of progenitor cell fate towards granulocyte differentiation. Thus, RBM15 is essential for B lymphocyte hematopoiesis and exerts an inhibitory effect in the bone marrow, megakaryocytes, and progenitor cells, influencing the fate and expansion of multipotent hematopoietic cells.28 RBM15 may also influence the development of hematopoietic stem cells (HSCs) by regulating c-Myc expression. This is evidenced by the increased long-term (LT) HSCs observed in both Myc-KO and RBM15-KO mice, as well as the production of abnormal, small-sized megakaryocytes in mutant progenitor cells. The increased megakaryocytes in RBM15-KO mice can be partially reversed by ectopic expression of c-Myc.29 Additionally, while conditional RBM15-deficient mice can maintain lifelong hematopoiesis, their HSCs exhibit a significant reduction in the G0 phase of the cell cycle and early onset of exhaustion under stress conditions. This suggests that RBM15 plays a crucial role in maintaining the quiescent state of HSCs. Furthermore, RBM15-deficient and aging HSCs share several common features, including reduced tolerance to replication stress, increased myeloid potential, elevated absolute numbers, and increased higher levels of reactive oxygen species.30 This may be attributed to RBM15’s role in regulating the alternative splicing of the Mpl-FL isoform into the dominant-negative Mpl-TR, thereby inhibiting HSC proliferation and dampening THPO signal transduction.31 These findings highlight RBM15’s critical role not only in early hematopoietic development but also in maintaining the quiescent hematopoietic environment under specific conditions even after maturation.
Research indicates that embryos with RBM15 gene deletion exhibit embryonic lethality by E10.5, highlighting its essential role in development. Rescue experiments with RBM15-deficient embryos confirm that RBM15 is essential for placental vascular branching, as well as for the development of the heart and spleen.32 Similarly, RBM15 influences nervous system development by mediating the degradation of BAF155 through methylation, which affects the layer formation of apical radial glial progenitor cells and consequently impacts cortical development,33 This suggests that RBM15 may have diverse, yet-to-be-discovered roles across various developmental systems. RBM15 has also been implicated in mediating the transcriptional silencing of lncRNA XIST,24 While some studies found that knocking down RBM15 does not block XIST-mediated gene silencing.34 Subsequent research demonstrated that both RBM15 and RBM15B show redundant functions in this process,24 This redundancy provides a plausible explanation for earlier findings. Interestingly, loss of RBM15 and METTL3/14 complex subunits through CRISPR/Cas9 has only a minor impact on XIST-mediated silencing.35 This may be due to the fact that, although the loss of the METTL3/14 complex can lead to secondary effects, the redundancy of RBM15B, along with variations in experimental models and silencing assays, may compensate for these changes and mask the effect of RBM15 knockdown on XIST silencing. These factors may collectively account for the observed discrepancies.36
RBM15 has been found to play a vital role in various diseases through m6A modification. Overexpression of RBM15 alleviates liver damage, inflammation, and oxidative stress in mice with non-alcoholic fatty liver disease. Conversely, RBM15 knockdown exacerbates these conditions, likely due to RBM15’s upregulation of RNF5, which subsequently ubiquitinates and inhibits ROCK1 expression.37 Similarly, silencing RBM15 in COVID-19 models significantly reduces lymphocyte death in vitro and markedly suppresses the expression of multiple target genes associated with programmed cell death. RBM15 modulates the immune response to SARS-CoV-2 by increasing the expression levels of these target genes, suggesting its potential as a therapeutic target for COVID-19.38 Additionally, offspring of mothers with gestational diabetes are more susceptible to impaired glucose intolerance and insulin resistance, which may be due to RBM15’s role in epigenetically repressing CLDN4, thereby increasing insulin resistance and reducing insulin sensitivity.39 Recent studies also indicate that RBM15 expression is significantly elevated in patients with myocardial infarction. RBM15 overexpression reduces myocardial cell apoptosis and improves cardiac function in post-infarction mice.40 In aortic aneurysm and dissection (AD), RBM15 upregulates glycolysis in macrophages and promotes AD progression by enhancing M1 macrophage polarization.41 These findings suggest that differential expression of RBM15 in various diseases may result from different mechanistic pathways. A summary of the pathophysiological functions of RBM15 and its associated molecular pathways is presented in Table 1.
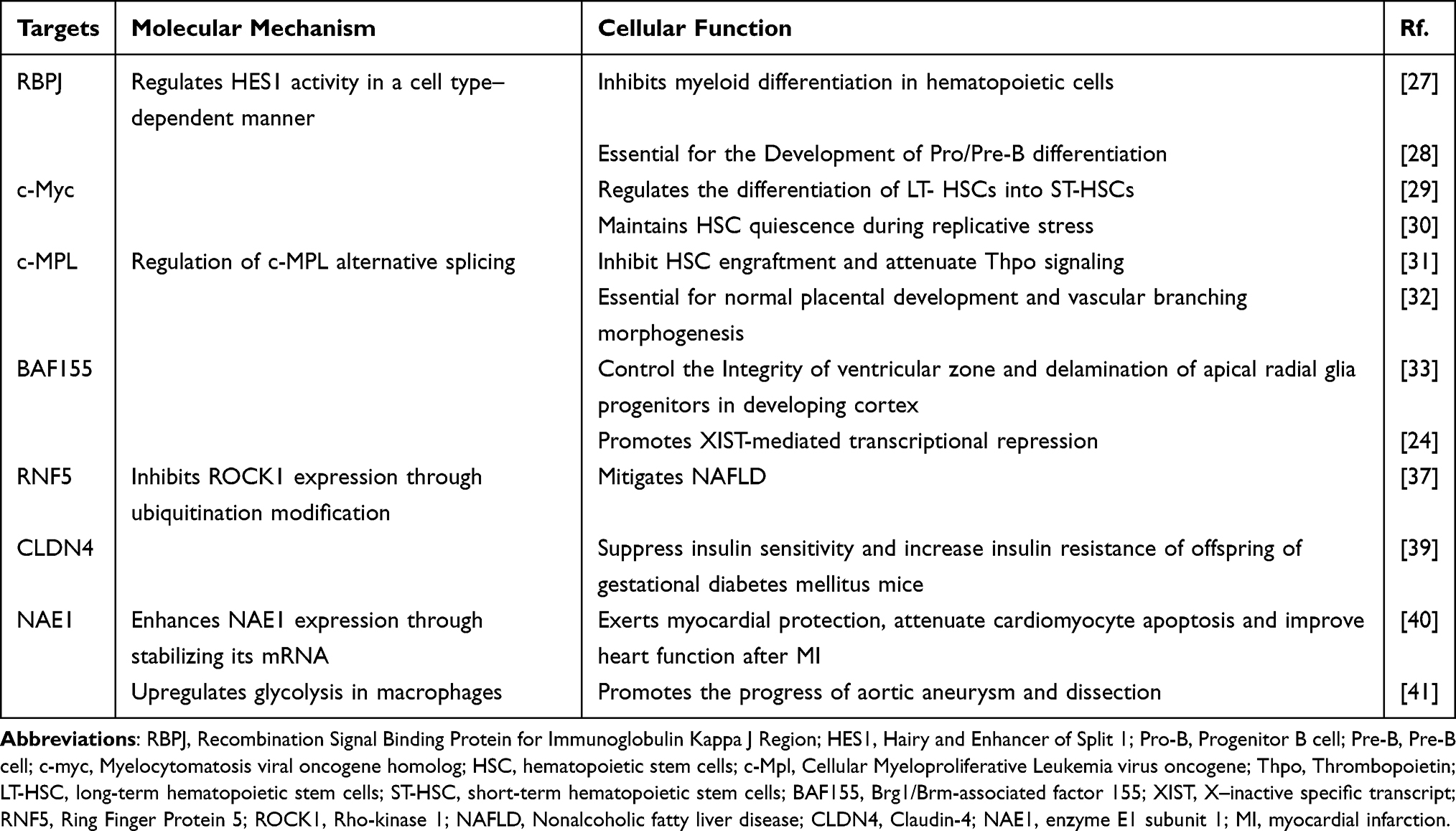 |
Table 1 The Physiological Function of RBM15 |
Overall, RBM15 is a pivotal RNA-binding protein whose structural configuration directly dictates its broad range of intracellular functions. It comprises three highly conserved RNA recognition motifs (RRMs) and a C-terminal SPOC (Spen paralog and ortholog C-terminal) domain. The RRMs primarily mediate the binding of RBM15 to uridine-rich sequences, thereby guiding it to specific target transcripts. The SPOC domain facilitates interactions with phosphorylated WTAP and the C-terminal domain (CTD) of RNA polymerase II, promoting the recruitment of the m^6A methyltransferase complex, including METTL3 and METTL14, to enable site-specific mRNA methylation. This structure–function coupling positions RBM15 as a key regulator of gene expression, alternative splicing, and mRNA nuclear export. Moreover, the SPOC domain also engages with nuclear export factor NXF1 and the RNA helicase DBP5, facilitating the efficient export of mRNA through the nuclear pore complex. Such multidomain versatility allows RBM15 to integrate a variety of RNA metabolic processes across diverse cellular contexts through domain-specific interactions. Although the functional consequences of RBM15 deficiency in adult hematopoiesis have been extensively characterized, a direct association between these phenotypic outcomes and changes in RNA methylation remains unconfirmed. Further investigations are warranted to elucidate how RBM15’s structural features mechanistically contribute to its roles in hematopoietic regulation. Unraveling these potential cross-cutting molecular mechanisms and how different structures function in different cellular environments could provide new therapeutic targets for related diseases.
RBM15 Functions in Tumor
In most cases, RBM15 functions as an oncogene, driving the initiation and progression of hematological malignancies and various solid tumors by modifying key transcriptional pathways through m6A methylation. Overall, as a component of the m6A methyltransferase complex, RBM15 frequently regulates tumor progression by mediating m6A modifications on key cancer-related mRNAs, thereby affecting their stability, translational efficiency, and decay (Figure 1).
 |
Figure 1 Molecular mechanisms underlying regulation of RBM15 on target genes in cancers. RBM15 methylates target transcripts, and the resulting m6A marks recruit m6A reader proteins to determine RNA fate. The target genes of RBM15 are indicated by underlined text. |
Acute Leukemia
Previous studies have shown that in acute megakaryoblastic leukemia (AMKL) characterized by the t(1;22)(p13;q13) translocation, a novel RBM15-MKL1 (Megakaryoblastic leukemia protein-1) fusion gene is formed by the joining of RBM15 and MKL1. This fusion protein disrupts RNA processing and the Hox and Ras/MAPK signaling pathways, leading to alterations in megakaryocyte proliferation and differentiation, thereby promoting leukemia development.26 Under normal conditions, MKL1 is typically localized in the cytoplasm.42 However, the RBM15-MKL1 fusion protein is exclusively nuclear, indicating that the aberrant fusion drastically alters MKL1’s subcellular localization. This mislocalization results in the loss of RBM15’s suppressive function and aberrant upregulation of MKL1-dependent transcriptional activation, ultimately contributing to the pathogenesis of AMKL.43 Interestingly, silencing the RBM15 gene typically results in the cytoplasmic depletion and nuclear accumulation of mRNA, suggesting that RBM15 plays a critical role in mRNA export.44 Furthermore, it has been observed that RBM15 expression is reduced in RBM15-MKL1 leukemia.45 Consistently, PRMT1 overexpression in AMKL cell lines reduces RBM15 protein levels. PRMT1 mediates this effect by methylating RBM15 at the R578 site, leading to its ubiquitin-mediated degradation. Notably, restoring RBM15 protein levels can rescue megakaryocyte terminal differentiation inhibited by PRMT1 overexpression.46
Cervical Carcinoma
It has been reported that RBM15 mediates the upregulation of the oncogene OTUB2, which promotes cervical cancer progression through the AKT/mTOR pathway.47 Additionally, two other studies have consistently found that RBM15 knockdown inhibits the proliferation of cervical cancer cells by affecting the JAK/STAT pathway and decorin (DCN) expression, and it also suppresses epithelial-to-mesenchymal transition (EMT) in cervical cancer. High levels of RBM15 are associated with poor clinical outcomes.48–50 Furthermore, the study indicates that HPV, a high-risk factor associated with cervical cancer, enhances the accumulation of RBM15 via HPV-E6. Specifically, RBM15 is degraded through autophagy, while HPV-E6 inhibits this process, resulting in increased RBM15 levels. This accumulation subsequently promotes the proliferation of cervical cancer cells by enhancing c-Myc protein levels.51
Liver Cancer
Research indicates that cFAM210A binds to YBX1 and inhibits its transactivation of the MET protein. cFAM210A expression is negatively correlated with tumor development in patients with HBV-associated hepatocellular carcinoma (HCC). However, the hepatitis B virus X protein (HBx) can promote the expression of RBM15, which in turn induces the degradation of cFAM210A, ultimately contributing to tumorigenesis.52
Colorectal Cancer
In colorectal cancer (CRC), RBM15 knockdown has been shown to inhibit tumor growth and liver metastasis. This effect is likely due to RBM15-mediated m6A modification of MyD88 mRNA, leading to reduced MyD88 expression, which is associated with poorer prognosis.53 However, another study found that MyD88 is highly expressed in CRC tissues and is linked to worse survival outcomes in CRC patients.54 This contradiction may be attributed to the dual role of MyD88 in CRC, its pro-tumorigenic effects are driven by cancer-associated inflammation and gut microbiota dysbiosis, which facilitate tumor invasion and promote the self-renewal capacity of cancer cells. In contrast, its anti-tumorigenic effects are achieved by preserving host–microbiota homeostasis, thereby inducing tumor cell cycle arrest and enhancing anti-tumor immune responses.55 Mechanistically, RBM15 knockdown increases the expression of KLF1 and SIN3 mRNAs, which, through their interaction with IGF2BP3, promote mRNA stability and subsequently enhance CRC cell proliferation.56
Renal Cancer
In clear cell renal carcinoma (ccRCC), the acetyltransferases EP300/CBP promote histone acetylation at the RBM15 promoter, thereby enhancing RBM15 transcription. RBM15, in turn, stabilizes CXCL11 mRNA in an m6A-dependent manner, which contributes to tumor growth, metastasis, and macrophage infiltration. Knockdown of RBM15 in ccRCC cells can inhibit EMT and reduce tumor metastasis.57 Additionally, from the perspective of cellular senescence, resveratrol has been found to affect the stability of CCNB1 mRNA mediated by RBM15 and inhibit cancer progression by suppressing EP300/CBP expression.58 These findings provide a basis for improving the prognosis of ccRCC and developing new combinatorial therapeutic strategies.
Prostate Cancer
AZGP1 has been identified as an independent predictor of biochemical recurrence following radical prostatectomy for prostate cancer,59–61 AZGP1P2, a pseudogene of AZGP1, has been found to act as a methyltransferase “writer” in castration-resistant prostate cancer (CRPC). It forms a complex with UBA1 and RBM15, regulating the ubiquitination and degradation of RBM15. This interaction decreases the stability of TPM1 mRNA, thereby inhibiting CRPC growth and metastasis. Moreover, RBM15 reduces CRPC’s sensitivity to docetaxel chemotherapy, suggesting that AZGP1P2 ultimately enhances CRPC’s sensitivity to docetaxel and exerts an antitumor effect.62 In summary, the research has unveiled a novel mechanism of docetaxel resistance CRPC, involving the AZGP1P2/UBA1/RBM15-TPM1-ERK1/2 signaling cascade. Conversely, a recent study has shown that overexpression of RBM15 significantly inhibits cell migration and proliferation, and it has proposed a new RBM15/IGF2BPs/DNMT1 cross-omic regulatory axis for m6A modulation.63 Whether the observed differences are due to new interactions between RNA m6A methylation and DNA methylation in the latter case requires further investigation. Additionally, a recent study indicates that RBM15 can also promote prostate cancer progression by regulating lncRNA expression.64
Lung Cancer
Studies have shown that upregulation of RBM15 in lung adenocarcinoma (LUAD) tissues is associated with poor patient survival outcomes.65 Additionally, RBM15 is linked to in vitro radiosensitivity by antagonizing SETD2.66 Knockdown of RBM15 inhibits LUAD cell proliferation, colony formation, migration, and invasion. Furthermore, the TGF-β activator (SRI-011381) reverses the suppressive effects of RBM15 silencing on tumor cell growth, possibly due to RBM15 silencing regulating the TGF-β/Smad2 pathway, thereby promoting ferroptosis in lung cancer cells.65,67 Additionally, studies have found that RBM15 suppresses RASSF8 protein levels through m6A modification and upregulates LINC01087, both of which enhance cell migration and invasion, thus accelerating LUAD progression.68,69 However, further research is needed to determine whether RBM15 can serve as a biomarker for lung cancer.
Osteosarcoma
RBM15 interacts with circular DNA, specifically with circ-CTNNB1, to promote the expression of hexokinase 2 (HK2), glucose-6-phosphate isomerase (GPI), and phosphoglycerate kinase 1 (PGK1) through N6-methyladenosine (m6A) modification. This interaction provides a metabolic survival advantage via aerobic glycolysis, thereby facilitating osteosarcoma progression.70 Interestingly, other studies have shown that in osteosarcoma (OS), RBM15 is negatively correlated with the expression of T cell γδ, while it is significantly positively correlated with natural killer (NK) cells, suggesting that RBM15 may serve as a potential immune target in OS.71
Other Cancers
Research on RBM15 in various tumors is relatively limited, with only a few relevant studies providing insights into its role, which may lead to discrepancies in findings. This review highlights some of these studies involving RBM15 and the involved mechanisms. For instance, in laryngeal squamous cell carcinoma, RBM15 mediates the m6A modification of TMBIM6 mRNA and enhances its stability in an IGF2BP3-dependent manner. This suggests that the RBM15/IGF2BP3/TMBIM6 axis may serve as a promising therapeutic target for laryngeal squamous cell carcinoma.72 Additionally, RBM15 expression has been reported to increase in testicular germ cell tumors73 and may also influence the progression of Kaposi’s sarcoma by promoting KHSV ORF59 expression.74 Moreover, a recent study in bladder cancer revealed that ENO1, an important glycolytic enzyme linked to tumorigenesis, is regulated by RBM15. RBM15 enhances the m6A modification of ENO1 mRNA through the Smad2/3 pathway, promoting tumor proliferation in bladder cancer.75 In another recent study, Su Hwan Park et al demonstrated that RBM15 levels are significantly elevated in basal-like breast cancer patients and are associated with poorer clinical outcomes, which is achieved through alterations in serine and glycine synthesis.76
The molecular regulatory pathways of RBM15 in cancers are shown in Table 2.
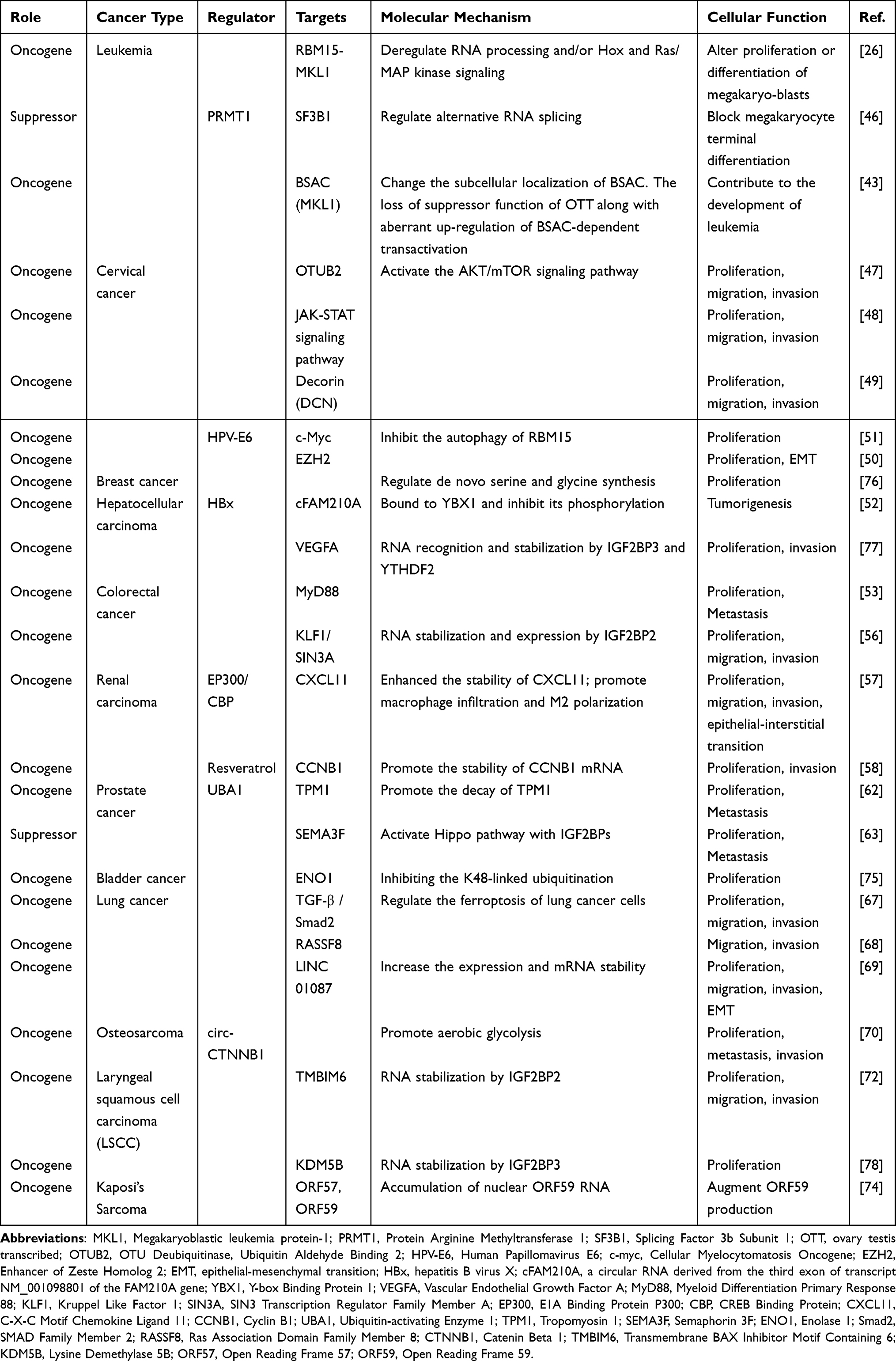 |
Table 2 Molecular Regulatory Pathways of RBM15 in Cancers |
Targeting of RBM15 for Potential Clinical Application
As previously mentioned, current research indicates that RBM15 primarily plays a pro-oncogenic role in cancer, while fewer studies demonstrated the anti-cancer effects of RBM15. Despite the limited available research, targeting RBM15 may hold clinical significance (Figure 2A–C). Notably, RBM15 promotes cell proliferation and inhibits myeloid differentiation in the normal hematopoietic system.27 Silencing RBM15 has been shown to enhance terminal differentiation of normal hematopoiesis, thereby inhibiting the proliferation of acute myeloid leukemia (AML). Interestingly, PRMT1 catalyzes the methylation of RBM15 at the R578 site, thereby promoting its ubiquitination and subsequent degradation. However, the enzymatic activity of PRMT1 V2 can enhance the stability of RBM15.46 To address the potential off-target effects of PRMT1 activator, the use of PRMT1 V1 isoform-specific activator may be an effective strategy46 (Figure 2E). The emergence of antisense RNA AS-RBM15 also offers a new avenue for research; studies have shown that AS-RBM15 enhances the translation of RBM15 protein in a CAP-dependent manner. Developing specific inhibitors targeting AS-RBM15 could more precisely influence the regulatory dynamics between RBM15 and AS-RBM1579 (Figure 2D). Additionally, reports indicate a direct interaction between RBM15-MKL1 and the Setd1b histone H3-Lys4 methyltransferase, which supports cell survival and subsequently enhances cell proliferation.45 Therefore, developing small molecule inhibitors and structural inhibitors that target the interaction between RBM15 and Setd1b may offer new therapeutic options for acute megakaryoblastic leukemia (AMKL). Similarly, recent studies have identified RBM15 as a critical component of the methyltransferase complex responsible for m6A modification of VEGFA, which is recognized and stabilized by IGF2BP3 and YTHDF2. Knockdown of RBM15 significantly inhibits tumor growth closely associated with VEGFA expression and angiogenesis, suggesting a potential alternative pathway for anti-angiogenic therapies targeting VEGFA.77 Furthermore, RBM15 is involved in tumor drug resistance; it promotes cisplatin resistance in laryngeal cancer by inhibiting ferroptosis. This indicates that combining cisplatin chemotherapy with RBM15 inhibitors could represent a more effective treatment strategy.78
 |
Figure 2 Clinical value and strategies of targeting RBM15 in cancer. (A) Targeting RBM15 to inhibit tumor angiogenesis by modulating VEGFA expression. (B) Targeting RBM15 to downregulate de novo serine and glycine biosynthesis in tumor tissues. (C) Targeting RBM15 to suppress glycolysis in tumor tissues. (D) Develop isoform-specific AS-RBM15 inhibitors to downregulate RBM15 translation and achieve targeted inhibition. (E) Design isoform-specific activators targeting PRMT1 V1 to induce ubiquitin-mediated degradation of RBM15. (F) Design bifunctional molecules leveraging PROTAC technology to induce the ubiquitin-mediated degradation of RBM15. Abbreviations: VEGFA, vascular endothelial growth factor A; PROTAC, Proteolysis-targeting chimeras. |
Clinically, high expression of RBM15 is significantly associated with poor overall survival in various cancer patients, including those with hepatocellular carcinoma(HCC),80 ovarian cancer,81 and pancreatic cancer.82 Thus, RBM15 holds promise as a prognostic marker across multiple tumors. Interestingly, inhibiting m6A modification can enhance tumor sensitivity to immunotherapy by altering the tumor microenvironment and increasing the recruitment of CD8+ tumor-infiltrating lymphocytes (TILs). In line with this, RBM15 has been reported to be negatively correlated with PD-L1,83 suggesting that the combined use of RBM15 inhibitors and anti-PD-L1 therapies may enhance the efficacy of immunotherapy. Moreover, inhibiting RBM15 has been shown to suppress pancreatic cancer progression and regulate macrophage phagocytosis, providing a more effective immunotherapeutic strategy for pancreatic cancer treatment.84 Reports indicate that RNA methyltransferases can influence the stability and expression of lncRNAs.85–87 Research has found that RBM15 promotes tumor stemness and enhances the stability of lncRNA HEIH, and lncRNA HEIH regulates EGFR expression through miR-802. Downregulation of miR-802 can impede the progression of cervical cancer.88 Therefore, RBM15 may serve as a potential target for cervical cancer therapy. Additionally, RBM15 contributes to the oncogenic activity of HPV and is a key factor in the growth of HPV-associated cervical cancer.51 Thus, targeting RBM15 could represent a novel approach for treating HPV-related cervical cancer.
A study analyzing the immune cell composition in osteosarcoma found a significant negative correlation between RBM15 and γδ T cells. Overexpression of the RBM15 gene led to reduced γδ T cell expression in osteosarcoma. Interestingly, RBM15 showed a significant positive correlation with activated natural killer (NK) cells, suggesting new directions for immunotherapy in osteosarcoma. This research also established a prognostic model based on the RNA-binding proteins RBM15 and YTHDC1, revealing that patients in the high-risk group had significantly lower survival rates compared to those in the low-risk group.71 These findings indicate that RBM15 and its m6A regulatory factors may serve as novel prognostic indicators for certain tumors.
Conclusions and Perspectives
M6A modification is one of the most prevalent RNA modifications, involving processes mediated by “writers”, “erasers”, and “readers”. Extensive research has highlighted its significant functional roles, not only in normal growth and development—such as hematopoiesis,27–31 embryonic development of the heart and spleen,32 and the transcriptional silencing of lncRNA XIST,24 but also in various human diseases, including azoospermia,89 heart failure,90 cancer.91 METTL3 is recognized as the sole catalytic subunit in the m6A methyltransferase complex (MTC), forming a methyltransferase complex in conjunction with METTL14.92,93 The primary role of METTL14 is to enhance the catalytic activity of this complex for m6A modification,8 whereas RBM15 contributes to this process by recruiting the MTC to U-rich regions of target mRNAs.10 Given the dual association of RBM15 with both drug resistance and favorable prognosis in certain cancers, the development of effective RBM15 inhibitors and activators holds promise as a novel anti-cancer strategy. Targeted therapies aimed at modulating RBM15, either alone or in combination with other agents, may offer new avenues for the treatment of various malignancies and other disease forms.
Previous investigations have primarily focused on the oncogenic role of RBM15, particularly its association with tumor proliferation, invasion, and migration. However, accumulating evidence suggests that RBM15 also contributes to other tumor-associated processes, including the inhibition of autophagy, angiogenesis, and metabolic regulation. Notably, some studies have revealed that RBM15 may exert tumor-suppressive effects in certain cancers and is associated with favorable prognostic outcomes. Whether these divergent roles stem from tissue specificity, variations in the key pathways through which RBM15 functions across different tumor types, or whether tumor-suppressor-related genes, within an immunosuppressive microenvironment, may paradoxically promote immune evasion.94 The underlying reasons for these divergent findings require further investigation. As a component of the MTC, RBM15’s functional dynamics within the complex warrant closer examination. Previous studies have demonstrated that METTL14, another core MTC component, exhibits tumor-suppressive activity in most cancer types.95 While this review focuses on RBM15, it is important to note that the METTL3–METTL14 heterodimer constitutes the core of the MTC, mediating m6A modifications. Research on the m6A-dependent roles of RBM15 should also consider the roles of METTL3 and METTL14. Nevertheless, how RBM15 interacts with other MTC components during tumor development, and how the interplay among MTC subunits collectively influences tumor progression and suppression, remains an open and compelling question.
In summary, RBM15 plays a pivotal role in various types of cancer, regardless of their dependency on m6A modification. Further investigations are warranted to develop RBM15-targeted therapies and promote their application in clinical settings. Existing studies suggest that RBM15 may undergo degradation via the ubiquitination pathway, highlighting the potential of designing bifunctional molecules using PROTAC (proteolysis-targeting chimera) technology to induce its ubiquitin-mediated degradation (Figure 2F).96 Finally, special attention should be paid to the dual regulatory role of RBM15 in tumor biology; the use of RBM15 activators or inhibitors should be approached with caution to avoid promoting tumorigenesis in other contexts.
Data Sharing Statement
No datasets were generated or analyzed during the current study.
Acknowledgment
Xiangyu Long and Bintao Hu are co-first authors for this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Desrosiers R, Friderici K, Rottman F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc Natl Acad Sci U S A. 1974;71(10):3971–3975. PubMed PMID: 4372599; PubMed Central PMCID: PMCPMC434308. doi:10.1073/pnas.71.10.3971
2. Wu Y, Xie L, Wang M, et al. Mettl3-mediated m(6)A RNA methylation regulates the fate of bone marrow mesenchymal stem cells and osteoporosis. Nat Commun. 2018;9(1):4772. PubMed PMID: 30429466; PubMed Central PMCID: PMCPMC6235890. doi:10.1038/s41467-018-06898-4
3. Frye M, Harada BT, Behm M, He C. RNA modifications modulate gene expression during development. Science. 2018;361(6409):1346–1349. PubMed PMID: 30262497; PubMed Central PMCID: PMCPMC6436390. doi:10.1126/science.aau1646
4. Barbieri I, Kouzarides T. Role of RNA modifications in cancer. Nat Rev Cancer. 2020;20(6):303–322. PubMed PMID: 32300195. doi:10.1038/s41568-020-0253-2
5. Wiener D, Schwartz S. The epitranscriptome beyond m(6)A. Nat Rev Genet. 2021;22(2):119–131. PubMed PMID: 33188361. doi:10.1038/s41576-020-00295-8
6. Roundtree IA, Evans ME, Pan T, He C. Dynamic RNA modifications in gene expression regulation. Cell. 2017;169(7):1187–1200. PubMed PMID: 28622506; PubMed Central PMCID: PMCPMC5657247. doi:10.1016/j.cell.2017.05.045
7. Zhao W, Qi X, Liu L, Ma S, Liu J, Wu J. Epigenetic regulation of m(6)A modifications in human cancer. Mol Ther Nucleic Acids. 2020;19:405–412.PubMed PMID: 31887551; PubMed Central PMCID: PMCPMC6938965. doi:10.1016/j.omtn.2019.11.022
8. Yang C, Hu Y, Zhou B, et al. The role of m(6)A modification in physiology and disease. Cell Death Dis. 2020;11(11):960. PubMed PMID: 33162550; PubMed Central PMCID: PMCPMC7649148. doi:10.1038/s41419-020-03143-z
9. Scholler E, Weichmann F, Treiber T, et al. Interactions, localization, and phosphorylation of the m(6)A generating METTL3-METTL14-WTAP complex. RNA. 2018;24(4):499–512. PubMed PMID: 29348140; PubMed Central PMCID: PMCPMC5855951. doi:10.1261/rna.064063.117
10. Huang W, Chen TQ, Fang K, Zeng ZC, Ye H, Chen YQ. N6-methyladenosine methyltransferases: functions, regulation, and clinical potential. J Hematol Oncol. 2021;14(1):117. PubMed PMID: 34315512; PubMed Central PMCID: PMCPMC8313886. doi:10.1186/s13045-021-01129-8
11. Zheng G, Dahl JA, Niu Y, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. 2013;49(1):18–29. PubMed PMID: 23177736; PubMed Central PMCID: PMCPMC3646334. doi:10.1016/j.molcel.2012.10.015
12. Duan X, Yang L, Wang L, et al. m6A demethylase FTO promotes tumor progression via regulation of lipid metabolism in esophageal cancer. Cell Biosci. 2022;12(1):60. PubMed PMID: 35568876; PubMed Central PMCID: PMCPMC9107638. doi:10.1186/s13578-022-00798-3
13. Wu R, Li A, Sun B, et al. A novel m(6)A reader Prrc2a controls oligodendroglial specification and myelination. Cell Res. 2019;29(1):23–41. PubMed PMID: 30514900; PubMed Central PMCID: PMCPMC6318280. doi:10.1038/s41422-018-0113-8
14. Huang H, Weng H, Sun W, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20(3):285–295. PubMed PMID: 29476152; PubMed Central PMCID: PMCPMC5826585. doi:10.1038/s41556-018-0045-z
15. Li F, Zhao D, Wu J, Shi Y. Structure of the YTH domain of human YTHDF2 in complex with an m(6)A mononucleotide reveals an aromatic cage for m(6)A recognition. Cell Res. 2014;24(12):1490–1492. PubMed PMID: 25412658; PubMed Central PMCID: PMCPMC4260351. doi:10.1038/cr.2014.153
16. Xu F, Guan Y, Ma Y, et al. Bioinformatic analyses and experimental validation of the role of m6A RNA methylation regulators in progression and prognosis of adrenocortical carcinoma. Aging. 2021;13(8):11919–11941. PubMed PMID: 33952721; PubMed Central PMCID: PMCPMC8109058. doi:10.18632/aging.202896
17. Zhang Q, Cai Y, Kurbatov V, et al. Gene alterations of N6-methyladenosine (m(6)A) regulators in colorectal cancer: a TCGA database study. Biomed Res Int. 2020;2020:8826456. PubMed PMID: 33415160; PubMed Central PMCID: PMCPMC7769650. doi:10.1155/2020/8826456
18. Wang T, Kong S, Tao M, Ju S. The potential role of RNA N6-methyladenosine in Cancer progression. Mol Cancer. 2020;19(1):88. PubMed PMID: 32398132; PubMed Central PMCID: PMCPMC7216508. doi:10.1186/s12943-020-01204-7
19. Knuckles P, Lence T, Haussmann IU, et al. Zc3h13/Flacc is required for adenosine methylation by bridging the mRNA-binding factor Rbm15/Spenito to the m(6)A machinery component Wtap/Fl(2)d. Genes Dev. 2018;32(5–6):415–429. PubMed PMID: 29535189; PubMed Central PMCID: PMCPMC5900714. doi:10.1101/gad.309146.117
20. Jenkins S, Addy M, Newcombe R. Studies on the effect of toothpaste rinses on plaque regrowth. (II). Triclosan with and without zinc citrate formulations. J Clin Periodontol. 1989;16(6):385–387. PubMed PMID: 2760251. doi:10.1111/j.1600-051x.1989.tb00009.x
21. Lunde BM, Moore C, Varani G. RNA-binding proteins: modular design for efficient function. Nat Rev Mol Cell Biol. 2007;8(6):479–490. PubMed PMID: 17473849; PubMed Central PMCID: PMCPMC5507177. doi:10.1038/nrm2178
22. Chen Y, Varani G. Protein families and RNA recognition. FEBS J. 2005;272(9):2088–2097. PubMed PMID: 15853794. doi:10.1111/j.1742-4658.2005.04650.x
23. Appel LM, Benedum J, Engl M, et al. SPOC domain proteins in health and disease. Genes Dev. 2023;37(5–6):140–170. PubMed PMID: 36927757; PubMed Central PMCID: PMCPMC10111866. doi:10.1101/gad.350314.122
24. Patil DP, Chen CK, Pickering BF, et al. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature. 2016;537(7620):369–373. PubMed PMID: 27602518; PubMed Central PMCID: PMCPMC5509218. doi:10.1038/nature19342
25. Uranishi H, Zolotukhin AS, Lindtner S, et al. The RNA-binding motif protein 15B (RBM15B/OTT3) acts as cofactor of the nuclear export receptor NXF1. J Biol Chem. 2009;284(38):26106–26116. PubMed PMID: 19586903; PubMed Central PMCID: PMCPMC2758010. doi:10.1074/jbc.M109.040113
26. Ma Z, Morris SW, Valentine V, et al. Fusion of two novel genes, RBM15 and MKL1, in the t(1;22)(p13;q13) of acute megakaryoblastic leukemia. Nat Genet. 2001;28(3):220–221. PubMed PMID: 11431691. doi:10.1038/90054
27. Ma X, Renda MJ, Wang L, et al. Rbm15 modulates Notch-induced transcriptional activation and affects myeloid differentiation. Mol Cell Biol. 2007;27(8):3056–3064. PubMed PMID: 17283045; PubMed Central PMCID: PMCPMC1899951. doi:10.1128/MCB.01339-06
28. Raffel GD, Mercher T, Shigematsu H, et al. Ott1(Rbm15) has pleiotropic roles in hematopoietic development. Proc Natl Acad Sci U S A. 2007;104(14):6001–6006. PubMed PMID: 17376872; PubMed Central PMCID: PMCPMC1851606. doi:10.1073/pnas.0609041104
29. Niu C, Zhang J, Breslin P, Onciu M, Ma Z, Morris SW. c-Myc is a target of RNA-binding motif protein 15 in the regulation of adult hematopoietic stem cell and megakaryocyte development. Blood. 2009;114(10):2087–2096. PubMed PMID: 19542301; PubMed Central PMCID: PMCPMC2744570. doi:10.1182/blood-2009-01-197921
30. Xiao N, Jani K, Morgan K, et al. Hematopoietic stem cells lacking Ott1 display aspects associated with aging and are unable to maintain quiescence during proliferative stress. Blood. 2012;119(21):4898–4907. PubMed PMID: 22490678; PubMed Central PMCID: PMCPMC3367894. doi:10.1182/blood-2012-01-403089
31. Xiao N, Laha S, Das SP, Morlock K, Jesneck JL, Raffel GD. Ott1 (Rbm15) regulates thrombopoietin response in hematopoietic stem cells through alternative splicing of c-Mpl. Blood. 2015;125(6):941–948. PubMed PMID: 25468569; PubMed Central PMCID: PMCPMC4319236. doi:10.1182/blood-2014-08-593392
32. Raffel GD, Chu GC, Jesneck JL, et al. Ott1 (Rbm15) is essential for placental vascular branching morphogenesis and embryonic development of the heart and spleen. Mol Cell Biol. 2009;29(2):333–341. PubMed PMID: 18981216; PubMed Central PMCID: PMCPMC2612519. doi:10.1128/MCB.00370-08
33. Xie Y, Castro-Hernandez R, Sokpor G, et al. RBM15 modulates the function of chromatin remodeling factor BAF155 through RNA methylation in developing cortex. Mol Neurobiol. 2019;56(11):7305–7320. PubMed PMID: 31020615. doi:10.1007/s12035-019-1595-1
34. McHugh CA, Chen CK, Chow A, et al. The Xist lncRNA interacts directly with SHARP to silence transcription through HDAC3. Nature. 2015;521(7551):232–236. PubMed PMID: 25915022; PubMed Central PMCID: PMCPMC4516396. doi:10.1038/nature14443
35. Nesterova TB, Wei G, Coker H, et al. Systematic allelic analysis defines the interplay of key pathways in X chromosome inactivation. Nat Commun. 2019;10(1):3129. PubMed PMID: 31311937; PubMed Central PMCID: PMCPMC6635394. doi:10.1038/s41467-019-11171-3
36. Brockdorff N, Bowness JS, Wei G. Progress toward understanding chromosome silencing by Xist RNA. Genes Dev. 2020;34(11–12):733–744. PubMed PMID: 32482714; PubMed Central PMCID: PMCPMC7263139. doi:10.1101/gad.337196.120
37. Li S, Lian S, Cheng W, Zhang T, Gong X. The role of N6-methyladenosine methyltransferase Rbm15 in nonalcoholic fatty liver disease. Shock. 2024;61(2):311–321. PubMed PMID: 38150369. doi:10.1097/SHK.0000000000002294
38. Meng Y, Zhang Q, Wang K, et al. RBM15-mediated N6-methyladenosine modification affects COVID-19 severity by regulating the expression of multitarget genes. Cell Death Dis. 2021;12(8):732. PubMed PMID: 34301919; PubMed Central PMCID: PMCPMC8298984. doi:10.1038/s41419-021-04012-z
39. Fang J, Wu X, He J, et al. RBM15 suppresses hepatic insulin sensitivity of offspring of gestational diabetes mellitus mice via m6A-mediated regulation of CLDN4. Mol Med. 2023;29(1):23. PubMed PMID: 36803098; PubMed Central PMCID: PMCPMC9942341. doi:10.1186/s10020-023-00615-8
40. Cheng H, Wu J, Li L, et al. RBM15 protects from myocardial infarction by stabilizing NAE1. JACC Basic Transl Sci. 2024;9(5):631–648. PubMed PMID: 38984049; PubMed Central PMCID: PMCPMC11228393. doi:10.1016/j.jacbts.2024.01.017
41. Tang M, Wang M, Wang Z, Jiang B. RBM15 activates glycolysis in M1-type macrophages to promote the progression of aortic aneurysm and dissection. Int J Med Sci. 2024;21(10):1976–1989. PubMed PMID: 39113895; PubMed Central PMCID: PMCPMC11302562. doi:10.7150/ijms.97185
42. Miralles F, Posern G, Zaromytidou AI, Treisman R. Actin dynamics control SRF activity by regulation of its coactivator MAL. Cell. 2003;113(3):329–342. PubMed PMID: 12732141. doi:10.1016/s0092-8674(03)00278-2
43. Sawada T, Nishiyama C, Kishi T, et al. Fusion of OTT to BSAC results in aberrant up-regulation of transcriptional activity. J Biol Chem. 2008;283(39):26820–26828. PubMed PMID: 18667423; PubMed Central PMCID: PMCPMC3258922. doi:10.1074/jbc.M802315200
44. Zolotukhin AS, Uranishi H, Lindtner S, Bear J, Pavlakis GN, Felber BK. Nuclear export factor RBM15 facilitates the access of DBP5 to mRNA. Nucleic Acids Res. 2009;37(21):7151–7162. PubMed PMID: 19786495; PubMed Central PMCID: PMCPMC2790900. doi:10.1093/nar/gkp782
45. Lee JH, Skalnik DG. Rbm15-Mkl1 interacts with the Setd1b histone H3-Lys4 methyltransferase via a SPOC domain that is required for cytokine-independent proliferation. PLoS One. 2012;7(8):e42965. PubMed PMID: 22927943; PubMed Central PMCID: PMCPMC3424240. doi:10.1371/journal.pone.0042965
46. Zhang L, Tran NT, Su H, et al. Cross-talk between PRMT1-mediated methylation and ubiquitylation on RBM15 controls RNA splicing. Elife. 2015;4. PubMed PMID: 26575292; PubMed Central PMCID: PMCPMC4775220. doi:10.7554/eLife.07938
47. Song Y, Wu Q. RBM15 m(6) A modification-mediated OTUB2 upregulation promotes cervical cancer progression via the AKT/mTOR signaling. Environ Toxicol. 2023;38(9):2155–2164. PubMed PMID: 37334762. doi:10.1002/tox.23852
48. Zhang C, Gu L, Xiao J, Jin F. Knockdown of RBM15 inhibits tumor progression and the JAK-STAT signaling pathway in cervical cancer. BMC Cancer. 2023;23(1):684. PubMed PMID: 37474926; PubMed Central PMCID: PMCPMC10360283. doi:10.1186/s12885-023-11163-z
49. Wang H, Li C, Wei Q, et al. RBM15 knockdown impairs the malignancy of cervical cancer by mediating m6A modification of decorin. Biochem Genet. 2024. PubMed PMID: 38429603. doi:10.1007/s10528-024-10757-x
50. Wang R, Tan W. RBM15-mediated N6-methyl adenosine (m6A) modification of EZH2 drives the epithelial-mesenchymal transition of cervical cancer. Crit Rev Eukaryot Gene Expr. 2024;34(5):15–29. PubMed PMID: 38842201. doi:10.1615/CritRevEukaryotGeneExpr.2024052205
51. Nie G, Tang B, Lv M, et al. HPV E6 promotes cell proliferation of cervical cancer cell by accelerating accumulation of RBM15 dependently of autophagy inhibition. Cell Biol Int. 2023;47(8):1327–1343. PubMed PMID: 37191290. doi:10.1002/cbin.12020
52. Yu J, Li W, Hou GJ, et al. Circular RNA cFAM210A, degradable by HBx, inhibits HCC tumorigenesis by suppressing YBX1 transactivation. Exp Mol Med. 2023;55(11):2390–2401. PubMed PMID: 37907737; PubMed Central PMCID: PMCPMC10689457. doi:10.1038/s12276-023-01108-8
53. Zhang Z, Mei Y, Hou M. Knockdown RBM15 inhibits colorectal cancer cell proliferation and metastasis via N6-Methyladenosine (m6A) modification of MyD88 mRNA. Cancer Biother Radiopharm. 2022;37(10):976–986. PubMed PMID: 34842457. doi:10.1089/cbr.2021.0226
54. Zhu G, Cheng Z, Huang Y, et al. MyD88 mediates colorectal cancer cell proliferation, migration and invasion via NF‑kappaB/AP‑1 signaling pathway. Int J Mol Med. 2020;45(1):131–140. PubMed PMID: 31746347; PubMed Central PMCID: PMCPMC6889924. doi:10.3892/ijmm.2019.4390
55. Wang L, Yu K, Zhang X, Yu S. Dual functional roles of the MyD88 signaling in colorectal cancer development. Biomed Pharmacother. 2018;107:177–184.PubMed PMID: 30086464. doi:10.1016/j.biopha.2018.07.139
56. Chen J. Regulatory mechanism of RNA binding motif protein 15-mediated N(6) methyladenosine modification in proliferation, invasion, and migration of colorectal cancer cells. Environ Toxicol. 2023;38(11):2545–2559. PubMed PMID: 37471637. doi:10.1002/tox.23883
57. Zeng X, Chen K, Li L, et al. Epigenetic activation of RBM15 promotes clear cell renal cell carcinoma growth, metastasis and macrophage infiltration by regulating the m6A modification of CXCL11. Free Radic Biol Med. 2022;184:135–147. PubMed PMID: 35381326. doi:10.1016/j.freeradbiomed.2022.03.031
58. Chen Z, Liu C, Ye T, Zhang Y, Chen Y. Resveratrol affects ccRCC cell senescence and macrophage polarization by regulating the stability of CCNB1 by RBM15. Epigenomics. 2023;15(18):895–910. PubMed PMID: 37909116. doi:10.2217/epi-2023-0150
59. Pearsey HM, Henson J, Sargeant JA, et al. Zinc-alpha2-glycoprotein, dysglycaemia and insulin resistance: a systematic review and meta-analysis. Rev Endocr Metab Disord. 2020;21(4):569–575. PubMed PMID: 32377863; PubMed Central PMCID: PMCPMC7557496. doi:10.1007/s11154-020-09553-w
60. Pelletier CC, Koppe L, Croze ML, et al. White adipose tissue overproduces the lipid-mobilizing factor zinc alpha2-glycoprotein in chronic kidney disease. Kidney Int. 2013;83(5):878–886. PubMed PMID: 23423258; PubMed Central PMCID: PMCPMC3938447. doi:10.1038/ki.2013.9
61. Liu Y, Wang T, Liu X, et al. Overexpression of zinc-alpha2-glycoprotein suppressed seizures and seizure-related neuroflammation in pentylenetetrazol-kindled rats. J Neuroinflammation. 2018;15(1):92. PubMed PMID: 29566716; PubMed Central PMCID: PMCPMC5863804. doi:10.1186/s12974-018-1132-6
62. Wang H, Liu J, Zhu X, Yang B, He Z, Yao X. AZGP1P2/UBA1/RBM15 cascade mediates the fate determinations of prostate cancer stem cells and promotes therapeutic effect of docetaxel in castration-resistant prostate cancer via TPM1 m6A modification. Research. 2023;6:0252.PubMed PMID: 37854295; PubMed Central PMCID: PMCPMC10581371. doi:10.34133/research.0252
63. Ying Y, Wu Y, Zhang F, et al. Co-transcriptional R-loops-mediated epigenetic regulation drives growth retardation and docetaxel chemosensitivity enhancement in advanced prostate cancer. Mol Cancer. 2024;23(1):79. PubMed PMID: 38658974; PubMed Central PMCID: PMCPMC11041046. doi:10.1186/s12943-024-01994-0
64. Hu B, Lin D, Liu Z, et al. Identification of RBM15 as a prognostic biomarker in prostate cancer involving the regulation of prognostic m6A-related lncRNAs. Eur J Med Res. 2024;29(1):411. PubMed PMID: 39118157; PubMed Central PMCID: PMCPMC11312177. doi:10.1186/s40001-024-02000-5
65. Ma M, Wang W, Wang B, et al. The prognostic value of N6-methyladenosine RBM15 regulators in lung adenocarcinoma. Cell Mol Biol. 2022;68(1):130–139. PubMed PMID: 35809319. doi:10.14715/cmb/2022.68.1.17
66. Zeng Z, Zhang J, Li J, et al. SETD2 regulates gene transcription patterns and is associated with radiosensitivity in lung adenocarcinoma. Front Genet. 2022;13:935601. PubMed PMID: 36035179; PubMed Central PMCID: PMCPMC9399372. doi:10.3389/fgene.2022.935601
67. Feng J, Li Y, He F, Zhang F. RBM15 silencing promotes ferroptosis by regulating the TGF-beta/Smad2 pathway in lung cancer. Environ Toxicol. 2023;38(4):950–961. PubMed PMID: 36715115. doi:10.1002/tox.23741
68. Ma M, Wang W, Li L, et al. RBM15 facilities lung adenocarcinoma cell progression by regulating RASSF8 stability through N6 Methyladenosine modification. Transl Oncol. 2024;46:102018. PubMed PMID: 38838436; PubMed Central PMCID: PMCPMC11214523. doi:10.1016/j.tranon.2024.102018
69. Zhang X, Wang DJ, Jia L, Zhang W. N6-methyladenosine-mediated LINC01087 promotes lung adenocarcinoma progression by regulating miR-514a-3p to upregulate centrosome protein 55. Kaohsiung J Med Sci. 2024;40(9):801–818. PubMed PMID: 39023191. doi:10.1002/kjm2.12879
70. Yang F, Liu Y, Xiao J, et al. Circ-CTNNB1 drives aerobic glycolysis and osteosarcoma progression via m6A modification through interacting with RBM15. Cell Prolif. 2023;56(1):e13344. PubMed PMID: 36181462; PubMed Central PMCID: PMCPMC9816931. doi:10.1111/cpr.13344
71. Jiang J, Qu H, Zhan X, et al. Identification of osteosarcoma m6A-related prognostic biomarkers using artificial intelligence: RBM15. Sci Rep. 2023;13(1):5255. PubMed PMID: 37002245; PubMed Central PMCID: PMCPMC10066227. doi:10.1038/s41598-023-28739-1
72. Wang X, Tian L, Li Y, et al. RBM15 facilitates laryngeal squamous cell carcinoma progression by regulating TMBIM6 stability through IGF2BP3 dependent. J Exp Clin Cancer Res. 2021;40(1):80. PubMed PMID: 33637103; PubMed Central PMCID: PMCPMC7912894. doi:10.1186/s13046-021-01871-4
73. Cong R, Ji C, Zhang J, et al. m6A RNA methylation regulators play an important role in the prognosis of patients with testicular germ cell tumor. Transl Androl Urol. 2021;10(2):662–679. PubMed PMID: 33718069; PubMed Central PMCID: PMCPMC7947426. doi:10.21037/tau-20-963
74. Majerciak V, Uranishi H, Kruhlak M, et al. Kaposi’s sarcoma-associated herpesvirus ORF57 interacts with cellular RNA export cofactors RBM15 and OTT3 to promote expression of viral ORF59. J Virol. 2011;85(4):1528–1540. PubMed PMID: 21106733; PubMed Central PMCID: PMCPMC3028919. doi:10.1128/JVI.01709-10
75. Shen C, Liu J, Xie F, et al. N6-Methyladenosine enhances the translation of ENO1 to promote the progression of bladder cancer by inhibiting PCNA ubiquitination. Cancer Lett. 2024;595:217002. PubMed PMID: 38823761. doi:10.1016/j.canlet.2024.217002
76. Park SH, Ju JS, Woo H, et al. The m(6)A writer RBM15 drives the growth of triple-negative breast cancer cells through the stimulation of serine and glycine metabolism. Exp Mol Med. 2024;56(6):1373–1387. PubMed PMID: 38825643; PubMed Central PMCID: PMCPMC11263342. doi:10.1038/s12276-024-01235-w
77. Xu X, Wu S, Zhang Y, et al. m6A modification of VEGFA mRNA by RBM15/YTHDF2/IGF2BP3 contributes to angiogenesis of hepatocellular carcinoma. Mol Carcinog. 2024;63(11):2174–2189. PubMed PMID: 39092767. doi:10.1002/mc.23802
78. Liang Y, Zhong H, Zhao Y, et al. Epigenetic mechanism of RBM15 in affecting cisplatin resistance in laryngeal carcinoma cells by regulating ferroptosis. Biol Direct. 2024;19(1):57. PubMed PMID: 39039611; PubMed Central PMCID: PMCPMC11264397. doi:10.1186/s13062-024-00499-6
79. Tran NT, Su H, Khodadadi-Jamayran A, et al. The AS-RBM15 lncRNA enhances RBM15 protein translation during megakaryocyte differentiation. EMBO Rep. 2016;17(6):887–900. PubMed PMID: 27118388; PubMed Central PMCID: PMCPMC5278610. doi:10.15252/embr.201541970
80. Fang Q, Chen H. The significance of m6A RNA methylation regulators in predicting the prognosis and clinical course of HBV-related hepatocellular carcinoma. Mol Med. 2020;26(1):60. PubMed PMID: 32552682; PubMed Central PMCID: PMCPMC7302147. doi:10.1186/s10020-020-00185-z
81. Wang Q, Zhang Q, Li Q, Zhang J, Zhang J. Clinicopathological and immunological characterization of RNA m(6) A methylation regulators in ovarian cancer. Mol Genet Genomic Med. 2021;9(1):e1547. PubMed PMID: 33225598; PubMed Central PMCID: PMCPMC7963423. doi:10.1002/mgg3.1547
82. Zhao Z, Ju Q, Ji J, Li Y, Zhao Y. N6-methyladenosine methylation regulator RBM15 is a potential prognostic biomarker and promotes cell proliferation in pancreatic adenocarcinoma. Front Mol Biosci. 2022;9:842833.PubMed PMID: 35223996; PubMed Central PMCID: PMCPMC8864094. doi:10.3389/fmolb.2022.842833
83. Liao X, Chen L, Liu J, et al. m(6)A RNA methylation regulators predict prognosis and indicate characteristics of tumour microenvironment infiltration in acute myeloid leukaemia. Epigenetics. 2023;18(1):2160134. PubMed PMID: 36567510; PubMed Central PMCID: PMCPMC9980463. doi:10.1080/15592294.2022.2160134
84. Wang W, He Y, Yao LC, et al. Identification of m6A modification patterns and RBM15 mediated macrophage phagocytosis in pancreatic cancer: an integrative analysis. Biochim Biophys Acta Mol Basis Dis. 2024;1870(7):167304. PubMed PMID: 38878830. doi:10.1016/j.bbadis.2024.167304
85. Li ZX, Zheng ZQ, Yang PY, et al. WTAP-mediated m(6)A modification of lncRNA DIAPH1-AS1 enhances its stability to facilitate nasopharyngeal carcinoma growth and metastasis. Cell Death Differ. 2022;29(6):1137–1151. PubMed PMID: 34999731; PubMed Central PMCID: PMCPMC9177844. doi:10.1038/s41418-021-00905-w
86. Meng L, Lin H, Huang X, Weng J, Peng F, Wu S. METTL14 suppresses pyroptosis and diabetic cardiomyopathy by downregulating TINCR lncRNA. Cell Death Dis. 2022;13(1):38. PubMed PMID: 35013106; PubMed Central PMCID: PMCPMC8748685. doi:10.1038/s41419-021-04484-z
87. Qin Y, Li L, Luo E, et al. Role of m6A RNA methylation in cardiovascular disease (Review). Int J Mol Med. 2020;46(6):1958–1972. PubMed PMID: 33125109; PubMed Central PMCID: PMCPMC7595665. doi:10.3892/ijmm.2020.4746
88. Quan Y, Zhou M, Li J, et al. The m6A methyltransferase RBM15 affects tumor cell stemness and progression of cervical cancer by regulating the stability of lncRNA HEIH. Exp Cell Res. 2024;436(2):113924. PubMed PMID: 38280435. doi:10.1016/j.yexcr.2024.113924
89. Chen Y, Wang J, Xu D, et al. m(6)A mRNA methylation regulates testosterone synthesis through modulating autophagy in Leydig cells. Autophagy. 2021;17(2):457–475. PubMed PMID: 31983283; PubMed Central PMCID: PMCPMC8007139. doi:10.1080/15548627.2020.1720431
90. Berulava T, Buchholz E, Elerdashvili V, et al. Changes in m6A RNA methylation contribute to heart failure progression by modulating translation. Eur J Heart Fail. 2020;22(1):54–66. PubMed PMID: 31849158. doi:10.1002/ejhf.1672
91. Liu T, Wei Q, Jin J, et al. The m6A reader YTHDF1 promotes ovarian cancer progression via augmenting EIF3C translation. Nucleic Acids Res. 2020;48(7):3816–3831. PubMed PMID: 31996915; PubMed Central PMCID: PMCPMC7144925. doi:10.1093/nar/gkaa048
92. Wang P, Doxtader KA, Nam Y. Structural Basis for Cooperative Function of Mettl3 and Mettl14 Methyltransferases. Mol Cell. 2016;63(2):306–317. PubMed PMID: 27373337; PubMed Central PMCID: PMCPMC4958592. doi:10.1016/j.molcel.2016.05.041
93. Wang X, Feng J, Xue Y, et al. Structural basis of N(6)-adenosine methylation by the METTL3-METTL14 complex. Nature. 2016;534(7608):575–578. PubMed PMID: 27281194. doi:10.1038/nature18298
94. Binnewies M, Roberts EW, Kersten K, et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat Med. 2018;24(5):541–550. PubMed PMID: 29686425; PubMed Central PMCID: PMCPMC5998822. doi:10.1038/s41591-018-0014-x
95. Guan Q, Lin H, Miao L, et al. Functions, mechanisms, and therapeutic implications of METTL14 in human cancer. J Hematol Oncol. 2022;15(1):13. PubMed PMID: 35115038; PubMed Central PMCID: PMCPMC8812173. doi:10.1186/s13045-022-01231-5
96. Wang Y, Jiang X, Feng F, Liu W, Sun H. Degradation of proteins by PROTACs and other strategies. Acta Pharm Sin B. 2020;10(2):207–238. PubMed PMID: 32082969; PubMed Central PMCID: PMCPMC7016280. doi:10.1016/j.apsb.2019.08.001
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.