Back to Journals » Infection and Drug Resistance » Volume 19
Research Progress on Antivirulence Agents Targeting the Accessory Gene Regulator (Agr) System of Staphylococcus Aureus
Authors Chen L, Huang M, Chen Y, Lin Y, Cong Y ![]()
Received 7 January 2026
Accepted for publication 13 March 2026
Published 24 March 2026 Volume 2026:19 594449
DOI https://doi.org/10.2147/IDR.S594449
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Hazrat Bilal
Lingwei Chen,1,2 Mintao Huang,1,2 Ying Chen,1,2 Yanzhen Lin,1,2 Yanguang Cong1– 4
1Dongguan Key Laboratory for Pathogenesis and Experimental Diagnosis of Infectious Diseases, The First Dongguan Affiliated Hospital, School of Medical Technology, Guangdong Medical University, Dongguan, 523710, People’s Republic of China; 2Department of Clinical Laboratory, The First Dongguan Affiliated Hospital, School of Medical Technology, Guangdong Medical University, Dongguan, 523710, People’s Republic of China; 3Dongguan Key Laboratory of Sepsis Translational Medicine, The First Dongguan Affiliated Hospital, Guangdong Medical University, Dongguan, 523710, People’s Republic of China; 4Guangdong Provincial Clinical Research Center for Laboratory Medicine, Guangzhou, 510280, People’s Republic of China
Correspondence: Yanguang Cong, Email [email protected]
Abstract: With global spread of drug-resistant strains such as methicillin-resistant Staphylococcus aureus, conventional antimicrobial strategies face significant challenges, which prompt the development of antiinfective agents with alternative strategies. The antivirulence approach targets bacterial virulence factors without directly killing the bacteria. The accessory gene regulator (Agr) system, a central quorum-sensing pathway in S. aureus, represents a primary target for such therapies. However, inhibition of the Agr system presents a dual nature: while it attenuates acute virulence, it may inadvertently promote biofilm formation and poses potential therapeutic trade-offs. This review provides an overview of the Agr system structure and functional mechanisms alongside recent advances in synthetic and natural quorum sensing inhibitors targeting the Agr system in S. aureus. By regulating primary virulence factors, this system serves as an ideal target for antivirulence therapies. Disrupting Agr can reduce pathogenicity without affecting bacterial growth. Current researches suggest that the Agr inhibitors hold significant potential as novel therapeutics, though their clinical application requires careful consideration of the dual effects.
Keywords: quorum sensing, Agr system, antivirulence, Agr inhibitors, quenching antibodies
Introduction
Staphylococcus aureus is a leading pathogen responsible for a wide range of infections from skin/soft tissue infections to life-threatening conditions such as pneumonia and sepsis.1 Over the past several decades, global spread of multidrug resistant strains, particularly methicillin-resistant S. aureus (MRSA), has posed significant challenges in the clinical treatment of infections caused by multidrug resistant S. aureus.1,2 MRSA infections cause over 100,000 deaths annually worldwide.3 Despite the development of novel antibiotics, the rapid evolution of drug resistance has significantly diminished the efficacy of traditional bactericidal strategies.4 Therefore, there is an urgent need to develop new antimicrobial approaches.
Quorum sensing (QS) is a crucial regulatory mechanism through which bacteria coordinate group behaviors by secreting and sensing signaling molecules.5 In S. aureus, QS plays a critical role in regulating the expression of virulence factors and biofilm formation, which are essential for establishing and maintaining infections.6,7 Interference with QS can markedly attenuate pathogenicity without inhibiting growth, thereby reducing selective pressure and delays resistance development.8 Unlike many Gram-negative bacteria that possess multiple QS circuits, S. aureus relies on a single system, the accessory gene regulator (Agr), which detects autoinducing peptide (AIP) gradients and dynamically modulates virulence gene expression.9,10 By contrast, Gram-negative pathogens, such as Pseudomonas aeruginosa, employ multiple interconnected QS systems (eg., las, rhl, and pqs systems), which complicate inhibitor design and may allow compensatory pathway activation upon single-target inhibition.11,12 The structural simplicity of the Agr system thus offers a more straightforward target for therapeutic intervention, with reduced risk of resistance bypass mechanisms. These characteristics make the Agr system a promising target for antivirulence therapy.
Compared with traditional bactericidal strategies, antivirulence approaches possess unique advantages.13 First, these approaches alleviate collateral damage to host tissues by inhibiting virulence factor rather than directly killing bacteria. Second, this disarmament strategy exerts lower selection pressure, and delays resistance emergence. Finally, Agr inhibitors may synergize with existing antibiotics to enhance their therapeutic efficacy.14 Among antivirulence targets, the Agr-targeted therapy occupies a distinctive position among antivirulence strategies: unlike direct virulence factor neutralization or ica-targeted anti-biofilm approaches that address single phenotypes, Agr inhibition offers broad-spectrum virulence suppression at the cost of potential biofilm enhancement;13,15 Conversely, alternative quorum sensing targets such as the sae system yield more modest effects due to their less central regulatory roles.16 The present article reviews the components and functions of the Agr system as well as recent advances in S. aureus quorum sensing inhibitors (QSIs), and provides perspectives on future development in this area.
Structure and Function of the Agr System
The agr gene cluster in S. aureus comprises two transcriptional units: RNAII and RNAIII. The RNAII operon consists of four genes (agrA, agrB, agrC, and agrD), which encode the core components of the Agr system. Specifically, agrD encodes the precursor of the autoinducing peptide; agrB encodes a membrane-bound protease responsible for processing and secreting the AIP precursor; while agrC and agrA encode a histidine kinase and a response regulator protein, respectively, which form a two-component signal transduction system. RNAIII serves as the primary effector molecule, which regulates the expression of target genes including those encoding virulence factors.5,9,10,17
AIP biosynthesis begins with the precursor peptide encoded by agrD, which is processed by AgrB and MroQ to produce a mature cyclic peptide.18 When AIP concentrations reach a threshold level, AIP binds to the transmembrane receptor AgrC, and triggers its autophosphorylation and subsequent transfer of the phosphate group to AgrA.19 Activated AgrA acts as a transcription factor that promotes the transcription of both RNAII and RNAIII, thereby establishing a positive feedback loop that amplifies the signal cascade20 (Figure 1).
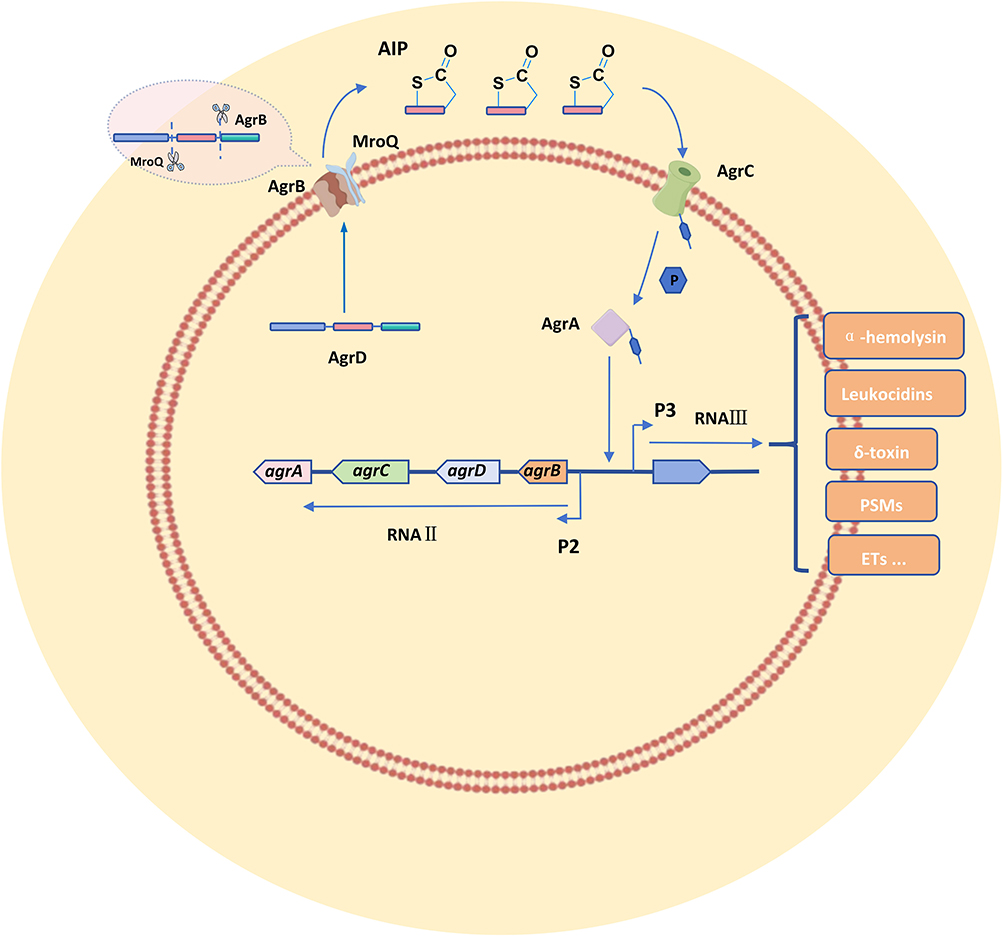 |
Figure 1 Regulatory mechanism of the Agr system in Staphylococcus aureus. The RNAII operon includes four genes: agrA, agrB, agrC, and agrD. The agrD gene encodes the AIP precursor (AgrD), which is processed by AgrB (the membrane-bound protease encoded by agrB) and MroQ to generate mature AIP. Upon reaching a threshold concentration, AIP activates the AgrC-AgrA two-component signal transduction system and drives positive feedback in RNAII transcription and RNAIII-mediated regulation of virulence genes such as α-hemolysin, leukocidins, δ-toxin, pore-forming small molecules (PSMs), and enterotoxins (ETs). |
The Agr system regulates the expression of associated genes through RNAIII. During the early stages of infection, when Agr activity is low, surface proteins such as Protein A and fibronectin-binding proteins are expressed and facilitate bacterial adhesion and colonization.21 As AIP concentrations increase, the activated Agr system upregulates secreted virulence factors like α-hemolysin and phenol-soluble modulins (PSMs), which promote tissue invasion and immune evasion.22,23 Simultaneously, the Agr system dynamically controls biofilm formation and dispersal by modulating the ica operon and protease expression. This temporal regulation mechanism enables S. aureus to adaptively adjust its behavior across different infection stages and microenvironments.14,24
Given the central role of the Agr system in modulating S. aureus virulence, its genetic or pharmacological inactivation generally attenuates toxin production and lessens tissue damage in multiple animal models.25 However, the same mutation frequently enhances biofilm formation, which impairs neutrophil-mediated clearance and leads to persistent colonization or chronic infection. Moreover, rapid compensatory up-regulation of non-Agr virulence determinants and the possible selection of Agr-negative strains have been observed in patients receiving long-term intensive care.16 These contradictory outcomes underscore that Agr inhibition is not a universally beneficial intervention and highlight the need for carefully timed or combination therapy.
Research Progress on Agr System Inhibitors
Synthetic Quorum Sensing Inhibitors
AIP Analogs
The discovery of AIP analogs as QSIs can be traced back to 1997, when cross-inhibiting AIPs were first shown to antagonize AgrC by interfering with AIP recognition.26 This finding established competitive receptor binding as a viable strategy for blocking QS signaling. Although rapidly cleared in vivo, AIP-2 was found to prevent abscess formation by disrupting the transient Agr-dependent initiation window.27,28
However, peptide-based QSIs face inherent pharmacokinetic limitations with susceptibility to proteolytic degradation and poor metabolic stability. To overcome these challenges, Tal-Gan et al undertook a systematic synthetic study of AIP-III analogs. Among them, four lead AgrC inhibitors (AIP-III D4A, tAIP-III D2A, AIP-III N2A/D4A, and AIP-III I1A/N2A/D4A) reduced the production of toxic shock syndrome toxin-1, a QS-regulated virulence factor, by over 80% at nanomolar concentrations. Recognizing that the thioester linkage in native AIPs is prone to hydrolysis, they subsequently developed amide-linked derivatives to enhance stability and solubility while retaining potent inhibitory activity against the AgrC receptor in S. aureus.29,30 Starting with t-AIP-II, they identified a peptidomimetic compound, n7OFF, which shares amphiphilic characteristics with native AIP-II and AIP-III D4A.31 Through further replacement of labile peptide bonds with non-hydrolyzable linkages, subsequent structural modifications of n7OFF yielded the highly active Agr inhibitor PhPr(3Br)-Bnc3, which represents the most potent peptidomimetic-type Agr inhibitor across all four S. aureus groups reported to date.32,33
RNAIII Inhibiting Peptide (RIP) Analogs
The RIP is produced by coagulase negative Staphylococcus with characteristic sequence YSPXTNF, where X can be a cysteine, a tryptophan, or a modified amino acid. RIP and its derivatives suppress RNAIII transcription, which in turn inhibits the production of QS-regulated toxins. Synthetic RIP analogs serve as potent suppressors of Agr-regulated exotoxin production in S. aureus in vitro.34,35 RIP had been demonstrated efficacy in treating various animal infection models including cellulitis in mice, keratitis in rabbits, osteomyelitis in rabbits, mastitis in cows, and septic arthritis in mice.36,37 The RIP treatment significantly reduced bacterial pathology and delayed the onset of disease symptoms.
Cirioni, Simonetti et al synthesized novel RIP derivatives including FS3, FS8, and FS10, which effectively inhibit the QS system of S. aureus. In a rat vascular graft infection model, the heptapeptide FS3 significantly enhanced S. aureus susceptibility to daptomycin.38 In a murine wound infection model, the tetrapeptide FS10 markedly increased MRSA susceptibility to tigecycline.39 These data indicated the potential of these RIP derivatives as antibiotic enhancers against MRSA infections.
Active Molecules from Compound Library Screening
High-throughput screening of compound libraries is a widely adopted strategy for drug discovery. This approach encompasses diverse methodologies including FDA-approved drug repurposing, combinatorial synthesis, phytochemical screening, and innovative display technologies.
FDA-Approved Drug Repurposing
Palaniappan et al performed in silico molecular docking with the C-LytTR domain of AgrA to screen an FDA-approved drug library. They identified the diuretic bumetanide with binding ability to AgrA. Bumetanide suppressed the expression of α-hemolysin, phenol-soluble modulins, and Panton-Valentine leukocidin. Animal studies demonstrated its efficacy in controlling ulcer progression and promoting wound healing.40
Salicylic acid (SAL), the primary metabolite of aspirin, was shown to bind to AgrA, thereby downregulating Agr-associated genes including agrA, agrC, rnaIII, and psmα/psmβ.41 However, SAL stabilized the mature biofilms of S. aureus, which may facilitate the bacterial persistence leading to therapeutic failure.42 Subinhibitory visomitin, a small-molecule antioxidant, reduces hemolysis, staphylococcal toxins, and colony spreading.43 Gliptins, a novel class of antidiabetics that are dipeptidyl peptidase-4 inhibitors (DPP-4) used to improve β-cell health and control blood glucose levels in type 2 diabetes mellitus, have been identified by Khayat et al as possessing antivirulence and anti-QS activity against S. aureus and P. aeruginosa.44
Synthetic and Natural Product Libraries
Sully et al identified the small-molecule inhibitor savirin via high-throughput screening. Savirin suppresses QS by targeting AgrA and leads to reduced tissue damage and enhanced bacterial clearance in two murine skin infection models. Notably, long-term in vitro passaging and in vivo studies revealed no resistance or tolerance to savirin in S. aureus, highlighting its stable anti-QS activity.14
Yu et al constructed a 148-member biarylhydroxyketone library via combinatorial synthesis and identified the bioactive compound 4f-12, which inhibits AgrA-DNA binding in electrophoretic mobility shift assays. The compound 4f-12 reduced rabbit erythrocyte hemolysis by 98.1 ± 0.1% compared to control without the 4f-12 treatment.45 Kuo et al demonstrated that three biarylhydroxyketone compounds (F1, F12, F19) act as QS inhibitors by targeting the AgrA response regulator. In an MRSA murine wound model, these compounds accelerated wound healing via suppression of virulence factor expression. In a Galleria mellonella infection model, they prolonged host survival and restored MRSA susceptibility to β-lactam antibiotics.46
Yamaguchi et al screened 577 phytochemicals and identified physalin H, physalin B, and isophysalin B as novel Agr-QS modulators. These compounds inhibit S. aureus QS by binding to the DNA-binding domain of AgrA, thereby blocking its interaction with the P3 promoter region.47 Khodaverdian et al screened 90,000 small molecules from the National Cancer Institute library and identified biaryl compounds that block AgrA’s phosphate-binding site. These compounds dose-dependently inhibit α-hemolysin and PSMα production without affecting bacterial growth.48 Todd et al demonstrated that succinic acid reduces MRSA-induced abscess formation in mice and validated its in vivo QS inhibitory activity.49
Structure-Based and Display Technologies
Kim et al employed click chemistry to covalently link QS modulators to surfaces. The surface-PEG10000-triazole-TrAIP-II competitively inhibited QS responses.50 Xie et al utilized the RaPID system with lipid nanodiscs to screen >1012 cyclic peptides and identified QQ-1 to QQ-4 as AgrC-binding inhibitors. QQ-3 and QQ-5 (a desulfurized analog) effectively suppressed S. aureus-induced hemolysis and demonstrated strong antivirulence potential. Although these cyclic peptides lack the thiolactone scaffold characteristic of native AIPs, they still dock into the AgrC ligand-binding pocket, competitively displace AIP and silence AgrC-mediated quorum sensing.51
Fatty Acids and Halogenated Scaffolds
Among the C18 unsaturated fatty acids tested (petroselinic, vaccenic, and oleic acids), petroselinic acid markedly suppresses the production of toxins, lipase, and α-hemolysin by downregulating agrA, rnaIII, hla, nuc1/nuc2, and saeR.52 The halogenated pyrimidine 2,4-dichloro-5-fluoropyrimidine (24DC5FP) inhibits agrA, rnaIII, hla, nuc1, and saeR expression.53
Despite promising in vitro activity, these synthetic QSIs generally lack human pharmacokinetic characterization and long-term toxicity evaluation, limiting their translational readiness.
Nanomaterial Applications for QSIs
Although a large number of studies have shown that QSIs have promising application potential in antiinfective therapy, they still face limitations in safety, solubility, and stability. Researchers have enhanced their performance through structural optimization and the integration of nanotechnology.29,30 For example, Kratochvil et al developed nanoporous, polymer-based superhydrophobic coatings as a reservoir for water-sensitive peptide QSIs, and enabled their prolonged release. These QSIs were continuously released into the surrounding medium for more than 8 mouths and potently inhibited the Agr quorum-sensing system in S. aureus for at least 40 days.54 Chen et al developed nanocomposite coatings with dihydropyrrol-2-ones (DHP) analogues, which can be applied to surfaces of any length scale and abolish S. aureus’s adhesion55 (Table 1).
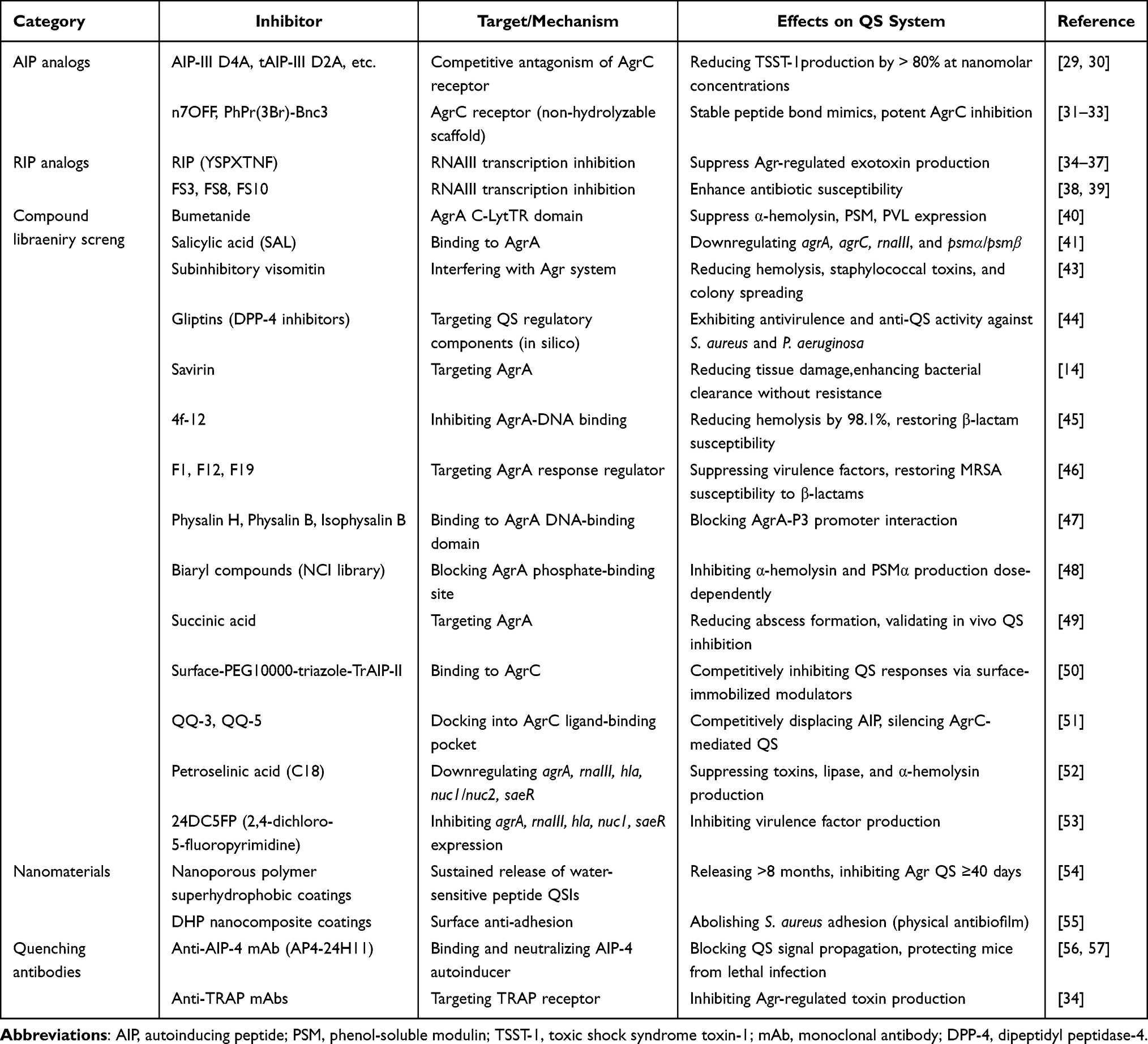 |
Table 1 Synthetic QS Inhibitors and Quorum Quenching Antibodies Targeting the S. aureus Agr System |
Nanomaterial-based delivery systems, while addressing solubility and stability limitations, introduce additional safety considerations regarding biodistribution, clearance, and potential inflammatory responses that remain incompletely characterized.
Natural Product-Derived QSIs
Evolutionary pressures from plant-microbe, microbe-microbe and animal-microbe interactions have driven the emergence of QSIs, which makes natural products a rich resource for the discovery of novel QSIs.
Microbial QSIs
A marine luminescent bacterium produces solonamides A and B, cyclic depsipeptides that interfere with the S. aureus Agr system and suppress QS-related genes including hla, rnaIII, and spa.58 Lactobacillus reuteri synthesizes cyclo (L-Phe-L-Pro) and cyclo (L-Tyr-L-Pro), both of which inhibit toxic shock syndrome toxin-1 (TSST-1) expression via Agr interference.59 An Arthrobacter strain, isolated from sandstone, produces arthroamide and turnagainlide A, both of which inhibit QS at submicromolar to micromolar concentrations.60
Desouky et al screened 54 actinomycete metabolites and identified phenalinolactones A-D, BU-4664LMe, 4,5-dehydrogeldamycin, and questinomycin A as inhibitors of QS and hemolysis.61 Bioactive peptides from Lactobacillus strains downregulate QS-controlled virulence mechanisms and restore MRSA sensitivity to cefoxitin when combined with β-lactams.62 Fungal cerebroside flavuside B reduces agrA expression in S. aureus and exhibits antioxidant and anti-inflammatory properties in S. aureus-infected skin wounds.63 The cyclodepsipeptide WS9326B, which is isolated from actinomycetes, suppresses S. aureus Agr activity and reduces corneal epithelial cell toxicity.64
Apicidin, a fungal metabolite, functions as a quorum-sensing inhibitor by directly antagonizing the AgrA response regulator of the S. aureus Agr system. This QS blockade suppresses the expression of major virulence determinants, thereby reduces bacterial pathogenicity in vivo. Consistently, apicidin treatment significantly decreased ulcer size and weight loss in murine MRSA skin infection models.65 Penicillium restrictum produces ω-hydroxyemodin (OHM), a polyhydroxyanthraquinone that binds AgrA to block its interaction with the P2 promoter. OHM treatment reduced skin necrosis, enhanced bacterial clearance, and lowered inflammatory cytokine levels in murine soft tissue infection models.66 The cyclic peptide avellanin C, derived from Hamigera ingelheimensis, inhibits QS signaling in S. aureus by targeting the agr P3 promoter, and potentially reduces the expression of Agr-regulated virulence factors.67
Plant-Derived QSIs
Medicinal plants are rich sources of QSIs. Echinatin, a natural compound derived from licorice, suppresses MRSA hemolysis via transcriptional regulation.68 Tanshinone IIB, a major bioactive compound extracted from the roots of Salvia miltiorrhiza, exhibits potent anti-virulence activity both in vitro and in vivo by reducing the expression of rnaIII and psmα.69 Thymol (2-isopropyl-5-methylphenol), a major constituent of thyme herb (Thymus vulgaris L.), possesses a wide spectrum of antimicrobial activity. Specifically, thymol reduces the production of PSMα and δ-toxin in S. aureus, and molecular docking studies indicate that thymol has binding affinity for the AgrA and AgrC receptors.70 Hamamelitannin, a polyphenol from Hamamelis virginiana bark, exerts anti-QS effects by inhibiting RNAIII without affecting bacterial growth. In rat graft models, it prevented device-related infections caused by MRSA and Staphylococcus epidermidis.71 Vermote et al demonstrated that hamamelitannin analogs increase the antibiotic susceptibility of S. aureus in both Caenorhabditis elegans and murine mammary infection models. These findings support their potential as antibiotic adjuvants.72,73
Hydrastis canadensis L. extract exhibits quorum quenching by suppressing the AgrC-AgrA two-component system, and reduces α-hemolysin production and MRSA-induced keratinocyte damage.74 The flavonoid-rich extract 430D-F5 from Schinus terebinthifolia (Brazilian Peppertree) inhibits Agr activity of S. aureus without growth suppression and reduces skin necrosis in murine models.75 Carnosic acid derived from Rosmarinus officinalis L. leaves suppresses agr expression.76
Hispidulin, a natural flavone, has been identified through a hemolysis activity assay as an inhibitor of hemolysis and hla transcription by binding to AgrC-AgrA. This action alleviates S. aureus-induced damage to A549 cells and reduces lactate dehydrogenase (LDH) release.77 Ajoene, a sulfur-rich garlic compound derived from allicin, suppresses QS in S. aureus and P. aeruginosa via inhibiting small RNA (sRNA).78
Animal-Derived QSIs
In addition to plant-derived and microbe-derived inhibitors, animals have co-evolved molecules capable of interfering with the staphylococcal Agr system. Li et al identified the milk-derived antimicrobial peptide BCp12, which downregulates agrA, agrB, agrC and psmβ, as well as markedly reduces MRSA α-hemolysin and phenol-soluble modulin (PSM) production.79 Notably, BCp12 acts upstream of the AgrC-AgrA two-component system, offering a complementary mode of action to RIP derivatives and flavonoids, making it a valuable partner for combination regimens. Colostrum oligosaccharides and lactoferrin hydrolysates have also been reported to possess quorum-quenching potential, indicating that the animal-derived bioactive library remains largely unexplored.80 Future efforts should integrate multi-omics screening, synthetic-biology expression and formulation protection to expand the structural diversity and in-vivo stability of such peptides (Table 2).
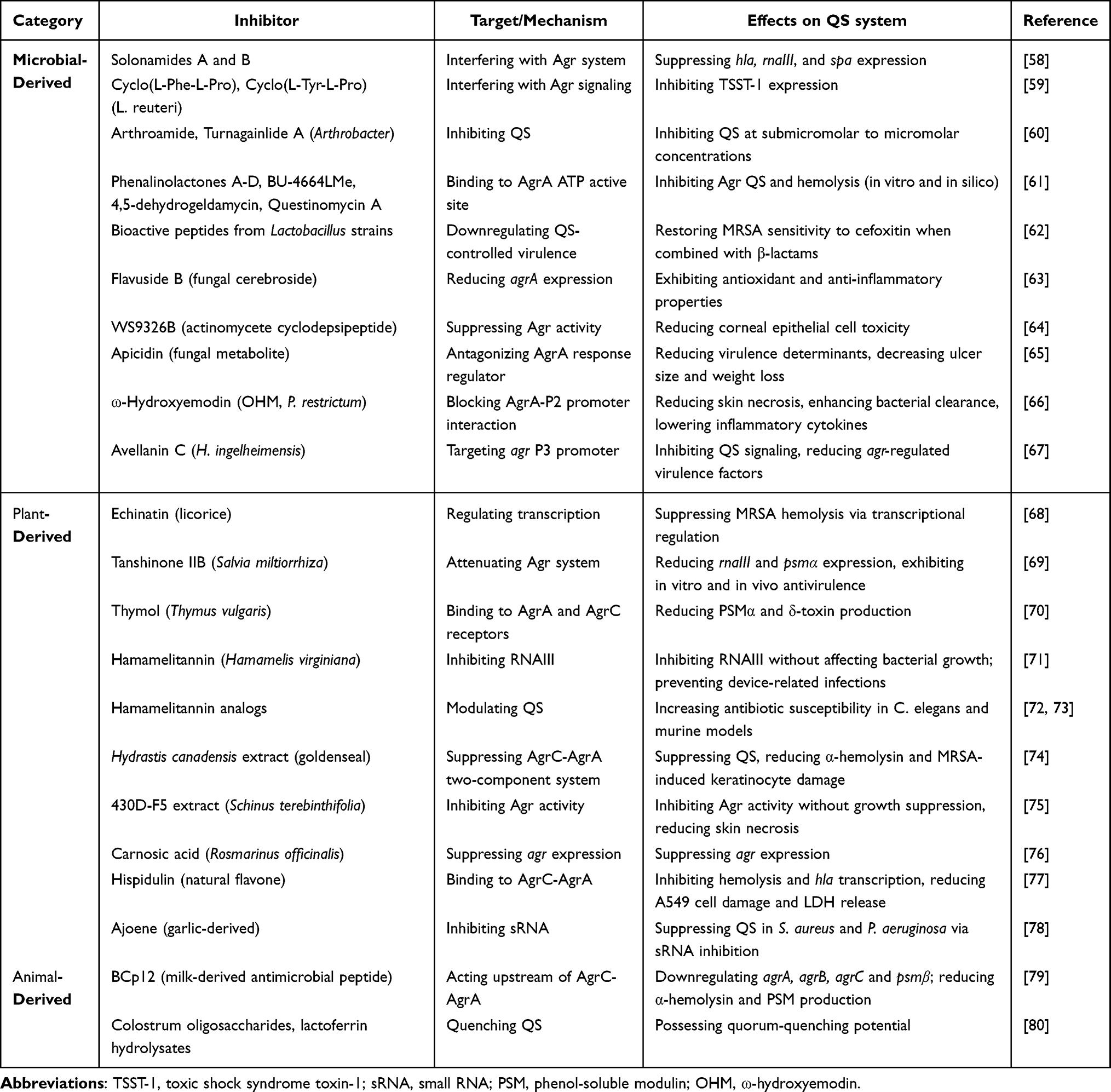 |
Table 2 Natural Product-Derived QSIs Targeting the S. aureus Agr System |
Quenching Antibodies
Quorum quenching antibodies specifically bind to and neutralize QS autoinducers (AIs), and block signal propagation. Park et al developed the anti-AIP-4 monoclonal antibody AP4-24H11, which inhibits S. aureus QS in vitro and protects mice from lethal infections in abscess models.56,57 Beyond targeting AIs directly, antibodies can also suppress QS by interfering with signal transduction components. For instance, anti-TRAP monoclonal antibodies inhibit Agr-regulated toxin production by targeting the TRAP receptor, which mediates RIP protein-dependent Agr suppression34 (Table 1).
Compared to small-molecule inhibitors, antibody-based approaches offer distinct pharmacological advantages. Monoclonal antibodies exhibit extended serum half-lives, enabling prolonged therapeutic coverage and less frequent dosing.81,82 Their high target specificity minimizes off-target effects and reduces resistance selection pressure, while the ability to provide immediate protection without invoking host immune responses renders them particularly attractive for passive immunization strategies, especially in immunocompromised patients who respond poorly to active vaccines.15,81,83
Challenges in Agr-Targeted Antivirulence Research
Despite over two decades of research on quorum sensing inhibition of S. aureus, no Agr-targeted antivirulence inhibitors have advanced to clinical application. This stagnation in clinical translation stems from three interconnected challenges, each defining specific research priorities grounded in current evidence:
First, robust in vivo efficacy data remain scarce, particularly from infection models that rigorously distinguish antivirulence effects from growth inhibition. While numerous Agr inhibitors exhibit potent activity at nanomolar concentrations in vitro, their protective efficacy in vivo is inconclusive. The predominant reliance on acute topical infection models or simple co-incubation assays fails to exclude confounding by nonspecific antibacterial activity, leaving the mechanism of protection ambiguous. Addressing this limitation necessitates prioritizing chronic and device-associated infection models—contexts where Agr exhibits its most complex and clinically relevant regulatory behavior—complemented by isogenic Agr-mutant controls and transcriptomic validation of on-target activity to establish mechanism-specific efficacy.
Second, the pleiotropic and context-dependent nature of Agr regulation creates inherent therapeutic trade-offs. Although Agr upregulates secreted toxins and immune evasion factors, it concurrently suppresses biofilm formation and promotes bacterial dissemination.23 Consequently, Agr inhibition risks exacerbating biofilm persistence, particularly in chronic and device-associated infections, thereby constraining its therapeutic utility. The most viable strategy emerging from existing data is the dual-activity design principle: selection of candidates that simultaneously inhibit Agr signaling and prevent biofilm formation. Current literature identifies three promising chemical classes: RIP derivatives (exemplified by FS8, which inhibits Agr signaling at nanomolar concentrations and synergizes with tigecycline to attenuate biofilm formation);84 natural flavonoids (luteolin, quercetin, and baicalein, which concurrently suppress Agr-regulated virulence factors and biofilm development at sub-inhibitory concentrations);85–87 and multifunctional natural products (carboxypyranoanthocyanins, baohuoside I, and cinnamaldehyde, which attenuate virulence without inducing biofilm rebound).88–92 These scaffolds constitute rational starting points for lead optimization and translational development.
Third, the clinical and regulatory pathway for non-bactericidal antivirulence agents remains ill-defined. The absence of rapid bactericidal activity complicates efficacy assessment and may deter commercial investment. To navigate these barriers, future efforts should prioritize combination therapeutic strategies that pair Agr inhibitors with conventional antibiotics or anti-biofilm agents—leveraging the documented synergy between FS8 and tigecycline as proof-of-concept. Beyond small-molecule approaches, nanomaterial-based delivery systems (eg, liposomal encapsulation, polymeric nanoparticles) offer promising avenues to enhance inhibitor stability, achieve sustained release at infection sites, and improve penetration of biofilm matrices. Additionally, biologic modalities including anti-AgrA monoclonal antibodies and AIP-targeted aptamers provide alternative targeting strategies with potentially superior specificity compared to small molecules. These technological innovations, integrated with the dual-activity scaffolds identified above, represent the most promising path toward overcoming current translational bottlenecks.
Conclusions and Future Directions
As the primary quorum sensing system in S. aureus, AgrA regulates the majority of virulence factors in this pathogen. This characteristic makes it an attractive target for screening anti-virulence drugs. To date, numerous candidate compounds targeting AgrA have been identified. These compounds not only exhibit significant inhibition of AgrA activity in vitro but have also demonstrated effective antiinfective effects in vivo in animal experiments, suggesting their potential for further development. However, since anti-AgrA compounds often simultaneously promote biofilm formation, a comprehensive and cautious evaluation of their potential side effects in vivo is necessary. Future efforts should focus on screening anti-virulence drugs that possess both anti-AgrA and biofilm inhibition properties. Further research should be conducted on the pharmacokinetics, toxicology, stability, and other characteristics of candidate drugs. Based on these findings, additional modifications, such as chemical stabilization through structural modification or the application of nanotechnology for controlled release, are essential to ultimately facilitate the practical application of these potential anti-virulence drugs against S. aureus.
Compliance with Ethics Requirements
This article does not contain any studies with human or animal subjects.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by Dongguan Key Laboratory for Pathogenesis and Experimental Diagnosis of Infectious Diseases (20231600400762). Talent Development Foundation of The First Dongguan Affiliated Hospital of Guangdong Medical University (GCC2023002).
Disclosure
The authors declare that they have no conflict of financial interests.
References
1. Turner NA, Sharma-Kuinkel BK, Maskarinec SA, et al. Methicillin-resistant Staphylococcus aureus: an overview of basic and clinical research. Nat Rev Microbiol. 2019;17(4):203–13. doi:10.1038/s41579-018-0147-4
2. Siddiqui AH, Koirala J. Methicillin-Resistant Staphylococcus aureus. In: StatPearls. StatPearls Publishing LLC.; 2025.
3. Antimicrobial Resistance C. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629–655. doi:10.1016/S0140-6736(21)02724-0
4. Chen CJ, Huang YC, Shie SS. Evolution of Multi-Resistance to Vancomycin, Daptomycin, and Linezolid in Methicillin-Resistant Staphylococcus aureus Causing Persistent Bacteremia. Front Microbiol. 2020;11:1414. doi:10.3389/fmicb.2020.01414
5. Vinodhini V, Kavitha M. Deciphering agr quorum sensing in Staphylococcus aureus: insights and therapeutic prospects. Mol Biol Rep. 2024;51(1):155. doi:10.1007/s11033-023-08930-3
6. Jenul C, Horswill AR. Regulation of Staphylococcus aureus Virulence. Microbiol Spectr. 2019;7(2):1. doi:10.1128/microbiolspec.GPP3-0031-2018
7. Otto M. Staphylococcal Biofilms. Microbiol Spectr. 2018;6(4):3. doi:10.1128/microbiolspec.GPP3-0023-2018
8. LaSarre B, Federle MJ. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol Mol Biol Rev. 2013;77(1):73–111. doi:10.1128/MMBR.00046-12
9. Novick RP, Geisinger E. Quorum sensing in staphylococci. Annu Rev Genet. 2008;42:541–564. doi:10.1146/annurev.genet.42.110807.091640
10. Queck SY, Jameson-Lee M, Villaruz AE, et al. RNAIII-independent target gene control by the agr quorum-sensing system: insight into the evolution of virulence regulation in Staphylococcus aureus. Mol Cell. 2008;32(1):150–158. doi:10.1016/j.molcel.2008.08.005
11. Papenfort K, Bassler BL. Quorum sensing signal-response systems in Gram-negative bacteria. Nat Rev Microbiol. 2016;14(9):576–588. doi:10.1038/nrmicro.2016.89
12. Haque S, Yadav DK, Bisht SC, et al. Quorum sensing pathways in Gram-positive and -negative bacteria: potential of their interruption in abating drug resistance. J Chemother. 2019;31(4):161–187. doi:10.1080/1120009x.2019.1599175
13. Dickey SW, Cheung GYC, Otto M. Different drugs for bad bugs: antivirulence strategies in the age of antibiotic resistance. Nat Rev Drug Discov. 2017;16(7):457–471. doi:10.1038/nrd.2017.23
14. Sully EK, Malachowa N, Elmore BO, et al. Selective chemical inhibition of agr quorum sensing in Staphylococcus aureus promotes host defense with minimal impact on resistance. PLoS Pathog. 2014;10(6):e1004174. doi:10.1371/journal.ppat.1004174
15. Varrone JJ, Li D, Daiss JL, Schwarz EM. Anti-Glucosaminidase Monoclonal Antibodies as a Passive Immunization for Methicillin-Resistant Staphylococcus aureus (MRSA) Orthopaedic Infections. Bonekey Osteovision. 2011;8:187–194. doi:10.1138/20110506
16. Otto M. Critical Assessment of the Prospects of Quorum-Quenching Therapy for Staphylococcus aureus Infection. Int J Mol Sci. 2023;24(4):4025. doi:10.3390/ijms24044025
17. Tan L, Li SR, Jiang B, Hu XM, Li S. Therapeutic Targeting of the Staphylococcus aureus Accessory Gene Regulator (agr) System. Front Microbiol. 2018;9:55. doi:10.3389/fmicb.2018.00055
18. Fang L, Cosgriff C, Alonzo III F. Determinants of maturation of the Staphylococcus aureus autoinducing peptide. J Bacteriol. 2024;206(9):e0019524. doi:10.1128/jb.00195-24
19. Wang B, Zhao A, Xie Q, et al. Functional Plasticity of the AgrC Receptor Histidine Kinase Required for Staphylococcal Virulence. Cell Chem Biol. 2017;24(1):76–86. doi:10.1016/j.chembiol.2016.12.008
20. Koenig RL, Ray JL, Maleki SJ, Smeltzer MS, Hurlburt BK. Staphylococcus aureus Agra binding to the RNAIII-agr regulatory region. J Bacteriol. 2004;186(22):7549–7555. doi:10.1128/jb.186.22.7549-7555.2004
21. Pader V, James EH, Painter KL, Wigneshweraraj S, Edwards AM. The Agr quorum-sensing system regulates fibronectin binding but not hemolysis in the absence of a functional electron transport chain. Infect Immun. 2014;82(10):4337–4347. doi:10.1128/IAI.02254-14
22. Bronner S, Monteil H, Prevost G. Regulation of virulence determinants in Staphylococcus aureus: complexity and applications. FEMS Microbiol Rev. 2004;28(2):183–200. doi:10.1016/j.femsre.2003.09.003
23. Periasamy S, Joo HS, Duong AC, et al. How Staphylococcus aureus biofilms develop their characteristic structure. Proc Natl Acad Sci U S A. 2012;109(4):1281–1286. doi:10.1073/pnas.1115006109
24. Boles BR, Horswill AR. Agr-mediated dispersal of Staphylococcus aureus biofilms. PLoS Pathog. 2008;4(4):e1000052. doi:10.1371/journal.ppat.1000052
25. Cheung GYC, Bae JS, Otto M. Pathogenicity and virulence of Staphylococcus aureus. Virulence. 2021;12(1):547–569. doi:10.1080/21505594.2021.1878688
26. Ji G, Beavis R, Novick RP. Bacterial interference caused by autoinducing peptide variants. Science. 1997;276(5321):2027–2030. doi:10.1126/science.276.5321.2027
27. Mayville P, Ji G, Beavis R, et al. Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proc Natl Acad Sci U S A. 1999;96(4):1218–1223. doi:10.1073/pnas.96.4.1218
28. Wright III JS, Jin R, Novick RP. Transient interference with staphylococcal quorum sensing blocks abscess formation. Proc Natl Acad Sci U S A. 2005;102(5):1691–1696. doi:10.1073/pnas.0407661102
29. Tal-Gan Y, Stacy DM, Foegen MK, Koenig DW, Blackwell HE. Highly potent inhibitors of quorum sensing in Staphylococcus aureus revealed through a systematic synthetic study of the group-III autoinducing peptide. J Am Chem Soc. 2013;135(21):7869–7882. doi:10.1021/ja3112115
30. Tal-Gan Y, Ivancic M, Cornilescu G, Yang T, Blackwell HE. Highly Stable, Amide-Bridged Autoinducing Peptide Analogues that Strongly Inhibit the AgrC Quorum Sensing Receptor in Staphylococcus aureus. Angew Chem Int Ed Engl. 2016;55(31):8913–8917. doi:10.1002/anie.201602974
31. Vasquez JK, Tal-Gan Y, Cornilescu G, Tyler KA, Blackwell HE. Simplified AIP-II Peptidomimetics Are Potent Inhibitors of Staphylococcus aureus AgrC Quorum Sensing Receptors. Chembiochem. 2017;18(4):413–423. doi:10.1002/cbic.201600516
32. Vasquez JK, Blackwell HE. Simplified Autoinducing Peptide Mimetics with Single-Nanomolar Activity Against the Staphylococcus aureus AgrC Quorum Sensing Receptor. ACS Infect Dis. 2019;5(4):484–492. doi:10.1021/acsinfecdis.9b00002
33. Zhao K, Vasquez JK, Blackwell HE. Potent pan-group quorum sensing inhibitors in Staphylococcus aureus revealed by N-terminal tailoring of peptidomimetics. Chem Commun. 2023;59(5):587–590. doi:10.1039/d2cc05733f
34. Vieira-da-Motta O, Ribeiro PD, Dias da Silva W, Medina-Acosta E. RNAIII inhibiting peptide (RIP) inhibits agr-regulated toxin production. Peptides. 2001;22(10):1621–1627. doi:10.1016/s0196-9781(01)00497-1
35. Balaban N, Novick RP. Autocrine regulation of toxin synthesis by Staphylococcus aureus. Proc Natl Acad Sci U S A. 1995;92(5):1619–1623. doi:10.1073/pnas.92.5.1619
36. Balaban N, Collins LV, Cullor JS, et al. Prevention of diseases caused by Staphylococcus aureus using the peptide RIP. Peptides. 2000;21(9):1301–1311. doi:10.1016/s0196-9781(00)00272-2
37. Balaban N, Goldkorn T, Nhan RT, et al. Autoinducer of virulence as a target for vaccine and therapy against Staphylococcus aureus. Science. 1998;280:5362):438–40. doi:10.1126/science.280.5362.438
38. Cirioni O, Mocchegiani F, Cacciatore I, et al. Quorum sensing inhibitor FS3-coated vascular graft enhances daptomycin efficacy in a rat model of staphylococcal infection. Peptides. 2013;40:77–81. doi:10.1016/j.peptides.2012.12.002
39. Simonetti O, Cirioni O, Cacciatore I, et al. Efficacy of the Quorum Sensing Inhibitor FS10 Alone and in Combination with Tigecycline in an Animal Model of Staphylococcal Infected Wound. PLoS One. 2016;11(6):e0151956. doi:10.1371/journal.pone.0151956
40. Palaniappan B, Solomon AP, DR C. Targeting Agra quorum sensing regulator by bumetanide attenuates virulence in Staphylococcus aureus - A drug repurposing approach. Life Sci. 2021;273:119306. doi:10.1016/j.lfs.2021.119306
41. Dotto C, Lombarte Serrat A, Ledesma M, et al. Salicylic acid stabilizes Staphylococcus aureus biofilm by impairing the agr quorum-sensing system. Sci Rep. 2021;11(1):2953. doi:10.1038/s41598-021-82308-y
42. Price CT, Lee IR, Gustafson JE. The effects of salicylate on bacteria. Int J Biochem Cell Biol. 2000;32(10):1029–1043. doi:10.1016/s1357-2725(00)00042-x
43. Wu R, Wu Y, Wu P, Li H, She P. Bactericidal and anti-quorum sensing activity of repurposing drug Visomitin against Staphylococcus aureus. Virulence. 2024;15(1):2415952. doi:10.1080/21505594.2024.2415952
44. Khayat MT, Abbas HA, Ibrahim TS, et al. Anti-Quorum Sensing Activities of Gliptins against Pseudomonas aeruginosa and Staphylococcus aureus. Biomedicines. 2022;10(5):1169. doi:10.3390/biomedicines10051169
45. Yu G, Kuo D, Shoham M, Viswanathan R. Combinatorial synthesis and in vitro evaluation of a biaryl hydroxyketone library as antivirulence agents against MRSA. ACS Comb Sci. 2014;16(2):85–91. doi:10.1021/co400142t
46. Kuo D, Yu G, Hoch W, et al. Novel quorum-quenching agents promote methicillin-resistant Staphylococcus aureus (MRSA) wound healing and sensitize MRSA to beta-lactam antibiotics. Antimicrob Agents Chemother. 2015;59(3):1512–1518. doi:10.1128/AAC.04767-14
47. Yamaguchi J, Manome T, Hara Y, et al. Physalin H, physalin B, and isophysalin B suppress the quorum-sensing function of Staphylococcus aureus by binding to Agra. Front Pharmacol. 2024;15:1365815. doi:10.3389/fphar.2024.1365815
48. Khodaverdian V, Pesho M, Truitt B, et al. Discovery of antivirulence agents against methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2013;57(8):3645–3652. doi:10.1128/AAC.00269-13
49. Todd DA, Parlet CP, Crosby HA, et al. Signal Biosynthesis Inhibition with Ambuic Acid as a Strategy To Target Antibiotic-Resistant Infections. Antimicrob Agents Chemother. 2017;61(8):2. doi:10.1128/AAC.00263-17
50. Kim MK, Zhao A, Wang A, et al. Surface-attached molecules control Staphylococcus aureus quorum sensing and biofilm development. Nat Microbiol. 2017;2:17080. doi:10.1038/nmicrobiol.2017.80
51. Xie Q, Wiedmann MM, Zhao A, et al. Discovery of quorum quenchers targeting the membrane-embedded sensor domain of the Staphylococcus aureus receptor histidine kinase, AgrC. Chem Commun. 2020;56(76):11223–11226. doi:10.1039/d0cc04873a
52. Lee JH, Kim YG, Lee J. Inhibition of Staphylococcus aureus Biofilm Formation and Virulence Factor Production by Petroselinic Acid and Other Unsaturated C18 Fatty Acids. Microbiol Spectr. 2022;10(3):e0133022. doi:10.1128/spectrum.01330-22
53. Sim M, Kim YG, Lee JH, Lee J. Antibiofilm Activities of Multiple Halogenated Pyrimidines Against Staphylococcus aureus. Int J Mol Sci. 2024;25(23):12830. doi:10.3390/ijms252312830
54. Kratochvil MJ, Tal-Gan Y, Yang T, Blackwell HE, Lynn DM. Nanoporous Superhydrophobic Coatings that Promote the Extended Release of Water-Labile Quorum Sensing Inhibitors and Enable Long-Term Modulation of Quorum Sensing in Staphylococcus aureus. ACS Biomater Sci Eng. 2015;1(10):1039–1049. doi:10.1021/acsbiomaterials.5b00313
55. Chen R, Maslekar N, Chakraborty S, et al. Quorum sensing inhibiting dihydropyrrol-2-ones embedded polymer/graphene oxide nanocomposite waterborne antimicrobial coatings. J Mater Chem B. 2024;12(34):8366–8375. doi:10.1039/d4tb01026d
56. Park J, Jagasia R, Kaufmann GF, et al. Infection control by antibody disruption of bacterial quorum sensing signaling. Chem Biol. 2007;14(10):1119–1127. doi:10.1016/j.chembiol.2007.08.013
57. Kaufmann GF, Park J, Mayorov AV, Kubitz DM, Janda KD. Generation of quorum quenching antibodies. Methods Mol Biol. 2011;692:299–311. doi:10.1007/978-1-60761-971-0_22
58. Mansson M, Nielsen A, Kjaerulff L, et al. Inhibition of virulence gene expression in Staphylococcus aureus by novel depsipeptides from a marine photobacterium. Mar Drugs. 2011;9(12):2537–2552. doi:10.3390/md9122537
59. Li J, Wang W, Xu SX, Magarvey NA, McCormick JK. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc Natl Acad Sci U S A. 2011;108(8):3360–3365. doi:10.1073/pnas.1017431108
60. Igarashi Y, Yamamoto K, Fukuda T, et al. Arthroamide, a Cyclic Depsipeptide with Quorum Sensing Inhibitory Activity from Arthrobacter sp. J Nat Prod. 2015;78(11):2827–2831. doi:10.1021/acs.jnatprod.5b00540
61. Desouky SE, Abu-Elghait M, Fayed EA, et al. Secondary Metabolites of Actinomycetales as Potent Quorum Sensing Inhibitors Targeting Gram-Positive Pathogens: in Vitro and In Silico Study. Metabolites. 2022;12(3):246. doi:10.3390/metabo12030246
62. Cella MA, Coulson T, MacEachern S, et al. Probiotic disruption of quorum sensing reduces virulence and increases cefoxitin sensitivity in methicillin-resistant Staphylococcus aureus. Sci Rep. 2023;13(1):4373. doi:10.1038/s41598-023-31474-2
63. Chingizova EA, Yurchenko EA, Starnovskaya SS, et al. Flavuside B exhibits antioxidant and anti-inflammatory properties in Staphylococcus aureus infected skin wound and affect the expression of genes controlling bacterial quorum sensing. J Appl Microbiol. 2025;136(1):318. doi:10.1093/jambio/lxae318
64. Desouky SE, Shojima A, Singh RP, et al. Cyclodepsipeptides produced by actinomycetes inhibit cyclic-peptide-mediated quorum sensing in Gram-positive bacteria. FEMS Microbiol Lett. 2015;362(14):109. doi:10.1093/femsle/fnv109
65. Parlet CP, Kavanaugh JS, Crosby HA, et al. Apicidin Attenuates MRSA Virulence through Quorum-Sensing Inhibition and Enhanced Host Defense. Cell Rep. 2019;27(1):187–198e6. doi:10.1016/j.celrep.2019.03.018
66. Daly SM, Elmore BO, Kavanaugh JS, et al. omega-Hydroxyemodin limits staphylococcus aureus quorum sensing-mediated pathogenesis and inflammation. Antimicrob Agents Chemother. 2015;59(4):2223–2235. doi:10.1128/AAC.04564-14
67. Igarashi Y, Gohda F, Kadoshima T, et al. Avellanin C, an inhibitor of quorum-sensing signaling in Staphylococcus aureus, from Hamigera ingelheimensis. J Antibiot (Tokyo). 2015;68(11):707–710. doi:10.1038/ja.2015.50
68. Zhang W, Gong Q, Tang Z, et al. The natural product, echinatin, protects mice from methicillin-resistant Staphylococcus aureus pneumonia by inhibition of alpha-hemolysin expression. Front Microbiol. 2023;14:1128144. doi:10.3389/fmicb.2023.1128144
69. Jiang JM, Chen G, Chen YY, et al. The anti-infective activity of Salvia miltiorrhiza against Staphylococcus aureus by attenuating accessory gene regulator system-mediated virulence. Food Funct. 2022;13(9):5050–5060. doi:10.1039/d1fo01888d
70. Lade H, Chung SH, Lee Y, et al. Thymol Reduces agr-Mediated Virulence Factor Phenol-Soluble Modulin Production in Staphylococcus aureus. Biomed Res Int. 2022;2022:8221622. doi:10.1155/2022/8221622
71. Brackman G, Breyne K, De Rycke R, et al. The Quorum Sensing Inhibitor Hamamelitannin Increases Antibiotic Susceptibility of Staphylococcus aureus Biofilms by Affecting Peptidoglycan Biosynthesis and eDNA Release. Sci Rep. 2016;6:20321. doi:10.1038/srep20321
72. Vermote A, Brackman G, Risseeuw MD, et al. Hamamelitannin Analogues that Modulate Quorum Sensing as Potentiators of Antibiotics against Staphylococcus aureus. Angew Chem Int Ed Engl. 2016;55(22):6551–6555. doi:10.1002/anie.201601973
73. Vermote A, Brackman G, Risseeuw MDP, Coenye T, Van Calenbergh S. Design, synthesis and biological evaluation of novel hamamelitannin analogues as potentiators for vancomycin in the treatment of biofilm related Staphylococcus aureus infections. Bioorg Med Chem. 2016;24(19):4563–4575. doi:10.1016/j.bmc.2016.07.058
74. Cech NB, Junio HA, Ackermann LW, Kavanaugh JS, Horswill AR. Quorum quenching and antimicrobial activity of goldenseal (Hydrastis canadensis) against methicillin-resistant Staphylococcus aureus (MRSA). Planta Med. 2012;78(14):1556–1561. doi:10.1055/s-0032-1315042
75. Muhs A, Lyles JT, Parlet CP, et al. Virulence Inhibitors from Brazilian Peppertree Block Quorum Sensing and Abate Dermonecrosis in Skin Infection Models. Sci Rep. 2017;7:42275. doi:10.1038/srep42275
76. Nakagawa S, Hillebrand GG, Nunez G. Rosmarinus officinalis L. (Rosemary) Extracts Containing Carnosic Acid and Carnosol are Potent Quorum Sensing Inhibitors of Staphylococcus aureus Virulence. Antibiotics. 2020;9(4):149. doi:10.3390/antibiotics9040149
77. Ren X, Guo X, Liu C, et al. Natural flavone hispidulin protects mice from Staphylococcus aureus pneumonia by inhibition of alpha-hemolysin production via targeting Agra(C). Microbiol Res. 2022;261:127071. doi:10.1016/j.micres.2022.127071
78. Jakobsen TH, Warming AN, Vejborg RM, et al. A broad range quorum sensing inhibitor working through sRNA inhibition. Sci Rep. 2017;7(1):9857. doi:10.1038/s41598-017-09886-8
79. Li Y, Li S, Yang K, et al. Antibiofilm mechanism of a novel milk-derived antimicrobial peptide against Staphylococcus aureus by downregulating agr quorum sensing system. J Appl Microbiol. 2022;133(4):2198–2209. doi:10.1111/jam.15653
80. Srivastava A, Singh BN, Deepak D, Rawat AK, Singh BR. Colostrum hexasaccharide, a novel Staphylococcus aureus quorum-sensing inhibitor. Antimicrob Agents Chemother. 2015;59(4):2169–2178. doi:10.1128/AAC.03722-14
81. Wang-Lin S, Balthasar J. Pharmacokinetic and Pharmacodynamic Considerations for the Use of Monoclonal Antibodies in the Treatment of Bacterial Infections. Antibodies. 2018;7(1):5. doi:10.3390/antib7010005
82. Deng R, Zhou C, Li D, et al. Preclinical and translational pharmacokinetics of a novel THIOMAB™ antibody-antibiotic conjugate against Staphylococcus aureus. MAbs. 2019;11(6):1162–1174. doi:10.1080/19420862.2019.1627152
83. Zhang J, Yang F, Zhang X, et al. Protective Efficacy and Mechanism of Passive Immunization with Polyclonal Antibodies in a Sepsis Model of Staphylococcus aureus Infection. Sci Rep. 2015;5(1):15553. doi:10.1038/srep15553
84. Simonetti O, Cirioni O, Mocchegiani F, et al. The efficacy of the quorum sensing inhibitor FS8 and tigecycline in preventing prosthesis biofilm in an animal model of staphylococcal infection. Int J Mol Sci. 2013;14(8):16321–16332. doi:10.3390/ijms140816321
85. Li Y, Dai J, Ma Y, et al. The mitigation potential of synergistic quorum quenching and antibacterial properties for biofilm proliferation and membrane biofouling. Water Res. 2024;255:121462. doi:10.1016/j.watres.2024.121462
86. Yuan Q, Feng W, Wang Y, et al. Luteolin attenuates the pathogenesis of Staphylococcus aureus by interfering with the agr system. Microb Pathog. 2022;165:105496. doi:10.1016/j.micpath.2022.105496
87. Chen Y, Liu T, Wang K, et al. Baicalein Inhibits Staphylococcus aureus Biofilm Formation and the Quorum Sensing System In Vitro. PLoS One. 2016;11(4):e0153468. doi:10.1371/journal.pone.0153468
88. Ferro TA, Araujo JM, Dos Santos Pinto BL, et al. Cinnamaldehyde Inhibits Staphylococcus aureus Virulence Factors and Protects against Infection in a Galleria mellonella Model. Front Microbiol. 2016;7:2052. doi:10.3389/fmicb.2016.02052
89. Kavanaugh NL, Ribbeck K. Selected antimicrobial essential oils eradicate Pseudomonas spp. and Staphylococcus aureus biofilms. Appl Environ Microbiol. 2012;78(11):4057–4061. doi:10.1128/AEM.07499-11
90. Brackman G, Cos P, Maes L, Nelis HJ, Coenye T. Quorum sensing inhibitors increase the susceptibility of bacterial biofilms to antibiotics in vitro and in vivo. Antimicrob Agents Chemother. 2011;55(6):2655–2661. doi:10.1128/AAC.00045-11
91. Coelho P, Oliveira J, Fernandes I, et al. Pyranoanthocyanins Interfering with the Quorum Sensing of Pseudomonas aeruginosa and Staphylococcus aureus. Int J Mol Sci. 2021;22(16):8559. doi:10.3390/ijms22168559
92. El-Sawy ER, Abdel-Aziz MS, Abdelmegeed H, Kirsch G. Coumarins: quorum Sensing and Biofilm Formation Inhibition. Molecules. 2024;29(19):4534. doi:10.3390/molecules29194534
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.