Back to Journals » Neuropsychiatric Disease and Treatment » Volume 21
Research Advances in Multimodal Framework-Based Identification and Therapeutic Management of Post-Stroke Depression via the Microbiota-Gut-Brain Axis
Authors Shen M ![]() , Yu P, Xiong Q, Sun Z, Zhang Y, Yang M
, Yu P, Xiong Q, Sun Z, Zhang Y, Yang M
Received 30 August 2025
Accepted for publication 9 December 2025
Published 16 December 2025 Volume 2025:21 Pages 2835—2853
DOI https://doi.org/10.2147/NDT.S564175
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rakesh Kumar
Mengyue Shen, Penglong Yu, Qian Xiong, Zhongli Sun, Yunhui Zhang, Menglin Yang
Department of Traditional Chinese Medicine, Chongqing Three Gorges Medical College, Chongqing, People’s Republic of China
Correspondence: Mengyue Shen, Chongqing Three Gorges Medical College, 366 Tianxing Road, Wanzhou District, Chongqing, 404120, People’s Republic of China, Email [email protected]
Abstract: Patients with post-stroke depression typically present with psychosocial impairments, including depressed mood and pessimism, accompanied by behavioral manifestations such as social withdrawal. These symptoms significantly impede rehabilitation compliance while elevating risks of self-harm and suicidal ideation. Recent advances in microbiota-gut-brain axis research have elucidated bidirectional communication pathways between the gut microbiota-mediated neuroendocrine-immune network and the central nervous system. Dysregulation of microbiota-gut-brain axis homeostasis may precipitate neuroinflammatory cascades and NE metabolic disturbances, potentially driving post-stroke depression pathogenesis. This paper attempts to propose a multimodal precision diagnosis and treatment framework for post-stroke depression based on the microbiota-gut-brain axis mechanism, representing the first effort to integrate perspectives from neurobiology, gut microbiota, and Traditional Chinese Medicine with modern scientific interpretation to construct such a framework. The paper encompasses four levels: mechanism, integration, evidence, and application. At the mechanism level, it explores the bidirectional regulatory mechanisms of the microbiota-gut-brain axis in post-stroke depression to identify potential therapeutic targets. At the integration level, it refines core principles for constructing a multimodal system applicable to post-stroke depression and builds an microbiota-gut-brain axis based multimodal research framework for post-stroke depression. At the evidence level, by integrating neuroimaging, metabolomics, and microbiomics technologies, it discusses the potential of a multimodal identification system, which is expected to aid in identifying molecular-cellular-circuit mechanisms associated with post-stroke depression. At the application level, it reviews research progress in treating post-stroke depression using the central nervous system interventions, gut microbiota modulation, pharmaceuticals, complementary medicine, and lifestyle interventions, summarizing them into multimodal interventional strategies to inform clinical practice for comprehensive treatment. By incorporating the roles of gut microbiota and oxidative stress in stroke-related complications and neuroimmune pathologies, this review offers a more comprehensive theoretical basis for the precise treatment of post-stroke depression. Future research should rely on large-scale cohorts and artificial intelligence to clarify the dynamic interactive networks of multiple biomarkers within critical time windows, ultimately facilitating the translation of this multimodal framework from theory to clinical practice.
Keywords: post-stroke depression, microbiota-gut-brain axis, multimodal system, identification, intervention
Introduction
Research Background
Post-stroke depression (PSD) is a prevalent neuropsychiatric consequence following a stroke, manifests clinically as persistent depressive affect and diminished motivation. The prevalence of PSD is significant within the initial three years following a stroke, with a worldwide incidence ranging from around 27.5% to 62.5%.1 The pathophysiology of PSD has been thoroughly investigated. Traditionally, evidence suggests that depression results from the loss of biogenic amine fibre bundles connecting prefrontal and limbic or striatal brain regions,2 which include distinct functional networks, as a direct consequence of stroke damage. Recently, the theory of the microbiota-gut-brain axis (MGBA) has gained attention in a broader set of neurological diseases, extending beyond PSD. Dysfunction of the MGBA caused by gut dysbiosis also plays a significant role in secondary brain injury after acute stroke, vascular cognitive impairment (VCI), and post-stroke dementia. Of particular significance is that VCI, the second most prevalent type of dementia worldwide, is closely linked to gut dysbiosis mediated by the MGBA. This paradigm strongly underscores the broad significance of the MGBA in post-stroke complications. In parallel, the bidirectional regulatory network of the MGBA in PSD has been elucidated: following an acute stroke, the central nervous system (CNS) engages descending pathways via the autonomic nervous system (ANS), the hypothalamic-pituitary-adrenal (HPA) axis, and immune signaling, leading to reduced gut microbial diversity, decreased levels of short-chain fatty acids (ScFAs), and enrichment of pro-inflammatory taxa. These alterations, in turn, form ascending regulatory circuits through neural, immune, and metabolic pathways, which feedback to influence central function, thereby establishing a reciprocal loop closely associated with PSD.
In this network, oxidative stress is recognized as a central hub. Specifically, gut microbiota-derived metabolites such as trimethylamine N-oxide exacerbate cerebrovascular risk by promoting atherosclerosis, whereas ScFAs exert protective effects by suppressing neuroinflammation. Concurrently, post-stroke disruption of the intestinal barrier results in the release of lipopolysaccharide (LPS), which activates microglia (MG) and induces oxidative damage. Similar mechanisms are observed in neuroimmune disorders such as multiple sclerosis, where microbial dysbiosis profoundly influences autoimmune progression and blood-brain barrier (BBB) integrity by modulating the balance between T helper 17 cell (Th17) and regulatory T cells (Treg) and related cytokine release. Collectively, these findings indicate that oxidative stress is a pervasive and central node in MGBA regulation and that gut-driven immune activation is critical in driving central pathophysiology. This study also provides a new perspective for PSD research: rather than being confined to localized brain injury, PSD should be viewed as a disease network mediated by the MGBA, involving multidimensional interactions among neural, immune, metabolic, and oxidative stress components. Emerging evidence further suggests that modulating the gut microbiota through dietary interventions, probiotics, or fecal microbiota transplantation (FMT) can alleviate the severity of PSD and improve functional recovery.3 This shift in perspective creates important opportunities for elucidating the mechanisms of PSD and integrating diagnostic and interventional strategies, making the construction of a multimodal research framework an imperative. Such a framework would employ longitudinal designs to dynamically dissect causal pathways, harness artificial intelligence (AI)-powered methods to map biomarker networks, and ultimately combine strategies from both TCM and Western medicine to overcome the limitations of conventional single-modality identifying and interventional models. However, current understanding of the specific MGBA mechanisms in PSD remains fragmented, hindering a comprehensive parsing of this complex network. Moreover, existing studies predominantly focus on validating isolated mechanisms or unimodal approaches, resulting in a lack of integration of these multimodal interactions. It is therefore imperative to construct a multimodal precision framework for PSD, bridging mechanistic insights to clinical intervention. This framework constitutes both the pressing need of current research and the central objective and innovation of this review.
Research Objectives
Against this backdrop, this review focuses on the two key fields of neurobiology and gut microbiota, integrates relevant research findings. We emphasize exploring the bidirectional regulatory mechanisms of the MGBA in PSD. Addressing the challenge that the complex and diverse clinical manifestations of PSD are poorly served by existing single identificatory or therapeutic modalities, it aims to construct a multimodal precision diagnosis and treatment framework based on the MGBA mechanism. This framework integrates multidisciplinary research findings, designs multimodal identification and treatment systems, and refines core principles for constructing multimodal systems, hoping to provide new ideas and methods for the comprehensive diagnosis and effective treatment of PSD. It is anticipated to break through the limitations of traditional diagnostic and therapeutic models and offer more precise intervention strategies for PSD patients.
Research Methods
Search Strategy
(Gastrointestinal Microbiome[MeSH Terms] OR Gut Microbiota[TIAB] OR Intestinal Microbiome[TIAB] OR Gut Bacteria[TIAB]) AND (Brain-Gut Axis[MeSH Terms] OR Gut-Brain Axis[TIAB] OR Microbiota-Gut-Brain Axis[TIAB]) AND (Stroke[MeSH Terms] OR Poststroke[TIAB] OR Post-stroke[TIAB] OR Cerebral Infarction[TIAB]) AND (Depression[MeSH Terms] OR Depressive Disorder[MeSH Terms] OR Poststroke Depression[TIAB] OR PSD[TIAB]). The time range was from December 2010 to July 2025. Databases searched: PubMed, Medline, Cochrane Library, Embase, Web of Science, CNKI, and Wanfang Database.
Inclusion Types
Original research, randomized controlled trials (RCTs), cohort studies, and meta-analyses were considered for inclusion.
Inclusion Criteria
Published in SCI journals (Q1-Q4) or core journals (Note: “core journals” is a common term for prestigious journals in China, like CSCD).
Exclusion Criteria
Conference abstracts, case reports, studies with non-standardized outcome measures, and those with insufficient statistical power were excluded.
Mechanism Layer: The Bidirectional Interaction Mechanism of the MGBA
The human intestinal tract harbors the body’s largest microbial ecosystem, predominantly composed of the Firmicutes and Bacteroidetes phyla.4 It is crucial for the synthesis of norepinephrine (NE), hormones, and vasoactive compounds, preserving the integrity of the intestinal barrier and the BBB and modulating the immune system. As a pivotal component of the gut-brain axis (GBA), The gut microbiota is capable of transmitting inputs to the CNS and influencing the function of the gut and other organs. Conversely, CNS activity regulates gut physiology through ANS signaling, the HPA axis activation, and cytokine-mediated immunomodulation, hence serving a significant bridging role. Intestinal inflammation, alterations in gut microbiota, and microbial metabolites produced by the microbiome can transmit signals via the gut-brain neural pathway to affect CNS activity.
Top-Down Regulatory Mechanisms of PSD on Gut Microbiota
Stroke-induced dysregulation of NE levels top-down disrupts gut microbiota composition through sympathetic and parasympathetic branches of ANS and HPA axis-mediated pathways. This neuroendocrine imbalance triggers immune homeostasis disruption under stress conditions, further exacerbating structural alterations in microbial communities. Compared to healthy individuals, patients with PSD exhibit gut dysbiosis characterized by elevated pro-inflammatory and pathogenic bacteria alongside a reduction in helpful microorganisms.
NE Level Fluctuations and Descending Signaling Pathways
Two predominant pathophysiological theories underlie the NE-associated mechanisms in PSD: the monoamine depletion hypothesis and the glutamate-mediated excitotoxicity hypothesis. More specifically, Neuronal necrosis following a stroke may diminish NE levels and peripheral sympathetic excitability in affected people. For instance, the concentrations and reactivity of monoamines (Noradrenaline (NA), Hydroxytryptamine (5-HT), and Dopamine (DA)), amino acids (Glutamine (Glu), γ-Aminobutyric Acid (GABA)), and alterations in the levels and sensitivity of NE. Abnormal expression of brain-derived neurotrophic factor (BDNF) impairs the regulation of neuronal plasticity. Furthermore, post-stroke, the BBB is compromised, allowing peripheral NE such as monoamines, neuropeptides, and amino acid to infiltrate the brain, thereby exacerbating CNS dysfunction and initiating a cascade effect via the ANS and HPA pathways, resulting in gut microbiota.
ANS fibre link the gastrointestinal tract to the CNS through the vagus nerve (VN) and spinal afferent fibres. During the stress condition of a stroke, the VN activates the neuroinflammatory pathway, suppresses the production of pro-inflammatory cytokines, safeguards the intestinal mucosal barrier downstream, and regulates the gut microbiota and local immunological milieu. Following stroke, the HPA axis becomes hyperactivated upon stimulation, affecting the synthesis and secretion of corticotropin-releasing hormone (CRH) in the hypothalamic paraventricular nucleus. This response affects the synthesis and secretion of CRH in the hypothalamic paraventricular nucleus, subsequently triggering the release of adrenocorticotropic hormone (ACTH) from the anterior pituitary. ACTH then stimulates the adrenal cortex to release excessive glucocorticoids (GCs), such as corticosterone (CORT). Consequently, the HPA axis hyperactivation may contribute to the occurrence of depressive-like behaviors. However, the causal relationship between HPA axis changes and PSD symptoms in stroke patients requires verification through large-sample longitudinal studies.This cascade disrupts the negative feedback regulatory mechanism of the HPA axis,5 exacerbates neurohormonal dysfunction, and impacts intestinal permeability and motility.6 The resultant stress reaction prompts intestinal T-cells to secrete substantial quantities of pro-inflammatory substances that modify the intestinal milieu. Consequently, the HPA axis hyperactivation may contribute to the occurrence of depressive-like behaviors. However, the causal relationship between HPA axis changes and PSD symptoms in stroke patients requires verification through large-sample longitudinal studies.
Immune-Mediated Microbial Remodeling
PSD recruits bidirectional neuroimmune crosstalk between the CNS and peripheral immune compartments. This interaction modulates gut microbiota composition and metabolic activity through the HPA axis signaling and the ANS regulation. In the context of immunological stress, pro-inflammatory and anti-inflammatory cytokines oppose one another and show differential expression during the early and chronic phases of PSD,7 inducing structural alterations in gut microbiota.
Acute stroke can provoke immunological activation in the body. MG are pivotal components in the CNS inflammatory response. Post-stroke, MG activation further stimulates astrocyte (AC) activation, resulting in a neurotoxic phenotype and the release of pro-inflammatory factors,8 which damages the BBB and exacerbates inflammatory injury to the frontal-limbic system (insula, cingulate gyrus, amygdala, hippocampus). Notably, activation of the HPA axis increases intestinal permeability, leading to the recruitment of peripheral immune cells (monocytes (MO) and macrophages (Mø)) that infiltrate the CNS, releasing pro-inflammatory cytokines, chemokines, and reactive oxygen species, thereby inducing a persistent inflammatory response.9 Consequently, following PSD, immune homeostasis becomes disrupted, resulting in altered levels of the hallmark pro-inflammatory factors interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6), which may exacerbate downstream inflammatory responses and influence the proliferation of Enterobacteriaceae while diminishing beneficial intestinal metabolites.10
Stroke may activate immune protective mechanisms to mitigate excessive inflammation and secondary tissue damage while maintaining gut microbial equilibrium, primarily through the modulation of regulatory Tregs and Th17, the secretion of anti-inflammatory cytokines (such as IL-10 and transforming growth factor-β (TGF-β)) to suppress the activity of other immune cells (such as MG and Tregs),11 and the induction of BDNF production. While this immunosuppressive state confers acute-phase benefits by preventing cytokine storms, its persistence induces detrimental sequelae. Research indicates that compromised B-cells maturation in immunosuppressed individuals results in a reduction in B-cell populations,12 hence diminishing humoral immunity and exacerbating susceptibility to secondary infections.
Bottom-up Feedback Mechanisms of Gut Microbiota on PSD
The gastrointestinal tract is the body’s second brain. Gut microbiota directly synthesize NE and account for approximately 90% of the 5-HT production.13 Additionally, they can indirectly produce NE through the metabolite and precursor synthesis, relaying bottom-up signals to the brain via the MGBA, immune pathways, and NE metabolic regulation, thereby facilitating the neuroendocrine-immune (NEI) network interaction mechanism to modulate PSD.
Neural and Immune Pathways
During PSD modulation, on the one hand, the gut microbiota and their neurotoxic byproducts may affect the CNS via the VN afferent pathway to the solitary tract nucleus and the dorsal midline nucleus,14 impairing emotional regulation. On the other hand, the microorganisms in the intestinal epithelial barrier produce endotoxins, like LPS, which activate the Toll-like receptor (TLR) 4 pathway and recruit Treg cells. This interaction degrades tight junction proteins (TJPs), weakening the intestinal mechanical barrier.15 Consequently, microorganisms and harmful metabolites stimulate associated immune pathways, cross the intestinal barrier into the bloodstream, and trigger pro-inflammatory responses via neural and humoral mechanisms, ultimately contributing to central depressive symptoms.
Regulation of NE Metabolism
The gut microbiota plays a critical role in the synthesis and metabolism of NE. Evidence indicates that ScFAs, beneficial metabolites produced by microbial fermentation of dietary fibre in the gut-act through neural (VNS), immune (Th17, Treg, MG), and chemical (Affects the expression of Tryptophan Hydroxylase (TPH)) pathways to regulate neuroinflammation and NE levels.16 Post-stroke dysbiosis reduces ScFAs levels, increasing susceptibility to neuroinflammation and depression. Specific gut bacteria produce enzymes that facilitate the production of NE or its precursors, which can cross the BBB into the brain and stimulate the manufacture of NE-related neurotransmitters such as Glu, GABA, 5-HT, and DA,17 which are transformed into functional chemicals. Furthermore, certain gut microbes interact with enteroendocrine cells (EECs) via metabolites, inducing NE synthesis and release,which can swiftly relay messages to the brain via the enteric nervous system (ENS) or VNS. For example, EECs are synaptically linked to the VN, and Glu synthesis rapidly relays metabolic signals to the brain within milliseconds.18 Indole derivatives (eg, Indole Propionic Acid, IPA) produced by Clostridium sporogenes can directly synthesize 5-HT or regulate tryptophan metabolism via oxidative stress, leading to 5-HT generation catalyzed by TPH. The signals are ultimately transmitted to the brain via the VNS.19 Collectively, gut microbiota-mediated changes in NE and precursor production may impact brain function and cognition.
In summary, a complex bidirectional pathological cycle exists between PSD and the MGBA. On one hand, post-stroke alterations in the NE system lead to HPA axis hyperactivation and induce immune dysregulation, which top-down regulate the gut environment, resulting in gut microbiota dysbiosis. On the other hand, the dysbiotic gut microbiota and its metabolites can, in turn, bottom-up regulate NE metabolism via the NEI and immune pathways, forming a bottom-up feedback loop to PSD (Figure 1). These intricate mechanisms reveal the close involvement of the MGBA in the pathogenesis and progression of PSD, involving a multi-level dynamic interactive system. Consequently, traditional unimodal research paradigms are inadequate for systematically parsing the spatiotemporal evolution of this complex network. Traditional unimodal research paradigms face limitations in tracking biological trajectories at key nodes, identifying pathological signals, and developing personalized interventions and efficacy evaluations. To address these limitations, we aim to use a multimodal precision diagnosis and treatment framework to thoroughly investigate PSD, decipher the aforementioned bidirectional regulatory network, and provide clear directions for the diagnosis and treatment of PSD.
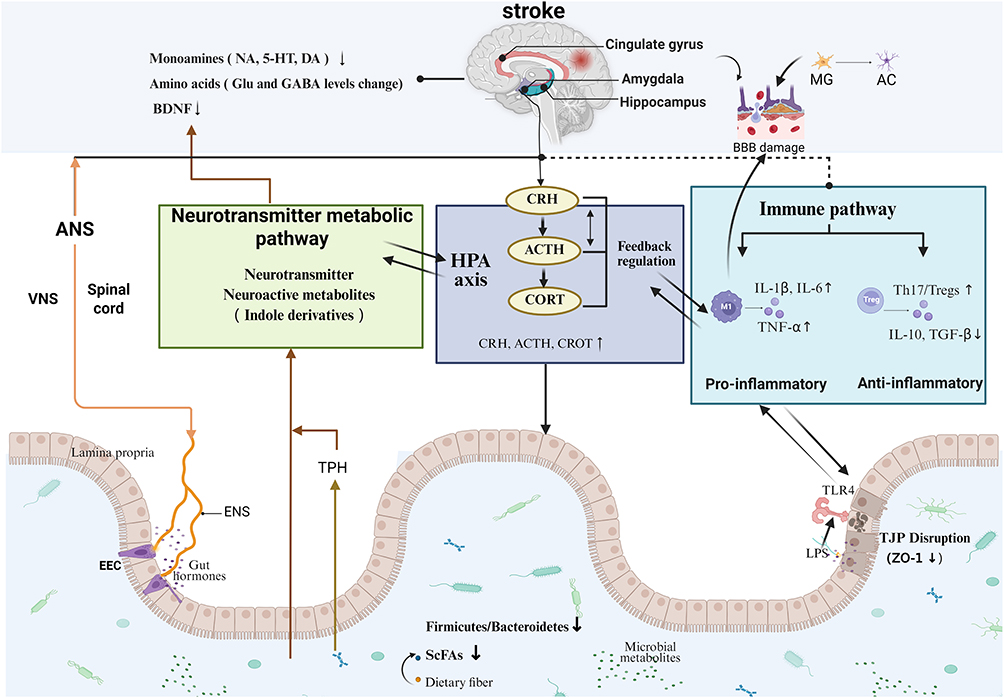 |
Figure 1 The Microbiota-Gut-Brain Axis-based bidirectional interaction mechanism in Post-Stroke Depression. Abbreviations: NA, Noradrenaline; 5-HT, 5-Hydroxytryptamine; DA, Dopamine; Glu, Glutamate; GABA, γ-Aminobutyric Acid; BDNF, Brain-Derived Neurotrophic Factor; BBB, Blood-Brain Barrier; MG, Microglia; AC, Adenylate Cyclase; ANS, Autonomic Nervous System; VNS, Vagus Nervous System; ENS, Enteric Nervous System; TPH, Tryptophan Hydroxylase; HPA, Hypothalamic-Pituitary-Adrenal axis; CRH, Corticotropin-Releasing Hormone; ACTH, Adrenocorticotropic Hormone; CORT, Corticosterone; M1, M1 Macrophage; IL-1β, Interleukin-1β; IL-6, Interleukin-6; TNF-α, Tumor Necrosis Factor-α; Treg, Regulatory T cell; Th17, T helper cell 17; IL-10, Interleukin-10; TGF-β, Transforming Growth Factor-β; EEC, Enteroendocrine Cell; ScFAs, Short-Chain Fatty Acids; TJP, Tight Junction Protein; ZO-1, Zonula Occludens-1; TLR4, Toll-Like Receptor 4; LPS, Lipopolysaccharide. |
Integration Layer: The Construction of a Multimodal PSD Research Framework Based on the MGBA
In the medical field, multimodal characterization is primarily applied in medical image recognition, developing auxiliary diagnostic systems, and designing personalized treatment plans. Building on multidisciplinary research, we innovatively propose the “Three Principles of Multimodal Integration for PSD”: Temporal Dynamics, Spatial Specificity, and Cross-domain Adaptability. These principles are intended to investigate PSD mechanisms, guide the design of multimodal identification and therapeutic systems, and establish a multimodal precision diagnosis and treatment framework for PSD.
Temporal Dynamics: Tracking the Biological Trajectory of PSD Evolution
PSD is the dynamic cumulative outcome of post-stroke neuroinflammation, metabolic disturbances, and dysregulated neuroplasticity. Dysbiosis, intestinal barrier damage, peripheral immune activation, brain network remodeling, and depressive phenotypes evolve progressively across different post-stroke stages (acute, subacute, and chronic phases). For example, the acute phase may drive elevated pro-inflammatory cytokines and reduced beneficial intestinal metabolites; the subacute phase may manifest persistent serum LPS elevation; and the chronic phase may associate default mode network connectivity abnormalities with consolidated long-term depressive symptoms. Additionally, the MG, Mø, T cells, and B cells transition from pro-inflammatory to anti-inflammatory over time, accompanied by Th17/Treg imbalance. Given the heterogeneity of PSD biological trajectories, we propose adopting a longitudinal tracking strategy for multimodal PSD research. Multimodal data—including behavioral scales, multi-sequence neuroimaging, serum inflammatory/metabolomic profiles, and intestinal microbiome sequencing—should be collected simultaneously at key time points (eg, 24 hours, 7 days, 1 month, 3 months, and 6 months). Through the combined application of vector autoregression and trajectory clustering, time-series modeling can be conducted to reveal causal sequential relationships among cross-modal events.
Spatial Specificity: Identifying Hierarchical Pathological Signals within the MGBA
The MGBA consists of anatomically and functionally distinct spatial hierarchies, including gut luminal microbial communities, the intestinal epithelial barrier, and the ENS, to portal vein circulation/vagal nerve pathways, the peripheral circulatory system, the BBB, and ultimately specific brain regions. Different modality data reflect pathological events at varying spatial scales. For example, microbiome data reveal gut microecological disturbances, peripheral inflammatory markers indicate intestinal barrier damage and systemic inflammation, and neuroimaging data capture changes in brain structure, network connectivity, and cerebral metabolites. Therefore, we propose utilizing the MGBA model to elucidate mechanisms of cross-hierarchical signal transduction in PSD. Integrating existing technical approaches, spatial registration and correlation analyses can correlate microbiome features and inflammatory markers with well-localized neuroimaging signatures, constructing multimodal hierarchical pathological signal profiles.
Cross-Domain Compatibility: Driving Synergistic Effects in Multimodal Therapy
The pathological mechanism of PSD involves multi-level interactions within the MGBA, rendering single-modality therapy potentially insufficient for global regulation. Meta-analytic evidence indicates that acupuncture plus rTMS had the highest probability of improving depressive symptoms, with a probability of 49.43% compared to oral Western medication alone.20 This suggests that neuromodulation, microbiota-directed interventions, pharmacotherapy, complementary medicine, and lifestyle management strategies for PSD could be integrated across domains, potentially yielding synergistic effects via multi-target, multi-level mechanisms. However, most current studies on combined therapies remain limited by small sample sizes, high heterogeneity in intervention protocols, and unclear long-term efficacy. Their effect mechanisms and clinical applicability require validation through more high-quality RCTs. Therefore, advocating for combinations of two or more therapeutic modalities must be supported by evidence-based medicine to avoid overinterpreting synergistic effects.
In summary, constructing a multimodal precision diagnosis and treatment framework for PSD based on the MGBA mechanism requires adherence to the Three Principles of Multimodal Integration: Temporal Dynamics emphasizes the implementation of longitudinal recognition strategies during critical time windows of disease progression; Spatial Specificity necessitates targeted interventions across the multiple anatomical and functional domains of the MGBA; and Cross-domain Adaptability advocates for multi-level regulation through combining two or more treatment modalities. However, it is worth noting that most of the current research focuses on validating combinations of macro-level therapeutic strategies, while unified standards and high-level evidence are still lacking for identification tools and treatments targeting key nodes. Although existing meta-analyses suggest potential synergism in multimodal combinations (eg, the established advantage of combined medication and psychotherapy for depressive disorders), research on multimodal interventions specifically for the PSD population, particularly those grounded in MGBA mechanisms, remains limited. We therefore recommend that future research focus on (1) establishing MGBA temporal feature-based staging diagnostic criteria for PSD, (2) developing multi-omics detection technologies with higher spatial resolution, and (3) validating dose-effect relationships of multimodal interventions through high-quality RCTs to mitigate risks of overtreatment or effect cancellation.
Evidence Layer: The MGBA Mechanism-Based Multimodal Framework for PSD Identification
The multimodal identification system integrates neuroimaging, metabolomics, and microbiomics, serving as a critical component of the MGBA-based multimodal precision diagnosis and treatment framework. It enables a comprehensive analysis of brain structure, function, metabolites, gene expression, and gut microbiota in patients with PSD, with multiple layers of data mutually corroborating and complementing each other to provide novel identificatory insights for clinical practice.
Imaging Identification
Multimodal magnetic resonance imaging (MRI) technology serves as one of the initial biomarkers for PSD. The primary multimodal MRI techniques in use currently include diffusion MRI (dMRI), functional MRI (fMRI), hydrogen proton magnetic resonance spectroscopy,1H-MRS and voxel-based morphometry (VBM). Multimodal MRI has revealed alterations in the brain structure, function, and metabolites of individuals with PSD, as well as modifications in the neural network connections within the relevant brain regions.
dMRI has revealed that individuals with PSD exhibit abnormal changes in the white matter microstructure (WMMS) of the prefrontal cortex and frontal-limbic regions associated with emotion, suggesting compromised integrity of the WMMS.21 fMRI study has revealed that the activity of emotion-related areas in patients with PSD is either abnormally hyperactive or hypoactive,22 primarily evidenced by changes in the projection function of the limbic system-cortex-striatum-globus pallidus-thalamus loop (LCSPT). VBM technology has revealed that the reduction in gray matter volume in PSD patients mainly occurs in the prefrontal-limbic areas of the impacted hemisphere.23 Clinically, a decrease in hippocampal volume has been observed in the affected hemisphere of PSD patients. This may pertain to the dysregulated HPA axis, which governs the neuronal activity of the hippocampus.1 H-MRS is used to identify and quantify levels of specific metabolites in brain tissue, including N-acetyl-aspartate (NAA), choline (Cho), phosphocreatine/creatine (PCr), and Glu, among others. Typically, changes in brain metabolites may precede anatomical changes. Research indicates that metabolic abnormalities develop to varying degrees in multiple brain regions following a stroke. A significant increase in Cho/Cr serves as a standard indicator of PSD.24,25 Moreover, the brain forms a complex neural network through the interconnection of multiple brain regions. Patients with PSD exhibit disruptions in associated brain networks, potentially linked to changes in the intrinsic functional connectivity of resting-state networks and increased connectivity between the default network and the salience/cognitive control network.26
Hematological Biomarker Assessment
The identification of neurological markers, serum inflammatory markers, or genetic polymorphisms has significant predictive value for the diagnosis of PSD. These markers can be utilized as important indicators for evaluating efficacy and therapeutic goals.
Patients with PSD exhibit abnormal levels NE. One study found that NE activity and plasma 5-HT levels are significantly reduced in patients with PSD and are closely associated with the severity of depressive symptoms.27 Additional research indicate that plasma glutamate levels in PSD patients are significantly lower than those in non-PSD patients 1 day after acute stroke.28 Furthermore, serum BDNF expression levels may serve as markers for diagnosis and disease evaluation. Reports indicate that blood BDNF levels in individuals with PSD 3–6 months post-stroke are lower than those in stroke patients without depression.29 Changes in NE levels suggest that PSD operates through a multifactorial mechanism. The simultaneous measurement of these markers may be used for early identification of high-risk patients.
The inflammatory response plays a critical role in the pathophysiology of PSD. The balance between pro-inflammatory and anti-inflammatory factors directly influences inflammatory outcomes and long-term patient prognosis. The Identifying serum markers, including levels of pro-inflammatory and anti-inflammatory factor, aids in the diagnosis and prognosis of PSD. Cytokines, such as IL-1, IL-6, and TNF-α, are rapidly produced following acute stroke, with their elevated levels correlating with the progression of PSD.30 C-reactive protein (CRP) is a widely used biomarker for evaluating the extent of inflammation. A cohort study found that elevated CRP levels during the phase of acute stroke signify an augmented risk of PSD.31 Conversely, IL-10 and TGF-β levels have been shown to be reduced in PSD.32 Th17/Treg cytokines influence the synthesis and metabolism of NE and affect cognitive function. Elevated Th17/Treg expression levels are associated with great depression severity in patients.32 Therefore, the inflammatory mechanism of PSD involves complex immune interactions related to oxidative stress, immune regulation, NE, and changes in synaptic plasticity. Unfortunately, inflammatory markers cannot be used as direct predictors for predicting the risk of PSD. A comprehensive multicenter longitudinal clinical studies are needed to develop a dependable prediction model.
Numerous studies have identified differential expression of depression-associated genes in individuals with PSD. Genetic susceptibility may contribute to specific PSD subtypes.33–36 PSD can be achieved through plasma gene expression profiling. The targeted sequencing of genes inside the miRNA-mRNA regulatory network of PSD patients revealed three genes (SPATA2, ZNF208, and YTHDC1) and four upstream miRNAs (miR-6883-5p,miR-6873-3p,miR-4776-3p,and miR-6738-3p), with these miRNAs exhibited distinct expression patterns in PSD. Genetic variations in the p11/tPA/BDNF pathway may lead to PSD. The polymorphisms of the 5-hydroxytryptamine receptor (5-HTR) gene, serotonin transporter gene-linked promoter region (5-HTTLPR), N-methyl-D-aspartate (NMDA) gene, estrogen receptor beta (ERβ) gene, and polymorphism of the apolipoprotein E (ApoE) gene are correlated with the occurrence of PSD. The 5-HTTLPR, ERβ, BDNF, and ApoE demonstrate potential associations with age-related and gender-specific indicators, suggesting that the cumulative genetic risk for PSD in elderly female patients requires special attention.
Gut Microbiota Sequencing
The acquisition of microbial species, kurtosis, and intestinal metabolic disorders through the MGBA approach holds significant importance for PSD identification.
PSD is characterized by increased pro-inflammatory and pathogenic bacteria, along with decreased beneficial bacteria, in terms of species composition and kurtosis. A clinical study demonstrated that the diversity of gut microbiota is elevated in individuals with PSD, marked by significant enrichment of Phascolarctobacterium, Escherichia, and Clostridium genera, while Bacteroides, Prevotella, and Ruminococcus genera showed significant reduction.37 Fundamental research revealed decreased gut microbiota abundance in PSD rat models, including Clostridium (phylum Firmicutes), Blautia, and Streptococcus genera, which showed negative correlation with depression severity.38
The biological activity of the gut in PSD is being investigated through microbial transcriptomics, fecal metabolomics, proteomics, and other methodologies. The gut microbiota possesses ten times the number of genes present in the human genome. The gene products enter the bloodstream, influencing host metabolism and contributing to the onset and progression of PSD. For example, Elevated acetylation of histone H3/H4 in the promoter region of the BDNF gene has been negatively correlated with depression severity.39 The fecal metabolomics characteristics of animal models exhibit elevated concentrations of amino acids, carbohydrates, lipids and lipid-like molecules, nucleotides, and organic heterocyclic compounds, with a particularly notable abundance of steroid production in PSD.40 Additionally, gut microbial metabolites, including ScFAs and NE, frequently influence PSD. A recent FMT study demonstrated that the gut microbiota of individuals with inflammatory depression showed elevated Bacteroides level and reduced Clostridium level.38 The abnormal expansion of ScFAs-producing bacteria lead to butyrate metabolic abnormalities, which can affect peripheral 5-HT levels and disrupt emotional modulation. These findings suggest that PSD incidence is linked to gut microbiota and metabolic dysregulation, highlighting the Firmicutes-ScFAs-lipid metabolism axis as a potential pathway within the MGBA framework. Proteomics studies have identified a connection between protein expression in the PSD host environment and an increased risk of depression, lipid problems, and infections. A clinical study found that PSD patients exhibited increased plasma levels of intestinal fatty acid-binding protein (iFABP),41 a biomarker of intestinal permeability, which significantly correlated with depression severity and lipid metabolism. Lipocalin-2 (LCN2), a key regulator of appetite and inflammation, was elevated in the blood and cerebrospinal fluid of stroke patients, with higher LCN2 levels serving as an independent risk factor for PSD.42 Individuals with severe brain injury often experience transient reductions in immunoglobulin G (IgG) levels,43 increasing the risk of secondary infection. In clinical practice, the gut microbial characteristics of PSD can be delineated through a multimodal approach integrating gene expression profiles, metabolite profiles, and protein markers to enhance diagnostic specificity and sensitivity, thereby informing personalized interventions.
TCM Identification
The concept of entirety and treatment based on syndrome differentiation are characteristics of TCM. As such, TCM has long been capable of identifying corresponding main, secondary, and concurrent symptoms and has accumulated substantial experience in differentiating various syndrome patterns, for example, “Liver Depression and Spleen Deficiency” and “Heart-Spleen Dual Deficiency Syndrome.” However, due to its heavy reliance on subjective clinical experience and a lack of objective, unified diagnostic criteria, the current role of TCM in the precise identification and subtyping of PSD remains limited. Therefore, integrating TCM syndrome differentiation with a multimodal identification system for PSD likely necessitates the establishment of objective diagnostic standards and quantifiable indicators. A promising strategy entails correlating TCM syndrome patterns with contemporary biomarkers, such as neuroimaging indicators and microbial profiles. This strategy aims to construct a translational bridge between the TCM disease mechanism pathway of the “Intestine-Spleen/Stomach-Brain Axis” with modern biological pathways.
Application Layer: The MGBA Mechanism-Based Multimodal Framework for PSD Intervention
The multimodal intervention framework closely aligns with the MGBA-based multimodal precision diagnosis and treatment framework for PSD. Targeting the core role of MGBA dysfunction in PSD pathogenesis, we integrate CNS interventions, gut microbiota modulation, pharmacotherapy, complementary medicine, and lifestyle interventions to address PSD at different hierarchical levels.
CNS Interventions
CNS interventions for PSD encompass invasive techniques, such as deep brain stimulation (DBS), transcutaneous vagus nerve stimulation (tVNS), and non-invasive brain stimulation methods (NBIS), such as repetitive transcranial magnetic stimulation (rTMS) and transcranial direct current stimulation (tDCS). DBS improves connectivity between the prefrontal-limbic areas, improves synaptic plasticity (eg, by increasing BDNF expression), and promotes hippocampal neurons. A study demonstrated that unilateral thalamic nucleus DBS significantly improved PSD symptoms.44 tVNS may stimulate both the upstream and downstream pathways of VNS. In the upstream pathway, VN-mediated 5-HT infiltrates hippocampal neurons through 5-HT1 receptors,45 potentially activating the BDNF pathway and protecting the BBB. Simultaneously, VN afferent fibres may stimulate GCs production via the HPA axis,46 exerting anti-inflammatory effects and alleviating depressive symptoms. Additionally, In the downstream pathway, tVNS enhances intestinal glial cell activation, reduces the levels of IL-6 and TNF-α, and improves intestinal barrier function. NIBS is a neuromodulation approach with the potential to transform PSD treatment paradigms. rTMS significantly improves PSD by modulating cortical circuit excitability and targeting specific brain regions, such as the left dorsolateral prefrontal cortex(DLPFC). Research studies that low-intensity rTMS, combined with antidepressant therapy, reduces IL-6 and TNF-α levels in individuals with PSD.47 The antidepressant mechanism of tDCS may involve enhancing motor cortical plasticity in the DLPFC. In neuroelectric stimulation therapies, flexible selection of stimulation sites, methods, and parameters can improve PSD by modulating neuronal excitability, promoting neural circuit remodelling, influencing gut microbiota and NE levels, and suppressing inflammation progression.
Gut Microbiota Interventions
Gut microbiota modulation in PSD patients is achieved through psychobiotics (including probiotics/prebiotics) and FMT, which may ameliorate MGBA function via a bottom-up mechanism and promote recovery from central mental disorders.
Psychobiotics may act through multiple pathways and brain signalling networks, primarily accomplished via the ENS, vagus nerve stimulation (VNS), and metabolic regulatory pathways, which modulate Th17/Treg immune balance and restore NE homeostasis. Currently, Lactobacillus and Bifidobacterium strains are the most widely used in therapeutic applications. In the early stages of stroke, adjunctive therapy with probiotics like Lactobacillus and Bifidobacterium can improve intestinal nutritional status, enhance immunological tolerance, reduce the occurrence of diarrhoea, and beneficial bacteria can lower the patient’s plasma LPS levels. Evidence suggests that beneficial bacteria may enhance ScFA production to promote intestinal tight junction protein (TJP, eg, ZO-1) expression, repair the intestinal mucosal barrier, and concurrently reduce glutamate excitotoxicity and upregulate GABA and BDNF levels,48 thereby exerting potent antidepressant effects. Clinical observations further indicate that Bacillus spp., Serratia marcescens, and Clostridium spp. can modulate gut microbiota and yield antidepressant effects. Therefore, the use of psychobiotics may be effective in treating PSD.
FMT can restore gut microbiota, reduce neuroinflammation, repair the intestinal barrier, and reestablish NE balance, making it a promising therapy for PSD. Animal studies have shown that the VNS may be a crucial communication pathway for FMT and that sigma-1 receptors (Sig-1R) and NLRP3 inflammasomes, which mediate neurochemical effects, may be significant targets for FMT efficacy.49,50 Numerous clinical studies have demonstrated that FMT markedly alleviates depression symptoms and effectively treats recurrent Clostridium difficile infections.51–53 However, despite being less invasive, the long-term safety of FMT (eg, risks of infection and metabolic abnormalities) remains uncertain, and the lack of standardized clinical protocols limits its current application.
Pharmacological Intervention
Current treatments for PSD primarily rely on antidepressant therapy. The modern approach to treating PSD has shifted from monoamine modulation to a multifaceted therapeutic strategy, utilizing monoamines to correct NE imbalance, glutamate modulators to protect neurons, and anti-inflammatory agents to reduce inflammatory damage.
Monoamine medications include tricyclic antidepressants (TCAs), selective serotonin reuptake inhibitors (SSRIs), serotonin-norepinephrine reuptake inhibitors (SNRIs), monoamine oxidase inhibitors (MAOIs), and norepinephrine-dopamine reuptake inhibitors (NDRIs).54 SSRIs, SNRIs, and TCAs are the most commonly prescribed and serve as first-line treatments due to their proven effectiveness. SSRIs, such as fluoxetine, paroxetine, and sertraline, improve cerebral blood flow by inhibiting 5-HT transporters and receptor targets,55 thereby reducing neuronal loss in the peri-infarct region. However, drugs within the same class may have additional pharmacological effects. For example, fluoxetine can inhibit post-ischemic brain inflammation and reverse cellular changes in the prefrontal cortex-limbic system;56 paroxetine has mild anti-Cho effects; and sertraline bind effectively to the DA transporter, significantly improving HPA axis dysfunction.57 Additionally, glutamatergic antidepressants (eg, AMPA receptor antagonists and NMDA receptor blockers) improve PSD outcomes primarily by stabilizing the glutamate system. Clinically, they are considered an optimal adjunct to monoamine-based therapies for PSD. Immunomodulation may be a component of MGBA targeted treatments. Nonsteroidal anti-inflammatory medications (NSAIDs) with anti-inflammatory properties may be an alternate therapy for PSD. Certain study found that statins, such as atorvastatin, may reduce the incidence of PSD by suppressing inflammatory mediators like IL-6.58 Given the wide variety of antidepressants, we suggest the necessity of distinguishing the pharmacological therapy of PSD from that of general depression to avoid inappropriate medication use. Before developing a PSD treatment plan, clinicians must carefully assess the patient’s stroke type (eg, ischemic or hemorrhagic) and PSD subtype (eg, inflammatory depression or NE imbalance type). For instance, NSAIDs are only recommended for inflammatory depression.
Complementary Medicine Therapy
Traditional Chinese medicine(TCM) and acupuncture can improve mood, cognitive function, and physical symptoms (such as fatigue, sleep problems, and anorexia) in patients with PSD, providing additional options and chances for these individuals.
The mechanism of TCM may operate through three primary pathways: Firstly, it suppresses overactivation activation of the HPA axis, thereby reducing levels of CRH, ACTH, and CORT. For example, extracts of St. John’s wort can increase the mRNA and protein expression of receptor-kinase-B(TrkB), 5-HTAR, and GR,59 restoring HPA axis balance. Second, TCM modulates neuroinflammation and regulates NE levels to support neuronal health. For instance, curcumin and geniposide,60,61 among other botanicals, can inhibit TLR4 by boosting AKT signalling through related pathways, thereby reducing stress-induced neuronal death. It targets lncRNA to modulate gene expression, enhance differential proteins in Glu/GABA synapses, BDNF, and mitogen-activated protein kinase (MAPK)signaling pathways, improving NE balance, synaptic plasticity, and cellular signaling to exert a synergistic anti-PSD effect. Third, TCM modulates the composition of gut microbiota, kurtosis, and ScFAs metabolites. For example, plant polysaccharides (eg, ginkgo polysaccharides) can alleviate PSD by reducing the abundance of depression-associated gut microbiota,62 increasing the presence of Lactobacillus and Clostridium. In clinical practice, the author observed that the Xiaoyao San compound can has been observed to have antidepressant effects. However, due to its complex composition and diverse active constituents, further research is needed to explore its targets and pharmacological mechanisms.
Acupuncture therapy may intervene PSD by facilitating the MGBA interaction, enabling bidirectional communication between the NEI networks. Acupuncture modulates the signalling pathway of the LCSPT neural circuit, attenuates hyperactivity of the HPA axis,63 regulates inflammatory mediators such as TNF-α, IL-1β, and LPS, increase Th2/ILC2-related cytokines, and restores the Treg/Th17 balance to prevent neuroinflammation.64,65 Furthermore, acupuncture exerts an antidepressant influence from the gastrointestinal tract to the brain. It may may exert antidepressant effects through the MGBA by restoring gut microbiota homeostasis. This mechanism could entail adjusting microbial proportions, distribution patterns (kurtosis), and abundance to protect the intestinal mucosal barrier, alongside modulating key metabolites (eg, DA, 5-HT, tryptophan, indole derivatives) and metabolic pathways via NE regulation, thereby preserving neural integrity.66 In clinical practice, acupoints such as Baihui (GV 20), Taichong (LR 3), and Siguan (EX-HN 1) may elicit antidepressant effects through complex neuroendocrine pathways. However, this involves complex NEI mechanisms, which warrant further exploration.
In summary, TCM and acupuncture exhibit unique potential for intervening in MGBA dysfunction and alleviating PSD symptoms due to their multi-component, multi-target, and holistic regulatory characteristics. This knowledge provides a foundation for the MGBA-based multimodal precision treatment framework for PSD proposed in this paper from the perspective of TCM. Nevertheless, efficacy varies among different TCM and acupuncture protocols due to heterogeneous herbal formula compositions, acupoint selections, and electroacupuncture parameters. Furthermore, the chemical complexity of multi-herbal TCM formulations presents significant challenges, as their active constituents, pharmacokinetic profiles, and in vivo targets remain largely uncharacterized, which consequently leads to considerable inter-individual variability in therapeutic responses. Moreover, the credibility of efficacy conclusions has long been constrained by methodological limitations, including non-standardized syndrome differentiation criteria and inadequate control of placebo effects, compounded by a scarcity of high-quality RCTs. We urgently need multinational, multicenter RCTs adhering to STRICTA standards to validate optimal intervention time windows and refine precision treatment strategies through multimodal research.
Lifestyle Interventions
Lifestyle interventions can serve as a primary treatment for patients with PSD who exhibit poor medication adherence, suboptimal response to medication, or are deemed unsuitable for pharmacological therapy. Psychological stress disorders are a key contributing factor to PSD. Psychological interventions include cognitive behavioural therapy (CBT), psychosocial therapies, and music therapy. CBT may help alleviate depression through cognitive restructuring and behavioral activation. Psychosocial therapies, such as motivational interviewing, transitional care, psychoeducation, and social support, can improve emotional well-being after a stroke. Music therapy may stimulate the senses, enhances brain activity, inhibits the HPA axis, and modulates the ANS to produce an immunomodulatory impact,67 thereby improving neurocognition in patients with PSD.
Exercise intervention is a widely used rehabilitation strategy that improves physical fitness, reduces fatigue, helps patients regain the ability to perform daily activities and social interactions, and decreases feelings of isolation. Research has shown that moderate exercise training may reduce CRH secretion in cerebral infarction lesions, restore HPA function, and prevent the excessive release of Glu,68 thereby reducing the incidence of PSD. Aerobic exercise may increase the expression of synaptic proteins in the hippocampus (eg, PSD-95, synaptophysin, and neurotrophin-related protein (GAP)-43) and influence the plasma and hypothalamus proteome,69 reducing neuroinflammation and alleviating PSD symptoms. Additionally, fMRI study has revealed that robot-assisted exercise induces changes in connectivity between the patient’s premotor cortex, motor cortex, primary motor cortex, and supplementary motor area,70 suggesting that supplementary motor activity may improve motor control and reduce physical symptoms of PSD. To minimize the risk of chronic depression, lifestyle modifications are recommended as a non-pharmacological approach for PSD, emphasizing early intervention and sustained adherence to treatment for optimal outcomes.
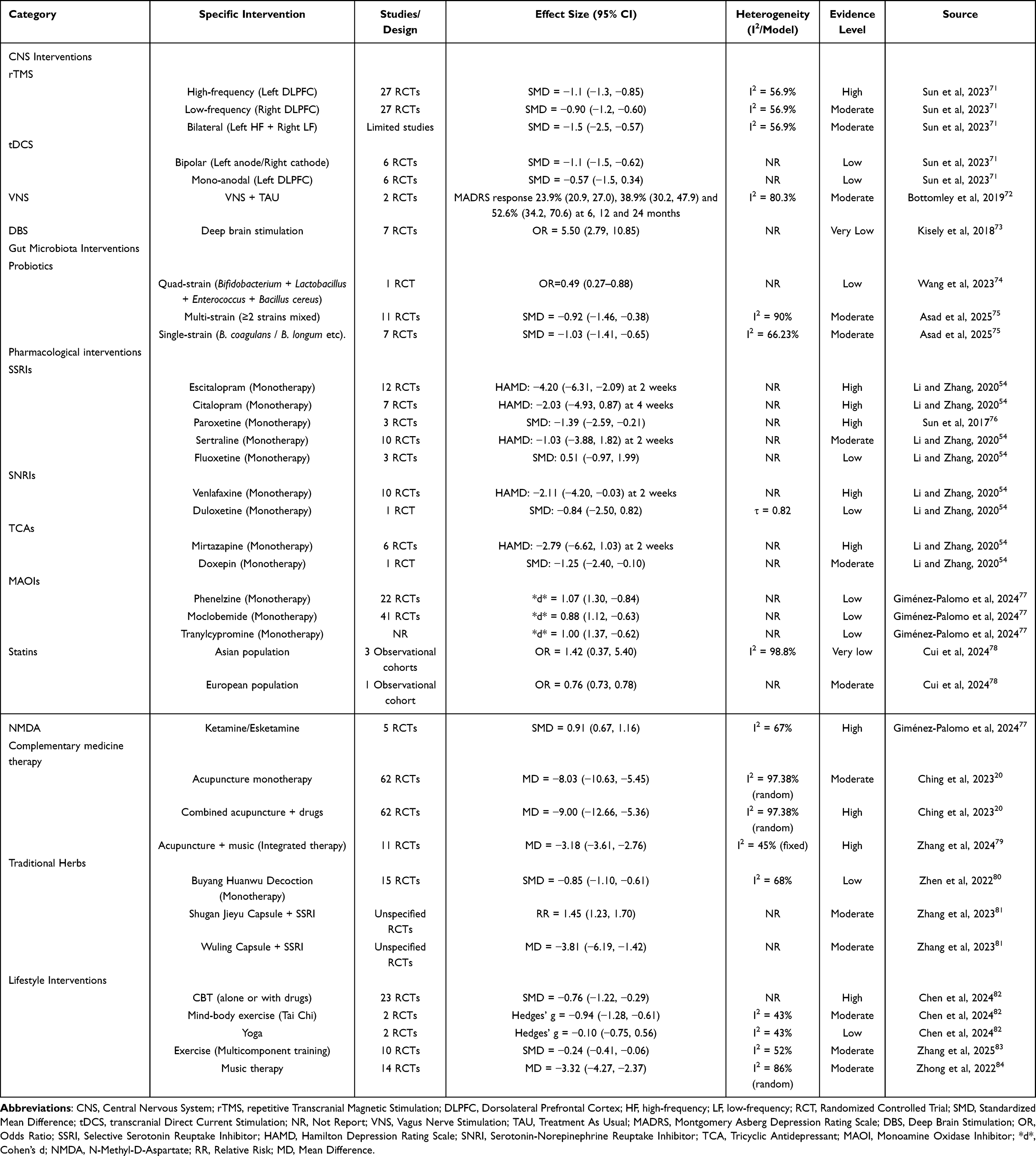 |
Table 1 Evidence Hierarchy for Multimodal Interventions in Post-Stroke Depression |
To further this review and integrate the current evidence on MGBA-based multimodal interventions for PSD, we synthesize findings from relevant meta-analyses and cohort studies to establish an evidence hierarchy matrix for PSD interventions (Table 1). This matrix aims to provide clinicians and researchers with a structured overview of the strength of evidence across various strategies, including CNS interventions, gut microbiota interventions, pharmacological interventions, complementary medicine therapy, lifestyle interventions. This table summarizes key aspects such as study design, effect size, heterogeneity, and evidence grading. It should be noted that the conclusions presented here are based on currently available data, and the strength of recommendations may evolve as future high-quality studies emerge. Individualized application to specific patient circumstances remains essential in clinical practice. It is hoped that this matrix will serve as a practical and structured reference to support evidence-based clinical decision-making and further research in the field.
Discussion
Research Contributions
The primary contribution of this review lies in constructing and elaborating an MGBA-based multimodal precision diagnosis and treatment framework for PSD from three core dimensions: neurobiology, gut microbiota interactions, and TCM interventions. It encompasses four levels: mechanism exploration, multimodal data integration principles, evidence identification, and clinical application. The framework designs a multimodal identification system that integrates neuroimaging, metabolic, and microbial multi-omics data, preliminarily outlining a potential MGBA-related biological network for PSD. At the same time, it proposes a multimodal intervention system that consolidates strategies encompassing CNS interventions, microbiota modulation, pharmacotherapy, complementary medicine, and lifestyle management. This framework not only deepens the understanding of PSD pathogenesis but also provides targeted and effective guidance for clinical practice.
Evidence Limitations
Current research exhibits several limitations: (1) Insufficient depth and breadth in MGBA mechanism studies for PSD: Most evidence derives from animal models or cross-sectional clinical observations, with a lack of large-sample longitudinal studies in human PSD patients, hindering the establishment of causal relationships between HPA axis abnormalities, microbiota-immune-neural circuit alterations, and PSD symptoms. (2) Technical challenges in multimodal data integration: The methods for acquiring, standardizing, and analyzing data from neuroimaging, metabolomics, and microbiomics are inconsistent. Moreover, the detection indicators for critical time-point biomarkers are not well-defined. These issues complicate the assessment of correlations between different datasets and hinder the construction and validation of biomarker panels. (3) Methodological shortcomings in TCM research: Although TCM and acupuncture show potential in regulating the MGBA and improving PSD symptoms, relevant studies have methodological shortcomings. These include non-uniform syndrome differentiation criteria, inadequate placebo control, and unstandardized stimulation parameters (eg, frequency and depth of electroacupuncture). Additionally, the high variability in herbal formulas and acupoint selections limits the reproducibility of results, and there is a lack of high-quality evidence-based support. (4) Inadequate interdisciplinary collaboration: Current research is often confined to individual disciplines, with limited integration across clinical medicine, basic science, bioinformatics, and policy-making. This lack of collaboration impedes the translation of multimodal systems from theoretical concepts to practical applications.
Conclusion
As a common and complex post-stroke complication, the pathogenesis of PSD involves intricate interactions between neurobiology and gut microbiota. To our knowledge, this review represents the first attempt to integrate three core dimensions—neurobiological mechanisms, gut microbiota interactions, and TCM interventions—within four analytical frameworks: mechanism elucidation, evidence synthesis, multimodal profiling, and clinical translation. By synthesizing multi-omics data, including neuroimaging, metabolomic, and microbial profiles, we propose a novel multimodal precision framework for PSD grounded in MGBA. Clinically, this framework facilitates early intervention through dynamic monitoring, precise therapeutic strategies, and standardized collaborative workflows. By incorporating integrative treatment modalities such as neuromodulation, gut microbiota-directed therapy, and TCM, it provides a structured approach to multimodal treatment design. Methodologically, it offers an operational framework for deciphering the “multi-component, multi-target” mechanisms of TCM formulas within the MGBA context and informs the design of high-quality RCTs. We anticipate that this framework will contribute to transforming diagnostic and therapeutic models for PSD, advance research paradigms in the field, and offer actionable insights for future investigation.
Future Perspectives
Future research could focus on clarifying the dynamic causal relationships within the MGBA in PSD through large-sample longitudinal cohorts. The development of high-resolution multi-omics technologies is essential to establish a PSD-specific biomarker panel. AI should be leveraged to integrate multimodal data and decipher temporal biomarker interaction networks during critical neuroplasticity windows. Meanwhile, efforts should focus on enhancing the standardization and global recognition of TCM research. This entails creating a standardized database for acupuncture parameters, implementing multinational RCTs to validate TCM efficacy, and defining integrated diagnostic-therapeutic criteria that harmonize TCM with Western medicine. Furthermore, high-quality clinical trials are needed to optimize multimodal treatment strategies, incorporating lifestyle interventions into the entire management process and advocating for medical insurance policies to cover microbial testing and multimodal assessment tools. Finally, interdisciplinary collaboration should be strengthened by encouraging international health organizations such as the WHO to prioritize research on the integration of traditional and modern medicine. Establishing global data-sharing platforms and implementing evidence-based policies will be crucial to achieving precision prevention and personalized therapeutic approaches for PSD.
Data Sharing Statement
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Acknowledgments
The authors would like to express their gratitude to Professor L.X. Wang from The First Affiliated Hospital of Dalian Medical University for his valuable suggestions and writing support.
Author Contributions
MS defined the core concepts, designed the framework, and drafted the initial manuscript (main body). QX and ZS conducted a comprehensive literature search and integration. YZ and MY wrote the Traditional Chinese Medicine section and oversaw structural integrity. PY performed review and revision. All authors consent to the submission and accept accountability for the work.
Funding
This study was supported by grants from Science and Technology Research Program of Chongqing Municipal Education Commission (Grant No. KJQN202402721), Science and Technology Research Program of Chongqing Municipal Education Commission (Grant No. KJQN202202706).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Taylor-Rowan M, Momoh O, Ayerbe L, Evans JJ, Stott DJ, Quinn TJ. Prevalence of pre-stroke depression and its association with post-stroke depression: a systematic review and meta-analysis. Psychol Med. 2019;49(4):685–696. doi:10.1017/s0033291718002003
2. Krick S, Koob JL, Latarnik S, et al. Neuroanatomy of post-stroke depression: the association between symptom clusters and lesion location. Brain Commun. 2023;5(2):fcad275. doi:10.1093/braincomms/fcad275
3. Shi M, Li Z, Tang Z, et al. Exploring the pathogenesis and treatment of PSD from the perspective of gut microbiota. Brain Res Bull. 2024;215:111022. doi:10.1016/j.brainresbull.2024.111022
4. Tyler Patterson T, Grandhi R. Gut Microbiota and Neurologic Diseases and Injuries. Adv Exp Med Biol. 2020;1238(4):73–91. doi:10.1007/978-981-15-2385-4_6
5. Datta A, Saha C, Godse P, Sharma M, Sarmah D, Bhattacharya P. Neuroendocrine regulation in stroke. Trends Endocrinol Metab. 2023;34(5):260–277. doi:10.1016/j.tem.2023.02.005
6. Ashique S, Mohanto S, Ahmed MG, et al. Gut-brain axis: a cutting-edge approach to target neurological disorders and potential synbiotic application. Heliyon. 2024;10(6):e34092. doi:10.1016/j.heliyon.2024.e34092
7. Makris K, Haliassos A, Chondrogianni M, Tsivgoulis G. Blood biomarkers in ischemic stroke: potential role and challenges in clinical practice and research. Crit Rev Clin Lab Sci. 2018;55(7):294–328. doi:10.1080/10408363.2018.1461190
8. Nagy EE, Frigy A, Szász JA, Horváth E. Neuroinflammation and microglia/macrophage phenotype modulate the molecular background of post-stroke depression: a literature review. Exp Ther Med. 2020;20(8):2510–2523. doi:10.3892/etm.2020.8933
9. Kang Y, Yang Y, Wang J, Ma Y, Cheng H, Wan D. Correlation between Intestinal Flora and Serum Inflammatory Factors in Post-stroke Depression in Ischemic Stroke. J Coll Physicians Surg Pak. 2021;31(9):1224–1227. doi:10.29271/jcpsp.2021.10.1224
10. Samuelson DR, Gu M, Shellito JE, et al. Intestinal Microbial Products From Alcohol-Fed Mice Contribute to Intestinal Permeability and Peripheral Immune Activation. Alcohol Clin Exp Res. 2019;43(10):2122–2133. doi:10.1111/acer.14176
11. Liesz A, Hu X, Kleinschnitz C, Offner H. Functional role of regulatory lymphocytes in stroke: facts and controversies. Stroke. 2015;46(5):1422–1430. doi:10.1161/strokeaha.114.008608
12. Alsbrook DL, Di Napoli M, Bhatia K, et al. Neuroinflammation in Acute Ischemic and Hemorrhagic Stroke. Curr Neurol Neurosci Rep. 2023;23(12):407–431. doi:10.1007/s11910-023-01282-2
13. Agus A, Planchais J, Sokol H. Gut Microbiota Regulation of Tryptophan Metabolism in Health and Disease. Cell Host Microbe. 2018;23(13):716–724. doi:10.1016/j.chom.2018.05.003
14. Margolis KG, Cryan JF, Mayer EA. The Microbiota-Gut-Brain Axis: from Motility to Mood. Gastroenterology. 2021;160(14):1486–1501. doi:10.1053/j.gastro.2020.10.066
15. Coyne MJ, Comstock LE. Type VI Secretion Systems and the Gut Microbiota. Microbiol Spectr. 2019;7(15):10–128. doi:10.1128/microbiolspec.PSIB-0009-2018
16. Silva YP, Bernardi A, Frozza RL. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front Endocrinol. 2020;11(16):25. doi:10.3389/fendo.2020.00025
17. Cryan JF, O’Riordan KJ, Sandhu K, Peterson V, Dinan TG. The gut microbiome in neurological disorders. Lancet Neurol. 2020;19(17):179–194. doi:10.1016/s1474-4422(19)30356-4
18. Kaelberer MM, Buchanan KL, Klein ME, et al. A gut-brain neural circuit for nutrient sensory transduction. Science. 2018;361(18):5236. doi:10.1126/science.aat5236
19. Stasi C, Sadalla S, Milani S. The Relationship Between the Serotonin Metabolism, Gut-Microbiota and the Gut-Brain Axis. Curr Drug Metab. 2019;20(19):646–655. doi:10.2174/1389200220666190725115503
20. Lam CW, HJ L, Guo J, et al. Acupuncture for post-stroke depression: a systematic review and network meta-analysis. BMC Psychiatry. 2023;23(7):314. doi:10.1186/s12888-023-04749-1
21. Oestreich LKL, Wright P, O’Sullivan MJ. Microstructural changes in the reward system are associated with post-stroke depression. Neuroimage Clin. 2020;28(20):102360. doi:10.1016/j.nicl.2020.102360
22. Egorova N, Veldsman M, Cumming T, Brodtmann A. Fractional amplitude of low-frequency fluctuations (fALFF) in post-stroke depression. Neuroimage Clin. 2017;16(21):116–124. doi:10.1016/j.nicl.2017.07.014
23. Hong W, Zhao Z, Wang D, et al. Altered gray matter volumes in post-stroke depressive patients after subcortical stroke. Neuroimage Clin. 2020;26(22):102224. doi:10.1016/j.nicl.2020.102224
24. Zhang L, Sui R, Zhang L, Zhang Z. Morphological and Metabolic Alteration of Cerebellum in Patients with Post-Stroke Depression. Cell Physiol Biochem. 2016;40(23):420–430. doi:10.1159/000452557
25. Zhang L, Li M, Sui R. Correlation between cerebellar metabolism and post-stroke depression in patients with ischemic stroke. Oncotarget. 2017;8(24):91711–91722. doi:10.18632/oncotarget.21063
26. Jaywant A, DelPonte L, Kanellopoulos D, O’Dell MW, Gunning FM. The Structural and Functional Neuroanatomy of Post-Stroke Depression and Executive Dysfunction: a Review of Neuroimaging Findings and Implications for Treatment. J Geriatr Psychiatry Neurol. 2022;35(25):3–11. doi:10.1177/0891988720968270
27. Song Y, Liu J, Zang DW, Tian Z, Lu Y. Correlation between neurotransmitter power and plasma 5-HT level with post-stroke depression. Chin J Gerontol. 2017;37(26):439–440. doi:10.3969/j.issn.1005-9202.2017.02.081
28. Geng LY, Qian FY, Qian JF, Zhang ZJ. The combination of plasma glutamate and physical impairment after acute stroke as a potential indicator for the early-onset post-stroke depression. J Psychosom Res. 2017;96(27):35–41. doi:10.1016/j.jpsychores.2017.01.006
29. Han Z, Wang Y, Qi L, et al. Differential Association of Serum BDNF With Poststroke Depression and Poststroke Anxiety. Arch Phys Med Rehabil. 2020;101(28):1355–1366. doi:10.1016/j.apmr.2020.04.011
30. Chen Y, Pu J, Liu Y, et al. Pro-inflammatory cytokines are associated with the development of post-stroke depression in the acute stage of stroke: a meta-analysis. Top Stroke Rehabil. 2020;27(29):620–629. doi:10.1080/10749357.2020.1755813
31. Yang Y, Zhu L, Zhang B, Gao J, Zhao T, Fang S. Higher levels of C-reactive protein in the acute phase of stroke indicate an increased risk for post-stroke depression: a systematic review and meta-analysis. Neurosci Biobehav Rev. 2022;134(30):104309. doi:10.1016/j.neubiorev.2021.08.018
32. Huang C, Zhang F, Li P, Song C. Low-Dose IL-2 Attenuated Depression-like Behaviors and Pathological Changes through Restoring the Balances between IL-6 and TGF-β and between Th17 and Treg in a Chronic Stress-Induced Mouse Model of Depression. Int J Mol Sci. 2022;23(31):13856. doi:10.3390/ijms232213856
33. Qiu H, Shen L, Shen Y, Mao Y. Identification of a miRNA-mRNA regulatory network for post-stroke depression: a machine-learning approach. Front Neurol. 2023;14(32):1096911. doi:10.3389/fneur.2023.1096911
34. Liang J, Yue Y, Jiang H, et al. Genetic variations in the p11/tPA/BDNF pathway are associated with post stroke depression. J Affect Disord. 2018;226(33):313–325. doi:10.1016/j.jad.2017.09.055
35. Wang Y, Liu H, Jiang Y, Shi X, Shao Y, Xu ZX. Meta-analysis of 5-hydroxytryptamine transporter gene promoter region polymorphism and post-stroke depression. J Int Med Res. 2020;48(34):300060520925943. doi:10.1177/0300060520925943
36. Zhang Z, Mu J, Li J, Li W, Song J. Aberrant apolipoprotein E expression and cognitive dysfunction in patients with poststroke depression. Genet Test Mol Biomarkers. 2013;17(35):47–51. doi:10.1089/gtmb.2012.0253
37. Fan WT, Yan YM, Bie YL, Wang Q. Diversity of intestinal microflora in patients with depression after stroke. Nan Fang Yi Ke Da Xue Xue Bao. 2016;36(36):1305–1311.
38. Liu P, Liu Z, Wang J, et al. Immunoregulatory role of the gut microbiota in inflammatory depression. Nat Commun. 2024;15(37):3003. doi:10.1038/s41467-024-47273-w
39. Fuchikami M, Yamamoto S, Morinobu S, Takei S, Yamawaki S. Epigenetic regulation of BDNF gene in response to stress. Psychiatry Invest. 2010;7(38):251–256. doi:10.4306/pi.2010.7.4.251
40. Jiang W, Gong L, Liu F, Ren Y, Mu J. Alteration of Gut Microbiome and Correlated Lipid Metabolism in Post-Stroke Depression. Front Cell Infect Microbiol. 2021;11(39):663967. doi:10.3389/fcimb.2021.663967
41. Zhong J, Chen J, Cao M, et al. Elevated plasma intestinal fatty acid binding protein and aberrant lipid metabolism predict post-stroke depression. Heliyon. 2022;8(40):e11848. doi:10.1016/j.heliyon.2022.e11848
42. Liu Y, Liu L, Zhi Z, et al. Higher serum lipocalin 2 is associated with post-stroke depression at discharge. BMC Neurol. 2023;23(41):294. doi:10.1186/s12883-023-03319-y
43. Liesz A, Roth S, Zorn M, Sun L, Hofmann K, Veltkamp R. Acquired Immunoglobulin G deficiency in stroke patients and experimental brain ischemia. Exp Neurol. 2015;271(42):46–52. doi:10.1016/j.expneurol.2015.04.021
44. Birchall EL, Walker HC, Cutter G, et al. The effect of unilateral subthalamic nucleus deep brain stimulation on depression in Parkinson’s disease. Brain Stimul. 2017;10(43):651–656. doi:10.1016/j.brs.2016.12.014
45. Shin HC, Jo BG, Lee CY, Lee KW, Namgung U. Hippocampal activation of 5-HT(1B) receptors and BDNF production by vagus nerve stimulation in rats under chronic restraint stress. Eur J Neurosci. 2019;50(44):1820–1830. doi:10.1111/ejn.14368
46. Creutzberg KC, Sanson A, Viola TW, et al. Long-lasting effects of prenatal stress on HPA axis and inflammation: a systematic review and multilevel meta-analysis in rodent studies. Neurosci Biobehav Rev. 2021;127(45):270–283. doi:10.1016/j.neubiorev.2021.04.032
47. Pan J, Li H, Wang Y, et al. Effects of low-frequency rTMS combined with antidepressants on depression in patients with post-stroke depression: a systematic review and meta-analysis. Front Neurol. 2023;14(46):1168333. doi:10.3389/fneur.2023.1168333
48. Gruenbaum BF, Merchant KS, Zlotnik A, Boyko M. Gut Microbiome Modulation of Glutamate Dynamics: implications for Brain Health and Neurotoxicity. Nutrients. 2024;16(47):4405. doi:10.3390/nu16244405
49. Kelly JR, Borre Y, OB C, et al. Transferring the blues: depression-associated gut microbiota induces neurobehavioural changes in the rat. J Psychiatr Res. 2016;82(48):109–118. doi:10.1016/j.jpsychires.2016.07.019
50. Xia CY, Guo YX, Lian WW, et al. The NLRP3 inflammasome in depression: potential mechanisms and therapies. Pharmacol Res. 2023;187(49):106625. doi:10.1016/j.phrs.2022.106625
51. Chinna meyyappan A, Forth E, Wallace CJK, Milev R. Effect of fecal microbiota transplant on symptoms of psychiatric disorders: a systematic review. BMC Psychiatry. 2020;20(50):299. doi:10.1186/s12888-020-02654-5
52. Green JE, Berk M, Loughman A, et al. FMT for psychiatric disorders: following the brown brick road into the future. Bipolar Disord. 2021;23(51):651–655. doi:10.1111/bdi.13124
53. Tian H, Cui J, Ye C, et al. Depletion of butyrate-producing microbes of the Firmicutes predicts nonresponse to FMT therapy in patients with recurrent Clostridium difficile infection. Gut Microbes. 2023;15(52):2236362. doi:10.1080/19490976.2023.2236362
54. Li X, Zhang C. Comparative efficacy of nine antidepressants in treating Chinese patients with post-stroke depression: a network meta-analysis. J Affect Disord. 2020;266(53):540–548. doi:10.1016/j.jad.2020.02.005
55. Jing D, Hou X, Guo X, et al. Astrocytes in Post-Stroke Depression: roles in Inflammation, Neurotransmission, and Neurotrophin Signaling. Cell Mol Neurobiol. 2023;43(54):3301–3313. doi:10.1007/s10571-023-01386-w
56. Zahrai A, Vahid-Ansari F, Daigle M, Albert PR. Fluoxetine-induced recovery of serotonin and norepinephrine projections in a mouse model of post-stroke depression. Transl Psychiatry. 2020;10(55):334. doi:10.1038/s41398-020-01008-9
57. Mead GE, Hsieh CF, Lee R, et al. Selective serotonin reuptake inhibitors for stroke recovery: a systematic review and meta-analysis. Stroke. 2013;44(56):844–850. doi:10.1161/strokeaha.112.673947
58. Kang HJ, Bae KY, Kim SW, et al. Effects of interleukin-6, interleukin-18, and statin use, evaluated at acute stroke, on post-stroke depression during 1-year follow-up. Psychoneuroendocrinology. 2016;72(57):156–160. doi:10.1016/j.psyneuen.2016.07.001
59. Grundmann O, Lv Y, Kelber O, Butterweck V. Mechanism of St. John’s wort extract (STW3-VI) during chronic restraint stress is mediated by the interrelationship of the immune, oxidative defense, and neuroendocrine system. Neuropharmacology. 2010;58(58):767–773. doi:10.1016/j.neuropharm.2009.12.014
60. Cai L, Li WT, Zhang LL, Lu XQ, Chen M, Liu Y. Long noncoding RNA GAS5 enhanced by curcumin relieves poststroke depression by targeting miR-10b/BDNF in rats. J Biol Regul Homeost Agents. 2020;34(59):815–823. doi:10.23812/20-113-a-25
61. Zou T, Sugimoto K, Zhang J, et al. Geniposide Alleviates Oxidative Stress of Mice With Depression-Like Behaviors by Upregulating Six3os1. Front Cell Dev Biol. 2020;8(60):553728. doi:10.3389/fcell.2020.553728
62. Sun Y, Cheng L, Zeng X, et al. The intervention of unique plant polysaccharides - Dietary fiber on depression from the gut-brain axis. Int J Biol Macromol. 2021;170(61):336–342. doi:10.1016/j.ijbiomac.2020.12.164
63. Zheng Y, He J, Guo L, et al. Transcriptome Analysis on Maternal Separation Rats With Depression-Related Manifestations Ameliorated by Electroacupuncture. Front Neurosci. 2019;13(62):314. doi:10.3389/fnins.2019.00314
64. Zhang Y, Ding N, Hao X, et al. Manual acupuncture benignly regulates blood-brain barrier disruption and reduces lipopolysaccharide loading and systemic inflammation, possibly by adjusting the gut microbiota. Front Aging Neurosci. 2022;14(63):1018371. doi:10.3389/fnagi.2022.1018371
65. Westfall S, Caracci F, Zhao D, et al. Microbiota metabolites modulate the T helper 17 to regulatory T cell (Th17/Treg) imbalance promoting resilience to stress-induced anxiety- and depressive-like behaviors. Brain Behav Immun. 2021;91(64):350–368. doi:10.1016/j.bbi.2020.10.013
66. Li P, Huang W, Yan YN, et al. Acupuncture Can Play an Antidepressant Role by Regulating the Intestinal Microbes and Neurotransmitters in a Rat Model of Depression. Med Sci Monit. 2021;27(65):e929027. doi:10.12659/msm.929027
67. Thoma MV, La Marca R, Brönnimann R, Finkel L, Ehlert U, Nater UM. The effect of music on the human stress response. PLoS One. 2013;8(66):e70156. doi:10.1371/journal.pone.0070156
68. Luo L, Li C, Du X, et al. Effect of aerobic exercise on BDNF/proBDNF expression in the ischemic hippocampus and depression recovery of rats after stroke. Behav Brain Res. 2019;362(67):323–331. doi:10.1016/j.bbr.2018.11.037
69. Zhan Y, Yang YT, You HM, et al. Plasma-based proteomics reveals lipid metabolic and immunoregulatory dysregulation in post-stroke depression. Eur Psychiatry. 2014;29(68):307–315. doi:10.1016/j.eurpsy.2014.03.004
70. Lazaridou A, Astrakas L, Mintzopoulos D, et al. fMRI as a molecular imaging procedure for the functional reorganization of motor systems in chronic stroke. Mol Med Rep. 2013;8(69):775–779. doi:10.3892/mmr.2013.1603
71. Yanyu S, Ying L, Kexin L, Jin W. Non-invasive brain stimulation for treating post-stroke depression: a network meta-analysis. Int J Geriatr Psychiatry. 2023;38(49):e5941. doi:10.1002/gps.5941
72. Bottomley JM, LeReun C, Diamantopoulos A, Mitchell S, Gaynes BN. Vagus nerve stimulation (VNS) therapy in patients with treatment resistant depression: a systematic review and meta-analysis. Compr Psychiatry. 2019;98(41):152156. doi:10.1016/j.comppsych.2019.152156
73. Kisely S, Li A, Warren N, Siskind D. A systematic review and meta-analysis of deep brain stimulation for depression. Depress Anxiety. 2018;35(42):468–480. doi:10.1002/da.22746
74. Wang Y, Zhang X, Wang Y, et al. Effects of Combined Live Bifidobacterium, Lactobacillus, Enterococcus and Bacillus Cereus Tablets on Post-Stroke Depression and Serum Inflammatory Factorse. Discov Med. 2023;35(33):312–320. doi:10.24976/Discov.Med.202335176.32
75. Asad A, Kirk M, Zhu S, Dong X, Gao M. Effects of Prebiotics and Probiotics on Symptoms of Depression and Anxiety in Clinically Diagnosed Samples: systematic Review and Meta-analysis of Randomized Controlled Trials. Nutr Rev. 2025;83(37):e1504–e1520. doi:10.1093/nutrit/nuae177
76. Sun Y, Liang Y, Jiao Y, et al. Comparative efficacy and acceptability of antidepressant treatment in poststroke depression: a multiple-treatments meta-analysis. BMJ Open. 2017;7(19):e016499. doi:10.1136/bmjopen-2017-016499
77. Giménez-Palomo A, Chamdal AK, Gottlieb N, et al. Efficacy and tolerability of monoamine oxidase inhibitors for the treatment of depressive episodes in mood disorders: a systematic review and network meta-analysis. Acta Psychiatr Scand. 2024;150(45MAOI):500–515. doi:10.1111/acps.13728
78. Cui C, Li J, Chen W. Statins and the incidence of post-stroke depression: a systematic review and meta-analysis. Front Neurol. 2024;15(12):1486367. doi:10.3389/fneur.2024.1486367
79. Zhang J, Zhao Y, Li H, Yang Y, Tang Q. Effectiveness of acupuncture plus music therapy for post-stroke depression: systematic review and meta-analysis. Medicine. 2024;103(46):e39681. doi:10.1097/md.0000000000039681
80. Zhen K, Shi H, Zhang X, et al. Efficacy and safety of Buyang Huanwu Decoction in the treatment of post-stroke depression: a systematic review and meta-analysis of 15 randomized controlled trials. Front Neurol. 2022;13(10):981476. doi:10.3389/fneur.2022.981476
81. Zhang J, Ming S, Chen X, et al. Herbal medicine as adjunctive therapy with antidepressants for post-stroke depression: a systematic review and network meta-analysis of randomized controlled trials. Front Pharmacol. 2023;14(11):1180071. doi:10.3389/fphar.2023.1180071
82. Chen R, Guo Y, Kuang Y, Zhang Q. Effects of home-based exercise interventions on post-stroke depression: a systematic review and network meta-analysis. Int J Nurs Stud. 2024;152(8):104698. doi:10.1016/j.ijnurstu.2024.104698
83. Zhang Y, Li G, Zheng W, et al. Effects of Exercise on Post-Stroke Depression: a Systematic Review and Meta-Analysis of Randomized Controlled Trials. Life. 2025;15(14):1. doi:10.3390/life15020285
84. Dayuan Z, Lan L, Hui C, Huanjie L, Deliang L, Yihui D. The effect of music as an intervention for post-stroke depression: a systematic review and meta-analysis. Complement Ther Med. 2022;71(47):102901. doi:10.1016/j.ctim.2022.102901
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.