Back to Journals » International Journal of Nanomedicine » Volume 20
Recent Advances in Plant-Derived Extracellular Vesicles as Nanoparticles for Cancer Drug Delivery
Authors Zhang Q, Duan H, Yang Y, Yu H, Wang W, Li B
Received 8 August 2025
Accepted for publication 26 November 2025
Published 12 December 2025 Volume 2025:20 Pages 14977—15016
DOI https://doi.org/10.2147/IJN.S559440
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Eng San Thian
Qiongdan Zhang, Huihong Duan, Yupei Yang, Huanghe Yu, Wei Wang, Bin Li
TCM and Ethnomedicine Innovation & Development International Laboratory, School of Pharmacy, Hunan·University of Chinese Medicine, Changsha, People’s Republic of China
Correspondence: Bin Li, Email [email protected] Wei Wang, Email [email protected]
Abstract: Plant-derived extracellular vesicles (PDEVs) have emerged as a highly promising and disruptive class of natural nanoparticles for anticancer drug delivery. This review provides a comprehensive analysis of PDEVs, positioning them within the broader landscape of nanomedicine through a direct comparison with conventional synthetic nanoparticles (eg, liposomes) and mammalian cell-derived extracellular vesicles (EVs). We highlight how the unique origin of PDEVs confers significant advantages, including superior natural biocompatibility, low immunogenicity, and the remarkable “dual-functionality” of acting as both inherent therapeutic agents and efficient drug carriers. The capacity of PDEVs to efficiently encapsulate a diverse range of therapeutic agents-from chemotherapeutic drugs and RNA interference molecules to gene-editing tools-is discussed in contrast to the more limited loading versatility and complex manufacturing of some alternative systems. The review systematically covers recent advances in PDEV isolation, characterization, and drug-loading techniques, emphasizing their demonstrated ability to cross biological barriers for targeted therapy and controlled release. Finally, we critically address the translational pathway, outlining key challenges in standardization and clinical translation, while forecasting their pivotal role in advancing personalized cancer nanomedicine. Through this comparative and functional perspective, PDEVs are poised to transition from a promising biological curiosity to a cornerstone of next-generation anticancer strategies.
Keywords: plant-derived extracellular vesicles, PDEVs, isolation, drug delivery, tumor therapy
Introduction
Cancer remains one of the leading causes of death worldwide, representing a major global health concern.1–3 Although significant progress has been made in treatment methods such as surgery, radiation, and chemotherapy, these therapies often come with serious side effects, diminished effectiveness over time, and the emergence of drug resistance.4–7 A key drawback of conventional cancer treatments is their lack of specificity in targeting tumor cells, which leads to collateral damage to healthy tissues. This challenge underscores the pressing need for more targeted and efficient drug delivery systems that can enhance therapeutic efficacy while reducing harmful side effects.
In recent years, nanomedicine has gained significant attention as a promising solution to a range of medical challenges, particularly through the development of advanced drug delivery systems (DDS). Nanoparticles, with their unique physicochemical properties, offer the potential to improve targeted drug delivery efficiency, allowing for precise control over drug release while reducing toxicity to healthy tissues.8–12 Among the various types of nanocarriers, extracellular vesicles (EVs)-naturally occurring nanoscale membrane-bound vesicles secreted by cells-have attracted considerable interest due to their biocompatibility, low immunogenicity, and ability to carry a diverse range of bioactive molecule, including proteins, lipids, nucleic acids, and small molecules.13–19 This field has expanded beyond the well-studied mammalian EVs to include vesicles derived from plants and even microorganisms, each offering a unique set of biological functions and therapeutic potentials. Moreover, EVs possess the exceptional ability to cross biological barriers, such as the blood-brain barrier, making them highly suitable for drug delivery applications.20–25
It is important to note that the biological kingdom of the source organism fundamentally shapes the properties and applications of its EVs. For instance, extracellular vesicles from microbial sources, including bacteria (outer membrane vesicles) and fungi, are gaining attention for their roles in pathogenesis and immune modulation, with emerging applications in vaccine development.26 However, their translation into systemic drug delivery can be complicated by potential safety concerns, such as the presence of endotoxins in gram-negative bacteria, which can trigger strong inflammatory responses.
In this evolving landscape, while mammalian cell-derived extracellular vesicles (EVs) have been extensively explored as drug delivery systems, plant-derived extracellular vesicles (PDEVs) are emerging as a promising alternative with several unique advantages. Crucially, the therapeutic potential of PDEVs extends far beyond their role as inert nanocarriers, constituting a fundamental distinction from most mammalian EVs. Whereas mammalian EVs often function primarily as signaling vehicles or drug delivery platforms, PDEVs are inherently bioactive therapeutics by design, owing to their plant origin. This intrinsic bioactivity stems from their unique cargo, which is enriched with plant-specific secondary metabolites-such as polyphenols, flavonoids, terpenoids, and lipids. These molecules are meticulously packaged into PDEVs, resulting in a synergistic, multi-targeted therapeutic entity. For instance, ginger-derived PDEVs, rich in gingerols and shogaols, not only facilitate cellular uptake but also directly induce apoptosis in cancer cells and ameliorate colitis by activating the Nrf2 pathway. Similarly, grapefruit-derived PDEVs exert inherent anti-inflammatory effects, while those from bitter melon exhibit specific pro-apoptotic activity against liver cancer. This dual functionality-where the vesicle acts simultaneously as both the delivery system and the active drug-is a paradigm shift from conventional nanomedicine. It eliminates the need for complex drug loading in certain applications and allows PDEVs to exert pleiotropic effects, including antioxidant, anti-inflammatory, and immunomodulatory activities, which are difficult to replicate with synthetic nanoparticles or mammalian EVs. Therefore, recognizing PDEVs as bioactive substances in their own right unveils their vast potential for preventative medicine, combination therapies, and as stand-alone natural product-based therapeutics.
Beyond their inherent bioactivity, the distinctive properties of PDEVs-including their abundance in bioactive compounds, low immunogenicity, and superior biodegradability compared to mammalian EVs-position them as highly promising candidates for the targeted and controlled release of anticancer drugs.27–34 This natural, non-toxic platform is exceptionally capable of delivering a diverse range of therapeutic cargo. Recent studies have demonstrated that PDEVs can successfully carry chemotherapeutic drugs, siRNAs, and gene-editing tools, while specifically targeting cancer cells.35–43 Furthermore, the ability to engineer PDEVs to improve their targeting accuracy, stability, and cargo capacity further strengthens their potential as advanced drug delivery vehicles.44–49 Therefore, recognizing PDEVs as bioactive substances in their own right, complemented by their advanced delivery capabilities, unveils their vast potential for preventative medicine, combination therapies, and as stand-alone natural product-based therapeutics.
This review seeks to offer a comprehensive analysis of recent advancements in the development and application of PDEVs as nanoparticles for cancer drug delivery. Specifically, we will cover: (1) the methods of isolating PDEVs for use in drug delivery; (2) their capacity for encapsulating and transporting a broad range of therapeutic agents; (3) the application of PDEVs in targeted drug delivery to specific cancer cells or tumor microenvironments. Furthermore, we will explore the future potential of plant-derived EVs in cancer therapy, considering both the current state of research and the exciting prospects ahead. As nanomedicine evolves, PDEVs offer a promising biologically derived alternative to synthetic nanoparticles, paving the way for the development of more effective and personalized cancer treatments.
Composition of Plant-Derived Extracellular Vesicles
Plant-derived extracellular vesicles (PDEVs) are intricate nanoscale structures that encapsulate a diverse array of bioactive molecules, each of which plays a pivotal role in mediating biological functions. These vesicles carry various types of nucleic acids, including RNA (such as microRNAs, small interfering RNAs, and messenger RNAs) and DNA, which are involved in gene regulation and intercellular communication. In addition to nucleic acids, PDEVs are enriched with a range of proteins that participate in cellular processes such as signaling, stress response, defense mechanisms, and cellular transport. The lipid content of PDEVs, which includes various phospholipids and glycosphingolipids, is essential for vesicle membrane integrity and the facilitation of molecular interactions. Moreover, PDEVs are known to harbor a range of metabolites, including polyphenols, flavonoids, carotenoids, and vitamins, all of which have antioxidant, anti-inflammatory, and protective properties. Together, these components enable PDEVs to mediate complex biological processes, such as stress tolerance, immune modulation, and cell-to-cell communication, both within plants and between plants and other organisms. The diverse composition of PDEVs highlights their potential to serve as vehicles for transferring molecular signals and bioactive substances, opening up possibilities for their use in therapeutic and biotechnological applications.
Nucleic Acids Content of PDEVs
Nucleic acids, key components of PDEVs, have attracted significant interest due to their potential as biomarkers and their functional roles. Techniques such as low-input RNA sequencing (RNA-Seq) and quantitative PCR (qPCR) are frequently used to detect specific nucleic acid sequences within PDEVs.50 In particular, RNA plays an essential role in regulating various biological processes in plants and may influence molecular interactions between different species.
Small RNAs (sRNAs) in plants, including microRNAs (miRNAs) and small interfering RNAs (siRNAs), are involved in gene silencing. These sRNAs can travel between plant cells, from roots to shoots, and even between plants and pathogens.50 Research has revealed that the ten most common miRNA families are present in a range of edible plants, and these miRNAs are particularly enriched in exosomes. Small RNA sequencing has shown that miRNAs, typically 20–22 nucleotides long, can regulate human mRNA.51 This suggests that sRNAs in PDEVs may facilitate long-distance communication within plants and could serve as mediators for cross-species interactions.52 Computational tools such as Target Scan predict that miRNAs from PDEVs may target mammalian genes involved in inflammation and cancer pathways. New evidence also highlights the immunomodulatory and anticancer properties of plant-derived miRNAs.52
While research on RNA within PDEVs has been the primary focus, recent studies are starting to investigate the role of DNA in these vesicles. Although DNA in PDEVs has been less extensively studied, there is growing evidence suggesting its potential for disease biomarker discovery and intercellular communication.53,54
Protein Content of PDEVs
Compared with the protein composition of animal-derived vesicles, PDEVs typically contain a narrower range of proteins, and these proteins are present at lower levels.55 Research has identified several key protein types within plant-derived vesicles, including those involved in glycolipid metabolism, guanosine triphosphatase (GTPase) activity, membrane structure organization, and vesicle formation.55 These proteins are crucial for maintaining the functionality of cellular membranes and play a significant role in cellular processes related to transport and signaling.
A detailed comparison between the protein species found in lemon-derived vesicles and those found in animal exosomes revealed a striking 56.7% overlap.56 This overlap suggests that a significant proportion of the proteins in plant exosome-like vesicles may share similarities with those involved in the formation and function of animal exosomes. This finding provides further support for the idea that certain proteins are conserved across species and may play a fundamental role in the biogenesis and functionality of exosome-like vesicles, regardless of their plant or animal origin. Moreover, the identified proteins in plant-derived exosome-like vesicles are likely to be involved in key cellular processes such as cytoskeleton organization, membrane transport, and vesicle trafficking.56 These processes are essential for maintaining cellular structure and facilitating the movement of molecules across membranes. Additionally, the proteins associated with these vesicles likely contribute to a variety of physiological functions, including metabolic regulation, cell communication, and intercellular signaling.57 Given the overlap between plant and animal exosome proteins, it is likely that these vesicles participate in similar biological pathways, including the transport of genetic material, lipids, and signaling molecules that influence cellular behavior and response to external stimuli.
In conclusion, while plant exosome-like vesicles differ from their animal counterparts in terms of protein composition, the proteins they carry play vital roles in physiological processes. Their involvement in membrane dynamics, signal transduction, and metabolic regulation underscores the importance of these vesicles in plant biology and suggests that they may share functional similarities with exosomes in animals.
Lipid Content of PDEVs
Lipids are fundamental components of the phospholipid bilayer that forms the structural basis of plant vesicles, and they are essential constituents of vesicles derived from both plants and animals. These lipid molecules play critical roles in maintaining the integrity of the vesicle membrane, facilitating vesicle trafficking, and participating in various cellular signaling processes. Numerous studies have highlighted the significant presence of several key lipids in plant-derived vesicles, including phosphatidic acid (PA), phosphatidylethanolamine (PE), phosphatidylinositol (PI), diacylglycerol (DAG), triacylglycerol (TAG), galactosyl diacylglycerol (DGDG), and mono-galactosyl diacylglycerol (MGDG).58
Among the lipids found in plant vesicles, phosphatidic acid (PA) is the most abundant. PA plays an integral role in cellular processes such as cell proliferation, membrane remodeling, and signal transduction. It is particularly important in regulating various aspects of membrane dynamics, including vesicle formation and fusion. Following PA, phosphatidylethanolamine (PE) is the second most prevalent lipid in plant vesicles. PE is essential for regulating membrane curvature and maintaining membrane fluidity, making it a key player in processes such as membrane fission, fusion, and endocytosis. In addition to PA and PE, other lipids, such as phosphatidylinositol (PI), diacylglycerol (DAG), and triacylglycerol (TAG), are also found in significant quantities within plant-derived vesicles. PI is involved in a wide range of cellular signaling pathways, particularly those related to inositol phosphate metabolism, which plays a pivotal role in regulating cell growth, differentiation, and stress responses. DAG and TAG are important lipid intermediates in the biosynthesis of various membrane lipids and energy storage molecules, contributing to the overall energy balance and lipid metabolism within cells. Furthermore, galactosyl diacylglycerol (DGDG) and mono-galactosyl diacylglycerol (MGDG) are specialized galactolipids that are predominantly found in the membranes of plant cells, especially in the chloroplasts. These lipids are crucial for maintaining the integrity and functionality of the thylakoid membrane, which plays a vital role in photosynthesis. While DGDG and MGDG are more commonly associated with plant cells, they have also been detected in plant-derived vesicles, indicating their importance in vesicle function and intercellular communication.59
Interestingly, the specific types and relative content of lipids in vesicles vary depending on the plant source. Different fruiting plants, such as citrus fruits, tomatoes, and grapes, exhibit unique lipid profiles in their vesicles.60 This variation can be attributed to factors such as the plant’s metabolic requirements, environmental conditions, and the specific roles these vesicles play in the plant’s physiology. For example, citrus-derived vesicles might be rich in certain lipids that are involved in stress responses, while vesicles from other plants may contain higher levels of lipids associated with nutrient storage or cell signaling.61
Overall, the lipid composition of plant-derived vesicles is both diverse and complex, with different lipid species playing specialized roles in membrane structure, signaling, and metabolic regulation. This diversity underscores the functional importance of lipids in plant vesicles and highlights the potential for exploiting plant-derived vesicles in biotechnological applications, such as drug delivery, intercellular communication, and environmental stress management. Furthermore, the study of lipid profiles in plant vesicles offers valuable insights into the mechanisms that govern plant cell function, growth, and response to environmental stimuli.
Other Content of PDEVs
In addition to lipids and proteins, PDEVs contain a variety of bioactive metabolites, including polyphenols, flavonoids, and other secondary metabolites,62 which contribute significantly to their therapeutic properties. These metabolites have been shown to enhance the inherent bioactivity of PDEVs, particularly in their antioxidant, anti-inflammatory, and anticancer effects. The presence of polyphenols, such as flavonoids, tannins, and phenolic acids, plays a critical role in the bioactivity of PDEVs, influencing cellular signaling pathways, modulating oxidative stress, and promoting apoptosis in cancer cells.
Polyphenols and flavonoids within PDEVs can act as natural antioxidants, neutralizing free radicals and reducing oxidative damage. This not only protects healthy cells from damage but also sensitizes cancer cells to therapeutic agents. Furthermore, these metabolites can modulate key inflammatory pathways, such as NF-κB and MAPK signaling, which are involved in tumor progression. The ability of PDEVs to deliver these bioactive compounds to target cells enhances their anticancer efficacy by disrupting the tumor microenvironment and reducing tumor growth and metastasis.
The metabolite profiles of PDEVs vary depending on the plant source, and this variability can influence the vesicles’ therapeutic potential. For example, ginger-derived vesicles may contain higher levels of specific polyphenols such as gingerols and shogaols, which have potent anticancer properties, while cabbage-derived vesicles may be rich in glucosinolates and their breakdown products, which are known for their chemoprotective effects. Understanding these variations can help optimize the selection of plant sources for PDEVs based on the specific metabolites that may offer synergistic benefits in cancer therapy.
These metabolites also contribute to the vesicles’ ability to interact with specific cancer cell receptors, modulating cellular processes like uptake, membrane fusion, and intracellular signaling. For instance, flavonoids found in PDEVs may improve their binding affinity to cancer cell receptors, enhancing the targeting and internalization of therapeutic agents.
In conclusion, the metabolites contained in PDEVs, particularly polyphenols and flavonoids, are integral to their bioactivity and therapeutic potential. Variations in the metabolite profiles of PDEVs from different plant sources can have significant implications for their effectiveness in cancer treatment, highlighting the importance of selecting the appropriate plant sources for targeted drug delivery systems.
Content and the Dual-Function Potential of PDEVs
The true promise of PDEVs lies in their potential to function in a dual capacity: as both drug delivery vehicles and providers of inherent therapeutic benefits through their natural cargo. For example, PDEVs can be engineered to not only deliver conventional chemotherapeutic drugs directly to cancer cells but also to enhance treatment outcomes by supplying additional bioactive compounds, such as anti-inflammatory flavonoids, from their natural cargo. This dual functionality could provide multiple layers of therapeutic effects, particularly in cancer treatment.
Dual-function PDEVs could have a profound impact on therapeutic strategies. As these vesicles deliver chemotherapeutic agents directly to tumor sites, the flavonoids and polyphenols naturally contained within the vesicles could simultaneously reduce inflammation, mitigate oxidative stress, and even enhance the efficacy of the drugs being delivered. For instance, flavonoids such as quercetin and kaempferol, known for their potent anti-inflammatory and antioxidative properties, can synergize with chemotherapy drugs to create a more effective, less toxic treatment regimen. In practice, dual-function PDEVs could be used to target tumor cells while also reducing the collateral damage to healthy tissues that is often seen in conventional cancer therapies. By incorporating both drug-delivery capabilities and natural bioactive compounds in a single vesicle, PDEVs can improve the targeting and uptake of drugs while providing a simultaneous therapeutic benefit through their inherent anti-inflammatory, antioxidant, and anticancer properties.
The metabolite profiles of PDEVs vary depending on the plant source, which can influence the selection of dual-function vesicles. For example, ginger-derived PDEVs, rich in gingerols and shogaols, could deliver anticancer drugs to the tumor site while also providing additional therapeutic effects due to the anti-inflammatory properties of these bioactive compounds. Similarly, cabbage-derived vesicles, which are rich in glucosinolates, could act as both delivery vehicles for chemotherapeutic agents and as an additional layer of protection through their chemoprotective effects. Understanding the variations in these metabolite profiles and how they can work synergistically with chemotherapeutic agents is crucial for the development of PDEVs as dual-function systems. By carefully selecting plant sources based on their bioactive compound content, it is possible to optimize the therapeutic benefits of PDEVs, tailoring them to provide both efficient drug delivery and enhanced therapeutic effects in cancer treatment.
In conclusion, the metabolites contained in PDEVs, particularly polyphenols and flavonoids, are integral to their bioactivity and therapeutic potential. When designed to function in dual roles, PDEVs can deliver chemotherapeutic drugs while simultaneously providing inherent therapeutic benefits from their natural cargo, such as reducing inflammation and oxidative stress. Variations in the metabolite profiles of PDEVs from different plant sources can have significant implications for their effectiveness in cancer treatment, highlighting the importance of selecting the appropriate plant sources for targeted drug delivery systems that offer both enhanced treatment efficacy and reduced side effects.
Isolation of Plant-Derived Extracellular Vesicles
PDEVs are present in various plant tissues such as leaves, roots, and seeds.63,64 However, isolating and purifying PDEVs from these complex plant matrices is challenging due to the presence of numerous cellular components, including proteins, lipids, and other biomolecules.65 The complexity of plant-derived biofluids makes it difficult to accurately isolate and analyze PDEVs.66 To address these difficulties, several techniques have been developed that leverage factors like size, density, and surface markers for the extraction and purification of PDEVs from plant materials. Commonly used methods include ultracentrifugation, density gradient centrifugation, and ultrafiltration centrifugation. Additionally, specialized kits and reagents that utilize techniques such as size-exclusion chromatography, immunoaffinity capture, or precipitation have been designed to selectively isolate PDEVs while minimizing contamination from other plant components. Table 1 provides a detailed overview of various methods used for the isolation of plant-derived extracellular vesicles.
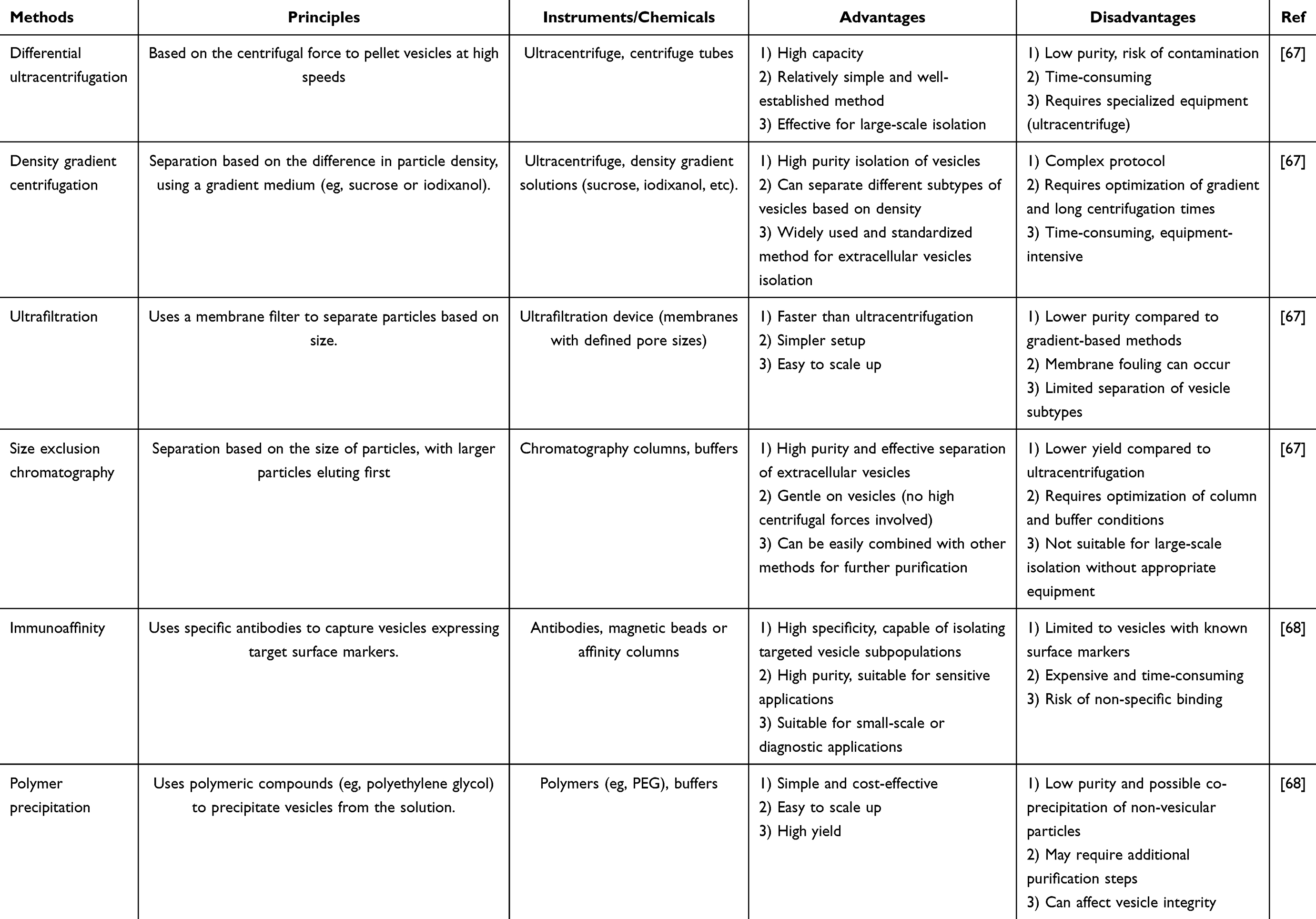 |
Table 1 An Overview on Different Isolation Methods for PDEVs |
Ultracentrifugation
Ultracentrifugation is a technique used to separate particles according to their size and density. It can be classified into two primary types: differential ultracentrifugation and density gradient ultracentrifugation. Both methods are widely applied for isolating PDEVs due to their simplicity and cost-effectiveness, and they are considered gold-standard techniques.69,70
Differential ultracentrifugation operates through a stepwise centrifugation process, where particles are separated according to their size and density. In this method, the sample is subjected to increasing centrifugal forces over several rounds, progressively removing larger and heavier particles like cells, cellular debris, apoptotic bodies, and microvesicles (Figure 1A).67 The initial low-speed centrifugation steps remove the largest components, while higher-speed steps isolate smaller vesicles such as PDEVs. This approach is particularly attractive due to its simplicity and ability to generate relatively quick results. However, one of its major limitations is the potential for contaminant co-sedimentation, especially with protein aggregates and other small particles that may share similar densities with PDEVs. Such contaminants can end up in the final vesicle preparation, compromising its purity and potentially affecting downstream applications. Despite this, differential ultracentrifugation remains a popular choice due to its ease of implementation and the reasonable purity it offers for many applications.
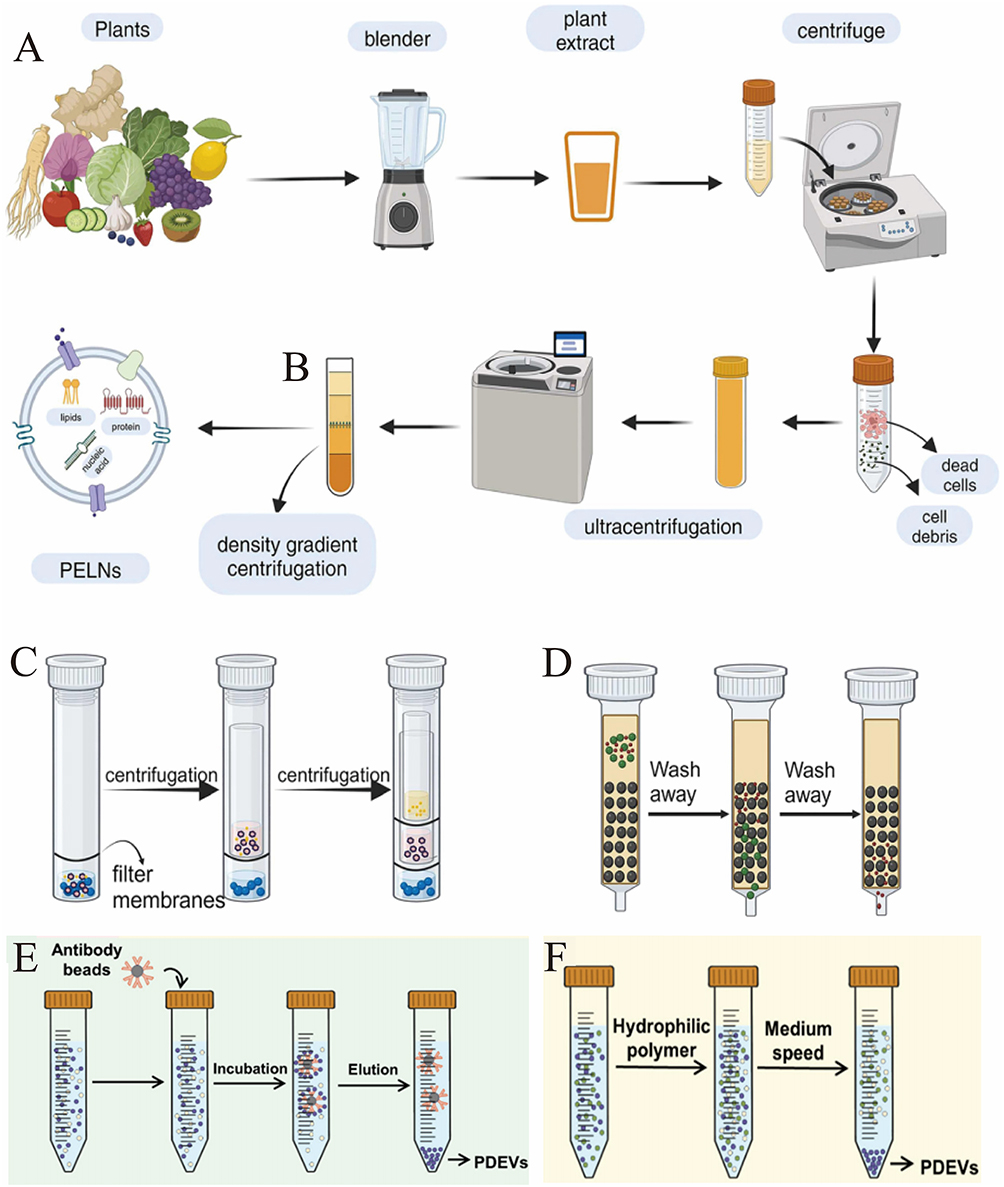 |
Figure 1 Extraction method of plant-derived extracellular vesicles (PDEVs). (A) Differential ultracentrifugation. (B) Density gradient centrifugation. (C) Ultrafiltration. (D) Size exclusion chromatography method. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 76 Copyright 2024 The Authors). (E) Immunoaffinity. (F) Polymer precipitation. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 82 Copyright 2024 The Authors). |
In contrast to differential ultracentrifugation, density gradient ultracentrifugation utilizes a gradient medium, often composed of sucrose or iodixanol, which is created by layering solutions of different densities. When the sample is centrifuged, particles within the sample migrate through the gradient and settle at the position where their density matches the surrounding medium. This method allows for much finer separation, as particles with even slight differences in density can be resolved into distinct layers, leading to the isolation of highly purified fractions (Figure 1B).67 The key advantage of density gradient ultracentrifugation is its ability to produce high-purity PDEVs, making it ideal for studies that require the vesicles to be free of contaminants that might interfere with downstream functional assays or molecular analyses.
However, despite its advantages in terms of purity, density gradient ultracentrifugation does come with its own set of challenges. One significant drawback is the relatively low yield of PDEVs compared to differential ultracentrifugation. The process involves multiple layers and longer centrifugation times, which can lead to a reduction in the quantity of vesicles isolated, making this method less ideal when large quantities are required. Moreover, the preparation of the density gradient medium itself can be time-consuming and technically demanding. As a result, while density gradient ultracentrifugation is preferred for the enrichment of PDEVs, it is often balanced by the trade-off of lower yields, making it more suitable for applications where purity is prioritized over quantity.71
Overall, both differential and density gradient ultracentrifugation are widely used for isolating plant-derived extracellular vesicles, each with its own set of advantages and limitations. Differential ultracentrifugation offers a simpler and faster method with a reasonable yield, though it may come with contamination risks. In contrast, density gradient ultracentrifugation excels in providing higher purity, though it often results in a lower vesicle yield. The choice between these two methods largely depends on the specific requirements of the study or application, such as the need for purity versus quantity of PDEVs.
Ultrafiltration Centrifugation
Ultrafiltration centrifugation is an effective technique that utilizes the unique properties of ultrafiltration membranes with a microporous structure to separate and isolate various components of a sample based on their size under centrifugal force (Figure 1C).67 In this process, the sample is passed through the ultrafiltration membrane, which serves as a selective barrier. It allows only particles smaller than the membrane’s pore size to pass through, while larger particles, such as PDEVs, are retained on the membrane surface. This results in a more concentrated and cleaner preparation of PDEVs, which is crucial for downstream applications like biological studies or therapeutic use.72
Ultrafiltration centrifugation is effective in preserving the biological activity of PDEVs, which is vital for their functional integrity in later experiments.73 However, although this technique is highly efficient in isolating and purifying the vesicles, the mechanical forces exerted during centrifugation may unintentionally alter their morphology. These morphological changes can be particularly significant when the structural integrity of the vesicles is crucial for specific applications. Therefore, while ultrafiltration centrifugation is a powerful method for purifying PDEVs, it is important to consider the potential impact on their physical structure during the procedure.
Size Exclusion Chromatography
Size-exclusion chromatography (SEC) is a powerful and widely employed technique for isolating and purifying PDEVs, leveraging their size and hydrodynamic properties (Figure 1D).67 The method works by using a chromatography column packed with a porous gel matrix, which acts as the stationary phase. Upon introducing a sample into the column, larger particles, like PDEVs, are excluded from entering the smaller pores of the gel, causing them to move through the column more rapidly and elute in earlier fractions. In contrast, smaller molecules-such as free proteins, nucleic acids, and other soluble contaminants-are able to enter the gel pores. These smaller particles experience a slower passage through the matrix, resulting in delayed elution. This size-dependent movement is key to SEC’s ability to separate EVs from smaller impurities. Consequently, PDEVs are eluted in the earlier fractions, while the smaller contaminants are retained in the gel matrix, ensuring their efficient separation and removal.
The selective separation mechanism of SEC efficiently isolates PDEVs, ensuring they are collected in a fraction with minimal contamination from smaller soluble components like free proteins, nucleic acids, and other molecular by-products.74 This is especially advantageous for applications that demand high purity of the EV fraction, such as functional studies or proteomics, where contamination by non-vesicular proteins could disrupt subsequent analyses.75 SEC provides a reliable, scalable, and highly effective approach for purifying PDEVs with minimal contamination, making it an indispensable tool in the study of PDEVs.
Immunoaffinity
Immunoaffinity capture (IA) methods are highly effective for isolating specific subpopulations of PDEVs by targeting the unique surface markers that define these vesicles. This technique relies on antibodies that bind to specific proteins, such as CD9, CD63, CD81, or plant-derived markers like TET8, which are commonly expressed on the surface of EVs (Figure 1E).68 The antibody-mediated recognition of these markers allows for the precise isolation of EV subsets, offering a level of specificity that surpasses other isolation techniques.
The strength of immunoaffinity capture (IA) lies in its unmatched ability to selectively isolate EVs with particular surface markers, making it an indispensable tool for investigating specific EV subpopulations and analyzing their molecular cargo.76 This high specificity is especially useful when exploring the functional roles of different EV types, as well as examining the diverse molecular content they carry, which can differ significantly between subpopulations. By concentrating on distinct surface markers, IA facilitates the enrichment of targeted EV populations, enabling more in-depth and focused analyses, including proteomic, transcriptomic, or lipidomic profiling.77
However, despite its high specificity, IA often results in lower yields compared to other physical separation methods, such as ultracentrifugation (UC), due to the inherently selective nature of antibody binding. This selectivity means that only the EVs bearing the targeted markers are captured, which can limit the overall quantity of isolated vesicles. Consequently, while IA is ideal for studies requiring precise characterization of specific EV subtypes, it is often combined with other separation techniques to enhance the yield. For instance, IA can be used in conjunction with methods like UC or size-exclusion chromatography (SEC) when the goal is to isolate a broad range of EVs while still maintaining specificity for particular subpopulations.78
Collectively, immunoaffinity capture is a highly specific and effective technique for isolating EV subpopulations, particularly when detailed molecular analyses are required. Nevertheless, its relatively low yield necessitates its use in combination with other isolation methods, ensuring both specificity and sufficient sample quantity for comprehensive investigations.
Polymer Precipitation
Polymer-based precipitation (PBP) is a widely adopted and well-established technique for isolating extracellular vesicles (EVs), particularly in laboratory environments. This method utilizes volume-excluding polymers, such as polyethylene glycol (PEG), to precipitate particles based on differences in solubility (Figure 1F).68 During the process, the polymer forms a network that selectively traps PDEVs, while allowing smaller, more soluble molecules to remain in the solution. The precipitated PDEVs are then collected by low-speed centrifugation, which facilitates the separation of the larger EVs from smaller, non-precipitated components. This principle is frequently utilized in commercial kits for EV isolation, making PBP an easily accessible and straightforward method for researchers.
PBP offers several advantages, including speed and simplicity, which make it particularly suitable for routine use in laboratory settings when quick isolation of PDEVs is needed. It is particularly effective for preparing PDEV samples in research environments where large volumes of biological samples must be processed in a time-efficient manner. Given its practicality, PBP is often used for isolating PDEVs in studies that require a relatively straightforward method without extensive instrumentation.79
However, one notable limitation of PBP is the risk of co-precipitating non-EV components, such as proteins, lipids, and other biomolecules that share similar solubility properties. These contaminating substances can lead to unwanted interference in downstream analyses, such as proteomic studies, lipidomics, or molecular profiling. For example, the presence of co-precipitated proteins may obscure the identification of EV-specific markers or complicate the interpretation of functional assays.80 As a result, while PBP is highly convenient and efficient for initial isolation, it may not always produce highly purified EV preparations.
To mitigate this issue and ensure a high level of purity for more sensitive analyses, additional purification steps-such as density-gradient centrifugation, size-exclusion chromatography (SEC), or immunoaffinity-based methods-are often required. While these techniques are universally applied in EV isolation from all biological sources, their application in PDEV research is uniquely shaped by the plant matrix. For instance, density-gradient centrifugation is critical for separating PDEVs from co-precipitating plant proteins and nucleic acids, while SEC must be optimized to avoid column fouling by abundant plant polysaccharides. These secondary methods are therefore indispensable for removing plant-specific contaminants and enabling precise downstream experiments. Therefore, while PBP remains a valuable and practical tool for initial EV isolation, researchers must carefully consider its limitations and potentially combine it with other purification techniques based on the specific needs of their studies.
In summary, selecting the most appropriate method for isolating PDEVs from complex plant matrices requires a dual consideration: the general principles of EV isolation and the specific challenges posed by the plant source. This approach ensures that the chosen technique will meet the necessary criteria for subsequent analyses. Researchers must not only evaluate universal factors such as efficiency, yield, and purity but also account for PDEV-specific considerations, such as the need for effective cell wall disruption during initial homogenization and the diligent removal of non-vesicular plant compounds (eg, pigments, organic acids, and secondary metabolites) that are not encountered when working with mammalian cell cultures. A thorough evaluation of both general and plant-specific factors is paramount before making a final methodological decision.
Advantages of Plant-Derived Extracellular Vesicles as Nano-Drug Delivery Systems
PDEVs have attracted considerable interest in recent years as promising candidates for nano-drug delivery systems, owing to their unique advantages. These vesicles, which are naturally secreted by plant cells, offer a range of unique properties that make them particularly well-suited for delivering therapeutic agents in a highly efficient and targeted manner. Below are some of the primary benefits that make PDEVs an exciting area of research and application in the field of drug delivery.
Biocompatibility and Low Toxicity
An ideal drug delivery system should exhibit both outstanding biocompatibility and non-toxicity. PDEVs meet these requirements and stand out for their exceptional safety profile and low cytotoxicity, particularly when compared to synthetic nanoparticles and liposomes.
One of the key advantages of PDEVs is their low immunogenicity, which significantly enhances their biocompatibility. This reduced immune response makes PDEVs highly attractive for medical applications and drug delivery systems, as it minimizes the risk of immune rejection-an essential factor in ensuring the long-term safety and effectiveness of therapies.
The minimal toxicity of PDEVs can be attributed to several factors. These vesicles are primarily derived from edible plants, which are naturally considered safe for human consumption. In contrast to exosomes from mammalian or bacterial sources, which could carry zoonotic or human pathogens, plant-derived vesicles do not present such risks.81 This significantly reduces the chance of transferring harmful genes or proteins, further improving their safety profile. In conclusion, PDEVs represent a highly promising approach for safe, biocompatible, and effective drug delivery.
Cost-Effective, Sustainable, and Scalable Production
A major advantage of PDEVs is their ability to be derived from a wide range of edible plants, which allows for cost-effective, sustainable, and large-scale production. This stands in stark contrast to synthetic nanoparticles or mammalian exosomes,82 which often require greater resource input and more complex manufacturing processes. A brief techno-economic analysis highlights this advantage: while comprehensive life-cycle assessments for PDEVs are still emerging, innovative production platforms like the Temporary Immersion Bioreactor System (TIBS) demonstrate the potential for significant cost and efficiency gains. TIBS enables the non-destructive, continuous collection of PDEVs from cultured plant tissues, reportedly reducing energy consumption by approximately 40% compared to traditional solid plant tissue culture methods. Furthermore, the absence of requirement for expensive culture supplements like fetal bovine serum, which is essential for mammalian cell-derived exosomes, substantially lowers raw material costs. In terms of scalability, while precise figures for grams of PDEVs per kg of biomass are still under characterization, the modular design of TIBS supports scalable production and has been successfully applied to a variety of plants, including ginger and licorice. When compared to the established manufacturing costs of Good Manufacturing Practice (GMP)-grade synthetic nanoparticles like liposomes, PDEVs benefit from a potentially simpler and less expensive upstream process by leveraging the inherent efficiency of plant biomass synthesis.
PDEVs not only offer higher production yields but also shorter extraction times compared to exosomes derived from animal or human sources, positioning them as a promising biological material with significant potential for broad applications. In contrast, mammalian exosomes are constrained by lower yields and limited sources, such as milk, urine, blood, and bile.83,84 Thus, the emergence of PDEVs provides an innovative solution to address these production limitations.
Wide Range of Sources
The Food and Agriculture Organization estimates that there are over 50,000 medicinal plants globally, many of which hold significant promise as natural resources for contemporary pharmaceutical research, although the active compounds in most of these plants remain largely unexplored.85 Some PDEVs derived from medicinal plants are thought to retain the therapeutic properties of their source plants. For example, exosomes from plants like ginseng,86 ginger,87 grapes,88 and honeysuckle89 may carry bioactive compounds similar to those in the original plants, pointing to potential pharmacological effects. This offers exciting possibilities for leveraging PDVEs as drug delivery systems or carriers, potentially broadening the scope and improving the effectiveness of pharmaceutical treatments.
Application of Plant-Derived Extracellular Vesicles for Drug Delivery
Drug Loading Methods
An effective drug delivery system should not only optimize the efficiency of drug delivery but also safeguard the integrity of the drug throughout the process. This is especially crucial in the development of drug delivery systems that utilize plant-derived vesicles. Successful loading requires careful optimization of several key factors, such as improving encapsulation efficiency, preserving the structural integrity of the vesicle, and ensuring that the drug retains its biological activity during the delivery process.
Various techniques can be employed to load drugs into plant vesicles, each offering distinct advantages and challenges. These include co-incubation, transfection, sonication, extrusion, and electroporation. The selection of the appropriate method typically depends on factors such as the type of drug, the characteristics of the vesicle system, and the desired therapeutic objectives. A schematic representation of these drug-loading methods is shown in Figure 2, and a detailed comparison of each technique is provided in Table 2.
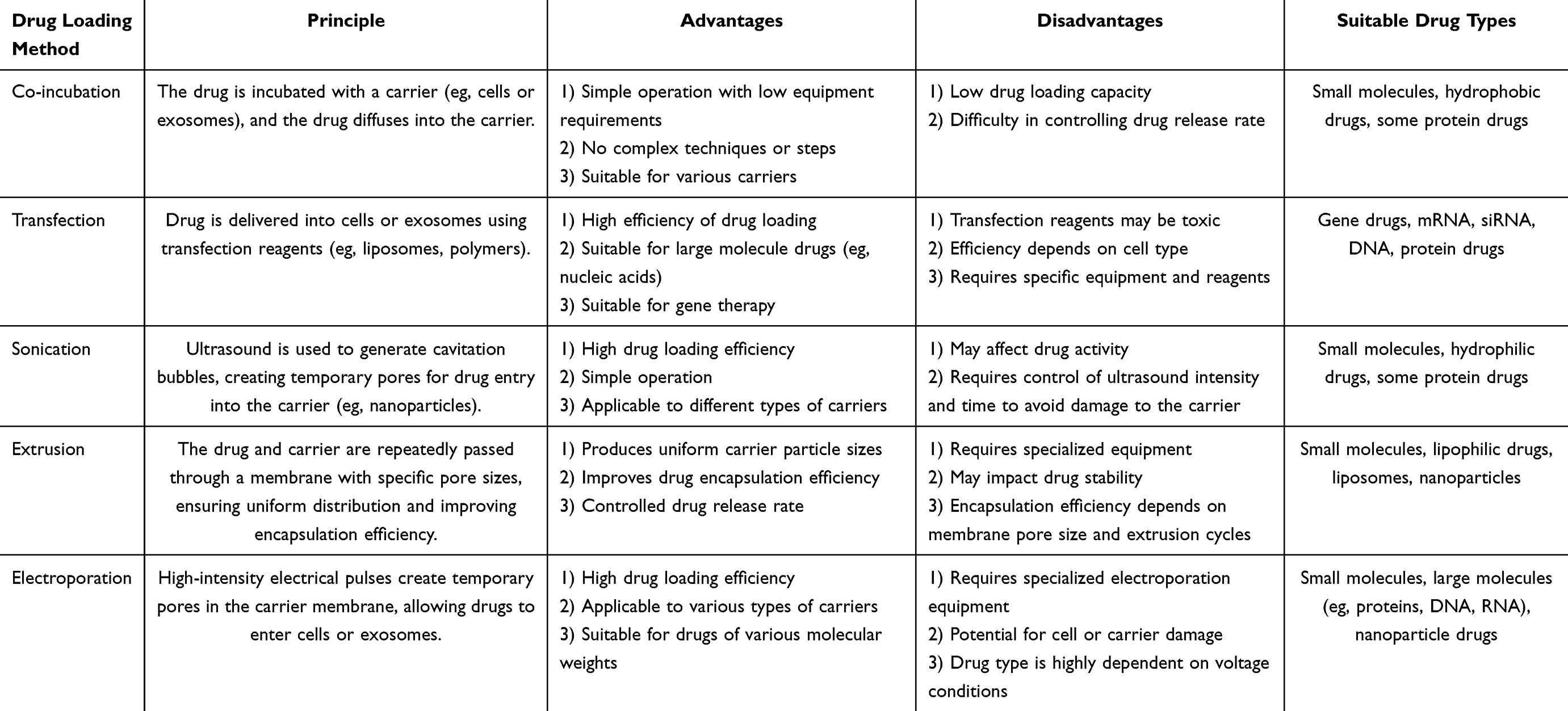 |
Table 2 Comparison of Various Plant Vesicle Drug Loading Methods |
 |
Figure 2 Schematic diagram of the drug-carrying methods of plant-derived extracellular vesicles. (A) Co-incubation method. (B) Transfection method. (C) Sonication. (D) Extrusion. (E) Electroporation. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 97 Copyright 2024 The Authors). |
Co-Incubation Method
The co-incubation method is the simplest and most commonly used approach for loading drugs into plant-derived extracellular vesicles. In this method, the drug is combined with the vesicles, allowing it to be encapsulated through mechanisms such as hydrophobic interactions, diffusion, or electrostatic forces (Figure 2A).90 The loading efficiency of this technique is mainly influenced by the hydrophobicity of the drug. Hydrophobic drugs, in particular, are more likely to interact with the lipid bilayer of the vesicle membrane, thereby enhancing the encapsulation efficiency.91
Transfection Method
Transfection is a technique used to introduce foreign substances, such as DNA, RNA, or drugs, into cells.92 In drug delivery, it facilitates the movement of drugs across the cell membrane or into extracellular vesicles like exosomes. This process typically employs chemical, physical, or biological approaches to enhance the transmembrane transport of drugs, thus boosting their cellular uptake and overall delivery efficiency (Figure 2B).90 This approach improves both the efficiency and targeting capability of drug delivery.93
Sonication
Sonication is a versatile and widely used method for preparing drug-loaded nanoparticles, exosomes, and other nanocarriers.94–96 This method utilizes high-frequency sound waves, typically ranging from 20 kHz to several megahertz, to induce mechanical vibrations in a liquid medium (Figure 2C).90 These vibrations create rapid pressure fluctuations that result in the formation of small cavitation bubbles. The collapse of these bubbles generates intense shear forces and localized heat, which effectively break down larger particles, enhance dispersion, and improve the uniform mixing of different components.
In drug delivery applications, the mechanical forces generated during sonication aid in breaking down drug particles and facilitating their uniform incorporation into nanocarriers. This process results in the creation of smaller, more uniform drug-loaded nanoparticles or liposomes, while also enhancing their stability, bioavailability, and targeting ability. Consequently, it improves the drug’s release profile and boosts its therapeutic effectiveness.
Extrusion
Extrusion is frequently combined with other techniques, such as sonication, to further enhance the properties of drug-loaded carriers. For example, in the preparation of liposomes, after the lipid bilayer is formed through methods like thin-film hydration or solvent injection, the formulation is typically extruded multiple times through a polycarbonate membrane with specific pore sizes (Figure 2D).90 This process ensures uniform liposome size and improves encapsulation efficiency. Extrusion is essential in the development of drug-loaded nanoparticles and liposomes, as it enhances drug loading efficiency, promotes consistency, and facilitates better-controlled release, all of which contribute to the stability and long-term effectiveness of the formulation.97
Electroporation
Electroporation is a method that involves applying a brief high-voltage pulse to generate hydrophilic pores in the membrane of PDEVs through an external electric field (Figure 2E).90 This increases the membrane’s permeability, facilitating the entry of substances like chemicals, DNA, RNA, or drugs into the plant-derived EVs.98 Once the drug is transported into the plant-derived EVs through the transient pores, the membrane quickly restores its integrity. This technique not only ensures effective encapsulation of the drug but also preserves the functional properties of the plant-derived EVs, positioning them as a highly promising platform for targeted drug delivery and therapeutic interventions.99
In summary, each method has its unique characteristics. The co-incubation technique is straightforward, scalable, and cost-effective, making it particularly well-suited for small molecule drugs. The transfection method plays a vital role in gene and RNA therapies, enabling efficient delivery of larger molecules. Sonication is effective for drug loading and can be used with various carrier systems, though it requires careful management to prevent damage. Extrusion provides precise control over both the size of carrier particles and drug loading, making it ideal for producing uniform drug carriers. Electroporation is commonly used in gene and protein therapies, but it demands precise execution to avoid cellular harm. The choice of the most appropriate method depends on factors such as the drug type, delivery system, and specific therapeutic objectives.
Application of Plant-Derived Extracellular Vesicles for Cancer Drug Delivery
Plant-Derived Extracellular Vesicles for Colon Cancer Drug Delivery
Colon cancer is the third most prevalent cancer worldwide, and its incidence has been steadily increasing in recent years, posing an escalating threat to human health and survival.100 Chemotherapy remains the primary treatment option for colon cancer patients.101 However, the toxic side effects of chemotherapeutic drugs limit their effectiveness. As a result, a nano-delivery system that can sustain the therapeutic effects of drugs and minimize their toxicity holds significant potential for improving treatment outcomes. Several currently available drugs have been loaded into extracellular vesicles derived from plants (Table 3) and studied.
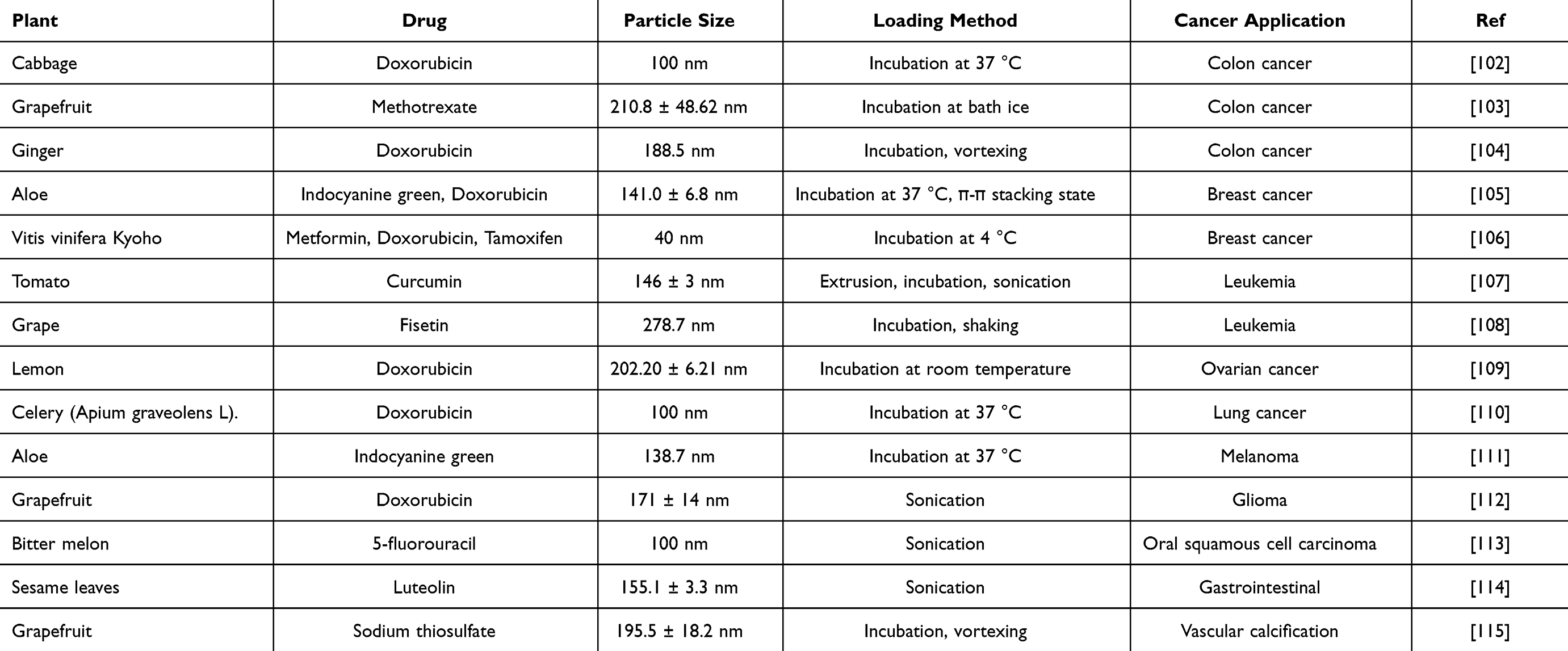 |
Table 3 Summary of the Most Recent Drug Loading Attempts on EVs of Plant Origin with Information |
You et al102 reported an approach for treating colon cancer using nanovesicles derived from cabbage (Cabex) and red cabbage (Rabex) to deliver the therapeutic agent doxorubicin (DOX). This approach effectively inhibited colon tumor growth in SW480 cell models (Figure 3A).102 In their study, NTA analysis of doxorubicin-loaded Cabex and Rabex demonstrated similar size distributions, regardless of the amount of doxorubicin incorporated (Figure 3B–F).102 Doxorubicin-loaded Cabex significantly inhibited colon cancer cell proliferation, reducing cell viability to 61.0% after treatment with doxorubicin alone, and to 57.5% following treatment with doxorubicin-loaded Cabex (Figure 3D).102 Furthermore, doxorubicin-loaded Rabex exhibited a more pronounced inhibitory effect on cell growth, with cell viability dropping to 39.5% after doxorubicin treatment alone, and further decreasing to 32.4% following treatment with doxorubicin-loaded Rabex (Figure 3G).102 The results indicate that plant-based nanovesicles, like Cabex and Rabex are effective drug carriers and can encapsulate large amounts of therapeutic substances and transport them efficiently to human cells. Cabex and Rabex hold promise as viable options for drug development. In addition, Wang et al103 proposed a novel strategy for maintaining colon macrophage homeostasis by utilizing grapefruit-derived nanovesicles (GDNs) to deliver methotrexate (MTX), which effectively reduced colon inflammation in a DSS-induced mouse model. NTA analysis revealed a significantly broader size distribution for methotrexate-loaded GDNs (GMTX) compared to unmodified GDNs, confirming the successful conjugation of MTX to the nanovesicles (Figure 4A).103 In terms of efficacy, GMTX-treated mice showed a marked reduction in body weight loss and colon length shortening induced by DSS (Figure 4B and C).103 H&E staining of colon tissues indicated that the severity of colon damage and inflammatory cell infiltration in GMTX-treated mice was notably less than in those treated with free MTX or PBS (Figure 4D and E).103 Interestingly, mice treated with free MTX exhibited worsened colitis symptoms, including more extensive colon tissue damage, greater colon length reduction, and reduced E-cadherin expression compared to PBS-treated mice (Figure 4F).103 MTX-loaded GDNs significantly alleviated the toxicity of MTX in comparison to free MTX, while significantly enhancing its therapeutic efficacy in DSS-induced colitis (Figure 4F).103 These findings suggest that GDNs could serve as effective immune modulators within the intestine, supporting macrophage homeostasis and mitigating inflammatory responses in colitis and similar conditions. Furthermore, Zhang et al104 reported a breakthrough in colon cancer treatment using ginger-derived nanovesicles (GDNVs) modified with folic acid (FA) to deliver the therapeutic agent doxorubicin (DOX) (Figure 5A).104 This approach successfully inhibited tumor growth in colon-26 xenograft models. In mice treated with FA-GDNVs or saline, tumors grew rapidly and progressively (Figure 5B),104 although no significant changes in body weight were observed (Figure 5C).104 In contrast, treatment with DOX-FA-GDNVs dramatically suppressed tumor growth, as shown in Figure 5D and E.104 Tumor weight in mice treated with Dox-FA-GDNVs was the lowest among all treatment groups (Figure 5F),104 indicating enhanced antitumor efficacy. H&E staining images (Figure 5G) 104 also showed a significant reduction in the number of cancerous cells in the Dox-FA-GDNVs group compared to the saline, free DOX, and FA-GDNVs groups. These findings demonstrate that FA-ligand-modified GDNVs effectively enhance the antitumor action of Dox through active targeting. Therefore, FA-GDNV represents an ideal drug delivery platform, effectively exerting anti-tumor effects while minimizing the adverse reactions associated with free-circulating drugs.
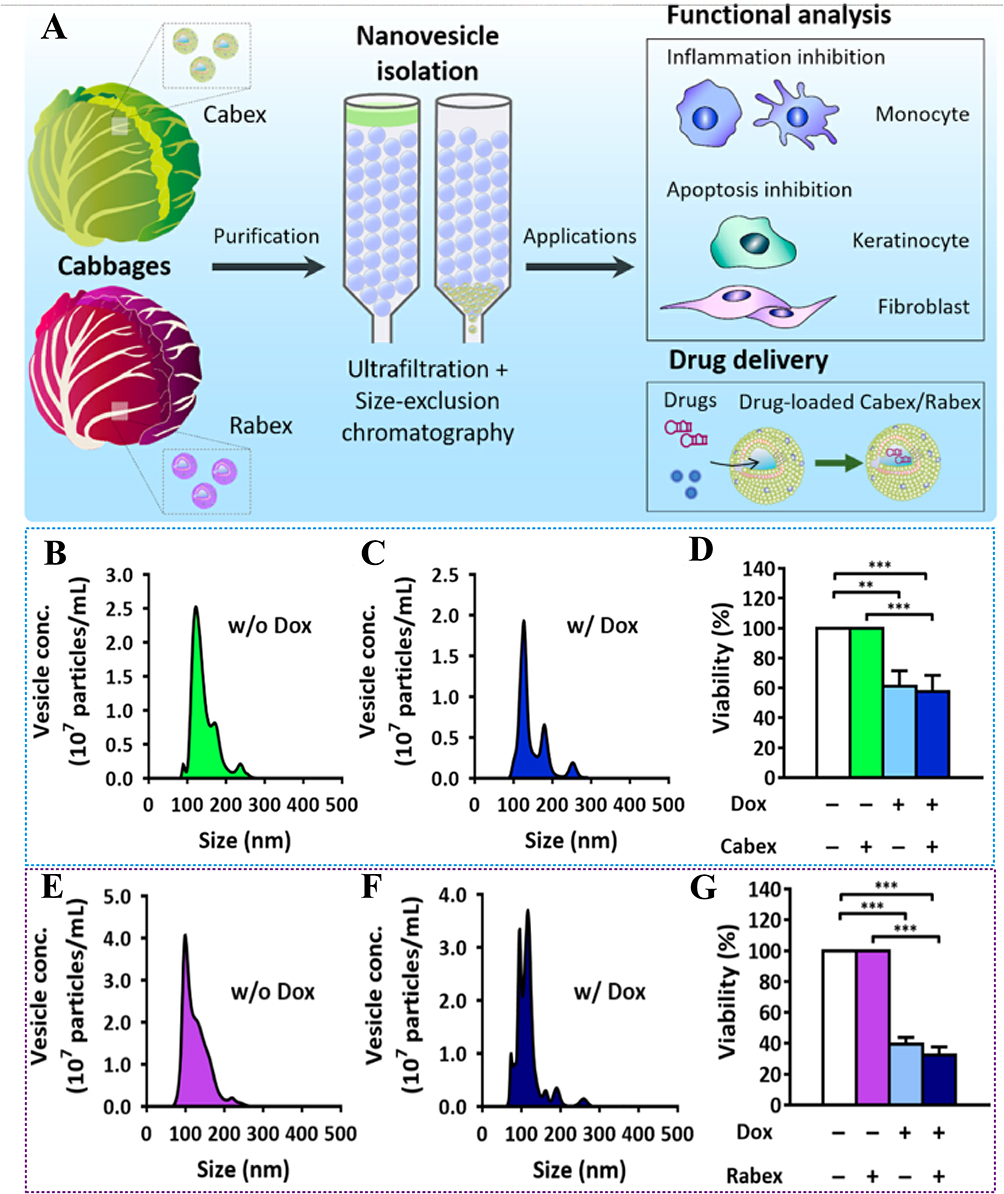 |
Figure 3 (A) Schematic illustration of exosome-like nanovesicle isolation from cabbage and the investigation of molecular functions and drug delivery applications of Cabex and Rabex. (B and C) Size distribution of Cabex (B) without and (C) with a chemotherapeutic (doxorubicin) drug load. (D) Anti-cancer effect of Cabex loaded with doxorubicin. (E and F) Size distribution of Rabex (E) without and (F) with a chemotherapeutic (doxorubicin) drug load. (G) Anti-cancer effect of Rabex loaded with doxorubicin. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 109 Copyright 2021 The Authors). Data are presented as mean ± SD. (**p < 0.01, ***p < 0.001, by t-test). |
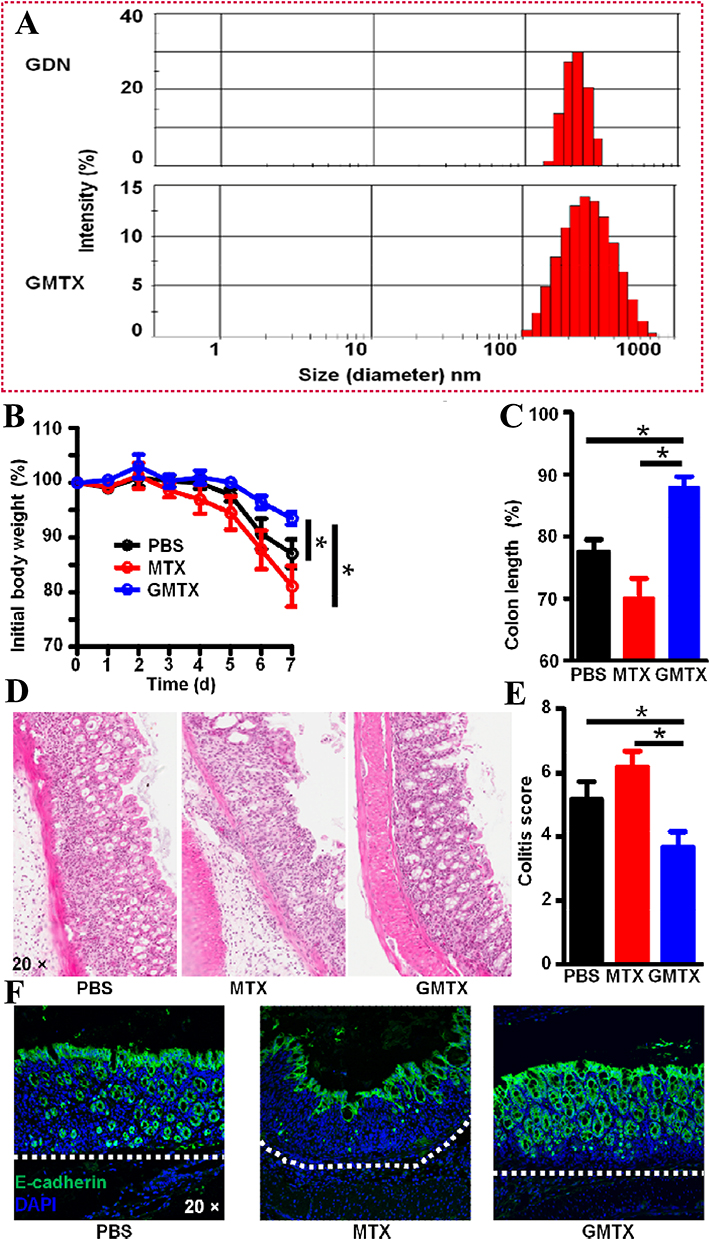 |
Figure 4 (A) The size of GMTX. The therapeutic effects of GMTX were evaluated by (B) body weight, (C) colon length, (D) pathology changes, (E) colitis score, (F) the epithelial integrity. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 110 Copyright 2013 The Authors). Data are presented as mean ± SD. (*p < 0.05, by t-test). |
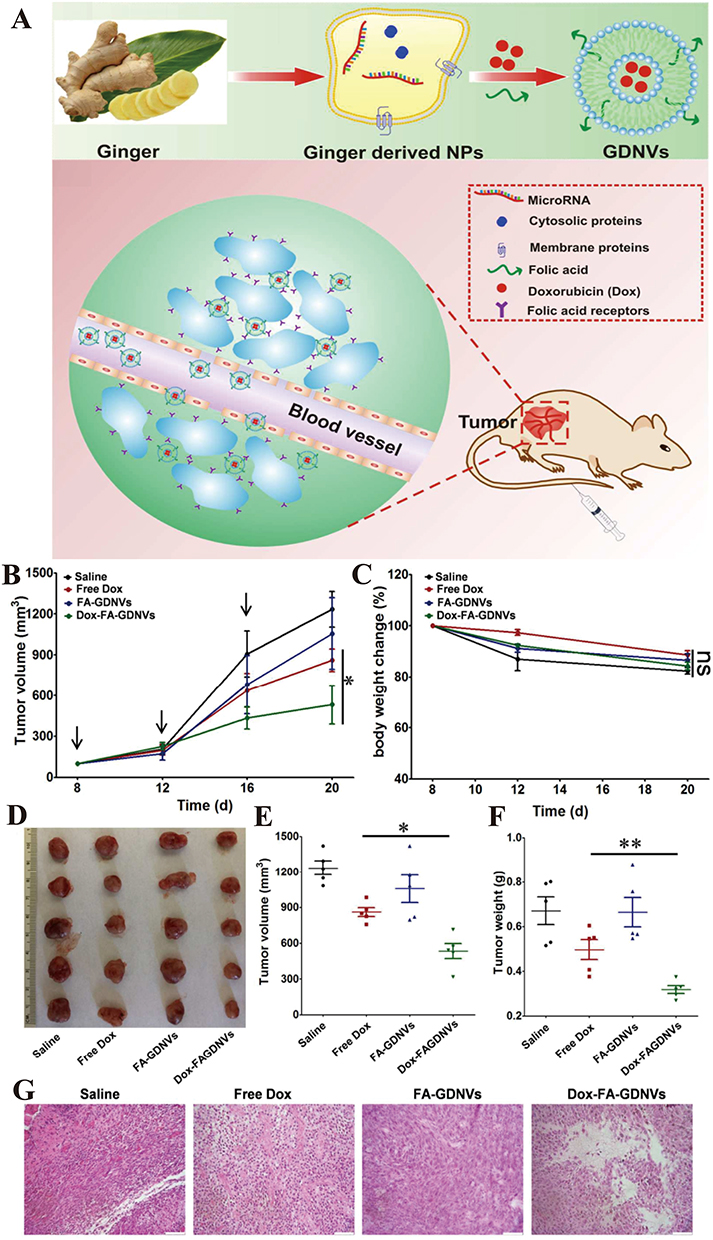 |
Figure 5 (A) Preparation of DOX-FA-GDNVs nanovectors from ginger derived lipids and schematic diagram of the targeted anti-tumor effect in vivo. (B) Tumor growth profiles in different treatment groups (saline, free Dox, FA-GDNVs and Dox-FA-GDNVs). (C) Body weight changes in different treatment groups. (D) Xenografts from each group were imaged and compared at the end of the experiments. (E) Tumor volumes at the end of the experiments were compared. (F) Tumor weights at the end of the experiment were compared. (G) Tumor tissues were removed, fixed and sectioned. H&E-staining of tumor tissues from each group were used to evaluate the anti-tumor effects. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 111 Copyright 2016 The Authors). Data are presented as mean ± SD. (*p < 0.05, **p < 0.01, by t-test). |
In summary, the varying efficacy of PDEVs from different sources, such as ginger and cabbage, in targeting various cancer types can be primarily attributed to differences in their lipid and protein profiles. These differences play a crucial role in determining the vesicles’ ability to target cancer cells, facilitate cellular uptake, and exert therapeutic effects. The lipid composition of PDEVs varies depending on the plant source, with lipids influencing the fluidity, stability, and interactions of the vesicle membranes with cancer cell membranes. Certain phospholipids or glycosphingolipids present in the vesicles can enhance their ability to fuse with specific cancer cell membranes or interact with particular receptors, leading to improved targeting. These lipid variations mean that PDEVs from different plants may preferentially interact with certain cancer cells, resulting in differences in therapeutic outcomes. Similarly, the proteins found on the surface of PDEVs, such as lectins, enzymes, and other membrane-associated proteins, play a vital role in their ability to recognize and bind to specific cancer cell receptors. These proteins often serve as mediators of cell-cell communication, enhancing the vesicles’ ability to target and be taken up by cancer cells. The protein profile differs between plant sources, so ginger-derived PDEVs may express different markers than those derived from cabbage, contributing to their varying effectiveness in targeting specific cancer types. Additionally, the carbohydrate and glycan structures on the surface of PDEVs can influence their interactions with cancer cells, with some plant vesicles having glycan profiles that are better suited for binding to cancer cell markers, thereby improving their targeting efficiency. The size and stability of PDEVs also vary by plant source and can affect their ability to penetrate tumor tissues or remain stable in the bloodstream. Smaller vesicles might be better at penetrating deep into tissues, while larger ones might be more stable for prolonged circulation, affecting drug delivery. Moreover, the way PDEVs interact with the tumor microenvironment-such as the acidic or hypoxic conditions commonly found in tumors-can influence their therapeutic efficacy. Some plant-derived vesicles may be more adept at navigating these conditions, which enhances their ability to deliver therapeutic compounds effectively. In conclusion, the lipid and protein profiles of PDEVs from different plant sources are key factors in their selective targeting and therapeutic outcomes. These profiles determine how the vesicles interact with cancer cells, and further research into the specific components of these vesicles could optimize their use for targeted drug delivery in cancer therapy. By leveraging the unique properties of plant-derived vesicles, this strategy provides an effective approach for the treatment of colon tumors, maximizing the therapeutic effects while minimizing adverse outcomes. Thus, PDEVs represent a promising candidate for enhancing targeted drug delivery in cancer therapy, offering significant advantages in terms of both efficacy and safety.
Plant-Derived Extracellular Vesicles for Breast Cancer Drug Delivery
Breast cancer is a major malignancy that poses a significant threat to women’s health worldwide.116 Particularly in the advanced stages, when metastasis occurs, the prognosis often becomes poor, resulting in limited life expectancy.117 At present, conventional treatments like chemotherapy and photothermal therapy continue to face several challenges, including tumor cell resistance, inadequate drug accumulation, and non-specific distribution, and toxicity to healthy organs. These limitations significantly hinder their effectiveness and broader application.118
To overcome the limitations of traditional treatment methods, Zeng et al105 demonstrated the potential of using a functional nanocarrier loaded with a combination of anticancer drugs for the treatment of breast cancer. They identified aloe vera gel-derived extracellular vesicles (gADNV) as a promising candidate for such a nanocarrier. gADNV was modified with an active integrin-targeted peptide, Arg-Gly-Asp (RGD), to effectively deliver the photothermal agent indocyanine green (ICG) and doxorubicin (DOX) for breast cancer treatment. The typically “competitive” interaction between ICG and DOX was transformed into a “cooperative” one through π-π stacking interactions within gADNVs, thereby enhancing the drug loading efficiency (Figure 6A).105 This dual-drug delivery system, named DIARs, demonstrated excellent stability, resistance to leakage, and strong targeting capabilities for breast tumors in vivo. DIARs exhibited high therapeutic efficacy in 4T1 tumor-bearing mouse models (Figure 6B and C),105 with no significant damage to other organs. Simultaneously, Farheen et al106 demonstrated that exosome-like nanoparticles (VKENPs) derived from Vitis vinifera Kyoho can serve as a multi-drug delivery system, effectively encapsulating and delivering various drugs for the treatment of breast tumors (BT) (Figure 7A).106 Through DLS and TEM analysis, it was shown that VKENPs could effectively carry various drugs, including metformin (MET), doxorubicin (DOX), and tamoxifen (TAX) (Figure 7B and C).106 VKENPs combined with MET were found to be non-toxic to 4T1 cells after 24 hours of incubation. In contrast, DOX caused a 20% reduction in cell survival, TAX led to a 9% reduction, and the combination of VKENPs with multiple drugs (VKE@MDT) resulted in a 27% decrease in 4T1 cell viability at a 100 μg/mL dose (Figure 7D).106 Cell death significantly increased when the incubation time was extended from 24 to 48 hours with the same concentrations (Figure 7E).106 After 48 hours of treatment, VKENPs decreased 4T1 cell survival by 18%, MET by 11%, DOX by 40%, TAX by 29%, and VKE@MDT treatment led to a dramatic 90% reduction in cell survival (Figure 7E).106 These results were consistent with findings from confocal laser scanning microscopy (Figure 7F).106 These findings highlight the selective and effective targeting role of VKENPs in delivering multiple therapeutic agents to BT cells.
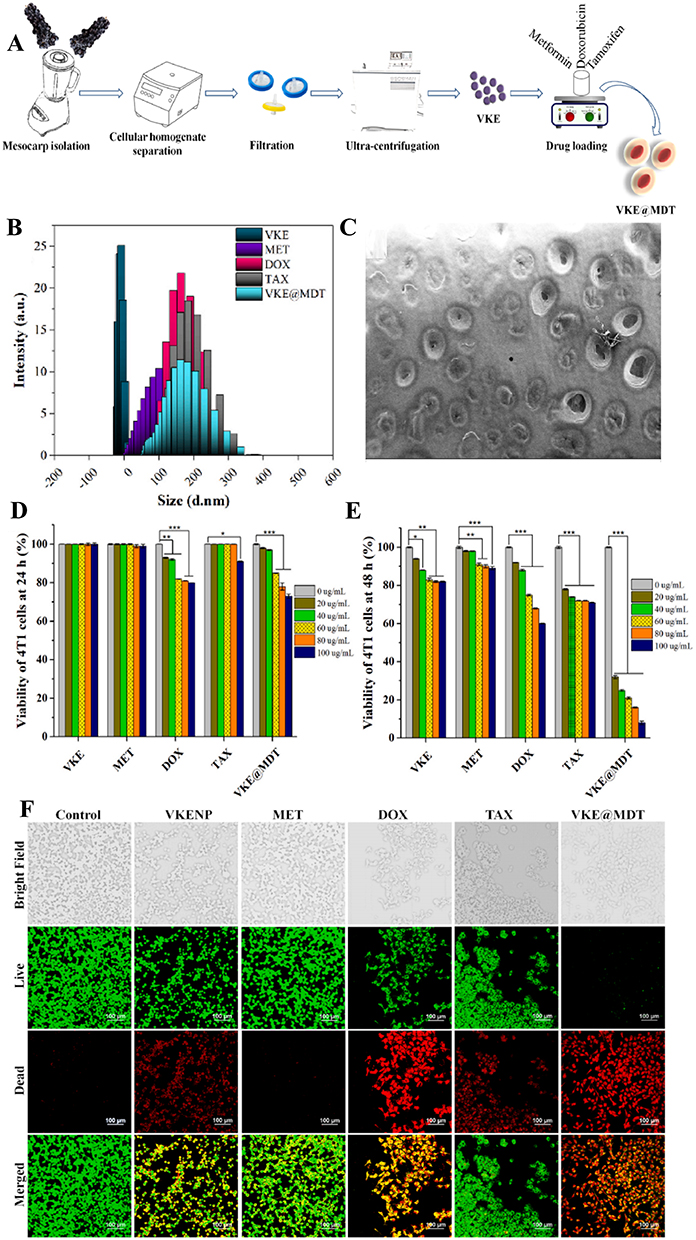 |
Figure 6 (A) Schematic diagram of the drug loading method. (B) Diagram of tumor changes in mice during treatment. (C) H&E staining of tumors of each treatment group. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 115 Copyright 2022 The Authors). |
 |
Figure 7 (A) Diagrammatic illustration of Vitis vinifera Kyoho-isolated exosome-like nanoparticles (VKENPs). (B) Size distribution of VKENPs, MET, DOX, TAX, and VKE@MDT. (C) Scanning electron micrographs of VKE@MDT. (D) 4T1 cell proliferation was examined with 20, 40, 60, 80, and 100 μg/mL VKENPs, metformin (MET), doxorubicin (DOX), tamoxifen (TAX), and VKE@MDT at 24 h and (E) 48 h. (F) Cytotoxicity assay of VKENPs and its drugs loaded cargo with 4T1 BT cells at 48 h. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 116 Copyright 2024 The Authors). Data are presented as mean ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, by t-test). |
Overall, plant-derived extracellular vesicles can serve as carriers to encapsulate multiple drugs simultaneously, such as chemotherapy agents, targeted therapy drugs, and photothermal therapy drugs, thereby constructing a dual or multimodal therapeutic system. This approach of co-delivering multiple drugs not only enables combination therapy but also improves treatment efficiency through synergistic actions. It offers new ideas and treatment options for breast cancer therapy, with the potential to play an important role in enhancing efficacy, reducing side effects, and overcoming drug resistance.
Plant-Derived Extracellular Vesicles for Leukemia Drug Delivery
Leukemia research is progressing swiftly, with innovative treatments such as targeted therapies, CAR-T cell therapy, and gene editing showing considerable promise. However, obstacles like drug resistance and treatment-related side effects remain significant challenges. Plant-derived exosomes offer a promising new approach to drug delivery, with the potential to enable targeted therapy, overcome drug resistance, and reduce side effects. As research continues to evolve, plant-derived nanovesicle-based drug delivery systems may become a crucial tool in leukemia treatment, either as standalone therapies or in combination with other treatment modalities.
Mammadova et al107 introduced an innovative method for treating leukemia-associated inflammation by utilizing nanovesicles derived from tomato to deliver curcumin using three distinct drug-loading techniques (extrusion, incubation, sonication) (Figure 8A).107 This approach effectively inhibited inflammatory responses in THP-1 cell models. The curcumin-loaded nanovesicles exerted anti-inflammatory effects by decreasing the mRNA levels of both IL-1β and IL-6 (Figure 8B).107 While all three drug loading methods resulted in similar reductions in IL-1β expression, the sample prepared by sonication produced a more significant decrease in IL-6 mRNA levels. These results suggest that different drug loading techniques can substantially influence the drug’s loading capacity and targeting efficiency, which in turn affects the therapeutic outcomes of plant-derived exosomes. Consequently, selecting the most appropriate drug-loading method should be based on a thorough evaluation of the drug’s characteristics, therapeutic goals, and the properties of the exosomes. Subsequently, Sarvarian et al108 proposed a novel approach for leukemia treatment by utilizing grape-derived nanovesicles (GDNs) to deliver the natural flavonoid Fisetin (FIS). This method effectively inhibited leukemia tumor growth in MOLT-4 cells. Their MTT assay demonstrated that the viability of cells treated with FIS-loaded grape-derived nanovesicles (FIS-GDNs) was significantly lower than that of cells treated with free FIS, suggesting a stronger cytotoxic effect of FIS-GDNs (Figure 8C–E).108 The incorporation of GDNs enhanced the solubility and bioavailability of FIS, promoting increased apoptosis in the cells. This enhanced apoptotic activity resulted in greater cytotoxicity and contributed to the improved antitumor efficacy of the FIS-GDNs nanovesicle formulation.
 |
Figure 8 (A) Schematic representation of the process to prepare curcumin loaded tomato nanovesicles by three methods: extrusion, direct loading by incubation followed by sucrose density gradient ultracentrifugation, and sonication. (B) (a) Pre-incubation and treatment conditions. Anti-inflammatory effect was evaluated on THP-1 cell line measuring the levels of pro-inflammatory cytokines (b) IL-1β; (c) IL-6. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 117 Copyright 2023 The Authors). (C) Cytotoxic effect of Fisetin (FIS) and Fisetin-loaded grape-derived nanoparticles (FIS-GDN) on MOLT-4 cells by MTT assay. IC50 values after 24 h treatment MOLT-4 cells with FIS and FIS-GDN. (D) IC50 values after 48 h treatment MOLT-4 cells with FIS and FIS-GDN. (E) Statistical analysis of comparisons between groups. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 118 Copyright 2023 The Authors). Data are presented as mean ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, by t-test). |
Plant-derived nanovesicles provide a versatile drug delivery platform, enabling the selection of the most effective loading methods for leukemia treatment. These nanovesicles can transport not only conventional chemotherapy agents but also natural compounds, offering significant potential for natural drugs delivery in leukemia therapy.
Plant-Derived Extracellular Vesicles for Ovarian Cancer Drug Delivery
Ovarian cancer is one of the most prevalent malignant tumors among women worldwide. Its early symptoms are often subtle and easily overlooked, leading to diagnosis at more advanced stages and contributing to high mortality rates. Despite interventions like surgery and chemotherapy, ovarian cancer frequently recurs, and drug resistance presents a major challenge. Multidrug resistance (MDR) is a key barrier in ovarian cancer therapy, with tumor cells evading chemotherapy drugs through various mechanisms such as drug efflux and metabolic changes, which diminish the efficacy of treatment and worsen the prognosis. Plant-derived extracellular vesicles have emerged as a promising natural drug delivery system to address drug resistance in ovarian cancer.
Xiao et al109 developed a biomimetic drug delivery system using lemon-derived extracellular vesicles (EVs) loaded with doxorubicin (DOX) and modified with heparin-cRGD to form HRED nanocomplexes, which effectively combated multidrug resistance in cancer (Figure 9A).109 In the preparation of HRED nanodrugs, Transmission electron microscopy (TEM) images revealed the characteristic vesicle structure of EVs, ED, HRE, and HRED, all displaying lipid bilayers (Figure 9B).109 The particle sizes of EVs, ED, HRE, and HRED were measured as 151.63 ± 5.20, 155.8 ± 12.71, 201.70 ± 5.07, and 202.20 ± 6.21 nm, respectively, confirming the successful encapsulation of the drug within the nanovesicles (Figure 9C).109 The HRED system exhibited a significant tumor inhibition effect compared to PBS, EVs, free DOX, and ED (Figure 9D–G).109 In contrast, free DOX had no inhibitory effect on tumor growth, indicating that the orthotopic SKOV3/DOX-Luc ovarian cancer xenograft nude mouse model was resistant to DOX treatment. This highlights that the functionalization of the EV surface with heparin-cRGD (HR) and encapsulation of doxorubicin (DOX) enables HRED to effectively target and penetrate doxorubicin-resistant cancer cells through multiple endocytic pathways, primarily caveolin-mediated endocytosis. The system also demonstrated excellent cellular uptake, attributed to its diverse endocytosis mechanisms. This contributes to reducing intracellular energy consumption and lowers adenosine triphosphate (ATP) production, effectively minimizing drug efflux. As a result, HRED exhibited strong anti-proliferative effects on doxorubicin-resistant ovarian cancer cells, successfully overcoming multidrug resistance.
 |
Figure 9 (A) Schematic illustration of lemon-derived extracellular vesicle (EV) nanodrugs for overcoming cancer multidrug resistance. (B) Transmission electron microscopy (TEM) images of lemon-derived EVs, ED, HRE, and HRED. (C) The size distribution of lemon-derived EVs, ED, HRE, and HRED. (D) IVIS bioluminescent imaging of orthotopic SKOV3/DOX-Luc ovarian cancer xenograft nude mouse from each group during treatment. (E) Protocol for tumor implantation and treatment used in this study. (F) The luminescent signal intensity of the mice in all groups. (G) Body weight of the mice in all groups. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 119 Copyright 2022 The Authors). |
In conclusion, by utilizing plant-derived extracellular vesicles-based delivery systems, it becomes possible to circumvent the resistance mechanisms of tumor cells, thereby boosting the effectiveness of chemotherapy and improving patient outcomes. Therefore, plant-derived extracellular vesicles show great potential for application in ovarian cancer therapy and are expected to become a novel strategy for overcoming chemotherapy resistance in the future.
Plant-Derived Extracellular Vesicles for Other Cancer Drug Delivery
Plant-derived extracellular vesicle-based drug delivery platforms are also extensively applied in the treatment of various cancers, including lung cancer, melanoma, glioma, oral squamous cell carcinoma, gastric cancer, and vascular calcification.
Lu et al110 highlighted the promising potential of celery exosome-like nanovesicles (CELNs) as a carrier for doxorubicin (DOX), an anticancer drug, in the treatment of lung cancer (Figure 10A).110 The CELNs demonstrated excellent drug-carrying capacity, successfully encapsulating DOX through co-incubation. The drug loading efficiency reached 87.03% (Figure 10B).110 Nanoparticle tracking analysis (NTA) showed that the median size of the DOX-loaded CELNs (CELNs-DOX) was 113.7 nm (Figure 10C),110 which was similar to that of CELNs without DOX. Additionally, transmission electron microscopy (TEM) images revealed that the overall structure of CELNs-DOX was similar to that of CELNs without the drug (Figure 10D).110 Subsequently, in vivo experiments demonstrated that CELNs-DOX exhibited superior antitumor efficacy and safety in the A549 subcutaneous tumor mouse model, outperforming free DOX treatment (Figure 10E–G).110 Mice in both the CELNs-DOX and liposome-DOX groups maintained stable body weight, indicating that encapsulating DOX in CELNs or liposomes significantly reduced its inherent toxicity. Notably, mice in the CELNs-DOX group had smaller tumor mass and volume than those in the liposome-DOX group, suggesting that CELNs-DOX may provide superior therapeutic effects (Figure 10H–J).110 The study concluded that CELNs are an efficient and safe biological drug carrier, with CELNs-DOX effectively reducing DOX-related toxicity. CELNs hold substantial potential as a drug carrier for various biomedical applications.
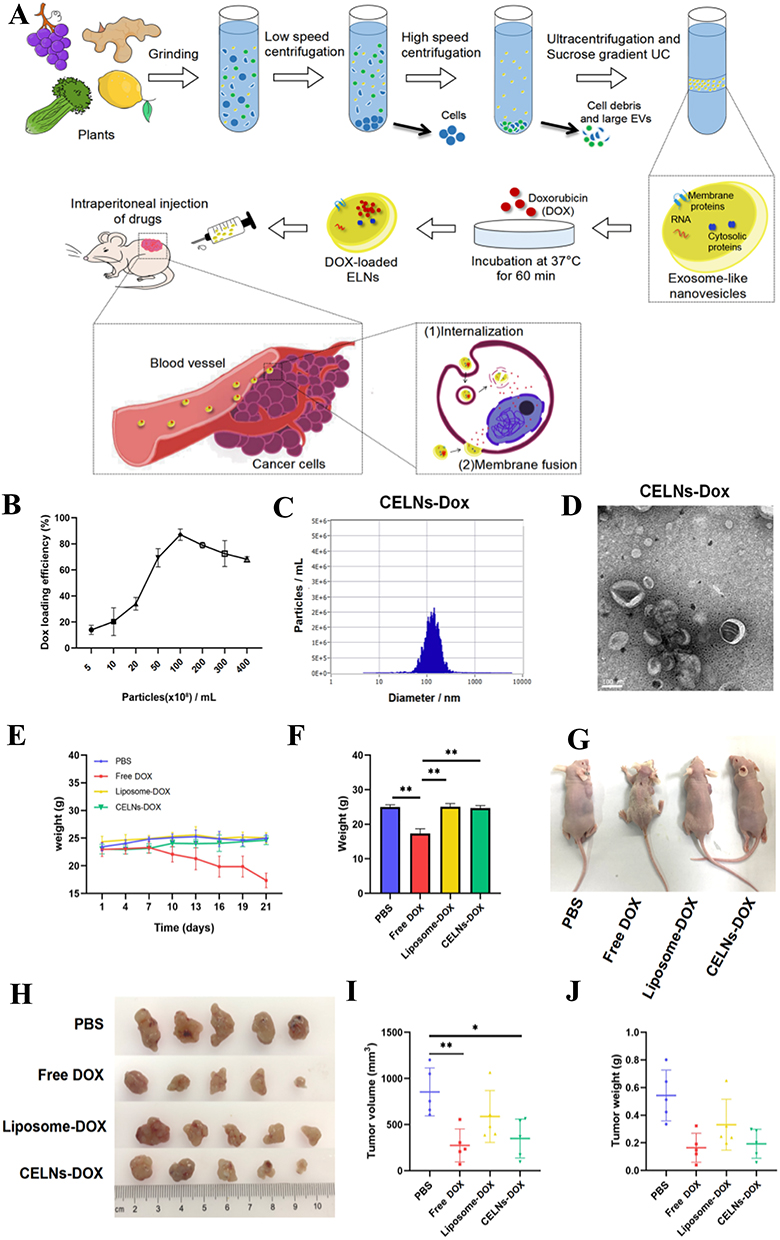 |
Figure 10 (A) Schematic diagram of the isolation of celery exosome-like nanovesicles (CELNs) from lemon. (B) Loading efficiency of doxorubicin in CELNs was measured. (C) Particle size distribution of CELNs-doxorubicin was measured by nanoparticle tracking analysis. (D) CELNs-doxorubicin were characterized by transmission electron microscopy; the scale bar indicates 100 nm. (E) Body weight changes of the mice in four groups (PBS, DOX, CELNs-DOX, and liposome-DOX). (F) Comparison of body weight of the four groups of mice at the end of the experiments. (G) Comparison of body size of the four groups of mice at the end. (H) Photographs were performed on each group of tumors. (I and J) Size and weight of the tumor in each group were evaluated. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 120 Copyright 2023 The Authors). Data are presented as mean ± SD. (*p < 0.05, **p < 0.01, by t-test). |
Zeng et al111 developed aloe gel-derived nanovesicles (gADNVs) that exhibited strong stability and leak-proof characteristics, making them highly effective for melanoma treatment (Figure 11A).111 When loaded with indocyanine green (ICG), the ICG/gADNVs exhibited outstanding stability, retaining more than 90% of the drug after 30 days of storage under both heat and serum conditions (Figure 11B–E).111 Even after this extended period, ICG/gADNVs remained potent in targeting melanoma cells and inhibiting tumor growth, outperforming both free ICG and ICG-loaded liposomes, which contributed to reduce production and tumor therapy costs. Furthermore, gADNVs demonstrated significant ability to penetrate mouse skin, highlighting their potential for noninvasive transdermal drug delivery. Overall, this study presents a promising, cost-effective, and safe strategy for ICG delivery in skin cancer therapy.
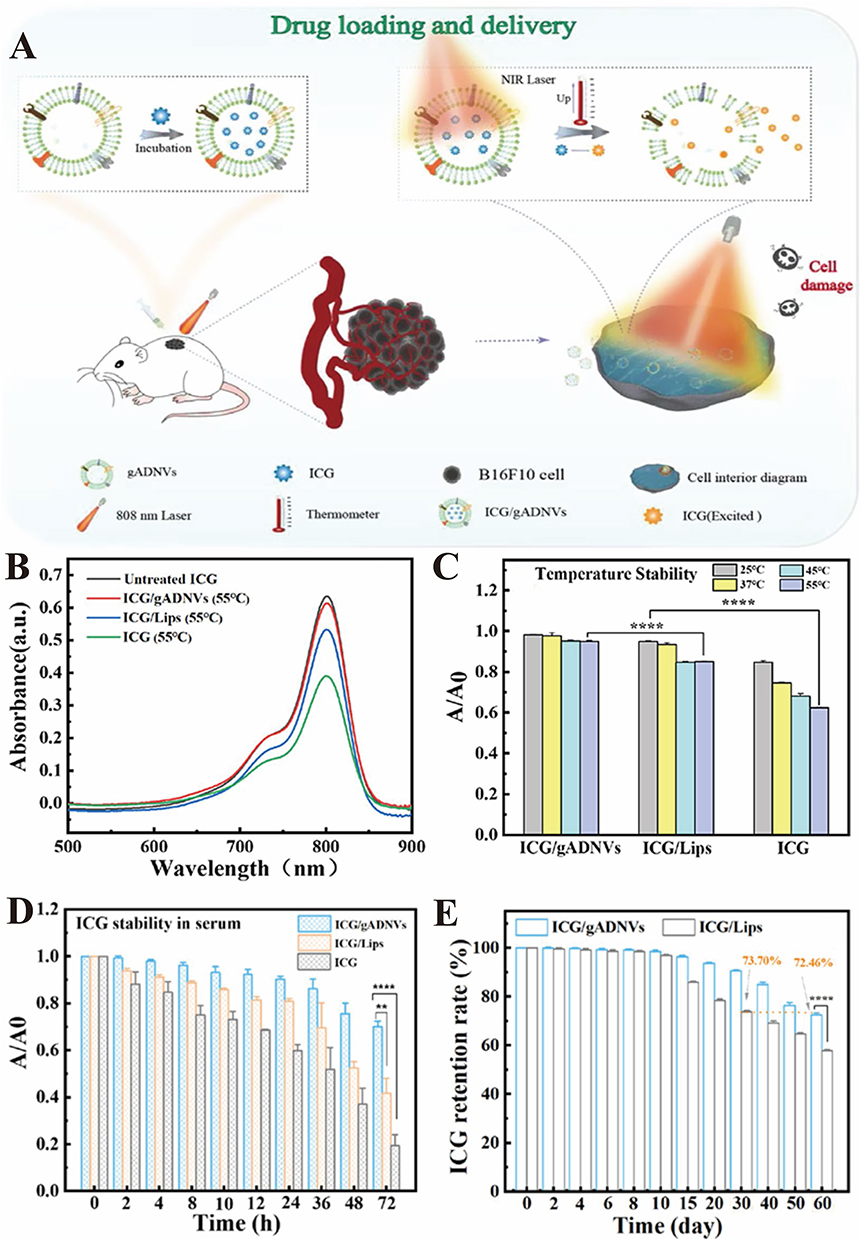 |
Figure 11 (A) The indocyanine green (ICG) loading and delivery based on gel-Aloe derived nanovesicle (gADNVs) for cancer therapy. (B) The UV-vis spectra of ICG/gADNVs, ICG/Lips and ICG after 12 h incubation in 55 °C compared with untreated ICG. (C) The thermal stability of ICG after loading in gADNVs and Lips. A/A0 was the ratio of ICG absorbance before and after heating. (D) The stability of ICG/gADNVs, ICG/Lips and ICG in serum. (E) The retention rate of ICG in gADNVs and Lips after storage for different time. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 121 Copyright 2021 The Authors). Data are presented as mean ± SD. (**p < 0.01, ***p < 0.001, by t-test). |
Moon et al112 proposed a nanocarrier system utilizing grapefruit-derived plant extracellular vesicles (pEVs) loaded with doxorubicin (DOX) and functionalized with aptamer-DSPE-PEG2000-Mal (Figure 12A).112 This formulation showed effective targeting capabilities against glioblastoma. The size of the doxorubicin-loaded pEVs was 198.4 ± 5.5 nm, which closely resembled the size of unmodified pEVs (Figure 12B and C).112 Approximately 45% of doxorubicin was released over 48 h at pH 7.4. In contrast, under acidic conditions (pH = 5), the release reached approximately 80% during the same period, after an initial rapid burst (Figure 12D).112 While the release at pH 7.4 slowed considerably after the initial burst, at pH 5, the release rate remained steady, continuing to increase throughout the 48 h. The sustained release of doxorubicin at pH 5 confirmed that pEVs effectively provide prolonged drug delivery. Moreover, by modifying the surface of pEVs with aptamers, proteins, miRNAs, and other biomolecules using techniques like hydrophobic insertion and click chemistry, it is possible to target a variety of cells or organs beyond endothelial cells. This strategy offers great potential for a wide range of applications in nanomedicine. Next, Yang’s group113 confirmed that bitter melon-derived extracellular vesicles (BMEVs) can enhance therapeutic efficacy and reduce resistance of oral squamous cell carcinoma (OSCC) to 5-fluorouracil (5-FU). The tumor volume in the BMEVs + 5-FU nanocarrier group was significantly smaller than that in the 5-FU and BMEVs groups (Figure 12E–G).113 TUNEL staining of tumor sections showed a higher number of apoptotic cells in the BMEVs + 5-FU group (Figure 12H).113 These findings present a novel strategy to enhance therapeutic efficacy and overcome drug resistance in cancer cells against chemotherapeutic agents, offering proof-of-concept for the future development of plant-derived extracellular vesicles (PDEVs)-enhanced therapies.
 |
Figure 12 (A) Schematic illustration of the synthesis and treatment of surface modified grapefruit plant-derived extracellular vesicles (pEVs) for a targeting delivery carrier and a drug loading carrier. (B) The average hydrodynamic size of pEVs before and after drug loading. (C) Concentration of pEV-doxorubicin after doxorubicin loading by probe sonication. (D) The drug release profile of pEV-Doxorubicin in PBS at pH 5 and 7.4, respectively. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 122 Copyright 2023 The Authors). (E) BMEVs enhanced the cytotoxic effect of 5-FU in vivo. Tumor growth curve of the control, 5-FU, BMEVs and BMEVs + 5-FU groups. (F) Representative images of CAL 27 tumors. (G) Quantification analysis of the weight of the tumors. (H) Representative TUNEL staining images of the tumors. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 123 Copyright 2021 The Authors). Data are presented as mean ± SD. (**p < 0.01, ***p < 0.001, by t-test). |
Jiang et al114 engineered exosome-like nanovesicles derived from sesame leaves (Exo) to deliver luteolin (Lu) (Exo@Lu), leveraging these nanovesicles as carriers to achieve anti-inflammatory effects in the gastrointestinal tract (Figure 13A).114 Exo and Exo@Lu show no adverse effects on the viability of RAW264.7 cells within the concentration range of 5 to 40 μg/mL, indicating excellent safety and biocompatibility (Figure 13B).114 Furthermore, Exo@Lu displayed efficient cellular uptake, as monitored by cellular ROS levels using CLSM (Figure 13C).114 The reduced green fluorescence of DCF in Exo@Lu suggested enhanced internalization, which facilitated the efficient release of Lu. Encapsulating Lu in Exo resulted in superior cellular uptake, antioxidant activity, and anti-inflammatory effects compared to both free Lu and Exo alone. Gastroenteritis can potentially progress into gastric cancer. With timely and effective intervention, the risk of developing gastric cancer can be greatly reduced, leading to better overall public health. This strategy shows promise as an early intervention strategy, aiming to lower the incidence of gastrointestinal cancers.
 |
Figure 13 (A) The extraction and purification of sesame leaf exosomes (Exo) to encapsulate luteolin (Lu). (B) Effects of Exo, Exo@Lu, and free Lu on cell viability. (C) CLSM images of Exo, free Lu and Exo@Lu incubated with RAW264.7 cells for 6 h under LPS exposure. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 124 Copyright 2024 The Authors). |
Feng et al115 fabricated a biomimetic nanocarrier system using grapefruit-derived extracellular vesicles (EVs) to encapsulate sodium thiosulfate (STS), which was further functionalized with a hydroxyapatite crystal-binding peptide (ESTP) for targeted delivery to vascular calcification (VC) (Figure 14A).115 Transmission electron microscopy (TEM) images confirmed the typical vesicular structure of EVs, ES, and ESTP, all displaying lipid bilayers (Figure 14B).115 The zeta potential and size distribution changes for EVs, ES, ESTP confirm the successful formulation of grapefruit-derived ESTP nanodrugs (Figure 14C, and D).115 Moreover, ESTP nanodrugs also exhibited efficient therapeutic efficacy for VC in vivo. Mice treated with ESTP showed a marked reduction in VC area. The anti-VC effect of ESTP was more pronounced than that of EVs or STS monotherapies alone, suggesting a synergistic effect between EVs and STS in treating VC (Figure 14E–G).115 Taken together, these results demonstrate that ESTP nanodrugs possess excellent biosafety and effectively inhibit VC progression by enhancing targeted accumulation at the site of calcification, making them promising for clinical translation as a treatment for VC.
 |
Figure 14 (A) Schematic illustration of ESTP for suppressing vascular calcification. Grapefruit-derived extracellular vesicle (EVs) nanodrugs (marked with ESTP) were fabricated by loading sodium thiosulfate (STS) in EVs and then modifying hydroxyapatite-binding peptide SP5-52-SH (TP) onto the surface of EVs using DSPE-(PEG)2000-maleimide as a linker. (B) Transmission electron microscopy (TEM) images of grapefruit-derived EVs, ES, and ESTP. (C) Zeta potential and (D) size distribution of EVs, ES, and ESTP measured by dynamic light scattering (DLS). (E) Representative Alizarin Red S staining and the (F) quantification of positive areas of calcified aortas (from the ascending aortic root to the iliac bifurcation). (G) Survival curves of mice with various treatments. (Reproduced with permission under Creative Commons CC BY 4.0 license from ref. 125 Copyright 2023 The Authors). Data are presented as mean ± SD. (*p < 0.05, **p < 0.01, ***p < 0.001, by t-test). |
In summary, plant-derived extracellular vesicles (PDEVs) are natural biomolecular carriers renowned for their exceptional biocompatibility and low immunogenicity. These vesicles are highly effective at encapsulating a wide range of drugs and therapeutic molecules, positioning them as ideal candidates for the development of efficient drug delivery systems and targeted therapies for various cancers. Therefore, plant-derived extracellular vesicles hold great potential for advancing clinical treatments in the future.
Comparative Analysis and Future Perspectives for PDEV Therapy
The diverse plant sources discussed in Application of Plant-Derived Extracellular Vesicles for Cancer Drug Delivery reveal a rich landscape of PDEVs with significant potential in anticancer therapy. A comparative analysis of their origins, lipid compositions, and demonstrated efficacies allows for the identification of key trends and the most promising candidates for future development.
Correlation Between PDEV Source, Lipid Composition, and Anticancer Efficacy
A clear correlation exists between the botanical source (and specifically the plant organ from which PDEVs are isolated), their inherent lipid composition, and their mechanism of action against cancer cells.
Fruit-Derived PDEVs (eg, Grapefruit, Lemon, Tomato, Bitter Melon, Vitis vinifera Kyoho, Grape): PDEVs from fruits often possess a lipid bilayer rich in phosphatidylcholine (PC) and other phospholipids that facilitate robust membrane fusion with mammalian cells. This property is crucial for the efficient delivery of bioactive cargo. Their efficacy appears to be broad-spectrum but with notable specificities. Grapefruit and Lemon PDEVs have demonstrated exceptional stability and a remarkable ability to serve as drug delivery vehicles for chemotherapeutics like paclitaxel and doxorubicin, significantly enhancing their antitumor effects against various cancers (eg, colon cancer, pancreatic cancer) while mitigating systemic toxicity. Their neutral lipid profile and surface proteins may contribute to their low immunogenicity and high biocompatibility.
Tomato PDEVs are particularly enriched in lycopene, a potent antioxidant. Their activity is often associated with antioxidant and anti-proliferative effects, making them promising for preventative strategies or for targeting cancers where oxidative stress plays a key role. Bitter Melon PDEVs exhibit a more targeted efficacy, showing strong pro-apoptotic effects specifically in liver cancer cells. This suggests the presence of unique lipids or embedded proteins that confer tissue tropism.
Vitis vinifera Kyoho and Grape PDEVs are noted for their high content of anthocyanins and other flavonoids, which contribute to their strong anti-inflammatory and anti-angiogenic properties, potentially inhibiting tumor growth and metastasis. Leaf-Derived PDEVs (eg, Cabbage, Sesame Leaves, Aloe, Celery): PDEVs isolated from leaves often carry a distinct phytochemical cargo reflective of their photosynthetic and defensive roles. Their lipid composition may include unique glycolipids and secondary metabolites. Ginger PDEVs (derived from the rhizome, a modified stem) are a standout example, demonstrating a unique lipid composition that includes phosphatidic acid (PA), which facilitates their preferential uptake by intestinal macrophages and subsequent induction of heme oxygenase-1 (HO-1). This underpins their potent anti-inflammatory and colitis-associated cancer preventive effects, a mechanism distinct from most fruit-derived PDEVs. Cabbage PDEVs have shown efficacy in ameliorating colitis, suggesting a similar niche in gastrointestinal health and potentially preventing inflammation-driven carcinogenesis. Aloe and Sesame Leaf PDEVs possess strong antioxidant and wound-healing properties. Their application in cancer therapy may be synergistic, particularly in mitigating the side effects (eg, oral mucositis, dermatitis) of conventional cancer treatments. Root-Derived PDEVs (eg, Ginger, as a rhizome, is often categorized here functionally): As exemplified by ginger, PDEVs from underground organs can have highly specialized interactions with the gut microbiome and the immune system, offering a unique, indirect pathway for anticancer activity that is less about direct cytotoxicity and more about modulating the tumor microenvironment.
In general, fruit-derived PDEVs appear to be superior as versatile, efficient, and biocompatible drug delivery platforms due to their stability and fusion competence. In contrast, leaf and root-derived PDEVs often exhibit more specialized, pharmacologically active properties rooted in their innate immunomodulatory and anti-inflammatory cargo, making them excellent candidates for cancer prevention and combination therapy to manage inflammation.
Identification of Promising Plant Sources
Based on the synthesized data, the most promising plant sources for future PDEV-based cancer therapy are Grapefruit and Lemon, Ginger, and Bitter Melon. Grapefruit and Lemon stand out as front-runners for drug delivery applications due to their well-documented ability to be loaded with diverse chemotherapeutic agents, their scalability of isolation, and their excellent safety profile in preclinical models. These characteristics position them as ideal candidates for clinical translation, with future work focusing on engineering their surfaces for active tumor targeting to enhance their precision and efficacy in drug delivery. Ginger, on the other hand, represents a shift from direct drug delivery to immunomodulation. Its efficacy in preventing colitis-associated cancer through a well-defined HO-1 pathway highlights its potential for cancer chemoprevention, particularly in individuals at high risk due to chronic inflammatory conditions. Ginger is thus a top candidate for preventative oncology and adjuvant therapy, offering new avenues for cancer prevention and supportive treatment. Meanwhile, Bitter Melon’s apparent organotropism toward liver tissue makes its PDEVs particularly promising for targeted therapies aimed at hepatocellular carcinoma. Investigating the specific ligand or lipid responsible for this organ specificity could lead to highly targeted therapies, either as native nanotherapeutics or as targeting moieties for engineered vesicles designed to treat liver cancer.
In conclusion, while each PDEV source offers unique advantages, Grapefruit and Lemon for delivery, Ginger for immunomodulation, and Bitter Melon for potential targeting stand out as the most immediately promising avenues in the evolving field of PDEV-based cancer therapy.
Current Bottlenecks in PDEV Standardization and Regulatory Challenges
Despite the burgeoning interest in Plant-Derived Extracellular Vesicles (PDEVs) for therapeutic applications, the field faces significant challenges in standardization that hinder the comparability of research findings and clinical translation. A primary obstacle is the lack of unified nomenclature. PDEVs are variously referred to in the literature as “plant exosomes”, “plant-derived extracellular vesicles”, or “extracellular vesicles-like particles (EVLPs)”.119 This terminological inconsistency creates confusion and complicates literature mining and scientific communication. Recent efforts have begun to address this issue by proposing standardized naming conventions that reflect the biological source and preparation method, which is a critical first step toward field-wide standardization.119
Furthermore, there is a pronounced heterogeneity in methods used for the isolation and characterization of PDEVs. Different research groups employ diverse techniques (eg, differential ultracentrifugation, precipitation, size-exclusion chromatography), each with varying yields, purity, and potential for co-isolating non-vesicular contaminants. This makes it exceedingly difficult to directly compare the biological activities or physicochemical properties of PDEVs reported in different studies. The problem extends to characterization, where minimal metrics for size, concentration, and marker expression are not universally agreed upon. Establishing a set of minimum information standards for PDEV characterization, akin to the MISEV guidelines for mammalian EVs, is an urgent need for the field. The Table 4 below summarizes the core challenges across the PDEV research and development pipeline:
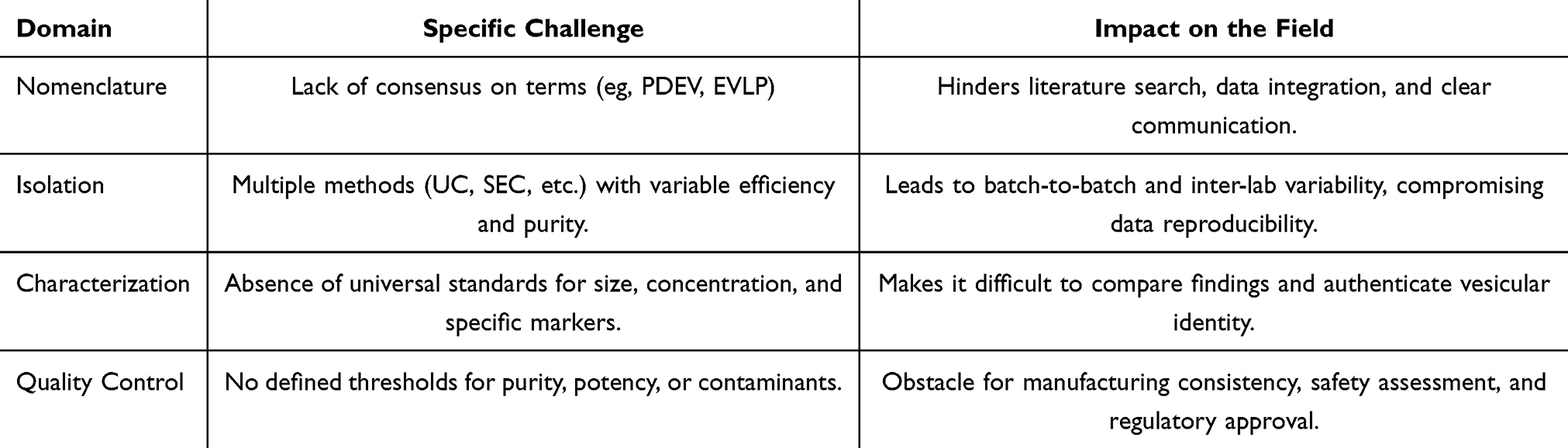 |
Table 4 Major Bottlenecks in PDEV Standardization and Their Implications |
Finally, the path to clinical application is obscured by undefined regulatory frameworks. As complex biological products, PDEVs require application-specific benchmarks for safety, purity, and potency. Critical parameters, such as long-term stability, immunogenicity, and in vivo pharmacokinetics, need to be systematically evaluated using standardized assays. The establishment of a robust manufacturing pipeline under Good Manufacturing Practice (GMP) conditions is paramount. Initiatives like the founding of the Chinese Herbal Vesicles Guangdong Provincial Engineering Research Centre aim to tackle these translational challenges, signaling a move towards centralized and standardized research efforts.119 Ultimately, close collaboration between researchers, industry, and regulatory bodies is essential to develop tailored guidelines that will enable PDEVs to transition from promising laboratory discoveries to approved therapeutics.
Future Perspectives
The exploration of Plant-Derived Extracellular Vesicles (PDEVs) has evolved from early descriptive observations to a promising frontier in nanomedicine. Initial research, particularly with vesicles derived from grapefruit and ginger, established PDEVs as dual-function entities-acting both as therapeutic agents and as drug delivery vehicles. Notably, PDEVs have shown potential in oral delivery for treating gut-related diseases, benefiting from their stability in the gastrointestinal tract and innate anti-inflammatory properties. These early studies highlighted the unique advantages of PDEVs over synthetic nanoparticles, opening the door to their broader application in medicine.
Despite these promising therapeutic outcomes, the clinical translation of PDEVs faces several significant challenges, primarily rooted in heterogeneity and the absence of standardized frameworks for their production. A major obstacle lies in the lack of scalable and reproducible biomanufacturing processes. Traditional methods of plant extraction led to considerable batch-to-batch variability in PDEV size, composition, and function. Emerging platforms like the Temporary Immersion Bioreactor System (TIBS) have made strides toward addressing this issue by enabling more controlled production, reducing energy consumption, and improving both cost-effectiveness and quality control. Compounding this issue is the widespread lack of standardized reporting for critical quality and functional attributes. Many studies, particularly earlier foundational ones, fail to report essential quantitative metrics-such as drug-loading efficiency, drug-to-lipid ratios, and in vitro leakage kinetics-that are indispensable for cross-study comparison, process optimization, and regulatory evaluation. However, the regulatory pathway for PDEVs remains undefined, as they are complex biological products that require specific benchmarks and robust manufacturing pipelines to ensure quality, safety, and efficacy.
One of the most critical gaps hindering the clinical application of PDEVs is the absence of comprehensive systemic biodistribution data, including blood half-life, clearance kinetics, and tissue accumulation profiles. This gap is not merely an oversight but a fundamental bottleneck in advancing PDEVs from preclinical success to clinical reality. The lack of pharmacokinetic and biodistribution studies, especially using advanced imaging techniques such as Positron Emission Tomography (PET) or In Vivo Imaging Systems (IVIS), stems from the early-stage nature of the field. Much of the initial research focused on validating the basic therapeutic potential of PDEVs in localized disease models, which bypass systemic circulation and thus do not require extensive biodistribution studies.
Moreover, reliably labeling PDEVs for tracking their biodistribution presents unique technical challenges. Unlike synthetic nanoparticles, which can be easily conjugated with fluorophores or radionuclides, labeling PDEVs without compromising their biological activity is complex. The vesicles’ natural composition, including lipid bilayers and surface proteins, must be preserved for their efficacy, making tracking difficult. Additionally, distinguishing between the signal from the vesicle and its cargo in vivo complicates the interpretation of the biodistribution data.
This gap in biodistribution knowledge significantly impedes the ability to determine appropriate dosing regimens, assess potential off-target toxicities, or optimize delivery methods, especially for systemic therapies such as intravenous (IV) delivery. Without clear data showing that PDEVs can avoid rapid uptake by the reticuloendothelial system (RES) and reach distant therapeutic targets, their potential for treating disseminated diseases like cancer remains speculative.
To address these challenges, a multi-pronged approach is essential. First, the development of robust and standardized labeling protocols for PDEVs is crucial, ensuring that their biological integrity is maintained while enabling reliable tracking in vivo. Comparative pharmacokinetic studies should assess the biodistribution of PDEVs from different plant sources (eg, grapefruit vs ginger) and compare them with synthetic nanocarriers like PEGylated liposomes. Surface engineering strategies, such as PEGylation or the use of CD47 mimetic peptides, should also be explored to improve circulation time, modulate RES uptake, and enhance tumor targeting. In parallel, the field must urgently establish minimum reporting standards (MRS) for critical PDEV characterization and drug-loading data. This includes mandating the reporting of loading efficiency, encapsulation efficiency, drug-to-lipid ratios, and stability metrics, which will be vital for ensuring reproducibility, facilitating technology transfer, and building a credible data foundation for regulatory submissions. Alongside these efforts, the field must develop scalable production methods for clinical-grade PDEVs that comply with Good Manufacturing Practice (GMP) guidelines.
Ultimately, advancing PDEVs into clinical applications will require rigorous validation through human clinical trials to demonstrate their safety, efficacy, and tolerability. By addressing the gaps in pharmacokinetics, biodistribution, standardized data reporting, and regulatory standards, PDEVs could become a powerful tool in the treatment of complex diseases such as cancer. The potential for innovative combination therapies, leveraging the unique strengths of different PDEVs, offers a sophisticated approach to tackling diseases and paves the way for their integration into clinical practice.
Conclusion
PDEVs have emerged as a promising and revolutionary class of natural nanoparticles for cancer drug delivery. Recent research highlights their exceptional potential for efficient drug encapsulation, targeted delivery, and controlled release, which collectively contribute to improved therapeutic efficacy and reduced systemic toxicity. Beyond their role as sophisticated nanocarriers, PDEVs exhibit inherent “dual-functionality”, acting as both innate bioactive therapeutics-owing to their cargo of plant-specific metabolites-and efficient delivery vehicles. This unique combination, coupled with their cost-effectiveness and potential for scalable production from renewable biomass, positions them as a superior alternative to many synthetic nanoparticles and mammalian cell-derived EVs.
However, despite highly encouraging preclinical results, several significant challenges must be overcome to successfully translate PDEVs into clinical applications. These include achieving standardized large-scale production, ensuring long-term stability, addressing the inherent heterogeneity of native PDEVs, and implementing functional modifications to enhance targeting precision and therapeutic reproducibility.
To address these hurdles and unlock the full clinical potential of PDEVs, future research must converge on a multi-pronged strategy. Priority should be given to several key technological advances: the establishment of GMP-compliant, scalable biomanufacturing platforms, such as Temporary Immersion Bioreactor Systems (TIBS), to ensure quality and consistency; the refinement of surface engineering techniques, including bio-orthogonal chemistry, for precision targeting; and a mandatory focus on conducting robust, quantitative biodistribution and pharmacokinetic studies using advanced imaging to de-risk clinical development. Furthermore, a multidisciplinary approach that integrates insights from nanomedicine, molecular biology, and immunology is crucial. Strategic combination of PDEVs with other therapeutic modalities, such as immunotherapy (eg, checkpoint inhibitors) or gene editing tools, presents a highly promising avenue to achieve synergistic antitumor efficacy and develop more personalized treatment regimens.
Ultimately, PDEVs hold a unique position at the convergence of two transformative trends in modern medicine: personalized medicine and green nanomedicine. The vast biodiversity of plants provides an unparalleled toolkit for tailoring therapies, enabling the rational design of personalized PDEV “cocktails” from specific sources (eg, ginger for anti-inflammation, grapefruit for delivery) matched to a patient’s disease profile. Simultaneously, their production from edible plants, coupled with inherent biodegradability and low toxicity, establishes PDEVs as an environmentally sustainable and biocompatible platform, aligning with the principles of green chemistry.
In conclusion, while still in the early stages of clinical application, PDEVs hold immense potential to revolutionize oncology. With continued advancements in their development, optimization, and a dedicated focus on overcoming translational challenges, PDEVs are poised to become a cornerstone of next-generation, sustainable, and personalized cancer therapeutics, paving the way for a new era of safer and more effective cancer treatments.
Acknowledgments
This work was supported in part by the National Natural Science Foundation of China (Grant No. 81703819 and 82074122), and Research Foundation of Education Bureau of Hunan Province (Grant No. 23A0280), and the Postgraduate Innovation Project of Hunan University of Chinese Medicine (Grant No. 2024CX087), and Key Research and Development Programs of Hunan Science and Technology Department (2023SK2046), and the Natural Science Foundation of Hunan Province (Grant No. 2024JJ8128).
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Tang PM-K, Zhang DM. Advance in targeted cancer therapy and mechanisms of resistance. Int J Mol Sci. 2023;24(23):16584. doi:10.3390/ijms242316584
2. Sarmento-Ribeiro AB, Scorilas A, Gonçalves AC, et al. The emergence of drug resistance to targeted cancer therapies: clinical evidence. Drug Resist Updat. 2019;47:100646. doi:10.1016/j.drup.2019.100646
3. Garg P, Malhotra J, Kulkarni P, et al. Emerging therapeutic strategies to overcome drug resistance in cancer cells. Cancers. 2024;16(13):2478. doi:10.3390/cancers16132478
4. Jin HJ, Wang LQ, Bernards R. Rational combinations of targeted cancer therapies: background, advances and challenges. Nat Rev Drug Discov. 2023;22:213–234.
5. Passirani C, Vessières A, Regina GL, et al. Modulating undruggable targets to overcome cancer therapy resistance. Drug Resist Updat. 2022;60:100788. doi:10.1016/j.drup.2021.100788
6. Aldea M, Andre F, Marabelle A, et al. Overcoming resistance to tumor-targeted and immune-targeted therapies. Cancer Discov. 2021;11(4):874–899. doi:10.1158/2159-8290.CD-20-1638
7. Chen C, Di Bartolomeo M, Corallo S, et al. Overcoming resistance to targeted therapies in gastrointestinal cancers: progress to date and progress to come. ASCO Educ Book. 2020;40:280871.
8. Liu Y, Yang GZ, Hui Y, et al. Microfluidic nanoparticles for drug delivery. Small. 2022;18(36):6580.
9. Beach MA, Nayanathara U, Gao YT, et al. Polymeric nanoparticles for drug delivery. Chem Rev. 2024;124(9):5505–5616. doi:10.1021/acs.chemrev.3c00705
10. Gutierrez-Millan C, Díaz CC, Lanao JM, et al. Advances in exosomes‐based drug delivery systems. Macromol Biosci. 2020;21(1):269.
11. Chu K, Liu J, Zhang X, et al. Herbal medicine-derived exosome-like nanovesicles: a rising star in cancer therapy. Int J Nanomed. 2024;19:7585–7603. doi:10.2147/IJN.S477270
12. Jiang XC, Gao JQ. Exosomes as novel bio-carriers for gene and drug delivery. Int J Pharm. 2017;521(1–2):167–175. doi:10.1016/j.ijpharm.2017.02.038
13. Witwer KW, Wolfram J. Extracellular vesicles versus synthetic nanoparticles for drug delivery. Nat Rev Mater. 2021;6(2):103–106. doi:10.1038/s41578-020-00277-6
14. Liu H, Luo GF, Shang Z. Plant-derived nanovesicles as an emerging platform for cancer therapy. Acta Pharm Sin B. 2024;14(1):133–154. doi:10.1016/j.apsb.2023.08.033
15. Wang L, Yu X, Zhou J, et al. Extracellular vesicles for drug delivery in cancer treatment. Biol Proced Online. 2023;25(1):28. doi:10.1186/s12575-023-00220-3
16. Yao CL, Wang C. Platelet-derived extracellular vesicles for drug delivery. Biomater Sci. 2023;11(17):5758–5768. doi:10.1039/D3BM00893B
17. Du S, Guan YC, Xie AH, et al. Extracellular vesicles: a rising star for therapeutics and drug delivery. J Nanobiotechnol. 2023;21:231.
18. Sun M, Zhang H, Liu JY, et al. Extracellular vesicles: a new star for gene drug delivery. Int J Nanomed. 2024;19:2241–2264. doi:10.2147/IJN.S446224
19. Sharma S, Masud MK, Kaneti YV, et al. Extracellular vesicle nanoarchitectonics for novel drug delivery applications. Small. 2021;17(42):2220. doi:10.1002/smll.202102220
20. Palazzolo S, Memeo L, Hadla M, et al. Cancer extracellular vesicles: next-generation diagnostic and drug delivery nanotools. Cancers. 2020;12(11):3165. doi:10.3390/cancers12113165
21. Wang J, Yin BH, Lian JB, et al. Extracellular vesicles as drug delivery system for cancer therapy. Pharmaceutics. 2024;16(8):1029. doi:10.3390/pharmaceutics16081029
22. Guo YY, Hu DS, Lian L, et al. Stem cell-derived extracellular vesicles: a promising nano delivery platform to the brain. Stem Cell Rev Rep. 2023;19(2):285–308. doi:10.1007/s12015-022-10455-4
23. Pauwels MJ, Vandendriessche C, Vandenbroucke RE. Special delEVery: extracellular vesicles as promising delivery platform to the brain. Biomedicines. 2021;9(11):1734. doi:10.3390/biomedicines9111734
24. Li JL, Zhang HW, Jiang YY, et al. The landscape of extracellular vesicles combined with intranasal delivery towards brain diseases. Nano Today. 2024;55:102169. doi:10.1016/j.nantod.2024.102169
25. Zhou LN, Kodidela S, Godse S, et al. Targeted drug delivery to the central nervous system using extracellular vesicles. Pharmaceuticals. 2022;15(3):358. doi:10.3390/ph15030358
26. Shahjin F, Chand S, Yelamanchili SV. Extracellular vesicles as drug delivery vehicles to the central nervous system. J Neuroimmune Pharmacol. 2020;15(3):443–458. doi:10.1007/s11481-019-09875-w
27. Qiang W, Li J, Ruan R, et al. Plant-derived extracellular vesicles as a promising anti-tumor approach: a comprehensive assessment of effectiveness, safety, and mechanisms. Phytomedicine. 2024;130:155750. doi:10.1016/j.phymed.2024.155750
28. Zhu Y, Zhao J, Ding H, et al. Applications of plant‐derived extracellular vesicles in medicine. MedComm. 2024;5(10):e741. doi:10.1002/mco2.741
29. Wang YC, Wang JF, Ma JQ, et al. Focusing on future applications and current challenges of plant derived extracellular vesicles. Pharmaceuticals. 2022;15(6):708. doi:10.3390/ph15060708
30. Wang XX, Xin CL, Zhou Y, et al. Plant-derived vesicle-like nanoparticles: the next-generation drug delivery nanoplatforms. Pharmaceutics. 2024;16(5):588. doi:10.3390/pharmaceutics16050588
31. Lo KJ, Wang MH, Ho CT, et al. Plant-derived extracellular vesicles: a new revolutionization of modern healthy diets and biomedical applications. J Agric Food Chem. 2024;72(6):2853–2878. doi:10.1021/acs.jafc.3c06867
32. Rome S. Biological properties of plant-derived extracellular vesicles. Food Funct. 2019;10(2):529–538. doi:10.1039/C8FO02295J
33. Zhang JY, Tian S, Guo L, et al. Chinese herbal medicine-derived extracellular vesicles as novel biotherapeutic tools: present and future. J Transl Med. 2024;22(1):1059. doi:10.1186/s12967-024-05892-3
34. Wu PJ, Wu WJ, Zhang S, et al. Therapeutic potential and pharmacological significance of extracellular vesicles derived from traditional medicinal plants. Front Pharmacol. 2023;14:1272241. doi:10.3389/fphar.2023.1272241
35. Edis Z, Wang JL, Waqas MK, et al. Nanocarriers-mediated drug delivery systems for anticancer agents: an overview and perspectives. Int J Nanomed. 2021;16:5099–5100. doi:10.2147/IJN.S330286
36. Alqosaibi AI. Nanocarriers for anticancer drugs: challenges and perspectives. Saudi J Biol Sci. 2022;29(6):103298. doi:10.1016/j.sjbs.2022.103298
37. Kenchegowda M, Rahamathulla M, Hani U, et al. Smart nanocarriers as an emerging platform for cancer therapy: a review. Molecules. 2022;27(1):146. doi:10.3390/molecules27010146
38. Kumar A, Mishra A. Nanocarrier-mediated delivery targeting for pancreatic cancer. Acad Press. 2024;9:233–255.
39. Dutta B, Barick KC, Hassan PA. Recent advances in active targeting of nanomaterials for anticancer drug delivery. Adv Colloid Interface Sci. 2021;296:102509. doi:10.1016/j.cis.2021.102509
40. Arafat M, Sakkal M, Beiram R, et al. Nanomedicines: emerging platforms in smart chemotherapy treatment-a recent review. Pharmaceuticals. 2024;17(3):315. doi:10.3390/ph17030315
41. Prakash S. Nano-based drug delivery system for therapeutics: a comprehensive review. Biomed Phys Eng Express. 2023;9(5):2002. doi:10.1088/2057-1976/acedb2
42. Kumar G, Virmani T, Sharma A, et al. Codelivery of phytochemicals with conventional anticancer drugs in form of nanocarriers. Pharmaceutics. 2023;15(3):889. doi:10.3390/pharmaceutics15030889
43. Liang Y, Liu ZY, Wang PY, et al. Nanoplatform-based natural products co-delivery system to surmount cancer multidrug-resistant. J Control Release. 2021;336:396–409. doi:10.1016/j.jconrel.2021.06.034
44. Cui LS, Perini G, Palmieri V, et al. Plant-derived extracellular vesicles as a novel frontier in cancer therapeutics. Nanomaterials. 2024;14(16):1331. doi:10.3390/nano14161331
45. Zhu Y, Zhou XN, Yao Z. A mini-review: advances in plant-derived extracellular vesicles as nano-delivery systems for tumour therapy. Front Bioeng Biotech. 2022;10:1076348. doi:10.3389/fbioe.2022.1076348
46. Alfieri M, Leone A, Ambrosone A. Plant-derived nano and microvesicles for human health and therapeutic potential in nanomedicine. Pharmaceutics. 2021;13(4):498. doi:10.3390/pharmaceutics13040498
47. Song H, Liu B, Dong B, et al. Exosome-based delivery of natural products in cancer therapy. Front Cell Dev Biol. 2021;9:650426. doi:10.3389/fcell.2021.650426
48. Wang RN, Zhang YJ, Guo YM, et al. Plant-derived nanovesicles: promising therapeutics and drug delivery nanoplatforms for brain disorders. Fund Res. 2023;9:7.
49. Ren LL, Zhang DM, Pang L, et al. Extracellular vesicles for cancer therapy: potential, progress, and clinical challenges. Front Bioeng Biotech. 2024;12:1476737. doi:10.3389/fbioe.2024.1476737
50. Samad AFA, Sajad M, Nazaruddin N, et al. MicroRNA and transcription factor: key players in plant regulatory network. Front Plant Sci. 2017;8:565. doi:10.3389/fpls.2017.00565
51. Xiao J, Feng SY, Wang X, et al. Identification of exosome-like nanoparticle-derived microRNAs from 11 edible fruits and vegetables. PeerJ. 2018;6:e5186. doi:10.7717/peerj.5186
52. Kim K, Yoo HJ, Jung JH, et al. Cytotoxic effects of plant sap derived extracellular vesicles on various tumor cell types. J Funct Biomater. 2020;11(2):22. doi:10.3390/jfb11020022
53. Cambier L, Stachelek K, Triska M, et al. Extracellular vesicle associated repetitive element DNAs as candidate osteosarcoma biomarkers. Sci Rep. 2021;11(1):94. doi:10.1038/s41598-020-77398-z
54. Clancy JW, Sheehan CS, Boomgarden AC, et al. Recruitment of DNA to tumor-derived microvesicles. Cell Rep. 2022;38(9):110443. doi:10.1016/j.celrep.2022.110443
55. Mathivanan S, Fahner CJ, Reid GE, et al. ExoCarta 2012: database of exosomal proteins, RNA and lipids. Nucleic Acids Res. 2012;40(D1):D1241–D1244. doi:10.1093/nar/gkr828
56. Théry C, Zitvogel L, Amigorena S. Exosomes: composition, biogenesis and function. Nat Rev Immunol. 2002;2(8):569–579. doi:10.1038/nri855
57. Jeppesen DK, Fenix AM, Franklin JL, et al. Reassessment of exosome composition. Cell. 2019;177(2):428–445. doi:10.1016/j.cell.2019.02.029
58. Hao SY, Yang HY, Hu JJ, et al. Bioactive compounds and biological functions of medicinal plant-derived extracellular vesicles. Pharmacol Res. 2024;200:107062. doi:10.1016/j.phrs.2024.107062
59. Zhang MZ, Viennois E, Prasad M, et al. Edible ginger-derived nanoparticles: a novel therapeutic approach for the prevention and treatment of inflammatory bowel disease and colitis-associated cancer. Biomaterials. 2016;101:321–340. doi:10.1016/j.biomaterials.2016.06.018
60. Sundaram K, Miller DP, Kumar A, et al. Plant-derived exosomal nanoparticles inhibit pathogenicity of porphyromonas gingivali. Iscience. 2019;21:308–327. doi:10.1016/j.isci.2019.10.032
61. Karamanidou T, Tsouknidas A. Plant-derived extracellular vesicles as therapeutic nanocarriers. Int J Mol Sci. 2022;23(1):191. doi:10.3390/ijms23010191
62. Lian MQ, Chng WH, Liang J, et al. Plant‐derived extracellular vesicles: recent advancements and current challenges on their use for biomedical applications. J Extracell Vesicles. 2022;11(12):12283. doi:10.1002/jev2.12283
63. Liu Y, Wu S, Koo Y, et al. Characterization of and isolation methods for plant leaf nanovesicles and small extracellular vesicles. Nanomed Nanotech Biol Med. 2020;29:102271. doi:10.1016/j.nano.2020.102271
64. Xu ZJ, Zeng SS, Gong ZC, et al. Exosome-based immunotherapy: a promising approach for cancer treatment. Mol Cancer. 2020;19(1):160. doi:10.1186/s12943-020-01278-3
65. El Andaloussi S, Mäger I, Breakefield XO, et al. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov. 2013;12(5):347–357. doi:10.1038/nrd3978
66. Kalra H, Drummen G, Mathivanan S. Focus on extracellular vesicles: introducing the next small big thing. Int J Mol Sci. 2016;17(2):170. doi:10.3390/ijms17020170
67. Bai C, Liu JR, Zhang XM, et al. Research status and challenges of plant-derived exosome-like nanoparticles. Biomed Pharmacother. 2024;174:116543. doi:10.1016/j.biopha.2024.116543
68. Zeng YB, Deng X, Shen LS, et al. Advances in plant-derived extracellular vesicles: isolation, composition, and biological functions. Food Funct. 2024;15(23):11319–11341. doi:10.1039/D4FO04321A
69. Li P, Kaslan M, Lee SH, et al. Progress in exosome isolation techniques. Theranostics. 2017;7(3):789–804. doi:10.7150/thno.18133
70. Hadizadeh N, Bagheri D, Shamsara M, et al. Extracellular vesicles biogenesis, isolation, manipulation and genetic engineering for potential in vitro and in vivo therapeutics: an overview. Front Bioeng Biotech. 2022;10:19821. doi:10.3389/fbioe.2022.1019821
71. Livshts MA, Khomyakova E, Evtushenko EG, et al. Isolation of exosomes by differential centrifugation: theoretical analysis of a commonly used protocol. Sci Rep. 2015;5(1):17319. doi:10.1038/srep17319
72. Northrop-Albrecht EJ, Taylor WR, Huang BQ, et al. Assessment of extracellular vesicle isolation methods from human stool supernatant. J Extracell Vesicles. 2022;11(4):e12208. doi:10.1002/jev2.12208
73. Doyle LM, Wang MZ. Overview of extracellular vesicles, their origin, composition, purpose, and methods for exosome isolation and analysis. Cells. 2019;8(7):727. doi:10.3390/cells8070727
74. Böing AN, Pol EVD, Grootemaat AE, et al. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J Extracell Vesicles. 2014;3(1):23430. doi:10.3402/jev.v3.23430
75. Stranska R, Gysbrechts L, Wouters J, et al. Comparison of membrane affinity-based method with size-exclusion chromatography for isolation of exosome like vesicles from human plasma. J Transl Med. 2018;16(1):1. doi:10.1186/s12967-017-1374-6
76. Shtam TA, Samsonov RB, Volnitskiy AV, et al. Isolation of extracellular microvesicles from cell culture medium: comparative evaluation of methods. Biochem Moscow Suppl Ser B. 2018;12(2):167–175. doi:10.1134/S1990750818020117
77. Huang YF, Wang S, Cai Q, et al. Effective methods for isolation and purification of extracellular vesicles from plants. J Integr Plant Biol. 2021;63(12):2020–2030. doi:10.1111/jipb.13181
78. Zarovni N, Corrado A, Guazzi P, et al. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using immunocapture approaches. Methods. 2015;87:46–58. doi:10.1016/j.ymeth.2015.05.028
79. Konoshenko MY, Lekchnov EA, Vlassov AV, et al. Isolation of extracellular vesicles: general methodologies and latest trends. BioMed Res Int. 2018;27:8545347.
80. Coumans FAW, Brisson AR, Buzas EI, et al. Methodological guidelines to study extracellular vesicles. Circ Res. 2017;120(10):10. doi:10.1161/CIRCRESAHA.117.309417
81. Nelemans LC, Gurevich L. Drug delivery with polymeric nanocarriers-cellular uptake mechanisms. Materials. 2020;13(2):366. doi:10.3390/ma13020366
82. Adamo G, Fierli D, Romancino DP, et al. Nanoalgosomes: introducing extracellular vesicles produced by microalgae. J Extracell Vesicles. 2021;10(6):e12081. doi:10.1002/jev2.12081
83. Bian DH, Wu Y, Song GD, et al. The application of mesenchymal stromal cells (MSCs) and their derivative exosome in skin wound healing: a comprehensive review. Stem Cell Res Ther. 2022;13(1):24. doi:10.1186/s13287-021-02697-9
84. Shokravi S, Borisov V, Zaman BA, et al. Mesenchymal stromal cells (MSCs) and their exosome in acute liver failure (ALF): a comprehensive review. Stem Cell Res Ther. 2022;13(1):192. doi:10.1186/s13287-022-02825-z
85. Salmerón-Manzano E, Garrido-Cardenas JA, Manzano-Agugliaro F. Worldwide research trends on medicinal plants. Int J Environ Res Public Health. 2020;17(10):3376. doi:10.3390/ijerph17103376
86. Xu XH, Yuan TJ, Dad HA, et al. Plant exosomes as novel nanoplatforms for microRNA transfer stimulate neural differentiation of stem cells in vitro and in vivo. Nano Lett. 2021;21(19):8151–8159. doi:10.1021/acs.nanolett.1c02530
87. Teng Y, Ren Y, Sayed M, et al. Plant-derived exosomal microRNAs shape the gut microbiota. Cell Host Microbe. 2018;24(5):637–652. doi:10.1016/j.chom.2018.10.001
88. Ju SW, Mu JY, Dokland T, et al. Grape exosome-like nanoparticles induce intestinal stem cells and protect mice from DSS-induced colitis. Mol Ther. 2013;21(7):1345–1357. doi:10.1038/mt.2013.64
89. Zhou LK, Zhou Z, Jiang XM, et al. Absorbed plant MIR2911 in honeysuckle decoction inhibits SARS-CoV-2 replication and accelerates the negative conversion of infected patient. Cell Discov. 2020;6(1):54. doi:10.1038/s41421-020-00197-3
90. Langellotto MD, Rassu G, Serri C, et al. Plant-derived extracellular vesicles: a synergetic combination of a drug delivery system and a source of natural bioactive compounds. Drug Delivery Trans Res. 2025;15(3):831–845. doi:10.1007/s13346-024-01698-4
91. Didiot MC, Hall LM, Coles AH, et al. Exosome-mediated delivery of hydrophobically modified siRNA for huntingtin mRNA silencing. Mol Ther. 2016;24(10):1836–1847. doi:10.1038/mt.2016.126
92. Mccann J, Sosa-Miranda CD, Guo HS, et al. Contaminating transfection complexes can masquerade as small extracellular vesicles and impair their delivery of RNA. J Extracell Vesicles. 2022;11(10):e12220. doi:10.1002/jev2.12220
93. Mcconnell RE, Youniss M, Gnanasambandam B, et al. Transfection reagent artefact likely accounts for some reports of extracellular vesicle function. J Extracell Vesicles. 2022;11(10):e12253. doi:10.1002/jev2.12253
94. Kimiz-Gebologlu I, Oncel SS. Exosomes: large-scale production, isolation, drug loading efficiency, and biodistribution and uptake. J Control Release. 2022;347:533–543. doi:10.1016/j.jconrel.2022.05.027
95. Ji XY, Ge LL, Liu C, et al. Capturing functional two-dimensional nanosheets from sandwich-structure vermiculite for cancer theranostics. Nat Commun. 2021;12(1):1124. doi:10.1038/s41467-021-21436-5
96. Yang Z, Gao D, Zhao J, et al. Thermal immuno-nanomedicine in cancer. Nat Rev Clin Oncol. 2023;20(2):116–134. doi:10.1038/s41571-022-00717-y
97. Hülsebusch L, Heyn TR, Amft J, et al. Extrusion of plant proteins: a review of lipid and protein oxidation and their impact on functional properties. Food Chem. 2025;470:142607. doi:10.1016/j.foodchem.2024.142607
98. Zhang YF, Shi JB, Li C, et al. Small extracellular vesicle loading systems in cancer therapy: current status and the way forward. Cytotherapy. 2019;21(11):1122–1136. doi:10.1016/j.jcyt.2019.10.002
99. Luan X, Sansanaphongpricha K, Myers I, et al. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol Sin. 2017;38(6):754–763. doi:10.1038/aps.2017.12
100. Sung JJY, Lau JYW, Goh KL, et al. Increasing incidence of colorectal cancer in Asia: implications for screening. Lancet Oncol. 2005;6(11):871–876. doi:10.1016/S1470-2045(05)70422-8
101. Gramont AD, Cutsem EV, Schmoll HJ, et al. Bevacizumab plus oxaliplatin-based chemotherapy as adjuvant treatment for colon cancer (AVANT): a Phase 3 randomised controlled trial. Lancet. 2012;13(12):1225–1233. doi:10.1016/S1470-2045(12)70509-0
102. You JY, Kang SJ, Rhee WJ. Isolation of cabbage exosome-like nanovesicles and investigation of their biological activities in human cells. Bioact Mater. 2021;6(12):4321–4332. doi:10.1016/j.bioactmat.2021.04.023
103. Wang BM, Zhuang XY, Deng ZB, et al. Targeted drug delivery to intestinal macrophages by bioactive nanovesicles released from grapefruit. Mol Ther. 2016;22(3):522–534. doi:10.1038/mt.2013.190
104. Zhang MZ, Xiao B, Wang H, et al. Edible ginger-derived nano-lipids loaded with doxorubicin as a novel drug-delivery approach for colon cancer therapy. Mol Ther. 2016;24(10):1783–1796. doi:10.1038/mt.2016.159
105. Zeng LP, Shi WH, Wang HY, et al. Codelivery of π-π stacked dual anticancer drugs based on aloe-derived nanovesicles for breast cancer therapy. ACS Appl Mater Interfaces. 2022;14(24):27686–27702. doi:10.1021/acsami.2c06546
106. Farheen J, Iqbal MZ, Lu Y, et al. Vitis vinifera kyoho-derived exosome-like nanoparticles-based drug delivery and therapeutic modalities for breast cancer therapy. J Drug Deliv Sci Technol. 2024;92:105332. doi:10.1016/j.jddst.2023.105332
107. Mammadova R, Maggio S, Fiume I, et al. Protein biocargo and anti-inflammatory effect of tomato fruit-derived nanovesicles separated by density gradient ultracentrifugation and loaded with curcumin. Pharmaceutics. 2023;15(2):333. doi:10.3390/pharmaceutics15020333
108. Sarvarian P, Samadi P, Gholipour E, et al. Fisetin-loaded grape-derived nanoparticles improve anticancer efficacy in MOLT-4 cells. Biochem Biophys Res Commun. 2023;658:69–79. doi:10.1016/j.bbrc.2023.03.040
109. Xiao Q, Zhao W, Wu C, et al. Lemon‐derived extracellular vesicles nanodrugs enable to efficiently overcome cancer multidrug resistance by endocytosis-triggered energy dissipation and energy production reduction. Adv Sci. 2022;9(20):2105274. doi:10.1002/advs.202105274
110. Lu X, Han Q, Chen J, et al. Celery (Apium graveolens L.) exosome-like nanovesicles as a new-generation chemotherapy drug delivery platform against tumor proliferation. J Agric Food Chem. 2023;71(22):8413–8424. doi:10.1021/acs.jafc.2c07760
111. Zeng L, Wang H, Shi W, et al. Aloe derived nanovesicle as a functional carrier for indocyanine green encapsulation and phototherapy. J Nanobiotechnol. 2021;19(1):1–24. doi:10.1186/s12951-021-01195-7
112. Moon K, Hur J, Kim KP, et al. Surface-functionalizable plant-derived extracellular vesicles for targeted drug delivery carrier using grapefruit. Adv Mater Interfaces. 2023;10(22):2300220. doi:10.1002/admi.202300220
113. Yang M, Luo Q, Chen X, et al. Bitter melon derived extracellular vesicles enhance the therapeutic effects and reduce the drug resistance of 5-fluorouracil on oral squamous cell carcinoma. J Nanobiotechnol. 2021;19(1):259. doi:10.1186/s12951-021-00995-1
114. Jiang D, Li Z, Liu H, et al. Plant exosome-like nanovesicles derived from sesame leaves as carriers for luteolin delivery: molecular docking, stability and bioactivity. Food Chem. 2024;438:137963. doi:10.1016/j.foodchem.2023.137963
115. Feng W, Teng Y, Zhong Q, et al. Biomimetic grapefruit-derived extracellular vesicles for safe and targeted delivery of sodium thiosulfate against vascular calcification. ACS nano. 2023;17(24):24773–24789. doi:10.1021/acsnano.3c05261
116. Yu WQ, Hu C, Gao HL. Advances of nanomedicines in breast cancer metastasis treatment targeting different metastatic stages. Adv Drug Deliv Rev. 2021;178:113909. doi:10.1016/j.addr.2021.113909
117. Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331(6024):1559–1564. doi:10.1126/science.1203543
118. Minko T, Rodriguez-Rodriguez L, Pozharov V, et al. Nanotechnology approaches for personalized treatment of multidrug resistant cancers. Adv Drug Deliv Rev. 2013;65(13–14):1880–1895. doi:10.1016/j.addr.2013.09.017
119. Zhao Q, Wang T, Wang H, et al. Consensus statement on research and application of Chinese herbal medicine derived extracellular vesicles-like particles. Chin Herb Med. 2023;16(1):3–12. doi:10.1016/j.chmed.2023.11.002
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Bubble Ticket Trip: Exploring the Mechanism of miRNA Sorting into Exosomes and Maintaining the Stability of Tumor Microenvironment
Wang L, Liu H, Chen G, Wu Q, Xu S, Zhou Q, Zhao Y, Wang Q, Yan T, Cheng X
International Journal of Nanomedicine 2024, 19:13671-13685
Published Date: 21 December 2024
Tumor Targeting with Peptide-Drug Conjugates: Showcasing Key Progress and Hurdles
Parang K, Do T, Dinh C, Davani-Davari D, Foroughi M, Khong T, Moazen M, Nasrolahi Shirazi A
Drug Design, Development and Therapy 2026, 20:562135
Published Date: 12 February 2026
Bioactive Flavonoids from Scutellaria baicalensis in Non-Small Cell Lung Cancer: Mechanistic Insights and Emerging Translational Perspectives
Yuan J, Wang Z
Drug Design, Development and Therapy 2026, 20:591381
Published Date: 8 April 2026
Recent Advances in Antitumor Nanomedicine Based on Covalent Organic Frameworks
Xu F, Wang R, Cheng Z, Zhang Z, Tian J
International Journal of Nanomedicine 2026, 21:378082
Published Date: 4 May 2026