Back to Journals » Drug Design, Development and Therapy » Volume 20
Bioactive Flavonoids from Scutellaria baicalensis in Non-Small Cell Lung Cancer: Mechanistic Insights and Emerging Translational Perspectives
Received 24 December 2025
Accepted for publication 28 March 2026
Published 8 April 2026 Volume 2026:20 591381
DOI https://doi.org/10.2147/DDDT.S591381
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Tuo Deng
Jiaqian Yuan,1 Zhen Wang2
1The First Clinical Medical College, Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China; 2Department of Respiratory and Critical Care Medicine, The First Affiliated Hospital of Zhejiang Chinese Medical University, Hangzhou, People’s Republic of China
Correspondence: Zhen Wang, Department of Respiratory and Critical Care Medicine, The First Affiliated Hospital of Zhejiang Chinese Medical University, No. 54, Youdian Road, Shangcheng District, Hangzhou, Zhejiang, People’s Republic of China, Email [email protected]
Abstract: Non-small cell lung cancer (NSCLC) remains a leading cause of cancer-related morbidity and mortality globally, and developing effective treatment strategies is a major challenge. In recent years, Scutellaria baicalensis Georgi has attracted increasing attention due to its multi-target pharmacological properties and relatively favorable safety profile. In this review, we systematically summarize the current literature from PubMed, Web of Science, and Embase, focusing on the pharmacokinetic characteristics, pharmacological mechanisms, and potential translational relevance of the bioactive constituents of S. baicalensis in NSCLC. The toxicity profiles of key flavonoids were also predicted using ADMETlab 2.0. Accumulating evidence, primarily derived from in vitro and in vivo preclinical studies, suggests that the flavonoid constituents of S. baicalensis exert anti-NSCLC effects across multiple stages of tumor development. These effects include early modulation of inflammation and oxidative stress, inhibition of tumor proliferation, epigenetic regulation, and suppression of metastasis and therapeutic resistance. The underlying mechanisms involve processes such as cell-cycle arrest, apoptosis, autophagy, ferroptosis, and remodeling of the tumor immune microenvironment. Overall, these compounds generally exhibit low systemic toxicity, although long-term or high-dose exposure may be associated with mild fluctuations in liver enzymes and lipid metabolism. Furthermore, nanocarrier-based delivery systems have been shown to improve bioavailability and tumor-targeting efficiency. Despite these promising findings, current evidence remains largely preclinical, and clinical validation is still limited. Therefore, further well-designed clinical studies and advanced drug-delivery strategies are required to facilitate the translation of S. baicalensis-derived compounds into clinical applications for NSCLC. Flowchart of Scutellaria baicalensis in pharmacokinetics, safety and anti-NSCLC mechanisms.It starts with Scutellaria baicalensis, showing its chemical structures. An arrow leads to Pharmacokinetics, depicting organs like the liver and kidneys and pills. From Pharmacokinetics, an arrow points to Anti-NSCLC Mechanisms, showing interactions with elements like Baicalein and pathways such as PI3K/AKT and mTOR. Another arrow connects Anti-NSCLC Mechanisms to Safety and Advanced Drug Delivery Systems, featuring lungs, a syringe and a hazard symbol. An arrow from Safety and Advanced Drug Delivery Systems loops back to Scutellaria baicalensis, completing the cycle.
Keywords: baicalin, baicalein, cancer, pharmacological mechanisms, drug delivery
Introduction
Over the years, lung cancer has remained one of the leading causes of cancer-related morbidity and mortality worldwide, with non-small cell lung cancer (NSCLC) accounting for approximately 85% of all lung cancer cases and posing a serious threat to human health and survival.1,2 In recent years, advances in modern medical approaches, such as surgery, radiotherapy, chemotherapy, molecular targeted therapy, and immunotherapy, have profoundly changed the treatment landscape for patients with advanced or locally advanced NSCLC. The introduction of immune checkpoint inhibitors (ICIs) and targeted agents has led to significant survival benefits in a subset of patients. Nevertheless, the overall five-year survival rate of NSCLC remains low, at around 17.8%, with more than half of patients dying within the first year after diagnosis.3 Drug resistance, adverse effects, tumor recurrence, and metastasis continue to be major clinical challenges.3,4 Consequently, developing novel, safe, and effective therapeutic strategies has become a crucial direction in NSCLC research.
In China, traditional Chinese medicine (TCM) has been employed for thousands of years for cancer prevention and treatment due to its multitarget characteristics, holistic regulation, mild toxicity, and cost-effectiveness. Increasing evidence suggests that TCM plays a unique role in improving quality of life, alleviating symptoms, and delaying tumor progression, particularly in patients who are intolerant to conventional chemotherapeutic regimens.5 Among these herbal medicines, Scutellaria baicalensis Georgi (Huangqin, S. baicalensis), a classical TCM herb, has been widely used to treat inflammatory, infectious, and cancer-related diseases.6 In the context of lung cancer, its historical use provides a traditional rationale for further pharmacological investigation. Modern pharmacological studies have further demonstrated that S. baicalensis contains multiple bioactive constituents, including flavonoids, terpenoids, polysaccharides, phenylethyl glycosides, and others, among which flavonoids constitute the major active group and exhibit potent anticancer effects in various lung cancer cell lines and animal models.6,7 Compared with current targeted therapies, such as tyrosine kinase inhibitors, and immunotherapies, which often face primary or acquired resistance in the context of tumor heterogeneity, these bioactive flavonoids may offer a complementary pharmacological approach because of their multi-component and multi-target characteristics. They exert multidimensional pharmacological activities by modulating key signaling pathways such as PI3K/Akt, MAPK, and NF-κB, thereby inhibiting tumor cell proliferation and invasion, inducing apoptosis and autophagy, and regulating the tumor microenvironment (TME).6,8 However, the currently available evidence remains largely preclinical, and many reported anticancer effects are based on in vitro or animal studies that may not fully translate into clinical efficacy in humans.
In recent years, the therapeutic potential of natural products in oncology has attracted increasing global attention, and research on S. baicalensis in NSCLC prevention and treatment has expanded accordingly. Accumulating studies suggest that its active constituents act through multiple molecular targets and pathways to suppress tumor progression, while flavonoids derived from S. baicalensis are not currently alternatives to standard-of-care therapies. Their further development is limited by poor oral bioavailability, rapid metabolism, and insufficient systemic exposure, highlighting the importance of drug delivery strategies for improving their translational potential. Nevertheless, existing studies are often limited to individual compounds or isolated mechanisms. In addition, current reviews lack a systematic and up-to-date integration of S. baicalensis flavonoids research in NSCLC. To the best of our knowledge, few reviews have simultaneously summarized both pharmacological mechanisms and clinical translation, particularly from the perspective of emerging drug delivery strategies. Therefore, this review aims to comprehensively summarize the research advances over the past two decades regarding the anti-NSCLC effects of S. baicalensis, with a focus on its active constituents, pharmacokinetic characteristics, multidimensional pharmacological mechanisms, and emerging translational perspectives. By providing an integrated overview, this work aims to furnish theoretical support and strategic guidance for future studies and the development of related anticancer agents.
Materials and Methods
This study systematically searched PubMed, Web of Science, and Embase for relevant publications from October 30, 2005, to October 30, 2025. The following search terms were used: “Scutellaria baicalensis”, “baicalin”, “baicalein”, “wogonin”, “wogonoside”, “oroxyloside”, “norwogonoside”, “oroxylin A”, “norwogonin”, “lung cancer”, “non-small cell lung cancer”, and “NSCLC”. To provide a supplementary and standardized assessment of the safety-related properties of major flavonoids from S. baicalensis, in silico prediction was performed using ADMETlab 2.0 (https://admetmesh.scbdd.com/). The evaluated endpoints included hERG liability, human hepatotoxicity (H-HT), drug-induced liver injury (DILI), respiratory toxicity (RT), carcinogenicity, and predicted median lethal dose (LD50). The prediction results were interpreted according to the default classification outputs provided by the platform.
Active Constituents and Pharmacokinetics
S. baicalensis, a commonly used traditional Chinese medicinal herb, is rich in diverse bioactive constituents whose pharmacokinetic characteristics and bioavailability largely determine their efficacy and clinical potential.9 Hundreds of compounds have been isolated from S. baicalensis, including flavonoids, terpenoids, volatile oils, polysaccharides, phenylethyl glycosides, amino acids, sterols, starches, alkaloids, organic acids, and trace elements.9,10 Among these, flavonoids represent the principal bioactive group, mainly including baicalin, baicalein, wogonin, wogonoside, oroxyloside, norwogonoside, oroxylin A (OA), and norwogonin, of which the first three have been most extensively investigated.10 Notably, flavonoids with clearly reported anti-NSCLC potential include baicalin, baicalein, wogonin, wogonoside, and OA. Quantitative phytochemical studies further support the predominance of baicalin in S. baicalensis. In one HPLC-based analysis of samples collected from different regions of China, the contents of baicalin, wogonoside, baicalein, wogonin, and OA in dried roots ranged from 76.2–160 mg/g, 17.1–25 mg/g, 1.13–4.67 mg/g, 0.28–2.31 mg/g, and 0.202–2.06 mg/g, respectively, indicating marked differences in constituent abundance.11 Consistent with this, other HPLC and UPLC-MS/MS studies have reported that baicalin accounts for approximately 8–20% of the dry root weight of S. baicalensis, generally representing the largest proportion among the major flavonoids.12,13 By comparison, wogonoside is present at approximately 2.52%, whereas baicalein, wogonin, and OA occur at substantially lower levels.13 In contrast to flavonoids, terpenoids, volatile oils, phenylethyl glycosides, sterols, alkaloids, and organic acids are generally regarded as relatively minor constituents of S. baicalensis.10,14 The detailed constituents are categorized and summarized in Supplementary Table 1.
Current evidence indicates that after oral administration, baicalin is hydrolyzed by intestinal β-glucuronidase into baicalein, which is absorbed into systemic circulation in its free form and can subsequently be reconjugated to baicalin via UDP-glucuronosyltransferases (UGTs).14,15 Baicalin then undergoes further glucuronidation and sulfation in the liver and intestine, existing predominantly in conjugated forms within plasma. Elimination occurs mainly through biliary excretion after hepatic uptake and metabolism, while a smaller fraction is excreted in urine.14–16 The major processes involved in the absorption, metabolism, and excretion of baicalin and baicalein are summarized in Figure 1. Due to its poor aqueous solubility and limited membrane permeability, baicalin exhibits low oral bioavailability; however, formulation optimization or co-administration with other drugs can significantly improve its absorption.10 For instance, Coptis chinensis has been shown to significantly inhibit the intestinal metabolism of flavonoid components from S. baicalensis, leading to increased plasma levels of the glycosides, hence prolonging their duration of action and enhancing therapeutic efficacy.17 Notably, the gut microbiota plays an essential role in mediating the pharmacological activity of S. baicalensis, enhancing baicalin hydrolysis and baicalein absorption, thus serving as an important endogenous factor of its bioavailability.10,18 In contrast, baicalein, owing to its higher lipophilicity, is rapidly absorbed via passive diffusion across cell membranes, but remains subject to significant first-pass metabolism, which similarly limits its oral bioavailability.19 In a Phase I study, single oral doses of baicalein tablets (200–800 mg) in healthy subjects produced parent-compound Cmax values of only 280.44–845.20 ng/mL, and systemic exposure increased in a less than dose-proportional manner, suggesting limited and nonlinear oral exposure.20
 |
Figure 1 The schematic diagram of the absorption, metabolism and excretion of baicalein and baicalin. Notes: This figure summarizes the major pharmacokinetic processes of baicalin and baicalein. After oral administration, baicalin is initially hydrolyzed in the intestine to baicalein by β-glucuronidase. Baicalein can then be reconjugated to baicalin by UGTs. Subsequently, compounds undergo further glucuronidation and sulfation in the liver and intestine, may circulate in the systemic circulation mainly in conjugated forms, and are ultimately excreted through bile or urine. The gut microbiota plays an important role throughout this process. Blue arrows indicate the general direction of the reported metabolic or transport processes. The black dotted circle represents the entire gastrointestinal tract, including both the small and large intestines. |
Wogonin also exhibits unfavorable pharmacokinetic characteristics. Its profile is marked by rapid systemic clearance and poor oral absorption, with low absolute bioavailability attributed mainly to limited aqueous solubility and extensive metabolism; only about 21% is excreted as the parent compound.21 OA appears even more restricted in this regard, with an absolute bioavailability of less than 2%.22 After either intravenous or oral administration, OA is eliminated rapidly, primarily through fecal excretion.22 In the first-in-human phase I study of OA, the compound appeared rapidly in plasma, but AUC increased less than dose proportionally and only moderate accumulation was observed after repeated dosing, indicating that dose escalation may not directly translate into proportional systemic exposure.23 Other flavonoids from S. baicalensis display specific tissue distribution and metabolic patterns. However, studies on these compounds remain scarce and warrant further investigation. In terms of tissue distribution, Scutellaria flavonoids show selective accumulation in the liver, lungs, and kidneys, while exhibiting poor permeability across the blood-brain barrier.10,19
By contrast, pharmacokinetic studies of the non-flavonoid constituents of S. baicalensis remain relatively scarce. Some reports suggest that these components may contribute to antitumor activity either directly or indirectly through immunomodulatory and anti-inflammatory effects, but their exposure profiles and in vivo relevance remain insufficiently characterized.24 At present, the pharmacokinetic evidence is more robust for the major flavonoids than for these minor constituents.
Collectively, the complex pharmacokinetic behavior and inherently low bioavailability of S. baicalensis constituents represent a bottleneck limiting clinical translation. One key issue underlying this limitation is that the exposure conditions under which anticancer effects are observed in preclinical models may not directly correspond to those achievable in humans, particularly in view of rapid metabolism, limited systemic exposure, and the predominance of circulating conjugated forms. At the same time, these challenges also present opportunities for innovation in modern pharmaceutics and combination therapy. Future research integrating pharmacokinetic modeling, gut microbiota interaction studies, and advanced drug-delivery systems may help bridge the gap between experimental findings and clinical application, ultimately promoting the effective translation of S. baicalensis-derived therapeutics for NSCLC treatment.
Pharmacological Mechanisms
Early Intervention in NSCLC
The initiation of tumors is a complex process driven by multiple factors. Among them, a persistent inflammatory microenvironment plays a pivotal role by disturbing the balance of immune cells and inflammatory mediators (such as IL-6, TNF-α, and IL-1β), in turn creating favorable conditions for tumor formation.25 Meanwhile, internal and external insults induce excessive production of free radicals, triggering oxidative stress. The resulting reactive oxygen species (ROS) and lipid peroxides directly cause DNA strand breaks or base modifications; if the intrinsic repair mechanisms fail, these lesions can be fixed as gene mutations.26 Moreover, ROS can induce epigenetic drift and metabolic reprogramming, further disrupting genomic stability and lowering the health threshold of the tissue microenvironment.26,27 In addition, ROS activate transcription factors NF-κB and AP-1, upregulating pro-inflammatory cytokines and promoting chronic inflammation, thus forming a vicious cycle.26 Notably, accumulating preclinical evidence suggests that S. baicalensis and its major bioactive flavonoids possess the potential capacity to suppress inflammatory amplification loops and alleviate oxidative chain reactions. In particular, they have been reported to exert complementary and synergistic effects between the NF-κB/STAT3 inflammatory axis and the Nrf2-ARE antioxidant axis, which may contribute to anti-tumor activity during early stages of lung carcinogenesis.28,29
The molecular basis of this early intervention effect has been progressively elucidated. Baicalin, a representative flavone glycoside, has been reported to exhibit anti-inflammatory modulation in the early stage of NSCLC. One study suggested that baicalin upregulates the expression of SOCS1 (suppressor of cytokine signaling 1), suppressing the hyperactivation of the NF-κB/STAT3 signaling pathway and reducing the transcription and secretion of downstream pro-inflammatory cytokines such as TNF-α and IL-6, thus disrupting the inflammation-cancer vicious cycle.30 In parallel, baicalein appears to exert a state-dependent dual role in oxidative stress regulation during the initiation of lung cancer. In a benzo[a]pyrene (B[a]P)-induced experimental lung carcinogenesis mouse model, long-term administration of baicalein significantly reduced ROS and lipid peroxide (LPO) levels while restoring mitochondrial enzymatic antioxidant systems and non-enzymatic antioxidants.31 This process mitigated oxidative damage and maintained mitochondrial functional stability.31 These findings suggest that baicalein may enhance endogenous antioxidant defenses and alleviate oxidative stress during the precancerous stage. However, in established tumor environments, where cancer cells depend on elevated ROS for metabolic and signaling activity, baicalein has been shown in vitro to induce excessive ROS production, activate the AMPK signaling pathway, trigger mitochondrial-dependent apoptosis, and ultimately eliminate malignant cells selectively.32 This dynamic shift from an antioxidant to a pro-oxidant role reflects a biphasic, precision regulatory mechanism by which baicalein acts according to the cellular oxidative stress threshold.
In genetically driven lung cancer models, this mechanism has been further explored. In the KrasG12D/p53flox/flox mouse lung cancer model, baicalein markedly reduced the formation of pulmonary tumor nodules.33 Mechanistically, baicalein alleviates mitochondrial dysfunction, reverses mutation-induced metabolic disturbances, and decreases mitochondrial DNA (mtDNA) leakage.33 Moreover, baicalein has been reported to interact with cGAS (cyclic GMP-AMP synthase) protein, inducing a “liquid-to-solid phase transition”, which may suppress excessive activation of the cGAS-STING (stimulator of interferon genes) innate immune signaling axis. Consequently, the downstream STING-TBK1-IRF3 (STING- TANK-binding kinase 1- interferon regulatory factor 3) pathway and type I interferon response are inhibited, interrupting the inflammation-driven tumorigenic cascade.33 These findings provide a potential mechanistic link between inflammation-oxidation interplay and tumorigenesis, although further validation is required. In addition, baicalein has been shown to inhibit the carcinogen-metabolizing enzyme CYP1A1 (cytochrome P450 family 1 subfamily a member 1), induces detoxification enzyme expression, and downregulates NF-κB signaling, accordingly achieving dual protection through anti-inflammatory and metabolic regulation.34,35 Together, these effects may contribute to restoring mitochondrial metabolic homeostasis and cellular redox balance.
By contrast, wogonin has been reported to exert both pro-oxidative apoptosis-inducing and anti-inflammatory gene-regulatory effects during the early intervention of lung cancer. Studies have demonstrated that wogonin selectively elevates intracellular ROS levels in lung cancer cells, surpassing their oxidative stress threshold, thereby co-activating caspase-dependent apoptotic and autophagic pathways to induce cancer cell death, while exhibiting minimal cytotoxicity toward normal epithelial cells.36 Additionally, wogonin suppresses ERK (extracellular signal-regulated kinase) pathway-mediated c-Jun (cellular Jun proto-oncogene) expression and AP-1 (activator protein-1) transcriptional activity in A549 cells, significantly inhibiting PMA (phorbol 12-myristate 13-acetate)-induced COX-2 (cyclooxygenase-2) transcription and reducing inflammatory mediator production.37 These findings support a potential role for wogonin in modulating inflammation-related pathways, although current evidence is largely derived from in vitro studies.
In summary, the active constituents of S. baicalensis have been reported to exert multi-level regulatory effects in early NSCLC intervention, involving anti-inflammatory modulation, oxidation and pro-oxidation balance regulation, and mitochondrial homeostasis maintenance (Table 1). However, it should be noted that most of these findings are derived from preclinical models, and their relevance under clinically achievable exposure conditions remains to be established. Importantly, the role of antioxidant activity in mediating these effects remains incompletely understood and appears to be context-dependent. Notably, Grigalius et al reported that baicalein lacks the potent ortho-dihydroxy structure, resulting in moderate antioxidant activity, which shows a limited correlation with its anticancer properties.38 This observation suggests that antioxidant capacity alone may not fully account for the antitumor effects of these compounds. Moreover, the reliance on antioxidant-based mechanisms in cancer prevention remains debated. While reducing oxidative stress may protect normal cells during early carcinogenesis, excessive suppression of ROS could potentially interfere with apoptosis and other ROS-dependent tumor-suppressive processes.39 Therefore, the biological effects of these flavonoids are likely to involve a dynamic balance between antioxidant and pro-oxidant activities, rather than a unidirectional protective mechanism. Taken together, these findings highlight the need for a more nuanced interpretation of redox-related mechanisms, as well as further investigation into other pathways such as apoptosis induction, metabolic regulation, and signaling pathway modulation.
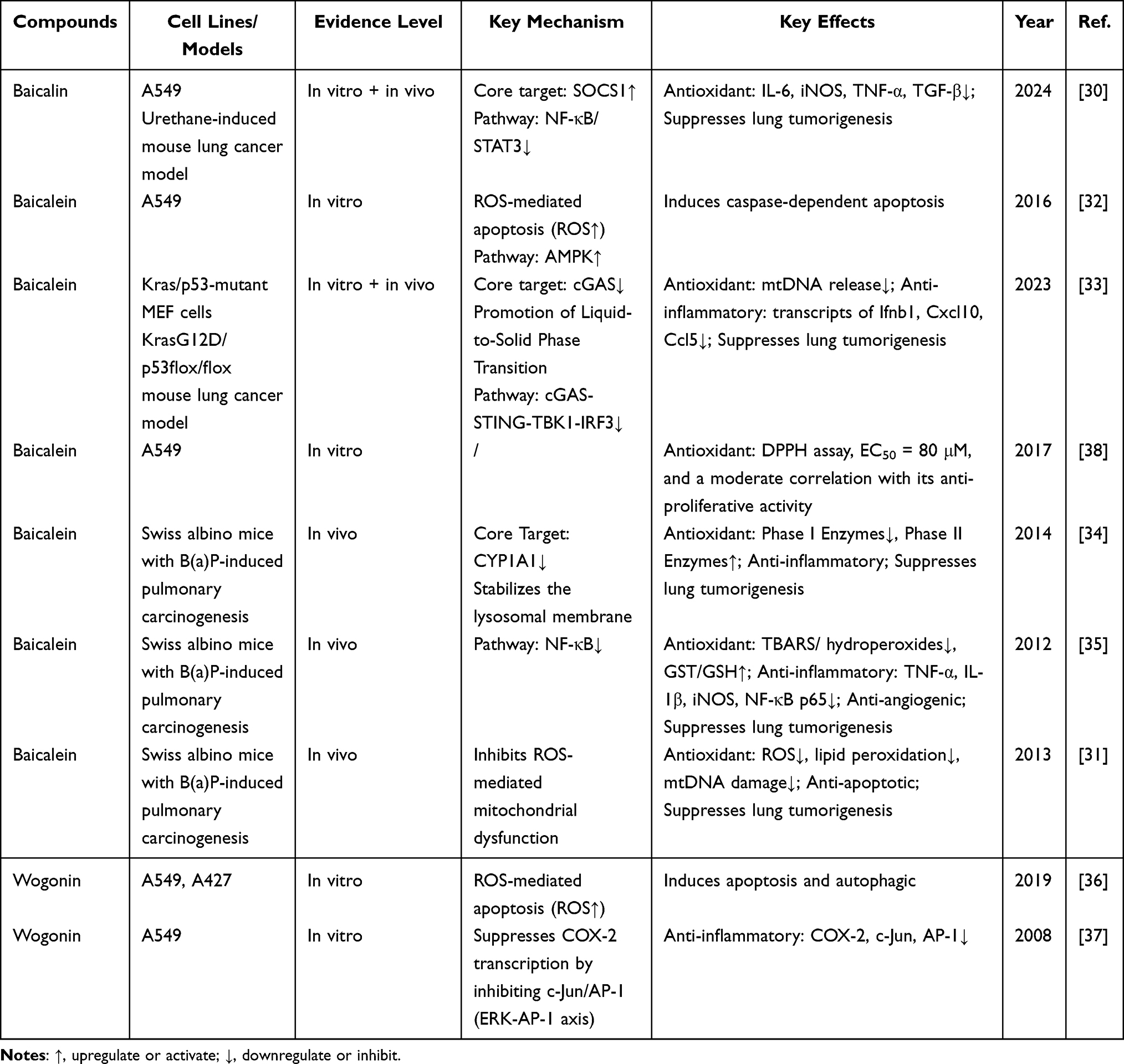 |
Table 1 Mechanistic Summary of Anti-Inflammatory and Anti/Pro-Oxidative Actions of Flavonoids in Early-Stage NSCLC Prevention |
Inhibition of Tumor Growth
The sustained growth of NSCLC is supported by persistent proliferative signaling, dysregulated cell-cycle progression, and evasion of programmed cell death. Current preclinical evidence suggests that the major flavonoids of S. baicalensis may interfere with these processes through partially overlapping mechanisms, although the strength of evidence is not evenly distributed across compounds or pathways. Overall, the most consistent support relates to cell-cycle regulation and apoptosis, whereas roles in autophagy, ferroptosis, and metabolic remodeling remain comparatively more context-dependent or newly emerging.
Among the major constituents, baicalin has been reported to exert relatively consistent growth-suppressive effects in NSCLC models, mainly through interference with cell-cycle control and associated survival signaling. Diao et al reported that baicalin inhibits PBK/TOPK (PDZ-binding kinase/T-LAK cell-originated protein kinase) activity, leading to decreased phosphorylation of histone H3 and ERK2 and consequent suppression of NSCLC cell proliferation.40 Additional studies further suggest that baicalin can also target mitotic control by inhibiting AURKB (Aurora kinase B) phosphorylation, thereby inducing G2/M-phase arrest and disrupting chromosome segregation-associated progression.41 Consistent with these anti-proliferative effects, baicalin has also been shown to suppress Akt/mTOR signaling, downregulate CDK2, CDK4, and Cyclin E2, and induce G1/S-phase arrest in lung cancer cells.42 In parallel, activation of the SIRT1/AMPK axis has been proposed to further contribute to inhibiting proliferation and migration while promoting apoptosis.43 Taken together, these studies suggest that baicalin may restrain NSCLC growth through coordinated effects on cell-cycle progression and survival-related signaling. However, the relative contribution of each proposed target remains unclear, and most evidence has been derived from a limited number of in vitro models and selected xenograft systems.
Baicalein likewise appears to affect NSCLC growth prominently at the level of cell-cycle regulation. It inhibits 12-LOX (12-lipoxygenase) and its metabolite 12(S)-HETE, inducing S-phase arrest accompanied by downregulation of CDK1 and Cyclin B1 and upregulation of p53 and Bax.44 Baicalein has also been reported to increase p21Cip1 while decreasing Cyclin D and Cyclin E expression, thereby inducing G0/G1-phase arrest, and to reduce PCNA (proliferating cell nuclear antigen) expression in lung cancer tissues in vivo.45 These findings indicate that baicalein may restrain tumor proliferation mainly by perturbing cell-cycle checkpoints and proliferative homeostasis.
At the level of programmed cell death, baicalein has also been one of the most extensively investigated flavonoids in NSCLC models. It activates the p53-Bax-Caspase-3 axis, resulting in mitochondrial membrane potential loss and cytochrome c release, which is consistent with intrinsic apoptosis.45 It has also been reported to activate death receptor-associated signaling through DR5/FasL/FADD, with upregulation of Caspase-8 and tBid, suggesting crosstalk between extrinsic and intrinsic apoptotic cascades.32 This process is accompanied by increased ROS generation and AMPK activation, indicating that oxidative and mitochondrial stress may participate in baicalein-associated apoptosis rather than representing completely independent mechanisms.32 Furthermore, baicalein promotes apoptosis by upregulating RUNX3 (Runt-related transcription factor 3) and FOXO3a (Forkhead box O3a) through crosstalk between AMPKα and MEK/ERK1/2 signaling.46 Overall, the apoptotic activity of baicalein is supported by multiple studies, although the hierarchy among these signaling events and their relevance under clinically achievable exposure conditions remain uncertain.
Beyond these effects on cell-cycle regulation and apoptosis, several more recent studies have proposed additional growth-suppressive mechanisms involving autophagy-associated stress, ferroptosis, and metabolic regulation. Baicalein activates the AMPK/Drp1 (dynamin-related protein 1) axis, promoting mitochondrial fission together with coordinated autophagy and apoptosis; inhibition of Drp1 significantly attenuates both processes, suggesting that mitochondrial dynamics may contribute to cell fate regulation in this setting.47 In another study, baicalein facilitated MAP4K3 ubiquitination and degradation, thereby activating TFEB-mediated autophagy and suppressing tumor growth in vitro and in vivo.48 By contrast, a more recent report suggested that baicalin targets the lysosomal channel MCOLN3, induces lysosomal alkalinization, blocks autophagic flux, and ultimately promotes mitochondrial apoptosis.49 These findings indicate that lysosomal-autophagic stress may represent a recurring theme, but they also suggest that the direction and consequence of autophagy modulation are context-dependent rather than fully unified.
Additional emerging studies have further linked Scutellaria flavonoids to ferroptosis and metabolic vulnerability in NSCLC. Baicalin was reported to induce ALOX12-mediated ferroptosis, characterized by ROS accumulation, intracellular iron overload, GSH (glutathione) depletion, increased MDA (Malondialdehyde) levels, GPX4 (glutathione peroxidase 4) downregulation, and ACSL4 (acyl-CoA synthetase long-chain family member 4) upregulation, thus providing a potential metabolic target for lung cancer therapy.50 Meanwhile, baicalein has been shown to suppress tumor cells’ dependence on glutamine metabolism by inhibiting ASCT2 (alanine-serine-cysteine transporter 2), LAT1 (L-type amino acid transporter 1), and GLS1, with downstream inhibition of mTOR signaling.51 Other recent reports additionally implicate the SMYD2/RPS7 axis and Notch signaling in the anti-proliferative activity of baicalein.52,53 These studies expand the mechanistic scope beyond cell-cycle and apoptosis, but most of these pathways remain newly emerging and await broader validation.
Compared with baicalin and baicalein, the evidence base for wogonin and wogonoside remains smaller and mechanistically less diverse. Wogonin upregulates Bad and cleaved caspase-3 while downregulating Bcl-2 and ErbB4, thereby inhibiting PI3K/Akt- and ErbB-related signaling and promoting apoptosis in NSCLC cells.54 It has also been reported to modulate the SGK1 (serum/glucocorticoid-regulated kinase 1)-p53 negative feedback loop, producing time-dependent responses that include cell-cycle arrest, senescence, and apoptosis.55 Wogonoside, in turn, activates AMPK and inhibits mTOR and p70S6K (substrate ribosomal protein S6 kinase beta-1) phosphorylation, resulting in G0/G1-phase arrest and mitochondria-mediated apoptosis; xenograft experiments further support a tumor-suppressive effect in vivo.56 Although these findings are supportive, they derive from fewer models and provide a narrower mechanistic basis than that currently available for baicalin and baicalein.
Beyond the mechanisms described above, a comparatively less explored layer of growth suppression involves epigenetic regulation. Several studies have suggested that major flavonoids from S. baicalensis may influence NSCLC cells through microRNA (miRNA)-associated regulation and histone acetylation-related mechanisms. Baicalin has been reported to upregulate miR-340-5p, which directly targets NET1 (neuroepithelial cell transforming gene 1), thereby suppressing proliferation and invasion of A549 and H1299 cells while inducing G1/S-phase arrest and apoptosis.57 These findings suggest that baicalin may also regulate cell-cycle progression and cell survival partly through a post-transcriptional mechanism. Functional experiments further validated these effects, supporting the involvement of the miR-340-5p/NET1 axis.57 Baicalein has also been reported to regulate miRNA-associated signaling at the post-transcriptional level. It downregulates miR-424-3p, restores PTEN (phosphatase and tensin homolog) activity, and suppresses PI3K/Akt signaling, leading to reduced proliferation and increased apoptosis in NSCLC cells.58 Rather than representing a completely distinct antitumor pathway, this observation may be viewed as an additional regulatory layer superimposed on signaling axes already discussed above. However, the current evidence remains limited, and the specific contribution of this miRNA-mediated regulation to the overall antitumor activity of baicalein has not yet been clearly defined. Similarly, Wogonin has been shown to exhibit epigenetic regulatory activity, mainly through modulation of histone acetylation. It downregulates HDAC1/2 (histone deacetylase 1/2), increases histone acetylation, and relieves transcriptional repression of tumor suppressor genes.59 In addition, wogonin interferes with the c-Myc/SKP2/Fbw7α signaling axis, revealing a possible link between chromatin regulation and protein stability control.59 Nevertheless, the supporting studies are few in number and are based mainly on in vitro cell models, with limited validation in vivo or in clinically relevant systems. Therefore, epigenetic regulation of flavonoids in NSCLC should be regarded as a supplementary and still emerging component of the reported antitumor profile.
Overall, currently available studies suggest that the major flavonoids of S. baicalensis may suppress NSCLC growth through interconnected effects on cell-cycle regulation, apoptosis, autophagy-associated stress, selected metabolic vulnerabilities, and an additional epigenetic regulatory layer (Tables 2 and 3). Importantly, these mechanisms should not be viewed as fully independent pathways, because many reported effects converge on a limited number of signaling hubs, particularly AMPK and PI3K/Akt/mTOR, or ultimately lead to similar phenotypic outcomes such as growth arrest and apoptosis. At the same time, the strength of evidence is clearly uneven. Cell-cycle arrest and apoptosis are supported by comparatively more consistent findings, whereas ferroptosis, lysosomal-autophagic regulation, glutamine metabolism, and other emerging pathways remain less extensively validated. In addition, much of the evidence derives from a limited set of in vitro models and xenograft systems, which may not fully capture the molecular heterogeneity and immune context of clinical NSCLC. Notably, considerable heterogeneity in experimental conditions across studies may limit direct comparison of the reported mechanisms, as cell models, treatment schedules, exposure concentrations, and endpoint definitions are not fully uniform. This issue is especially relevant for the epigenetic studies. Therefore, the growth-suppressive effects of these flavonoids are best interpreted as promising but still predominantly preclinical mechanisms rather than clinically established antitumor effects. Schematic overview of reported growth-suppressive pathways modulated by major flavonoids in NSCLC is shown in Figure 2.
 |
Table 2 Mechanistic Summary of the Tumor Growth Inhibitory Actions of Flavonoids in NSCLC |
 |
Table 3 Mechanistic Summary of Epigenetic Modulatory Actions of Flavonoids in NSCLC |
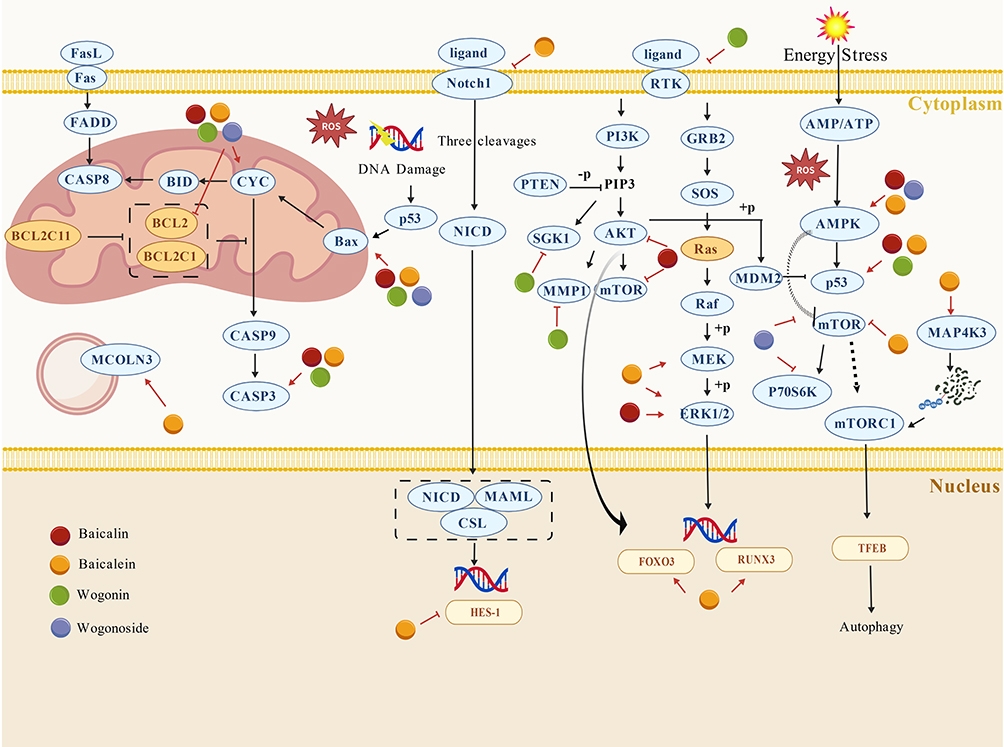 |
Figure 2 Schematic overview of reported growth-suppressive pathways modulated by major flavonoids in NSCLC. Notes: The schematic mainly highlights mitochondrial apoptotic signaling involving BCL2 family proteins, cytochrome c release, and caspase-9/3 activation; death receptor-associated apoptotic signaling through the Fas/FasL/FADD/caspase-8 axis; Notch1-related signaling; RTK/PI3K/AKT/mTOR and RAS/RAF/MEK/ERK pathways; AMPK-associated energy stress responses; MAP4K3-mTORC1-TFEB-mediated autophagy regulation; and MCOLN3-related lysosomal dysfunction. Overall, the figure is intended to show that these flavonoids may inhibit tumor growth through coordinated regulation of apoptosis, survival signaling, energy stress, and autophagy-related processes. This schematic integrates findings from different published studies and experimental models, the depicted interactions should be interpreted as reported regulatory relationships rather than universally validated direct causal interactions within a single biological context. Red arrow-headed and bar-headed lines indicate activation and inhibition, respectively. |
Inhibition of Tumor Metastasis and Invasion
In the advanced stages of tumor development, cell migration, invasion, and angiogenesis constitute critical processes that determine the malignancy and prognosis of NSCLC. Modern pharmacological studies have suggested that major flavonoids from S. baicalensis may interfere with these processes.
Among the major constituents, baicalin has been reported to inhibit migration and invasion of NSCLC cells while partially reversing EMT (epithelial-mesenchymal transition)-related phenotypes. In A549 and NCI-H460 cells, baicalin increased E-cadherin expression and reduced vimentin levels, together with inhibition of the PDK1/AKT signaling pathway, which deprives tumor cells of the driving force for transitioning to a mesenchymal phenotype.63 In addition, baicalin was shown to display SOD (superoxide dismutase)-mimetic activity and suppress HIF-1α-related responses under hypoxic conditions, with reduced migratory and angiogenic behavior in lung cancer models.64 Taken together, these findings support a role for baicalin in modulating both EMT-linked plasticity and hypoxia-associated adaptation. Although these studies primarily describe these pathways in the context of anti-metastatic effects, many of these pathways are also involved in other stages of tumor progression, including proliferation and oxidative stress regulation, suggesting that a single mechanism may influence multiple aspects of tumor biology.
Baicalein appears to show a broader mechanistic profile in this context. It has been reported to inhibit Src (proto-oncogene tyrosine-protein kinase Src) phosphorylation and downstream Id1 (DNA-binding 1) expression, thereby reversing EMT-associated changes and VEGF-A (vascular endothelial growth factor A)- mediated angiogenic signaling.65 More importantly, baicalein disrupts the iNOS (inducible nitric oxide synthase)-ezrin mechanotransduction axis in the inflammatory microenvironment.66 By reducing the S-nitrosylation level of ezrin, it decreases cytoskeletal tension, accordingly impairing pseudopodia formation and leader cell establishment.66 This biomechanical interference interrupts the physical foundation of collective tumor cell migration. Furthermore, vasculogenic mimicry (VM), a process in which tumor cells form vessel-like channels independent of endothelial cells, has been closely associated with invasiveness, metastasis, and poor prognosis. Recent evidence has demonstrated that baicalein inhibits the RhoA/ROCK pathway, weakening the actin-myosin cytoskeletal network and reducing myosin light chain (MLC) phosphorylation, ultimately preventing VM formation.67 In vivo experiments further confirmed that baicalein significantly decreases the number of VM channels and enlarges necrotic areas within tumors, underscoring its destructive effects on tumor structural and metabolic support systems.67 These findings broaden the proposed anti-metastatic profile of baicalein, although some mechanisms, particularly those involving mechanotransduction and VM, are currently supported by a limited number of studies.
Compared with baicalin and baicalein, the available evidence for wogonin and wogonoside is more limited, but it remains supportive of anti-invasive activity. Wogonin suppresses the IL-6/STAT3 pathway, blocking STAT3 phosphorylation and nuclear translocation, which in turn downregulates the EMT-associated transcription factors Snail and Twist.68 As a result, E-cadherin expression is restored while mesenchymal markers are reduced.68 These data support a possible role for wogonin in limiting inflammation-driven EMT and invasion. On the other hand, Wogonoside inhibits A549 cell invasion, migration, and microtubule bundle formation in a dose-dependent manner, leading to EMT reversal; in vivo study further demonstrates that wogonoside reduces the proportions of VEGF, vimentin, and cluster of CD34-positive cells, suggesting that it restrains tumor spread through dual mechanisms involving EMT inhibition and angiogenesis blockade.69
Another major flavonoid, OA, appears to act through EMT regulation and metabolic adaptation. It promotes Snail degradation by suppressing ERK signaling and relieving inhibition of GSK-3β (glycogen synthase kinase 3 beta), thereby reversing EMT in NSCLC cells.70 In addition, OA disrupts the c-Src/AKT-HK II/VDAC axis, attenuates glycolytic activity, reduces mitochondrial membrane potential, and restores sensitivity to anoikis (the apoptosis triggered by detachment from the extracellular matrix).71 This pattern is of particular interest because it links metastatic potential not only to EMT-associated transcriptional regulation but also to metabolic support and resistance to anoikis. Even so, the current evidence remains concentrated in relatively few experimental systems, and the extent to which these findings generalize across genetically distinct NSCLC subtypes has not yet been established.
At the integrated level, a reconstructed flavonoid complex termed reTFAE, composed of baicalein, wogonin, and OA at a ratio of 65.8%:21.2%:13.0%, exhibits potential anti-metastatic efficacy.72 ReTFAE inhibits the PI3K/AKT-TWIST1 signaling pathway, synergistically downregulating EMT-associated factors and markedly suppressing cell migration and invasion.72 Proteomic analysis further revealed that reTFAE downregulates several key glycolytic enzymes, including ALDOA, PKM, and LDHA, suggesting that it impairs metastatic potential by suppressing metabolic reprogramming.72 This observation is useful because it implies that combined flavonoid exposure may perturb metastatic behavior through coordinated effects on EMT-associated signaling and metabolic reprogramming, rather than through a single dominant target alone. However, because the formulation contains multiple components, the relative contribution of each constituent to the observed phenotype remains difficult to disentangle.
In summary, currently available studies suggest that S. baicalensis flavonoids may inhibit NSCLC metastasis and invasion through partially interconnected processes on EMT suppression, angiogenesis, cytoskeletal remodeling, and metabolic adaptation (Table 4). However, the strength of evidence differs across mechanisms. EMT-related pathways are supported by relatively consistent observations, whereas processes such as VM and mechanotransduction have been reported in fewer studies. Importantly, EMT itself is increasingly recognized as a dynamic and reversible process, so changes in a limited set of epithelial or mesenchymal markers should not automatically be equated with comprehensive blockade of metastasis. Although most available evidence is supported by both in vitro experiments and in vivo animal models, representing a relatively robust preclinical evidence base, these systems may still not fully capture the complexity of metastatic progression in clinical NSCLC. Taken together, current findings provide a biologically plausible and increasingly supported mechanistic framework, although further validation in more clinically relevant models and settings remains warranted.
 |
Table 4 Mechanistic Summary of Anti-Metastatic Actions of Flavonoids in NSCLC |
Overcoming Therapeutic Resistance
The major obstacles in NSCLC treatment lie in drug resistance and the complexity of the TME. On the one hand, tumor cells sustain survival through mechanisms such as DNA damage repair, hypoxia adaptation, and activation of pro-survival signaling; on the other hand, chronic inflammation and immune suppression within the TME collectively weaken therapeutic efficacy and amplify toxicity. Accumulating studies indicate that major flavonoids from S. baicalensis may interfere with several of these resistance-associated processes across chemotherapy, radiotherapy, targeted therapy, and immunotherapy settings.
In chemotherapy-related resistance, baicalin has been reported to enhance cisplatin (DDP) sensitivity by downregulating the expression of XRCC1 (X-ray cross complementing protein 1), in turn interfering with DNA repair.73 Combined treatment with DDP significantly suppresses the viability of both A549 and DDP-resistant A549/DDP cells, leading to S-phase arrest and accumulation of DNA damage.73 Baicalin has also been shown to inhibit MARK2 (microtubule affinity-regulating kinase 2) and attenuate a downstream prosurvival kinase cascade associated with resistance maintenance, thereby restoring DDP responsiveness.74 In parallel, baicalin regulates the KEAP1-NRF2/HO-1 axis, promotes ferritinophagy and lipid peroxidation, thereby triggering apoptosis and ferroptosis in tumor cells, which in turn contributes to enhanced DDP efficacy and partial remodeling of the TME.75 Likewise, baicalein enhances the activity of DDP through coordinated pro-apoptotic and anti-inflammatory effects, including modulation of Bcl-2 family proteins, caspase-3 activation, and suppression of TNF-α and IL-6 release.76 In addition, baicalein has shown synergistic activity with docetaxel (DOC), producing enhanced anti-proliferative and anti-angiogenic effects in association with β-catenin-related regulation, without increasing hepatic or renal toxicity.77 Wogonin has also been reported to counter inflammation-associated chemoresistance by suppressing IL-6-induced AKR1C1/1C2 expression and blocking PKC-dependent resistant phenotypes.78 Furthermore, Wogonin also promotes intracellular H2O2 accumulation, augmenting DDP-induced apoptosis.79 It should be noted that all related experiments on wogonin have so far been conducted only at the cellular level, and the level of evidence therefore remains preliminary. Another key compound, OA, targets the HIF-1α-XPC axis to suppress nucleotide excision repair (NER) activity, effectively reversing hypoxia-induced DDP resistance.80 Overall, these findings suggest that Scutellaria flavonoids may sensitize NSCLC cells to chemotherapy through partially convergent mechanisms involving DNA damage persistence, stress adaptation failure, and restoration of apoptotic responsiveness, rather than through a single dominant pathway. At the same time, several limitations warrant attention. Although some resistance-related effects have been supported in animal studies, the evidence base remains dominated by reductionist experimental systems, and only a limited proportion of mechanisms has been validated across both resistant cell models and in vivo settings. Moreover, some reported endpoints focus on pathway modulation without sufficient evidence that the observed molecular changes are the main drivers of resensitization. Accordingly, the relative importance of each proposed mechanism remains incompletely defined.
In the context of radiotherapy, baicalin exhibits both tissue-protective and response-modifying properties. Ionizing radiation induces EMT and mitochondrial ultrastructural damage in alveolar epithelial cells. Baicalin counteracts these effects by suppressing the TGF-β/SMAD and ERK/GSK-3β pathways, thereby preventing Snail nuclear translocation and alleviating epithelial injury.81 Additionally, baicalin attenuates radiation-induced pulmonary inflammation and fibrosis through inhibition of the CysLTs/CysLT1 axis and reduction of inflammatory cytokines such as TGF-β, TNF-α, IL-1β, and IL-6.82 These observations support a potentially useful adjunctive role during radiotherapy; however, protection of normal tissue and sensitization of tumor tissue are not equivalent biological outcomes, and their balance has not yet been adequately resolved in NSCLC-specific therapeutic models.
In targeted therapy, baicalein has been reported to enhance the efficacy of Almonertinib in resistant NSCLC cells by increasing ROS accumulation, suppressing resistance-associated survival signaling, and reinforcing apoptotic execution processes.83 This finding extends the potential application of Scutellaria flavonoids beyond conventional chemotherapy, but the current evidence is still confined to specific resistant models, and it remains unclear how broadly these results apply across the heterogeneous spectrum of clinically acquired resistance.
With respect to immune evasion, baicalein has been shown to directly bind to PD-L1 (CD274), facilitating its interaction with MAP1LC3B (microtubule-associated protein 1 light chain 3 beta) and promoting PD-L1 degradation through the autophagy-lysosome pathway.84 This process disrupts PD-1/PD-L1 binding, restores T cell-mediated immune responses, and significantly increases infiltration of CD4+ and CD8+ T cells in tumor tissues.84 It also enhances the expression of cytotoxic effector molecules GZMB and PRF1, achieving immune enhancement comparable to anti-PD-L1 antibodies.84 In addition, baicalin combined with tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) activates the p38-MAPK pathway and promotes ROS accumulation, significantly enhancing apoptotic sensitivity.85 Wogonin has also been shown to potentiate TRAIL’s antitumor activity in vivo.86 Further research indicates that wogonin suppresses catalase activity, leading to H2O2 accumulation, and blocks TNF-induced NF-κB activation. This results in downregulation of anti-apoptotic factors such as cellular c-FLIP and MnSOD, along with enhanced caspase-3 activation and PARP cleavage, ultimately sensitizing cells to TNF-induced apoptosis without causing cytotoxicity to normal bronchial epithelial cells.87 Meanwhile, OA inhibits the NF-κB signaling pathway and decreases TGF-β1 secretion, thereby blocking the differentiation of immunosuppressive Treg cells, effectively reshaping the immune microenvironment and mitigating immune evasion.88 These findings collectively indicate that the effects of Scutellaria flavonoids are not limited to enhancing cytotoxic responses, but also involve modulation of immune escape and sensitivity to death receptor-mediated signaling. Nevertheless, most immunoregulatory evidence remains preclinical, and the complexity of immune checkpoint response in patients is unlikely to be fully captured by the currently available models.
In general, S. baicalensis flavonoids have been reported to exert multi-layered pharmacological actions that target critical barriers in NSCLC therapy (Table 5). Their mechanisms encompass inhibition of DNA repair, modulation of oxidative stress, degradation of immune checkpoints, and reprogramming of signaling pathways. These actions underscore their potential to enhance efficacy, reduce adverse effects, and facilitate combination therapy translation. At present, although several major flavonoids have been implicated in resistance modulation, direct evidence for wogonoside in overcoming therapeutic resistance remains lacking, indicating that this aspect of its pharmacological profile is still insufficiently explored.
 |
Table 5 Mechanistic Summary of the Sensitizing and Resistance-Reversing Actions of Flavonoids in NSCLC |
To integrate the mechanistic evidence summarized above, we further compiled the major flavonoids into a unified schematic overview (Figure 3). Importantly, only pathways and mechanistic categories supported by both in vitro and in vivo studies were included, in order to highlight the most robust findings. As shown in Figure 3, these compounds converge on a limited number of recurrent signaling hubs, while also exhibiting partially distinct mechanistic profiles. Baicalin and baicalein display the broadest mechanistic coverage, spanning inflammatory regulation, oxidative stress modulation, cell-cycle control, apoptosis, and additional processes such as ferroptosis, DNA damage-related regulation, mitochondrial regulation, angiogenesis, or metabolic regulation. By contrast, wogonin, wogonoside, and OA appear to act through comparatively narrower but still biologically relevant mechanisms, particularly EMT-related regulation and selected stress-response or metabolic pathways. This pattern suggests that, the antitumor actions of S. baicalensis flavonoids may not represent fully independent mechanisms, but rather context-dependent perturbations of several shared regulatory hubs. Despite the apparent diversity of reported targets, most mechanistic studies remain confined to preclinical models, and there is a lack of well-controlled clinical studies. At the same time, the mechanistic breadth is highly uneven across compounds, with baicalin and baicalein being extensively studied, whereas wogonin, wogonoside, and OA are supported by comparatively limited in vivo data, potentially biasing the overall mechanistic landscape. Additionally, many studies focus on single pathways in isolation, while the extent to which these signaling networks interact or dominate under physiological conditions remains insufficiently defined. Further studies could focus on these aspects to overcome the current gaps.
 |
Figure 3 Key pathways and regulatory mechanisms of the major flavonoids in NSCLC. Notes: Only pathways and mechanisms validated in both in vitro and in vivo studies were included in this figure Purple hexagons represent flavonoid compounds, yellow diamonds represent key pathways, and the green circles represent that the compound acts via the corresponding pathway. |
Toxicity and Safety
Although S. baicalensis flavonoids have demonstrated broad therapeutic potential in preclinical studies, its clinical safety remains an important consideration for further translational development. First, the drug toxicity characteristics of key active constituents were evaluated using ADMETlab 2.0. As summarized in Table 6, toxicity predictions indicated that all five compounds were predicted to be non-blockers of hERG, suggesting a low predicted risk of hERG-related cardiotoxicity. Predictions for H-HT, RT, and carcinogenicity were all negative. Notably, the DILI prediction for all five major flavonoids was moderately positive, implying a mild risk of hepatocellular stress during long-term or high-dose exposure. The predicted LD50 ranged from 4.688 to 6.061 mg/kg, indicating low acute toxicity and a wide therapeutic window. However, these in silico predictions should be regarded only as supplementary evidence and cannot serve as independent proof of safety, nor do they replace experimental toxicology or clinical evaluation. Therefore, they should be interpreted together with published in vitro, in vivo, and clinical observations.
 |
Table 6 Summary of Predicted Toxicity of the Key Compounds |
Growing evidence further supports these findings. High-dose baicalin (≥400 mg/kg/day) has been reported to induce renal injury and fibrosis in rodents, whereas in a subchronic toxicity model employing a baicalin-standardized formulation, administration up to 2000 mg/kg/day caused no apparent organ toxicity, suggesting that its toxic profile may depend on formulation type and exposure route.90,91 In addition, baicalin was found to reduce plasma exposure to rosuvastatin in an OATP1B1 (organic anion transporting polypeptide 1B1) haplotype-dependent manner, indicating potential drug-drug interactions when co-administered with modern therapeutic agents.92 Nevertheless, this study included only a small sample size (n = 18), consisting exclusively of male volunteers, and its findings should be interpreted with caution. A phase I clinical study of baicalein chewable tablets in healthy Chinese volunteers (n = 70) demonstrated that systemic exposure to baicalin, the major glucuronide metabolite, was markedly higher than that of the parent compound; however, pharmacokinetic parameters remained stable, showing no accumulation or compound-specific toxicity.93 In the same study, single oral doses of baicalein up to 2800 mg were well tolerated, and no dose-limiting toxicity was observed.93 These data are broadly consistent with the ADMET predictions, suggesting an overall favorable safety profile with mainly mild and reversible effects. Across single-dose and dose-escalation studies (100–800 mg), no serious adverse events occurred; vital signs and electrocardiograms remained stable, and no cumulative toxicity was noted.20,94,95 Occasional gastrointestinal discomfort (such as bloating or constipation) was reported.20,94,95 Studies also documented slight elevations in serum triglycerides, suggesting that baicalein may influence lipid metabolism, although its clinical relevance and underlying mechanisms remain unclear.20,94 A small number of participants exhibited transient increases in high-sensitivity C-reactive protein (hs-CRP), alanine aminotransferase (ALT), or aspartate aminotransferase (AST), and brief proteinuria; all abnormalities were reversible and showed no clear dose correlation.20,94,95 These observations are also broadly in line with the ADMET predictions (non-hERG blockade but DILI-positive), while indicating the need for monitoring mild hepatocellular effects during long-term administration.
For wogonin, integrated toxicological studies show that it is generally safe at low concentrations but carries potential risks under high-dose or chronic exposure. A reproductive and developmental toxicity study found that intravenous administration of high-dose wogonin (40 mg/kg) in pregnant rats caused maternal weight loss and fetal skeletal abnormalities, while in vitro assays revealed slight chromosomal aberrations at high concentrations (up to 16 µg/mL).96 Nevertheless, subchronic toxicity studies confirmed a broad therapeutic window. In a 90-day intravenous study in beagle dogs, doses up to 60 mg/kg/day produced no significant histopathological damage to the heart, liver, or kidneys, with only minor increases in platelet volume and serum lipids. The no-observed-adverse-effect level (NOAEL) was determined to be 30 mg/kg, indicating low acute toxicity and reversibility, though chronic administration at very high doses (120 mg/kg) was associated with cardiac injury risk.96
OA likewise exhibited a favorable safety profile in both animal and human studies. In animal models, OA at 50 mg/kg induced no weight loss or organ pathology and markedly alleviated temozolomide-induced leukopenia and pulmonary injury, suggesting a potential chemoprotective effect.97 In a phase I clinical trial, OA was well tolerated at doses ranging from 400 mg to 2400 mg, with most adverse events being mild gastrointestinal discomfort or asymptomatic laboratory abnormalities.23 Only a few participants experienced grade 2–3 ALT/AST elevations, all of which resolved upon discontinuation.23 These findings are consistent with ADMET results, indicating that such changes likely reflect reversible pharmacological stress rather than sustained hepatotoxicity. Available clinical studies primarily evaluating the safety and tolerability of these compounds are summarized in Table 7.
 |
Table 7 Summary of Safety of the Key Compounds in Clinical Trials |
Collectively, both ADMET-based predictions and in vivo/in vitro evidence converge to show that Scutellaria-derived flavonoids exhibit low acute toxicity and high systemic safety. The principal risks involve mild, reversible fluctuations in liver enzymes and metabolic indices. Furthermore, the currently available clinical evidence remains limited, with clinical studies reported mainly for baicalein and OA, while other major flavonoids still lack rigorous clinical validation. All studies are early-phase (Phase I), single-center trials conducted in healthy volunteers, with relatively small sample sizes and limited statistical power. In addition, population representativeness is restricted, and the effects of factors such as sex, age, and disease status have not been adequately evaluated. Further well-designed Phase II/III clinical trials in NSCLC patients are needed, with larger sample sizes, repeated and long-term dosing regimens, to provide more robust evidence for safety and to evaluate the clinical translatability of these flavonoids in NSCLC treatment.
Novel Strategies for Clinical Translation: Advanced Drug Delivery Systems
The multidimensional pharmacological activities of flavonoids from S. baicalensis against NSCLC have been extensively investigated, revealing their potential to inhibit tumor growth, metastasis, and drug resistance through multiple molecular mechanisms. However, their clinical application is often limited by poor solubility, low oral bioavailability, and rapid systemic metabolism. In recent years, the emergence of nanotechnology-based drug delivery systems has created new opportunities for the modernization of TCM and precision anticancer therapy.
A lung-targeted baicalin-loaded nanoliposome formulation achieved preferential pulmonary distribution in vivo and significantly prolonged survival in orthotopic lung cancer models without causing hemolysis or pulmonary tissue injury.98 Similarly, transferrin-modified, pH-sensitive solid lipid nanoparticles co-delivering DOC and baicalin showed synergistic antitumor activity in drug-resistant NSCLC cells.99 In vivo, this system exhibited tumor suppression with relatively low systemic toxicity, suggesting that targeted co-delivery nanoplatforms may enhance chemosensitivity while mitigating adverse effects.99 These findings suggest a potential improvement in local drug exposure and tolerability; however, whether such advantages can be maintained under clinically relevant conditions remains to be determined.
A dual-targeting nano-dandelion system (Que-S-S-oHA-Man-FA, QHMF) based on an oligomeric hyaluronic acid (oHA) backbone was designed to co-deliver curcumin and baicalin for the treatment of NSCLC. This system simultaneously targets CD44 receptors on tumor cells and CD206 receptors on tumor-associated macrophages (TAMs), enabling reduction-responsive drug release under high GSH conditions.100 It achieved high tumor accumulation and pronounced antitumor efficacy in vivo, ultimately realizing synergistic chemo-immunotherapeutic outcomes.100 While this strategy highlights the feasibility of multi-target delivery, the increased structural complexity may introduce challenges in formulation stability and large-scale production. In addition, baicalin has been incorporated into gene delivery systems. A baicalin-modified polyethyleneimine (PEI) vector delivering miR-34a demonstrated significant inhibition of lung cancer cell proliferation and migration, induced cell death, and showed no observable toxicity in major organs.101
Baicalein-based co-delivery systems also demonstrate significant advantages in achieving synergistic effects and toxicity reduction. Biomimetic selenium-baicalein nanoparticles (ACM-Sse-BE), cloaked with tumor cell membranes for homologous targeting, exhibited pH-responsive release that triggered ROS generation and apoptosis, achieving a tumor inhibition rate of 84.52%, underscoring the therapeutic promise of biomimetic design strategies.102 Furthermore, a dual-ligand self-assembled nanoparticles co-delivering paclitaxel and baicalein effectively overcame multidrug resistance and enhanced tumor-selective cytotoxicity, providing a promising approach for personalized therapy.103 Optimization of administration routes has further improved translational potential. Oral solid lipid nanoparticles (SLNB) significantly increased the oral bioavailability of baicalein (by approximately 3-fold) and exhibited a dual function in radiotherapy by both protecting normal cells from radiation-induced damage and sensitizing NSCLC cells to radiation, demonstrating promising clinical application prospects.104 However, achieving a balance between radioprotection of normal tissues and radiosensitization of tumor cells remains inherently challenging due to underlying biological constraints.
In conclusion, nanocarrier-based delivery systems for S. baicalensis flavonoids have shown potential to improve pharmacokinetic behavior, enhance tumor targeting, and support combination therapy strategies. Nevertheless, it should be noted that most of these delivery systems remain at the preclinical stage, and their ability to overcome fundamental pharmacokinetic limitations in humans has not yet been fully validated. Challenges such as large-scale manufacturing stability, long-term safety, and behavior within complex human microenvironments still require further investigation.
Conclusion and Future Prospects
As a classical traditional Chinese herb, S. baicalensis has attracted increasing attention in NSCLC research due to its multi-component and multi-target pharmacological characteristics. The major flavonoid constituents of S. baicalensis have been reported to modulate a range of biological processes relevant to NSCLC, including early intervention of inflammation and oxidative stress, inhibition of tumor growth, regulation of epigenetic modifications, blockade of metastasis, and reversal of therapeutic resistance. These effects are associated with the regulation of key signaling pathways such as NF-κB, STAT3, PI3K/Akt, and cGAS-STING, as well as cellular processes including cell cycle arrest, apoptosis, autophagy induction, ferroptosis promotion, and TME remodeling. Notably, these multi-target activities also suggest a potential role of these flavonoids as adjuvant agents that may enhance the efficacy of existing therapies, highlighting a relatively realistic pathway for clinical translation.
Despite these encouraging findings, the currently available evidence is still largely based on in vitro and animal studies, and the consistency of these findings across different experimental models has not yet been fully established. Moreover, the exposures associated with promising preclinical effects may be difficult to achieve or maintain in a clinical setting, further complicating clinical translation. Current human pharmacokinetic data from phase I studies have shown that systemic exposure to the parent compounds is limited and increases in a less than dose-proportional manner, with Cmax values often orders of magnitude lower than the concentrations required for in vitro anticancer effects. This discrepancy highlights a potential pharmacokinetic-pharmacodynamic mismatch, in which mechanistic effects observed under experimental conditions may not directly translate into clinical efficacy. This mismatch is largely driven by inherent pharmacokinetic limitations, including low oral bioavailability, rapid metabolism, and restricted tissue distribution, which collectively limit systemic exposure to bioactive forms.
In this context, nanocarrier-based drug delivery systems, including strategies such as targeted modification, stimuli-responsive release, and co-delivery, have shown potential to improve poor bioavailability, as well as to enhance pulmonary distribution and tumor accumulation. These approaches may also strengthen synergistic effects with chemotherapy, targeted therapy, and immunotherapy, although they are still at an early stage of development. Beyond nanocarrier-based delivery, the translational potential of S. baicalensis flavonoids may also lie in their roles as chemosensitizers, immunomodulatory agents, and multi-target regulators, supported by their favorable safety profiles and opportunities for pharmacokinetic optimization.
In terms of safety, available evidence suggests that S. baicalensis constituents are generally well tolerated. Both ADMET-based predictions and existing clinical observations support a relatively favorable safety profile. However, potential adverse effects, such as mild elevations in liver enzymes and alterations in lipid metabolism, should be carefully monitored, particularly during long-term or high-dose administration.
Future research should place greater emphasis on bridging the gap between preclinical findings and clinical application. Well-designed, multicenter clinical trials are needed to rigorously evaluate the efficacy and safety of S. baicalensis flavonoids under clinically achievable exposure conditions. In addition, further studies should clarify the mechanisms underlying their potential synergistic effects with immunotherapy and targeted therapy. There is also a need to develop more efficient, lung-targeted, and intelligent drug delivery systems to improve bioavailability and tissue distribution. At the same time, greater efforts are required to reinterpret the compatibility theory of TCM within a modern biomedical framework, in order to systematically elucidate the interactions among multiple components in compound formulations.
In summary, S. baicalensis flavonoids demonstrate considerable potential in the prevention and treatment of NSCLC. The integration of its multidimensional pharmacological mechanisms with advanced drug delivery technologies provides new perspectives for developing effective integrative therapeutic strategies that bridge TCM and modern oncology. However, a more rigorous and evidence-based approach is required to support its further clinical development in NSCLC.
Funding
This study was supported by the National Key R&D Program of China (2018YFC200 2500).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Zhou H, Zheng Z, Fan C, Zhou Z. Mechanisms and strategies of immunosenescence effects on non-small cell lung cancer (NSCLC) treatment: a comprehensive analysis and future directions. Semin Cancer Biol. 2025;109:44–25. doi:10.1016/j.semcancer.2025.01.001
2. Thai AA, Solomon BJ, Sequist LV, Gainor JF, Heist RS. Lung cancer. Lancet. 2021;398(10299):535–554. doi:10.1016/S0140-6736(21)00312-3
3. Li Y, Tuerxun H, Zhao Y, et al. The new era of lung cancer therapy: combining immunotherapy with ferroptosis. Crit Rev Oncol Hematol. 2024;198:104359. doi:10.1016/j.critrevonc.2024.104359
4. Rahal Z, El Darzi R, Moghaddam SJ, Cascone T, Kadara H. Tumour and microenvironment crosstalk in NSCLC progression and response to therapy. Nat Rev Clin Oncol. 2025;22(7):463–482. doi:10.1038/s41571-025-01021-1
5. Shi S, Zhao S, Tian X, et al. Molecular and metabolic mechanisms of bufalin against lung adenocarcinoma: new and comprehensive evidences from network pharmacology, metabolomics and molecular biology experiment. Comput Biol Med. 2023;157:106777. doi:10.1016/j.compbiomed.2023.106777
6. Xiang L, Gao Y, Chen S, Sun J, Wu J, Meng X. Therapeutic potential of Scutellaria baicalensis Georgi in lung cancer therapy. Phytomedicine. 2022;95:153727. doi:10.1016/j.phymed.2021.153727
7. Liao H, Ye J, Gao L, Liu Y. The main bioactive compounds of Scutellaria baicalensis Georgi. For alleviation of inflammatory cytokines: a comprehensive review. Biomed Pharmacother. 2021;133:110917. doi:10.1016/j.biopha.2020.110917
8. Yu P, Li J, Luo Y, et al. Mechanistic role of Scutellaria baicalensis Georgi in breast cancer therapy. Am J Chin Med. 2023;51(2):279–308. doi:10.1142/S0192415X23500155
9. Zhao T, Tang H, Xie L, et al. Scutellaria baicalensis Georgi. (Lamiaceae): a review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. J Pharm Pharmacol. 2019;71(9):1353–1369. doi:10.1111/jphp.13129
10. Song JW, Long JY, Xie L, et al. Applications, phytochemistry, pharmacological effects, pharmacokinetics, toxicity of Scutellaria baicalensis Georgi. And its probably potential therapeutic effects on COVID-19: a review. Chin Med. 2020;15(1):102. doi:10.1186/s13020-020-00384-0
11. Liu J, Meng F, Zhang S, et al. Simultaneous determination of baicalin, wogonoside, baicalein, wogonin, and oroxylin-A in Scutellaria baicalensis by UPLC. Chin Traditional Herbal Drugs. 2014;45(10):1477–1480.
12. Jiang Y, Zhao Z, Xiao Y, Xu H. Identifying the habitat quality of Scutellaria baicalensis based on baicalin content using the fuzzy matter element model. Ecol Indic. 2022;141:109033. doi:10.1016/j.ecolind.2022.109033
13. Bochoráková H, Paulová H, Slanina J, Musil P, Táborská E. Main flavonoids in the root of Scutellaria baicalensis cultivated in Europe and their comparative antiradical properties. Phytother Res. 2003;17(6):640–644. doi:10.1002/ptr.1216
14. Wang ZL, Wang S, Kuang Y, Hu ZM, Qiao X, Ye M. A comprehensive review on phytochemistry, pharmacology, and flavonoid biosynthesis of Scutellaria baicalensis. Pharm Biol. 2018;56(1):465–484. doi:10.1080/13880209.2018.1492620
15. Yang JY, Li M, Zhang CL, Liu D. Pharmacological properties of baicalin on liver diseases: a narrative review. Pharmacol Rep. 2021;73(5):1230–1239. doi:10.1007/s43440-021-00227-1
16. Si L, Lai Y. Pharmacological mechanisms by which baicalin ameliorates cardiovascular disease. Front Pharmacol. 2024;15:1415971. doi:10.3389/fphar.2024.1415971
17. Zheng W, Sun G, Chen J, et al. Inhibitory effects of Coptidis Rhizoma on the intestinal absorption and metabolism of Scutellariae Radix. J Ethnopharmacol. 2021;270:113785. doi:10.1016/j.jep.2021.113785
18. Wen Y, Wang Y, Zhao C, Zhao B, Wang J. The pharmacological efficacy of baicalin in inflammatory diseases. Int J Mol Sci. 2023;24(11):9317. doi:10.3390/ijms24119317
19. Lai J, Li C. Review on the pharmacological effects and pharmacokinetics of scutellarein. Arch Pharm. 2024;357(9):e2400053. doi:10.1002/ardp.202400053
20. Dong R, Li L, Gao H, et al. Safety, tolerability, pharmacokinetics, and food effect of baicalein tablets in healthy Chinese subjects: a single-center, randomized, double-blind, placebo-controlled, single-dose phase I study. J Ethnopharmacol. 2021;274:114052. doi:10.1016/j.jep.2021.114052
21. Tuli HS, Rath P, Chauhan A, et al. Wogonin, as a potent anticancer compound: from chemistry to cellular interactions. Exp Biol Med. 2023;248(9):820–828. doi:10.1177/15353702231179961
22. Ren G, Chen H, Zhang M, et al. Pharmacokinetics, tissue distribution and excretion study of Oroxylin A, Oroxylin A 7-O-glucuronide and Oroxylin A sodium sulfonate in rats after administration of Oroxylin A. Fitoterapia. 2020;142:104480. doi:10.1016/j.fitote.2020.104480
23. Yang F, Meng X, Qin S, Guo Q, Wei L, Zhao D. Safety and pharmacokinetics evaluation of oroxylin A in Chinese healthy volunteers: a phase I, double-blind, placebo-controlled, single ascending dose, multiple dose, and food effect study. Clin Transl Oncol. 2026;28(3):1016–1029. doi:10.1007/s12094-025-04017-6
24. Wang L, Ni B, Wang J, et al. Research progress of Scutellaria baicalensis in the treatment of gastrointestinal cancer. Integr Cancer Ther. 2024;23:15347354241302049. doi:10.1177/15347354241302049
25. Wu J, He Z, Zhuang W, et al. Mechanisms of inflammation-driven lung cancer: from external influences to internal regulation. Clin Immunol. 2025;279:110572. doi:10.1016/j.clim.2025.110572
26. Jomova K, Raptova R, Alomar SY, et al. Reactive oxygen species, toxicity, oxidative stress, and antioxidants: chronic diseases and aging. Arch Toxicol. 2023;97(10):2499–2574. doi:10.1007/s00204-023-03562-9
27. Qi C, Sun SW, Xiong XZ. From COPD to lung cancer: mechanisms linking, diagnosis, treatment, and prognosis. Int J Chron Obstruct Pulmon Dis. 2022;17:2603–2621. doi:10.2147/COPD.S380732
28. Sabry K, Jamshidi Z, Emami SA, Sahebka A. Potential therapeutic effects of baicalin and baicalein. Avicenna J Phytomed. 2024;14(1):23–49. doi:10.22038/AJP.2023.22307
29. Ahmadi A, Mortazavi Z, Mehri S, Hosseinzadeh H. Protective and therapeutic effects of Scutellaria baicalensis and its main active ingredients baicalin and baicalein against natural toxicities and physical hazards: a review of mechanisms. Daru. 2022;30(2):351–366. doi:10.1007/s40199-022-00443-x
30. Guo L, Yue M, Ma C, Wang Y, Hou J, Li H. Baicalin reduces inflammation to inhibit lung cancer via targeting SOCS1/NF-κB/STAT3 axis. Heliyon. 2024;10(8):e29361. doi:10.1016/j.heliyon.2024.e29361
31. Naveenkumar C, Raghunandhakumar S, Asokkumar S, Devaki T. Baicalein abrogates reactive oxygen species (ROS)-mediated mitochondrial dysfunction during experimental pulmonary carcinogenesis in vivo. Basic Clin Pharmacol Toxicol. 2013;112(4):270–281. doi:10.1111/bcpt.12025
32. Kim HJ, Park C, Han MH, et al. Baicalein induces caspase-dependent apoptosis associated with the generation of ROS and the activation of AMPK in human lung carcinoma A549 cells. Drug Dev Res. 2016;77(2):73–86. doi:10.1002/ddr.21298
33. Zheng T, Liu H, Hong Y, et al. Promotion of liquid-to-solid phase transition of cGAS by Baicalein suppresses lung tumorigenesis. Signal Transduct Target Ther. 2023;8(1):133. doi:10.1038/s41392-023-01326-6
34. Naveenkumar C, Raghunandakumar S, Asokkumar S, et al. Mitigating role of baicalein on lysosomal enzymes and xenobiotic metabolizing enzyme status during lung carcinogenesis of Swiss albino mice induced by benzo(a)pyrene. Fundam Clin Pharmacol. 2014;28(3):310–322. doi:10.1111/fcp.12036
35. Chandrashekar N, Selvamani A, Subramanian R, Pandi A, Thiruvengadam D. Baicalein inhibits pulmonary carcinogenesis-associated inflammation and interferes with COX-2, MMP-2 and MMP-9 expressions in-vivo. Toxicol Appl Pharmacol. 2012;261(1):10–21. doi:10.1016/j.taap.2012.02.004
36. Wang C, Cui C. Inhibition of lung cancer proliferation by wogonin is associated with activation of apoptosis and generation of reactive oxygen species. Balkan Med J. 2019;37(1):29–33. doi:10.4274/balkanmedj.galenos.2019.2019.7.75
37. Chen LG, Hung LY, Tsai KW, et al. Wogonin, a bioactive flavonoid in herbal tea, inhibits inflammatory cyclooxygenase-2 gene expression in human lung epithelial cancer cells. Mol Nutr Food Res. 2008;52(11):1349–1357. doi:10.1002/mnfr.200700329
38. Grigalius I, Petrikaite V. Relationship between antioxidant and anticancer activity of trihydroxyflavones. Molecules. 2017;22(12):2169. doi:10.3390/molecules22122169
39. Wu K, El Zowalaty AE, Sayin VI, Papagiannakopoulos T. The pleiotropic functions of reactive oxygen species in cancer. Nat Cancer. 2024;5(3):384–399. doi:10.1038/s43018-024-00738-9
40. Diao X, Yang D, Chen Y, Liu W. Baicalin suppresses lung cancer growth by targeting PDZ-binding kinase/T-LAK cell-originated protein kinase. Biosci Rep. 2019;39(4):BSR20181692. doi:10.1042/BSR20181692
41. Noor S, Choudhury A, Raza A, et al. Probing Baicalin as potential inhibitor of Aurora kinase B: a step towards lung cancer therapy. Int J Biol Macromol. 2024;258(Pt 1):128813. doi:10.1016/j.ijbiomac.2023.128813
42. Sui X, Han X, Chen P, et al. Baicalin induces apoptosis and suppresses the cell cycle progression of lung cancer cells through downregulating Akt/mTOR signaling pathway. Front Mol Biosci. 2021;7:602282. doi:10.3389/fmolb.2020.602282
43. You J, Cheng J, Yu B, Duan C, Peng J. Baicalin, a Chinese herbal medicine, inhibits the proliferation and migration of human non-small cell lung carcinoma (NSCLC) cells, A549 and H1299, by activating the SIRT1/AMPK signaling pathway. Med Sci Monit. 2018;24:2126–2133. doi:10.12659/MSM.909627
44. Leung HW, Yang WH, Lai MY, Lin CJ, Lee HZ. Inhibition of 12-lipoxygenase during baicalein-induced human lung nonsmall carcinoma H460 cell apoptosis. Food Chem Toxicol. 2007;45(3):403–411. doi:10.1016/j.fct.2006.08.021
45. Chandrashekar N, Subramanian R, Thiruvengadam D. Baicalein inhibits cell proliferation and enhances apoptosis in human A549 cells and benzo(a)pyrene-induced pulmonary carcinogenesis in mice. J Biochem Mol Toxicol. 2022;36(7):e23053. doi:10.1002/jbt.23053
46. Zheng F, Wu J, Zhao S, et al. Baicalein increases the expression and reciprocal interplay of RUNX3 and FOXO3a through crosstalk of AMPKα and MEK/ERK1/2 signaling pathways in human non-small cell lung cancer cells. J Exp Clin Cancer Res. 2015;34(1):41. doi:10.1186/s13046-015-0160-7
47. Deng X, Liu J, Liu L, Sun X, Huang J, Dong J. Drp1-mediated mitochondrial fission contributes to baicalein-induced apoptosis and autophagy in lung cancer via activation of AMPK signaling pathway. Int J Biol Sci. 2020;16(8):1403–1416. doi:10.7150/ijbs.41768
48. Li J, Yan L, Luo J, et al. Baicalein suppresses growth of non-small cell lung carcinoma by targeting MAP4K3. Biomed Pharmacother. 2021;133:110965. doi:10.1016/j.biopha.2020.110965
49. Dong X, Liu X, Lin D, et al. Baicalin induces cell death of non-small cell lung cancer cells via MCOLN3-mediated lysosomal dysfunction and autophagy blockage. Phytomedicine. 2024;133:155872. doi:10.1016/j.phymed.2024.155872
50. Jin Y, Wen J, Geng Z, et al. Baicalin inhibits lung cancer cell proliferation and migration via ALOX12-mediated ferroptosis. Anticancer Agents Med Chem. 2025;25(20):1642–1659. doi:10.2174/0118715206342238250428115441
51. Li J, Zhang D, Wang S, et al. Baicalein induces apoptosis by inhibiting the glutamine-mTOR metabolic pathway in lung cancer. J Adv Res. 2025;68:341–357. doi:10.1016/j.jare.2024.02.023
52. Gu L, Xu F, Zhang X, Gu Z. Baicalein inhibits the SMYD2/RPS7 signaling pathway to inhibit the occurrence and metastasis of lung cancer. J Oncol. 2022;2022:3796218. doi:10.1155/2022/3796218
53. Su G, Chen H, Sun X. Baicalein suppresses non small cell lung cancer cell proliferation, invasion and Notch signaling pathway. Cancer Biomark. 2018;22(1):13–18. doi:10.3233/CBM-170673
54. Wang L, Zhang J, Shan G, et al. Establishment of a lung cancer discriminative model based on an optimized support vector machine algorithm and study of key targets of Wogonin in lung cancer. Front Pharmacol. 2021;12:728937. doi:10.3389/fphar.2021.728937
55. Shi G, Wang Q, Zhou X, et al. Response of human non-small-cell lung cancer cells to the influence of Wogonin with SGK1 dynamics. Acta Biochim Biophys Sin. 2017;49(4):302–310. doi:10.1093/abbs/gmx006
56. Luo M, Mo J, Yu Q, et al. Wogonoside induces apoptosis in human non-small cell lung cancer A549 cells by promoting mitochondria dysfunction. Biomed Pharmacother. 2018;106:593–598. doi:10.1016/j.biopha.2018.06.077
57. Zhao F, Zhao Z, Han Y, Li S, Liu C, Jia K. Baicalin suppresses lung cancer growth phenotypes via miR-340-5p/NET1 axis. Bioengineered. 2021;12(1):1699–1707. doi:10.1080/21655979.2021.1922052
58. Lu C, Wang H, Chen S, Yang R, Li H, Zhang G. Baicalein inhibits cell growth and increases cisplatin sensitivity of A549 and H460 cells via miR-424-3p and targeting PTEN/PI3K/Akt pathway. J Cell Mol Med. 2018;22(4):2478–2487. doi:10.1111/jcmm.13556
59. Chen XM, Bai Y, Zhong YJ, et al. Wogonin has multiple anti-cancer effects by regulating c-Myc/SKP2/Fbw7α and HDAC1/HDAC2 pathways and inducing apoptosis in human lung adenocarcinoma cell line A549. PLoS One. 2013;8(11):e79201. doi:10.1371/journal.pone.0079201
60. Cathcart MC, Useckaite Z, Drakeford C, et al. Anti-cancer effects of baicalein in non-small cell lung cancer in-vitro and in-vivo. BMC Cancer. 2016;16(1):707. doi:10.1186/s12885-016-2740-0
61. Guo J, Jin G, Hu Y, et al. Wogonin restrains the malignant progression of lung cancer through modulating MMP1 and PI3K/AKT signaling pathway. Protein Pept Lett. 2023;30(1):25–34. doi:10.2174/0929866530666221027152204
62. Wang SJ, Zhao JK, Ren S, Sun WW, Zhang WJ, Zhang JN. Wogonin affects proliferation and the energy metabolism of SGC-7901 and A549 cells. Exp Ther Med. 2019;17(1):911–918. doi:10.3892/etm.2018.7023
63. Chen J, Yuan CB, Yang B, Zhou X. Baicalin inhibits EMT through PDK1/AKT signaling in human nonsmall cell lung cancer. J Oncol. 2021;2021:4391581. doi:10.1155/2021/4391581
64. Du G, Han G, Zhang S, et al. Baicalin suppresses lung carcinoma and lung metastasis by SOD mimic and HIF-1alpha inhibition. Eur J Pharmacol. 2010;630(1–3):121–130. doi:10.1016/j.ejphar.2009.12.014
65. Zhao Z, Liu B, Sun J, et al. Baicalein inhibits orthotopic human non-small cell lung cancer Xenografts via Src/Id1 pathway. Evid Based Complement Alternat Med. 2019;2019:9806062. doi:10.1155/2019/9806062
66. Zhang X, Ruan Q, Zhai Y, et al. Baicalein inhibits non-small-cell lung cancer invasion and metastasis by reducing ezrin tension in inflammation microenvironment. Cancer Sci. 2020;111(10):3802–3812. doi:10.1111/cas.14577
67. Zhang Z, Nong L, Chen M, et al. Baicalein suppresses vasculogenic mimicry through inhibiting RhoA/ROCK expression in lung cancer A549 cell line. Acta Biochim Biophys Sin. 2020;52(9):1007–1015. doi:10.1093/abbs/gmaa075
68. Zhao Y, Yao J, Wu XP, et al. Wogonin suppresses human alveolar adenocarcinoma cell A549 migration in inflammatory microenvironment by modulating the IL-6/STAT3 signaling pathway. Mol Carcinog. 2015;54 Suppl 1:E81–E93. doi:10.1002/mc.22182
69. Yan Y, Yao L, Sun H, et al. Effects of wogonoside on invasion and migration of lung cancer A549 cells and angiogenesis in xenograft tumors of nude mice. J Thorac Dis. 2020;12(4):1552–1560. doi:10.21037/jtd-20-1555
70. Wei L, Yao Y, Zhao K, et al. Oroxylin A inhibits invasion and migration through suppressing ERK/GSK-3β signaling in snail-expressing non-small-cell lung cancer cells. Mol Carcinog. 2016;55(12):2121–2134. doi:10.1002/mc.22456
71. Wei L, Dai Q, Zhou Y, et al. Oroxylin A sensitizes non-small cell lung cancer cells to anoikis via glucose-deprivation-like mechanisms: c-Src and hexokinase II. Biochim Biophys Acta. 2013;1830(6):3835–3845. doi:10.1016/j.bbagen.2013.03.009
72. Cao HJ, Zhou W, Xian XL, et al. A mixture of Baicalein, Wogonin, and Oroxylin-A inhibits EMT in the A549 cell line via the PI3K/AKT-TWIST1-glycolysis pathway. Front Pharmacol. 2022;12:821485. doi:10.3389/fphar.2021.821485
73. Yin Z, Chen E, Cai X, et al. Baicalin attenuates XRCC1-mediated DNA repair to enhance the sensitivity of lung cancer cells to cisplatin. J Recept Signal Transduction Res. 2022;42(3):215–224. doi:10.1080/10799893.2021.1892132
74. Xu Z, Mei J, Tan Y. Baicalin attenuates DDP (cisplatin) resistance in lung cancer by downregulating MARK2 and p-Akt. Int J Oncol. 2017;50(1):93–100. doi:10.3892/ijo.2016.3768
75. Chen Y, Bao S, Wang Z, Fang Z, Tang H. Baicalin promotes the sensitivity of NSCLC to cisplatin by regulating ferritinophagy and macrophage immunity through the KEAP1-NRF2/HO-1 pathway. Eur J Med Res. 2024;29(1):387. doi:10.1186/s40001-024-01930-4
76. Kiartivich S, Wei Y, Liu J, et al. Regulation of cytotoxicity and apoptosis-associated pathways contributes to the enhancement of efficacy of cisplatin by baicalein adjuvant in human A549 lung cancer cells. Oncol Lett. 2017;13(4):2799–2804. doi:10.3892/ol.2017.5746
77. Lu L, Zhang M, Wang X, et al. Baicalein enhances the antitumor efficacy of docetaxel on nonsmall cell lung cancer in a β-catenin-dependent manner. Phytother Res. 2020;34(1):104–117. doi:10.1002/ptr.6501
78. Wang HW, Lin CP, Chiu JH, et al. Reversal of inflammation-associated dihydrodiol dehydrogenases (AKR1C1 and AKR1C2) overexpression and drug resistance in nonsmall cell lung cancer cells by wogonin and chrysin. Int J Cancer. 2007;120(9):2019–2027. doi:10.1002/ijc.22402
79. He F, Wang Q, Zheng XL, et al. Wogonin potentiates cisplatin-induced cancer cell apoptosis through accumulation of intracellular reactive oxygen species. Oncol Rep. 2012;28(2):601–605. doi:10.3892/or.2012.1841
80. Liu Y, Wang X, Li W, et al. Oroxylin A reverses hypoxia-induced cisplatin resistance through inhibiting HIF-1α mediated XPC transcription. Oncogene. 2020;39(45):6893–6905. doi:10.1038/s41388-020-01474-x
81. Lu J, Zhong Y, Lin Z, et al. Baicalin alleviates radiation-induced epithelial-mesenchymal transition of primary type II alveolar epithelial cells via TGF-β and ERK/GSK3β signaling pathways. Biomed Pharmacother. 2017;95:1219–1224. doi:10.1016/j.biopha.2017.09.037
82. Bao WA, Wang YZ, Zhu X, et al. Baicalin ameliorates radiation-induced lung injury by inhibiting the CysLTs/CysLT1 signaling pathway. Evid Based Complement Alternat Med. 2022;2022:2765354. doi:10.1155/2022/2765354
83. Chen T, Zhang P, Cong XF, et al. Synergistic antitumor activity of baicalein combined with almonertinib in almonertinib-resistant non-small cell lung cancer cells through the reactive oxygen species-mediated PI3K/Akt pathway. Front Pharmacol. 2024;15:1405521. doi:10.3389/fphar.2024.1405521
84. Hao B, Lin S, Liu H, et al. Baicalein tethers CD274/PD-L1 for autophagic degradation to boost antitumor immunity. Autophagy. 2025;21(5):917–933. doi:10.1080/15548627.2024.2439657
85. Zhang L, Wang X, Wang R, et al. Baicalin potentiates TRAIL‑induced apoptosis through p38 MAPK activation and intracellular reactive oxygen species production. Mol Med Rep. 2017;16(6):8549–8555. doi:10.3892/mmr.2017.7633
86. Yang L, Wang Q, Li D, et al. Wogonin enhances antitumor activity of tumor necrosis factor-related apoptosis-inducing ligand in vivo through ROS-mediated downregulation of cFLIPL and IAP proteins. Apoptosis. 2013;18(5):618–626. doi:10.1007/s10495-013-0808-8
87. Yang L, Zheng XL, Sun H, et al. Catalase suppression-mediated H(2)O(2) accumulation in cancer cells by wogonin effectively blocks tumor necrosis factor-induced NF-κB activation and sensitizes apoptosis. Cancer Sci. 2011;102(4):870–876. doi:10.1111/j.1349-7006.2011.01874.x
88. Shen L, Zhang LL, Li H, et al. Oroxylin A inhibits the generation of Tregs in non-small cell lung cancer. Oncotarget. 2017;8(30):49395–49408. doi:10.18632/oncotarget.17218
89. Lee E, Enomoto R, Suzuki C, et al. Wogonin, a plant flavone, potentiates etoposide-induced apoptosis in cancer cells. Ann N Y Acad Sci. 2007;1095(1):521–526. doi:10.1196/annals.1397.056
90. Cai Y, Ma W, Xiao Y, et al. High doses of baicalin induces kidney injury and fibrosis through regulating TGF-β/Smad signaling pathway. Toxicol Appl Pharmacol. 2017;333:1–9. doi:10.1016/j.taap.2017.08.003
91. Capó X, Kumar R, Mishra AP, et al. Baicalein and baicalin in cancer therapy: multifaceted mechanisms, preclinical evidence, and translational challenges. Semin Oncol. 2025;52(5):152377. doi:10.1016/j.seminoncol.2025.152377
92. Fan L, Zhang W, Guo D, et al. The effect of herbal medicine baicalin on pharmacokinetics of rosuvastatin, substrate of organic anion-transporting polypeptide 1B1. Clin Pharmacol Ther. 2008;83(3):471–476. doi:10.1038/sj.clpt.6100318
93. Li M, Shi A, Pang H, et al. Safety, tolerability, and pharmacokinetics of a single ascending dose of baicalein chewable tablets in healthy subjects. J Ethnopharmacol. 2014;156:210–215. doi:10.1016/j.jep.2014.08.031
94. Li L, Gao H, Lou K, et al. Safety, tolerability, and pharmacokinetics of oral baicalein tablets in healthy Chinese subjects: a single-center, randomized, double-blind, placebo-controlled multiple-ascending-dose study. Clin Transl Sci. 2021;14(5):2017–2024. doi:10.1111/cts.13063
95. Pang H, Xue W, Shi A, et al. Multiple-ascending-dose pharmacokinetics and safety evaluation of baicalein chewable tablets in healthy chinese volunteers. Clin Drug Investig. 2016;36(9):713–724. doi:10.1007/s40261-016-0418-7
96. Banik K, Khatoon E, Harsha C, et al. Wogonin and its analogs for the prevention and treatment of cancer: a systematic review. Phytother Res. 2022;36(5):1854–1883. doi:10.1002/ptr.7386
97. Wang PX, Mu XN, Huang SH, Hu K, Sun ZG. Cellular and molecular mechanisms of oroxylin A in cancer therapy: recent advances. Eur J Pharmacol. 2024;969:176452. doi:10.1016/j.ejphar.2024.176452
98. Wei Y, Liang J, Zheng X, et al. Lung-targeting drug delivery system of baicalin-loaded nanoliposomes: development, biodistribution in rabbits, and pharmacodynamics in nude mice bearing orthotopic human lung cancer. Int J Nanomed. 2016;12:251–261. doi:10.2147/IJN.S119895
99. Li S, Wang L, Li N, Liu Y, Su H. Combination lung cancer chemotherapy: design of a pH-sensitive transferrin-PEG-Hz-lipid conjugate for the co-delivery of docetaxel and baicalin. Biomed Pharmacother. 2017;95:548–555. doi:10.1016/j.biopha.2017.08.090
100. Wang B, Zhang W, Zhou X, et al. Development of dual-targeted nano-dandelion based on an oligomeric hyaluronic acid polymer targeting tumor-associated macrophages for combination therapy of non-small cell lung cancer. Drug Deliv. 2019;26(1):1265–1279. doi:10.1080/10717544.2019.1693707
101. Wang Y, Wang B, Xiao Y, et al. Baicalin-modified polyethylenimine for miR-34a efficient and safe delivery. Front Bioeng Biotechnol. 2023;11:1290413. doi:10.3389/fbioe.2023.1290413
102. Shi H, Wang B, Ma H, et al. Preparation of biomimetic selenium-baicalein nanoparticles and their targeted therapeutic application in nonsmall cell lung cancer. Mol Pharm. 2024;21(9):4476–4489. doi:10.1021/acs.molpharmaceut.4c00390
103. Wang W, Xi M, Duan X, Wang Y, Kong F. Delivery of baicalein and paclitaxel using self-assembled nanoparticles: synergistic antitumor effect in vitro and in vivo. Int J Nanomed. 2015;10:3737–3750. doi:10.2147/IJN.S80297
104. Joshi HA, Patwardhan RS, Sharma D, Sandur SK, Devarajan PV. Pre-clinical evaluation of an innovative oral nano-formulation of baicalein for modulation of radiation responses. Int J Pharm. 2021;595:120181. doi:10.1016/j.ijpharm.2020.120181
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Bubble Ticket Trip: Exploring the Mechanism of miRNA Sorting into Exosomes and Maintaining the Stability of Tumor Microenvironment
Wang L, Liu H, Chen G, Wu Q, Xu S, Zhou Q, Zhao Y, Wang Q, Yan T, Cheng X
International Journal of Nanomedicine 2024, 19:13671-13685
Published Date: 21 December 2024
Recent Advances in Plant-Derived Extracellular Vesicles as Nanoparticles for Cancer Drug Delivery
Zhang Q, Duan H, Yang Y, Yu H, Wang W, Li B
International Journal of Nanomedicine 2025, 20:14977-15016
Published Date: 12 December 2025
Tumor Targeting with Peptide-Drug Conjugates: Showcasing Key Progress and Hurdles
Parang K, Do T, Dinh C, Davani-Davari D, Foroughi M, Khong T, Moazen M, Nasrolahi Shirazi A
Drug Design, Development and Therapy 2026, 20:562135
Published Date: 12 February 2026
Recent Advances in Antitumor Nanomedicine Based on Covalent Organic Frameworks
Xu F, Wang R, Cheng Z, Zhang Z, Tian J
International Journal of Nanomedicine 2026, 21:378082
Published Date: 4 May 2026