Back to Journals » Clinical Pharmacology: Advances and Applications » Volume 15
Recent Advances in Messenger Ribonucleic Acid (mRNA) Vaccines and Their Delivery Systems: A Review
Authors Yihunie W ![]() , Nibret G
, Nibret G ![]() , Aschale Y
, Aschale Y
Received 26 May 2023
Accepted for publication 28 July 2023
Published 3 August 2023 Volume 2023:15 Pages 77—98
DOI https://doi.org/10.2147/CPAA.S418314
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Arthur E. Frankel
Wubetu Yihunie,1 Getinet Nibret,1 Yibeltal Aschale2
1Department of Pharmacy, College of Health Sciences, Debre Markos University, Debre Markos, Ethiopia; 2Department of Medical Laboratory Science, College of Health Sciences, Debre Markos University, Debre Markos, Ethiopia
Correspondence: Wubetu Yihunie, Department of Pharmacy, College of Health Sciences, Debre Markos University, Debre Markos, Ethiopia, Tel + 251910112391, Email [email protected]
Abstract: Messenger ribonucleic acid (mRNA) was found as the intermediary that transfers genetic information from DNA to ribosomes for protein synthesis in 1961. The emergency use authorization of the two covid-19 mRNA vaccines, BNT162b2 and mRNA-1273, is a significant achievement in the history of vaccine development. Because they are generated in a cell-free environment using the in vitro transcription (IVT) process, mRNA vaccines are risk-free. Moreover, chemical modifications to the mRNA molecule, such as cap structures and changed nucleosides, have proved critical in overcoming immunogenicity concerns, achieving sustained stability, and achieving effective, accurate protein production in vivo. Several vaccine delivery strategies (including protamine, lipid nanoparticles (LNPs), polymers, nanoemulsions, and cell-based administration) were also optimized to load and transport RNA into the cytosol. LNPs, which are composed of a cationic or a pH-dependent ionizable lipid layer, a polyethylene glycol (PEG) component, phospholipids, and cholesterol, are the most advanced systems for delivering mRNA vaccines. Moreover, modifications of the four components that make up the LNPs showed to increase vaccine effectiveness and reduce side effects. Furthermore, the introduction of biodegradable lipids improved LNP biocompatibility. Furthermore, mRNA-based therapies are expected to be effective treatments for a variety of refractory conditions, including infectious diseases, metabolic genetic diseases, cancer, cardiovascular and cerebrovascular diseases. Therefore, the present review aims to provide the scientific community with up-to-date information on mRNA vaccines and their delivery systems.
Keywords: mRNA vaccines, in vitro transcription, lipid nanoparticles, transfection efficiency
Introduction
About This Review
Currently, there is considerable research being done on messenger ribonucleic acid (mRNA) vaccines to treat cancer, infectious diseases, gene therapy, and other disorders. The favorable safety and efficacy of the already developed and authorized BNT162b2 and mRNA-127 Covid-19 vaccines have boosted the promise for future vaccines to be based on mRNA. The high molecular weight and negative charge of mRNA vaccines, their susceptibility to ribonucleases, and the existence of intracellular and extracellular barriers are only a few of the hurdles they face despite their many benefits. To overcome these challenges, chemical modifications to the mRNA molecule, such as cap structures and modified nucleosides, and using novel drug delivery systems are crucial. Hence, this review provides updated information on mRNA vaccines overview, in vitro transcription method, the role of structural elements of mRNA vaccines, challenges and adverse effects of mRNA vaccines and methods how to overcome the challenges, mRNA vaccine delivery systems, entry, and endosomal escape of nanoparticle construct of mRNA, routes of mRNA vaccine administration and application of mRNA vaccines in different diseases (disorders).
mRNA Vaccines Overview
Vaccines are classified as whole-pathogen, subunit, nucleic acid, or viral vector based on the antigen used in their synthesis.1 In 1961, messenger ribonucleic acid (mRNA) has been identified as the mechanism by which genetic information is passed from DNA to ribosomes to produce proteins.2 Moreover, mRNA vaccines can be synthesized in a laboratory using easily accessible components.3,4 However, because of the extremely unstable nature of the mRNA molecule, the first protein was synthesized in vitro from isolated mRNA in 1969.5,6
In 1987, researchers devised a highly successful approach for in vitro mRNA production by encapsulating mRNA in cationic lipids and injecting it into eukaryotic cells.4 Furthermore, activating anti-influenza cytotoxic T lymphocytes (CTLs) in the host after immunizing mice with liposome-encapsulated mRNA expressing the influenza-virus nucleoprotein (NP) marked a significant step forward in the development of the first mRNA vaccine.7 Furthermore, Pfizer-BioNTech’s (BNT162b2) and Moderna’s (mRNA-1273)8,9 mRNA vaccines have both been granted emergency use authorizations (EUA).10 These vaccines have been a huge success, with excellent protective effectiveness of more than 90%.11
Why mRNA Vaccines are Preferable to DNA Vaccines?
In comparison to DNA, mRNA therapy has numerous benefits. While viral vectors are needed for high transfection efficiency in DNA treatments, non-viral vectors (such as lipids and polymers) can be used for mRNA delivery and still achieve extremely strong transfection efficiency.12–18 Because they are generated in a cell-free environment via in vitro transcription, mRNA vaccines are also harmless. MRNA cannot induce vector- or carrier-specific immunogenicity, unlike viral vectors or virus-like particles (VLPs).15,16,19
Antigens encoded in mRNA vaccines can be expressed more expeditiously since mRNA can be functional in the cytoplasm whereas DNA must enter the nucleus and be transcribed before proteins can be created.20 In cells, mRNA undergoes a series of molecular changes, such as deadenylation and decapping, before being hydrolyzed by RNase.21 These processes ensure that exogenous mRNA treatments are only expressed briefly, which makes it to be safer.22
In vitro Transcription (IVT) Method and Role of Structural Elements of mRNA Vaccines
In vitro Transcription (IVT) Method
The in vitro transcription (IVT) technique is used to synthesize self-amplifying mRNA (saRNA) and conventional mRNA in a cell-free system. In this approach, the production of plasmid DNA (pDNA) carrying the sequence for a DNA-dependent RNA polymerase promoter (T7 or SP6), followed by the sequence matching to the mRNA construct is required.14,23–28 T7 RNA polymerase can accurately integrate pseudouridine triphosphate and other modified nucleotides and make RNAs longer than 20,000 nucleotides.29–31
The pDNA can act as a template for mRNA transcription utilizing a DNA-dependent RNA polymerase after being linearized by an enzyme. After the transcription reaction is finished, the pDNA is treated with DNase to degrade it. Enzymatic post-transcription capping and simultaneous capping by an extra cap analog in the transcription mixture are the two main capping techniques used in IVT reactions.32 Moreover, guanylyl transferase and 2′-O-methyltransferase can be employed to introduce a Cap 0 (N7MeGpppN) or Cap 1 (N7MeGpppN2′-OMe) structure, respectively.14,23,24
Poly-A-tailed IVT mRNA is typically produced in two ways. One method involves attaching a poly-A tail to the 3’ end of IVT mRNA after recombinant poly-A polymerase has synthesized the mRNAs. This method creates a varied length of poly-A and has less reliable batch controls, making it challenging to meet the standards.33 The second technique involves utilizing a DNA template with poly-T nucleotides to co-transcribe the poly-A tail amid IVT mRNA production, producing homologous mRNA products. Producing fixed and repeatable poly(A) length is a benefit of DNA template-encoded poly(A).34 However, because of its propensity for recombination, its poly-A tail’s length is shortened.35
Role of Structural Elements of mRNA Vaccines
Mature eukaryotic mRNA is composed of the 5’ cap structure (m7GpppN or m7Gp3N (N can be any nucleotide)), the 5’ untranslated region (5’UTR), an open reading frame (ORF), the 3’ untranslated region (3’UTR), and a poly(A) tail. These fundamental structural domains influence the stability, immunogenicity, and translation efficacy of mRNA vaccines.36,37 An mRNA ORF determines the target protein’s basic sequence as well as higher-order RNA structures that influence translation efficiency. Unexpectedly, mRNA coding regions capable of generating secondary structures were found to be associated with highly expressed mRNAs.38
The 5’ UTR is essential for ribosome binding and serves as the location of protein translation preinitiation complex formation.39 Moreover, according to a scanning model of RNA translation, mRNA stability, and translation efficiency are influenced by the 5’ UTR sequence and secondary structures.39–43 Even a 5’ cap-independent protein expression pathway is possible due to the presence of an internal ribosome entry site (IRES) for the encephalomyocarditis virus in the 5’ UTR.44,45 Additionally, the primary function of the 5′ UTR is to translate its downstream ORF sequence.46,47 To boost translation efficiency, the Kozak sequence is usually inserted next to the 5′ UTR sequence.24,48
Likewise, the role of the 3′ UTR is to keep mRNA stable.49,50 Most eukaryotic mRNAs have 3’ UTR mRNA degradation signals that govern the stability of mRNA. The presence of AU-rich regions in the 3’ UTR of mRNA has been shown to aid in the cleavage of the poly (A) tail during mRNA degradation.51,52 As a result, the half-life of mRNAs could be enhanced by replacing their AU-rich regions with 3’UTR sequences.53 Furthermore, the iron-responsive elements (IREs) are another essential mRNA stability-regulating segment within the 3’UTR and control mRNA translation.54
With a few exceptions (like histone), all cellular proteins that encode mRNAs consist of a poly(A) tail.55 Most actively translated mRNAs in mammalian cells have a poly(A) tail containing 100–250 adenosine residues.56 Additionally, the poly-A tail is necessary for the stability of the mRNA, translation, and recognition by the poly-A binding protein (PABP), which joins with the translation initiation complex (eIF4G) to form a loop-like conformation.55,57 The cytoplasmic translocation of mature mRNA is mediated by the poly(A) tail.58 In addition, the poly (A) tail modulates translation efficiency and mRNA breakdown.59–61 Additionally, a poly(A) tail with the proper length can increase mRNA stability and translation efficiency55,56 (Figure 1).
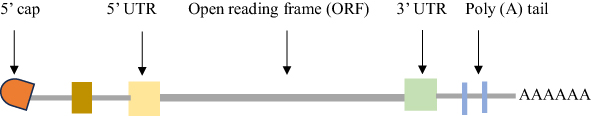 |
Figure 1 Structural elements of mRNA vaccines. |
Types of mRNA Technologies
To develop mRNA vaccines, conventional (non-replicating) mRNA (nrRNA) and self-amplifying mRNA (saRNA) have been proposed.62–64 Between its 5’ UTR and ORF, saRNA has additional virus replication components than nrRNA. Furthermore, alphaviruses, flaviviruses, measles viruses, or rhabdoviruses could be the sources of the viral replicase of saRNA. As a result, saRNA might produce a lot of antigen protein while activating the immune system quickly and effectively.14,63–65 While viral genes holding information for replication machinery proteins are intact in saRNA, genes encoding therapeutic proteins replace those for structural proteins.14,66 Additionally, saRNA vaccines can carry genetic material encoding the desired antigen in addition to other genes, such as viral RNA polymerase, which enables mRNA to multiply on its own.67,68 Safe trans-amplifying RNA (taRNA) vaccines have been optimized and produced based on saRNA technology.65 However, because of its longer length, saRNA delivery is more difficult than nrRNA.69
Non-replicating mRNA vaccinations only provide genetic information that codes for the target antigen.70 Also, the use of a simple structure and shorter-length RNA molecule is one of the advantages of nrRNA vaccines. Furthermore, a modified or tweaked mRNA can have significantly increased its efficacy.71 Conventional mRNA has several advantages over saRNA, including its smaller size (2–3 kb vs 10 kb), lack of viral genes, which reduces the risk of showing unwanted immunogenicity, its easiness and scalable manufacturing techniques, as well as the ease with which its sequence can be altered to improve therapeutic efficacy and minimize any unwanted effects.14,70
Challenges and Adverse Effects
Due to mRNA’s physical properties such as high molecular weight, negative charge, vulnerability to ribonucleases (RNases),15,21,70,72–76 and the presence of extracellular and intracellular barriers,21,77 it is difficult for mRNA to be successfully uptaken into cells and translated to targeted antigens. Furthermore, after entry, enormous amounts of mRNA are ambushed in endosomes and cannot reach into the cytoplasm to perform its functions.75,78,79 Furthermore, as indicated by a short half-life (5 min) in sera, IVT mRNA transcripts are unstable and extremely sensitive to nuclease destruction.76,80
Despite, immunogenicity is undesirable for several mRNA uses, such as protein replacement therapy and genome editing, it can be useful for vaccination techniques and may even replace the use of adjuvants.37,81–84 Unmodified mRNA may activate innate immune systems through the endosomal recognition of pattern-recognition receptors (PRRs), such as Toll-like receptors (TLRs)85–87 and the RIG-1- like receptor families, like RIG-1, MDA5, and LGP2, resulting in the production of pro-inflammatory cytokines and type I interferons.15,88–90
High frequencies of adverse reactions to vaccinations, including pericarditis, myocarditis, inflammation of the nervous system, and autoimmune hepatitis, had been associated with mRNA side effects.91–94 Even though, studies suggested that these side effects were unrelated to the vaccine itself.95 Moreover, in young adults and male adolescents who received BNT162b2 and mRNA-273 Coronavirus disease 2019 (COVID-19) vaccines, cardiomyopathy, myocarditis, and pericarditis were reported within a week of the second vaccination.96,97 Furthermore, infarction, allergies, heart failure, and renal failure are a few possible side effects of mRNA vaccinations.98 In addition, studies reported that the most common cause of acute myocarditis is a viral infection. For example, healthy people who have received smallpox or influenza vaccines have been known to manifest the adverse effect.99
According to studies, the bulk of adverse effects are caused by lipid nanoparticles (LNPs) ingredients like PEG and ionized lipids.100,101 The necessity to improve the LNP delivery platform is highlighted by the reports of negative effects brought on by LNPs for mRNA-based COVID-19 vaccines.102,103 PEG-lipids may cause allergic responses by activating the complement system.75,104,105 Additionally, by hastening blood clearance, anti-PEG antibodies may cause fast systemic elimination of subsequently administered PEGylated nanoparticles.104,105 In addition, rodents have been shown to suffer liver and lung damage as a result of LNP administration in vivo,106,107 which could be explained by the delivery of LNP materials’ cytotoxicity and the production of pro-inflammatory factors.108,109
How to Enhance the Stability and Suppress the Immunity of mRNA Vaccines?
The amount of mRNA that degrades can be considerably decreased by adding a 5’-cap, modifying nucleosides, adjusting 30-poly(A) tail length and structure, and optimizing nucleoside sequences.36,110,111 To boost mRNA’s stability and stop it from degrading, LNPs and alternative delivery systems like polymers, peptides, and cationic nano-emulsions (CNEs) can be used.67,112 Moreover, mRNA-based vaccines can be lyophilized (freeze-dried), making them more stable and preserving their biological action.113 Additionally, Kariko and colleagues were the first to change certain nucleosides (cytidine and uridine) with 5-methylcytidine and pseudouridine, respectively, to make the resulting mRNA molecules more stable intracellularly and less immunogenic.37,114–116
To overcome immunogenicity issues, achieve sustained stability, and achieve effective and precise protein production in vivo, chemical modifications to the mRNA molecule, such as cap structures and modified nucleosides, are essential.70,117 In 2005, Karikó and colleagues revealed that mRNA generated with modified uridine might withstand immune system recognition and destruction, significantly improving mRNA stability and immunogenicity in vivo.115 The uses of mRNA technology in the biomedical field are expanding because of improvements in delivery technology and the application of modified nucleosides to escape innate immune recognition.21 By reducing the usage of uridine in the codons,37,118–120 and altering the nucleotides used in IVT mRNA,21,28,37,70,76,115,121,122 it is possible to block TLR recognition of mRNA. 5-methylcytosine (m5C), 5-methyluridine (m5U), 2-thiouridine (s2U), or pseudouridine (ψ) modifications such as 1-methyl pseudouridine (m1 ψ) are the most widely used alterations.123 One of these changes, the substitution of pseudouridine for uridine, has been demonstrated to improve mRNA efficacy and decrease immunogenicity.39,124 In place of each uridine residue in the coding region and UTRs identified by the ribosome, the Pfizer/BioNTech BNT162b2 mRNA contains N1-methyl pseudouridine (m1ψ).25,117,123
By rigorously purifying mRNA using high-performance liquid chromatography (HPLC), which can eliminate the aberrant RNAs produced in the IVT reaction, immunogenicity could be further reduced.37,125,126 The constituents of the LNPs were being altered by researchers to increase vaccine effectiveness and reduce side effects.100,101,127 Furthermore, biodegradable lipids may be used to increase the biocompatibility of lipid nanoparticles.128–132
Purification of IVT mRNA
Abortive initiation products and double-stranded RNA produced by DNA-dependent RNA polymerases can trigger the production of type I IFN and inflammatory cytokines when PRRs bind to them.133 DNase treatment can easily remove pDNA, however, several chromatographic methods are available to get rid of the remaining impurities.134 While polyacrylamide gel electrophoresis can be used to remove short RNA moieties.135 Chromatography, namely HPLC, is the sole method that can purge contaminants from longer mRNA preparations.82 As a result, great translatability is made possible by mRNA purification via HPLC without inducing IFN1 and proinflammatory cytokine responses.70,76,82,136 For large-scale mRNA production and Good Manufacturing Practice (GMP) procedures, purification using fast protein liquid chromatography (FPLC) or HPLC may be carried out.82,137–140
mRNA Vaccine Delivery
Viral and non-viral vector delivery techniques have been used to deliver mRNA vaccines.141 To increase the safety and effectiveness of mRNA-based immunotherapy, delivery systems can be modified to offer tissue or cell specificity.142 Bangham made the initial discovery of lipid-based systems in the 1960s when cationic LNPs (cLNPs), also known as liposomes, were seen to generate vesicles spontaneously in aqueous solutions.143–146 Doxil®, a liposomal-formulated doxorubicin, was authorized in the US thirty years later. Since then, the FDA has approved several liposome146 and LNP medications (comprising ionizable cationic lipids, iLNPs) for clinical use.147,148
To traverse membrane lipids and efficiently deliver RNA vaccines into the cytosol, a variety of vaccine delivery techniques (including protamine, LNPs, polymers, nanoemulsions, and cell-based administration) were optimized.149 Therefore, both viral and non-viral delivery systems must be used to prevent RNases from degrading mRNA and to improve their intracellular effectiveness. Viral vector delivery system may result in the induction of immune response to the antibody, vector pre-existing immunity inhibiting transduction, potential safety issues with long-term expression, and risk of insertional mutagenesis.15,150 Hence, in this review, non-viral delivery methods have been discussed in detail.
Protamine
Cell-penetrating peptides (CPPs) showed a good safety profile and effective transfection capabilities.151–153 Peptides have also been employed for mRNA administration because they include cationic or amphipathic amine groups, such as arginine, that can electrostatically attach to negatively charged mRNA and form nano complexes.154 Moreover, a cationic peptide called protamine can stop lysosomal degradation while RNA is being delivered. It has been demonstrated that protamine-based delivery activates TLR 7 to cause a potent immunological response.155,156
Protamine, an arginine-rich cationic peptide, can attach to mRNA and efficiently transfer it into the cytosol.157 Hence, it was utilized in the development of the self-adjuvanted RNActive vaccination platform, which has proven effective against several infectious illnesses and malignancies.115,156,158,159 Furthermore, protamine protects mRNA from being degraded by serum RNases151 and is found to guard against severe storage conditions for the mRNA rabies vaccine.160
To elicit an immunological response against the rabies virus, CureVac investigated the protamine-mRNA combination in 2016. In that investigation, the mRNA encoding the non-replicating rabies glycoprotein (RABVG) was tailored to induce strong virus-neutralization in mice and domestic pigs.158 A brand-new lipid/protamine/mRNA nanoparticle technology was recently developed and widely used for systemic tumor administration. In this technique, 1,2-dioleoyl-3-trimethylammonium propane (DOTAP) liposomes have been tested to enclose protamine-complexed mRNA before being coated with 1, 2-Distearoyl-sn-glycero-3-phosphoethanolamine-Poly(ethylene glycol) (DSPE- PEG) and DSPE-PEG-anisamide.161
Lipid Nanoparticles (LNPs)
Due to their ability to preserve mRNA from enzymatic cleavage, efficiently carry mRNA into cell cytoplasm,93,129,162 have minimal immunogenicity, are biocompatible, and have a high encapsulation rate,163 lipids have been used for exogenous mRNA administration. In addition, LNPs are lipid-based spherical vesicles and can self-assemble into precise structures like cell membranes.164 Furthermore, LNPs are lipid-based, and nanoscale carriers that can effectively transfer mRNA intracellularly and safeguard it from RNAase during systemic circulation.165 Moreover, LNPs are now the most cutting-edge method of administering mRNA vaccines.129,149,166–169 (Figure 2)
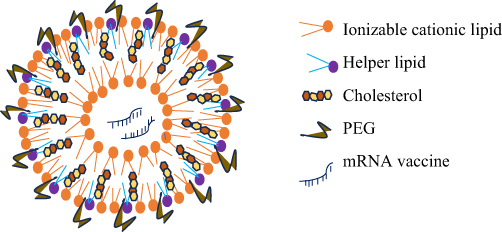 |
Figure 2 mRNA encapsulated in Lipid nanoparticle. PEG, Polyethylene glycol. |
Modifying the lipid structure130 and particle surface has been regarded as a strategy to increase the efficient delivery of mRNA into the cytosol.14,170 It was discovered that LNPs having multilamellar, faceted, and lamellar lipid phases have better mRNA transfection effectiveness.171,172 N-[1-(2,3-dioleyloxy) propyl]-N,N,N trimethylammonium chloride (DOTMA), for example, was a first-generation permanently charged lipid that was toxic,173 subpar, and relied on non-scalable methods. At an acidic pH (where the amino lipids were positively charged), in the presence of ethanol, ionizable amino lipids were employed to create nucleic acids.174,175
Eminently, LNP delivery materials are employed in the existing mRNA vaccines approved by FDA.57,176 The numerous advantages of lipid-nanoparticle-based mRNA delivery systems, such as their high stability, transfection efficiency, efficacy, safety, and low-cost production techniques, have facilitated the rapid development of mRNA vaccines and medicines, providing a powerful disease-fighting tool.177
Components of LNP
LNPs are made up of a cationic or a pH-dependent ionizable lipid layer, a PEG component, phospholipids, and cholesterol.14,165,178–183 The lipid-anchored PEG ensures vial and storage stability, the ionizable lipid is essential for cellular absorption and endosomal escape, enabling mRNA to enter into the cytosol, and the phospholipid and sterol are essential for the stabilization of the LNP.182,184,185 The ratio of the components can be changed depending on the target tissue, and the lipid content can also be changed to alter the physical characteristics of LNPs like particle size, shape, encapsulation effectiveness, and surface charge.186,187 Numerous encapsulation devices can be created using lipids and lipid-based nanoparticles, including liposomes, LNPs, microbubbles, micelles, lipid implants, and emulsions.71,165,188–193
Cationic Lipids
Cationic lipids either have a quaternary nitrogen atom that permanently gives them a positive charge or a primary amine that gives them a positive charge at or below physiological pH. However, it was discovered that cationic lipids with such a long-lasting positive charge were more toxic, less effective, and non-biodegradable.17,194–197 The addition of structural lipids like DOPE may decrease cytotoxicity while increasing endosomal release. Among the earliest LNP preparations that have been successful in the in vivo translation of mRNA is Lipofectin, which is made by combining DOTMA and DOPE.198–200 The systemic toxicity of this combination, however, made it ineffective.201
Whereas net positively charged complexes have been demonstrated to improve mRNA stability in vitro, cationic complexes may interact with negatively charged serum proteins, resulting in clumps, clots, and rapid clearance.202–204 Although there are certain disadvantages to utilizing cationic lipids, the positive charge traps nucleic acids very well. This approach was used to create pH-sensitive ionizable cationic LNPs for enhanced RNA delivery.194
Ionizable Lipids
Cullis developed the first pH-responsive cationic lipid (ionizable lipids (ILs)) in the early 1990s, which gained a net positive charge in an acidic pH while remaining neutral in a physiological pH. The reticuloendothelial system (RES) cannot break down ILs because of their pH-sensitive characteristics, which prolongs their half-life.205–207 ILs have polar head groups that contain ionizable amines, a hydrophobic tail that promotes self-assembly, and linkers that join the head groups to the hydrophobic tail. After being carried into an endosome, they are assumed to be ionized negatively once more upon acidification, which helps to build hexagonal phase structures and, eventually, makes it easier for mRNA to escape from the endosome and enter the cytosol.70,129,179,201,208–213
Nanoparticles made with ILs have a low positive charge density in the bloodstream, resulting in better biocompatibility and less off-target accumulation.214 In the COVID-19 vaccinations, the cationic lipids employed are SM-102 and ALC-0315 in the Moderna and Pfizer/BioNtech vaccines, respectively.215 Ionizable lipids’ enhanced capacity for biodegradation Due to quick metabolic breakdown and clearance, there is less exposure to adjacent tissues and a reduction in inflammation at the injection site.216 Additionally, ILs have a better safety profile since they are less likely to stimulate the immune system or interact with serum proteins.17,78
Biodegradable functional groups were employed in the subsequent synthesis of lipids to speed up clearance. The incorporation of ester moieties is one method of increasing biodegradability.217–219 The incorporation of disulfide bonds into the backbone of lipids is another method of imparting biodegradability. Disulfide bonds are bio-reduced in the cell by glutathione (GSH) or other disulfide-reductases.220 Further modifications to ionizable lipid structures result in altered physicochemical properties that can influence the selective delivery of mRNA to different tissues.221
Cholesterol
Cholesterol can improve particle stability and affects the efficacy and biodistribution of in vivo mRNA injection. C-24 alkyl phytosterols, for example, improved the delivery effectiveness of LNP mRNA in vivo.171 By preventing excessive amounts of endogenous cholesterol from being sequestered inside LNPs while they are in circulation, cholesterol helps maintain stability.194,222 When compared to standard cholesterol, nanoparticles containing a -sitosterol substitute increased endosomal entry of mRNA in the cytoplasm by a factor of ten.170,171,223 Therefore, cholesterol and its derivatives play a crucial role in the general stability of LNPs in circulation and may facilitate endosomal escape, improving mRNA entry into the cytoplasm.224
Furthermore, cholesterol is required for LNP to transition from the lamellar to the hexagonal phase. The hexagonal phase is required for mRNA to be released from LNPs and transported over the endosomal membrane into the cytosol.225 The fluid phospholipid bilayer was strengthened by cholesterol and LNPs’ content loss was reduced.226 When employed in the optimum concentration, it may also aid in the fusing of the membrane for LNPs and gene transfer.227
PEG
It has been widely used to layer PEG on lipid carriers to slow down aggregation and lengthen blood circulation.228–231 Additionally, lipid-anchored PEGs primarily form a barrier on the LNP surface, sterically stabilizing the LNP and decreasing specific protein binding.179 Moreover, PEGylated nanoparticles are widely referred to as stealth nanoparticles due to their ability to avoid opsonization by serum proteins and detection by the reticuloendothelial system (RES).172,232–235 PEG-lipids restrict LNP uptake while decreasing opsonization by serum proteins and reticuloendothelial clearance, extending LNP circulation lifetime.236 Furthermore, By supplying a hydrophilic exterior coating, PEG regulates the lipid nanoparticle in many ways, including nanoparticle formation, inhibiting nanoparticle aggregation, and extending particle blood circulation.,104,194,237–241 and by avoiding their physical aggregation in solution, may potentially improve the LNPs’ storage stability.201,208
It’s also crucial to take into account the so-called PEG dilemma, which is the decreased fusogenicity of PEG lipids and may prevent mRNA from being released from endosomes. A practical method for effective mRNA intracellular delivery is cleavable PEGylation.237,241–245 PEG on LNP surfaces has the potential to trigger anti-PEG IgM antibodies to be induced, especially after repeated doses.246 PEG-lipid, unlike other components of LNP, is engineered to eventually dissociate and shed the PEG to avoid the potential generation of PEG-specific antibodies that would cause rapid systemic clearance of successive doses of PEGylated nanoparticles via the accelerated blood clearance (ABC) phenomenon.105
Helper Lipids
Phospholipids, also known as helper lipids, are frequently utilized to give LNP structure, which enhances formulation stability and may facilitate endosomal escape.181,182,247,248 Phospholipids, such as DSPC and DPPC, are typically neutral and provide bilayer structural stability to LNPs. Additionally, phospholipids contribute to the fusogenicity and biodistribution of LNPs.247 Moreover, LNPs, which were incorporated in the mRNA-1273 and BNT162b, might have their structural integrity stabilized by the DSPC.249–251 In addition, DOPE was employed for mRNA and siRNA administration in vivo as an alternative to DSPC since it could destabilize the endosomal members and encourage mRNA entry into the cytosol.182 Endosomal escape is made possible by phospholipids’ induction of a transition from lamellar to hexagonal endosomal architecture by disrupting the lipid bilayer.252
Polymers
Early gene delivery attempts relied heavily on poly (ethylene imine) (PEI), poly(L-lysine) (PLL), and poly (amidoamine) (PAMAM). Though PEI was the only polymer employed for mRNA delivery.253 Although optimized PEI structures have high cationic charge density, they are toxic.254 In 1987, polylysine (PLL) was announced as the first non-viral cationic polymer vector to efficiently transfect plasmid DNA.255 Because of their high net positive charges and inability to dissolve in physiological conditions, they could cause harmful levels of bioaccumulation, raising concerns about their limited efficacy and potential toxicity.224
Several polycationic systems were used to enhance the entry of mRNA into the cytosol, including DEAE (diethylamino-ethyl)-dextran, DOPE (1,2-dioleoyl-3-phosphoethanolamine), poly L-lysine, PEI (polyethylene imine), and DOTAP.36 Self-amplifying -mRNA nanoparticles were also delivered using chitosan and PEI.256 Chitosan has several advantages, such as biodegradability, biocompatibility, and cationic charge that allows nucleic acid binding, but it also has disadvantages, such as poor water solubility and limited target capability.164
A polyethyleneimine copolymer (PVES) treated with vitamin E succinate is used in the self-assembled polymeric micelle delivery technique. When VE binds to PEI, a conjugated polymer capable of self-assembling into stable micelles is formed.257,258 Charge-altering releasable transporters are originally positively charged polymers that can efficiently load mRNA and improve physical characteristics by degradative, charge-neutralizing intramolecular rearrangement, releasing functional mRNA, and translating protein in cells.259
PEI’s toxicity and transfection effectiveness both rise as its molecular weight does. To address such limitations, different modifications to PEI have been researched, including ones that use polysaccharides and polyethylene glycol to boost biocompatibility and transfection effectiveness and wrap PEI in neutral or anionic liposomes to lessen non-specific adhesion.260 Blakney et al created pABOL, a bioreducible, cationic polymer that improved transfection efficacy but not cytotoxicity at higher molecular weights.261
Nanoemulsions
The emulsions are often water-in-oil emulsions made of squalene, sorbitan trioleate, polysorbate 80, and DOTAP, much like the licensed MF59 adjuvant.215 Above all, the main benefit of this platform is that MF59 is safe.262,263 CNEs (cationic nanoemulsions) were proposed as a possible means of delivering nucleic acids in 1990.264 Moreover, the presence of cationic lipids in the formulation is essential for nucleic acid complexation via electrostatic interactions, which also promotes nucleic acid stability and transfection efficiency while protecting them from nuclease degradation.265 Anderluzzi et al discovered that CNE induced the highest number of antibodies against rabies when compared to DOTAP polymeric nanoparticles, DOTAP liposomes, and DDA liposomes.266
Moreover, Gennova Biopharmaceuticals Ltd in conjunction with HDT Biotech Corporation developed a lipid inorganic nanoparticle, which is called LION®, for the delivery of SARS-CoV2 vaccine candidate HGCO10 (self-amplifying RNA). Genova reported LION ® is a very stable cationic lipid (DOTAP)-squalene emulsion akin to CNEs with 15 nm superparamagnetic iron oxide (Fe3O4) nanoparticles (SPIO) implanted in the hydrophobic oil phase (which offers therapeutic and imaging functions). When stored between 4° and 25 °C, this formulation was found to have colloidal stability for at least three months.267,268
Ex vivo Loading of mRNA to Dendritic Cells
This method can be accomplished using either mild electroporation269 or lipid-derived carriers.270 By reducing potential off-target effects, electroporation can boost mRNA transport to the target cells, resulting in a reduction in the amount of mRNA that is required.166,271,272 Additionally, this strategy is generally employed for cancer immunotherapy since the majority of ex vivo loaded dendritic cells demonstrate cell-mediated immunity.273 Nanoparticle formulations were required to improve dendritic cell targeting.274
Dendritic cells used in cancer immunotherapy may be transfected with total tumor RNA or tumor-associated antigens (TAAs) encoding mRNA.272 Yet, the drawbacks of this strategy include a lack of known TAAs for various malignancies, and selecting TAAs may be difficult because not all recognized TAAs generate antitumor immunity. Furthermore, TAA mRNAs were found to induce antitumor immunity in experimental studies.275
Entry, and Endosomal Escape of Nanoparticle Construct of mRNA
The internalization of RNA-loaded lipid-based nanoparticles which involves endocytosis, micropinocytosis, macropinocytosis, and phagocytosis is mediated by caveolae and clathrin.184,276–280 Because clathrin-mediated uptake is thought to be faster than caveolin-mediated uptake, targeting the caveolin pathway should result in more effective delivery and more time for the drug to escape endosomes than clathrin-mediated uptake, which can cause significant buildup in late endosomes and lysosomes.281,282 With the aid of nanoparticles, several processes are required for mRNAs to enter the cytoplasm, including endocytosis, lysosomal escape, and mRNA release.162 The availability of mRNA in the cytoplasm may be increased by stimulating endosomal escape and scavenger receptor activation to increase mRNA absorption.73
In the endosomes, nanoparticles undergo a pH gradient, beginning with neutral extracellular pH (7.4) and moving to gradual acidification in early endosomes (pH 6.3), late endosomes (pH 5.5), and finally lysosomes (pH 4.5).283 Following cellular uptake, mRNA must escape endosomes to reach the cytosol (pH 7.2) for mRNA translation, which is a limiting step for productive mRNA delivery. For example, only 1–2.5% of mRNA was detected in the cytosol after transfection of human epithelial cells with mRNA, and this varies by cell type.129,184,284,285 Additionally, the methods utilized to avoid this terminal degradation rely on units that are activated by acidic pH.283,286
Furthermore, ionizable units and/or fusogenic lipids in mRNA nanocarriers destabilize the endosomal membrane, enabling mRNA to enter into the cytosol.285 As a result, endosomal escape, which has a high association with transfection effectiveness, is another crucial step in the delivery of mRNA to ribosomes.287 Three scenarios are widely accepted among numerous techniques to induce nanocarrier endosomal escape: (1) destabilization of the endosomal membrane, (2) osmotic rupture of the endosomes via the “proton sponge” effect, and (3) endosome rupture via particle swelling. Moreover, distinct nano constructs use distinct pathways, such as pH-responsive endosomal escape and proton sponge effect.288,289
In addition, pH-responsive endosomal escape results from conformational changes brought on by protonation or the breakdown of a polymer link at endosomes.290 Endosomal escape may be facilitated by interactions between cationic lipids and the negatively charged endosomal membrane. Following the protonation of its head group under acidic conditions, DOPE turns fusogenic, causing the formation of a hexagonal (HII) phase and momentarily destabilizing the endosomal membrane. The proton sponge effect, in which endocytosed polyplexes produce osmotic swelling of the endosome due to proton influx and eventually rupture the endosome, is hypothesized to be how cationic polyplexes undertake endosomal escape.291 Flow cytometry appears to be the most efficient and informative tool for studying the cellular uptake and trafficking of nanoparticles.292 Using nanoparticles that have been fluorescein-labeled, different information about particle localization on the cell surface, inside the cell, or into the acidic compartment, where the acidic pH quenches fluorescein’s fluorescence, can be learned.293,294
Routes of Administration of mRNA Vaccines
The method of administration and formulation of mRNA vaccines have a crucial role in regulating the rate and amount of antigen expression as well as the effectiveness of the immune response.34,291 The route of administration can have a considerable impact on the organ distribution, expression kinetics, and therapeutic effects of LNP-mRNA formulations.165,295–298 The most popular routes of mRNA vaccines administration are intramuscular (IM), subcutaneous (SC), intradermal (ID), and intravenous (IV).10,17,70,165,253,291,299–301
The highest amount of encoded protein synthesis in the body can be achieved with IV injections of mRNA therapies. Moreover, the liver is typically the target of intravenous mRNA therapies, which effectively transfect endothelial, Kupffer, and hepatocyte cells.17,75,302 It is possible to generate adaptive immune responses from IV–injected mRNA vaccines by transfecting the spleen as a site of transfection.274,303 Some of the disadvantages of I, V administration include impediments to vaccine transport in the bloodstream caused by plasma proteins, enzymes, and mechanical forces.304 Additionally, systemic adverse effects such as spleen damage and lymphocyte depletion may be brought on by the mRNA and its delivery vehicles.21 LNP-mRNA vaccination IV injections are less frequent due to the possibility of systemic side effects. Infusing immunogenic material into the bloodstream may cause a cytokine storm, or the overwhelming synthesis of cytokines, which can result in shock and death.305
The most popular method of administering vaccines to patients is through intramuscular injection.306,307 After IM injection, the LNPs are efficiently taken up by the myocytes before the cytoplasmic release of the mRNAs for S protein translation.308 IM injection allows for a higher volume to be injected than the ID and SC routes, which may result in fewer unpleasant injection site reactions but increased systemic absorption.130 In addition, SC injection-based mRNA vaccines allow for a relatively higher injection volume, which minimizes pressure and pain at the injection site.309 However, one downside of SC injection is that the rate of absorption is slow, and inadvertent mRNA destruction may occur.309,310 Furthermore, with mRNA-LNP vaccines, the intradermal (ID) method of delivery has been found to successfully produce a Th1-type immune response and cytotoxic T-cell activation.311,312 According to certain studies, IM and ID delivery of LNP-mRNA vaccines led to longer-lasting protein expression than IV.70,313,314
Therapeutic protein augmentation in certain organs, such as heart,315,316 eyes,317–319 and brain, 320,321 is made possible by local injection of LNP-mRNA compositions. Additionally, immune stimulator-coding LNP-mRNA formulations can be injected directly into cancer tissue by intratumoral injection.322–325 It has been noted that the intranodal (IN) injection of naked mRNA-encoding antigens causes a strong T-cell response.326
Utilizing the potential of mucosal immunity, intranasal (IN) vaccine delivery to the mucosal layers, like the nasal and pulmonary mucosa, is a practical, noninvasive method of vaccine administration.253,289,327,328 Pathogen-specific antibodies that are produced in the mucus via mucosal vaccination can neutralize pathogens at the earliest stages of infection.329 Furthermore, mucosal delivery of mRNA vaccines can result in the release of immunoglobulin A (IgA), which can neutralize bacterial toxins and viruses.330 M cells move the LNPs from the nasal epithelium to the underlying nasal-associated lymphoid tissue, which is home to significant numbers of B cells, T cells, and DCs.327
Stability, and Storage of mRNA Vaccines
A cold chain is often required for vaccine storage and shipping, but the supply chain for mRNA vaccines may require an even colder cold chain331 than the conventional vaccines which can commonly be stored at 4–8°C.332 Spikevax and Comirnaty, two currently licenced COVID-19 mRNA vaccines, require storage temperatures of −20°C and between −80°C and −60°C, respectively.333
Inadequate mRNA storage can lead to chemical instability through reactions including oxidation and hydrolysis, changing the physical properties of the therapeutic product and perhaps its functionality.334,335 Furthermore, pH, buffer composition and concentration, metal cation presence, non-viral vector formulation composition, and physiochemical properties all have a major impact on stability.333 Currently, a cryoprotectant is used to store LNP-mRNA medications and vaccines for an extended time to avoid aggregation. Additionally, non-permeable cryoprotectants like sucrose and trehalose are used to permit vitrification of the surrounding aqueous solution.336,337
For mRNA-based vaccines, lyophilization, also known as freeze drying, is a common alternative storage technique that may support long-term stability at higher temperatures.338,339 Lyophilized mRNA-LNPs were stable for 6 months at 4°C and 3 months at room temperature.340 An interim ultra-cold chain storage device called Cryo-Vacc, created by the South African company Renergen, has been developed.66 Because lyophilization is an expensive, time-consuming, and high-energy process, additional drying techniques including spray drying and supercritical drying should also be investigated.201
According to a study on the long-term storage conditions of mRNA-loaded lipid materials by Zhao et al, lipid-like nanoparticles (mRNA-LLNs) maintained in an aqueous solution undergo size changes and lose efficacy in vivo.341 In comparison, freeze-dried mRNA LLNs kept their efficiency after being lyophilized with a 5% cryoprotectant solution and switched the preferential organ absorption from the liver to the spleen after being lyophilized with a 20% cryoprotectant solution.289
Application of mRNA Vaccines
mRNA-based medicines are projected to be effective treatments for a wide range of refractory disorders, including infectious diseases, metabolic genetic diseases, cancer, cardiovascular and cerebrovascular diseases, and others.342 mRNA vaccines have been extensively researched over the last two decades for infectious disease prevention as well as cancer prophylaxis and therapy.70,343
The delivery of tumor-associated antigens (TAAs) expressing mRNA is the most fundamental application of mRNA vaccines in oncology.343,344 In addition to being utilized in cellular therapies to ex vivo transfect patient-derived cells before reinserting transfected cells into patients, mRNAs may be employed therapeutically to immunize patients. The TAA of interest is expressed by patient-derived DCs after they have been transfected with the mRNA encoding it, and TAA-derived peptides are then presented to stimulate antigen-specific T cells in vivo.345,346
Prophylactic or therapeutic mRNA vaccines against infectious illnesses could be produced. mRNA vaccines that express an infectious pathogen’s antigen elicit both strong and powerful T cell and humoral immune responses.20,70,163,347 Vaccines are made from in vitro transcribed mRNAs encoding viral antigens, whereas immunotherapy is made from mRNAs encoding antibodies or immune modulators. Because of their interactions with cellular RNA sensors such as Toll-like receptors (TLRs), PKR, and RIG-I, some structural characteristics of mRNA have been identified as immunostimulatory.348–350 The extraordinarily rapid development of mRNA vaccine candidates for the recent global COVID-19 pandemic highlights its clinical value.351
In pre-clinical and clinical investigations, IVT mRNAs are being examined to supplement missing or faulty proteins caused by hereditary diseases, or where the delivered protein could have a therapeutic effect. Among the studies are the use of IVT mRNA to cure hepatic disorders,352 regenerate cardiac tissues,353 and generate human stem cells354 (Table 1).
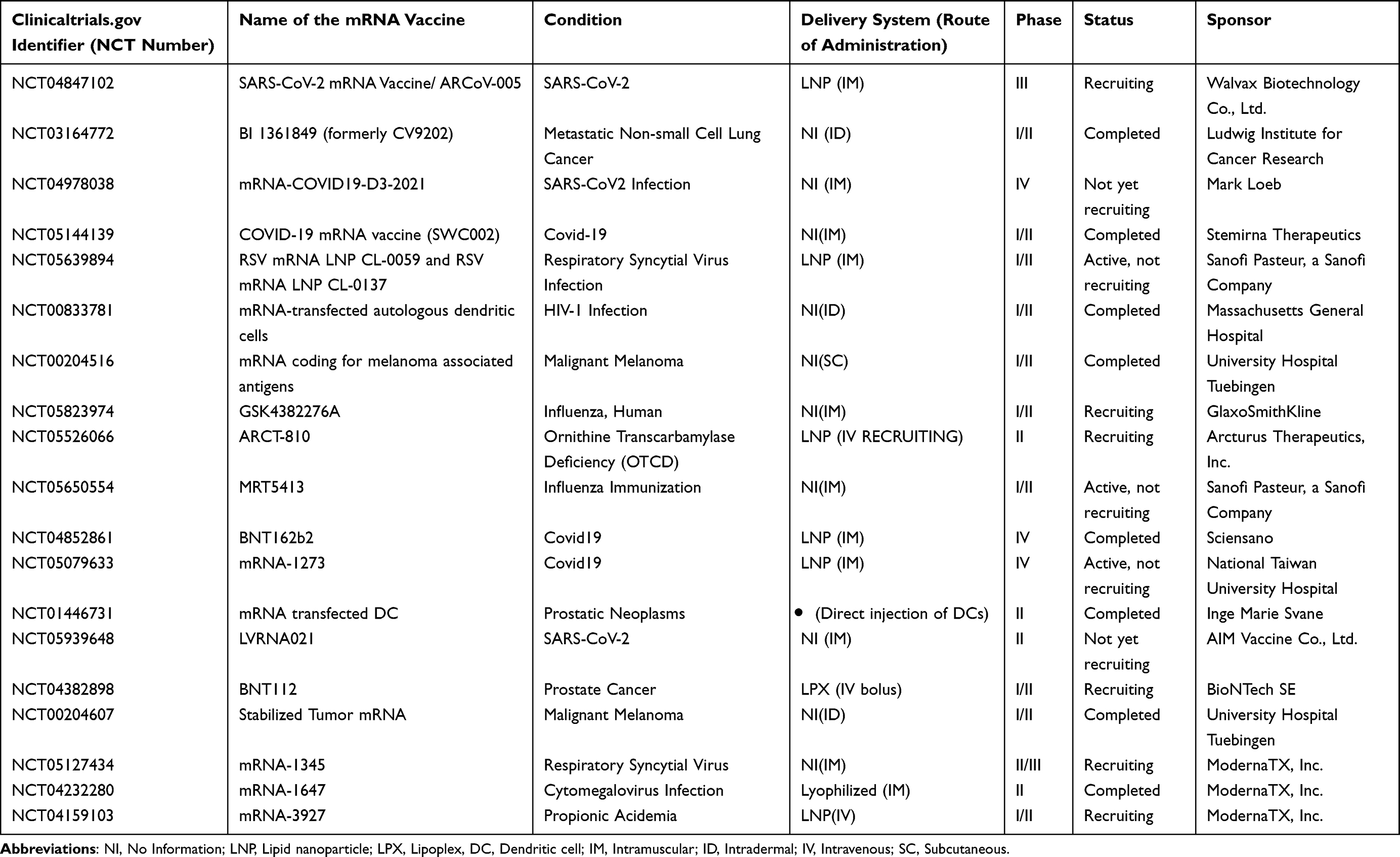 |
Table 1 Examples of mRNA vaccines (candidates) for infectious diseases, cancer, and other disorders currently in clinical trials.355 (as of July 14, 2023) |
Future Direction and Conclusions
Even though mRNA vaccines may be quickly made with commonly available materials and are relatively safer, there are still numerous difficulties. Bio-incompatibility, ineffective targeted delivery, poor transfection efficiency, immunogenicity, and instability are still a problem. Besides modification of the structure of the mRNA molecule, much emphasis has to be given to the delivery systems too.
Most of the adverse events observed from mRNA vaccines were also reported from RTS, S malaria vaccine, and other vaccines were also shared by other vaccines.356 Therefore, as far as mRNA vaccines are the current choice of vaccine development, further research is required to optimize the in vitro transcribed RNA vaccine and delivery materials, notably lipid nanoparticles, to address the aforementioned difficulties. Use of biodegradable lipids, changing (optimization) of the four components of LNPs, use of effective purification techniques, chemical modifications to the mRNA molecule, such as cap structures and modified nucleosides, choice of appropriate delivery materials, use of cryoprotectants and lyophilization technique, and appropriate implementation of cold chain requirements of mRNA vaccines are among the strategies to increase the effectiveness of mRNA vaccines.
Abbreviations
PEG, Polyethylene-glycol; cLNPs, Cationic Lipid Nanoparticles; CNEs, Cationic nano-emulsions; CPPs, Cell-penetrating peptides; CTLs, Cytotoxic T lymphocytes; EUA, Emergency Use Authorization; HPLC, high-performance liquid chromatography; IRES, Internal ribosome entry site; IREs, iron-responsive elements; IRES, Internal Ribosome Entry Site; IVT, In Vitro Transcription; LNPs, lipid nanoparticles; mRNA, Messenger Ribonucleic Acid; nrRNA, non-replicating mRNA; PABP, poly-A binding protein; pDNA, plasmid DNA; PRRs, Pattern-Recognition Receptors; saRNA, self-amplifying mRNA; TLRs, Toll-like receptors; VLPs, virus-like particles.
Disclosure
The authors declared that they have no competing interests in this work.
References
1. Karch CP, Burkhard P. Vaccine technologies: from whole organisms to rationally designed protein assemblies. Biochem Pharmacol. 2016;120:1–4.
2. Brenner S, Jacob F, Meselson M. An unstable intermediate carrying information from genes to ribosomes for protein synthesis. Nature. 1961;190:576–581.
3. Centers for disease control and prevention. Understanding mRNA COVID-19 vaccines; 2021.
4. Thran M, Mukherjee J, Pönisch M, et al. mRNA mediates passive vaccination against infectious agents, toxins, and tumors. EMBO Mol Med. 2017;9(10):1434–1447.
5. Cao L, Zheng ZC, Zhao YC, et al. Gene therapy of Parkinson’s disease model rat by direct injection of plasmid DNA–lipofectin complex. Hum Gene Ther. 1995;6(11):1497–1501.
6. Xia S, Zhang Y, Wang Y, et al. Safety and immunogenicity of an inactivated SARS-CoV-2 vaccine, BBIBP-CorV: a randomized, double-blind, placebo-controlled, Phase 1/2 trial. Lancet Infect Dis. 2021;21(1):39–51.
7. Martinon F, Krishnan S, Lenzen G, et al. Induction of virus‐specific cytotoxic T lymphocytes in vivo by liposome‐entrapped mRNA. Eur J Immunol. 1993;23(7):1719–1722.
8. Barda N, Dagan N, Lipsitch M, et al. Effectiveness of a third dose of the BNT162b2 mRNA COVID-19 vaccine for preventing severe outcomes in Israel: an observational study. Lancet. 2021;398(10316):2093–2100.
9. Tang P. BNT162b2 and mRNA-1273 COVID-19 vaccine effectiveness against the SARS-CoV-2 Delta variant in Qatar. Nat Med. 2021;27:2136–2143.
10. Chaudhary N, Weissman D, Whitehead KA. mRNA vaccines for infectious diseases: principles, delivery, and clinical translation. Nat Rev Drug Discov. 2021;20(11):817–838.
11. Krieg PA, Melton DA. in vitro transcription of cloned cDNAs. Nucleic Acids Res. 1984;12:7057–7070.
12. Velikova T, Georgiev T. SARS-CoV-2 vaccines and autoimmune diseases amidst the COVID-19 crisis. Rheumatol Int. 2021;41(3):509–518.
13. Damiati LA, El-Messeiry S. An Overview of RNA-Based Scaffolds for Osteogenesis. Front Mol Biosci. 2021;8:682581. doi:10.3389/fmolb.2021.682581
14. Kowalski PS, Rudra A, Miao L, Anderson DG. Delivering the Messenger: Advances in Technologies for Therapeutic mRNA Delivery. Mol Ther. 2019;27(4):710–728. doi:10.1016/j.ymthe.2019.02.012
15. Zhang C, Maruggi G, Shan H, Li J. Advances in mRNA vaccines for infectious diseases. Front Immunol. 2019;10:594.
16. Ho W, Gao M, Li F, Li Z, Zhang XQ, Xu X. Next‐generation vaccines: nanoparticle‐mediated DNA and mRNA delivery. Adv Healthcare Mater. 2021;10(8):2001812.
17. Granot Y, Peer D. Delivering the right message: challenges and opportunities in lipid nanoparticles-mediated modified mRNA therapeutics—An innate immune system standpoint. Seminars Immunol. 2017;34:68–77.
18. Guevara ML, Persano S, Persano F. Lipid-based vectors for therapeutic mRNA-based anti-cancer vaccines. Curr Pharm Des. 2019;25(13):1443–1454.
19. Fang E, Liu X, Li M, et al. Advances in COVID-19 mRNA vaccine development. Signal Transduction Targeted Therapy. 2022;7(1):94.
20. Iavarone C, O’hagan DT, Yu D, Delahaye NF, Ulmer JB. Mechanism of action of mRNA-based vaccines. Expert Rev Vaccines. 2017;16(9):871–881.
21. Sahin U, Karikó K, Türeci Ö. mRNA-based therapeutics—developing a new class of drugs. Nat Rev Drug Discov. 2014;13(10):759–780.
22. Tourriere H, Chebli K, Tazi J. mRNA degradation machines in eukaryotic cells. Biochimie. 2002;84(8):821–837.
23. Zhong Z, Mc Cafferty S, Combes F, et al. mRNA therapeutics deliver a hopeful message. Nano Today. 2018;23:16–39.
24. Weissman D. mRNA transcript therapy. Expert Rev Vaccines. 2015;14(2):265–281.
25. Nance KD, Meier JL. Modifications in an emergency: the role of N1-methyl pseudouridine in COVID-19 vaccines. ACS Central Sci. 2021;7(5):748–756.
26. Chamberlin M, Mcgrath J, Waskell L. New RNA polymerase from Escherichia coli infected with bacteriophage T7. Nature. 1970;228:227–231.
27. Chamberlin M, Kingston R, Gilman M, Wiggs J, De Vera A. Isolation of bacterial and bacteriophage RNA polymerases and their use in the synthesis of RNA in Vitro. Methods Enzymol. 1983;101:540–568.
28. Krieg PA, Melton DA. Functional messenger RNAs are produced by SP6 in vitro transcription of cloned cDNAs. Nucleic Acids Res. 1984;12(18):7057–7070. doi:10.1093/nar/12.18.7057
29. Li B, Luo X, Dong Y. Effects of chemically modified messenger RNA on protein expression. Bioconjug Chem. 2016;27(3):849–853.
30. Kimoto M, Meyer AJ, Hirao I, Ellington AD. Genetic alphabet expansion transcription generating functional RNA molecules containing a five-letter alphabet including modified unnatural and natural base nucleotides by thermostable T7 RNA polymerase variants. Chem Commun. 2017;53(91):12309–12312.
31. Milisavljevič N, Perlíková P, Pohl R, Hocek M. Enzymatic synthesis of base-modified RNA by T7 RNA polymerase. A systematic study and comparison of 5-substituted pyrimidine and 7-substituted 7-deazapurine nucleoside triphosphates as substrates. Org Biomol Chem. 2018;16(32):5800–5807.
32. Stepinski J, Waddell C, Stolarski R, Darzynkiewicz E, Rhoads RE. Synthesis and properties of mRNAs containing the novel “anti-reverse” cap analogs 7-methyl (3′-O-methyl) GpppG and 7-methyl (3′-deoxy) GpppG. RNA. 2001;7(10):1486–1495.
33. Fath S, Bauer AP, Liss M, et al. Multiparameter RNA, and codon optimization: a standardized tool to assess and enhance autologous mammalian gene expression. PLoS One. 2011;6(3):e17596.
34. Holtkamp S, Kreiter S, Selmi A, et al. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood. 2006;108(13):4009–4017.
35. Trepotec Z, Geiger J, Plank C, Aneja MK, Rudolph C. Segmented poly (A) tails significantly reduce recombination of plasmid DNA without affecting mRNA translation efficiency or half-life. RNA. 2019;25(4):507–518.
36. To KK, Cho WC. An overview of rational design of mRNA-based therapeutics and vaccines. Expert Opin Drug Discov. 2021;16(11):1307–1317.
37. Karikó K, Muramatsu H, Welsh FA, et al. Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol Therapy. 2008;16(11):1833–1840.
38. Mauger DM, Cabral BJ, Presnyak V, et al. mRNA structure regulates protein expression through changes in functional half-life. Proce National Acad Sci. 2019;116(48):24075–24083.
39. Hinnebusch AG, Ivanov IP, Sonenberg N. Translational control by 5′-untranslated regions of eukaryotic mRNAs. Science. 2016;352(6292):1413–1416.
40. Babendure JR, Babendure JL, Ding JH, Tsien RY. Control of mammalian translation by mRNA structure near caps. RNA. 2006;12(5):851–861.
41. Kozak M. Regulation of translation via mRNA structure in prokaryotes and eukaryotes. Gene. 2005;361:13–37.
42. Sonenberg N, Hinnebusch AG. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 2009;136(4):731–745.
43. Haizel SA, Bhardwaj U, Gonzalez RL, Mitra S, Goss DJ. 5′-UTR recruitment of the translation initiation factor eIF4GI or DAP5 drives cap-independent translation of a subset of human mRNAs. J Biol Chem. 2020;295(33):11693–11706.
44. Elroy-Stein O, Fuerst TR, Moss B. Cap-independent translation of mRNA conferred by encephalomyocarditis virus 5’sequence improves the performance of the vaccinia virus/bacteriophage T7 hybrid expression system. Proce National Acad Sci. 1989;86(16):6126–6130.
45. Tan X, Wan Y. Enhanced protein expression by IRES-driven mRNA translation as a novel approach for in vitro loading dendritic cells with antigens. Human Immunol. 2008;69(1):32–40.
46. Balzer Le S, Onsager I, Lorentzen JA, Lale R. Dual UTR-A novel 5′ untranslated region design for synthetic biology applications. Synthetic Biol. 2020;5(1):ysaa006.
47. Leppek K, Das R, Barna M. Functional 5′ UTR mRNA structures in eukaryotic translation regulation and how to find them. Nat Rev Mol Cell Biol. 2018;19(3):158–174.
48. Asrani KH, Farelli JD, Stahley MR, et al. Optimization of mRNA untranslated regions for improved expression of therapeutic mRNA. RNA Biol. 2018;15(6):756–762.
49. Mayr C. Regulation by 3′-untranslated regions. Annu Rev Genet. 2017;51:171–194.
50. Matoulkova E, Michalova E, Vojtesek B, Hrstka R. The role of the 3’untranslated region in post-transcriptional regulation of protein expression in mammalian cells. RNA Biol. 2012;9(5):563–576.
51. Barreau C, Paillard L, Osborne HB. AU-rich elements and associated factors: are there unifying principles?. Nucleic Acids Res. 2005;33(22):7138–7150.
52. Jiang Y, Xu XS, Russell JE. A nucleolin-binding 3′ untranslated region element stabilizes β-globin mRNA in vivo. Mol Cell Biol. 2006;26(6):2419–2429.
53. Eberhardt W, Doller A, Akool ES, Pfeilschifter J. Modulation of mRNA stability as a novel therapeutic approach. Pharmacol Ther. 2007;114(1):56–73.
54. Wilkinson N, Pantopoulos K. The IRP/IRE system in vivo: insights from mouse models. Front Pharmacol. 2014;5:176.
55. Yu S, Kim VN. A tale of non-canonical tails: gene regulation by post-transcriptional RNA tailing. Nat Rev Mol Cell Biol. 2020;21(9):542–556.
56. Tang TT, Passmore LA. Recognition of poly (A) RNA through its intrinsic helical structure. Cold Spring Harbor Symposia Quantitative Biol. 2019;84:21–30.
57. Goldstrohm AC, Wickens M. Multifunctional deadenylase complexes diversify mRNA control. Nat Rev Mol Cell Biol. 2008;9(4):337–344.
58. Natalizio BJ, Wente SR. Postage for the messenger: designating routes for nuclear mRNA export. Trends Cell Biol. 2013;23(8):365–373.
59. Bresson SM, Conrad NK. The human nuclear poly (a)-binding protein promotes RNA hyper adenylation and decay. PLoS Genet. 2013;9(10):e1003893.
60. Wu HY, Ke TY, Liao WY, Chang NY. Regulation of coronaviral poly (A) tail length during infection. PLoS One. 2013;8(7):e70548.
61. Gallie D. The cap and poly (A) tail function synergistically to regulate mRNA translational efficiency. Genes Dev. 1991;5(11):2108–2116.
62. Bloom K, van den Berg F, Arbuthnot P. Self-amplifying RNA vaccines for infectious diseases. Gene Ther. 2021;28(3–4):117–129.
63. McCullough KC, Milona P, Thomann-Harwood L, et al. Self-amplifying replicon RNA vaccine delivery to dendritic cells by synthetic nanoparticles. Vaccines. 2014;2(4):735–754.
64. Sandbrink JB, Shattock RJ. RNA vaccines: a suitable platform for tackling emerging pandemics?. Front Immunol. 2020;11:608460.
65. Beissert T, Perkovic M, Vogel A, et al. A trans-amplifying RNA vaccine strategy for induction of potent protective immunity. Mol Therapy. 2020;28(1):119–128.
66. Kairuz D, Samudh N, Ely A, Arbuthnot P, Bloom K. Advancing mRNA technologies for therapies and vaccines: An African context. Front Immunol. 2022;1:13.
67. Blakney AK, Ip S, Geall AJ. An update on self-amplifying mRNA vaccine development. Vaccines. 2021;9(2):97.
68. Kim DY, Atasheva S, McAuley AJ, et al. Enhancement of protein expression by alphavirus replicons by designing self-replicating subgenomic RNAs. Proc Natl Acad Sci U S A. 2014;111(29):10708–10713. doi:10.1073/pnas.1408677111
69. Lundstrom K. Self-replicating RNA viruses for RNA therapeutics. Molecules. 2018;23(12):3310.
70. Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines—a new era in vaccinology. Nat Rev Drug Discov. 2018;17(4):261–279.
71. Kwon H, Kim M, Seo Y, et al. The emergence of synthetic mRNA: In vitro synthesis of mRNA and its applications in regenerative medicine. Biomaterials. 2018;156:172–193.
72. Dowdy SF. Overcoming cellular barriers for RNA therapeutics. Nat Biotechnol. 2017;35(3):222–229.
73. Lorenz C, Fotin-Mleczek M, Roth G, et al. Protein expression from exogenous mRNA: uptake by receptor-mediated endocytosis and trafficking via the lysosomal pathway. RNA Biol. 2011;8(4):627–636.
74. Diken M, Kreiter S, Selmi A, et al. Selective uptake of naked vaccine RNA by dendritic cells is driven by macropinocytosis and abrogated upon DC maturation. Gene Ther. 2011;18(7):702–708.
75. Reichmuth AM, Oberli MA, Jaklenec A, Langer R, Blankschtein D. mRNA vaccine delivery using lipid nanoparticles. Ther Deliv. 2016;7(5):319–334.
76. Hajj KA, Whitehead KA. Tools for translation: non-viral materials for therapeutic mRNA delivery. Nat Rev Materials. 2017;2(10):1–7.
77. Wadhwa A, Aljabbari A, Lokras A, Foged C, Thakur A. Opportunities and challenges in the delivery of mRNA-based vaccines. Pharmaceutics. 2020;12(2):102.
78. Schlich M, Palomba R, Costabile G, et al. Cytosolic delivery of nucleic acids: The case of ionizable lipid nanoparticles. Bioeng Translational Med. 2021;6(2):e10213.
79. Delehedde C, Even L, Midoux P, Pichon C, Perche F. Intracellular routing and recognition of lipid-based mRNA nanoparticles. Pharmaceutics. 2021;13(7):945.
80. Islam MA, Xu Y, Tao W, et al. Restoration of tumor-growth suppression in vivo via systemic nanoparticle-mediated delivery of PTEN mRNA. Nature Biomed Eng. 2018;2(11):850–864.
81. Iavarone C, Ramsauer K, Kubarenko AV, et al. A point mutation in the amino terminus of TLR7 abolishes signaling without affecting ligand binding. J Immunol. 2011;186(7):4213–4222.
82. Karikó K, Muramatsu H, Ludwig J, Weissman D. Generating the optimal mRNA for therapy: HPLC purification eliminates immune activation and improves translation of nucleoside-modified, protein-encoding mRNA. Nucleic Acids Res. 2011;39(21):e142.
83. Lokugamage MP, Gan Z, Zurla C, et al. Mild innate immune activation overrides efficient nanoparticle‐mediated RNA delivery. Adv Mater. 2020;32(1):1904905.
84. Pollard C, Rejman J, De Haes W, et al. Type I IFN counteracts the induction of antigen-specific immune responses by lipid-based delivery of mRNA vaccines. Mol Therapy. 2013;21(1):251–259.
85. Zaki AM, Van Boheemen S, Bestebroer TM, Osterhaus AD, Fouchier RA. Isolation of a novel coronavirus from a man with pneumonia in Saudi Arabia. N Eng J Med. 2012;367(19):1814–1820.
86. world health organization (WHO). Middle East respiratory syndrome. MERS situation; 2021.
87. Mu X, Hur S. Immunogenicity of in vitro-transcribed RNA. Acc Chem Res. 2021;54(21):4012–4023.
88. Hadas Y, Katz MG, Bridges CR, Zangi L. Modified mRNA as a therapeutic tool to induce cardiac regeneration in ischemic heart disease. Wiley Interdiscip Rev Syst Biol Med. 2017;9(1):e1367.
89. Sartorius R, Trovato M, Manco R, D’Apice L, De Berardinis P. Exploiting viral sensing mediated by Toll-like receptors to design innovative vaccines. NPJ Vaccines. 2021;6(1):127.
90. Ganesan P, Narayanasamy D. Lipid nanoparticles: different preparation techniques, characterization, hurdles, and strategies for the production of solid lipid nanoparticles and nanostructured lipid carriers for oral drug delivery. Sustainable Chem Pharmacy. 2017;6:37–56.
91. Richner JM, Himansu S, Dowd KA, et al. Modified mRNA vaccines protect against Zika virus infection. Cell. 2017;168(6):1114–1125.
92. Freyn AW, da Silva JR, Rosado VC, et al. A multi-targeting, nucleoside-modified mRNA influenza virus vaccine provides broad protection in mice. Mol Therapy. 2020;28(7):1569–1584.
93. Zhuang X, Qi Y, Wang M, et al. mRNA vaccines encoding the HA protein of influenza A H1N1 virus delivered by cationic lipid nanoparticles induce protective immune responses in mice. Vaccines. 2020;8(1):123.
94. Pardi N, LaBranche CC, Ferrari G, et al. Characterization of HIV-1 nucleoside-modified mRNA vaccines in rabbits and rhesus macaques. Mol Therapy Nucleic Acids. 2019;15:36–47.
95. Moyo N, Vogel AB, Buus S, et al. Efficient induction of T cells against conserved HIV-1 regions by mosaic vaccines delivered as self-amplifying mRNA. Mol Therapy Methods Clin Dev. 2019;12:32–46.
96. Mansanguan S, Charunwatthana P, Piyaphanee W, Dechkhajorn W, Poolcharoen A, Mansanguan C. Cardiovascular manifestation of the BNT162b2 mRNA COVID-19 vaccine in adolescents. Trop Med Infect Dis. 2022;7(8):196.
97. Sinagra G, Merlo M, Porcari A. Exploring the possible link between myocarditis and mRNA COVID-19 vaccines. Eur J Intern Med. 2021;92:28–30.
98. Liang Y, Huang L, Liu T. Development and delivery systems of mRNA vaccines. Front Bioeng Biotechnol. 2021;9:718753.
99. Kuntz J, Crane B, Weinmann S, Naleway AL; Vaccine Safety Datalink Investigator Team. Myocarditis and pericarditis are rare following live viral vaccinations in adults. Vaccine. 2018;36(12):1524–1527.
100. Tarab-Ravski D, Stotsky-Oterin L, Peer D. Delivery strategies of RNA therapeutics to leukocytes. J Controlled Release. 2022;342:362–371.
101. Gutkin A, Rosenblum D, Peer D. RNA delivery with a human virus-like particle. Nat Biotechnol. 2021;39(12):1514–1515.
102. Igyártó BZ, Jacobsen S, Ndeupen S. Future considerations for the mRNA-lipid nanoparticle vaccine platform. Curr Opin Virol. 2021;48:65–72.
103. Ndeupen S, Qin Z, Jacobsen S, Bouteau A, Estanbouli H, Igyártó BZ. The mRNA-LNP platform’s lipid nanoparticle component used in preclinical vaccine studies is highly inflammatory. Science. 2021;24(12):103479.
104. Knop K, Hoogenboom R, Fischer D, Schubert US. Poly (ethylene glycol) in drug delivery: pros and cons as well as potential alternatives. Angewandte chemie int edition. 2010;49(36):6288–6308.
105. Lila AS, Kiwada H, Ishida T. The accelerated blood clearance (ABC) phenomenon: clinical challenge and approaches to manage. J Controlled Release. 2013;172(1):38–47.
106. Kedmi R, Ben-Arie N, Peer D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials. 2010;31(26):6867–6875.
107. Sedic M, Senn JJ, Lynn A, et al. Safety evaluation of lipid nanoparticle–formulated modified mRNA in the Sprague-Dawley rat and cynomolgus monkey. Vet Pathol. 2018;55(2):341–354.
108. Dokka S, Toledo D, Shi X, Castranova V, Rojanasakul Y. Oxygen radical-mediated pulmonary toxicity induced by some cationic liposomes. Pharm Res. 2000;17:521–525.
109. Li S, Wu SP, Whitmore M, et al. Effect of immune response on gene transfer to the lung via systemic administration of cationic lipidic vectors. Am J Physiol Lung Cell Mol Physiol. 1999;276(5):L796–804.
110. Kowalzik F, Schreiner D, Jensen C, Teschner D, Gehring S, Zepp F. mRNA-Based Vaccines. Vaccines. 2021;9(4):390. doi:10.3390/vaccines9040390.
111. Wollner CJ, Richner JM. mRNA vaccines against flaviviruses. Vaccines. 2021;9(2):148.
112. Wang Y, Zhang Z, Luo J, Han X, Wei Y, Wei X. mRNA vaccine: a potential therapeutic strategy. Mol Cancer. 2021;20(1):33.
113. Petsch B, Schnee M, Vogel AB, et al. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat Biotechnol. 2012;30(12):1210–1216.
114. Anderson BR, Muramatsu H, Nallagatla SR, et al. Incorporation of pseudouridine into mRNA enhances translation by diminishing PKR activation. Nucleic Acids Res. 2010;38(17):5884–5892.
115. Karikó K, Buckstein M, Ni H, Weissman D. Suppression of RNA recognition by Toll-like receptors: the impact of nucleoside modification and the evolutionary origin of RNA. Immunity. 2005;23(2):165–175.
116. Warren L, Manos PD, Ahfeldt T, et al. Highly efficient reprogramming to pluripotency and directed differentiation of human cells with synthetic modified mRNA. Cell stem cell. 2010;7(5):618–630.
117. Nelson J, Sorensen EW, Mintri S, et al. Impact of mRNA chemistry and manufacturing process on innate immune activation. Sci Adv. 2020;6(26):eaaz6893.
118. Thess A, Grund S, Mui BL, et al. Sequence-engineered mRNA without chemical nucleoside modifications enables an effective protein therapy in large animals. Mol Therapy. 2015;23(9):1456–1464.
119. Al-Saif M, Khabar KS. UU/UA dinucleotide frequency reduction in coding regions results in increased mRNA stability and protein expression. Mol Therapy. 2012;20(5):954–959.
120. Vaidyanathan S, Azizian KT, Haque AA, et al. Uridine depletion and chemical modification increase Cas9 mRNA activity and reduce immunogenicity without HPLC purification. Mol Therapy Nucleic Acids. 2018;12:530–542.
121. Krienke C, Kolb L, Diken E, et al. A noninflammatory mRNA vaccine for the treatment of experimental autoimmune encephalomyelitis. Science. 2021;371(6525):145–153.
122. Starostina EV, Sharabrin SV, Antropov DN, et al. Construction and immunogenicity of modified mRNA-vaccine variants encoding influenza virus antigens. Vaccines. 2021;9(5):452.
123. Schoenmaker L, Witzigmann D, Kulkarni JA, et al. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int J Pharm. 2021;601:120586.
124. Yang Q, Yu CH, Zhao F, et al. eRF1 mediates codon usage effects on mRNA translation efficiency through premature termination at rare codons. Nucleic Acids Res. 2019;47(17):9243–9258.
125. Triana-Alonso FJ, Dabrowski M, Wadzack J, Nierhaus KH. Self-coded 3′-extension of run-off transcripts produces aberrant products during in vitro transcription with T7 RNA polymerase. J Biol Chem. 1995;270(11):6298–6307.
126. Gholamalipour Y, Karunanayake Mudiyanselage A, Martin CT. 3′ end additions by T7 RNA polymerase are RNA self-templated, distributive, and diverse—RNA-Seq analyses. Nucleic Acids Res. 2018;46(18):9253–9263.
127. Wadman M. Public needs to prep for vaccine side effects. Science. 2020;370(6520):1022. doi:10.1126/science.370.6520.1022
128. Maier MA, Jayaraman M, Matsuda S, et al. Biodegradable lipids enabling rapidly eliminated lipid nanoparticles for systemic delivery of RNAi therapeutics. Mol Therapy. 2013;21(8):1570–1578.
129. Sabnis S, Kumarasinghe ES, Salerno T, et al. A novel amino lipid series for mRNA delivery: improved endosomal escape and sustained pharmacology and safety in non-human primates. Mol Therapy. 2018;26(6):1509–1519.
130. Hassett KJ, Benenato KE, Jacquinet E, et al. Optimization of lipid nanoparticles for intramuscular administration of mRNA vaccines. Mol Therapy Nucleic Acids. 2019;15:1.
131. Zhang X, Zhao W, Nguyen GN, et al. Functionalized lipid-like nanoparticles for in vivo mRNA delivery and base editing. Sci Adv. 2020;6(34):eabc2315.
132. Zhang X, Li B, Luo X, et al. Biodegradable amino-ester nanomaterials for Cas9 mRNA delivery in vitro and in vivo. ACS Appl Mater Interfaces. 2017;9(30):25481–25487.
133. Jackson NA, Kester KE, Casimiro D, Gurunathan S, DeRosa F. The promise of mRNA vaccines: a biotech and industrial perspective. NPJ Vaccines. 2020;5:11.
134. Slater RJ. The purification of poly (A)-containing RNA by affinity chromatography. Nucleic Acids. 1984;117–120.
135. Summer H, Grämer R, Dröge P. Denaturing urea polyacrylamide gel electrophoresis (Urea PAGE). J Visualized Exp. 2009;29(32):e1485.
136. Aldosari BN, Alfagih IM, Almurshedi AS. Lipid nanoparticles as delivery systems for RNA-based vaccines. Pharmaceutics. 2021;13(2):206.
137. Pascolo S. Vaccination with messenger RNA. DNA Vaccines. 2006;10:23–40.
138. Weissman D, Pardi N, Muramatsu H, Karikó K. Synthetic messenger RNA and cell metabolism modulation, methods, and protocols. Methods Mol Biol. 2012;969:43–54.
139. Baiersdörfer M, Boros G, Muramatsu H, et al. A facile method for the removal of dsRNA contaminant from in vitro-transcribed mRNA. Mol Therapy Nucleic Acids. 2019;15:26–35.
140. Weissman D, Pardi N, Muramatsu H, Karikó K. HPLC purification of in vitro transcribed long RNA. Synthetic Messenger RNA Cell Metab Modulation. 2013;43–54.
141. Zhang RX, Ahmed T, Li LY, Li J, Abbasi AZ, Wu XY. Design of nanocarriers for nanoscale drug delivery to enhance cancer treatment using hybrid polymer and lipid building blocks. Nanoscale. 2017;9(4):1334–1355.
142. Deering RP, Kommareddy S, Ulmer JB, Brito LA, Geall AJ. Nucleic acid vaccines: prospects for non-viral delivery of mRNA vaccines. Expert Opin Drug Deliv. 2014;11(6):885–899.
143. Stump WT, Hall KB. SP6 RNA polymerase efficiently synthesizes RNA from short double-stranded DNA templates. Nucleic Acids Res. 1993;21(23):5480–5484.
144. Edelmann A, Kirchberger J, Naumann M, Kopperschläger G. Generation of catalytically active 6‐phosphofructokinase from Saccharomyces cerevisiae in a cell‐free system. Eur J Biochem. 2000;267(15):4825–4830.
145. Working PK. Pharmacological-toxicological expert report. CAELYX^. (Stealth^(! R) liposomal doxorubicin HCl). Hum Exp Toxicol. 1996;15:751–785.
146. Bulbake U, Doppalapudi S, Kommineni N, Khan W. Liposomal formulations in clinical use: an updated review. Pharmaceutics. 2017;9(2):12.
147. FDA Approves first-of-its-kind targeted RNA-based therapy to treat a rare disease. FDA. Available from: https://www.fda.gov/news-events/press-announcements/fda-approves-first-its-kind-targeted-rna-based-therapy-treat-rare-disease.
148. FDA Approves first COVID-19 vaccine. FDA. Available from:https://www.fda.gov/news-events/press-announcements/fda-approves-first-covid-19-vaccine.
149. Working PK. Pharmacological-toxicological expert report. Hum Exp Toxicol. 1996;15:751–785.
150. Deal CE, Carfi A, Plante OJ. Advancements in mRNA encoded antibodies for passive immunotherapy. Vaccines. 2021;9(2):108.
151. Hoerr I, Obst R, Rammensee HG, Jung G. In vivo, the application of RNA leads to the induction of specific cytotoxic T lymphocytes and antibodies. Eur J Immunol. 2000;30(1):1–7.
152. Hoyer JA, Neundorf I. Peptide vectors for the nonviral delivery of nucleic acids. Acc Chem Res. 2012;45(7):1048–1056.
153. Nakase I, Akita H, Kogure K, et al. Efficient intracellular delivery of nucleic acid pharmaceuticals using cell-penetrating peptides. Acc Chem Res. 2012;45(7):1132–1139.
154. Islam MA, Reesor EK, Xu Y, Zope HR, Zetter BR, Shi J. Biomaterials for mRNA delivery. Biomater sci. 2015;3(12):1519–1533.
155. Li H, Tsui TY, Ma W. Intracellular delivery of molecular cargo using cell-penetrating peptides and the combination strategies. Int J Mol Sci. 2015;16(8):19518–19536.
156. Kallen KJ, Heidenreich R, Schnee M, et al. A novel, disruptive vaccination technology: self-adjuvanted RNActive® vaccines. Hum Vaccin Immunother. 2013;9(10):2263–2276.
157. Scheel B, Teufel R, Probst J, et al. Toll‐like receptor‐dependent activation of several human blood cell types by protamine‐condensed mRNA. Eur J Immunol. 2005;35(5):1557–1566.
158. Schnee M, Vogel AB, Voss D, et al. An mRNA vaccine encoding rabies virus glycoprotein induces protection against lethal infection in mice and correlates of protection in adult and newborn pigs. PLoS Negl Trop Dis. 2016;10(6):e0004746.
159. Weide B, Pascolo S, Scheel B, et al. Direct injection of protamine-protected mRNA: results of a phase 1/2 vaccination trial in metastatic melanoma patients. J Immunother. 2009;32(5):498–507.
160. Stitz L, Vogel A, Schnee M, et al. A thermostable messenger RNA-based vaccine against rabies. PLoS Negl Trop Dis. 2017;11(12):e0006108.
161. Wang Y, Su HH, Yang Y, et al. Systemic delivery of modified mRNA encoding herpes simplex virus 1 thymidine kinase for targeted cancer gene therapy. Mol Therapy. 2013;21(2):358–367.
162. Midoux P, Pichon C. Lipid-based mRNA vaccine delivery systems. Expert Rev Vaccines. 2015;14(2):221–234.
163. Pardi N, Hogan MJ, Naradikian MS, et al. Nucleoside-modified mRNA vaccines induce potent T follicular helper and germinal center B cell responses. J Exp Med. 2018;215(6):1571–1588.
164. Van Meer G, Voelker DR, Feigenson GW. Membrane lipids: where they are and how they behave. Nat Rev Mol Cell Biol. 2008;9(2):112–124.
165. Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Materials. 2021;6(12):1078–1094.
166. Kranz LM, Diken M, Haas H, et al. Systemic RNA delivery to dendritic cells exploits antiviral defense for cancer immunotherapy. Nature. 2016;534(7607):396–401.
167. Oberli MA, Reichmuth AM, Dorkin JR, et al. Lipid nanoparticle assisted mRNA delivery for potent cancer immunotherapy. Nano Lett. 2017;17(3):1326–1335.
168. Persano S, Guevara ML, Li Z, et al. Lipopolyplex potentiates anti-tumor immunity of mRNA-based vaccination. Biomaterials. 2017;125:81–89.
169. Miao L, Lin J, Huang Y, et al. Synergistic lipid compositions for albumin receptor-mediated delivery of mRNA to the liver. Nat Commun. 2020;11(1):2424.
170. Patel S, Ashwanikumar N, Robinson E, et al. Naturally occurring cholesterol analogs in lipid nanoparticles induce polymorphic shape and enhance intracellular delivery of mRNA. Nat Commun. 2020;11(1):983.
171. Eygeris Y, Patel S, Jozic A, Sahay G. Deconvoluting lipid nanoparticle structure for messenger RNA delivery. Nano Lett. 2020;20(6):4543–4549.
172. Guevara ML, Persano F, Persano S. Advances in lipid nanoparticles for mRNA-based cancer immunotherapy. Front chem. 2020;8:589959.
173. Felgner PL, Gadek TR, Holm M, et al. Lipofection: a highly efficient, lipid-mediated DNA-transfection procedure. Proce National Acad Sci. 1987;84(21):7413–7417.
174. Semple SC, Klimuk SK, Harasym TO, et al. Efficient encapsulation of antisense oligonucleotides in lipid vesicles using ionizable amino lipids: formation of novel small multilamellar vesicle structures. Biochimica et Biophysica Acta. 2001;1510(1–2):152–166.
175. Semple SC, Klimuk SK, Harasym TO, Hope MJ. Lipid-based formulations of antisense oligonucleotides for systemic delivery applications. Methods Enzymol. 2000;313:322–341.
176. Kahvejian A, Svitkin YV, Sukarieh R, M’Boutchou MN, Sonenberg N. Mammalian poly (A)-binding protein is a eukaryotic translation initiation factor, which acts via multiple mechanisms. Genes Dev. 2005;19(1):104–113.
177. Tang Z, Zhang X, Shu Y, Guo M, Zhang H, Tao W. Insights from nanotechnology in COVID-19 treatment. Nano Today. 2021;36:101019.
178. Fobian SF, Cheng Z, Ten Hagen TL. Smart lipid-based nanosystems for therapeutic immune induction against cancers: perspectives and outlooks. Pharmaceutics. 2022;14(1):26.
179. Linares-Fernández S, Lacroix C, Exposito JY, Verrier B. Tailoring mRNA vaccine to balance innate/adaptive immune response. Trends Mol Med. 2020;26(3):311–323.
180. Guan S, Rosenecker J. Nanotechnologies in delivery of mRNA therapeutics using nonviral vector-based delivery systems. Gene Ther. 2017;24(3):133–143.
181. Eygeris Y, Gupta M, Kim J, Sahay G. Chemistry of lipid nanoparticles for RNA delivery. Acc Chem Res. 2021;55(1):2–12.
182. Kauffman KJ, Dorkin JR, Yang JH, et al. Optimization of lipid nanoparticle formulations for mRNA delivery in vivo with fractional factorial and definitive screening designs. Nano Lett. 2015;15(11):7300–7306.
183. Kuntsche E, Kuntsche S, Knibbe R, et al. Cultural and gender convergence in adolescent drunkenness: evidence from 23 European and North American countries. Arch Pediatr Adolesc Med. 2011;165(2):152–158.
184. Gilleron J, Querbes W, Zeigerer A, et al. Image-based analysis of lipid nanoparticle-mediated siRNA delivery, intracellular trafficking, and endosomal escape. Nat Biotechnol. 2013;31(7):638–646.
185. Li B, Luo X, Deng B, et al. An orthogonal array optimization of lipid-like nanoparticles for mRNA delivery in vivo. Nano Lett. 2015;15(12):8099–8107.
186. Vu MN, Kelly HG, Wheatley AK, et al. Cellular interactions of liposomes and PISA nanoparticles during human blood flow in a microvascular network. Small. 2020;16(33):2002861.
187. Magar KT, Boafo GF, Li X, Chen Z, He W. Liposome-based delivery of biological drugs. Chine Chem Lett. 2022;33(2):587–596.
188. Guimarães D, Cavaco-Paulo A, Nogueira E. Design of liposomes as drug delivery system for therapeutic applications. Int J Pharm. 2021;601:120571.
189. Wanigasekara J, de Carvalho AM, Cullen PJ, Tiwari B, Curtin JF. Converging technologies: targeting the hallmarks of cancer using ultrasound and microbubbles. Trends Cancer. 2021;7(10):886–890.
190. Tenchov R, Bird R, Curtze AE, Zhou Q. Lipid nanoparticles─ from liposomes to mRNA vaccine delivery, a landscape of research diversity and advancement. ACS Nano. 2021;15(11):16982–17015.
191. Kallen KJ, Theß A. A development that may evolve into a revolution in medicine: mRNA as the basis for novel, nucleotide-based vaccines, and drugs. Advances Vaccines. 2014;2(1):10–31.
192. Read ML, Singh S, Ahmed Z, et al. A versatile reducible polycation-based system for efficient delivery of a broad range of nucleic acids. Nucleic Acids Res. 2005;33(9):e86.
193. Müller RH, Mäder K, Gohla S. Solid lipid nanoparticles (SLN) for controlled drug delivery - a review of the state of the art. Eur J Pharm Biopharm. 2000;50(1):161–177. doi:10.1016/s0939-6411(00)00087-4
194. Sato Y, Hatakeyama H, Sakurai Y, Hyodo M, Akita H, Harashima H. A pH-sensitive cationic lipid facilitates the delivery of liposomal siRNA and gene-silencing activity in vitro and in vivo. J Controlled Release. 2012;163(3):267–276.
195. Malone RW, Felgner PL, Verma IM. Cationic liposome-mediated RNA transfection. Proce National Acad Sci. 1989;86(16):6077–6081.
196. Dalby B, Cates S, Harris A, et al. Advanced transfection with Lipofectamine 2000 reagent: primary neurons, siRNA, and high-throughput applications. Methods. 2004;33(2):95–103.
197. Zhang Y, Sun C, Wang C, Jankovic KE, Dong Y. Lipids and lipid derivatives for RNA delivery. Chem Rev. 2021;121(20):12181–12277.
198. Wasungu L, Hoekstra D. Cationic lipids, lipoplexes and intracellular delivery of genes. J Controlled Release. 2006;116(2):255–264.
199. Hirsch-Lerner D, Zhang M, Eliyahu H, Ferrari ME, Wheeler CJ, Barenholz Y. Effect of “helper lipid” on lipoplex electrostatics. Biochimica et Biophysica Acta. 2005;1714(2):71–84.
200. Wasungu L, Stuart MC, Scarzello M, Engberts JB, Hoekstra D. Lipoplexes formed from sugar-based gemini surfactants undergo a lamellar-to-micellar phase transition at acidic pH. Evidence for a non-inverted membrane-destabilizing hexagonal phase of lipoplexes. Biochimica et Biophysica Acta. 2006;1758(10):1677–1684.
201. Ramachandran S, Satapathy SR, Dutta T. Delivery strategies for mRNA vaccines. Pharmaceut Med. 2022;36(1):11–20.
202. de Ilarduya CT, Arangoa MA, Düzgüneş N. Transferrin-lipoplexes with protamine-condensed DNA for serum-resistant gene delivery. Methods Enzymol. 2003;373:342–356.
203. Hoekstra D, Scherphof G. Effect of fetal calf serum and serum protein fractions on the uptake of liposomal phosphatidylcholine by rat hepatocytes in primary monolayer culture. Biochimica et Biophysica Acta. 1979;551(1):109–121.
204. Chonn A, Semple SC, Cullis PR. Association of blood proteins with large unilamellar liposomes in vivo. Relation to circulation lifetimes. J Biol Chem. 1992;267(26):18759–18765.
205. Whitehead KA, Langer R, Anderson DG. Knocking down barriers: advances in siRNA delivery. Nat Rev Drug Discov. 2009;8(2):129–138.
206. Bailey AL, Cullis PR. Modulation of membrane fusion by asymmetric trans bilayer distributions of amino lipids. Biochemistry. 1994;33(42):12573–12580.
207. Żak MM, Zangi L. Lipid nanoparticles for organ-specific mRNA therapeutic delivery. Pharmaceutics. 2021;13(10):1675.
208. Thevenot J, Troutier AL, David L, Delair T, Ladavière C. Steric stabilization of lipid/polymer particle assemblies by poly (ethylene glycol)-lipids. Biomacromolecules. 2007;8(11):3651–3660.
209. Li M, Wang Z, Xie C, Xia X. Advances in mRNA vaccines. Int Rev Cell Mol Biol. 2022:65.
210. Pardi N, Hogan MJ, Weissman D. Recent advances in mRNA vaccine technology. Curr Opin Immunol. 2020;65:14–20.
211. Leung AK, Hafez IM, Baoukina S, et al. Lipid Nanoparticles Containing siRNA Synthesized by Microfluidic Mixing Exhibit an Electron-Dense Nanostructured Core. J Phys Chem C Nanomater Interfaces. 2012;116(34):18440–18450. doi:10.1021/jp303267y
212. Ross J. mRNA stability in mammalian cells. Microbiol Rev. 1995;59(3):423–450.
213. Geall AJ, Verma A, Otten GR, et al. Nonviral delivery of self-amplifying RNA vaccines. Proc Natl Acad Sci U S A. 2012;109(36):14604–14609. doi:10.1073/pnas.1209367109
214. Tam YY, Chen S, Cullis PR. Advances in lipid nanoparticles for siRNA delivery. Pharmaceutics. 2013;5(3):498–507.
215. Brito LA, Chan M, Shaw CA, et al. A cationic nanoemulsion for the delivery of next-generation RNA vaccines. Mol Therapy. 2014;22(12):2118–2129.
216. Jeeva S, Kim KH, Shin CH, Wang BZ, Kang SM. An update on mRNA-based viral vaccines. Vaccines. 2021;9(9):965.
217. Gordon S, Plüddemann A, Mukhopadhyay S. Sinusoidal immunity: macrophages at the lymphohematopoietic interface. Cold Spring Harb Perspect Biol. 2015;7(4):a016378.
218. Heesters BA, van der Poel CE, Das A, Carroll MC. Antigen presentation to B cells. Trends Immunol. 2016;37(12):844–854.
219. Ionescu L, Urschel S. Memory B cells and long-lived plasma cells. Transplantation. 2019;103(5):890–898.
220. Tanaka H, Sakurai Y, Anindita J, Akita H. Development of lipid-like materials for RNA delivery based on intracellular environment-responsive membrane destabilization and spontaneous collapse. Adv Drug Deliv Rev. 2020;154:210–226.
221. Fenton OS, Kauffman KJ, Kaczmarek JC, et al. Synthesis and biological evaluation of ionizable lipid materials for the in vivo delivery of messenger RNA to B lymphocytes. Adv Mater. 2017;29(33):1606944.
222. Rodrigueza WV, Wheeler JJ, Klimuk SK, Kitson CN, Hope MJ. Transbilayer movement and net flux of cholesterol and cholesterol sulfate between liposomal membranes. Biochemistry. 1995;34(18):6208–6217.
223. Herrera M, Kim J, Eygeris Y, Jozic A, Sahay G. Illuminating endosomal escape of polymorphic lipid nanoparticles that boost mRNA delivery. Biomater Sci. 2021;9(12):4289–4300.
224. Zadory M, Lopez E, Babity S, Gravel SP, Brambilla D. Current knowledge on the tissue distribution of mRNA nanocarriers for therapeutic protein expression. Biomater Sci. 2022.
225. Zuhorn IS, Bakowsky U, Polushkin E, et al. Nonbilayer phase of lipoplex–membrane mixture determines endosomal escape of genetic cargo and transfection efficiency. Mol Therapy. 2005;11(5):801–810.
226. Allen TM, Cullis PR. Liposomal drug delivery systems: from concept to clinical applications. Adv Drug Deliv Rev. 2013;65(1):36–48.
227. Buschmann MD, Carrasco MJ, Alishetty S, Paige M, Alameh MG, Weissman D. Nanomaterial delivery systems for mRNA vaccines. Vaccines. 2021;9(1):65.
228. Heyes J, Hall K, Tailor V, Lenz R, MacLachlan I. Synthesis and characterization of novel poly (ethylene glycol)-lipid conjugates suitable for use in drug delivery. J Controlled Release. 2006;112(2):280–290.
229. Torchilin VP. Recent advances with liposomes as pharmaceutical carriers. Nat Rev Drug Discov. 2005;4(2):145–160.
230. Woodle MC. Controlling liposome blood clearance by surface-grafted polymers. Adv Drug Deliv Rev. 1998;32(1–2):139–152.
231. Leung AK, Tam YY, Cullis PR. Lipid nanoparticles for short interfering RNA delivery. Adv Genet. 2014;88:71–110.
232. Li SD, Huang L. Nanoparticles evading the reticuloendothelial system: role of the supported bilayer. Biochimica et Biophysica Acta. 2009;1788(10):2259–2266.
233. Noble GT, Stefanick JF, Ashley JD, Kiziltepe T, Bilgicer B. Ligand-targeted liposome design: challenges and fundamental considerations. Trends Biotechnol. 2014;32(1):32–45.
234. Kumar V, Qin J, Jiang Y, et al. Shielding of lipid nanoparticles for siRNA delivery: impact on physicochemical properties, cytokine induction, and efficacy. Mol Therapy Nucleic Acids. 2014;3:e210.
235. Samaridou E, Heyes J, Lutwyche P. Lipid nanoparticles for nucleic acid delivery: Current perspectives. Adv Drug Deliv Rev. 2020;154:37–63.
236. Tabrez S, Jabir NR, Adhami VM, et al. Nanoencapsulated dietary polyphenols for cancer prevention and treatment: successes and challenges. Nanomedicine. 2020;15(11):1147–1162.
237. Zhu X, Tao W, Liu D, et al. Surface De-PEGylation controls nanoparticle-mediated siRNA delivery in vitro and in vivo. Theranostics. 2017;7(7):1990.
238. Semple SC, Akinc A, Chen J, et al. Rational design of cationic lipids for siRNA delivery. Nat Biotechnol. 2010;28(2):172–176.
239. Love KT, Mahon KP, Levins CG, et al. Lipid-like materials for low-dose, in vivo gene silencing. Proce National Acad Sci. 2010;107(5):1864–1869.
240. Whitehead KA, Sahay G, Li GZ, et al. Synergistic silencing: combinations of lipid-like materials for efficacious siRNA delivery. Mol Therapy. 2011;19(9):1688–1694.
241. Hatakeyama H, Akita H, Harashima H. The polyethyleneglycol dilemma: advantage and disadvantage of PEGylation of liposomes for systemic genes and nucleic acids delivery to tumors. Biol Pharm Bull. 2013;36(6):892–899.
242. Fang Y, Xue J, Gao S, et al. Cleavable PEGylation: a strategy for overcoming the “PEG dilemma” inefficient drug delivery. Drug Deliv. 2017;24(2):22–32.
243. Harvie P, Wong FM, Bally MB. Use of poly (ethylene glycol)–lipid conjugates to regulate the surface attributes and transfection activity of lipid–DNA particles. J Pharm Sci. 2000;89(5):652–663.
244. Hatakeyama H, Akita H, Kogure K, et al. Development of a novel systemic gene delivery system for cancer therapy with a tumor-specific cleavable PEG-lipid. Gene Ther. 2007;14(1):68–77.
245. Song LY, Ahkong QF, Rong Q, et al. Characterization of the inhibitory effect of PEG-lipid conjugates on the intracellular delivery of plasmid and antisense DNA mediated by cationic lipid liposomes. Biochimica et Biophysica Acta. 2002;1558(1):1–3.
246. Judge A, McClintock K, Phelps JR, MacLachlan I. Hypersensitivity and loss of disease site targeting caused by antibody responses to PEGylated liposomes. Mol Therapy. 2006;13(2):328–337.
247. Cheng X, Lee RJ. The role of helper lipids in lipid nanoparticles (LNPs) designed for oligonucleotide delivery. Adv Drug Deliv Rev. 2016;99:129–137.
248. Cheng Q, Wei T, Jia Y, et al. Dendrimer‐based lipid nanoparticles deliver therapeutic FAH mRNA to normalize liver function and extend survival in a mouse model of hepatorenal tyrosinemia type I. Adv Mater. 2018;30(52):1805308.
249. Koltover I, Salditt T, Rädler JO, Safinya CR. An inverted hexagonal phase of cationic liposome-DNA complexes related to DNA release and delivery. Science. 1998;281(5373):78–81.
250. Baden LR, El Sahly HM, Essink B, et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N Eng J Med. 2021;384(5):403–416.
251. Polack FP, Thomas SJ, Kitchin N, et al.; C4591001 Clinical Trial Group. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N Engl J Med. 2020;383(27):2603–2615. doi:10.1056/NEJMoa2034577
252. Xiao Y, Tang Z, Huang X, et al. Emerging mRNA technologies: delivery strategies and biomedical applications. Chem Soc Rev. 2022.
253. Zeng C, Zhang C, Walker PG, Dong Y. Formulation and delivery technologies for mRNA vaccines. Cham: Springer International Publishing; 2020.
254. Li M, Li Y, Peng K, et al. Engineering intranasal mRNA vaccines to enhance lymph node trafficking and immune responses. Acta biomaterials. 2017;64:237–248.
255. Wu GY, Wu CH. Receptor-mediated in vitro gene transformation by a soluble DNA carrier system. J Biol Chem. 1987;262(10):4429–4432.
256. Démoulins T, Milona P, Englezou PC, et al. Polyethylenimine-based polyplex delivery of self-replicating RNA vaccines. Nanomedicine. 2016;12(3):711–722.
257. Ghosh B, Biswas S. Polymeric micelles in cancer therapy: state of the art. J Controlled Release. 2021;332:127–147.
258. Ghezzi M, Pescina S, Padula C, et al. Polymeric micelles in drug delivery: An insight of the techniques for their characterization and assessment in biorelevant conditions. J Controlled Release. 2021;332:312–336.
259. McKinlay CJ, Vargas JR, Blake TR, et al. Charge-altering releasable transporters (CARTs) for the delivery and release of mRNA in living animals. Proce National Acad Sci. 2017;114(4):E448–56.
260. Jere D, Jiang HL, Arote R, et al. Degradable polyethyleneimine as DNA and small interfering RNA carriers. Expert Opin Drug Deliv. 2009;6(8):827–834.
261. Blakney AK, Zhu Y, McKay PF, et al. Big is beautiful: enhanced saRNA delivery and immunogenicity by a higher molecular weight, bioreducible, cationic polymer. ACS Nano. 2020;14(5):5711–5727.
262. Ansaldi F, Canepa P, Parodi V, et al. Adjuvanted seasonal influenza vaccines and perpetual viral metamorphosis: the importance of cross-protection. Vaccine. 2009;27(25–26):3345–3348.
263. O’Hagan DT, Ott GS, Nest GV, Rappuoli R, Giudice GD. The history of MF59® adjuvant: a phoenix that arose from the ashes. Expert Rev Vaccines. 2013;12(1):13–30.
264. Teixeira H, Dubernet C, Puisieux F, Benita S, Couvreur P. Submicron cationic emulsions as a new delivery system for oligonucleotides. Pharm Res. 1999;16:30–36.
265. Teixeira HF, Bruxel F, Fraga M, et al. Cationic nanoemulsions as nucleic acids delivery systems. Int J Pharm. 2017;534(1–2):356–367.
266. Anderluzzi G, Lou G, Gallorini S, et al. Investigating the impact of delivery system design on the efficacy of self-amplifying RNA vaccines. Vaccines. 2020;8(2):212.
267. Erasmus JH, Khandhar AP, O’Connor MA, et al. An Alphavirus-derived replicon RNA vaccine induces SARS-CoV-2 neutralizing antibody and T cell responses in mice and nonhuman primates. Sci Transl Med. 2020;12(555):eabc9396.
268. Erasmus JH, Khandhar AP, Walls AC, et al. Single-dose replicating RNA vaccine induces neutralizing antibodies against SARS-CoV-2 in nonhuman primates. bioRxiv. 2020;2020–2025.
269. Boczkowski D, Nair SK, Snyder D, Gilboa E. Dendritic cells pulsed with RNA are potent antigen-presenting cells in vitro and in vivo. J Exp Med. 1996;184(2):465–472.
270. Tateshita N, Miura N, Tanaka H, et al. Development of a lipoplex-type mRNA carrier composed of an ionizable lipid with a vitamin E scaffold and the KALA peptide for use as an ex vivo dendritic cell-based cancer vaccine. J Controlled Release. 2019;310:36–46.
271. Van Tendeloo VF, Ponsaerts P, Lardon F, et al. Highly efficient gene delivery by mRNA electroporation in human hematopoietic cells: superiority to lipofection and passive pulsing of mRNA and to electroporation of plasmid cDNA for tumor antigen loading of dendritic cells. Blood J Am Soc Hematol. 2001;98(1):49–56.
272. McNamara MA, Nair SK, Holl EK. RNA-Based Vaccines in Cancer Immunotherapy. J Immunol Res. 2015;2015:9.
273. Benteyn D, Heirman C, Bonehill A, Thielemans K. mRNA-based dendritic cell vaccines. Expert Rev Vaccines. 2015;14(2):161–176.
274. Perche F, Benvegnu T, Berchel M, et al. Enhancement of dendritic cells transfection in vivo and of vaccination against B16F10 melanoma with mannosylated histidylated lipo polyplexes loaded with tumor antigen messenger RNA. Nanomedicine. 2011;7(4):445–453.
275. Nencioni A, Grünebach F, Schmidt SM, et al. The use of dendritic cells in cancer immunotherapy. Crit Rev Oncol Hematol. 2008;65(3):191–199.
276. Hillaireau H, Couvreur P. Nanocarriers’ entry into the cell: relevance to drug delivery. Cell Mol Life Sci. 2009;66:2873–2896.
277. Doherty GJ, McMahon HT. Mechanisms of Endocytosis. Annu Rev Biochem. 2009;78:857–902.
278. Wang Y, Huang L. A window onto siRNA delivery. Nat Biotechnol. 2013;31(7):611–612.
279. Rzigalinski BA, Carfagna CS, Ehrich M. Cerium oxide nanoparticles in neuroprotection and considerations for efficacy and safety. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2017;9(4):e1444.
280. Kim J, Narayana A, Patel S, Sahay G. Advances in intracellular delivery through supramolecular self-assembly of oligonucleotides and peptides. Theranostics. 2019;9(11):3191.
281. Rejman J, Bragonzi A, Conese M. Role of clathrin-and caveolae-mediated endocytosis in gene transfer mediated by lipo-and polyplexes. Mol therapy. 2005;12(3):468–474.
282. Fujimoto T, Kogo H, Nomura R, Une T. Isoforms of caveolin-1 and caveolar structure. J Cell Sci. 2000;113(19):3509–3517.
283. Casey JR, Grinstein S, Orlowski J. Sensors and regulators of intracellular pH. Nat Rev Mol Cell Biol. 2010;11(1):50–61.
284. Maugeri M, Nawaz M, Papadimitriou A, et al. Linkage between the endosomal escape of LNP-mRNA and loading into EVs for transport to other cells. Nat Commun. 2019;10(1):4333.
285. Uchida S, Perche F, Pichon C, Cabral H. Nanomedicine-based approaches for mRNA delivery. Mol Pharm. 2020;17(10):3654–3684.
286. Varkouhi AK, Scholte M, Storm G, Haisma HJ. Endosomal escape pathways for delivery of biologicals. J Controlled Release. 2011;151(3):220–228.
287. Jiang Y, Lu Q, Wang Y, et al. Quantitating endosomal escape of a library of polymers for mRNA delivery. Nano Lett. 2020;20(2):1117–1123.
288. Wu Z, Li T. Nanoparticle-mediated cytoplasmic delivery of messenger RNA vaccines: challenges and future perspectives. Pharm Res. 2021;38:473–478.
289. Kim J, Eygeris Y, Gupta M, Sahay G. Self-assembled mRNA vaccines. Adv Drug Deliv Rev. 2021;170:83–112.
290. Kanamala M, Wilson WR, Yang M, Palmer BD, Wu Z. Mechanisms and biomaterials in pH-responsive tumor-targeted drug delivery: a review. Biomaterials. 2016;85:152–167.
291. Li B, Zhang X, Dong Y. Nanoscale platforms for messenger RNA delivery. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2019;11(2):e1530.
292. Midoux P, Roche AC, Monsigny M. Quantitation of the binding, uptake, and degradation of fluoresceinylated neoglycoproteins by flow cytometry. Cytometry. 1987;8(3):327–334.
293. Murphy RF, Powers S, Cantor CR. Endosome pH measured in single cells by dual fluorescence flow cytometry: rapid acidification of insulin to pH 6. J Cell Biol. 1984;98(5):1757–1762.
294. Sago CD, Lokugamage MP, Lando GN, et al. Modifying a commonly expressed endocytic receptor retargets nanoparticles in vivo. Nano Lett. 2018;18(12):7590–7600.
295. Pardi N, Tuyishime S, Muramatsu H, et al. Expression kinetics of nucleoside-modified mRNA delivered in lipid nanoparticles to mice by various routes. J Controlled Release. 2015;217:345–351.
296. Melo M, Porter E, Zhang Y, et al. Immunogenicity of RNA replicons encoding HIV Env immunogens designed for self-assembly into nanoparticles. Mol Therapy. 2019;27(12):2080–2090.
297. Karer M, Stiasny K, Zeitlinger M, Jilma B. Subcutaneous injection of mRNA vaccines against severe acute respiratory syndrome coronavirus 2: an option for severe bleeding disorders or anticoagulated patients?. Blood Coagulation Fibrinolysis. 2021;32(6):423.
298. Chen Q, Qi R, Chen X, et al. A targeted and stable polymeric nanoformulation enhances systemic delivery of mRNA to tumors. Mol Therapy. 2017;25(1):92–101.
299. Lindgren G, Ols S, Liang F, et al. Induction of robust B cell responses after influenza mRNA vaccination is accompanied by circulating hemagglutinin-specific ICOS+ PD-1+ CXCR3+ T follicular helper cells. Front Immunol. 2017;8:1539.
300. Zhang L, Wang W, Wang S. Effect of vaccine administration modality on immunogenicity and efficacy. Expert Rev Vaccines. 2015;14(11):1509–1523.
301. Lindsay KE, Bhosle SM, Zurla C, et al. Visualization of early events in mRNA vaccine delivery in non-human primates via PET–CT and near-infrared imaging. Nature Biomed Eng. 2019;3(5):371–380.
302. Schlake T, Thess A, Fotin-Mleczek M, Kallen KJ. Developing mRNA-vaccine technologies. RNA Biol. 2012;9(11):1319–1330.
303. Mockey M, Bourseau E, Chandrashekhar V, et al. mRNA-based cancer vaccine: prevention of B16 melanoma progression and metastasis by systemic injection of MART1 mRNA histidylated lipo polyplexes. Cancer Gene Ther. 2007;14(9):802–814.
304. Homayun B, Lin X, Choi HJ. Challenges and recent progress in oral drug delivery systems for biopharmaceuticals. Pharmaceutics. 2019;11(3):129.
305. Mauro VP, Chappell SA. A critical analysis of codon optimization in human therapeutics. Trends Mol Med. 2014;20(11):604–613.
306. Reddy ST, Rehor A, Schmoekel HG, Hubbell JA, Swartz MA. In vivo targeting of dendritic cells in lymph nodes with poly (propylene sulfide) nanoparticles. J Controlled Release. 2006;112(1):26–34.
307. Nicholson LB. The immune system. Essays Biochem. 2016;60:275–301.
308. Wang F, Kream RM, Stefano GB. An evidence-based perspective on mRNA-SARS-CoV-2 vaccine development. Med sci monitor. 2020;26:e924700–1.
309. Turner MR, Balu-Iyer SV. Challenges and opportunities for the subcutaneous delivery of therapeutic proteins. J Pharm Sci. 2018;107(5):1247–1260.
310. Usach I, Martinez R, Festini T, Peris JE. Subcutaneous injection of drugs: a literature review of factors influencing pain sensation at the injection site. Adv Ther. 2019;36:2986–2996.
311. Hess PR, Boczkowski D, Nair SK, Snyder D, Gilboa E. Vaccination with mRNAs encoding tumor-associated antigens and granulocyte-macrophage colony-stimulating factor efficiently primes CTL responses, but is insufficient to overcome tolerance to a model tumor/self-antigen. Cancer Immunol Immunother. 2006;55(6):672–683.
312. Tan L, Zheng T, Li M, et al. Optimization of an mRNA vaccine assisted with cyclodextrin-polyethyleneimine conjugates. Drug Deliv Transl Res. 2020;10:678–689.
313. Bahl K, Senn JJ, Yuzhakov O, et al. Preclinical and clinical demonstration of immunogenicity by mRNA vaccines against H10N8 and H7N9 influenza viruses. Mol Therapy. 2017;25(6):1316–1327.
314. Feldman RA, Fuhr R, Smolenov I, et al. mRNA vaccines against H10N8 and H7N9 influenza viruses of pandemic potential are immunogenic and well tolerated in healthy adults in phase 1 randomized clinical trials. Vaccine. 2019;37(25):3326–3334.
315. Zangi L, Lui KO, Von Gise A, et al. Modified mRNA directs the fate of heart progenitor cells and induces vascular regeneration after myocardial infarction. Nat Biotechnol. 2013;31(10):898–907.
316. Turnbull IC, Eltoukhy AA, Fish KM, et al. Myocardial delivery of lipidoid nanoparticle-carrying modRNA induces rapid and transient expression. Mol Therapy. 2016;24(1):66–75.
317. Patel S, Ryals RC, Weller KK, Pennesi ME, Sahay G. Lipid nanoparticles for delivery of messenger RNA to the back of the eye. J Controlled Release. 2019;303:91–100.
318. Ryals RC, Patel S, Acosta C, McKinney M, Pennesi ME, Sahay G. The effects of PEGylation on LNP-based mRNA delivery to the eye. PLoS One. 2020;15(10):e0241006.
319. Devoldere J, Peynshaert K, Dewitte H, et al. Non-viral delivery of chemically modified mRNA to the retina: Subretinal versus intravitreal administration. J Controlled Release. 2019;307:315–330.
320. Nabhan JF, Wood KM, Rao VP, et al. Intrathecal delivery of frataxin mRNA encapsulated in lipid nanoparticles to dorsal root ganglia as a potential therapeutic for Friedreich’s ataxia. Sci Rep. 2016;6(1):20019.
321. Anderson DM, Hall LL, Ayyalapu AR, Irion VR, Nantz MH, Hecker JG. Stability of mRNA/cationic lipid lipoplexes in human and rat cerebrospinal fluid: methods and evidence for nonviral mRNA gene delivery to the central nervous system. Hum Gene Ther. 2003;14(3):191–202.
322. Li Y, Su Z, Zhao W, et al. Multifunctional oncolytic nanoparticles deliver self-replicating IL-12 RNA to eliminate established tumors and prime systemic immunity. Nature Cancer. 2020;1(9):882–893.
323. Li Y, Teague B, Zhang Y, et al. In vitro evolution of enhanced RNA replicons for immunotherapy. Sci Rep. 2019;9(1):1.
324. Haabeth OA, Blake TR, McKinlay CJ, et al. Local delivery of Ox40l, Cd80, and Cd86 mRNA kindles global anticancer immunity. Cancer Res. 2019;79(7):1624–1634.
325. Hewitt SL, Bai A, Bailey D, et al. Durable anticancer immunity from intratumoral administration of IL-23, IL-36γ, and OX40L mRNAs. Sci Transl Med. 2019;11(477):eaat9143.
326. Kreiter S, Selmi A, Diken M, et al. Intranodal Vaccination with Naked Antigen-Encoding RNA Elicits Potent Prophylactic and Therapeutic Antitumoral ImmunityAntitumoral Immunity by Intranodal Vaccination with RNA. Cancer Res. 2010;70(22):9031–9040.
327. Mutsch M, Zhou W, Rhodes P, et al. Use of the inactivated intranasal influenza vaccine and the risk of Bell’s palsy in Switzerland. N Eng J Med. 2004;350(9):896–903.
328. Mai Y, Guo J, Zhao Y, Ma S, Hou Y, Yang J. Intranasal delivery of cationic liposome-protamine complex mRNA vaccine elicits effective anti-tumor immunity. Cell Immunol. 2020;354:104143.
329. Shakya AK, Chowdhury MY, Tao W, Gill HS. Mucosal vaccine delivery: Current state and a pediatric perspective. J Controlled Release. 2016;240:394–413.
330. Woof JM, Kerr MA. The function of immunoglobulin A in immunity. J Pathol. 2006;208(2):270–282.
331. Baba M, Itaka K, Kondo K, Yamasoba T, Kataoka K. Treatment of neurological disorders by introducing mRNA in vivo using polyplex nanomicelles. J Controlled Release. 2015;201:41–48.
332. Kulkarni JA, Darjuan MM, Mercer JE, et al. On the formation and morphology of lipid nanoparticles containing ionizable cationic lipids and siRNA. ACS Nano. 2018;12(5):4787–4795.
333. Crommelin DJ, Anchordoquy TJ, Volkin DB, Jiskoot W, Mastrobattista E. Addressing the cold reality of mRNA vaccine stability. J Pharm Sci. 2021;110(3):997–1001.
334. Pogocki D, Schöneich C. Chemical stability of nucleic acid–derived drugs. J Pharm Sci. 2000;89(4):443–456.
335. Holm MR, Poland GA. Critical aspects of packaging, storage, preparation, and administration of mRNA and adenovirus-vectored COVID-19 vaccines for optimal efficacy. Vaccine. 2021;39(3):457.
336. Chen S, Ren J, Chen R. Cryopreservation and desiccation preservation of cells. Compreh Biotechnol. 2019;5:157–166.
337. Alberer M, Gnad-Vogt U, Hong HS, et al. Safety and immunogenicity of an mRNA rabies vaccine in healthy adults: an open-label, non-randomized, prospective, first-in-human phase 1 clinical trial. Lancet. 2017;390(10101):1511–1520.
338. Abdelwahed W, Degobert G, Stainmesse S, Fessi H. Freeze-drying of nanoparticles: formulation, process and storage considerations. Adv Drug Deliv Rev. 2006;58(15):1688–1713.
339. Dolgin E. How COVID unlocked the power of RNA vaccines. Nature. 2021;589(7841):189–192.
340. Muramatsu H, Lam K, Bajusz C, et al. Lyophilization provides long-term stability for a lipid nanoparticle-formulated, nucleoside-modified mRNA vaccine. Mol Therapy. 2022;30(5):1941–1951.
341. Zhao P, Hou X, Yan J, et al. Long-term storage of lipid-like nanoparticles for mRNA delivery. Bioactive Materials. 2020;5(2):358–363.
342. Qin S, Tang X, Chen Y, et al. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduction Targeted Therapy. 2022;7(1):166.
343. Fiedler K, Lazzaro S, Lutz J, Rauch S, Heidenreich R. mRNA cancer vaccines. Curr Strategies Cancer Gene Therapy. 2016;2:61–85.
344. Coulie PG, Van den Eynde BJ, van der Bruggen P, et al. TumourantigensrecognizedbyTlymphocytes: At the core of cancer immunotherapy. Nat Rev Cancer. 2014;14:135–146.
345. Laureano RS, Sprooten J, Vanmeerbeerk I, et al. Trial watch Dendritic cell (DC)-based immunotherapy for cancer. Oncoimmunology. 2022;11(1):2096363.
346. Poorebrahim M, Abazari MF, Sadeghi S, et al. Genetically modified immune cells targeting tumor antigens. Pharmacol Ther. 2020;214:107603.
347. Restifo NP, Ying H, Hwang L, Leitner WW. The promise of nucleic acid vaccines. Gene Ther. 2000;7(2):89–92.
348. Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. Recognition of double-stranded RNA and activation of NF-κB by Toll-like receptor 3. Nature. 2001;413(6857):732–738.
349. Kato H, Takeuchi O, Sato S, et al. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature. 2006;441(7089):101–105.
350. Nallagatla SR, Hwang J, Toroney R, Zheng X, Cameron CE, Bevilacqua PC. 5’-triphosphate-dependent activation of PKR by RNAs with short stem-loops. Science. 2007;318(5855):1455–1458.
351. Jackson LA, Anderson EJ, Rouphael NG, et al. An mRNA vaccine against SARS-CoV-2—preliminary report. N Eng J Med. 2020;383(20):1920–1931.
352. Trepotec Z, Lichtenegger E, Plank C, Aneja MK, Rudolph C. Delivery of mRNA therapeutics for the treatment of hepatic diseases. Mol Therapy. 2019;27(4):794–802.
353. Magadum A, Kaur K, Zangi L. mRNA-based protein replacement therapy for the heart. Mol Therapy. 2019;27(4):785–793.
354. Warren L, Lin C. mRNA-based genetic reprogramming. Mol Therapy. 2019;27(4):729–734.
355. National Library of Medicine. Clinical trials.gov. Available from: https://clinicaltrials.gov/.
356. Yihunie W, Kebede B, Tegegne BA, et al. Systematic Review of Safety of RTS, S with AS01 and AS02 Adjuvant Systems Using Data from Randomized Controlled Trials in Infants, Children, and Adults. Clin Pharmacol. 2023;15:21–32.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.