Back to Journals » International Journal of Nanomedicine » Volume 20
Recent Advances in Biomimetic Drug Delivery for Rheumatoid Arthritis Treatment
Authors Xiang Y, Pan Z, Tian D, Zhang L, Dang W, Ye J, Xu X, Zhang Q
Received 16 July 2025
Accepted for publication 14 November 2025
Published 28 November 2025 Volume 2025:20 Pages 14253—14282
DOI https://doi.org/10.2147/IJN.S553826
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Jie Huang
Yucheng Xiang,1,2,* Zongyu Pan,1,* Deyong Tian,2 Lie Zhang,1,3,4 Wantai Dang,3,5 Jing Ye,2 Xiaohong Xu,2,6 Quan Zhang1– 3,7
1Department of Neurosurgery, The First Affiliated Hospital of Chengdu Medical College, Chengdu, People’s Republic of China; 2Key Laboratory of Structure-Specific Small Molecule Drugs, Institute of Materia Medica, School of Pharmacy, Chengdu Medical College, Chengdu, People’s Republic of China; 3Development and Regeneration Key Laboratory of Sichuan Province, Chengdu Medical College, Chengdu, People’s Republic of China; 4School of Clinical Medicine, Chengdu Medical College, Chengdu, People’s Republic of China; 5Department of Rheumatology and Immunology, The First Affiliated Hospital of Chengdu Medical College, Chengdu, People’s Republic of China; 6Xichang Medical College, Xichang, People’s Republic of China; 7Chengdu Nature’s Grace Biological Technology Co., Ltd, Chengdu, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaohong Xu, Email [email protected] Quan Zhang, Email [email protected]
Abstract: Rheumatoid arthritis (RA) affects approximately 1% of the global population, causing debilitating joint pain and often leading to severe disability. Although conventional treatments can control the initial symptoms of RA, there is no curative treatment strategy for RA. Biomimetic nanomedicine has emerged as a promising therapeutic approach, leveraging the integration of nanoparticles with natural biomaterials to achieve targeted drug delivery and improved treatment outcomes. Beyond exogenous nano-delivery systems, the natural biomimetic strategy might offer superior biocompatibility and lower immunogenicity. This review summarizes the latest advancements in biomimetic drug delivery systems for RA and highlights the underlying mechanisms for these biomimetic carriers. We also discuss the critical factors influencing the transition of these biomimetic nanomedicines from laboratory research to clinical implementation. By emphasizing the transformative potential of biomimetic strategies in RA treatment, this review aims to provide new insights and directions for future research and clinical applications in this field.
Keywords: rheumatoid arthritis, biomimetic drug delivery system, targeting therapy, biomaterials
Introduction
Rheumatoid arthritis (RA) is an autoimmune disease characterized by synovitis and cartilage damage.1,2 The exact mechanisms causing RA remain unclear, but it might be triggered by both genetic and environmental factors. Growing evidence demonstrated that the interplay between the innate and adaptive immune systems is closely associated with the pathogenesis of RA.3 Various immune cells continuously infiltrate into the synovial membrane of the joints, including T cells, B cells, macrophages, synovial fibroblasts, mast cells, endothelial cells, neutrophils, dendritic cells (DCs), and natural killer cells. This cellular infiltration promotes secretion, leading to synovitis. Ultimately, invasive pannus and cartilage destruction would occur.4,5
1% of the global population suffer from RA, causing significant socioeconomic burden worldwide.6,7 Due to the complex pathogenic mechanism of RA, there is no satisfactory cure yet. Current treatments are based on surgery and pharmacotherapy (Figure 1) as The treatment mainly includes disease-modifying antirheumatic drugs (DMARDs), biologics, nonsteroidal anti-inflammatory drugs (NSAIDs), and glucocorticoids, and gene therapy.8 However, there are various limitations including low absorption rates at the site of injury, poor targeting specificity, and potential immune system impairment, which severely hinder their therapeutic efficacy.1 The mechanisms of these medications usually act on the immune response suppression or the characteristic inflammatory factor inhibition, resulting into the RA associated symptoms improvement. Due to the inherent short circulation time and uncontrollable dissemination throughout the body, the risky side effects of these medications on extra-articular organs are inevitable Typically, glucocorticoids which have remarkable anti-inflammatory and immunomodulatory effects are widely used to treat rheumatoid arthritis. However, weight gain cushinoid facies, peptic ulcer and even psychosis were observed in patients treated with glucocorticoids.9 Moreover, methotrexate which functions as an analog of folic acid might cause liver and pulmonary damage. Immunosuppressive sulfasalazine might cause gastrointestinal, central nervous system, and hematologic side effects. Even the kinase inhibitors including Tofacitinib might result into zoster infection and other potential side effects.10 Therefore, more advanced drug delivery technology is required to liberate patients suffered from RA.
 |
Figure 1 Summary of traditional treatment strategies and therapeutic targets for rheumatoid arthritis. Traditional treatment strategies: (A) Glucocorticoids; (B) Nonsteroidal Anti-inflammatory Drugs; (C) Disease-Modifying Antirheumatic Drugs; (D) Biologics. Therapeutic target for rheumatoid arthritis; (E) Cartilage; (F) Vasculature; (G) Synovial; (H) Immune Cells; (I) Osteoclasts. |
The management of RA usually require long-term drug administration which might amplify the side effects. Thus, developing nanomedicine which can enhance the efficacy and reduce unwanted adverse effects is beneficial for RA treatment. Although multiple organic and inorganic nanoparticles have been engineered to promote the drug delivery, few of them march toward clinical trials. In this process, advances in comprehending how biological particles including cells, pathogens, and organelles interact with bodily and cellular mechanisms have motivated attempts to mimic their structures and behaviors for the design of innovative nanomedicine systems. Biomimetic nanotechnology, which combines the advantages of nanoparticles with natural biomaterials, holding significant promise for future applications.11,12 Generally, Nanoparticles can be designed to respond upon specific pathological factors, thereby realizing precise drug delivery. Additionally, biomimetic technology enhances molecular imaging and improves the bioavailability of drugs. Furthermore, biomimetic nanoparticles possess the ability to selectively bind to pathological targets associated with RA. Moreover, compared with conventional nano-delivery systems, the utilization of natural bionic drug delivery systems might have additional higher biocompatibility. The conventional chemically synthesized systems often showed unexpected immunogenicity, which is risky in clinical administration. By contrast, nanomedicines based on endogenous materials such as albumin have negligible immunogenicity and prolonged circulation time. In addition, biomimetic nanocarriers have already enter clinical trials including NCT00110695 (albumin-based nanoparticle), NCT01159288 (dendritic cell-derived exosomes) and NCT04592484 (nanoscale exosome-based delivery system).13 When it comes to RA treatment, several preclinical studies showed great potential. The pharmacokinetic study revealed that the biomimetic dual-target drug delivery system developed by Na Jia et al showed over 10 times higher accumulation in inflamed joints after 24h injection, compared with free small-molecular group.14 Huichao Xie et al used macrophage membrane-based strategy realized 35.12 times higher accumulation in the paws of the collagen-induced arthritis model after 2h administration, compared with healthy group.15 This review will discuss the limitations of traditional therapeutic drugs, summarize various therapeutic targets of RA in biomimetic drug delivery systems, and outlines commonly used biomimetic nanocarriers while proposing strategies to advance the field of RA nano-therapy.
Traditional Pharmacological Treatment Strategies
Currently, commonly used medications for RA include glucocorticoids, NSAIDs, synthetic DMARDs and certain biologics such as tumor necrosis factor (TNF) inhibitors, co-stimulatory modulators, interleukin-6 (IL-6) inhibitors, and B-cell depleting agents. (as shown in Figure 1) However, these treatments have notable drawbacks, including significant adverse reactions, poor stability, and nonspecific targeting, which still need to be addressed.6,16,17
Glucocorticoids
Glucocorticoids, such as dexamethasone, methylprednisolone, prednisone, and triamcinolone, are a class of chemicals with potent anti-inflammatory and immunosuppressive properties. They can bind to glucocorticoid receptors, forming glucocorticoid-receptor complex. This complex then binds to the glucocorticoid response units in the nucleus, changing the transcription of genes associated with the inflammatory response and inhibiting the release of phospholipids which contribute to joint inflammation.1,18 However, long-term use of glucocorticoids result in various adverse effects, including osteoporosis, diabetes, metabolic syndrome, cataracts, cardiovascular diseases, and peptic ulcers.19
NSAIDs
NSAIDs can be classified as follows: non-selective NSAIDs (for example ibuprofen and naproxen), semi-selective NSAIDs (for example diclofenac and indomethacin), and selective COX-2 inhibitors (for example rofecoxib and celecoxib).8 These drugs are used to manage early pain and stiffness in RA patients due to their anti-inflammatory, analgesic, and antipyretic effects. Their mechanism of action primarily involves the inhibition of inflammatory mediators in the cyclooxygenase (COX) pathway. However, the efficacy of these drugs is limited, only providing significant pain relief primarily in the early stages of the disease. Furthermore, NSAIDs might induce a variety of adverse reactions, including blood pressure fluctuations, thrombocytopenia, renal insufficiency, gastrointestinal discomfort, and an increased risk of cardiovascular events.20
DMARDs
Synthetic DMARDs include methotrexate (MTX), hydroxychloroquine, sulfasalazine and leflunomide. These drugs exhibit specific antirheumatic activity and are often used in combination with other therapeutic agents, (for example) glucocorticoids.6 However, the underlying mechanisms for DMARDs are not fully understood, recent studies reported that DMARDs can initiate multiple signaling pathways improving RA. Due to their low cost and favorable efficacy, DMARDs are widely preferred in clinical. However, their use is associated with several adverse effects, including liver dysfunction, pneumonia, bone marrow suppression, nephrotoxicity, allergic reactions, and retinal changes.21
Biologics
Biologics can inhibit specific molecular pathways involved in inflammatory process of RA.8 Cytokine antagonists, such as infliximab, etanercept, adalimumab, and golimumab, block TNF, which are beneficial for RA treatment. Other biologics, including anakinra (which blocks the IL-1 receptor), tocilizumab (an IL-6 receptor inhibitor), rituximab (a B-cell depleting agent), abatacept (a T-cell co-stimulation modulator), and kinase inhibitors (such as p38 MAPK inhibitors, Syk inhibitors, and JAK inhibitors like tofacitinib, and IkB inhibitors), can also be used to treat RA.6 During early RA, biologics combined with methotrexate demonstrate high efficacy. However, in patients with long-term condition, the efficacy of biologics is less satisfactory. Furthermore, there are several challenges in biologic therapies including the risk of tuberculosis, hepatitis B reactivation and high treatment costs.22
RA Therapeutic Targets
Recent studies have indicated that both the innate and adaptive immune systems play pivotal roles in the pathogenesis of RA. A variety of inflammatory molecules and immune cells are involved in the complex pathological process. Inflammatory molecules such as interleukin-4 (IL-4), interleukin-10 (IL-10), interleukin-15 (IL-15), interleukin-17 (IL-17), interleukin-18 (IL-18), interleukin-23 (IL-23), interleukin-1 receptor-associated kinase (IRAK)-4, as well as small molecule metabolites such as prostaglandins (PGs), lipoxins (LXs), platelet-activating factor (PAF), leukotrienes (LTs), nitric oxide (NO), and reactive oxygen species (ROS), are all critical participants in the pathophysiology of RA.6,7 Additionally, immune cells, such as neutrophils, can induce the production of self-antigens, thereby affecting the immune response.23 DCs, upon stimulation by specific environmental factors, can initiate innate immune responses. This activation leads to the subsequent stimulation of T cells, B cells, macrophages, synovial cells, chondrocytes, and osteoclasts. Consequently, pro-inflammatory cytokines and cartilage-degrading factors, such as interleukin-1 beta (IL-1β), interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and matrix metalloproteinases (MMPs), are secrete.24 Therefore, targeting the molecular and cellular components involved in the inflammatory process of RA represents a key therapeutic strategy.
Cartilage
Erosion of articular cartilage is a hallmark feature of RA. Under the attack of cytokines, the cartilage matrix is degraded by MMPs and other related enzymes, leading to metabolic disturbances and damage to chondrocytes.25 Since cartilage damage is irreversible, it is crucial to address and repair cartilage damage in the early stages of RA. Articular cartilage is primarily composed of water and two key solid components: cartilage collagen and aggrecan. The collagen in cartilage consists mainly of type II collagen, with a smaller proportion of type I collage.26 In 2008, Rothenfluh et al identified a peptide, WYRGRL, which binds to the α1 chain of type II collagen, making it one of the most widely studied collagen-targeting molecules.27–29 Chondrocytes, the primary cellular component of cartilage tissue, are also essential targets for RA therapies. Recent studies have shown that NF-κB plays a crucial role in regulating both the normal development and pathological destruction of cartilage in joint inflammation, providing a potential therapeutic target for RA.30
Vasculature
Angiogenesis is a critical pathological feature in early RA, contributing to synovial proliferation. It also plays a key role in both the maintenance and exacerbation of RA. Neovascularization arises from the sprouting of endothelial cells (ECs) in capillaries. During the highly inflammatory phase, immune cells such as CD4+ T cells and B cells continuously infiltrate into the inner layer of the synovium, forming pannus. In this pathological environment, these infiltrating cells produce a large number of pro-angiogenic signals that stimulate endothelial cell proliferation. The rate of cellular proliferation in this environment often exceeds the supply of nutrients and oxygen, leading to the secretion of growth factors, cytokines, and chemokines, which further promote vascular growth.31 At this stage (the vascular phase), the density of capillaries increased significantly, providing nutrition and oxygen to the pannus. However, this phenomenon destruction of cartilage and bone tissue, thereby worsening the RA condition.32,33 Angiogenesis in RA is regulated by pro-angiogenic and anti-angiogenic mediators, including growth factors, cytokines, chemokines, cell adhesion molecules, and MMPs. These findings open up new possibilities for targeting angiogenesis. Vascular endothelial growth factor (VEGF) is a critical signaling protein involved in angiogenesis and expressed in RA synovial tissue, bodily fluids, and serum.34 Among the VEGF subtypes, VEGF165 plays a central role in the pathological process of RA, interacting with VEGF-R1 and VEGF-R2. High expression levels of VEGF have been observed in sub-synovial macrophages, fibroblasts surrounding micro-vessels, vascular smooth muscle cells, and synovial lining cells.35 Therefore, targeting the VEGF pathway might offer potential therapeutic benefits for RA.
Synovial Fluid
The synovium is a connective tissue lining the inner surface of the joint capsule and served as an essential pathway for drug delivery. Synovitis is also a major pathological feature of RA, making synovial tissue an important target for RA therapies. The synovium consists of a lining layer consist of macrophages and fibroblast-like synoviocytes (FLS), and a layer composed of vascularized connective tissue. Synovial cells are generally classified into two main types: macrophage/monocytes and fibroblast-like synoviocytes.36
Macrophages
Macrophages produce both pro-inflammatory and anti-inflammatory mediators via the activation of Toll-like receptors (TLRs), facilitating the recognition, phagocytosis, and destruction of pathogens.37 Under normal conditions, synovial macrophages are important in maintaining the homeostasis of joint tissue. However, in RA, macrophages derived from synovial lining cells and monocytes participate in immune responses, releasing cytokines and enzymes related to inflammation. This further activates osteoclasts and fibroblasts, exacerbating the disease.38,39 Macrophages exhibit multi-faceted effect and can turn into different phenotypes, producing distinct subsets of mediators.3 The M1 and M2 phenotypes represent two extremes of macrophage activation.40,41 M1 macrophages are pro-inflammatory and are considered as major contributors to RA progression.42 The interaction between activated M1 macrophages and helper T cells (Th1) leads to the production of a variety of pro-inflammatory mediators, promoting the progression of RA.3 In contrast, M2 macrophages secrete anti-inflammatory factors. The shift from M1 to M2 macrophages is influenced by various metabolic pathways, surface markers, and cytokine production.43 Therefore, inducing apoptosis in M1 macrophages or reprogramming them to the M2 phenotype has emerged as an important therapeutic strategy. Current targeting strategies for RA focus on receptors such as the folate receptor (FA), mannose receptor, scavenger receptor (SR-A), p65, and matrix metalloproteinase.44–48
Fibroblast-Like Synoviocytes
Fibroblast-like synoviocytes (FLS) are specialized mesenchymal cells found in the synovial lining of joints. Under normal conditions, FLS regulate the composition of the extracellular matrix (ECM) and synovial fluid by secreting hyaluronic acid and lubricin, thereby effectively lubricating and nourishing the articular cartilage.49,50 However, during the progression of RA, FLS exhibit more invasive behaviors.51 Studies have demonstrated that FLS extracted from RA patients display pathogenic characteristics when transplanted into mice.52 During active RA, the number of FLS increases significantly, and these cells resist apoptosis through endoplasmic reticulum-related mechanisms.53 Moreover, FLS function as immune modulators, promoting the secretion of cytokines, chemokines, and angiogenic factors, thereby stimulating the proliferation and infiltration of immune cells and the formation of new blood vessels.32,51,54 Upon the pannus-cartilage interface in RA-affected joints, FLS are responsible for the overproduction of MMPs (such as MMP-1, MMP-3, and MMP-13), which further damage the collagen structure of joint tissues.34,51 In RA, FLS express various surface receptors and markers, including cadherin-11 (CDH11), podoplanin (PDPN), CD55+, and CD90+, which are highly expressed on FLS.34,55
Immune Cells
RA is an autoimmune disorder characterized by dysregulated immune responses that result in joint tissue damage. Immune cells, including T cells, B cells, and neutrophils, play critical roles in the pathogenesis of RA.
T Cells
Autoimmunity in RA is generally believed to be initiated by the production of antibodies against post-translationally modified proteins. T cells are known to interact with these modified proteins, such as citrullinated and carbamylated proteins.4 Among the 100 non-HLA genes associated with RA susceptibility, many of them are involved in T cell selection, maturation, and function, thereby underscoring the pivotal role of T cells in the disease’s pathogenesis.56,57 A significant reduction in the frequency of naive CD4+ T cells, which developed into pro-inflammatory T cells with enhanced tissue invasiveness and proliferative capacity.58 At this stage, insufficient regulation of mitochondrial DNA (mtDNA) leads to abnormal differentiation of CD4+ T cells, which develop into pro-inflammatory T cells with heightened tissue invasiveness and proliferative capacity.59 Short-lived effector T cells that differentiate from naive CD4+ T cells enter the synovial tissue, where they undergo pyroptosis, triggering a robust inflammatory response.59–61 The abnormal activation and immune damage of these T cells not only induce immunogenic cell death and mitochondrial DNA damage, but also promote T cell pyroptosis and an imbalance in reactive oxygen species (ROS), These phenomena further exacerbate RA inflammation.62 Consequently, therapeutic strategies targeting helper T cells and regulatory T cells are common in RA treatment.63,64
B Cells
B cells exert multiple functions in the complex pathophysiology of RA. In addition to antibody production, B cells contribute to antigen presentation and cytokine secretion, which are closely linked to the development of RA.65,66 B cells are capable of engulfing and processing antigens, thereby stimulating CD4+ T cells and initiating the immune response.67,68 Furthermore, B cells produce autoantibodies, such as rheumatoid factor (RF) and anti-citrullinated protein antibodies (ACPA), which promote osteoclast differentiation and the destruction of joint bone structure.69–71 B cells also secrete a range of cytokines that interact with other immune cells, thereby promoting the inflammatory cascade in RA. Notable cytokines include TNF-α and IL-6. TNF-α activates inflammatory cells, while IL-6 is involved in both B cell differentiation and antibody production, as well as in modulating T cell responses.3,72,73 During their development, B cells pass through multiple checkpoints, which influence their function through receptors on the B cell surface and their corresponding ligands.66 For example, checkpoints such as IL-6, IL-21, FcγRIIB, and CD38 play critical roles in regulating B cell development and function. These intricate mechanisms collectively facilitate the onset and progression of RA.74–78 Notably, as reported by Espéli et al and Barlev et al, there are limitations of targeting FcγRIIB. Firstly, FcγRIIB is widely expressed not only on B cells but also on myeloid cells and other cells. Rashly targeting FcγRIIB solely might lead to immune dysregulation. Secondly, the expression and function of FcγRIIB vary across B cell subsets. Some B cells respond to FcγRIIB inhibition weakly.
Neutrophils
Neutrophils are essential in the pathogenesis of RA, which are also abundant in the inflamed joints of RA patients.79 Early in the disease, neutrophils become abnormally activated by cytokines, chemokines, and autoantibodies, initiating a cascade of inflammatory responses and tissue damage.80,81 Activated neutrophils release additional cytokines and chemokines, which exacerbate the inflammatory response, creating a vicious cycle of tissue damage.82 Furthermore, neutrophils contain a variety of soluble proteases, such as neutrophil elastase (NE) and proteinase 3 (PR3).82 Upon activation, these proteases are released extracellularly, contributing to the degradation of articular cartilage. Neutrophils also increase various receptor levels, including the interleukin-6 receptor, which might influence the inflammatory microenvironment in the joint and affect signaling between pathological cells.83 In brief, interleukin-6 (IL-6) is a key player in chronic inflammation including RA and cancer, particularly via the IL-6–signal transducer and activator of transcription 3 (STAT3) signaling axis and the IL-6 amplifier (IL-6 Amp) mechanism. Thus, weakening the IL-6 activation via competitive inhibition or downregulating receptor expression is a potential strategy to benefit RA treatment. Under certain conditions, neutrophils release neutrophil extracellular traps (NETs), which provide autoantigens.84 NETs, composed of DNA, histones, and antimicrobial peptides, serve to capture and eliminate pathogens. However, in the context of RA, excessive NET release can lead to self-tissue damage, thereby promoting RA inflammation. Consequently, several potential therapeutic targets have emerged, including the Fc γ RIIIb receptor, PI3 kinase, and granulocyte-macrophage colony-stimulating factor (GM-CSF).85–88 These targets offer novel avenues for RA treatment.
Osteoclasts
Bone homeostasis in the joints is maintained through a dynamic equilibrium between bone resorption by osteoclasts and bone formation by osteoblasts.89 However, in RA, the activation of the immune system directly affects bone cells or stimulates joint cells (such as fibroblasts), disrupting bone homeostasis. Osteoclasts are the only cells in the human body capable of resorbing and degrading bone tissue.90 The differentiation and formation of osteoclasts are regulated by various factors, with receptor activator of nuclear factor kappa-B ligand (RANKL), a TNF family cytokine, and macrophage colony-stimulating factor (M-CSF) being the primary regulators.89 In RA, as previously mentioned, cytokines such as TNF-α and IL-6 can directly promote the differentiation of osteoclast precursors into multinucleated osteoclasts.91,92 Pathologically, the differentiation pathways diverge such cytokine-induced osteoclasts contributing to joint invasion, while RANKL-differentiated osteoclasts are primarily involved in osteoporosis.93,94 As highlighted by Iwamoto and Kawakami, the differentiation of monocytes into osteoclasts represents a pivotal mechanism in joint destruction during rheumatoid arthritis. Future research could therefore focus on targeting this process. The osteoclast-associated receptor (OSCAR) is upregulated in patients with RA and the OSCAR-collagen recognition stimulates osteoclastogenesis. Thus, ligands based on type I collagen (COLI) and type II collagen (COLII) have been investigated as potential targets for osteoclasts.95 Moreover, modulating M1/M2 ratio and regulating monocyte-derived dendritic cells (Mo-DC) differentiation are also promising.
Biomimetic Drug Delivery Strategies
Nanocarrier Materials
Due to the highly complex pathological environment in RA, the efficacy of free drugs is often suboptimal. However, the inflamed or acidic microenvironment may facilitate the passive or active accumulation of nanoparticles (NPs).96 Nanocarrier drug delivery systems can specifically target therapeutic agents to the site of disease while protecting the drugs from degradation (as shown in Table 1 and Figure 2). In addition, encapsulating free drugs within delivery systems can extend their circulation time, reduce systemic side effects. Furthermore, it also enables drugs with environment-responsive release properties tailored to the unique physiological and pathological features of RA. Despite the considerable potential of nanodrug delivery systems, several challenges remain. For instance, plasma proteins might adsorb onto nanoparticle surface, hindering their effective migration to the target site.97 Moreover, NPs that accumulate in inflamed joints probably release the drug to various cells within the joint cavity.98 Consequently, minimizing the clearance rate of nanoparticles from the body and enhancing their selective accumulation in target areas are critical for the clinical application of nanotechnology.
 |
Figure 2 Schematic diagrams of nanodrug delivery system therapeutic strategies for rheumatoid arthritis treatment. |
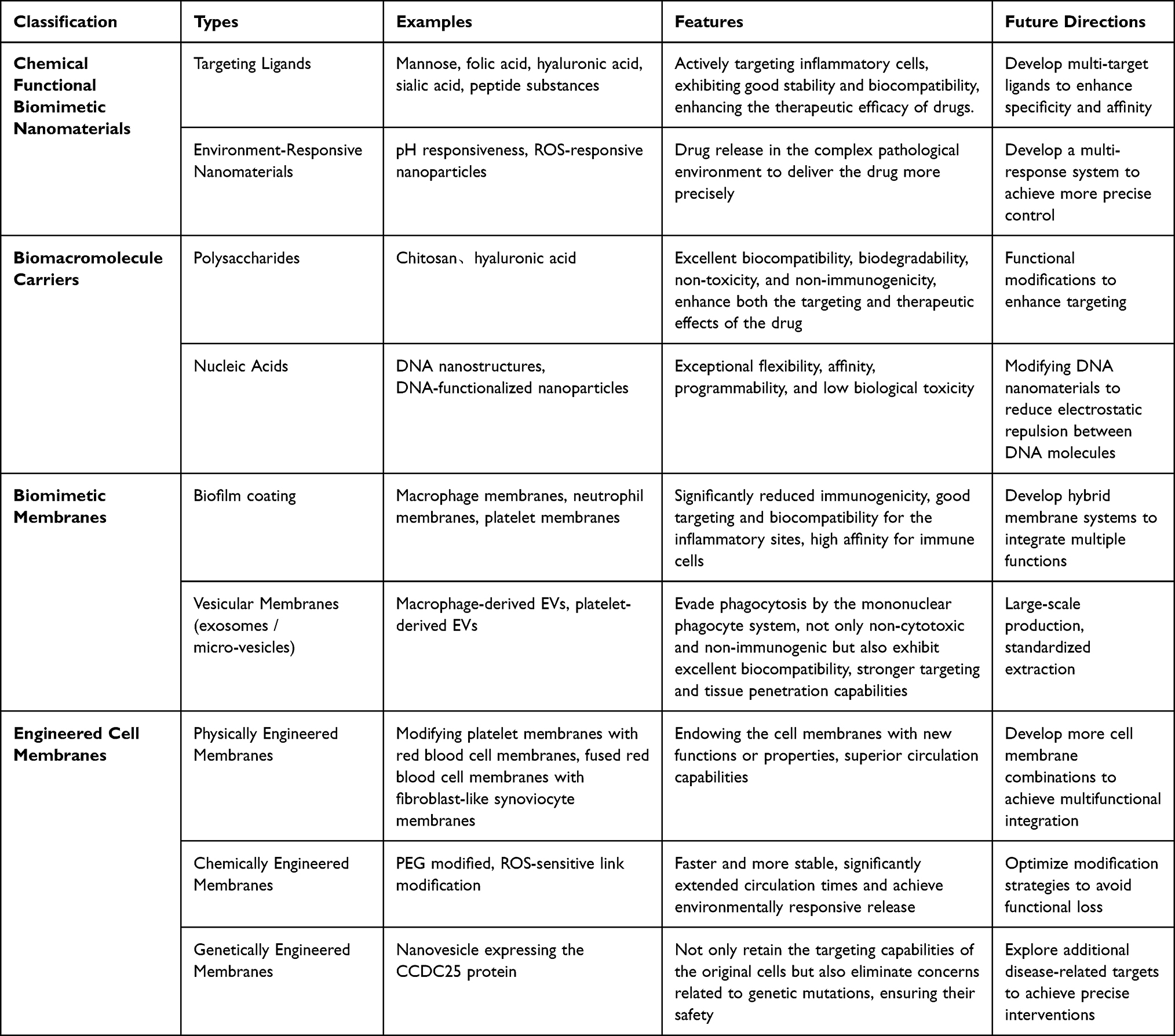 |
Table 1 Current Research Status and Future Directions of Biomimetic Drug Delivery in Rheumatoid Arthritis Treatment |
Chemical Functional Biomimetic Nanomaterials
Targeting Ligands
In RA, certain receptors are overexpressed on the surface of immune cells. Consequently, small molecules ligands (mannose,99 folic acid100), macromolecules ligands (hyaluronic acid, sialic acid101) or peptide substances102 have been employed to develop ligands for the active targeting of inflammatory cells. Yang et al developed a mannose-conjugated polymeric (Man-PMTX) nanoformulation capable of effectively targeting macrophages and delivering the anti-rheumatic drug methotrexate (MTX).103 Compared with free MTX, Man-PMTX exhibited significantly enhanced targeting and was more readily taken up by macrophages. In in vivo biodistribution assay, the images revealed that the Cy5 labeled Man-PMTX exhibited about 2 times higher accumulation in the knees of rheumatoid arthritis mice models after 4h administration, compared with non-modified nanoparticle treated group. There was no obvious damage observed in major organs among free methotrexate and Man-PMTX groups. The in vivo model results demonstrated that this nanoformulation promoted the polarization of macrophages from the M1 to the M2 phenotype while reducing the release of inflammatory factors. These findings provide novel insights into the treatment of RA, indicating that a nanodrug delivery system targeting macrophages can effectively modulate immune responses and alleviate inflammation. To specifically target rhein (RH) to inflamed synovial macrophages, Zhou et al designed a cerium oxide nanoenzyme and hyaluronic acid-complexed ROS-responsive nanomicelle.104 This nanomicelle leverages the redox properties of cerium oxide nanoenzymes to neutralize reactive oxygen species (ROS) produced by M1 macrophages, while rhein induces the polarization of M1 macrophages by inhibiting their signaling pathways. In vivo models demonstrated that this nanomicelle significantly reduced the number of M1 macrophages, while the proportion of M2 macrophages increased. Moreover, the levels of inflammatory factors, such as TNF-α and IL-6, were notably reduced. Guo et al developed a sialic acid (SA)-poly(lactic-co-glycolic acid) (PLGA) nanoformulation carrying antagomiR-7.105 In animal models, this nanoformulation was shown to specifically target B cells in systemic lupus erythematosus (an autoimmune disease) and effectively suppress B cell activation (Figure 3).
 |
Figure 3 (A) Schematic illustration of SA-PLGA@antagomiR-7 inhibits B cell overresponse to alleviate systemic lupus erythematosus. (B) TEM images of PLGA@antagomiR-7 (Left) and SA-PLGA@antagomiR-7 (Right). (Scale bar, 100 nm). (C) Serum levels of anti-dsDNA Ab, IL-6, IgG, and IL-10 were determined by ELISA. (D) In vivo fluorescence images of C57BL/6 mice at different times after intravenous injection of DIR-PLGA@antagomiR-7 or DIR-SA-PLGA@antagomiR-7. (E) Semi-quantitative analysis of fluorescence images of C57BL/6 mice (n=3). The white label corresponds to the fluorescence intensity value at that point. nsP > 0.05 indicated no significant difference, *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001. Referenced from Ref.105 |
Common nanomedicines, such as liposomes and solid lipid nanoparticles are also widely used in the treatment of RA. These materials exhibit good stability and biocompatibility, enhancing the therapeutic efficacy of drugs. Li et al combined liposomes with folic acid to develop an oral nano-delivery system (FA-LP-GCK) for encapsulating a ginsenoside mixture K (GCK) to treat RA.106 The results indicated that this nanoformulation significantly reduced the levels of inflammatory factors and effectively inhibited synovial hyperplasia. Recently, Mirchandani et al designed a solid lipid nanoparticle (SLN) combined with a hyaluronic acid nanoformulation (HA-MTX-SLN) for loading methotrexate (MTX).107 In adjuvant-induced mouse models, this nanoparticle demonstrated high encapsulation efficiency and effective targeting, ensuring localization to the inflammatory site while significantly reducing drug toxicity (Figure 4). Other studies have shown that mannose-modified morin liposomes (ML-Morin) can effectively inhibit inflammatory responses and the generation of osteoclasts.108
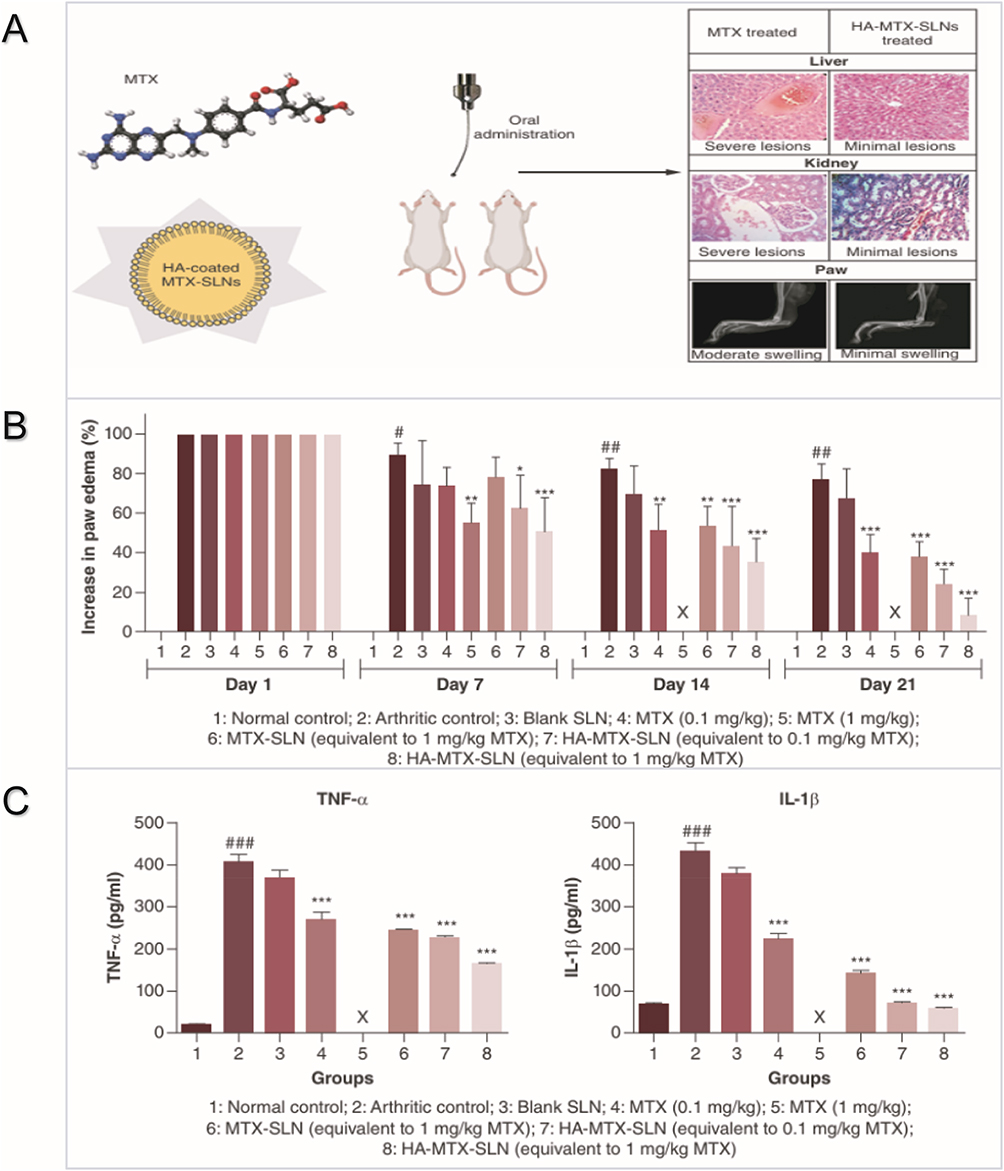 |
Figure 4 (A) Solid lipid nanoparticles (SLNs) as carriers for MTX to achieve improved efficacy in RA treatment at reduced doses, thus decreasing the potential toxicity of the drug. (B) Effect of MTX, MTX-SLN and HA-MTX-SLN formulations on percentage increase in paw edema in CFA-induced arthritic rats from day 1 to day 21. Values are expressed as mean ± standard deviation. #P < 0.05 and ##P < 0.01 considered significant with respect to arthritic control group on day1; X indicates that the rats have died. *P< 0.05; **P < 0.01; ***P < 0.001 considered significant with respect to arthritic control group on respective days. (C) Effect of MTX, MTX-SLNs and HA-MTX-SLNs on TNF-α and IL-1β levels in arthritis-induced rats. Values are expressed as mean ± standard deviation. ###P < 0.001 considered significant with respect to normal control group; ***P < 0.001 considered significant with respect to normal control group. Referenced from Ref.107 |
Environment-Responsive Nanomaterials
Given the complex pathological environment of RA, which includes inflammation and an acidic pH. Researchers have developed various multifunctional nanomaterials responsive to these environmental conditions. To improve the bioavailability and targeting of gentistic acid (the main component of Gentiana), Jia et al designed a dual-targeting nanoparticle formulation (RBA-NPs) with pH responsiveness and the ability to target CD44 and folate receptors.14 In a rat model of RA, this nanoparticle formulation was able to deliver the drug more precisely to the site of action compared to the free drug, thereby enhancing targeting. Additionally, RBA-NPs promoted the repolarization of M1 macrophages, demonstrating favorable therapeutic effects. Li et al designed a hyaluronic acid hydrogel nanoparticle (RGD-conjugated gold nanoparticles) loaded with triptolide (TP).109 This nanoparticle not only targets drug delivery via hyaluronic acid but also alleviates inflammation in RA through photothermal therapy. In a collagen-induced mouse model, low doses of the hydrogel nanoparticle combined with near-infrared irradiation exhibited significant therapeutic effects, reducing the migration and joint-erosion capabilities of synovial fibroblasts (FLS) (Figure 5). Other studies have shown that enzyme-responsive nanoparticles modified with arginine-glycine-aspartate (RGD) peptides, in combination with αvβ3 integrin-loaded celastrol, can promote apoptosis of inflammatory macrophages and osteoclasts, thereby increasing the bioavailability and therapeutic efficacy of celastrol.110,111
 |
Figure 5 (A) Preparation process of TP-PLGA-Au@RGD/HA hydrogels and schematic illustration of the anti-inflammatory effect of TP-PLGA-Au@RGD/HA hydrogels in collagen-induced arthritis mice. (B) Profiles of TP release from TP-PLGA-Au@RGD/HA hydrogels and TP-PLGA-Au@RGD NPs under initial NIR irradiation for 10 min. Profiles of TP release from TP-PLGA-Au@RGD/HA hydrogels with or without NIR irradiation for 10 min. Data are expressed as mean values for n = 3, and error bars represent the standard deviation of the means (*P < 0.05). (C) Histological findings of synovial tissues from healthy mice and CIA mice on day 28 after different treatments. H&E (synovial inflammation, original magnifications × 100, and immunohistochemical staining for IL-1β, IL-6, and TNF-α, original magnifications × 20). The arrow indicates the area with significant infiltration of inflammatory cells. Semiquantitative analysis of histopathological evaluation (synovial inflammation and immunohistochemical staining for IL-1β, IL-6, and TNF-α). Asterisks (*) represent significance compared with the untreated mice at *P < 0.05. Referenced from Ref.109 |
Biological Macromolecular Carriers
In recent years, the use of biomacromolecules as carriers in the construction of nanodrug delivery systems (NDDS) has garnered increasing attention. Natural biomacromolecules such as albumin, lipoproteins, and polysaccharides exhibit favorable biocompatibility, renewability, targeting capabilities, and low biological toxicity.112–123 These natural materials are considered safer and more stable compared to synthetic alternatives. For example, Ting Gong et al designed a human serum albumin-based nanoparticle named HSA-HS15. In biodistribution assay, the joint accumulation of DiD labeled HSA-HS15 is much higher than that of free DiD (over 4 times higher) after 4h injection, exhibiting inflammatory targeting ability. After loading celastrol as therapeutic agent, celastrol loaded HSA-HS15 showed enhanced biosafety, compared with free celastrol. After treatment, obvious nephrotoxicity, cardiotoxicity and potential hepatotoxicity was observed in rats received free celastrol treatment. In contrast, the major organs in rats treated with celastrol loaded HSA-HS15 were not damaged and the biochemical indicators were similar to normal rats.
Polysaccharides
Polysaccharides, including chitosan and hyaluronic acid, are natural materials known for their hydrophilicity, mechanical stability, tunability, and renewability, making them widely used in the medical field as nano-delivery materials.124 This section focuses on the application of chitosan and hyaluronic acid as biomimetic carriers in the treatment of RA.125
Chitosan is a natural polymer with excellent biocompatibility, biodegradability, non-toxicity, and non-immunogenicity.126 As a naturally cationic polymer, chitosan shares physicochemical properties and biological characteristics with glycosaminoglycans which are major components of the extracellular matrix (ECM), Thus, the chitosan exhibits a strong affinity for certain cell membranes. Consequently, chitosan is frequently used as a biomimetic carrier for the treatment of various diseases including RA. Numerous studies have reported the use of chitosan in combination with commonly used anti-rheumatic drugs, such as methotrexate (MTX).127,128 Jabbari et al developed a nanoformulation encapsulating eugenol within chitosan, exploiting eugenol’s antioxidant properties for the treatment of RA.129 In a comparative study with MTX in a mouse model, the eugenol nanoformulation was shown to reduce malondialdehyde (MDA) levels (a marker of oxidative stress) and the expression of FOXO3 protein, while inhibiting the expression of inflammatory mediators such as TGF-β and MCP-1. To enhance the functionality of chitosan, Siddiqui et al designed a folate-modified chitosan (CHI)-chondroitin sulfate (CHS) nanoparticle encapsulating leflunomide (LEF) in a hydrogel nanoparticle formulation. The chitosan-chondroitin sulfate composite itself possesses anti-rheumatic and anti-inflammatory properties, which further synergizes with the drug to achieve improved therapeutic outcomes. In a mouse model, this nanoparticle formulation demonstrated higher drug accumulation and superior therapeutic effects compared to free LEF and LEF-NPs. Similarly, Shafiq et al combined core-shell Pluronic F127 with chitosan to load methotrexate, resulting in the design of an oral nanoparticle formulation that enhanced pharmacokinetic properties, including an 8-fold increase in half-life and a 3.48-fold increase in mean residence time. The oral nanoparticle sustained drug release for two days at pH 6.8, significantly improving the bioavailability of MTX.130 In animal models, this nanoparticle demonstrated effective targeting and therapeutic efficacy.
Hyaluronic acid (HA) is a glycosaminoglycan widely present in connective and epithelial tissues, primarily composed of D-glucuronic acid and N-acetylglucosamine.131 HA is renowned for its excellent biocompatibility and plays a crucial role in cell communication. Receptors or molecules that interact with HA include Neurocan, CD44, hyaluronic acid-mediated motility receptor (RHAMM), glial HA-binding protein (GHAP), LYVE-1 (lymphatic endothelial HA receptor 1), Versican, Aggrecan, and TSG6 (TNF-stimulated gene 6), with CD44 being a well-known surface receptor of M1 macrophages. Consequently, the use of hyaluronic acid in combination with drugs for precision therapy in RA holds considerable promise.
Wang et al focused on the acidity and reactive oxygen species (ROS) reactivity of HA to crosslink HA with β-cyclodextrin (β-CD), forming nanoparticles (HCPC) loaded with dexamethasone (DEX) for the treatment of RA.132 This nanoparticle formulation, utilizing the properties of HA, enhances both the targeting and therapeutic effects of the drug (Figure 6). In vitro and in vivo experiments demonstrated that these nanoparticles exhibited excellent targeting to M1 macrophages, significantly increasing drug retention in RA joints and further inhibiting inflammation, thereby demonstrating substantial therapeutic effects. After loading fluorescence probe IR780, the labeled nanoparticles showed excellent inflamed joint targeting ability. After 6h administration, the fluorescence intensity of IR780 labeled HCPC was approximately seven times higher than that of free IR780. To evaluate the safety, the major organ tissues including (heart, live, spleen, lung, kidney and brain) were sliced and analyzed. No substantive lesions or inflammation was observed. Zhou et al combined HA with solid lipid nanoparticles (SL) to deliver the glucocorticoid prednisolone (PD), resulting in nanoparticles (HA-SLN/PD) for RA treatment.133 In the collagen-induced arthritis (CIA) mouse model, the HA-modified HA-SLN/PD formulation exhibited more pronounced therapeutic effects compared to free drugs or SLN nanoparticles.
 |
Figure 6 (A) Schematic diagram illustrating the synthesis of HCPC NPs and its applications in enhanced treatment of RA via passive targeting, M1 macrophage targeting, and responsive drug release. (B) Micro-CT analysis of the hind paws. (a) The representative visual 3D images of ankle joints (yellow arrows indicate osteophyte and hyperplasia, green arrows show bone destruction, red arrows show trabecular osteopenia, blue arrows indicate subchondral bone sclerosis). Quantitative results of bone histomorphometry parameters including (b) BV/TV, (c) Tb.Th, (d) Tb.N, and (e) BMD. Data are mean ± standard deviation, n = 6, *P < 0.05, **P < 0.01 vs AA group. Referenced from Ref.132 |
Certain HA derivatives, while retaining the original physicochemical properties of HA, possess additional functionalities. For example, Shang et al designed a hyaluronic acid derivative (PAM-HA) that can adsorb to subchondral bone, remain in inflamed joints for extended periods, and respond to the clearance of ROS.134 They employed PAM-HA as a biomimetic carrier to encapsulate sinomenine (Sin), resulting in a new nanoparticle formulation (PAM-HA@Sin NP). In vivo experiments showed that this nanoparticle effectively cleared ROS, reduced the levels of pro-inflammatory factors, and enhanced joint lubrication.
Nucleic Acids
Nucleic acids, including deoxyribonucleic acid (DNA) and ribonucleic acid (RNA), are biological macromolecules composed of nucleotide monomers. Specifically, nanomaterials based on DNA, have gained increasing attention in the medical field due to their exceptional flexibility, affinity, programmability, and low biological toxicity.135
Currently, DNA nanomaterials are categorized into two primary types: DNA nanostructures (DNA-Nss) and DNA-functionalized nanoparticles (DNA-NPs).136 In 1996, Mirkin et al pioneered the development of DNA-functionalized nanoparticles (DNA-NPs), where DNA is highly oriented and densely functionalized with covalent bonds, forming a nanoparticle structure. Compared to free DNA, DNA-NPs exhibit significantly increased resistance to nuclease degradation, thereby enhancing the stability of nucleic acids in the body.135,137
Huang et al designed a complex of DNA and cationic polyethyleneimine nanoparticles (DNPs), which were found to stimulate the upregulation of indoleamine 2,3-dioxygenase (IDO) activity, further inducing DCs and regulatory T cells (Tregs) to suppress immune overexpression in inflamed joints, demonstrating strong immune-modulatory properties.138 Zhang et al found that interleukin-10 (IL-10) can improve the inflammatory environment of RA through macrophage metabolic reprogramming.139 They developed a multifunctional nanoparticle (bPEI-SS-PEG-T/NLS/DNA NPs) composed of NLS (a peptide with a nuclear localization signal) and IL-10 plasmid DNA (pDNA) in combination with polyethyleneimine-dithiobis (succinimidyl propionate) (bPEI-SS-PEG-T). This formulation responds to intracellular glutathione levels and promotes macrophage uptake, thereby increasing drug accumulation (Figure 7). In vivo models showed that this nanoparticle demonstrated excellent IL-10 transfection efficiency, and inhibitory effects on RA arthritis. This DNA based nanoparticles realized remarkable inflammatory joint accumulation ability. Compared with unmodified nanoparticles, the modified nanoparticles exhibited about 10 times higher accumulation in rat joints after 2.5h administration. Beyond efficacy, compared with normal group, the body weight, renal and hepatic functions were not damaged.
 |
Figure 7 Schematic illustration of the macrophage-hitchhiking IL-10 pDNA delivery system against RA. (A) Illustration of formation of bPEI-SS-PEGT/NLS/DNA NPs. (B) Illustration of RA microenvironment. (C) Schematic of CIA rats and treatment with intra-peritoneal injection. After the intraperitoneal injection, nanoparticles could effectively accumulate at inflammatory sites of RA due to the macrophage migration to the inflammatory joints. (D) The intracellular fate of bPEI-SS-PEG-T/NLS/DNA NPs. (E) The mechanism of action of bPEI-SS-PEG-T/NLS/DNA NPs in vivo. NPC: Nuclear pore complex. (F) In vivo imaging of unmodified NPs and bPEI-SS-PEG-T/NLS/ DNA NPs in CIA rats and bPEI-SS-PEG-T/NLS/DNA NPs in normal rats at different times. The inflammatory sites are circled in red. (G) Average radiant efficiency of paws. The values reported are mean ± standard deviation (n = 3), **P < 0.01, ***P < 0.001 and ****P < 0.0001 compared with arthritic rats with modified NPs. Referenced from Ref.139 |
DNA nanostructures (DNA-Nss) can be engineered with single-stranded DNA arranged based on regional or origami structures. These structures are widely used in anti-inflammatory and anti-tumor therapies due to their ability to exhibit arbitrary sizes, shapes, and surface features.135 Wang et al designed a self-assembled DNA tetrahedron (TDs) coupled with NF-κB decoy oligonucleotides (dODNs) and VCAM-1-targeting peptides (P), forming a novel nanoformulation (TD-P-dODN).140 In an adjuvant-induced animal model, this nanoformulation demonstrated superior drug retention and therapeutic effects compared to free dODN (Figure 8). To target pro-inflammatory molecules such as TNF-α and NF-κB, Liu et al designed a nanoparticle (siRNA@M(Cage-dODN)) (siMCO) coated with macrophage cytoplasmic membranes, carrying anti-NF-κB decoy oligonucleotides (dODN) and anti-TNF-α siRNA for RA treatment. Both in vitro and in vivo results showed that siMCO significantly increased drug accumulation and reduced the expression of inflammatory factors.141
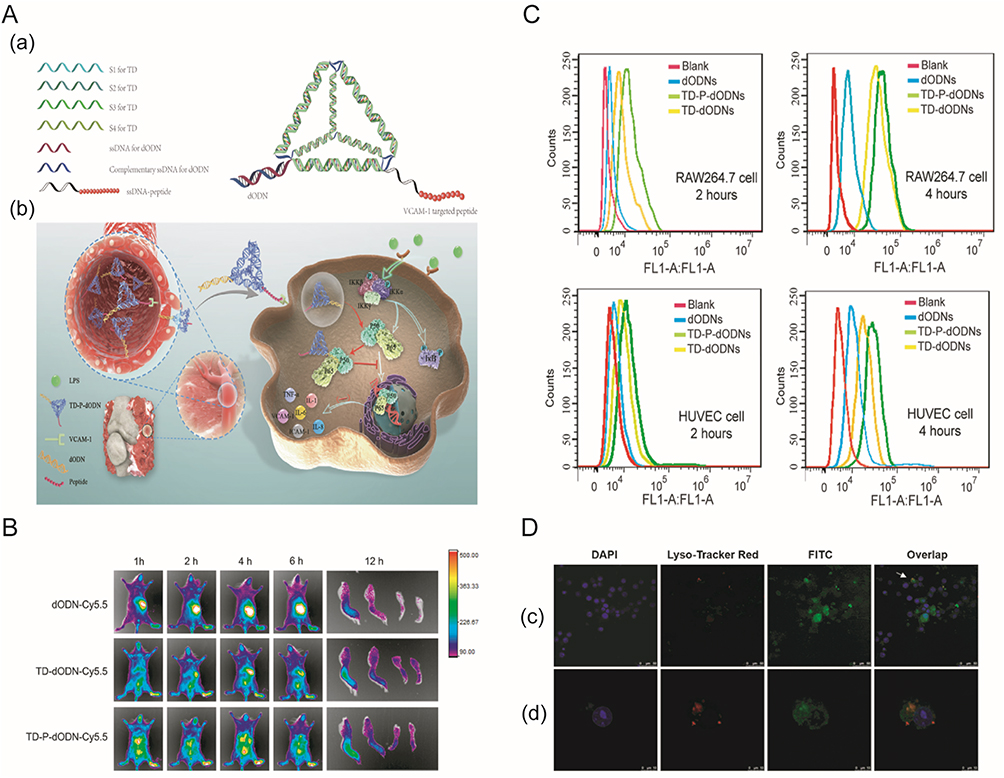 |
Figure 8 (A) Schematic illustration of the engineered DNA nanodrugs. An illustration of the self-assembly of a TD-based DNA nanostructure (a). In vivo delivery of TD-P-dODN and the intracellular interaction of TD-P dODN with its targeted protein (P50/P65) (b). (B) In vivo distribution images of dODNs-Cy5.5, TD-dODN-Cy5.5 and TD-P-dODN-Cy5.5. (C) Flow cytometry analysis of the intracellular uptake of the listed treatment after 2 h and 4 h incubation. (D) Cell images that represent the cellular uptake of FITC-labeled TD-P-dODNs by RAW264.7 cells (c). (Scale bar = 50 μm) (d) Enlarged cell images of the arrow pointed cells in (D) (c). (Scale bar = 10 μm) (after 4 h coculture). Referenced from Ref.140 |
Overall, DNA-based nanomaterials offer significant advantages in terms of infectivity, immunogenicity, and cytotoxicity. However, challenges remain. For example, the negative charge of DNA molecules, which leads to electrostatic repulsion and impairs the passive transport of nanoparticles in tissues.142 Consequently, the modification of DNA nanomaterials may enhance their clinical applications.
Biomimetic Membranes/Extracellular Vesicles Coating Strategies
The utilization of biomimetic technology offers new possibilities for drug in RA therapy. By modifying cell membranes or incorporating biomimetic proteins (Figure 2), drug delivery systems can acquire biomimetic camouflage. Thereby, achieve reduced clearance rate, improved biocompatibility, and prolonged drug retention in the body. Given that the initiation of RA involves the infiltration of multiple immune cells, biomimetic strategies based on immune cell membranes, associated proteins, or cell-derived exosomes are naturally more advantageous than traditional modification methods. The biomimetic approach is expected to enhance the targeting and therapeutic efficacy of nanodrugs, thereby opening new avenues for RA treatment.
Biomimetic Membranes Biofilm Coating
Leukocytes, including lymphocytes, monocytes, neutrophils, eosinophils, and basophils, are collectively known as white blood cells. These cells play a central role in the infiltration of the synovium in RA joints, contributing to the initiation of inflammation. Therefore, nanoparticles that can bind to leukocytes are particularly effective in penetrating the affected joints in RA and targeting drug delivery to inflammatory sites. Recently, cell membrane-coated nanoparticles have emerged as a novel technology to enhance the efficacy of traditional drugs.143 Nanoparticles incorporating natural cell membranes inherit the physicochemical properties and membrane functions of the original cell membrane, including membrane-anchored proteins, antigens, and immune characteristics.144,145
Macrophage Membranes
Macrophages are among the most predominant cells involved in the infiltration of RA joints. Targeted delivery of anti-inflammatory drugs using macrophage membranes for the treatment of inflammation has shown significant progress.146 Flavia et al developed a hybrid nanosystem (TCPSi@KG-1) modified with KG-1 macrophage membranes on porous silicon (Psi).147 In vitro experiments revealed that undecylenic acid-modified hydrophobic thermally carbonized porous silicon (UnTHCPSi) nanoparticles were highly stable in physiological solutions and biological fluids. Cell experiments indicated that the membrane-coated UnTHCPSi nanoparticles expressed CD80 more closely to the control group than unmodified nanoparticles, suggesting that the macrophage membrane-modified nanosystem significantly reduced immunogenicity (Figure 9).
 |
Figure 9 (A) Schematic representation of the nanoplatforms. PSi nanoparticles are processed together with cytoplasmic membranes isolated from KG-1 macrophages. Image created with Servier Medical Art. (B) TEM images of a) APTS-TCPSi@KG-1, b) UnTHCPSi@KG-1, and c) TCPSi@KG-1. (C) Stability of UnTCHPSi, UnTHCPSi@KG-1, TCPSi, and TCPSi@KG-1 systems in d, e) human fresh frozen plasma and f, g) simulated synovial fluid, up to 2 h. The results are presented as mean ± standard deviation (n = 3). The results were analyzed with two-way ANOVA, followed by Bonferroni’s posttest and the levels of significance were set at the probabilities of *P < 0.05, **P < 0.01, and ***P < 0.001. Referenced from Ref.147 |
Song et al combined metal-phenolic networks (MPN) with epigallocatechin gallate (EGCG), exploiting their properties of scavenging reactive oxygen species (ROS) and inhibiting inflammatory cytokines.148 They further incorporated EGCG with cerium (IV) ions coated with macrophage membranes to prepare MPN nanoparticles (M-EC). In vitro experiments demonstrated that the EC concentration-dependent clearance of reactive nitrogen species (RNS) by the M-EC nanoparticles increased, with nearly complete clearance of ROS at 100μg/mL MTT assays revealed that, at concentrations up to 100 μg/mL, EC had minimal cytotoxicity on RAW264.7 cells, HUVEC cells, and chondrocytes. But EC exhibited significant cytotoxicity to inflammatory macrophages, suggesting good safety and protective effects against harmful ROS. Fluorescence labeling studies revealed that the coated nanoparticles had a faster targeting speed and could evade capture by normal macrophages compared to unmodified nanoparticles. Both nanoparticle formulations were efficiently internalized by inflammatory macrophages, indicating effective targeting. In a mouse model, the M-EC group showed reduced expression of IL-1β, promoted the repolarization of M1 macrophages, and effectively inhibited joint inflammation compared to the PBS and EC groups. Other studies suggest that macrophage membranes may competitively bind with inflammatory cytokines to further inhibit osteoclast generation.149 Ruixiang Li et al used macrophage-derived microvesicle (MMV)-coated nanoparticles (MNP) realized about 15 times higher accumulation in arthritic paws after 12h injection, compared with free small-molecular group. No obvious pathological changes were observed in the liver, spleen and kidney for both free tacrolimus treated group and correspond membrane coated nanoparticle treated group.
Neutrophil Membranes
Neutrophils play a crucial role in the pathogenesis of RA. As the most abundant type of white blood cells, neutrophils accumulate in inflamed joints, where they can significantly contribute to the progression of RA inflammation.150,151 The LFA-1 receptor on neutrophil membranes can specifically bind to activated chondrocytes and the overexpressed intercellular adhesion molecule 1 (ICAM-1) on human umbilical vein endothelial cells (HUVECs), providing an important theoretical basis for the biomimetic application of neutrophil membranes.152
Zhang et al employed neutrophil membrane-modified poly(lactic-co-glycolic acid) (PLGA) polymer nanoparticles in a mouse femoral allograft model. Fluorescence labeling revealed the accumulation of neutrophil membrane nanoparticles in the distal region of the femoral head and surrounding tissues, indicating that the nanoparticles exhibited excellent targeting and chondrocyte penetration.153 In a collagen-induced arthritis (CIA) model, histopathological analysis revealed no significant degeneration of chondrocytes in the cartilage in the neutrophil membrane nanoparticle-treated group, The number of fibroblast-like synoviocytes (FLS) positive for staining was significantly reduced. Serum tests demonstrated that the membrane-modified nanoparticles significantly decreased the expression of inflammatory factors such as IL-1β and TNF-α. These results highlight the potential of neutrophil membranes in addressing RA inflammation.
Yu et al coated polyethylene glycolated l-arginine nanoparticles (LANPs) with neutrophil membranes and loaded them with a reactive oxygen species (ROS) inducer (Ru), creating a nitric oxide (NO) biomimetic delivery system (NM-LANPs@Ru).154 Studies in animal models showed that this delivery system demonstrated good targeting and biocompatibility for the inflammatory sites. Compared with the control group, NM-LANPs@Ru exhibited enhanced therapeutic characteristics.
Platelet Membranes
Platelets, small cytoplasmic fragments responsible for hemostasis, are considered a crucial component of the innate immune system due to their ability to interact with bacteria, viruses, and complement proteins.154–156 Increasing evidence suggests that the dysregulation of platelet activation plays a significant role in the pathogenesis of RA.157,158 Upon activation by ligands such as collagen and thrombin, platelets release various inflammatory mediators, including prothrombotic agents, cytokines, chemokines, and growth factors.159 These secretions exacerbate the inflammatory response in RA, promote neovascularization, and facilitate immune cell aggregation, thereby intensifying disease progression.160
It is evident that platelets express immune-related receptors, thereby linking interactions between immune cells. For instance, CD44 is a surface glycoprotein widely present on many RA inflammatory cells. In 1993, Stone et al reported that P-selectin, overexpressed on the membrane of activated platelets, can specifically recognize CD44.161 Additionally, glycoprotein VI (GPVI) on platelet membranes can specifically interact with type IV collagen.162 This suggests that platelets have a high affinity for immune cells involved in RA pathogenesis.
To validate this hypothesis, He et al loaded PLGA nanoparticles (PNPs) with FK506, a potent immunosuppressive agent used to treat RA in patients unresponsive to methotrexate, and modified them with platelet membranes.163 The in vitro experiments showed that human RA synovial cells (MH7A), cultured in a collagen IV-rich medium, exhibited significant binding with DiD-labeled PNPs under confocal microscopy. In vivo experiments revealed that the fluorescence signal intensity of PNPs in inflamed joints was markedly higher than in other organs, as shown by in vivo imaging and localization analysis. Immunofluorescence staining confirmed the co-localization of DiD-labeled PNPs with CD44 and collagen IV, further validating the targeting of platelet membranes. The group treated with drug-loaded PNPs exhibited the lowest arthritis index (AI), indicating excellent therapeutic effects. Li et al utilized platelet membrane-coated mesoporous silica nanoparticles (MSNs) encapsulating vascular disruption agents (VDAs) to create a novel type of nanoparticle. In animal models, this formulation demonstrated significant vascular disruption and effective anti-angiogenesis, yielding promising therapeutic outcomes.162
Vesicular Membranes
Extracellular vesicles (EVs) are lipid-enveloped vesicles secreted by various cells, including macrophages. And they are involved in cell-to-cell communication. EVs can be categorized into apoptotic bodies, microvesicles (MVs), and exosomes, with exosomes being particularly important in the pathogenesis of RA.164–166 Exosomes are capable of regulating osteoclast formation and modulating immune responses, thereby reducing inflammation associated with RA. Mesenchymal stem cells (MSCs), associated with EVs, can regenerate and replace damaged cells within tissues, offering protective effects, while also exerting immunomodulatory actions through the release of EVs and paracrine factors, such as cytokines and growth factors.167,168
EVs are not only non-cytotoxic and non-immunogenic but also exhibit excellent biocompatibility. When combined with nanoparticles, they can evade phagocytosis by the mononuclear phagocyte system (MPS).169 The attachment of molecular therapeutic agents, such as nucleic acids, proteins, and lipids, to EVs for the treatment of RA represents a promising therapeutic strategy.
Macrophage-derived EVs: Li et al employed cytochalasin B (CB) to develop microvesicles (MMVs) derived from macrophages. By coating these MMVs with PLGA nanoparticles and loading them with tacrolimus, they created a novel nanoparticle formulation.149 In a mouse model, this formulation demonstrated significantly better targeting of RA inflammatory sites compared to drug-loaded uncoated nanoparticles and red blood cell membrane-coated nanoparticles (RNPs). Yan et al modified macrophage-derived exosomes with a folate-polyethylene glycol-cholesterol complex and loaded them with dexamethasone phosphate (Dex), resulting in a biomimetic nanoformulation (FPC-Exo/Dex).170 The in vitro experiments showed that this formulation significantly reduced levels of TNF-α and IL-1β, indicating favorable drug accumulation and anti-inflammatory properties. In another study, Li et al used M2 macrophage-derived exosomes (M2 Exo) to encapsulate anti-inflammatory cytokine IL-10 pDNA and the anti-inflammatory drug betamethasone phosphate (BSP), creating a biomimetic drug delivery system. This system was able to downregulate the secretion of pro-inflammatory cytokines (IL-1β, TNF-α) and upregulate IL-10 expression.171 In a mouse model, this drug delivery system increased drug accumulation in the joints and promoted the repolarization of macrophages from M1 to M2, exhibiting promising therapeutic effects.
Platelet-derived EVs: Tessandier et al demonstrated that during RA inflammation, vascular leakage facilitates the entry of platelet-derived extracellular vesicles (EVs) into the lymphatic system, whereas EVs from other cells, such as red blood cells and white blood cells, encounter greater difficulty passing through the lymphatic system. This suggests that platelet-derived EVs possess stronger targeting and tissue penetration capabilities.172 Based on this finding, Ma et al developed a targeted delivery system for RA-inflamed joints by utilizing platelet-derived EVs as biomimetic carriers (PEVs) loaded with the anti-inflammatory drug berberine (BBR) to modulate the joint pathological environment.173 In RA animal models, combined with fluorescence imaging, it was observed that after intravenous injection, PEVs accumulated significantly in the inflamed joints and paws of RA mice, indicating efficient targeting and drug accumulation. The PEVs realized about 9 times higher accumulation in the inflamed joints, compared with small molecular group. There was no obvious pathological change in the heart, liver, spleen, lung and kidney in both free berberine group and berberine loaded PEVs group. Micro-computed tomography (micro-CT) analysis of bone tissue revealed that, compared to free drugs, BBR-loaded PEVs enhanced bone density and significantly inhibited bone erosion. Furthermore, analysis of immune cells in the mice demonstrated that a marked reduction in the expression levels of pro-inflammatory receptors (CD68 and CD80), pro-inflammatory cytokines (TNF-α and IL-6), and MMPs (MMP-3), further confirming the therapeutic efficacy of BBR-loaded PEVs.
Other Cell-Derived EVs: DCs-derived extracellular vesicles (EVs) have also been extensively investigated. For example, Kim et al demonstrated in animal models that FasL gene-modified bone marrow-derived DC exosomes (Exo/FasL) can suppress joint inflammation and autoimmune diseases.174 Additionally, Wu et al explored the impact of exosomes secreted by granulocyte-myeloid derived suppressor cells (G-MDSCs) on humoral immunity in mice, confirming that these exosomes promote the generation of IL-10+ Breg cells, thereby alleviating inflammation.175 Song et al found that exosomes secreted by endothelial cells (EC-exosomes) exhibited greater bone-targeting properties compared to exosomes from other cell sources, with the endogenous cargo miR-155 in EC-exosomes inhibiting osteoclast activity.176
Engineered Cell Membranes
The concept of “engineered cell membranes” has emerged to enhance the functionality or characteristics of cell membranes. Engineered cell membranes refer to the process of modifying or replicating cell membranes using physical, chemical, or genetic engineering techniques. This technology typically involves the incorporation of specific molecules, proteins, or drugs onto cell membranes to fulfill specific requirements. Based on the modification approach, engineered cell membranes can be classified into physically engineered membranes, chemically engineered membranes, and genetically engineered membranes.
Physically Engineered Membranes
Physically engineered membrane technology is an advanced biotechnological method that integrates specific macromolecules or fragments of cell membranes onto target cell membranes using physical techniques, thereby endowing the cell membranes with new functions or properties. For instance, modification of red blood cell membranes can significantly extend the circulation time of drug delivery systems in the body, up to 120 days. Furthermore, by modifying the cell surface marker CD47, these systems can effectively evade clearance by the immune system.177 This section discusses the application of physically engineered cell membranes from two perspectives: the modification of target membranes with specific cell membranes, and the modification of cell membranes with macromolecules.
Dehaini et al validated the feasibility of modifying platelet membranes with red blood cell membranes. They conducted membrane fusion studies, labeling platelet membranes with two different fluorescent dyes (DOPE-RhB and C6-NBD), and mixing them with red blood cell membranes in varying proportions at high temperatures. Fluorescence recovery through Förster Resonance Energy Transfer (FRET) confirmed the fusion of the two membranes. Subsequently, poly(lactic-co-glycolic acid) (PLGA) nanoparticles were coated with the mixed membrane to obtain [RBC-P]NPs.178 Protein characteristic analysis revealed that the fused membrane contained specific protein markers from both original cell types, and functional analysis demonstrated that [RBC-P]NPs exhibited a low uptake rate by THP-1 macrophages, similar to RBCNPs and PNPs. In animal models, the [RBC-P]NPs showed superior circulation capabilities compared to RBCNPs and PNPs.
Fibroblast-like synoviocyte membranes exhibit significantly stronger targeting capabilities than platelet membranes, due to the variety of markers present on fibroblast-like synoviocytes. Therefore, Yu et al fused red blood cell membranes with fibroblast-like synoviocyte membranes (RFM) to modify Prussian blue nanoparticles (PB NPs) loaded with Schisanlactone E (SE), and chemically modified hyaluronic acid was grafted onto the RPS NPs through DSPE-PEG2000-NH2 to create a multifunctional nanodelivery system (HA@RFM@PB@SE, HRPS NPs)179(Figure 10). Dialysis bag experiments revealed that at pH 5.0, the release rate of SE was high, which is suitable for the acidic pathological environment of RA. In photothermal tests, HRPS NPs demonstrated efficient photothermal conversion and stability, as the solution temperature increased under different laser powers. The targeting efficiency of the nanoparticles was assessed using fluorescence labeling under laser irradiation, showing a higher uptake rate by fibroblast-like synoviocytes in the HRPS NPs group. In RA mouse models, histopathological scoring indicated a significant reduction in joint inflammation and synovial hyperplasia in the HRPS NPs group, which was further confirmed by Micro-CT and MRI imaging techniques. ELISA assays revealed significant reductions in inflammatory factors such as TNF-α, IL-1β, and IL-6. Safety assessments showed that the nanodelivery system did not cause significant organ damage or functional disorders, demonstrating good biosafety.
 |
Figure 10 Overall Synthesis Process of HRPS NPs and Their Application for RA Therapy: (A) Synthesis process of HRPS NPs; (B) Application of HRPS NPs for RA therapy; (C) Related mechanisms of HRPS NPs against RA. The red arrow depicts signaling from the NF-κB pathway/IL-23/IL-17 axis targeting RAFLS cells. The green arrow indicates the action of TNF-α, IL-6, and IL-1β—produced upon activation of the NF-κB pathway/IL-23/IL-17 axis—on osteoclasts. (D) Fluorescence images of cellular uptake and lysosomal escape of PS NPs, RPS NPs, and HRPS NPs in RAFLS cells; the yellow fluorescence in the merged image showed the colocalization of NPs and lysosomes. Scale bar = 10 μm. (E) Mean integrated density of PS NPs, RPS NPs, and HRPS NPs in RAFLS cells. (F) Fluorescence images of cellular uptake of HRPS NPs and HA solution + HRPS NPs in RAFLS cells. Scale bar = 10 μm. (G) Mean integrated density of HRPS NPs and HA solution + HRPS NPs in RAFLS cells. (H) TEM images of PB NPs. Scale bar = 100 nm. (I) SEM images of PB NPs. Scale bar = 100 nm. (J) TEM images of PB@SE NPs (PS NPs) showed an approximate irregular square. Scale bar = 100 nm. (K) TEM images of PS NPs and the RFM was covering the PS NPs with larger magnification. Scale bar = 50 nm. The red curve and arrow indicate outer film structure of the RFM@PB@SE NPs (RPS NPs). (L) Cryo-TEM image of PS NPs. Scale bar = 80 nm. (M) Cryo-TEM image of PS NPs with larger magnification. Scale bar = 50 nm. The red curve corresponds to the presence of an RFM coating on the PS NPs. (N) Immunoelectron microscopy (IEM) images of PS NPs and RFM of 10 nm gold-labeled particles coated with CD44 and CD 47 antibodies. Scale bar = 100 nm. The red arrows highlight the specific expression of CD44 and CD47 on the external side of the Representative Membrane (RFM). (O) TEM images of HA@ RFM@PB@SE NPs (HRPS NPs) showed an approximate spherical shape. Scale bar = 100 nm. The red curve corresponds to the nanospherical architecture of the HA@RFM@PB@SE (HRPS NPs) core-shell nanoparticles. All studies were performed at least three times. Data presented as mean ± standard deviation. Statistical analysis was performed using independent samples t test. **P ≤ 0.01. Referenced from Ref.179 |
In addition to red blood cells, Zhao et al designed hybrid nanovesicles (HNVs) by the extrusion method, coated with M1 macrophage membranes and derived from M2 macrophage vesicular membranes. These nanovesicles were subsequently coated with black phosphorus (BP) nanosheets to obtain HNV@BP.180 In macrophage activation experiments, the HNV@BP was co-cultured with RAW macrophages (RAW264.7) and M1 macrophages (induced by lipopolysaccharide and interferon-γ). Flow cytometry was used to detect the expression of CD206 (a marker of M2 macrophages) and CD86 (a marker of M1 macrophages). The results showed that HNV@BP increased the proportion of M2 macrophage subtypes (M2a), and ELISA tests confirmed that HNV@BP effectively bound to IL-6 and TNF-α, reducing their expression levels. Cytotoxicity assays demonstrated good biocompatibility, and under near-infrared (NIR) light stimulation, HNV@BP significantly inhibited the survival of L929 cells (a mouse fibroblast cell line). In animal models, H&E staining and Safranin-O/fast green staining showed that HNV@BP, in combination with NIR irradiation, significantly reduced damage to articular cartilage and the proliferation of synovial tissue. These nanovesicles inherited the cytokine receptors of the two macrophage subtypes, and the combination of BP nanosheets and NIR irradiation led to significant suppression of RA inflammation.
Certain large molecular substances, such as liposomes, are also widely utilized in physical engineering membrane technology. This method involves the anchoring of glycosylphosphatidylinositol (GPI) via lipid interactions on the cell membrane, enabling GPI-anchored fusion proteins to be physically modified and attached to the membrane.181 This technique is based on the structure of lipids and the fluidity of the membrane.182 Hao et al employed liposomes to modify red blood cell membrane vesicles, resulting in hybrid vesicles loaded with the immunosuppressive agent mycophenolic acid (MPA) for targeting DCs in the treatment of systemic lupus erythematosus (SLE).183 In animal models, these biomimetic vesicles demonstrated good targeting and therapeutic effects, with no significant drug toxicity observed. Furthermore, Molinaro et al reported that liposomes enriched with white blood cell membrane proteins could be used to modify cell vesicles.184 The modified vesicles not only retained the biological characteristics of the original white blood cells but also overcame vascular barriers in inflammatory environments. Molinaro et al further found that these liposome-modified vesicles containing white blood cell membrane proteins could evade capture by the mononuclear phagocyte system and specifically target the delivery of dexamethasone to inflammatory sites. This discovery broadens the potential applications of physically engineered membranes.
Chemically Engineered Membranes
Chemical engineering strategies typically involve the use of amine and thiol groups on membrane proteins, as well as hydroxyl groups on polysaccharides, as active sites for various covalent conjugations.185 For example, polymers, nucleic acids, and proteins can be directly conjugated to cell membranes through amide bond formation reactions mediated by multifunctional 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide (EDC), which are faster and more stable than conventional EDC-mediated amidation reactions.186–188 Using NHS-PEG2-maleimide technology, Zhou et al successfully modified red blood cell membranes with human recombinant hyaluronidase PH20 (rHuPH20) and coated nanoparticles.189 Compared to unmodified red blood cell membrane nanoparticles (RBCM-NPs), the coated nanoparticles exhibited significantly extended circulation times. Moreover, to clear free circulating DNA (cfDNA) in the inflammatory environment of RA, Zhe Wang et al manufactured an exosome-based system called MEX+cP including anti-inflammatory (M2) macrophage-derived exosomes (MEX), positively charged oligolysine and PEG fragment (cP). In biodistribution assay, the in vivo fluorescence images revealed that the accumulation of fluorescence labeled MEX+cP showed obvious inflamed paw accumulation. After 24h injection, the accumulation of MEX+cP is over 3 times higher than that of correspond control group with blockade antibody. Compared with phosphate buffer saline treated group, there was no significant change in lactic dehydrogenase, aspartate aminotransferase, alanine aminotransferase and alkaline phosphatase levels. No obvious damage was observed in major organs.190 The PEG modification prolonged circulation time and enabled the directional accumulation of C─C motif chemokine ligands around inflamed joints. Upon MMP degradation, the positively charged oligolysine could clear cfDNA, and the exosome promoted macrophage polarization. In animal models, the chemically modified exosomes exhibited promising therapeutic effects and potential for clinical application. Additionally, Lee et al modified dendritic cell-derived exosomes (TKDex) with reactive oxygen species-sensitive polyethylene glycol linkers. Studies indicated that TKDex could be internalized by mature DCs, significantly reducing the expression of CD40 and IL-6, as well as the production of tumor necrosis factor TNF-α, thereby demonstrating its potential in targeted therapy.191
Genetically Engineered Membranes
Genetic engineering allows for the selective introduction of desired proteins or peptides into cell membranes via transfection or via non-viral or viral vectors, a strategy widely applied in the treatment of RA.185,192 Deng et al utilized platelet microvesicle membranes (PMM) modified with a Golgi apparatus-targeting peptide to encapsulate poly(D, L-lactic-co-glycolic acid) (PLGA) nanoparticles loaded with atorvastatin (Atra). This formulation aimed to suppress the secretion of pathological substances by synovial fibroblasts (FLS) in RA joints, thus promoting RA inflammation.193 The system successfully targeted the delivery of atorvastatin to the Golgi apparatus of FLS, leading to disruption of the Golgi apparatus and inhibition of pathogenic protein expression. In both in vitro and in vivo models, this nanoformulation significantly increased drug accumulation in the Golgi apparatus of FLS, reduced the expression of pathogenic proteins and mediators, and alleviated joint inflammation in RA. Considering the specific role of CCDC25 protein and NETs, Wang et al developed a genetically engineered nanovesicle expressing the CCDC25 protein, which was then modified with a liposome and loaded with DNase I (a DNA enzyme) to form a new nanoformulation.194 The hybrid vesicles successfully targeted NETs and inhibited the aggregation of neutrophils. Additional studies have indicated that mesenchymal stem cell (MSC) membranes can also be modified using genetic engineering techniques.195 MSCs inherently possess the ability to regulate inflammatory processes, repair damaged cells within joints, and migrate to injured tissues. Genetically modified MSC membranes not only retain the targeting capabilities of the original cells but also eliminate concerns related to genetic mutations, ensuring their safety.
Conclusion
This article summarizes the traditional treatment strategies, associated targets, and recent advances in biomimetic therapeutic strategies for RA. Traditional therapeutic drugs failed to achieve the expected goals in RA joint therapy due to significant adverse reactions and a lack of distribution specificity. Some biomimetic nanocarriers for cancer therapy have been studied in clinical trials including albumin-based nanoparticle (NCT00110695), dendritic cell-derived exosomes (NCT01159288) and nanoscale exosome-based delivery system (NCT04592484). But in RA treatment, the challenges still remain between preclinical study and actual clinical study for biomimetic technology. However, with the continuous development of biomimetic nanotechnology and deeper research into RA pathological targets, the clinical treatment prospects are promising. Typically, after biomimetic strategy modification, the tissue-specific accumulation can be greatly improved to 35 times. The toxicity can be minimized. In summary, compared with chemically synthesized nanomaterials, biomimetic nanomaterials exhibit superior targeting, prolonged blood circulation, immune tolerance, and reduced biological toxicity.
Prospects
Despite the significant advancements in bionic nanotechnology observed in current research, substantial challenges remain before true clinical translation can be achieved. The author identifies several key factors contributing to this gap:
Firstly, most animal models currently employed for bionic nanoparticles do not fully replicate the actual pathological environment of RA in humans. This disparity creates uncertainty regarding the long-term stability and safety of the nanomaterials. Although bionic nanoparticles have shown promising therapeutic effects in vitro and in animal models, their application in humans is subject to various considerations, including immune system responses, potential long-term toxicity, and interactions with existing therapeutic drugs.
Secondly, within RA lesions, numerous targets, such as white blood cells and synovial cells, exist. Compared to single-target therapies, a multi-level targeting strategy which can interact with two or more targets simultaneously might be more suited for the pathological environment of RA. For example, the interleukin-6, Tyrosine kinase 2 and CD20 surface maker on B lymphocytes all play vital roles in the progression in RA. Intervening each of these targets alone is not sufficient. Therefore, designing drug delivery systems which can inhibit all three targets might be beneficial. Currently, computer-assisted drug candidates have been developed to simultaneously blocked the three mentioned pathways.196 As for tissue perspective, multi-level targeting allows drug delivery systems to bind specifically to multiple target cells at once, significantly enhancing the targeting efficiency of drug delivery systems (DDS) and possibly yielding synergistic effects that improve therapeutic efficacy. At the cellular level, nanoparticles employing multi-level targeting strategies can achieve precise targeting of cells, cell membranes, and organelles, reducing damage to normal cells and minimizing side effects, thereby enhancing treatment safety. Overall, multi-level targeting can intervene in the complex microenvironment of RA at multiple levels, significantly improving drug efficacy.
Thirdly, although the treatment strategy for RA primarily focuses on preventing chronic joint erosion caused by the immune system, ie, targeting drug delivery, optimizing pharmacokinetics, enhancing anti-inflammatory effects, and minimizing systemic side effects, it does not imply that researchers should overlook the mechanical friction between joints. The ability of nanoparticles to withstand wear due to joint movement is essential for maintaining their physical stability. However, to date, there is no literature providing evidence regarding the performance of bionic nanomaterials in friction tests. Similarly, nanoparticles circulating within the bloodstream may be subjected to shear forces from blood flow, potentially leading to surface or structural wear. These factors introduce uncertainties that could affect the stability of bionic nanoparticles in the human body.
Lastly, RA is not only an intra-articular disease. It also involves a range of extra-articular manifestations or complications, including rheumatoid nodules, vasculitis, and disorders affecting the cardiovascular (CV), pulmonary, neurological, gastrointestinal, renal, and hematological systems.197 As such, RA might be regarded as a syndrome. Nevertheless, many current bionic therapeutic strategies remain limited anti-inflammatory and immune cell targeting ability within the joint, which is clearly insufficient to address the broader scope of the disease as a systemic syndrome.
These challenges drive requirements for advanced bionic nanoparticles, particularly pathology-responsive designs targeting RA’s unique microenvironment. These intelligent designs would enhance therapeutic efficacy for rheumatic diseases and enable integrated approaches—including photothermal (PTT) and photodynamic therapy (PDT)—for RA combination treatment. As bionic technologies evolve, they will enable more precise and safer RA therapeutic strategies.
Acknowledgments
This study was supported by the National Natural Science Foundation of China (grant number 81603045, China), the Sichuan Provincial Natural Science Foundation of China (grant number 2023NSFSC1683, China), the Scientific Research Project of Sichuan Provincial Administration of Traditional Chinese Medicine (grant number 2024MS034, China), the Engineering Research Center for Pharmaceuticals and Equipments of Sichuan Province Fund (grant number ERCPEF2303, China), the Open Fund of Development and Regeneration Key Laboratory of Sichuan Province (grant number 23LHPDZYB17 and 24LHFYSZ1-46, China), the Regional Preparation Center Joint Research Project (grant number 2024GJQY01, China) and the High Level Talent Cultivation Project of Chengdu Medical College (grant number KYPY24-03, China).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Dolati S, Sadreddini S, Rostamzadeh D, Ahmadi M, Jadidi-Niaragh F, Yousefi M. Utilization of nanoparticle technology in rheumatoid arthritis treatment. Biomed Pharmacother. 2016;80:30–41. doi:10.1016/j.biopha.2016.03.004
2. van Vollenhoven R. Treat-to-target in rheumatoid arthritis — are we there yet? Nat Rev Rheumatol. 2019;15(3):180–186. doi:10.1038/s41584-019-0170-5
3. McInnes IB, Schett G. Pathogenetic insights from the treatment of rheumatoid arthritis. Lancet. 2017;389(10086):2328–2337. doi:10.1016/S0140-6736(17)31472-1
4. Weyand CM, Goronzy JJ. The immunology of rheumatoid arthritis. Nat Immunol. 2021;22(1):10–18. doi:10.1038/s41590-020-00816-x
5. Xiang Y, Chen L, Liu C, Yi X, Li L, Huang Y. Redirecting chemotherapeutics to the endoplasmic reticulum increases tumor immunogenicity and potentiates Anti-PD-L1 therapy. Small. 2022;18(6):2104591. doi:10.1002/smll.202104591
6. Huang J, Fu X, Chen X, Li Z, Huang Y, Liang C. Promising therapeutic targets for treatment of rheumatoid arthritis. Review. 2021;12. doi:10.3389/fimmu.2021.686155.
7. Finckh A, Gilbert B, Hodkinson B, et al. Global epidemiology of rheumatoid arthritis. Nat Rev Rheumatol. 2022;18(10):591–602. doi:10.1038/s41584-022-00827-y
8. Ben Mrid R, Bouchmaa N, Ainani H, El Fatimy R, Malka G, Mazini L. Anti-rheumatoid drugs advancements: new insights into the molecular treatment of rheumatoid arthritis. Biomed Pharmacother. 2022;151:113126. doi:10.1016/j.biopha.2022.113126
9. Conn DL. The story behind the use of glucocorticoids in the treatment of rheumatoid arthritis. Seminars Arthritis Rheumatism. 2021;51(1):15–19. doi:10.1016/j.semarthrit.2020.09.016
10. Guo Q, Wang Y, Xu D, Nossent J, Pavlos NJ, Xu J. Rheumatoid arthritis: pathological mechanisms and modern pharmacologic therapies. Bone Res. 2018;6(1):15. doi:10.1038/s41413-018-0016-9
11. Wu G, Su T, Zhou P, et al. Engineering M2 macrophage-derived exosomes modulate activated T cell cuproptosis to promote immune tolerance in rheumatoid arthritis. Biomaterials. 2025;315:122943. doi:10.1016/j.biomaterials.2024.122943
12. Wu G, Zhu J, Huang R, et al. Photodriven PtPdCo-TiO2 heterostructure modified with hyaluronic acid and folic acid enhances antioxidative stress through efficient hydrogen/oxygen delivery and thermal effects in rheumatoid arthritis therapy. Int J Biol Macromol. 2025;307:142014. doi:10.1016/j.ijbiomac.2025.142014
13. Ahmed T, Alam KT. Biomimetic nanoparticle based targeted mrna vaccine delivery as a novel therapy for glioblastoma multiforme. AAPS PharmSciTech. 2025;26(3):68. doi:10.1208/s12249-025-03065-z
14. Jia N, Gao Y, Li M, et al. Metabolic reprogramming of proinflammatory macrophages by target delivered roburic acid effectively ameliorates rheumatoid arthritis symptoms. Signal Transduction Targeted Ther. 2023;8(1):280. doi:10.1038/s41392-023-01499-0
15. Xie H, Huang X, Li B, et al. Biomimetic nanoplatform for targeted rheumatoid arthritis therapy: modulating macrophage niches through self-sustaining positive feedback-driven drug release mechanisms. Adv Sci. 2025;12(15):2416265. doi:10.1002/advs.202416265
16. Lamers-Karnebeek FBG, Jacobs JWG, Radstake TRDJ, van Riel PLCM, Jansen TL. Adalimumab drug and antidrug antibody levels do not predict flare risk after stopping adalimumab in RA patients with low disease activity. Rheumatology. 2019;58(3):427–431. doi:10.1093/rheumatology/key292
17. Scott LJ. Tocilizumab: a Review in Rheumatoid Arthritis. Drugs. 2017;77(17):1865–1879. doi:10.1007/s40265-017-0829-7
18. Maity S, Misra A, Wairkar S. Novel injectable carrier based corticosteroid therapy for treatment of rheumatoid arthritis and osteoarthritis. J Drug Delivery Sci Technol. 2021;61:102309. doi:10.1016/j.jddst.2020.102309
19. Sarzi-Puttini P, Ceribelli A, Marotto D, Batticciotto A, Atzeni F. Systemic rheumatic diseases: from biological agents to small molecules. Autoimmunity Rev. 2019;18(6):583–592. doi:10.1016/j.autrev.2018.12.009
20. Thakur S, Riyaz B, Patil A, Kaur A, Kapoor B, Mishra V. Novel drug delivery systems for NSAIDs in management of rheumatoid arthritis: an overview. Biomed Pharmacother. 2018;106:1011–1023. doi:10.1016/j.biopha.2018.07.027
21. van Vollenhoven RF. Treatment of rheumatoid arthritis: state of the art 2009. Nat Rev Rheumatol. 2009;5(10):531–541. doi:10.1038/nrrheum.2009.182
22. Salliot C, Finckh A, Katchamart W, et al. Indirect comparisons of the efficacy of biological antirheumatic agents in rheumatoid arthritis in patients with an inadequate response to conventional disease-modifying antirheumatic drugs or to an anti-tumour necrosis factor agent: a meta-analysis. Annals Rheumatic Dis. 2011;70(2):266–271. doi:10.1136/ard.2010.132134
23. O’Neil LJ, Kaplan MJ. Neutrophils in rheumatoid arthritis: breaking immune tolerance and fueling disease. Trends Mol Med. 2019;25(3):215–227. doi:10.1016/j.molmed.2018.12.008
24. Eisenbarth SC. Dendritic cell subsets in T cell programming: location dictates function. Nat Rev Immunol. 2019;19(2):89–103. doi:10.1038/s41577-018-0088-1
25. Scherer HU, Häupl T, Burmester GR. The etiology of rheumatoid arthritis. J Autoimmunity. 2020;110:102400. doi:10.1016/j.jaut.2019.102400
26. Grillet B, Pereira RVS, Van Damme J, Abu El-Asrar A, Proost P, Opdenakker G. Matrix metalloproteinases in arthritis: towards precision medicine. Nat Rev Rheumatol. 2023;19(6):363–377. doi:10.1038/s41584-023-00966-w
27. DiDomenico CD, Lintz M, Bonassar LJ. Molecular transport in articular cartilage — what have we learned from the past 50 years? Nat Rev Rheumatol. 2018;14(7):393–403. doi:10.1038/s41584-018-0033-5
28. Rothenfluh DA, Bermudez H, O’Neil CP, Hubbell JA. Biofunctional polymer nanoparticles for intra-articular targeting and retention in cartilage. Nat Mat. 2008;7(3):248–254. doi:10.1038/nmat2116
29. Bajpayee AG, Grodzinsky AJ. Cartilage-targeting drug delivery: can electrostatic interactions help? Nat Rev Rheumatol. 2017;13(3):183–193. doi:10.1038/nrrheum.2016.210
30. Yang Y, Zhang H. Intra-articular injection of nanomaterials for the treatment of osteoarthritis: from lubrication function restoration to cell and gene therapy. Adv Funct Mat. 2024;34(30):2401547. doi:10.1002/adfm.202401547
31. Jimi E, Huang F, Nakatomi C. NF-κB signaling regulates physiological and pathological chondrogenesis. Int J Mol Sci. 2019;20(24). doi:10.3390/ijms20246275
32. Leblond A, Allanore Y, Avouac J. Targeting synovial neoangiogenesis in rheumatoid arthritis. Autoimmunity Rev. 2017;16(6):594–601. doi:10.1016/j.autrev.2017.04.005
33. Konisti S, Kiriakidis S, Paleolog EM. Angiogenesis in Rheumatoid Arthritis. In: Dulak J, Józkowicz A, Łoboda A, editors. Angiogenesis and Vascularisation: Cellular and Molecular Mechanisms in Health and Diseases. Springer Vienna; 2013:339–365.
34. Nygaard G, Firestein GS. Restoring synovial homeostasis in rheumatoid arthritis by targeting fibroblast-like synoviocytes. Nat Rev Rheumatol. 2020;16(6):316–333. doi:10.1038/s41584-020-0413-5
35. Pérez-Gutiérrez L, Ferrara N. Biology and therapeutic targeting of vascular endothelial growth factor A. Nat Rev Mol Cell Biol. 2023;24(11):816–834. doi:10.1038/s41580-023-00631-w
36. Asif Amin M, Fox DA, Ruth JH. Synovial cellular and molecular markers in rheumatoid arthritis. Seminars Immunopathol. 2017;39(4):385–393. doi:10.1007/s00281-017-0631-3
37. Degboé Y, Poupot R, Poupot M. Repolarization of unbalanced macrophages: unmet medical need in chronic inflammation and cancer. Int J Mol Sci. 2022;23(3). doi:10.3390/ijms23031496
38. Udalova IA, Mantovani A, Feldmann M. Macrophage heterogeneity in the context of rheumatoid arthritis. Nat Rev Rheumatol. 2016;12(8):472–485. doi:10.1038/nrrheum.2016.91
39. Ross EA, Devitt A, Johnson JR. Macrophages: the good, the bad, and the gluttony. Review. 2021;12. doi:10.3389/fimmu.2021.708186.
40. Mosser DM, Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. 2008;8(12):958–969. doi:10.1038/nri2448
41. Tardito S, Martinelli G, Soldano S, et al. Macrophage M1/M2 polarization and rheumatoid arthritis: a systematic review. Autoimmun Rev. 2019;18(11):102397. doi:10.1016/j.autrev.2019.102397
42. McInnes IB, Schett G. Cytokines in the pathogenesis of rheumatoid arthritis. Nat Rev Immunol. 2007;7(6):429–442. doi:10.1038/nri2094
43. Mills CD, Kincaid K, Alt JM, Heilman MJ, Hill AM. Pillars article: m-1/M-2 macrophages and the Th1/Th2 paradigm. J Immunol. 2017;199(7):2194–2201. doi:10.4049/jimmunol.1701141
44. Yang Y, Guo L, Wang Z, et al. Targeted silver nanoparticles for rheumatoid arthritis therapy via macrophage apoptosis and Re-polarization. Biomaterials. 2021;264:120390. doi:10.1016/j.biomaterials.2020.120390
45. Chandrupatla DMSH, Molthoff CFM, Lammertsma AA, van der Laken CJ, Jansen G. The folate receptor β as a macrophage-mediated imaging and therapeutic target in rheumatoid arthritis. Drug Delivery Transl Res. 2019;9(1):366–378. doi:10.1007/s13346-018-0589-2
46. Cummings RD. The mannose receptor ligands and the macrophage glycome. Current Opinion Struct Biol. 2022;75:102394. doi:10.1016/j.sbi.2022.102394
47. Taban Q, Mumtaz PT, Masoodi KZ, Haq E, Ahmad SM. Scavenger receptors in host defense: from functional aspects to mode of action. Cell Commun Signaling. 2022;20(1):2. doi:10.1186/s12964-021-00812-0
48. Martín-Sabroso C, Torres-Suárez AI, Alonso-González M, Fernández-Carballido A, Fraguas-Sánchez AI. Active targeted nanoformulations via folate receptors: state of the art and future perspectives. Pharmaceutics. 2022;14(1). doi:10.3390/pharmaceutics14010014
49. Han Y, Pang X, Pi G. Biomimetic and bioinspired intervention strategies for the treatment of rheumatoid arthritis. Adv Funct Mat. 2021;31(38):2104640. doi:10.1002/adfm.202104640
50. Watkins AR, Reesink HL. Lubricin in experimental and naturally occurring osteoarthritis: a systematic review. Osteoarthritis Cartilage. 2020;28(10):1303–1315. doi:10.1016/j.joca.2020.05.009
51. Bartok B, Firestein GS. Fibroblast-like synoviocytes: key effector cells in rheumatoid arthritis. Immunolog Rev. 2010;233(1):233–255. doi:10.1111/j.0105-2896.2009.00859.x
52. Bottini N, Firestein GS. Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat Rev Rheumatol. 2013;9(1):24–33. doi:10.1038/nrrheum.2012.190
53. Kato M, Ospelt C, Gay RE, Gay S, Klein K. Dual role of autophagy in stress-induced cell death in rheumatoid arthritis synovial fibroblasts. Arthritis Rheumatol. 2014;66(1):40–48. doi:10.1002/art.38190
54. Noss EH, Brenner MB. The role and therapeutic implications of fibroblast-like synoviocytes in inflammation and cartilage erosion in rheumatoid arthritis. Immunolog Rev. 2008;223(1):252–270. doi:10.1111/j.1600-065X.2008.00648.x
55. Hua P, Liang R, Yang S, Tu Y, Chen M. Microneedle-assisted dual delivery of PUMA gene and celastrol for synergistic therapy of rheumatoid arthritis through restoring synovial homeostasis. Bioact Mat. 2024;36:83–95. doi:10.1016/j.bioactmat.2024.02.030
56. Amariuta T, Luo Y, Knevel R, Okada Y, Raychaudhuri S. Advances in genetics toward identifying pathogenic cell states of rheumatoid arthritis. Immunolog Rev. 2020;294(1):188–204. doi:10.1111/imr.12827
57. Chen Z, Bozec A, Ramming A, Schett G. Anti-inflammatory and immune-regulatory cytokines in rheumatoid arthritis. Nat Rev Rheumatol. 2019;15(1):9–17. doi:10.1038/s41584-018-0109-2
58. Ponchel F, Burska AN, Hunt L, et al. T-cell subset abnormalities predict progression along the inflammatory arthritis disease continuum: implications for management. Scient Rep. 2020;10(1):3669. doi:10.1038/s41598-020-60314-w
59. Li Y, Goronzy JJ, Weyand CM. DNA damage, metabolism and aging in pro-inflammatory T cells: rheumatoid arthritis as a model system. Exp Gerontol. 2018;105:118–127. doi:10.1016/j.exger.2017.10.027
60. Weyand CM, Shen Y, Goronzy JJ. Redox-sensitive signaling in inflammatory T cells and in autoimmune disease. Free Radical Biol Med. 2018;125:36–43. doi:10.1016/j.freeradbiomed.2018.03.004
61. Weyand CM, Goronzy JJ. Immunometabolism in early and late stages of rheumatoid arthritis. Nat Rev Rheumatol. 2017;13(5):291–301. doi:10.1038/nrrheum.2017.49
62. Li Y, Shen Y, Jin K, et al. The DNA repair nuclease MRE11A functions as a mitochondrial protector and prevents T cell pyroptosis and tissue inflammation. Cell Metabol. 2019;30(3):477–492.e6. doi:10.1016/j.cmet.2019.06.016
63. Semerano L, Minichiello E, Bessis N, Boissier M-C. Novel immunotherapeutic avenues for rheumatoid arthritis. Trends Mol Med. 2016;22(3):214–229. doi:10.1016/j.molmed.2016.01.005
64. van Hamburg JP, Tas SW. Molecular mechanisms underpinning T helper 17 cell heterogeneity and functions in rheumatoid arthritis. J Autoimmunity. 2018;87:69–81. doi:10.1016/j.jaut.2017.12.006
65. Volkov M, van Schie KA, van der Woude D. Autoantibodies and B cells: the ABC of rheumatoid arthritis pathophysiology. Immunolog Rev. 2020;294(1):148–163. doi:10.1111/imr.12829
66. Bugatti S, Vitolo B, Caporali R, Montecucco C, Manzo A. B cells in rheumatoid arthritis: from pathogenic players to disease biomarkers. BioMed Res Int. 2014;2014(1):681678. doi:10.1155/2014/681678
67. Scherer HU, van der Woude D, Toes REM. From risk to chronicity: evolution of autoreactive B cell and antibody responses in rheumatoid arthritis. Nat Rev Rheumatol. 2022;18(7):371–383. doi:10.1038/s41584-022-00786-4
68. Sun W, Meednu N, Rosenberg A, et al. B cells inhibit bone formation in rheumatoid arthritis by suppressing osteoblast differentiation. Nat Commun. 2018;9(1):5127. doi:10.1038/s41467-018-07626-8
69. Fillatreau S, Manfroi B, Dörner T. Toll-like receptor signalling in B cells during systemic lupus erythematosus. Nat Rev Rheumatol. 2021;17(2):98–108. doi:10.1038/s41584-020-00544-4
70. Laurent L, Anquetil F, Clavel C, et al. IgM rheumatoid factor amplifies the inflammatory response of macrophages induced by the rheumatoid arthritis-specific immune complexes containing anticitrullinated protein antibodies. Annals Rheumatic Dis. 2015;74(7):1425. doi:10.1136/annrheumdis-2013-204543
71. Harre U, Lang SC, Pfeifle R, et al. Glycosylation of immunoglobulin G determines osteoclast differentiation and bone loss. Nat Commun. 2015;6(1):6651. doi:10.1038/ncomms7651
72. Yanaba K, Bouaziz J-D, Haas KM, Poe JC, Fujimoto M, Tedder TF. A regulatory B cell subset with a unique CD1dhiCD5+ phenotype controls t cell-dependent inflammatory responses. Immunity. 2008;28(5):639–650. doi:10.1016/j.immuni.2008.03.017
73. Thiolat A, Semerano L, Pers YM, et al. Interleukin-6 receptor blockade enhances CD39+ regulatory T cell development in rheumatoid arthritis and in experimental arthritis. Arthritis Rheumatol. 2014;66(2):273–283. doi:10.1002/art.38246
74. Wong PKK, Quinn JMW, Sims NA, van Nieuwenhuijze A, Campbell IK, Wicks IP. Interleukin-6 modulates production of T lymphocyte–derived cytokines in antigen-induced arthritis and drives inflammation-induced osteoclastogenesis. Arthritis Rheumatism. 2006;54(1):158–168. doi:10.1002/art.21537
75. Yeo L, Toellner K-M, Salmon M, et al. Cytokine mRNA profiling identifies B cells as a major source of RANKL in rheumatoid arthritis. Annals Rheumatic Dis. 2011;70(11):2022–2028. doi:10.1136/ard.2011.153312
76. Chen Z, Cui Y, Yao Y, et al. Heparan sulfate regulates IL-21 bioavailability and signal strength that control germinal center B cell selection and differentiation. Sci Immunol. 2023;8(80):eadd1728. doi:10.1126/sciimmunol.add1728
77. Espéli M, Smith KGC, Clatworthy MR. FcγRIIB and autoimmunity. Immunolog Rev. 2016;269(1):194–211. doi:10.1111/imr.12368
78. Barlev AN, Malkiel S, Kurata-Sato I, Dorjée AL, Suurmond J, Diamond B. FcγRIIB regulates autoantibody responses by limiting marginal zone B cell activation. J Clin Investig. 2022;132(17). doi:10.1172/JCI157250
79. Cascão R, Rosário HS, Souto-Carneiro MM, Fonseca JE. Neutrophils in rheumatoid arthritis: more than simple final effectors. Autoimmun Rev. 2010;9(8):531–535. doi:10.1016/j.autrev.2009.12.013
80. Wright HL, Moots RJ, Bucknall RC, Edwards SW. Neutrophil function in inflammation and inflammatory diseases. Rheumatology. 2010;49(9):1618–1631. doi:10.1093/rheumatology/keq045
81. Wright HL, Lyon M, Chapman EA, Moots RJ, Edwards SW. Rheumatoid arthritis synovial fluid neutrophils drive inflammation through production of chemokines, reactive oxygen species, and neutrophil extracellular traps. Orig Res. 2020;11. doi:10.3389/fimmu.2020.584116.
82. Pham CTN. Neutrophil serine proteases: specific regulators of inflammation. Nat Rev Immunol. 2006;6(7):541–550. doi:10.1038/nri1841
83. Hirano T. IL-6 in inflammation, autoimmunity and cancer. Int Immunol. 2021;33(3):127–148. doi:10.1093/intimm/dxaa078
84. Kaplan MJ. Neutrophils in the pathogenesis and manifestations of SLE. Nat Rev Rheumatol. 2011;7(12):691–699. doi:10.1038/nrrheum.2011.132
85. Humphrey MB, Nakamura MC. A comprehensive review of immunoreceptor regulation of osteoclasts. Clin Rev Allergy Immunol. 2016;51(1):48–58. doi:10.1007/s12016-015-8521-8
86. Camps M, Rückle T, Ji H, et al. Blockade of PI3Kγ suppresses joint inflammation and damage in mouse models of rheumatoid arthritis. Nat Med. 2005;11(9):936–943. doi:10.1038/nm1284
87. Plater-Zyberk C, Joosten LAB, Helsen MMA, Koenders MI, Baeuerle PA, van den Berg WB. Combined blockade of granulocyte-macrophage colony stimulating factor and interleukin 17 pathways potently suppresses chronic destructive arthritis in a tumour necrosis factor α-independent mouse model. Annals Rheumatic Dis. 2009;68(5):721. doi:10.1136/ard.2007.085431
88. Nair JR, Edwards SW, Moots RJ. Mavrilimumab, a human monoclonal GM-CSF receptor-α antibody for the management of rheumatoid arthritis: a novel approach to therapy. Expert Opinion Biol Ther. 2012;12(12):1661–1668. doi:10.1517/14712598.2012.732062
89. Takayanagi H. Osteoimmunology: shared mechanisms and crosstalk between the immune and bone systems. Nat Rev Immunol. 2007;7(4):292–304. doi:10.1038/nri2062
90. Jacome-Galarza CE, Percin GI, Muller JT, et al. Developmental origin, functional maintenance and genetic rescue of osteoclasts. Nature. 2019;568(7753):541–545. doi:10.1038/s41586-019-1105-7
91. Axmann R, Böhm C, Krönke G, Zwerina J, Smolen J, Schett G. Inhibition of interleukin-6 receptor directly blocks osteoclast formation in vitro and in vivo. Arthritis Rheumatism. 2009;60(9):2747–2756. doi:10.1002/art.24781
92. Lam J, Takeshita S, Barker JE, Kanagawa O, Ross FP, Teitelbaum SL. TNF-α induces osteoclastogenesis by direct stimulation of macrophages exposed to permissive levels of RANK ligand. J Clin Investig. 2000;106(12):1481–1488. doi:10.1172/JCI11176
93. Yokota K. Osteoclast differentiation in rheumatoid arthritis. Immunolog Med. 2024;47(1):6–11. doi:10.1080/25785826.2023.2220931
94. Yokota K, Sato K, Miyazaki T, et al. Characterization and function of tumor necrosis factor and interleukin-6–induced osteoclasts in rheumatoid arthritis. Arthritis Rheumatol. 2021;73(7):1145–1154. doi:10.1002/art.41666
95. Iwamoto N, Kawakami A. The monocyte-to-osteoclast transition in rheumatoid arthritis: recent findings. Review. 2022;13. doi:10.3389/fimmu.2022.998554.
96. Anita C, Munira M, Mural Q, Shaily L. Topical nanocarriers for management of Rheumatoid Arthritis: a review. Biomed Pharmacother. 2021;141:111880. doi:10.1016/j.biopha.2021.111880
97. Wu J, Xing L, Zheng Y, et al. Disease-specific protein Corona formed in pathological intestine enhances the oral absorption of nanoparticles. Acta Pharmaceutica Sinica B. 2023;13(9):3876–3891. doi:10.1016/j.apsb.2023.02.012
98. Chen Z, Xing L, Fan Q, et al. Drug-bearing supramolecular filament hydrogels as anti-inflammatory agents. Research paper. Theranostics. 2017;7(7):2003–2014. doi:10.7150/thno.19404
99. Deng X, Liu X, Zhang Y, et al. Changes of serum IgG glycosylation patterns in rheumatoid arthritis. Clin Proteomics. 2023;20(1):7. doi:10.1186/s12014-023-09395-z
100. Ma S, Xue F, Yang L, et al. Pharmacokinetic and pharmacodynamic study of folic acid-modified chitosan–stearic acid nanomicelles loaded with tetrandrine for rheumatoid arthritis. Pharmaceutics. 2025;17(2). doi:10.3390/pharmaceutics17020169
101. Kratz EM, Kacperczyk M, Kokot I, et al. Glycosylation pattern of serum clusterin in psoriatic arthritis and rheumatoid arthritis—the search for new diagnostic glycomarkers. Int J Mol Sci. 2024;25(23). doi:10.3390/ijms252313060
102. Park SB, Yang Y, Bang SI, Kim TS, Cho D. AESIS-1, a rheumatoid arthritis therapeutic peptide, accelerates wound healing by promoting fibroblast migration in a CXCR2-dependent manner. Int J Mol Sci. 2024;25(7). doi:10.3390/ijms25073937
103. Yang L, Sha Y, Wei Y, et al. Mannose-mediated nanodelivery of methotrexate to macrophages augments rheumatoid arthritis therapy. Biomat Sci. 2023;11(6):2211–2220. doi:10.1039/D2BM02072F
104. Zhou F, Li M, Chen M, et al. Redox homeostasis strategy for inflammatory macrophage reprogramming in rheumatoid arthritis based on ceria oxide nanozyme-complexed biopolymeric micelles. ACS Nano. 2023;17(5):4358–4372. doi:10.1021/acsnano.2c09127
105. Guo H, Ma J, Zhang Y, et al. Delivery of AntagomiR-7 through polymer nanoparticles for assisting B cell to alleviate systemic lupus erythematosus. Orig Res. 2023;11. doi:10.3389/fbioe.2023.1180302
106. Li Z, Han S, Cui G, et al. Oral liposomes encapsulating ginsenoside compound K for rheumatoid arthritis therapy. Int J Pharmaceut. 2023;643:123247. doi:10.1016/j.ijpharm.2023.123247
107. Mirchandani Y, Patravale VB, Brijesh S. Hyaluronic acid-coated solid lipid nanoparticles enhance antirheumatic activity and reduce toxicity of methotrexate. Nanomedicine. 2022;17(16):1099–1114. doi:10.2217/nnm-2022-0009
108. Sultana F, Neog MK, Rasool M. Targeted delivery of morin, a dietary bioflavanol encapsulated mannosylated liposomes to the macrophages of adjuvant-induced arthritis rats inhibits inflammatory immune response and osteoclastogenesis. Eur J Pharmaceut Biopharmaceut. 2017;115:229–242. doi:10.1016/j.ejpb.2017.03.009
109. Li C, Liu R, Song Y, et al. Hyaluronic acid hydrogels hybridized with au-triptolide nanoparticles for intraarticular targeted multi-therapy of rheumatoid arthritis. Orig Res. 2022;13. doi:10.3389/fphar.2022.849101
110. Rehman A, John P, Bhatti A. Biogenic selenium nanoparticles: potential solution to oxidative stress mediated inflammation in rheumatoid arthritis and associated complications. Nanomaterials. 2021;11(8). doi:10.3390/nano11082005
111. Deng C, Zhang Q, He P, et al. Targeted apoptosis of macrophages and osteoclasts in arthritic joints is effective against advanced inflammatory arthritis. Nat Commun. 2021;12(1):2174. doi:10.1038/s41467-021-22454-z
112. Zhang Y, Sun T, Jiang C. Biomacromolecules as carriers in drug delivery and tissue engineering. Acta Pharmaceutica Sinica B. 2018;8(1):34–50. doi:10.1016/j.apsb.2017.11.005
113. Hu H, Quintana J, Weissleder R, Parangi S, Miller M. Deciphering albumin-directed drug delivery by imaging. Adv Drug Delivery Rev. 2022;185:114237. doi:10.1016/j.addr.2022.114237
114. X-x F, M-z X, Leung EL-H, Jun C, Yuan Z, Liu L. ROS-responsive berberine polymeric micelles effectively suppressed the inflammation of rheumatoid arthritis by targeting mitochondria. Nano-Micro Lett. 2020;12(1):76. doi:10.1007/s40820-020-0410-x
115. Liu L, Hu F, Wang H, et al. Secreted protein acidic and rich in cysteine mediated biomimetic delivery of methotrexate by albumin-based nanomedicines for rheumatoid arthritis therapy. ACS Nano. 2019;13(5):5036–5048. doi:10.1021/acsnano.9b01710
116. Gong T, Zhang P, Deng C, Xiao Y, Gong T, Zhang Z. An effective and safe treatment strategy for rheumatoid arthritis based on human serum albumin and Kolliphor® HS 15. Nanomedicine. 2019;14(16):2169–2187. doi:10.2217/nnm-2019-0110
117. Yan F, Li H, Zhong Z, et al. Co-delivery of prednisolone and curcumin in human serum albumin nanoparticles for effective treatment of rheumatoid arthritis. Int J Nanomed. 2019;14(null):9113–9125. doi:10.2147/IJN.S219413
118. Zheng X, Yang H, Zhang Z, et al. pH-responsive size-adjustable liposomes induce apoptosis of fibroblasts and macrophages for rheumatoid arthritis treatment. Acta Biomaterialia. 2024;179:256–271. doi:10.1016/j.actbio.2024.03.006
119. Busatto S, Walker SA, Grayson W, et al. Lipoprotein-based drug delivery. Adv Drug Delivery Rev. 2020;159:377–390. doi:10.1016/j.addr.2020.08.003
120. Alolga RN, Opoku-Damoah Y, Alagpulinsa DA, et al. Metabolomic and transcriptomic analyses of the anti-rheumatoid arthritis potential of xylopic acid in a bioinspired lipoprotein nanoformulation. Biomaterials. 2021;268:120482. doi:10.1016/j.biomaterials.2020.120482
121. Pozzi FS, Maranhão RC, Guedes LK, et al. Plasma kinetics of an LDL-like non-protein nanoemulsion and transfer of lipids to high-density lipoprotein (HDL) in patients with rheumatoid arthritis. J Clin Lipidol. 2015;9(1):72–80. doi:10.1016/j.jacl.2014.10.004
122. García-Gómez MC, Padró T, Muñoz-García N, et al. Dysfunctional antioxidant capacity of high-density lipoprotein in rheumatoid arthritis. Eur J Clin Investig. 2023;53(8):e13999. doi:10.1111/eci.13999
123. VanEvery H, Yang W, Su J, et al. Low-density lipoprotein cholesterol and the risk of rheumatoid arthritis: a prospective study in a Chinese cohort. Nutrients. 2022;14(6). doi:10.3390/nu14061240
124. Meng Q, Zhong S, Gao Y, Cui X. Advances in polysaccharide-based nano/microcapsules for biomedical applications: a review. Int J Biol Macromol. 2022;220:878–891. doi:10.1016/j.ijbiomac.2022.08.129
125. Meng Q, Zhong S, Xu L, et al. Review on design strategies and considerations of polysaccharide-based smart drug delivery systems for cancer therapy. Carbohydrate Polymers. 2022;279:119013. doi:10.1016/j.carbpol.2021.119013
126. Qindeel M, Ahmed N, Khan GM, Rehman A. Ligand decorated chitosan as an advanced nanocarrier for targeted delivery: a critical review. Nanomedicine. 2019;14(12):1623–1642. doi:10.2217/nnm-2018-0490
127. Zhang X, Chen F, Ni J. A novel method to prepare magnetite chitosan microspheres conjugated with methotrexate (MTX) for the controlled release of MTX as a magnetic targeting drug delivery system. Drug Delivery. 2009;16(5):280–288. doi:10.1080/10717540902989555
128. Shakeran Z, Keyhanfar M, Varshosaz J, Sutherland DS. Biodegradable nanocarriers based on chitosan-modified mesoporous silica nanoparticles for delivery of methotrexate for application in breast cancer treatment. Mat Sci Eng. 2021;118:111526. doi:10.1016/j.msec.2020.111526
129. Jabbari N, Eftekhari Z, Roodbari NH, Parivar K. Evaluation of encapsulated eugenol by Chitosan nanoparticles on the aggressive model of rheumatoid arthritis. Int Immunopharmacol. 2020;85:106554. doi:10.1016/j.intimp.2020.106554
130. Shafiq A, Madni A, Khan S, et al. Core-shell Pluronic F127/chitosan based nanoparticles for effective delivery of methotrexate in the management of rheumatoid arthritis. Int J Biol Macromol. 2022;213:465–477. doi:10.1016/j.ijbiomac.2022.05.192
131. Vasvani S, Kulkarni P, Rawtani D. Hyaluronic acid: a review on its biology, aspects of drug delivery, route of administrations and a special emphasis on its approved marketed products and recent clinical studies. Int J Biol Macromol. 2020;151:1012–1029. doi:10.1016/j.ijbiomac.2019.11.066
132. Wang P, Zhang Y, Lei H, et al. Hyaluronic acid-based M1 macrophage targeting and environmental responsive drug releasing nanoparticle for enhanced treatment of rheumatoid arthritis. Carbohydrate Polymers. 2023;316:121018. doi:10.1016/j.carbpol.2023.121018
133. Zhou M, Hou J, Zhong Z, Hao N, Lin Y, Li C. Targeted delivery of hyaluronic acid-coated solid lipid nanoparticles for rheumatoid arthritis therapy. Drug Delivery. 2018;25(1):716–722. doi:10.1080/10717544.2018.1447050
134. Shang W, Sun Q, Zhang C, et al. Drug in therapeutic polymer: sinomenine-loaded oxidation-responsive polymeric nanoparticles for rheumatoid arthritis treatment. ACS Appl Mat Interfaces. 2023;15(40):47552–47565. doi:10.1021/acsami.3c10562
135. Zhao Y, Shi L, Kuang H, Xu C. DNA-driven Nanoparticle Assemblies for Biosensing and Bioimaging. In: Fan C, Ke Y, editors. DNA Nanotechnology: From Structure to Functionality. Springer International Publishing; 2020:267–299.
136. Nicolson F, Ali A, Kircher MF, Pal S. DNA nanostructures and DNA-functionalized nanoparticles for cancer theranostics. Adv Sci. 2020;7(23):2001669. doi:10.1002/advs.202001669
137. Seferos DS, Prigodich AE, Giljohann DA, Patel PC, Mirkin CA. Polyvalent DNA nanoparticle conjugates stabilize nucleic acids. Nano Lett. 2009;9(1):308–311. doi:10.1021/nl802958f
138. Huang L, Lemos HP, Li L, et al. Engineering DNA nanoparticles as immunomodulatory reagents that activate regulatory T cells. J Immunol. 2012;188(10):4913–4920. doi:10.4049/jimmunol.1103668
139. Zhang X, Liu Y, Liu W, et al. Macrophage-hitchhiking interleukin-10 plasmid DNA delivery system modulates rheumatoid arthritis microenvironment via the re-polarization of macrophages. Nano Today. 2024;54:102068. doi:10.1016/j.nantod.2023.102068
140. Wang Z, Chu X, Li N, Fu L, Gu H, Zhang N. Engineered DNA nanodrugs alleviate inflammation in inflammatory arthritis. Int J Pharmaceut. 2020;577:119047. doi:10.1016/j.ijpharm.2020.119047
141. Liu F, Dong Z, Li M, et al. A macrophage plasma membrane-coated and DNA structured nanomedicine targets to alleviate rheumatoid arthritis via dual inhibition to TNF-α and NF-κB. Int J Pharmaceut. 2023;642:123188. doi:10.1016/j.ijpharm.2023.123188
142. Fernandes JC, Wang H, Jreyssaty C, et al. Bone-protective effects of nonviral gene therapy with folate–Chitosan DNA nanoparticle containing interleukin-1 receptor antagonist gene in rats with adjuvant-induced arthritis. Mol Ther. 2008;16(7):1243–1251. doi:10.1038/mt.2008.99
143. You DG, Lim GT, Kwon S, et al. Metabolically engineered stem cell–derived exosomes to regulate macrophage heterogeneity in rheumatoid arthritis. Sci Adv. 2021;7(23):eabe0083. doi:10.1126/sciadv.abe0083
144. Gao W, Hu C-MJ, Fang RH, Luk BT, Su J, Zhang L. Surface functionalization of gold nanoparticles with red blood cell membranes. Adv Mat. 2013;25(26):3549–3553. doi:10.1002/adma.201300638
145. Fang RH, Hu C-MJ, Zhang L. Nanoparticles disguised as red blood cells to evade the immune system. Expert Opinion Biol Ther. 2012;12(4):385–389. doi:10.1517/14712598.2012.661710
146. Sun T, Kwong CHT, Gao C, et al. Amelioration of ulcerative colitis via inflammatory regulation by macrophage-biomimetic nanomedicine. Research Paper. Theranostics. 2020;10(22):10106–10119. doi:10.7150/thno.48448
147. Fontana F, Albertini S, Correia A, et al. Bioengineered porous silicon nanoparticles@macrophages cell membrane as composite platforms for rheumatoid arthritis. Adv Funct Mat. 2018;28(22):1801355. doi:10.1002/adfm.201801355
148. Song X, Zheng Z, Ouyang S, et al. Biomimetic epigallocatechin gallate–cerium assemblies for the treatment of rheumatoid arthritis. ACS Appl Mat Interfaces. 2023;15(28):33239–33249. doi:10.1021/acsami.3c02768
149. Li R, He Y, Zhu Y, et al. Route to rheumatoid arthritis by macrophage-derived microvesicle-coated nanoparticles. Nano Lett. 2019;19(1):124–134. doi:10.1021/acs.nanolett.8b03439
150. Chu D, Dong X, Shi X, Zhang C, Wang Z. Neutrophil-based drug delivery systems. Adv Mat. 2018;30(22):1706245. doi:10.1002/adma.201706245
151. Rao L, Bu -L-L, Xu J-H, et al. Red blood cell membrane as a biomimetic nanocoating for prolonged circulation time and reduced accelerated blood clearance. Small. 2015;11(46):6225–6236. doi:10.1002/smll.201502388
152. Zhang Q, Dehaini D, Zhang Y, et al. Neutrophil membrane-coated nanoparticles inhibit synovial inflammation and alleviate joint damage in inflammatory arthritis. Nat Nanotechnol. 2018;13(12):1182–1190. doi:10.1038/s41565-018-0254-4
153. Yu Q, Huang Y, Chen X, et al. A neutrophil cell membrane-biomimetic nanoplatform based on l-arginine nanoparticles for early osteoarthritis diagnosis and nitric oxide therapy. 10.1039/D2NR02601E. Nanoscale. 2022;14(32):11619–11634. doi:10.1039/D2NR02601E
154. Portier I, Campbell RA. Role of platelets in detection and regulation of infection. Arteriosclerosis Thrombosis Vasc Biol. 2021;41(1):70–78. doi:10.1161/ATVBAHA.120.314645
155. Antoniak S, Mackman N. Platelets and viruses. Platelets. 2021;32(3):325–330. doi:10.1080/09537104.2021.1887842
156. Nording H, Langer HF. Complement links platelets to innate immunity. Seminars Immunol. 2018;37:43–52. doi:10.1016/j.smim.2018.01.003
157. Jenne CN, Kubes P. Platelets in inflammation and infection. Platelets. 2015;26(4):286–292. doi:10.3109/09537104.2015.1010441
158. Jiang SZ, To JL, Hughes MR, McNagny KM, Kim H. Platelet signaling at the nexus of innate immunity and rheumatoid arthritis. Mini Rev. 2022;13. doi:10.3389/fimmu.2022.977828.
159. Golebiewska EM, Poole AW. Platelet secretion: from haemostasis to wound healing and beyond. Blood Rev. 2015;29(3):153–162. doi:10.1016/j.blre.2014.10.003
160. O-o O-A, Page MJ, Soma P, Pretorius E. Platelets: emerging facilitators of cellular crosstalk in rheumatoid arthritis. Nat Rev Rheumatol. 2019;15(4):237–248. doi:10.1038/s41584-019-0187-9
161. Stone JP, Wagner DD. P-selectin mediates adhesion of platelets to neuroblastoma and small cell lung cancer. J Clin Investig. 1993;92(2):804–813. doi:10.1172/JCI116654
162. C-mj H, Fang RH, Wang K-C, et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature. 2015;526(7571):118–121. doi:10.1038/nature15373
163. He Y, Li R, Liang J, et al. Drug targeting through platelet membrane-coated nanoparticles for the treatment of rheumatoid arthritis. Nano Res. 2018;11(11):6086–6101. doi:10.1007/s12274-018-2126-5
164. Liu H, Li R, Liu T, Yang L, Yin G, Xie Q. Immunomodulatory effects of mesenchymal stem cells and mesenchymal stem cell-derived extracellular vesicles in rheumatoid arthritis. Mini Rev. 2020;11. doi:10.3389/fimmu.2020.01912.
165. Tavasolian F, Moghaddam AS, Rohani F, et al. Exosomes: effectual players in rheumatoid arthritis. Autoimmun Rev. 2020;19(6):102511. doi:10.1016/j.autrev.2020.102511
166. Bruno MC, Cristiano MC, Celia C, et al. Injectable drug delivery systems for osteoarthritis and rheumatoid arthritis. ACS Nano. 2022;16(12):19665–19690. doi:10.1021/acsnano.2c06393
167. Spees JL, Lee RH, Gregory CA. Mechanisms of mesenchymal stem/stromal cell function. Stem Cell Res Ther. 2016;7(1):125. doi:10.1186/s13287-016-0363-7
168. Huang Y, Wu Q, Tam PK. Immunomodulatory mechanisms of mesenchymal stem cells and their potential clinical applications. Int J Mol Sci. 2022;23(17). doi:10.3390/ijms231710023
169. Belhadj Z, He B, Deng H, et al. A combined “eat me/don’t eat me” strategy based on extracellular vesicles for anticancer nanomedicine. J Extracell Vesicles. 2020;9(1):1806444. doi:10.1080/20013078.2020.1806444
170. Yan F, Zhong Z, Wang Y, et al. Exosome-based biomimetic nanoparticles targeted to inflamed joints for enhanced treatment of rheumatoid arthritis. J Nanobiotechnol. 2020;18(1):115. doi:10.1186/s12951-020-00675-6
171. Li H, Feng Y, Zheng X, et al. M2-type exosomes nanoparticles for rheumatoid arthritis therapy via macrophage re-polarization. J Controlled Release. 2022;341:16–30. doi:10.1016/j.jconrel.2021.11.019
172. Tessandier N, Melki I, Cloutier N, et al. Platelets disseminate extracellular vesicles in lymph in rheumatoid arthritis. Arteriosclerosis Thrombosis Vasc Biol. 2020;40(4):929–942. doi:10.1161/ATVBAHA.119.313698
173. Ma Q, Bai J, Xu J, et al. Reshaping the inflammatory environment in rheumatoid arthritis joints by targeting delivery of berberine with platelet-derived extracellular vesicles. Adv NanoBiomed Res. 2021;1(11):2100071. doi:10.1002/anbr.202100071
174. Hee kim S, Bianco N, Menon R, et al. Exosomes derived from genetically modified DC expressing FasL are anti-inflammatory and immunosuppressive. Mol Ther. 2006;13(2):289–300. doi:10.1016/j.ymthe.2005.09.015
175. Wu X, Zhu D, Tian J, et al. Granulocytic myeloid-derived suppressor cell exosomal prostaglandin E2 ameliorates collagen-induced arthritis by enhancing IL-10+ B cells. Orig Res. 2020;11. doi:10.3389/fimmu.2020.588500
176. Song H, Li X, Zhao Z, et al. Reversal of osteoporotic activity by endothelial cell-secreted bone targeting and biocompatible exosomes. Nano Lett. 2019;19(5):3040–3048. doi:10.1021/acs.nanolett.9b00287
177. Ding H, Lv Y, Ni D, et al. Erythrocyte membrane-coated NIR-triggered biomimetic nanovectors with programmed delivery for photodynamic therapy of cancer. Nanoscale. 2015;7(21):9806–9815. doi:10.1039/C5NR02470F
178. Li C, Wang X, Li R, et al. Resveratrol-loaded PLGA nanoparticles functionalized with red blood cell membranes as a biomimetic delivery system for prolonged circulation time. J Drug Delivery Sci Technol. 2019;54:101369. doi:10.1016/j.jddst.2019.101369
179. Yu H, Fan J, Shehla N, et al. Biomimetic hybrid membrane-coated xuetongsu assisted with laser irradiation for efficient rheumatoid arthritis therapy. ACS Nano. 2022;16(1):502–521. doi:10.1021/acsnano.1c07556
180. Zhao C, Song W, Ma J, Wang N. Macrophage-derived hybrid exosome-mimic nanovesicles loaded with black phosphorus for multimodal rheumatoid arthritis therapy. 10.1039/D2BM01274J. Biomat Sci. 2022;10(23):6731–6739. doi:10.1039/D2BM01274J
181. Saha S, Anilkumar AA, Mayor S. GPI-anchored protein organization and dynamics at the cell surface. J Lipid Res. 2016;57(2):159–175. doi:10.1194/jlr.R062885
182. Escribá PV, Busquets X, J-i I, et al. Membrane lipid therapy: modulation of the cell membrane composition and structure as a molecular base for drug discovery and new disease treatment. Progress Lipid Res. 2015;59:38–53. doi:10.1016/j.plipres.2015.04.003
183. Hao X, Gan J, Cao J, Zhang D, Liang J, Sun L. Biomimetic liposomes hybrid with erythrocyte membrane modulate dendritic cells to ameliorate systemic lupus erythematosus. Mat Today Bio. 2023;20:100625. doi:10.1016/j.mtbio.2023.100625
184. Parodi A, Quattrocchi N, van de Ven AL, et al. Synthetic nanoparticles functionalized with biomimetic leukocyte membranes possess cell-like functions. Nat Nanotechnol. 2013;8(1):61–68. doi:10.1038/nnano.2012.212
185. Yan H, Shao D, Lao Y-H, Li M, Hu H, Leong KW. Engineering cell membrane-based nanotherapeutics to target inflammation. Adv Sci. 2019;6(15):1900605. doi:10.1002/advs.201900605
186. Giraldo JA, Molano RD, Rengifo HR, et al. The impact of cell surface PEGylation and short-course immunotherapy on islet graft survival in an allogeneic murine model. Acta Biomaterialia. 2017;49:272–283. doi:10.1016/j.actbio.2016.11.060
187. Lutterotti A, Yousef S, Sputtek A, et al. Antigen-specific tolerance by autologous myelin peptide–coupled cells: a Phase 1 trial in multiple sclerosis. Sci Transl Med. 2013;5(188):188ra75–188ra75. doi:10.1126/scitranslmed.3006168
188. Cheng H, Byrska-Bishop M, Zhang CT, et al. Stem cell membrane engineering for cell rolling using peptide conjugation and tuning of cell–selectin interaction kinetics. Biomaterials. 2012;33(20):5004–5012. doi:10.1016/j.biomaterials.2012.03.065
189. Zhou H, Fan Z, Lemons PK, Cheng H. A facile approach to functionalize cell membrane-coated nanoparticles. Theranostics. 2016;6(7):1012–1022. doi:10.7150/thno.15095
190. Wang Z, Zhang C, Meng J, et al. A targeted exosome therapeutic confers both CfDNA scavenging and macrophage polarization for ameliorating rheumatoid arthritis. Adv Mat. 2023;35(48):2302503. doi:10.1002/adma.202302503
191. Lee ES, Sul JH, Shin JM, et al. Reactive oxygen species-responsive dendritic cell-derived exosomes for rheumatoid arthritis. Acta Biomaterialia. 2021;128:462–473. doi:10.1016/j.actbio.2021.04.026
192. Abbina S, Siren EMJ, Moon H, Kizhakkedathu JN. Surface engineering for cell-based therapies: techniques for manipulating mammalian cell surfaces. ACS Biomat Sci Eng. 2018;4(11):3658–3677. doi:10.1021/acsbiomaterials.7b00514
193. Deng C, Zhao X, Chen Y, et al. Engineered platelet microparticle-membrane camouflaged nanoparticles for targeting the golgi apparatus of synovial fibroblasts to attenuate rheumatoid arthritis. ACS Nano. 2022;16(11):18430–18447. doi:10.1021/acsnano.2c06584
194. Wang Z, Chen C, Shi C, et al. Cell membrane derived liposomes loaded with DNase I target neutrophil extracellular traps which inhibits colorectal cancer liver metastases. J Controlled Release. 2023;357:620–629. doi:10.1016/j.jconrel.2023.04.013
195. Shin MJ, Park JY, Lee DH, Khang D. Stem cell mimicking nanoencapsulation for targeting arthritis. Int J Nanomed. 2021;16(null):8485–8507. doi:10.2147/IJN.S334298
196. Mansouri P, Mansouri P, Najafipour S, Kouhpayeh SA, Farjadfar A, Behmard E. Multi-target drug discovery for rheumatoid arthritis: a comprehensive computational approach using bioactive compounds. Scient Rep. 2025;15(1):27516. doi:10.1038/s41598-025-12666-4
197. Figus FA, Piga M, Azzolin I, McConnell R, Iagnocco A. Rheumatoid arthritis: extra-articular manifestations and comorbidities. Autoimmun Rev. 2021;20(4):102776. doi:10.1016/j.autrev.2021.102776
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.