Back to Journals » OncoTargets and Therapy » Volume 11
Radix Tetrastigma hemsleyani flavone exhibits antitumor activity in colorectal cancer via Wnt/β-catenin signaling pathway
Authors Wu X, Yu N, Zhang Y, Ye Y, Sun W, Ye L, Wu H, Yang Z, Wu L, Wang F
Received 23 April 2018
Accepted for publication 23 June 2018
Published 4 October 2018 Volume 2018:11 Pages 6437—6446
DOI https://doi.org/10.2147/OTT.S172048
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Samir Farghaly
Xiaowei Wu,1,* Na Yu,1,* Yuping Zhang,2 Yuanning Ye,2 Wenrong Sun,1 Lei Ye,1 Huimin Wu,3 Zhao Yang,2 Lin Wu,1 Fangyu Wang3
1Department of Gastroenterology and Hepatology, Jinling Hospital, Nanjing, China; 2Department of Gastroenterology and Hepatology, Jinling Hospital, Southern Medical University, Guangzhou, China; 3Department of Gastroenterology and Hepatology, Jinling Hospital, Medical School of Nanjing University, Nanjing, China
*These authors contributed equally to this work
Background: Radix Tetrastigma hemsleyani flavone (RTHF) is extracted from a traditional Chinese medicinal herb T. hemsleyani, which is conventionally used as a folk medicine for its anti-inflammation activity and antiviral activity. In this study, the effects of RTHF on inhibiting malignant biological properties in colorectal cancer (CRC) were evaluated by conducting both in vitro and in vivo experiments, and the underlying mechanism was investigated.
Materials and methods: Cell Counting Kit-8, colony formation, and flow cytometry assays were performed to evaluate the proliferation of RTHF-treated colon tumor cells. Migration and invasion capacities were also tested by cell wound scratch assay and Transwell invasion assay. Moreover, the antitumor effects of RTHF on azoxymethane/dextran sulfate sodium-induced colitis-related CRC were investigated in C57BL/6 mice. In addition, Western blot and/or quantitative reverse transcription polymerase chain reaction analysis were used to evaluate the expressions of Lgr5, Cyclin D1, c-Myc, and E-cadherin.
Results: These experiments showed that RTHF could decrease the cell growth kinetics and clone-forming capacity. RTHF could also dose dependently induce cell cycle arrest at G0/G1 phase and inhibit epithelial -mesenchymal transition process. Furthermore, downregulation of β-catenin activation and downstream protein expression were detected in CRC cells after being treated with RTHF. RTHF daily gavage suppressed the number and size of CRC in mice and inhibited Lgr5 and Cyclin D1 expressions in tumor tissue.
Conclusion: In conclusion, RTHF treatment inhibits colorectal tumor growth, decreases Wnt/β-catenin pathway activity, and downregulates target genes’ expression.
Keywords: radix Tetrastigma hemsleyani, flavone, colorectal cancer, Wnt/β-catenin, proliferation, migration
Introduction
Colorectal cancer (CRC), as common malignancy, is one of the most common causes of death around the world. Besides conventional therapies of CRC, traditional Chinese medicines play a potentially role in CRC treatment.1,2 Flavonoids are the main active ingredients of many Chinese herbal medicines. Many studies have shown that flavonoids can inhibit the growth of tumor cells with low cytotoxicity to normal cells.3,4 For example, procyanidin B1 acetylated analogs can inhibit the proliferation of HeLa cell line.5 Rutin decreases cellular viability and proliferation and increases cell differentiation and apoptosis of CRC. Rutin also regulates cell adherence and chemoresistance in leukemic cells.6 Quercetin displays a stronger cytotoxicity on HepG2 cells.7 Quercitrin shows antiapoptotic and antiproliferative effects in in lung cancer cells by modulating the immune response.8 Another flavonoid, isoquercitrin, inhibits CRC cells growth by targeting the Wnt/β-catenin pathway.9
Tetrastigma hemsleyani, which is mainly distributed in southwest China, is an endangered precious traditional Chinese medicinal herb. The herb is widely used for pneumonia, hepatitis, scrofula, infantile febrile convulsion, menstrual disorders, asthma, and rheumatism.10 Primary medicinal parts of T. hemsleyani, its roots, are rich in flavonoids.11,12 Radix T. hemsleyani flavone (RTHF), which is considered to be an important effective component of the herb, has been demonstrated to have anti-inflammatory, analgesic, antipyretic, hepatoprotective, antiviral, and immunomodulatory activities.13,14
In this investigation, we treated CRC cells with different dosages of RTHF in vitro and evaluated whether RTHF could suppress CRC growth in mice. Additionally, we also investigated potential molecular mechanisms underlying the therapeutic effects of RTHF against CRC.
Materials and methods
Reagents and cell culture
RTHF was obtained from Jiangsu Zelang Medical Technology Company Limited. HT29 and SW620 cells lines were purchased from Shanghai Cell Collection. All cells were cultured in DMEM (Thermo Fisher Scientific, Waltham, MA, USA). The medium was supplemented with 10% fetal bovine serum and 1% penicillin and streptomycin (Thermo Fisher Scientific). Cells were incubated in an atmosphere of humidified 5% CO2 and at 37°C. RTHF was dissolved in dimethyl sulfoxide (Sigma-Aldrich Co., St Louis, MO, USA). The percentage of dimethyl sulfoxide was <0.1% in the medium. When cells were cultured with RTHF, the medium was changed every 48 hours.
Cell Counting Kit-8 assay
HT29 and SW620 cells were seeded in 96-well plates at a density of 5×103 cells/well and treated with different concentration of RTHF (0, 1.6, 3.2, and 6.4 mg/mL, respectively).14–16 Following incubation for 0, 24, 48, and 72 hours, 10 μL Cell Counting Kit-8 solution (Beyotime, Shanghai, China) was added to each well and plates were incubated at 37°C for 1 hour. Then, absorbance at 450 nm was recorded using a microplate reader (Bio-Rad Laboratories Inc., Hercules, CA, USA). Cell viability was calculated as the average of three duplicate wells.
Colony formation assay
Cells were seeded in six-well plates at a density of 3×102 cells/well and treated with RTHF for 12 days at 37°C. For each concentration of RTHF, three duplicate wells were prepared. Cells were fixed with 4% paraformaldehyde for 15 minutes and stained with crystal violet for 10 minutes. Plates were imaged using a light microscope (Olympus Corporation, Tokyo, Japan). Cluster containing >50 cells was treated as a clone.
Flow cytometry
Cells (1×106 cells/well) were harvested and fixed with 70% ethanol at 4°C for 1 hour. Then, the cells were centrifuged (100×g, 5 minutes), resuspended, and incubated with RNaseA (1 mg/mL) at 37°C for 30 minutes, followed by staining with propidium iodide (0.04 mg/mL) at 4°C for another 30 minutes. BD FACS Flow Cytometry Calibur (BD Biosciences, San Jose, CA, USA) was used to analyze DNA content of cell suspension. Experiments were performed in triplicate.
Cell wound scratch assay
Cells were seeded in 24-well plates at a density of 5×105 cells/well. After 24 hours of incubation, a tip of plastic micropipette was used to scratch straight lines in the cell monolayer of HT29 and SW620 cells. Then, wells were flushed three times with PBS and cultured in a medium containing RTHF for another 48 hours. Experiments were performed in triplicate and images were acquired using a microscope.
Transwell invasion experiments
Cells (1×104 cells/well) were seeded and cultured with serum-starved medium which contained RTHF in the upper well of Matrigel-coated chambers (BD Biosciences), while the medium containing 10% fetal bovine serum and RTHF was added to the bottom well of chambers. After 24 hours, cells which did not migrate were removed from the upper face of filters by cotton swabs. Migrated cells were fixed with 4% paraformaldehyde and stained by crystal violet solution. The number of invading cells was counted as an average of three duplicate wells and photographed using a light microscope.
Immunofluorescence
Paraformaldehyde-fixed SW620 and HT29 cells were blocked by goat serum and incubated with diluted antibodies against E-cadherin (1:200, Abcam) and vimentin (1:100, Abcam) at 4°C overnight. Fluorescent secondary antibody was incubated at 37°C for 40 minutes. Then, slides of cells were visualized by 4′,6-diamidino-2-phenylindole (Beyotime) and sealed by glycerol. Three duplicate wells were prepared for each concentration of RTHF. Images were observed under a laser scanning confocal microscope.
Western blot
Total protein was extracted from cells treated with RTHF for 48 hours using 99% Radio immunoprecipitation assay (RIPA)+1%Phenylmethanesulfonyl fluoride (PMSF) (Beyotime). For nuclear protein extraction, EpiQuik Nuclear Extraction Kit (EpiGentek, Farmingdale, NY, USA) was used. Cell lysates (50 μg) were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a nitrocellulose membrane (Bio-Rad). Membranes were blocked in 5% bovine serum albumin (Sigma-Aldrich) in TBS containing 0.05% Tween-20 (TBST) for 2 hours and incubated with primary antibodies (active β-catenin [1:1,000; EMD Millipore, Billerica, MA, USA], E-cadherin [1:1,000, Abcam], vimentin [1:1,000, Abcam], Cyclin D1 [1:1,000, Abcam], c-Myc [1:1,000, Abcam], Lgr5 [1:1,000; Abcam], and β-actin [1:5,000, Epitomics]) at 4°C overnight. Antibodies were detected with horseradish peroxidase-conjugated secondary antibodies (1:5,000, Epitomics), followed by visualization with Immobilon Western Chemiluminescent HRP Substrate detection reagents (EMD Millipore). Each experiment was performed three times. Photographs were taken using a ChemiDoc Imaging System (Tanon, Shanghai, China).
Quantitative reverse transcription-polymerase chain reaction analysis (qRT-PCR)
Total RNA was extracted from cells using TRIzol reagent (Thermo Fisher Scientific) and reverse transcribed into cDNA using PrimeScript™ RT kit (Takara, Kusatsu, Japan). SYBR premix Ex Taq™ Green II (Takara) was used on a real-time PCR system (Bio-Rad) to determine the relative levels of target gene mRNA transcripts. Each experiment was performed three times to ensure experimental accuracy. Primers used for qRT-PCR were as follows: Lgr5: forward sequence (FS): CCTGCTTGACTTTGAGGAAGACC, reverse sequence (RS): CCAGCCATCAAGCAGGTGTTCA; Cyclin D1: FS: CTACACCGACAACTCCATCCG, RS: TCTGGCATTTTGGAGAGGAAGTG; c-Myc: FS: CCTGGTGCTCCATGAGGAGAC, RS: CAGACTCTGACCTTTTGCCAGG. Relative mRNA levels of target gene were calculated with respect to the control gene β-actin and expressed in terms of fold change.
Animals
C57BL/6 mice at ~4–6 weeks and weighing 23–25 g were housed in pathogen-free plastic cages with a daylight cycle of 12 hours every day at 21°C–23°C. Mice were fed a standard laboratory rodent chow diet (Nanjing Huaxin Biological Technology Co. Ltd, Nanjing City, China) and had free access to water. Animals were purchased from the Department of Comparative Medicine in Jinling Hospital. The protocols of animal studies were approved by Jinling Hospital Animal Care Committee. All mice received humane care in accordance with the National Institutes of Health animal use guidelines.
CRC model
Forty male C57BL/6 mice were randomly divided into five groups, including blank control (Ctr) group, tumor model (Mod) group, vehicle control (Veh) group, low concentration treatment (Low) group, and high concentration treatment (Hig) group. Each group of mice, except for Ctrl group, was given an intraperitoneal injection of azoxymethane (AOM; Sigma-Aldrich) at a dose of 10 mg/kg body weight. The day after injection, mice received 2% dextran sulfate sodium (DSS; Sigma-Aldrich) solution as drinking water for 5 days, following which they were given distilled water for 14 days. After five cycles of DSS treatment, mice were given a standard diet and distilled water for an additional 19 days.17,18
RTHF was suspended in 0.5% sodium carboxymethyl cellulose. After the first day of the second DSS cycle, mice of Low group and Hig group were given RTHF (30 and 60 mg/kg, respectively) by gavage until the end of experiment. Dosage of RTHF was calculated depending on traditional prescriptions which contained the raw herb and other studies.13,14 In parallel, the Veh group was given the same volume of 0.5% carboxymethyl cellulose. All mice were euthanized. Colons were excised, flushed with ice-cold PBS, and fixed in 4% paraformaldehyde overnight.
Histological analysis
Colorectal sections were stained with H&E. For immunohistochemistry, paraffin-embedded colorectal tissue sections were autoclaved for 2 minutes in citrate buffer to promote antigen retrieval. After blocking in 5% bovine serum albumin, sections were incubated with primary antibodies against Lgr5 (1:100, Abcam) and Cyclin D1 (1:100, Abcam) at 4°C for overnight. Sections were then incubated with secondary antibodies for 30 minutes at 37°C, visualized with Diaminobenzidine (DAB) (Dako Denmark A/S, Glostrup, Denmark), and then counterstained with hematoxylin and observed under a light microscope using Olympus Camedia Software. Two pathologists analyzed the expressions of Lgr5 and Cyclin D1 in a blinded manner. For quantification of the immunohistochemical staining, relative protein expression of Lgr5 and Cyclin D1 was quantified by Image Pro Plus version 6.0 software (Media Cybernetics, Inc., Rockville, MD, USA) and was defined as follows: relative protein expression=IOD/area sum, where IOD represents integrated OD.
Statistical analysis
The outcomes were expressed as mean±SD or percentage. Statistical significance was determined with independent sample t-test and one-way analysis of variance. P<0.05 was considered as a significant difference. Graph generation and statistical analyses were performed with SPSS 19.0 software (IBM Corporation, Armonk, NY, USA).
Results
RTHF suppresses CRC cells proliferation
Human CRC cell lines SW620 and HT29 were incubated in RTHF for 0, 24, 48, and 72 hours. Results showed that RTHF inhibited cell proliferation in a dose-dependent manner (Figure 1A). Additionally, plate clone formation assay was used to analyze the colony formation of RTHF-treated SW620 and HT29 cells (Figure 1B). When cultured with 1.6 mg/mL RTHF, the number of clones was significantly suppressed. With higher concentration of RTHF, no cell clone was formed. These results suggest that RTHF inhibits tumor cells proliferation in a dosage-dependent manner in vitro.
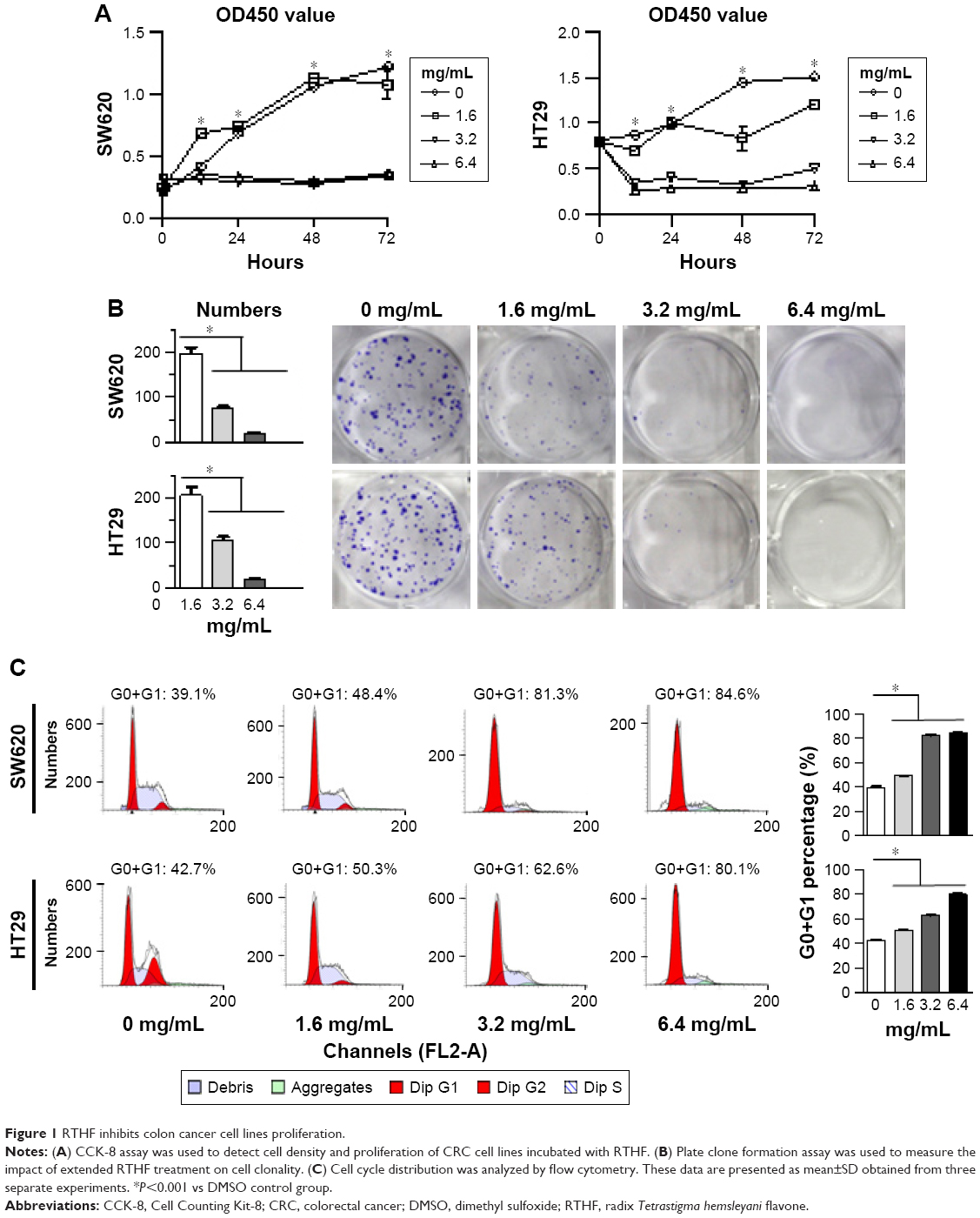 | Figure 1 RTHF inhibits colon cancer cell lines proliferation. |
RTHF induces cell cycle arrest at G0/G1 phases
To examine the impact of RTHF on cell cycle, flow cytometry was used to analyze the cell cycle of SW620 and HT29 cells (Figure 1C). With increased RTHF concentration, G0+G1 proportions of SW620 and HT29 cells were (39.1%, 48.4%, 81.3%, and 84.6%) and (42.7%, 50.3%, 62.6%, and 80.1%), respectively. G0+G1 proportions were significantly increased, suggesting that tumor cells were trapped in G0/G1 phase in a dose-dependent manner.
RTHF inhibits migration and invasion of CRC cells
Cell wound scratch assay was used to determine the rate of cell migration. When cells were cultured with 1.6 mg/mL RTHF as indicated, the motility of tumor cells was decreased (Figure 2A). Besides, tumor cell invasion was evaluated by Transwell invasion experiments. Compared with control groups, the number of invading cells was reduced significantly by RTHF treatment (Figure 2B).
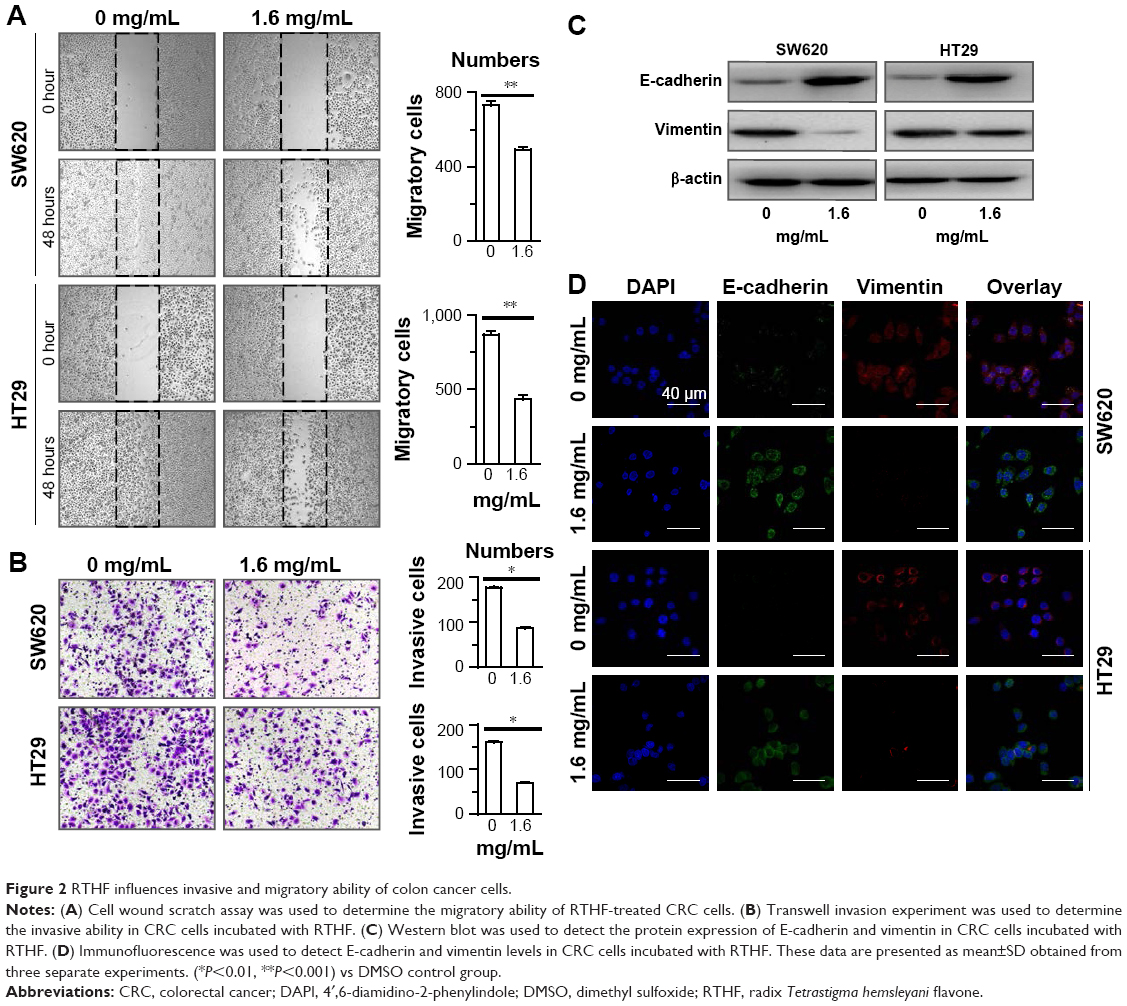 | Figure 2 RTHF influences invasive and migratory ability of colon cancer cells. |
RTHF induces mesenchymal–epithelial transition of CRC cells
Furthermore, expressions of epithelial–mesenchymal transition (EMT) process associated protein, E-cadherin and vimentin, were detected by Western blot (Figure 2C). E-cadherin was increased in the presence of RTHF, while vimentin expression was significantly decreased. Again, immunofluorescence also demonstrated the upregulation of E-cadherin and downregulation of vimentin expression (Figure 2D), indicating that RTHF may suppress EMT in vitro.
RTHF inhibits Wnt/β-catenin pathway activity in CRC cells
Western blot showed that β-catenin level was significantly reduced in RTHF-treated group. RTHF also inhibited Lgr5, c-Myc, and Cyclin D1 expression in a dose-dependent manner (Figure 3A and B). Results of Western blot were further confirmed by qRT-PCR (Figure 3C and D). All these results suggest that RTHF downregulates Wnt/β-catenin pathway activation.
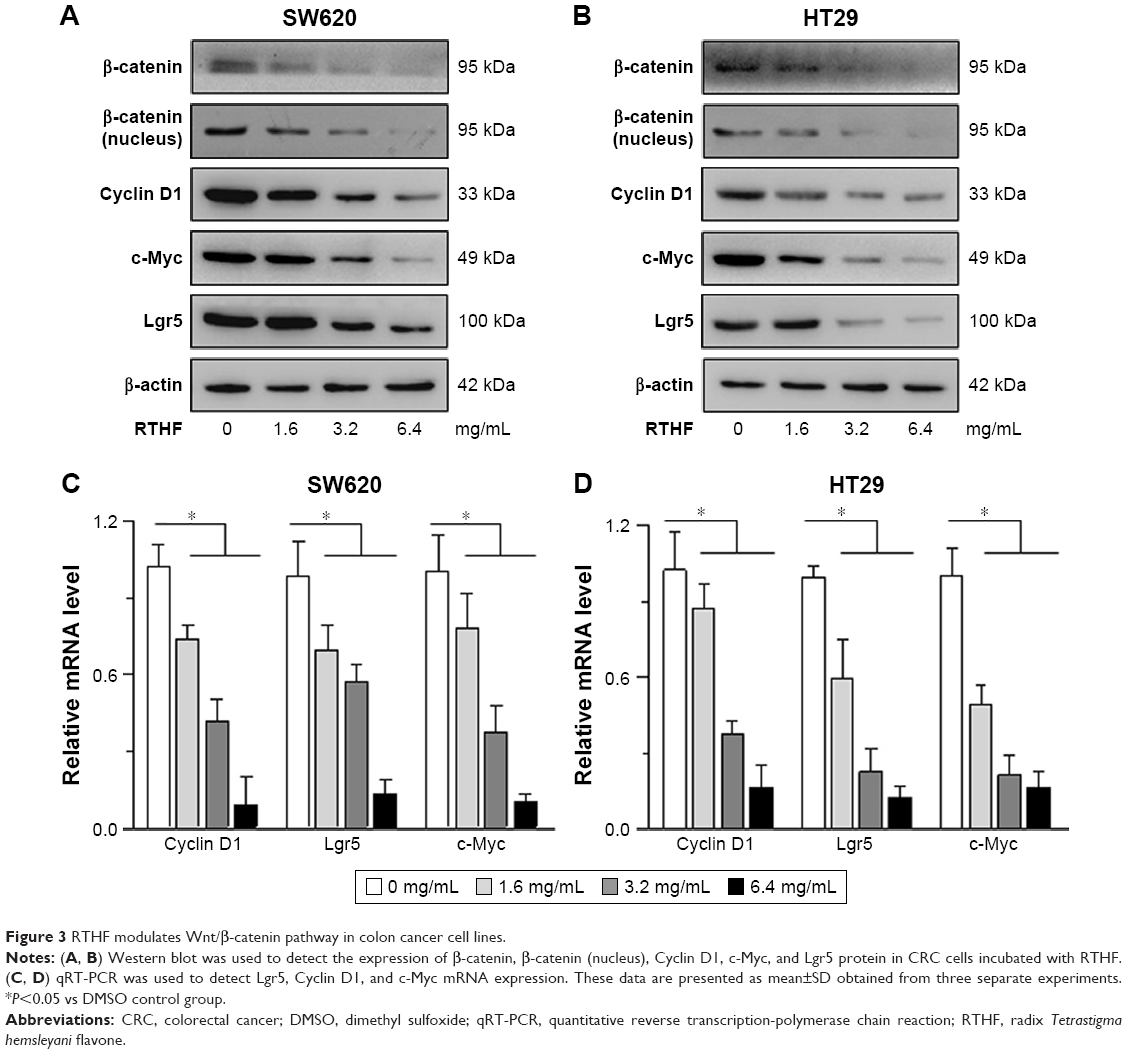 | Figure 3 RTHF modulates Wnt/β-catenin pathway in colon cancer cell lines. |
RTHF inhibits tumor formation in an inflammation-driven spontaneous CRC model in C57BL/6 mice
Next we wanted to know if RTHF can prevent AOM/DSS-induced CRC in mice. Mice in Mod group and Veh group showed significant weight loss, extensive adenomatous polyps, and tumor growth, which were mitigated in Low group and Hig group. No adenomatous polyps and tumor were observed in Ctr group (Figure 4A and B). Furthermore, we found that the average number of tumors, maximum tumor diameter, and total tumor volume were significantly greater in Mod and Veh groups compared with Hig group and to a lesser extent in Low group (Figure 4C–E).
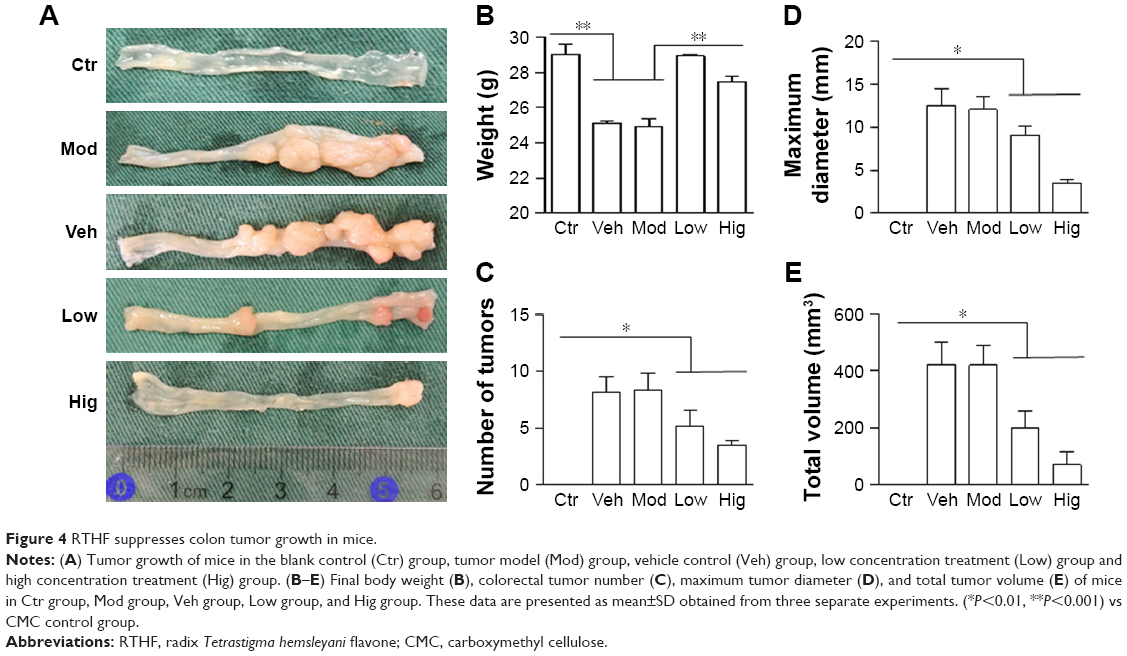 | Figure 4 RTHF suppresses colon tumor growth in mice. |
Tumors from Mod group and Veh group were highly dysplastic with glandular architectural distortion, cauliflower-like surfaces, and cytological alterations, including pleomorphic nuclei and pathological mitosis.19 However, mice treated with low and high dose of RTHF displayed milder pathological changes (Figure 5A–E).
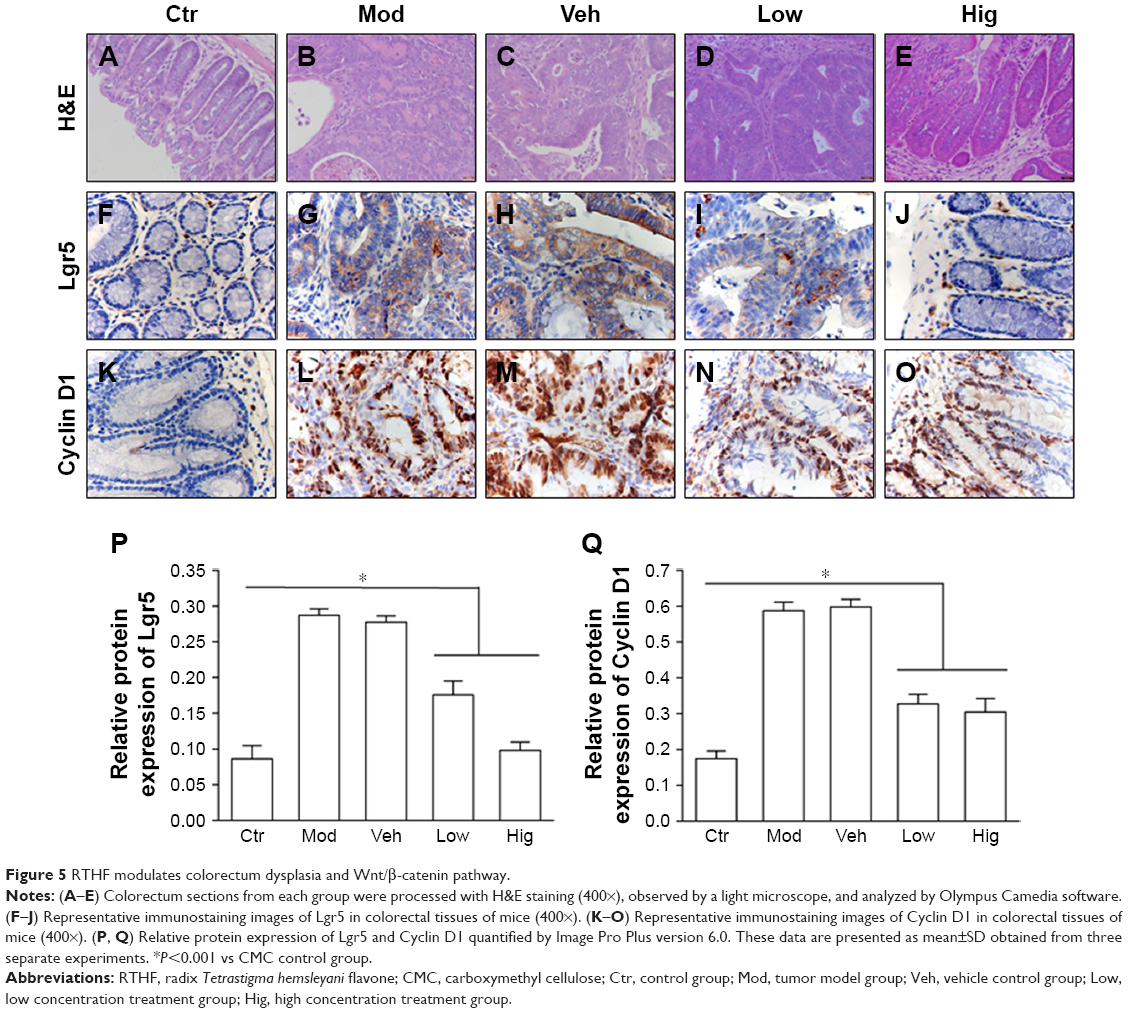 | Figure 5 RTHF modulates colorectum dysplasia and Wnt/β-catenin pathway. |
RTHF inhibits CRC cells proliferation and downregulates the expressions of β-catenin–responsive genes in vivo
In AOM/DSS models, immunostaining of CRC tissue revealed that Lgr5 was expressed in the cytoplasm. Lgr5 expression, which is rarely seen in normal colon tissue, was correlated with tumor heteromorphism.20 Compared with Hig group, Low group mice expressed higher Lgr5 levels, while the highest level of Lgr5 was observed in Mod and Veh groups (Figure 5F–J). Similar to Lgr5, Cyclin D1 was also accumulated in the cell nuclei compared with Ctr group. However, after RTHF treatment, Cyclin D1 expression signifdecreased, especially in Hig group (Figure 5K–O). Relative protein expression of Lgr5 and Cyclin D1 were presented in Figure 5P and Figure 5Q. Collectively, RTHF downregulated β-catenin activation and modulated Lgr5, c-Myc, and Cyclin D1 expressions in a dose-dependent manner. These data indicate that Wnt/β-catenin signaling is involved in RTHF regulation of CRC.
Discussion
Flavonoids are widely present in plants, and the general structure of flavonoids is a 15-carbon skeleton consisting of two phenyl rings and a heterocyclic ring. It has been demonstrated that a variety of flavonoids can inhibit proliferation and induce apoptosis of tumor cells.21 Zhong et al15 reported that RTHF exerted an antiapoptosis and anti-growth activity against lung cancer A549 cells. Furthermore, they explored and found that RTHF exhibited anti-growth and antimetastasis activity in lung cancer A549 cells by regulating the expression of MMPs and tissue inhibitors of metalloproetienases.22
In this effort, the in situ colon adenocarcinoma cell line HT29 and the peritoneal lymph node metastasis tumor cell line SW620 were cultured with different doses of RTHF. Our results showed that RTHF inhibited multiple malignant biological behaviors of CRC cells in a dose-dependent manner. For the first time, this study found that RTHF can reduce activated β-catenin level in CRC cells, indicating that RTHF may inhibit Wnt pathway activation. Wnt/β-catenin pathway is one of the necessary signaling pathways in regulating cell proliferation, differentiation, migration, adhesion, and self-renewal.23,24 Mutations in Wnt/β-catenin pathway are detected in 90% of CRC cases and correlate with the poor prognosis of CRC.25,26 Activated β-catenin serves as a transcription cofactor to activate multiple downstream target genes, such as Lgr5, Cyclin D1, c-Myc, and E-cadherin, which are major genes involved in the malignant progression of CRC.27–29
Lgr5 is a marker of cancer stem cells in many malignant diseases. Cancer stem cells, considered to be the origin of tumor self-renewal and differentiation, are essential in tumor progression, development, and chemotherapy resistance.30,31 Many studies investigated that overexpression of Lgr5 was positively correlated with CRC initiation and 5-fluorouracil–based chemotherapy resistance. High level of Lgr5 expression also indicated less overall survival and deeper invasion in CRC patients.24 Interestingly, RTHF suppressed Lgr5 expression, which was negatively correlated with the dephosphorylation of β-catenin, indicating that the self-renewal capacity of tumor cells might be reduced by RTHF.
In the present work, RTHF-cultured tumor cells were arrested in G0/G1 phase in a dose-dependent manner. Moreover, Cyclin D1 and c-Myc were diminished at both protein and transcriptional levels in CRC cells cultured with RTHF. Cell cycle is driven by the sequential activation of cyclins and cyclin-dependent kinases in all eukaryotic cells.32,33 Wnt/β-catenin signaling promotes cell cycle progression by upregulating target genes, such as c-Myc and Cyclin D1.34,35 Our data indicated that RTHF might, therefore, regulate mitotic events by means of inhibiting Wnt/β-catenin pathway.
Metastasis is the main cause of death in patients with cancer. Several studies demonstrate that aberrant reactivation of EMT can potently drive tumor metastasis.36,37 EMT is a unique process that transforms immotile epithelial cells to motile mesenchymal cells, leading to an augment of migration and invasion in malignant diseases. The EMT process is characterized by a decrease of cell–cell contact–associated protein expression, such as E-cadherin and γ-catenin, as well as an increase of mesenchymal marker expression, such as vimentin, N-cadherin, and fibronectin.38,39 Wnt/β-catenin pathway is associated with the regulation of EMT. Activated β-catenin protein binds to E-cadherin and modulates the EMT process.40 We found that RTHF could increase E-cadherin expression and decrease vimentin expression in cultured CRC cells, which meant that RTHF might suppress EMT progression.
Furthermore, we analyzed the effects of RTHF on colitis-associated CRC in mice. Compared with control groups, RTHF-treated mice had substantially higher body weight, fewer colorectal tumors, and lesser tumor volume. Colorectal sections demonstrated that RTHF-treated mice showed minor heteromorphism and invasion depth. Also, target genes of Wnt/β-catenin pathway were evaluated in colorectal tissue. Decrease in the expression of both Lgr5 and Cyclin D1 was found in RTHF-treated mice, suggesting that the RTHF inhibitory effects on CRC growth might be mediated by inactivating Wnt pathway, thereby reducing CRC proliferation in mice.
Conclusion
RTHF inhibits the growth and migration of CRC in vitro and in vivo, suggesting that RTHF is potentially applicable for treating CRC in clinical settings. Wnt/β-catenin pathway participates in cancer generation and development; therefore, inactivation of Wnt/β-catenin pathway by RTHF deserves to be further tested in CRC.
Acknowledgments
We are indebted to Dr Lianming Liao (Fujian Academy of Integrative Medicine, Fujian University of Traditional Chinese Medicine, Fuzhou, China) for helpful discussions. This work was supported by grants from the National Natural Science Foundation of China (NSFC No 81270453).
Disclosure
The authors report no conflicts of interest in this work.
References
Chen H, Tian T, Miao H, Zhao YY. Traditional uses, fermentation, phytochemistry and pharmacology of Phellinus linteus: a review. Fitoterapia. 2016;113:6–26. | ||
Chen M, May BH, Zhou IW, Sze DM, Xue CC, Zhang AL. Oxaliplatin-based chemotherapy combined with traditional medicines for neutropenia in colorectal cancer: a meta-analysis of the contributions of specific plants. Crit Rev Oncol Hematol. 2016;105:18–34. | ||
Mohammed RS, Abou Zeid AH, El Hawary SS, Sleem AA, Ashour WE. Flavonoid constituents, cytotoxic and antioxidant activities of Gleditsia triacanthos L. leaves. Saudi J Biol Sci. 2014;21(6):547–553. | ||
Kim YA, Kim H, Seo Y. Antiproliferative effect of flavonoids from the halophyte Vitex rotundifolia on human cancer cells. Nat Prod Commun. 2013;8(10):1405–1408. | ||
Okamoto S, Ishihara S, Okamoto T, et al. Inhibitory activity of synthesized acetylated procyanidin B1 analogs against HeLa S3 cells proliferation. Molecules. 2014;19(2):1775–1785. | ||
Bourogaa E, Bertrand J, Despeaux M, et al. Hammada scoparia flavonoids and rutin kill adherent and chemoresistant leukemic cells. Leuk Res. 2011;35(8):1093–1101. | ||
Barcelos GR, Grotto D, Angeli JP, et al. Evaluation of antigenotoxic effects of plant flavonoids quercetin and rutin on HepG2 cells. Phytother Res. 2011;25(9):1381–1388. | ||
Cincin ZB, Unlu M, Kiran B, Bireller ES, Baran Y, Cakmakoglu B. Molecular mechanisms of quercitrin-induced apoptosis in non-small cell lung cancer. Arch Med Res. 2014;45(6):445–454. | ||
Amado NG, Predes D, Fonseca BF, et al. Isoquercitrin suppresses colon cancer cell growth in vitro by targeting the Wnt/β-catenin signaling pathway. J Biol Chem. 2014;289(51):35456–35467. | ||
Dai Y, Shen Z, Liu Y, Wang L, Hannaway D, Lu H. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg. Environ Exp Bot. 2009;65(2–3):177–182. | ||
Xu W, Fu ZQ, Lin J, et al. Rapid simultaneous determination of ten major flavonoids in Tetrastigma hemsleyanum by UPLC-MS/MS. Yao Xue Xue Bao. 2014;49(12):1711–1717. | ||
Lin Z, Chen L, Qiu Q, Guo S. Isolation and identification of antiproliferative compounds from the roots of Tetrastigma hemsleyanum against MDA-MB-435S cell lines. Pak J Pharm Sci. 2016;29(4):1171–1175. | ||
Feng Z, Hao W, Lin X, Fan D, Zhou J. Antitumor activity of total flavonoids from Tetrastigma hemsleyanum Diels et Gilg is associated with the inhibition of regulatory T cells in mice. Onco Targets Ther. 2014;7:947–956. | ||
Xu CJ, Ding GQ, Fu JY, Meng J, Zhang RH, Lou XM. Immunoregulatory effects of ethyl-acetate fraction of extracts from Tetrastigma hemsleyanum Diels et. Gilg on immune functions of ICR mice. Biomed Environ Sci. 2008;21(4):325–331. | ||
Zhong LR, Chen X, Wei KM. Radix Tetrastigma hemsleyani flavone induces apoptosis in human lung carcinoma a549 cells by modulating the MAPK pathway. Asian Pac J Cancer Prev. 2013;14(10):5983–5987. | ||
Peng X, Zhuang DD, Guo QS. Induction of S phase arrest and apoptosis by ethyl acetate extract from Tetrastigma hemsleyanum in human hepatoma HepG2 cells. Tumour Biol. 2015;36(4):2541–2550. | ||
Cooks T, Pateras IS, Tarcic O, et al. Mutant p53 prolongs NF-κB activation and promotes chronic inflammation and inflammation-associated colorectal cancer. Cancer Cell. 2013;23(5):634–646. | ||
Dupaul-Chicoine J, Yeretssian G, Doiron K, et al. Control of intestinal homeostasis, colitis, and colitis-associated colorectal cancer by the inflammatory caspases. Immunity. 2010;32(3):367–378. | ||
Ou J, Miao H, Ma Y, et al. Loss of abhd5 promotes colorectal tumor development and progression by inducing aerobic glycolysis and epithelial–mesenchymal transition. Cell Rep. 2014;9(5):1798–1811. | ||
Barker N, van Es JH, Kuipers J, et al. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature. 2007;449(7165):1003–1007. | ||
López de Las Hazas MC, Mosele JI, Macià A, Ludwig IA, Motilva MJ. Exploring the colonic metabolism of grape and strawberry anthocyanins and their in vitro apoptotic effects in HT-29 colon cancer cells. J Agric Food Chem. 2017;65(31):6477–6487. | ||
Zhong L, Zheng J, Sun Q, Wei K, Hu Y. Radix Tetrastigma hemsleyani flavone inhibits proliferation, migration, and invasion of human lung carcinoma A549 cells. Onco Targets Ther. 2016;9:635–641. | ||
Anastas JN, Moon RT. WNT signalling pathways as therapeutic targets in cancer. Nat Rev Cancer. 2013;13(1):11–26. | ||
Lei ZJ, Wang J, Xiao HL, et al. Lysine-specific demethylase 1 promotes the stemness and chemoresistance of Lgr5(+) liver cancer initiating cells by suppressing negative regulators of β-catenin signaling. Oncogene. 2015;34(24):3188–3198. | ||
Li VS, Ng SS, Boersema PJ, et al. Wnt signaling through inhibition of β-catenin degradation in an intact Axin1 complex. Cell. 2012;149(6):1245–1256. | ||
Rogler G. Chronic ulcerative colitis and colorectal cancer. Cancer Lett. 2014;345(2):235–241. | ||
Huch M, Dorrell C, Boj SF, et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature. 2013;494(7436):247–250. | ||
Qi L, Sun B, Liu Z, Cheng R, Li Y, Zhao X. Wnt3a expression is associated with epithelial–mesenchymal transition and promotes colon cancer progression. J Exp Clin Cancer Res. 2014;33(1):107. | ||
Leem SH, Li XJ, Park MH, Park BH, Kim SM. Genome-wide transcriptome analysis reveals inactivation of Wnt/β-catenin by 3,3′-diindolylmethane inhibiting proliferation of colon cancer cells. Int J Oncol. 2015;47(3):918–926. | ||
Hirsch D, Barker N, Mcneil N, et al. LGR5 positivity defines stem-like cells in colorectal cancer. Carcinogenesis. 2014;35(4):849–858. | ||
Schepers AG, Snippert HJ, Stange DE, et al. Lineage tracing reveals Lgr5+ stem cell activity in mouse intestinal adenomas. Science. 2012;337(6095):730–735. | ||
Wang H, Wang H, Makki MS, et al. Overexpression of β-catenin and cyclinD1 predicts a poor prognosis in ovarian serous carcinomas. Int J Clin Exp Pathol. 2014;7(1):264–271. | ||
Żuryń A, Litwiniec A, Safiejko-Mroczka B, et al. The effect of sulforaphane on the cell cycle, apoptosis and expression of cyclin D1 and p21 in the A549 non-small cell lung cancer cell line. Int J Oncol. 2016;48(6):2521–2533. | ||
Wu S, Guo Z, Hopkins CD, et al. Bis-cyclopropane analog of disorazole C1 is a microtubule-destabilizing agent active in ABCB1-overexpressing human colon cancer cells. Oncotarget. 2015;6(38):40866–40879. | ||
Davidson G, Niehrs C. Emerging links between CDK cell cycle regulators and Wnt signaling. Trends Cell Biol. 2010;20(8):453–460. | ||
Han XY, Wei B, Fang JF, et al. Epithelial–mesenchymal transition associates with maintenance of stemness in spheroid-derived stem-like colon cancer cells. PLoS One. 2013;8(9):e73341. | ||
Tsai JH, Yang J. Epithelial–mesenchymal plasticity in carcinoma metastasis. Genes Dev. 2013;27(20):2192–2206. | ||
Gujral TS, Chan M, Peshkin L, Sorger PK, Kirschner MW, Macbeath G. A noncanonical Frizzled2 pathway regulates epithelial–mesenchymal transition and metastasis. Cell. 2014;159(4):844–856. | ||
Choi YJ, Kim N, Chang H, et al. Helicobacter pylori-induced epithelial–mesenchymal transition, a potential role of gastric cancer initiation and an emergence of stem cells. Carcinogenesis. 2015;36(5):553–563. | ||
Liu CC, Cai DL, Sun F, et al. FERMT1 mediates epithelial–mesenchymal transition to promote colon cancer metastasis via modulation of β-catenin transcriptional activity. Oncogene. 2017;36(13):1779–1792. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.