Back to Journals » Infection and Drug Resistance » Volume 12
Quorum quenching activity of Bacillus cereus isolate 30b confers antipathogenic effects in Pseudomonas aeruginosa
Authors Raafat MM ![]() , Ali-Tammam M, Ali AE
, Ali-Tammam M, Ali AE
Received 6 August 2018
Accepted for publication 11 April 2019
Published 7 June 2019 Volume 2019:12 Pages 1583—1596
DOI https://doi.org/10.2147/IDR.S182889
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Sahil Khanna
Marwa M Raafat,1 Marwa Ali-Tammam,1 Amal E Ali1
1Department of Microbiology & Immunology, Faculty of Pharmaceutical Sciences & Pharmaceutical Industries, Future University in Egypt (FUE), New Cairo, Egypt
Background: Quorum quenching, the interference of a Quorum sensing (QS) system that contributes to the pathogenesis through triggering the production of various virulence determinants, is among the newly suggested antivirulence strategies.
Purpose: This study aimed at screening of N-Acyl homoserine lactonase activity from local bacterial isolate, and investigating its effect on Pseudomonas aeruginosa (P. aeruginosa) virulence and biofilm formation.
Materials and methods: Soil bacteria were screened for aiiA gene coding for lactonase enzyme by Polymerase Chain reaction and sequencing of aiiA gene homologs. Lactonase activity and spectrum were assessed in the cell-free lysate by well diffusion assay using Agrobacterium tumafaciens KYC55. A bacterial isolate showing the highest N-acyl-homoserine lactones degradation percentage was identified by gene amplification and sequencing of the 16S rRNA gene and its aiiA gene homolog. High performance liquid chromatography was used to confirm N-acyl-homoserine lactone degradation. The effect of cell-free lysate on the biofilm formation ability and cytotoxicity of P. aeruginosa PAO1 and P. aeruginosa clinical isolates from different clinical sources were assessed by static microtiter plate and viability assay, respectively
Results: Lactonase gene and activity were identified in three Bacillus spp. isolates. They showed broad catalytic activities against tested N-acyl-homoserine lactones. However, The lactonase activity in the cell-free lysate of isolate 30b showed the highest significant degradation percentage on all tested signals; N-butanoyl-L-homoserine lactone (71%), N-hexanoyl-l-homoserine lactone (100%), N-decanoyl-homoserine lactone (100%), N-(3-oxohexanoyl)-L-homoserine lactone (37.5%), N-(oxodecanoyl)-L-homoserine lactone (100%), and N-(3-oxododecanoyl)-L-homoserine lactone (100%). Alignment of the amino acid sequences of AiiA protein of isolate 30b showed 96% identity with Bacillus cereus (B. cereus) homologous lactonases in the GenBank database, and the isolate was designated as B. cereus isolate 30b. Cell-free lysate of B. cereus isolate 30b reduced biofilm formation significantly in 93% of P. aeruginosa isolates. The highest mean percentage of reduction in the biofilm was 86%. Moreover, the viability percentage of human lung carcinoma A549 cells infected by P. aeruginosa and treated with cell-free lysate of B. cereus isolate 30b increased up to 15%.
Conclusion: The results of this study highlight the potential of lactonases as a promising strategy to combat Pseudomonas aeruginosa virulence.
Keywords: lactonase, antivirulence, Pseudomonas aeruginosa, biofilm, cytotoxicity, quorum quenching, AHL
Introduction
Quorum sensing (QS) is a signaling system based on cell–to-cell communication mechanism which depends on cell population density in bacteria.1,2 This QS system regulates the expression of diverse physiological activities such as bioluminescence, antibiosis, and biofilm development.3–6 The QS system allows bacteria to compete and dominate the bacterial community in which they develop strategies to increase their chances for co-existence. The QS system monitoring is achieved via small signal molecules.7
There is a wide range of QS signals, but only two are the most widely studied: N-acyl–L- homoserine lactones (AHLs) produced by more than 70 species of Gram negative bacteria, which has been termed autoinducers, while Gram positive bacteria uses autoinducing peptides as signaling molecules.8–10 In Gram negative bacteria, the AHLs signals diffuse across the cell membrane and bind to regulatory proteins.
AHLs are synthesized in the bacteria by a member of the LuxI protein family, and diffuse outside the bacteria. When AHL concentration reaches a threshold value due to accumulation of AHL derived from each bacterial cell, the AHL receptor protein belonging to LuxR protein family binds and regulates the expression of many virulence genes.1,5,11
However, interfering with the microbial QS system by quorum quenching (QQ) has been considered as an important strategy for disease control, as it is based on shutting down the virulence expression in pathogenic bacteria rather than inhibiting cell growth,12–15 This has shown the potential to overcome drug toxicities and antibiotic resistance.
QQ is mediated by enzymes known as QQ enzymes.16 QQ enzymes have been identified in a number of bacteria as quorum quenchers since AHL-lactonase AiiA from Bacillus spp.was first identified.17
QQ enzymes have been classified according to their enzymatic mechanism into three major types: AHL lactonase, AHL acylase, and AHL oxidase and reductase.18 AHL lactonases encoded by the gene aiiA, which hydrolyze the lactone ring of AHL of Gram negative pathogens and are found in numerous Bacillus spp.19–22
P. aeruginosa has become an important cause of hospital-acquired infections such as; pneumonia, urinary tract infections, skin and soft tissue infections. The problem is aggravated by the appearance of multi-drug resistant strains leading to high mortality rates.23 In addition, P. aeruginosa forms biofilms in host tissues. Being protected by extracellular polymeric substances, biofilm-forming cells can evade the immune host response and are considered 1,000-fold more resistant to conventional antibiotics compared to planktonic cells.24 P. aeruginosa produces two AHLs: N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C12-HSL) and N-butanoyl-L-homoserine lactone (C4-HSL). These signals control hundreds of genes, including those involved in the production of extracellular virulence factors and biofilm formation.25,26 In addition to their role in the upregulation of the genes involved in the production of destructive virulence factors such as proteases and toxins that affect human cells,27 QS signals themselves can modulate the production of pro-inflammatory cytokines and induce apoptosis in human cells. Studies demonstrated that 3-oxo-C12-HSL inhibits the production of IL-12 and TNFα by LPs-activated macrophages.28
Accordingly, investigating new antivirulence and anti-biofilm strategies is of major importance in limiting P. aeruginosa infections. Although lactonase activity of many bacteria has been thoroughly studied, limited research was conducted on its antipathogenic effect on clinical isolates of P. aeruginosa.29
In this present study, we aimed at isolating QQ lactonase enzymes from Bacillus spp., examining the effect of cell-free lysate from three Bacillus spp. on a range of QS signals, including those produced by P. aeruginosa. The highest spectrum and percentage of AHLs-degradation activity was obtained with the cell-free lysate of B. cereus isolate 30b. Additionally, we confirmed the antibiofilm effect and reduced cytotoxicity of both reference and clinical isolates of P. aeruginosa treated with the cell-free lysate of B. cereus isolate 30b.
Materials and methods
Isolation of soil bacilli
Soil samples were collected from agricultural areas at different Egyptian governorates. Sampling was done at the subsurface, where about 10 grams of soil samples were suspended in 20 mL of sterile distilled water in sterile 50 mL falcon tubes. Soil suspensions were vortexed and placed in a water bath at 80°C for 30 minutes. Heat-treated suspensions were placed at room temperature for 2 hours, then were serially diluted to 10–1, 10–2 in sterile saline, and were streaked on brain heart agar (Oxoid) plates. Bacilli isolates with different morphology were picked up and isolated for pure colonies on brain heart agar (Oxoid) plates.30
Screening for AHL-lactonase – positive bacterial isolates
Bacterial isolates were screened for the presence of lactonase gene by Polymerase Chain reaction (PCR) amplification of aiiA homologs using the extracted genomic DNA of each isolate as a PCR template, Taq DNA polymerase (Promega) and primer pairs: aiiA-F1 (5ʹ-ACG TGG ATC CCG CAG GAT CCA TAT GAC AGT AAA GAA GCT-3ʹ), and aiiA-R1 (5ʹ-GCT GGT CGA CCG TCG ACT ATA TAT ATT CAG GGA A-3’).31 The thermal cycling conditions were carried out with modification according to Rajesh and Ravishankar Rai32 as follows: initial denaturation at 94°C for 45 seconds, followed by five cycles of 94°C for 45 seconds, 44°C for 45 seconds, 72°C for 1 minute, then followed by another 30 cycles of 94°C for 45 seconds, 60°C for 45 seconds, 72°C for 1 minute, and final extension at 72°C for 8 minutes. Genomic DNA obtained from Bacillus weihenstephanensis isolate P65 – producing lactonase previously identified by Sakr et al33 – was used as a positive control. Detection of the amplified products was done by agarose gel electrophoresis, and the expected size of DNA fragment was determined in comparison to the DNA ladder (GeneRuler 1 kb Thermo Fisher Scientific).
Preparation of crude cell- free lysate
Isolates were incubated in brain heart broth media (Oxoid) overnight at 30°C, with shaking at 180 rpm. Cultures were centrifuged at 6,000 rpm for 10 minutes. Cell pellets were collected and were washed 3 -times with 50 mM Tris buffer (pH 7). Cells were incubated in lysis buffer containing: 50 mM Tris buffer (pH 7), 200 mM NaCl, 5 mM MgCl2, 1 mg/mL lysozyme (Sigma-Aldrich),1 mM Phenyl methanesulfonyl fluoride (Sigma-Aldrich), and 1 mM Dithiothreitol (Sigma-Aldrich) at 4°C for 20 minutes.34 Cells were disrupted by sonication using a sonicator probe for 5×2 minute pulses at 60 Hz. Cell lysates were centrifuged at 13,000 rpm at 4°C, and supernatant was collected and filtered through a 0.22 µm sterile filter, then stored at −20°C. Protein content was determined by Bradford reagent (Sigma-Aldrich) using bovine serum albumin as standard.35
Confirmation of quorum quenching activity in cell-free lysate using AHL inactivation assay
The AHL inactivation assay was carried out by well diffusion method using A. tumefaciens KYC55 as a reporter strain. Agar plates were prepared as follows: 1 mL of overnight culture of KYC55 was added to 50 mL of AT minimal medium supplemented with Gentamycin (100 µg/mL) (Sigma- Aldrich), Tetracycline (4 µg/mL) (Sigma-Aldrich), Spectinomycin (100 µg/mL) (Sigma- Aldrich),36 and was grown at 28ºC and 160 rpm overnight and was poured into 100 ml AT minimal medium (1.2% agar) maintained at 46ºC. X-Gal (Biomatik, Canada) was added to a final concentration of 60 µg/ml. The agar-culture solution was immediately poured as 20 ml portions in petri dishes. Cell-free lysate of the isolates showing positive aiiA PCR products were adjusted to the same protein content of 1500 µg/mL, and were incubated for 3 hours with N-hexanoyl-
Assay of the spectrum of the AHLs degradation activity of the cell-free lysates
The AHLs degradation activity of cell-free lysates of Bacillus spp. isolates 30b, 32C, and 6 was assessed against different AHL signals using the well diffusion assay. AHL signals tested were; 100 µM C4-HSL (Cayman), 50 µM C6-HSL, 50 µM N- decanoyl-homoserine lactone (C10-HSL), 25 µM N-(3-oxohexanoyl)-L-homoserine lactone (3-oxo-C6-HSL), 25 µM N-(3-oxododecanoyl)-L-homoserine lactone (3-oxo-C10-HSL), 25 µM 3-oxo-C12-HSL (Sigma-Aldrich). The relative degradation activity was calculated for each AHL signal by calculating the residual AHL from a calibration curve constructed using different concentrations of standard AHL and their induced zone diameter using two AHLs reporter strains; Chromobacterium violaceum (C. violaceum) CV026 and A. tumafaciens KYC55, which respond to short chain and medium-to-long chain AHLs, respectively.39–42 The maximum degrading activity is defined as 100% relative degradation activity.43
Identification of bacterial isolate, sequencing of aiiA gene homolog, and phylogenetic analysis
Identification of the bacterial isolate possessing the most significant degradation percentage against different AHLs signals was done by 16S rRNA gene amplification and sequencing. Genomic DNA of the isolate was extracted using a DNA extraction kit (Qiagen). The 16S rRNA gene was amplified by PCR using universal primers 27F: (5‘-AGA GTT TGA TCC TGG CTC AG-3‘) and 1492R (5‘-ACG GCT ACC TTG TTA CGC TT- 3‘) by standard procedure.32 The purified PCR product (~1,426 bp) was sequenced, and the edited sequence data was assembled into a final consensus sequence. The obtained consensus was blasted in the nucleotide National Center for Biotechnology Information (NCBI) GenBank database and submitted into GenBank under accession number (MF671984). The PCR product of aiiA gene (~720 bp) of the same isolate was purified using QIA quick PCR purification kit (Qiagen) and was sequenced. The open reading frame of the lactonase enzyme was obtained using CLC Main Workbench 5 (CLC Bio, Aarhus, Denmark). Alignment of the amino acid sequence of the identified lactonase enzyme with AHL homologous lactonases retrieved from the NCBI database was done using CLC Main Workbench 5. The neighbor-joining method was used to construct the phylogenetic tree using the MEGA 5 software. Cell-free lysate was analyzed by SDS-PAGE and was visualized by Coomassie brilliant blue staining (Sigma-Aldrich).
High performance liquid chromatography (HPLC) analysis
HPLC analysis was carried out by a modified method of Wang et al.44To analyze AHL-degradation products, C6-HSL at a final concentration of 50 µM was mixed with an equal volume of the cell-free lysate of B. cereus strain 30b (adjusted to a protein content of 1500 µg/mL), and was incubated at 30°C for 3 hours. A negative control without the cell-free lysate containing final concentration of 50 µM C6-HSL in Tris buffer (pH 7) was also included. After incubation, the reaction was stopped by heating at 95°C for 10 minutes and the reaction mixture was centrifuged to pellet the precipitated protein. Then the reaction mixture was extracted with an equal volume of ethyl acetate (Sigma-Aldrich). The mixture was shaken vigorously for 30 seconds, and the phases were allowed to separate. The shaking was repeated three times before the ethyl acetate fraction was removed and another fraction was added. The whole extraction process was repeated three times. The combined ethyl acetate fractions were evaporated to dryness and reconstituted in 0.2 mL of acetonitrile (Sigma-Aldrich), nd chromatographed on HPLC system (Shimadzu class-VP) with a UV/Vis detector at 205 nm using an Agilent C18 column (4.6x250 mm). Samples were eluted isocratically with water-acetonitrile (50:50, pH 6.5) [vol/vol] at 1 mL/min. The amount of AHL was estimated by comparing the reduction in peak areas for a given retention time with a C6-HSL standard calibration curve.
P. aeruginosa isolates and culture conditions
A total of 33 clinical isolates of P. aeruginosa previously identified with API 20NE System (Biomerieux, France) were included in this study. They were selected from our culture collection based on the presence of one or more virulence factors, as described previously.45 (Table S1). The isolates had been taken from different specimens: Abscess (10), urine (10), sputum (6), wound (5), and stool (2). They were obtained from the bacteriology laboratory of Al-Azhar University and Kasr Al Ainy hospitals, Egypt. All experiments in this study using the clinical isolates were conducted in accordance with and approval of the ethical committee at the Faculty of Pharmaceutical Sciences & Pharmaceutical Industries, Future University in Egypt.
P. aeruginosa PAO1 ( ATCC 15692) and clinical isolates were cultured in Luria Bertani (LB) broth (Oxoid). The cultures were incubated at 37°C for 24 hours in a rotary shaker at 160 rpm.
Effect of cell-free lysate on P. aeruginosa biofilm
The biofilm-forming ability of P. aeruginosa clinical isolates and the anti-biofilm activity of cell-free lysate of B. cereus isolate 30b were assessed using static microtiter plate assay according to Cady et al,46 with slight modifications. P. aeruginosa PAO1 and clinical isolates were grown individually in LB broth for 18 hours at 37°C with a rotary shaker at 150 rpm. The overnight culture was resuspended in LB media to get approximately 1.5×107 CFU/mL (determined by optical density). Then 100 µL of culture along with 0.22 µm filter sterilized 100 µL cell-free lysate were dispensed into a 96-well microtiter plate. The assay controls were maintained without cell-free lysate treatment using Phosphate Buffered Saline (PBS). The plate was incubated at 37°C for 6 hours without agitation. The planktonic cells were removed and read at 600 nm. Wells were washed twice with 150 µL of PBS. Biofilms formed were detected by staining with 100 µL of crystal violet [0.2% (w/v), incubated for 15 minutes at room temperature and then washed four times with PBS. One hundred microliters of ethanol (95%) was used to remove crystal violet from the biofilm onto a new microtiter plate, and absorbance was measured at 590 nm. The ratio of the OD 590/OD 600 was used to normalize the amount of biofilm formed to the growth of bacteria in the presence and absence of the cell-free lysate.
Effect of cell-free lysate of B. cereus isolate 30b on P. aeruginosa cytotoxicity to human lung carcinoma A549 cells
The human lung carcinoma A549 cell line was purchased from Vacsera (Egypt). The cell line was cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum, 100 U/mL of penicillin, and 100 µg/mL of streptomycin, and incubated at 37°C with 5% CO2 till confluency. The cells were diluted to 2.0×105 cells/mL, and 100 µL aliquots were dispensed in a 96-well plate and incubated overnight. The cells were washed and were infected with P. aeruginosa PAO1 or clinical isolates at a concentration of 1×106 CFU/well or with a combination of non-cytotoxic dose of cell-free lysate of B. cereus isolate 30b (250 µg/mL protein) and P. aeruginosa PAO1/clinical isolates. Negative control wells with normal saline were also included. The cytotoxicity was assessed by measuring the amount of lactate dehydrogenase (LDH) from viable cells by treatment with 3- (4,5- Dimethylthiazol-2-yl)-2.5-Diphenyltetrazolium Bromide (MTT) dye (0.5 mg/mL) (Sigma-Aldrich) at 37°C for 4 hours, then MTT was discarded. Plates were washed three times with PBS. Dimethyl sulfoxide (BDH, England) was added as 50 μL/well. Plates were shaken for 30 minutes to dissolve the produced intracellular blue MTT-Formazan complex. Optical densities (ODs) were measured at 570 nm using an ELISA plate reader.47
Statistical analysis
All data were presented as the mean value of three independent measurements±standard deviation. GraphPad Prism (La Jolla, CA, USA) software was used to calculate the standard error and hypothesis testing to determine the level of significance. Significant differences between means were tested by unpaired t-test and one-way analysis of variance (ANOVA) with Tukey’s Multiple Comparison post-hoc test. Differences were considered significant at P˂0.05.
Results and discussion
Owing to the continuous emergence of resistant strains to commonly used antibiotics, there is a crucial need for exploring new therapeutic approaches. Lactonases are among the QQ enzymes that had received much more attention in the last few years as promising alternatives targeting AHL molecules of Gram negative pathogens without affecting cell growth or imposing selective pressure on microorganisms.48 They exert their effect by hydrolyzing the ester bond of AHLs, and give the corresponding AHL molecule,19,49 thus preventing the interaction between AHL and its cognate receptor protein and the subsequent expression of virulence phenotypes in pathogenic bacteria. The main focus of this study was to screen for AHL-lactonase activity from local Bacillus spp., as well as investigate its inhibitory effect on P. aeruginosa cytotoxicity and biofilm-forming ability.
Isolation of soil bacilli, screening for aiiA gene
Out of 97 soil samples collected from agricultural areas of different Egyptian governorates, 209 soil bacilli isolates were recovered. All isolates were screened for the presence of AHLs-lactonase (aiiA) homolog gene. The aiiA PCR product was detected in only three isolates (3/209; 1.43%).
Confirmation of AHL degradation activity by well diffusion assay
The AHL-lactonase activity of the cell-free lysate of aiiA-positive isolates was tested for their ability to degrade C6-HSL, using a sensitive indirect assay for AHL-lactonase activity. In this assay, we utilized a specific bacterial biosensor,50 A. tumafaciens KYC55 strain, which was reported to have high sensitivity and permit detection of a broad range of AHL derivatives using a well diffusion method. A. tumafaciens KYC55 produces blue coloration in response to C6-HSL. The cell-free lysates exhibiting quorum quenching activity degrade C6-HSL, which in turn inhibits the development of the blue color. Based on the size of the blue coloration around the wells, the three isolates showed different AHL lactonase degradation activity (Figure 1A). The cell-free lysate of isolates 32C and 6 showed only partial degradation of C6-HSL, as indicated by the blue halos around wells, whereas the cell-free lysate of isolate 30b showed complete degradation of C6-HSL. Since the hydrolytic cleavage of the homoserine lactone ring by AHL-lactonase is reversible and is similar to pH-mediated lactonolysis, the effect of acidification of the reaction products after incubation of C6-HSL was tested with the cell-free lysate. The appearance of the blue color around the wells confirms the presence of lactonase enzyme in cell-free extracts and excludes the possibility of the presence of acylase type-QQ enzymes (Figure 1B).12 The three isolates were subjected to subsequent assay to investigate their spectrum of AHLs degradation activity.
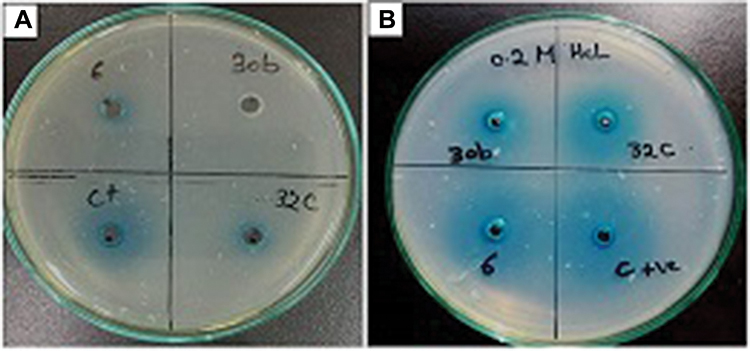 | Figure 1 (A) Confirmation of AHL degradation activity of the cell-free lysates of Bacillus sp. isolates 30b, 32c, 6 by well diffusion assay using A. tumefaciens KYC55. (B). Effect of acidification with 0.2M HCL on the reaction products.Abbreviation: AHL, N-acyl-L-homoserine lactone; A. tumefaciens, Agrobacterium tumefaciens. |
Assay of the spectrum of the AHLs degradation activity of the cell-free lysate
We expanded our analysis for the AHL degradation activity to a broader range of synthetic signals. The ability of the cell-free lysates of the three bacterial isolates to degrade different types of AHLs was studied. We used a detectable concentration for each AHL signal, which was between 25–100 µM, and this concentration was determined according to the sensitivity of the reporter strain towards different AHLs. In this study, all three isolates showed a broad substrate spectrum. All signals were significantly degraded by the cell-free lysate of the three isolates, except 3-oxo-C6-HSL, which was significantly reduced only by 30b cell-free lysate. By examination of the percentage degradation of all signals, significant difference in the mean degradation percentage was found between 30b and 32C or 6 cell-free lysates. In general, 30b cell-free lysate exhibited the highest degradation percentage of all tested signals which reached 100%, except for C4-HSL, which was 71%. However, 32C and 6 cell-free extract showed lower percentages of degradation against all signals (Figure 2). These results were in agreement with previous studies on lactonase enzyme from Bacillus spp. which reported that the AHL-lactonase AiiA from Bacillus exhibited the same enzymatic activity toward AHLs of variable length and irrespective to the nature of the substitution at the C3 position of the acyl chain.51,52 However, in other studies AiiA from Bacillus sp. 240B1 showed better activity against AHLs without 3-oxo substitutions than against the substituted derivatives.53 Such discrepancies could reflect strain-specific variations.
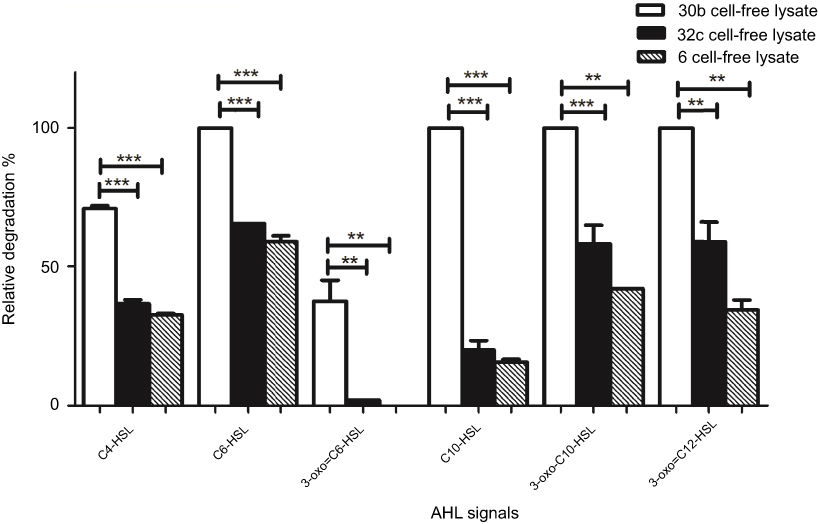 | Figure 2 AHL-degrading activity of cell-free lysates of Bacillus sp. isolates 30b, 32c, 6 against different substrates. The results are means of three independent experiments, and error bars show standard deviation.Notes: Results are the means of three independent experiments and the error bars show SD. (*) P<0.05, (**) p<0.001 and (***) p<0.001 according to student’s t-test.Abbreviation: AHL, N-acyl-L-homoserine lactone. |
In the current study, the increased catalytic efficiency of 30b cell-free lysate towards 3-oxo-C12-HSL, which is reported as the main signal that controls the expression of the subsequent QS circuit in P. aeruginosa,26,54,55 led us to investigate the effect of 30b cell-free extract on P. aeruginosa biofilm formation and cytotoxicity to the human cell line.
Identification of the bacterial isolate, sequencing of its aiiA gene, and phylogenetic analysis
The isolate was identified using 16S rRNA gene amplification and sequencing as B. cereus isolate 30b (accession number MF671984). Alignment of the nucleotide/amino acid sequence of the obtained lactonase from B. cereus isolate 30b (accession number MF979526) revealed 96% identity with B. cereus homologous in the GenBank database. The phylogenetic tree based on amino acid sequence showed its close clustering to other lactonases from different Bacillus spp. (Figure 3). In the same context, Huma et al31 reported that among 42 Bacillus strains possessing AHL-lactonase gene aiiA, 45% were found to be B. cereus; however, lactonase gene aiiA was also detected in various other Bacillus spp.20,21,56–58 Moreover, lactonase production has been reported in other microorganisms such as Agrobacterium,59 Rhodococcus,60 Streptomyces,61 Pseudomonas,62 Arthrobacter, and Klebsiella spp.63
 | Figure 3 Phylogenetic tree deriving from neighbor-joining based on amino acid sequence of AHL-lactonase showing the evolutionary relationship between B. cereus isolate 30b AHL-lactonase (accession MF979526) and the closet BLAST hits, represented by numbers written on branches. The tree was generated by Mega 5 software. Abbreviations: AHL, N-acyl-L-homoserine lactone; B. cereus, Bacillus cereus. |
HPLC analysis
To further confirm the lactonase activity in B. cereus 30b cell-free lysate, C6-HSL substrate was incubated with the cell-free lysate, and the reaction products were analyzed with HPLC. To exclude any alkaline lactonolysis due to pH, the pH of the mobile phase was adjusted at 6.5, and the pH of cell free lysate was 7. The negative control of C6-HSL without the cell-free lysate gave one major peak at a retention time of 3.3 minutes and a solvent peak at 1.7 minutes (Figure 4A). After 3 hours incubation of the cell-free lysate with C6-HSL, the HPLC profile of the reaction mixture revealed a peak at a retention time of 2.2 minutes, which might correspond to hydrolyzed products of C6-HSL and a small peak corresponding to the remaining C6-HSL at 3.5 minutes, in addition to the solvent peak at 1.8 minutes (Figure 4B). From the standard calibration curve, and the equation obtained having an R2=0.9991, the peak area of the remaining C6-HSL was equivalent to 2 µM C6-HSL, indicating that the degradation percentage reached 96% of 50 µM of C6-HSL after 3 hours of incubation.
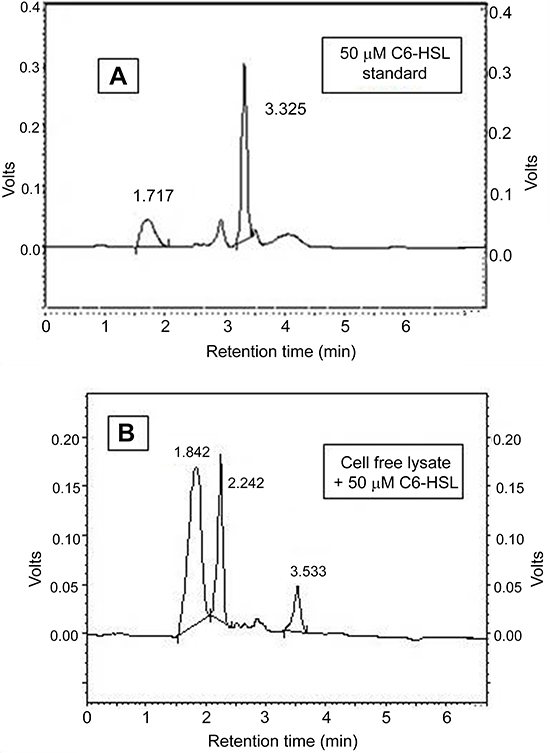 | Figure 4 HPLC profile. (A) Negative control C6-HSL, (B) AHL-degrading activity of cell free lysate of B. cereus isolate 30b against C6-HSL.Abbreviations: AHL, N-acyl-L-homoserine lactone; B. cereus, Bacillus cereus; HPLC, high performance liquid chromatography. |
Effect of cell-free lysate on P. aeruginosa biofilm-formation ability and cytotoxicity to human lung carcinoma A549 cells
The ability of P. aeruginosa to establish infections relies on the production of various QS-mediated virulence factors such as enzymes and toxins. In addition, P. aeruginosa forms biofilms that impair host defense and increases resistance to antimicrobial treatment. Three quorum systems are operating hierarchically in P. aeruginosa, LasR, rhlR, and PqsR systems with LasR operating at the top to control the expression of different QS genes and regulates the expression of virulence factors and biofilm.64 We investigated the effect of the 30b cell-free lysate on attenuation of the virulence of P. aeruginosa PAO1 and clinical isolates exhibiting different virulence phenotypes, by measuring its effect on biofilm formation and the cytotoxicity to human lung carcinoma A549 cells.
PAO1 was used as a positive control for biofilm formation. The isolates giving normalized OD values ≤ half of that of PAO1 were considered as biofilm-deficient.64 Among the clinical isolates investigated, 15 (46%) were biofilm forming and 18 (54%) were non-biofilm forming in the conditions tested (Figure 5A). The deficiency in biofilm-forming phenotype in some clinical strains of P. aeruginosa was also reported in previous studies.64,65 This finding might not reflect the loss of lasR or rhlR genes, as demonstrated by Wang et al,64 who reported that isolates deficient in biofilm formation carried intact lasR and rhlR genes, but exhibited some mutations and were different in their ability to produce QS-mediated virulence factors. A second study, however, reported the absence of all virulence factors in six P. aeruginosa isolates, yet still caused urinary tract infections.65 We examined the potential of cell-free lysate of B. cereus isolate 30b in inhibiting biofilm formation by P. aeruginosa PAO1 and biofilm-forming isolates by static microtiter plate method. To exclude any inhibitory effect of cell-free lysate on bacterial growth, planktonic cells, and biofilm cells were measured and normalized OD was compared. Biofilm was significantly reduced in eight clinical isolates and PAO1 by 72–86%, and by 47–70% in six clinical isolates, while it was non-significantly reduced in one isolate (Figure 5B). Although the process of biofilm formation in P. aeruginosa is a complex process and is regulated by QS system in addition to other signaling systems,66 significant reduction in biofilm formation by cell-free lysate might be the result of effective degradation of the produced AHLs.
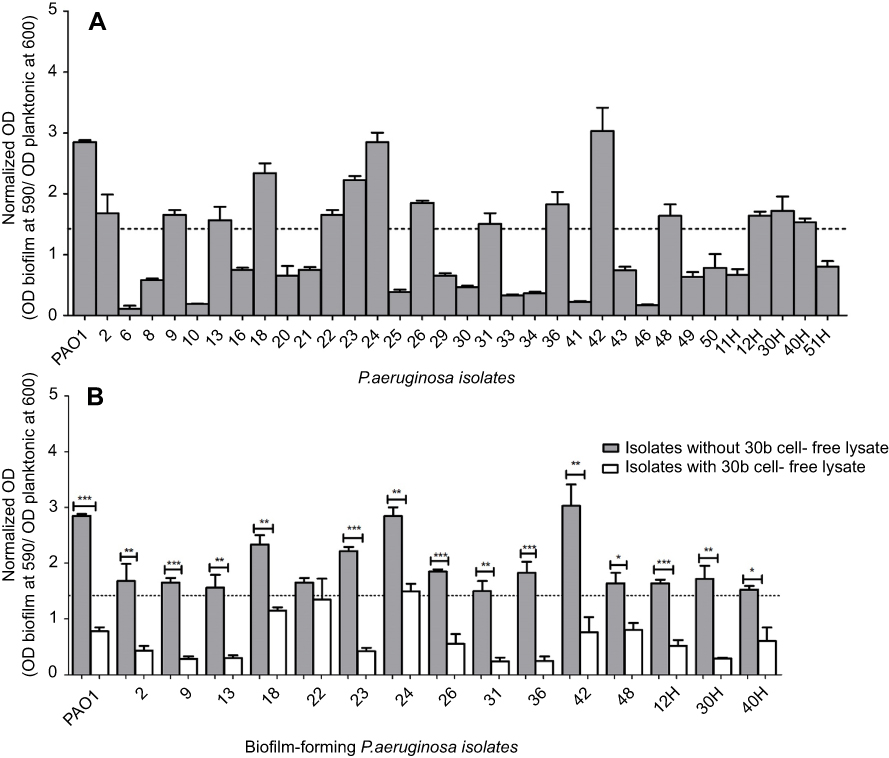 | Figure.5 (A) Biofilm formation ability of P. aeruginosa isolates. The break line indicates a normalized OD value ≤1/2 of that of PAO1. The isolates which had the normalized OD value ≤1/2 of that in PAO1 are defined as biofilm deficient. The results are means of three independent experiments, and error bars show standard deviation. (B) Biofilm activity of treated and untreated culture of each P. aeruginosa isolate. The results are means of three independent experiments, and error bars show standard deviation. Notes: Results are the means of three independent experiments and the error bars show SD. (*) P<0.05, (**) p<0.001 and (***) p<0.001 according to student’s t-test.Abbreviation: P. aeruginosa, Pseudamonas aeruginosa. |
The planktonic cells in cell lysate-treated P. aeruginosa PAO1 was slightly increased than in non-treated P. aeruginosa PAO1, thus indicating that cell free lysate targets AHLs degradation rather than growth inhibition and suggesting that no resistance mechanism could arise due to enzyme treatment. The increase in planktonic cells could be explained by the effect of AHL- lactonase in cell-free lysate to convert biofilm-forming cells to planktonic cells. Thus, this may increase the sensitivity to antibiotic treatment. These findings were in the same line with those of Kiran et al,67 who reported the transformation of (71.2%) of biofilm forms to planktonic forms due to lactonase (1 Unit) treatment of P. aeruginosa PA2 strain. The inhibitory effect of lactonases on virulence factors of P. aeruginosa was previously reported by Ng et al68, and Pei and Lamas-Samanamud.69 Hraiech et al70 reported that SsoPox-I, which is a hyperthermostable lactonase, exhibited in vitro inhibition of P. aeruginosa PAO1 pyocyanin synthesis, proteolytic activity, and biofilm formation, and further reduced the mortality of rats with acute pneumonia.
In this study, the human cancer cell line was used as a host model for assessment of the pathogenicity of P. aeruginosa. This approach has been used in previous studies.47,71,72
We selected representative biofilm and non-biofilm-forming isolates of a different virulence pattern for evaluation of their cytotoxic effect on human cell lines with and without treatment with cell-free lysate of B. cereus isolate 30b. The isolates tested were: 18, 22, 23, 50, 12H, 30H, 48H, 51H, and P. aeruginosa PAO1.
In our study, viability increased significantly in four P. aeruginosa isolates (50, 12H, 30H, 51H), by 7–15%, and in PAO1 (P<0.001, P<0.005) (Figure 6). These findings could be explained by the effect of lactonase enzyme that acts by disruption of the QS system. Our results were in agreement with those of Migiyama et al,47 who reported the reduction of cytotoxicity of P. aeruginosa PAO1 by half as a result of expression of AiiA lactonase, and attributed this reduction in cytotoxicity to the decreased accumulation of AHL signals and AHL-mediated virulence factors such as pyocyanin and elastase reported to cause cytotoxicity and tissue damage. However, our results showed that the cytotoxicity of two P. aeruginosa isolates (23 and 48 H) was unaffected by treatment with cell-free lysate, while it was increased in two isolates (18 and 22). These observations suggest that these strains might produce high concentrations of AHLs which couldn’t be counteracted by the low concentration of lactonase enzyme in the crude cell-free lysate. In the same context, Guendouze et al29 reported that resistance to lactonase enzyme may arise as a result of an increase in AHL production by bacterial cell, the presence of an enzyme inhibitor, or the bacteria modifying the AHL molecule to prevent its recognition by the enzyme.
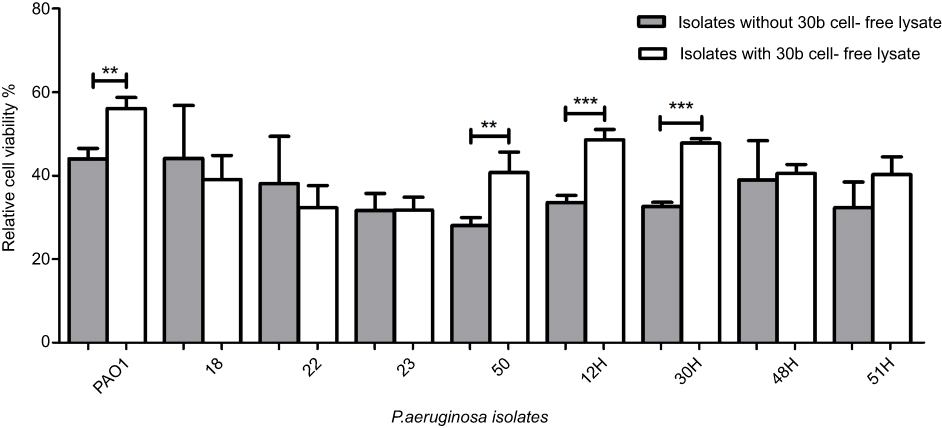 | Figure 6 Percentage viability of human lung carcinoma A549 cells after infection with treated and untreated culture for each P. aeruginosa isolate. The results are means of three independent experiments, and error bars show standard deviation.Notes: Results are the means of three independent experiments and the error bars show SD. (*) P<0.05, (**) p<0.001 and (***) p<0.001 according to student’s t-test.Abbreviation: P. aeruginosa, Pseudomonas aeruginosa. |
In conclusion, cell-free lysate of B. cereus strain 30b demonstrated AHL-lactonase activity against a wide range of AHLs signals. Moreover, it decreased the cytotoxicity of P. aeruginosa to the human lung carcinoma cell line by 7–15%, and exhibited significant biofilm inhibitory activity of 47–86%. These results emphasize the potential use of respective lactonase enzyme as antipathogenic therapy, particularly against clinically relevant pathogens such as P. aeruginosa. Work is in progress to purify the lactonase enzyme from the cell-free lysate. Further studies on the purified enzyme should be extended on a larger number of clinical isolates, on the effect of the purified enzyme on different virulence factors of P. aeruginosa, and on its possible use as a combined therapy for treatment of antibiotic-resistant strains of P. aeruginosa.
Data availability
The figures used to support the findings of this study are included within the article. DNA sequences generated from this study were deposited at GenBank with the accession numbers listed in the article.
Acknowledgment
This work was supported by the Academy of Scientific Research & Technology (JESOR Program-2015), Egypt.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Diggle SP, Griffin AS, Campbell GS, West SA. Cooperation and conflict in quorum-sensing bacterial populations. Nature. 2007;450(7168):411–414. doi:10.1038/nature06279
2. de Kievit TR, Iglewski BH. Bacterial quorum sensing in pathogenic relationships. Infect Immun. 2000;68(9):4839–4849.
3. Schuster M, Sexton DJ, Diggle SP, Greenberg EP. Acyl-homoserine lactone quorum sensing: from evolution to application. Annu Rev Microbiol. 2013;67:43–63. doi:10.1146/annurev-micro-092412-155635
4. Whitehead NA, Barnard AM, Slater H, Simpson NJ, Salmond GP. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol Rev. 2001;25(4):365–404. doi:10.1111/j.1574-6976.2001.tb00583.x
5. Hawver LA, Jung SA, Ng W-L. Specificity and complexity in bacterial quorum-sensing systems. FEMS Microbiol Rev. 2016;40(5):738–752. doi:10.1093/femsre/fuw014
6. Rutherford ST, Bassler BL. Bacterial quorum sensing: its role in virulence and possibilities for its control. Cold Spring Harb Perspect Med. 2012;2(11):a012427–a012427. doi:10.1101/cshperspect.a012427
7. Camilli A, Bassler BL. Bacterial small-molecule signaling pathways. Science. 2006;311(5764):1113–1116. doi:10.1126/science.1121357
8. Pedroza CJ, Flórez AM, Ruiz OS, Orduz S. Enzymatic hydrolysis of molecules associated with bacterial quorum sensing using an acyl homoserine lactonase from a novel Bacillus thuringiensis strain. Antonie Van Leeuwenhoek. 2014;105(1):253–264. doi:10.1007/s10482-013-0072-5
9. Papenfort K, Bassler BL. Quorum sensing signal-response systems in Gram-negative bacteria. Nat Rev Microbiol. 2016;14(9):576–588. doi:10.1038/nrmicro.2016.89
10. Monnet V, Juillard V, Gardan R. Peptide conversations in Gram-positive bacteria. Crit Rev Microbiol. 2016;42(3):339–351. doi:10.3109/1040841X.2014.948804
11. Morohoshi T, Kato M, Fukamachi K, Kato N, Ikeda T. N-acylhomoserine lactone regulates violacein production in chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol Lett. 2008;279(1):124–130. doi:10.1111/j.1574-6968.2007.01016.x
12. Chen F, Gao Y, Chen X, Yu Z, Li X. Quorum quenching enzymes and their application in degrading signal molecules to block quorum sensing-dependent infection. Int J Mol Sci. 2013;14(9):17477–17500. doi:10.3390/ijms140917477
13. Allen RC, Popat R, Diggle SP, Brown SP. Targeting virulence: can we make evolution-proof drugs? Nat Rev Microbiol. 2014;12(4):300–308. doi:10.1038/nrmicro3232
14. Whiteley M, Diggle SP, Greenberg EP. Progress in and promise of bacterial quorum sensing research. Nature. 2017;551(7680):313–320. doi:10.1038/nature24624
15. Defoirdt T. Quorum-sensing systems as targets for antivirulence therapy. Trends Microbiol. 2018;26(4):313–328. doi:10.1016/j.tim.2017.10.005
16. Rasmussen TB, Givskov M. Quorum-sensing inhibitors as anti-pathogenic drugs. Int J Med Microbiol IJMM. 2006;296(2–3):149–161. doi:10.1016/j.ijmm.2006.02.005
17. Li X, Du G, Chen J. Use of enzymatic biodegradation for protection of plant against microbial disease. Biotechnology. 2008;4:1–12.
18. Lin Y-H, Xu J-L, Hu J, et al. Acyl-homoserine lactone acylase from ralstonia strain XJ12B represents a novel and potent class of quorum-quenching enzymes. Mol Microbiol. 2003;47(3):849–860.
19. Dong YH, Wang LH, Xu JL, Zhang HB, Zhang XF, Zhang LH. Quenching quorum-sensing-dependent bacterial infection by an N-acyl homoserine lactonase. Nature. 2001;411(6839):813–817. doi:10.1038/35081101
20. Dong Y-H, Gusti AR, Zhang Q, Xu J-L, Zhang L-H. Identification of quorum-quenching N-acyl homoserine lactonases from bacillus species. Appl Environ Microbiol. 2002;68(4):1754–1759.
21. Lee SJ, Park S-Y, Lee -J-J, Yum D-Y, Koo B-T, Lee J-K. Genes encoding the N-acyl homoserine lactone-degrading enzyme are widespread in many subspecies of Bacillus thuringiensis. Appl Environ Microbiol. 2002;68(8):3919–3924.
22. Easwaran N, Karthikeyan S, Sridharan B, Gothandam KM. Identification and analysis of the salt tolerant property of AHL lactonase (AiiATSAWB) of Bacillus species. J Basic Microbiol. 2015;55(5):579–590. doi:10.1002/jobm.201400013
23. Bassetti M, Vena A, Croxatto A, Righi E, Guery B. How to manage pseudomonas aeruginosa infections. Drugs Context. 2018;7:212527. doi:10.7573/dic.212527
24. Luo J, Dong B, Wang K, et al. Baicalin inhibits biofilm formation, attenuates the quorum sensing-controlled virulence and enhances Pseudomonas aeruginosa clearance in a mouse peritoneal implant infection model. PLoS One. 2017;12(4):e0176883. doi:10.1371/journal.pone.0176883
25. Smith RS, Iglewski BH. P. aeruginosa quorum-sensing systems and virulence. Curr Opin Microbiol. 2003;6(1):56–60.
26. Donabedian H. Quorum sensing and its relevance to infectious diseases. J Infect. 2003;46(4):207–214.
27. Moradali MF, Ghods S, Rehm BHA. Pseudomonas aeruginosa lifestyle: a paradigm for adaptation, survival, and persistence. Front Cell Infect Microbiol. 2017;7:39. doi:10.3389/fcimb.2017.00039
28. Martínez JL. Interkingdom signaling and its consequences for human health. Virulence. 2014;5(2):243–244. doi:10.4161/viru.28073
29. Guendouze A, Plener L, Bzdrenga J, et al. Effect of quorum quenching lactonase in clinical isolates of pseudomonas aeruginosa and comparison with quorum sensing inhibitors. Front Microbiol. 2017;8:227. doi:10.3389/fmicb.2017.00227
30. Ng K-GC. Rapid isolation method of soil bacilli and screening of their quorum quenching activity. Asia Pac J Mol Biol Biotechnol. 2007;15(3):153–156.
31. Huma N, Shankar P, Kushwah J, et al. Diversity and polymorphism in AHL-lactonase gene (aiiA) of Bacillus. J Microbiol Biotechnol. 2011;21(10):1001–1011.
32. Rajesh PS, Ravishankar Rai V. Quorum quenching activity in cell-free lysate of endophytic bacteria isolated from pterocarpus santalinus linn., and its effect on quorum sensing regulated biofilm in pseudomonas aeruginosa PAO1. Microbiol Res. 2014;169(7–8):561–569. doi:10.1016/j.micres.2013.10.005
33. Sakr MM, Aboshanab KMA, Aboulwafa MM, Hassouna NA-H. Characterization and complete sequence of lactonase enzyme from Bacillus weihenstephanensis isolate P65 with potential activity against acyl homoserine lactone signal molecules. Biomed Res Int. 2013;2013:192589. doi:10.1155/2013/192589
34. Straight PD, Fischbach MA, Walsh CT, Rudner DZ, Kolter R. A singular enzymatic megacomplex from Bacillus subtilis. Proc Natl Acad Sci U S A. 2007;104(1):305–310. doi:10.1073/pnas.0609073103
35. Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254.
36. Tempé J, Petit A, Holsters M, Montagu M, Schell J. Thermosensitive step associated with transfer of the Ti plasmid during conjugation: possible relation to transformation in crown gall. Proc Natl Acad Sci U S A. 1977;74(7):2848–2849.
37. Ghani NA, Norizan SNM, Chan XY, Yin W-F, Chan K-G. Labrenzia sp. BM1: a quorum quenching bacterium that degrades N-acyl homoserine lactones via lactonase activity. Sensors. 2014;14(7):11760–11769. doi:10.3390/s140711760
38. Ravn L, Christensen AB, Molin S, Givskov M, Gram L. Methods for detecting acylated homoserine lactones produced by Gram-negative bacteria and their application in studies of AHL-production kinetics. J Microbiol Methods. 2001;44(3):239–251.
39. McLean RJC, Pierson LS, Fuqua C. A simple screening protocol for the identification of quorum signal antagonists. J Microbiol Methods. 2004;58(3):351–360. doi:10.1016/j.mimet.2004.04.016
40. Chen R, Zhou Z, Cao Y, Bai Y, Yao B. High yield expression of an AHL-lactonase from Bacillus sp. B546 in pichia pastoris and its application to reduce aeromonas hydrophila mortality in aquaculture. Microb Cell Fact. 2010;93:39. doi:10.1186/1475-2859-9-39
41. O’Connor G, Knecht LD, Salgado N, Strobel S, Pasini P, Daunert S. Whole-cell biosensors as tools for the detection of quorum-sensing molecules: uses in diagnostics and the investigation of the quorum-sensing mechanism. Adv Biochem Eng Biotechnol. 2015. doi:10.1007/10_2015_337
42. Quorum sensing vs quorum quenching: a battle with no end in sight | vipin Chandra Kalia | springer. Available from:
43. Seo M-J, Lee B-S, Pyun Y-R, Park H. Isolation and characterization of N-acylhomoserine lactonase from the thermophilic bacterium, geobacillus caldoxylosilyticus YS-8. Biosci Biotechnol Biochem. 2011;75(9):1789–1795. doi:10.1271/bbb.110322
44. Wang W-Z, Morohoshi T, Ikenoya M, Someya N, Ikeda T. AiiM, a novel class of N-acylhomoserine lactonase from the leaf-associated bacterium microbacterium testaceum. Appl Environ Microbiol. 2010;76(8):2524–2530. doi:10.1128/AEM.02738-09
45. Raafat MM, Ali- Tammam M, Ali AE. Phenotypic and genotypic characterization of pseudomonas aeruginosa isolates from Egyptian hospitals. Afr J Microbiol Res. 2016;10(39):1645–1653. doi:10.5897/AJMR2016.8254
46. Cady NC, McKean KA, Behnke J, et al. Inhibition of biofilm formation, quorum sensing and infection in pseudomonas aeruginosa by natural products-inspired organosulfur compounds. PLoS One. 2012;7(6):e38492. doi:10.1371/journal.pone.0038492
47. Migiyama Y, Kaneko Y, Yanagihara K, et al. Efficacy of AiiM, an N-acylhomoserine lactonase, against pseudomonas aeruginosa in a mouse model of acute pneumonia. Antimicrob Agents Chemother. 2013;57(8):3653–3658. doi:10.1128/AAC.00456-13
48. Tay SB, Chow JY, Go MK, Yew WS. Anti-virulent disruption of pathogenic biofilms using engineered quorum-quenching lactonases. J Vis Exp JoVE. 2016;(107). doi:10.3791/53243
49. Dong YH, Xu JL, Li XZ, Zhang LH. AiiA, an enzyme that inactivates the acylhomoserine lactone quorum-sensing signal and attenuates the virulence of Erwinia carotovora. Proc Natl Acad Sci U S A. 2000;97(7):3526–3531. doi:10.1073/pnas.060023897
50. Steindler L, Venturi V. Detection of quorum-sensing N-acyl homoserine lactone signal molecules by bacterial biosensors. FEMS Microbiol Lett. 2007;266(1):1–9. doi:10.1111/j.1574-6968.2006.00501.x
51. Liu D, Momb J, Thomas PW, et al. Mechanism of the quorum-quenching lactonase (AiiA) from Bacillus thuringiensis. 1. Product-bound structures. Biochemistry (Mosc). 2008;47(29):7706–7714. doi:10.1021/bi800368y
52. Kalia VC, Raju SC, Purohit HJ. Genomic analysis reveals versatile organisms for quorum quenching enzymes: acyl-homoserine lactone-acylase and -lactonase. Open Microbiol J. 2011;5:1–13. doi:10.2174/1874285801105010001
53. Wang L-H, Weng L-X, Dong Y-H, Zhang L-H. Specificity and enzyme kinetics of the quorum-quenching N-Acyl homoserine lactone lactonase (AHL-lactonase). J Biol Chem. 2004;279(14):13645–13651. doi:10.1074/jbc.M311194200
54. Wagner VE, Bushnell D, Passador L, Brooks AI, Iglewski BH. Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J Bacteriol. 2003;185(7):2080–2095.
55. Chun CK, Ozer EA, Welsh MJ, Zabner J, Greenberg EP. Inactivation of a Pseudomonas aeruginosa quorum-sensing signal by human airway epithelia. Proc Natl Acad Sci U S A. 2004;101(10):3587–3590. doi:10.1073/pnas.0308750101
56. Ishida T, Ikeda T, Takiguchi N, Kuroda A, Ohtake H, Kato J. Inhibition of quorum sensing in pseudomonas aeruginosa by N-acyl cyclopentylamides. Appl Environ Microbiol. 2007;73(10):3183–3188. doi:10.1128/AEM.02233-06
57. Augustine N, Kumar P, Thomas S. Inhibition of vibrio cholerae biofilm by AiiA enzyme produced from Bacillus spp. Arch Microbiol. 2010;192(12):1019–1022. doi:10.1007/s00203-010-0633-1
58. Molina L, Constantinescu F, Michel L, Reimmann C, Duffy B, Défago G. Degradation of pathogen quorum-sensing molecules by soil bacteria: a preventive and curative biological control mechanism. FEMS Microbiol Ecol. 2003;45(1):71–81. doi:10.1016/S0168-6496(03)00125-9
59. Carlier A, Uroz S, Smadja B, et al. The Ti plasmid of Agrobacterium tumefaciens harbors an attM-paralogous gene, aiiB, also encoding N-Acyl homoserine lactonase activity. Appl Environ Microbiol. 2003;69(8):4989–4993.
60. Park S-Y, Hwang B-J, Shin M-H, Kim J-A, Kim H-K, Lee J-K. N-acylhomoserine lactonase producing rhodococcus spp. with different AHL-degrading activities. FEMS Microbiol Lett. 2006;261(1):102–108. doi:10.1111/j.1574-6968.2006.00336.x
61. Park S-Y, Kang H-O, Jang H-S, Lee J-K, Koo B-T, Yum D-Y. Identification of extracellular N-acylhomoserine lactone acylase from a Streptomyces sp. and its application to quorum quenching. Appl Environ Microbiol. 2005;71(5):2632–2641. doi:10.1128/AEM.71.5.2632-2641.2005
62. Sio CF, Otten LG, Cool RH, et al. Quorum quenching by an N-acyl-homoserine lactone acylase from pseudomonas aeruginosa PAO1. Infect Immun. 2006;74(3):1673–1682. doi:10.1128/IAI.74.3.1673-1682.2006
63. Park S-Y, Lee SJ, Oh T-K, et al. AhlD, an N-acylhomoserine lactonase in arthrobacter sp., and predicted homologues in other bacteria. Microbiol Read Engl. 2003;149(Pt6):1541–1550. doi:10.1099/mic.0.26269-0
64. Wang Y, Gao L, Rao X, et al. Characterization of lasR-deficient clinical isolates of pseudomonas aeruginosa. Sci Rep. 2018;8(1):13344. doi:10.1038/s41598-018-30813-y
65. Senturk S, Ulusoy S, Bosgelmez-Tinaz G, Yagci A. Quorum sensing and virulence of pseudomonas aeruginosa during urinary tract infections. J Infect Dev Ctries. 2012;6(6):501–507.
66. Turkina MV, Vikström E. Bacteria-host crosstalk: sensing of the quorum in the context of pseudomonas aeruginosa infections. J Innate Immun. 2018;1–17. doi:10.1159/000494069
67. Kiran S, Sharma P, Harjai K, Capalash N. Enzymatic quorum quenching increases antibiotic susceptibility of multidrug resistant pseudomonas aeruginosa. Iran J Microbiol. 2011;3(1):1–12.
68. Ng FSW, Wright DM, Seah SYK. Characterization of a phosphotriesterase-like lactonase from sulfolobus solfataricus and its immobilization for disruption of quorum sensing. Appl Environ Microbiol. 2011;77(4):1181–1186. doi:10.1128/AEM.01642-10
69. Pei R, Lamas-Samanamud GR. Inhibition of biofilm formation by T7 bacteriophages producing quorum-quenching enzymes. Appl Environ Microbiol. 2014;80(17):5340–5348. doi:10.1128/AEM.01434-14
70. Hraiech S, Hiblot J, Lafleur J, et al. Inhaled lactonase reduces Pseudomonas aeruginosa quorum sensing and mortality in rat pneumonia. PLoS One. 2014;9(10):e107125. doi:10.1371/journal.pone.0107125
71. Ichikawa JK, Norris A, Bangera MG, et al. Interaction of pseudomonas aeruginosa with epithelial cells: identification of differentially regulated genes by expression microarray analysis of human cDNAs. Proc Natl Acad Sci U S A. 2000;97(17):9659–9664. doi:10.1073/pnas.160140297
72. Lee K-M, Yoon MY, Park Y, Lee J-H, Yoon SS. Anaerobiosis-induced loss of cytotoxicity is due to inactivation of quorum sensing in pseudomonas aeruginosa. Infect Immun. 2011;79(7):2792–2800. doi:10.1128/IAI.01361-10
Supplementary material
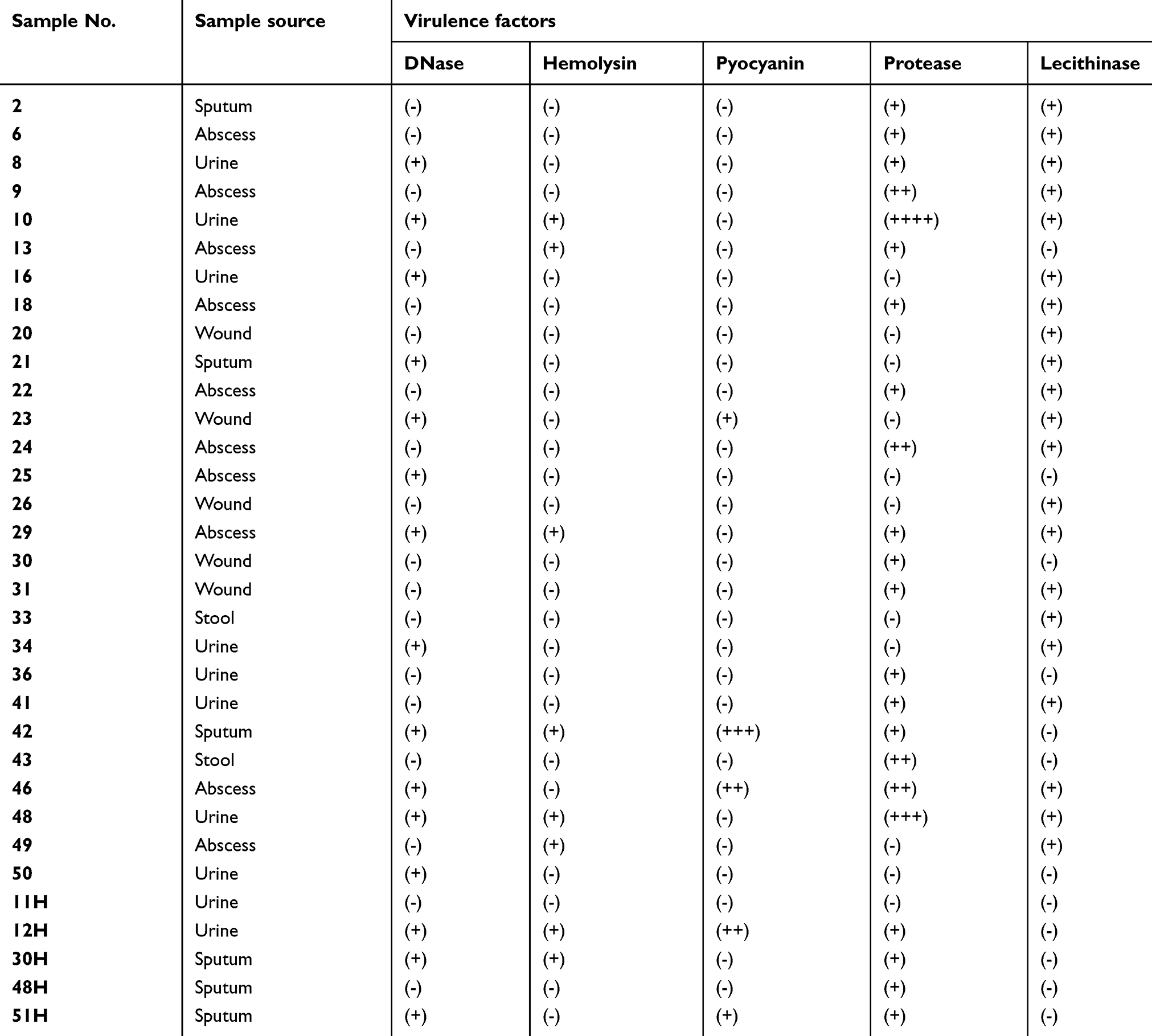 | Table S1 Clinical isolates of P. aeruginosa used in this study with their sources and identified virulence factors |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.