Back to Journals » Infection and Drug Resistance » Volume 12
Quinolone resistance mechanisms among third-generation cephalosporin resistant isolates of Enterobacter spp. in a Bulgarian university hospital
Authors Markovska R ![]() , Stoeva T
, Stoeva T ![]() , Dimitrova D
, Dimitrova D ![]() , Boyanova L
, Boyanova L ![]() , Stankova P
, Stankova P ![]() , Mihova K, Mitov I
, Mihova K, Mitov I ![]()
Received 25 February 2019
Accepted for publication 7 April 2019
Published 28 May 2019 Volume 2019:12 Pages 1445—1455
DOI https://doi.org/10.2147/IDR.S204199
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sahil Khanna
Rumyana Markovska,1 Temenuga Stoeva,2 Dobromira Dimitrova,2 Lyudmila Boyanova,1 Petya Stankova,1 Kalina Mihova,3 Ivan Mitov1
1Department of Medical Microbiology, Medical University of Sofia, Sofia, Bulgaria; 2Department of Microbiology, University Hospital “Saint Marina”, Medical University, Varna, Bulgaria; 3Molecular Medicine Center, Medical University of Sofia, Sofia, Bulgaria
Background: There have been no reports in Bulgaria about quinolone resistance determinants among Enterobacter spp.
Aims: To investigate plasmid and chromosomal quinolone resistance rates among 175 third-generation cephalosporin resistant Enterobacter spp. isolates (167 Enterobacter cloacae complex and eight Enterobacter aerogenes isolates) collected at a university hospital in Varna, Bulgaria, as well as to reveal their association with ESBL/AmpC production and a carriage of specific plasmid replicon types.
Methods: PCR, isoelectric focusing, replicon typing, sequencing, and epidemiology typing were carried out.
Results: A high level of combined third-generation cephalosporin and quinolone resistant Enterobacter spp. was found − 79.4%. The ESBL production rate was 87%, consisting mainly of CTX-M-15 among E. cloacae complex (in 76%) and CTX-M-3 among E. aerogenes (in 88%). Plasmid mediated quinolone resistance (PMQR) determinants were identified in 57% of the isolates. The most commonly detected PMQR determinants were qnrB (90%), consisting mainly of qnrB1 (in 61%), and qnrB9 (in 27%) of the isolates. Both alleles were transferred with CTX-M-15 genes; transconjugants showed HI2 replicons (for qnrB1 positive transconjugants) and were non-typeable (for qnrB9). One Enterobacter spp. isolate produced qnrB4. QnrA1, qnrS1, and aac(6ʹ)-Ib-cr were detected in single isolates only. QnrC, qnrD, qepA, and oqxAB genes were not found. QnrB was associated with CTX-M-15 production, and qnrS1 was linked to CTX-M-3. Alterations in 83 and 87 positions of gyrB in quinolone-resistance determining regions, and 80 position of parC were detected in high level quinolone resistant isolates. Among all the Enterobacter spp. isolates tested, one predominant clone A was identified (53%).
Conclusion: Our data showed the necessity of more prudent use of quinolones and third-generation cephalosporins, because of the risk of promoting dissemination, and selection of multiple resistance determinants (ESBL, PMQR) among Enterobacter spp. isolates in Bulgaria.
Keywords: quinolone resistance, Enterobacter spp., PMQR, Bulgaria
Introduction
Enterobacter spp. is а common cause of hospital infections such as urinary tract, blood-stream, lower respiratory tract, and soft tissues.1 These organisms have been associated with treatment failure and high mortality rate, mainly in patents admitted to intensive care units.2 Recently, a WHO expert group defined a Priority Pathogen List for research and development of new antimicrobials active against multidrug- and extensively drug-resistant Gram-negative bacteria. Third-generation cephalosporin resistant Enterobacter spp. were included in the first priority list, named “Priority 1: Critical”, which shows their significance.3 The treatment of infections associated with Enterobacter spp. is difficult due to overexpression of chromosomal AmpC enzymes, as well as production of ESBL (most commonly CTX-M, but also SHV and TEM).1 During the last decade, increasing numbers of carbapenemase producing Enterobacter isolates have been reported.4 Quinolones and aminoglycosides are treatment alternatives for infections caused by ESBL/AmpC or carbapenemase producing Enterobacter spp. susceptible to these antimicrobials.6 The quinolones such as ciprofloxacin and especially levofloxacin and moxifloxacin possess a broad spectrum of activity and can be used in a wide range of infections. The increased inappropriate usage of quinolones has led to emergence of chromosomal point mutations in the quinolone-resistance determining regions (QRDR) (gyrA and/or parC genes), which encode the subunits of gyrase and topoisomerase IV enzymes.5,6 Over the last decades, plasmid mediated quinolone resistance (PMQR) mechanisms have appeared. They perform target protection (gyrase and type IV topoisomerases) by qnr family proteins, enzymatic target modification by the aminoglycoside modifying enzyme aac(6ʹ)Ib-cr and by efflux mechanisms (qepA and oqxAB).5–7 In Bulgaria, there have been no reports about quinolone resistance determinants in Enterobacter spp.
The aim of this study was to determine the plasmid and chromosomal mechanisms of quinolone resistance in Enterobacter isolates resistant to third-generation cephalosporins from the University Hospital “Saint Marina” - Varna, Bulgaria, as well as to assess their association with ESBL/AmpC presence and plasmid replicon types.
Materials and methods
Bacterial isolates
A total of 175 isolates of Enterobacter spp. resistant to at least one third-generation cephalosporin were collected between January 2014 and January 2017 from hospitalized patients in the University Multiprofile Hospital for Active Treatment (UMHAT) “Saint Marina”-Varna during routine hospital laboratory work. Species' identification was done by Phoenix (BD, Franklin Lakes, NJ, USA). A previously published DHA-1 producing Enterobacter spp. isolate were also included in this study.8
Antimicrobial susceptibility testing
The antimicrobial susceptibility was determined by the disc diffusion method according to EUCAST, 2017 guidelines (http://www.eucast.org/clinical_breakpoints/). The following antimicrobial agents were tested: amoxicillin-clavulanate, cefotaxime, ceftazidime, cefepime, piperacillin-tazobactam, imipenem, meropenem, tobramycin, gentamicin, amikacin, trimethoprim/sulfamethoxazole (Oxoid, Basingstoke, UK). MICs of nalidixic acid, ciprofloxacin, and levofloxacin were determined by MIC strips (Liofilchem, Roseto degli Abruzzi, Italy).
Transfer of resistance determinants and plasmid replicon typing
Conjugative plasmid transfer was performed on BBL™ Mueller-Hinton agar II (Becton Dickinson, Sparks, MD, USA) using Escherichia coli K12:W3110 lac− resistant to rifampicin as recipient. Transconjugants were selected on MacConkey agar (Becton Dickinson, Sparks, MD, USA) containing 50 mg/L rifampicin and 2 mg/L cefotaxime. Plasmid replicon types were determined by the PCR-based replicon typing scheme described by Carattoli et al,9 using whole-cell DNA extracted from transconjugants. The replicon types were detected using primers described by Garcia-Fernandes et al.10
PCR detection and sequencing of ESBL and AmpC genes
PCR was performed to detect the presence of blaSHV and blaCTX-M genes as previously described.11 Plasmid-mediated AmpC genes –blaCMY, blaFOX, blaDHA, and blaAAC were screened as described by Perez-Perez et al.12 For sequencing of entire genes, primers binding outside the coding region were used for blaSHV, blaCTX-M-1-group,11 and blaDHA13 The amplicons were purified and sequenced with ABI 3130xl Genetic Analyzer. The nucleotide and deduced amino acid sequences were analyzed and multiple alignments were performed using Chromas Lite 2.01 (Technelysium Pty Ltd, Brisbane, Australia) and DNAMAN version 8.0 Software (Lynnon BioSoft, Vaudreuil-Dorion, Canada).
Isoelectric focusing (IEF) and bioassay
Beta-lactamase production of representative SHV and CTX-M positive isolates and all CTX-M and SHV negative isolates was analyzed by IEF as described previously.11 The isolates with known beta-lactamases (TEM-1, OXA-1, SHV-3, SHV-12, CTX-M-3, and CTX-M-15) were used as controls. The hydrolytic activity of individual beta-lactamase bands was assessed by bioassay.11 Two consecutive agar overlays were laid on the gel: first 0.5% tryptic soy agar (Becton Dickinson) containing the respective beta-lactam (cefotaxime 2 mg/L or ceftazidime 1 mg/L), followed (after 2hours of incubation at 35°C) by a second tryptic soy agar overlay containing the susceptible indicator strain E. coli K12:W3110 (RifR lac-, 1.2×107 CFU/mL). After an overnight incubation at 35°C, the growth of the indicator strain on the gel was determined the bands with hydrolytic activity.
Epidemiological typing
Whole-cell DNA was prepared by the GFX Genomic DNA Purification Kit (GE Healthcare UK Ltd, Little Chalfont, UK) and was used in random amplified polymorphic DNA analysis with ERIC 1 and ERIC 2A primers.11 The genetic similarity was determined using Dice coefficient as similarity measure and the unweighted pair group method with arithmetic mean (http://genomes.urv.cat/UPGMA/).
Quinolone resistance determining regions (QRDR) and PMQR detection
QRDR in gyrA and parC were amplified by PCR and sequenced as described previously.14 Plasmid mediated quinolone resistant determinants qnrA, qnrB, qnrC, qnrD, qnrS, and qepA were amplified14 and sequenced with ABI 3130xl Genetic Analyzer. PCRs for qepA, aac(6ʹ)-Ib,14 and oqxAB15 were carried out. The exact allele of aac(6ʹ)-Ib enzymes was determined with restriction with BtsCI (New England Biolabs, UK). If the allele Ib-cr was present, the PCR product was cut into two segments –270 bp and 210 bp.
The nucleotide sequences were analyzed with Chromas Lite 2.01 (Technelysium Pty Ltd) DNAMAN version 8.0 Software (Lynnon BioSoft) and NCBI Blast tool (http://www.ncbi.nlm.nih.gov). Mutations in QRDR of gyrA and parC were identified with comparison with DNA sequence of QRDR regions of E. cloacae ATCC 13047 (GenBank accession numbers D88980 and D88981 for gyrA and parC, respectively).16
Statistical analysis
Differences were assessed with the chi-squared test or Fisher’s exact test (https://www.graphpad.com/quickcalcs/contingency1.cfm).
Results
Bacterial isolates and antimicrobial susceptibility testing
During the period 2014–2017, a total of 167 Enterobacter cloacae complex and eight Enterobacter aerogenes isolates resistant to at least one third-generation cephalosporin were collected from patients hospitalized in different hospital wards: intensive care units (39), surgery wards (21), hemodialysis (eight), hematology (17), pediatric wards (13), nephrology/urology wards (31), and other internal wards (46). The isolates were recovered from different clinical samples: urine (46), blood (45), central venous catheters (five), wounds (35), tracheal secretions (14), bronchoalveolar lavage (two), sputum (eight), bile (one), drainage fluids (six), throat swabs (eight), and synovial fluid (one).
The results of the antimicrobial susceptibility testing exhibited high resistance rates (resistant and intermediately susceptible) to third-generation cephalosporins (selection criteria) (ceftazidime, 98.9%; cefotaxime; 100%), aminoglycosides (tobramycin, 89.7%; gentamicin, 82.3%), and fluoroquinolones (ciprofloxacin, 79.4%; levofloxacin, 39.4%). The resistance rates to trimethoprim/sulfamethoxazole and piperacillin/tazobactam were 58.2% and 72%, respectively. The isolates were highly susceptible to imipenem (100%), meropenem (100%), and amikacin (88%). Twenty four isolates (13.7%) were susceptible to cefepime. MIC50 and MIC90 of nalidixic acid, ciprofloxacin, and levofloxacin are shown in Table 1.
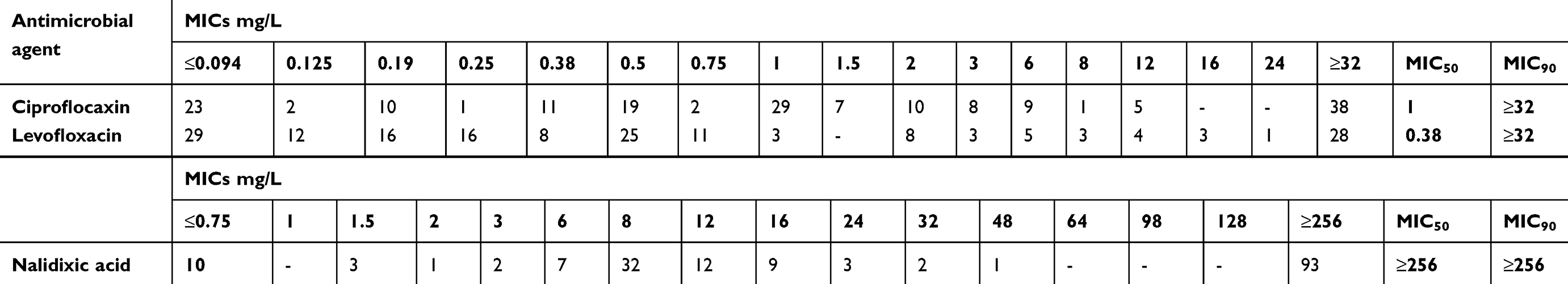 | Table 1 MICs of nalidixic acid, ciprofloxacin, and levofloxacin in 175 Enterobacter spp. isolates |
Beta-lactamase identification
The PCR study showed positive results for blaSHV and blaCTX-M group specific genes. The sequence analysis identified blaSHV-12, blaCTX-M-3, and blaCTX-M-15. ESBL producing isolates were identified in 87% (152/175). CTX-M-15 was the predominant ESBL, detected in 73% (128/175) of the isolates, predominantly in E. cloacae complex isolates – 76%. CTX-M-3 production was found in 18 isolates (10%) and SHV-12 was present in only six (3%) isolates. All E. aerogenes isolates (except one) produced CTX-M-3 ESBL – 88%. Twenty three isolates were negative for the major types of ESBLs and were assumed as probable AmpC hyperproducers. All but one isolates were negative for blaCMY, blaFOX, blaAAC, and blaDHA. A single isolate of E.cloacae complex, co-producer of CTX-M-3 and DHA-1, has been previously reported.8
IEF and bioassay
IEF was performed with 48 isolates, representative for the three detected enzyme groups (CTX-M-15, CTX-M-3, and SHV-12). In addition, eight isolates, negative for both CTX-M and SHV enzymes were also studied. Beta-lactamases with pIs 5.4, 7.4, 7.8, 8.2, 8.4, 8.8, and ≥9.0 were found. Considering their cefotaxime-hydrolyzing activity determined by bioassay, the enzymes were assigned to different groups of beta-lactamases. All tested isolates positive for blaCTX-M-15 gave one band with cefotaxime hydrolytic activity at pI 8.8. The tested isolates with blaCTX-M-3 showed band with cefotaxime hydrolytic activity at pI 8.4. We did not find bands corresponding to TEM type ESBLs. The beta-lactamase with pI 7.4, with no cefotaxime hydrolytic activity was assumed to be an OXA-type broad-spectrum beta-lactamase. The beta-lactamases (pI 5.4) which did not hydrolyze ceftazidime or cefotaxime were deemed broad-spectrum TEM-1. For CTX-M and SHV PCR negative isolates, single beta-lactamases with pI 7.8 or ≥9.0 and no cefotaxime or ceftazidime hydrolytic activity were detected. These isolates were considered as chromosomal type AmpC enzymes.
Conjugation experiments and replicon typing
The conjugation experiments were successful for 40 (28 positive for blaCTX-M-15 and 12 positive for blaCTX-M-3) of 175 donor isolates (23%). CTX-M-3 and CTX-M-15 producers transferred the ESBL determinants in 67% (12/18) and 22% (28/128), respectively. The resistotypes of the transconjugant strains are shown in Table 2. Plasmid replicon typing showed the presence of IncL/M replicon type in all transconjugants that carried blaCTX-M-3 (Table 2).
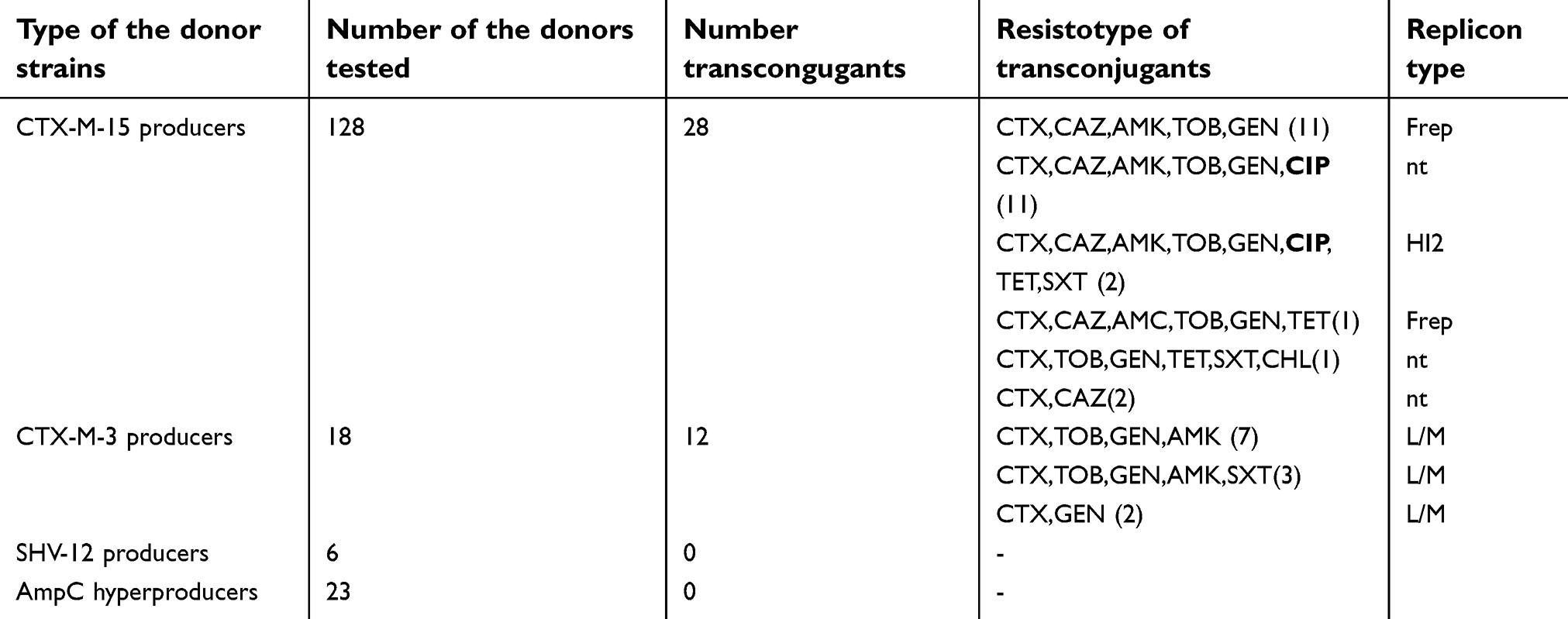 | Table 2 Resistotypes and replicon types of 40 transconjugant isolates of Enterobater spp. |
Regarding blaCTX-M-15 gene, according to the resistance profile, seven types of transconjugants were found: three of them were positive for F replicon type and two (with identical resistance profile) were HI2 positive (Table 2). The other transconjugants were not typеable. The blaSHV-12 positive Enterobacter spp. did not produce transconjugants. The aminoglycoside resistant determinants were most commonly co-transferred with ESBL genes. Quinolone resistance was observed in 13 transconjugates (33%), which were separated into two groups on the basis of their replicon type. (Table 2).
Molecular typing
ERIC PCR typing assigned all E. cloacae complex isolates to 15 different types. One dominant clone (A, n=92), consisting of three ERIC types (A, n=56; Aa, n=32; Ab, n=4) with similarity >0.8 was identified. ERIC types E (n=3) and V (n=19) also defined a separate clone (similarity 0.8). The other ERIC types represented different clones with similarity between 0.2 and 0.7: clone C (n=32), B (n=3), F (n=5), K (n=2), and L (n=6). ERIC types D, J, I, H, and P were represented by single E. cloacae complex isolates. E. aerogenes demonstrated three different ERIC profiles: g (n=6), m (n=1), and q (n=1) with similarity of 0.5–0.6.
Quinolone resistance determinants
PMQR determinants
All 175 isolates were tested for presence of PMQR determinants. Totally, 103 isolates (59%) had PMQR determinants. The frequency of PMQR determinants in the group of ciprofloxacin resistant isolates was significantly higher (69%, 99/143 isolates) than in the susceptible isolates (13%, 4/32 isolates, p<0.0001).
All 103 isolates harbored qnr determinants. Five of them (2%) were positive for aac(6ʹ)-Ib-cr variant. No qnrC, qnrD, qepA, and oqxAB gene positive isolates were found. Three different qnr families were identified – qnrA, qnrB, and qnrS. Sequencing showed three qnrB variants. The first qnrB variant was identical to the published sequence DQ351241 of qnrB117 and is number 1 in qnr nomenclature in Lachey.org (https://www.lahey.org/qnrstudies/); the second one was identical to published sequence EF526508 of qnrB9 and the third one was similar to published sequence accession number DQ303921 of qnrB4 allele18 with one silent mutation difference. QnrA and qnrS amplicons were identical to qnrA1 (accession number AY07023519 and CP031576.1 (Enterobacter hormaechei)) and qnrS1 (accession number AB187515).20
QnrB1 and qnrB9 were detected in 63 and 28 isolates, respectively. Combinations of determinants were found: qnrA1, qnrS1, aac(6ʹ)-Ib-cr and qnrB9, qnrS1 in two isolates (Table 3). One isolate produced qnrB4. Eight isolates were qnrA1 positive and four of them also harbored aac(6ʹ)-Ib-cr variant (Table 3). Three isolates were positive only for qnrS1.
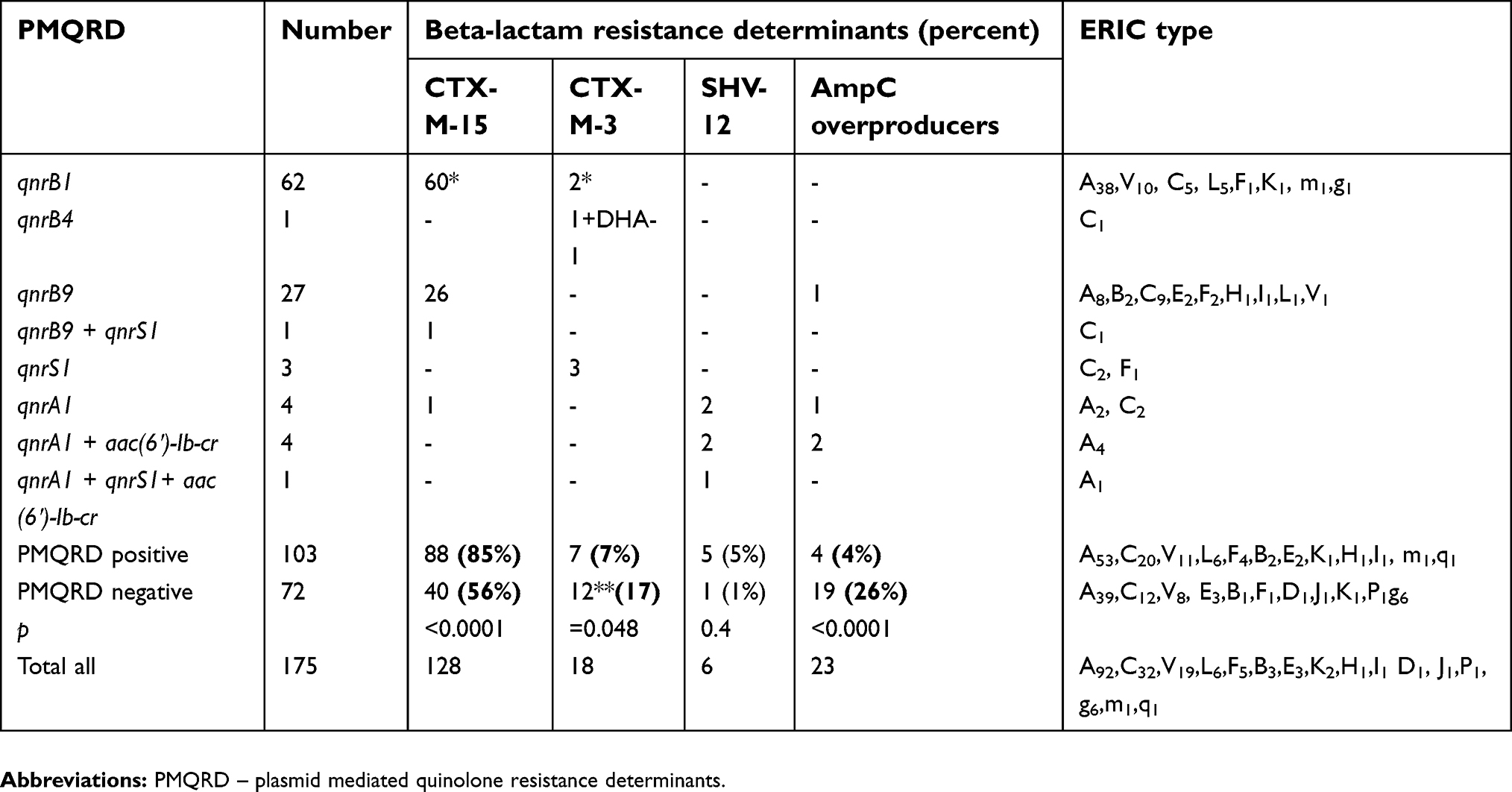 | Table 3 Distribution of plasmid mediated quinolone resistance (PMQR) alleles in Enterobacter spp. isolates according to the ESBL/AmpC enzymes and the ERIC type |
The association between ESBL and PMQR determinants in Enterobacter spp. isolates is presented on Table 3. CTX-M-15 ESBL was associated predominantly with qnr determinants, mostly qnrB alleles. Eighty-seven qnrB positive isolates (96%, 87/91) carried blaCTX-M-15, while only a single Enterobacter spp. isolate from a group of 16 positive for other qnr alleles isolates (6%) was a CTX-M-15 producer (p<0.0001).
The presence of blaCTX-M-15 was significantly lower in the group of PMQR negative isolates (56%; 40 from 72) in comparison with the PMQR positive group (Table 3). CTX-M-3 and AmpC hyperproducers were not associated with presence of PMQR determinants. The SHV-12 producers carried qnrA1 alleles. The single qnrB4 positive isolate was positive for blaCTX-M-3 and blaDHA-1.
QnrB1 was successfully transferred only in two conjugation experiments. The transconjugants demonstrated resistance to cefotaxime, ceftazidime, amikacin, tobramycin, gentamycin, ciprofloxacin, tetracycline, and co-trimoxazole. Replicon typing detected the presence of HI2 replicons. QnrB9 alleles were successfully transferred in eleven conjugation experiments. Resistance to ceftazidime, amikacin, tobramycin, gentamycin, and ciprofloxacin was detected in the transconjugants and they were nontypable in replicon typing. Some qnrB1 postive transconjugants showed elevated MICs of ciprofloxacin (0.25–0.5 mg/L) and levofloxacin (0.12–0.38 mg/L), which were slightly higher than those found in the qnrB9 positive transconjugants (MICof ciprofloxacin 0.12−0.25 mg/L; MICof levofloxacin 0.06−0.12 mg/L). The MICs for nalidixic acid were in the range 16–24 mg/L. Six of the eight E. aerogenes isolates were CTX-M-3 producers and PMQR negative. PMQR determinants, represented by qnrB1, were detected in two E. aerogenes isolates only.
QRDR
QRDR were studied in 130 Enterobacter isolates (127 E. cloacae and threeE. aerogenes isolates). Substitutions in the QRDR of gyrA or/and parC were found in 53% of them (Table 4). Fifteen isolates (all members of clone V) with the highest ciprofloxacin and levofloxacin MICs harbored two substitutions in the QRDR of gyrA (Ser83Tyr and Asp87Ala) and one in parC (Ser80Ile) (Table 4). Two substitutions (one in gyrA and one in parC) were detected in 15 isolates of E. cloacae complex (Table 4). These isolates were highly resistant to ciprofloxacin and those positive for qnrB4, qnrS1, and qnrA1 were also highly resistant to levofloxacin. A single substitution in gyrA region (Ser83Phe) was found in 35 isolates (Table 4). In general, the presence of at least one GyrA substitution led to a higher MIC of nalidixic acid (>256 mg/L), while a higher level of levofloxacin resistance was associated with a presence of two or more substitutions. Three E. aerogenes isolates showed presence of threonine in 83 positions of gyrA gene, which is typical for the wild type of E. aerogenes strains.21
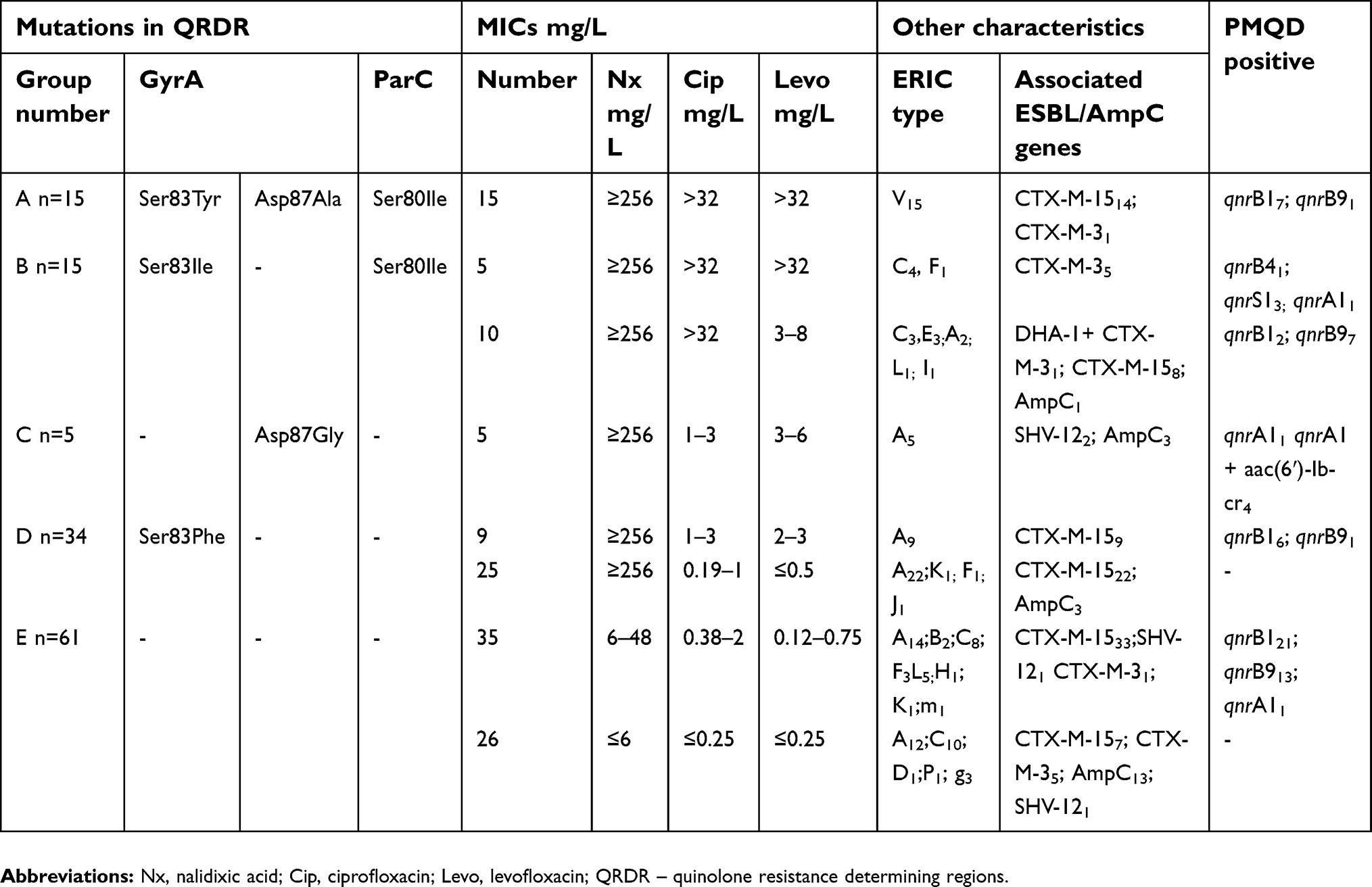 | Table 4 Distribution of 130 Enterobacter spp. isolates according to point mutations in quinolone-resistance determining regions (QRDR) of gyrA and parC, plasmid mediated quinolone resistance (PMQR), MICs of quinolones, ERIC types, and ESBLs |
Discussion
Quinolones are among the most commonly used antimicrobials to treat bacterial infections including those associated with Enterobacter spp. The prolonged usage of third-generation cephalosporins can lead to AmpC overexpression in Enterobacter spp. The present study demonstrated a high level (79.4%) of combined third-generation cephalosporins and ciprofloxacin non-suceptibility.
PMQR determinants have been increasingly reported in various Enterobacteriaceae species, especially in E. coli, Enterobacter spp., Klebsiella spp. and Salmonella spp.6 The published data greatly depend on the selection criteria and the number of investigated PMQR determinants. The present study demonstrated a high frequency of PMQR positive Enterobacter spp. (59%) in a large collection of third-generation cephalosporin resistant clinical isolates of Enterobacter spp. Similar results were reported for ESBL producing Enterobacter spp. isolates from Tunisia (50%), Mexico (61%), and Argentina (66%),22,23 but lower rates for isolates from Cote d’Ivoire (42.9%).24
PMQR determinants typically induce low-level quinolone resistance with ciprofloxacin MIC of ≥ 0.25 μg/mL. They facilitate the selection of chromosomal mutants with higher level quinolone resistance and promote treatment failure.5,7 In previous investigations, the presence of PMQR determinants in E. coli isolates was found to reduce ciprofloxacin activity in urinary25 and respiratory tract26 infections in a murine model.
The aac(6ʹ)-Ib-cr enzyme and the qnr proteins are the most widely distributed PMQR determinants worldwide.5,7 In the present study, the qnrB was the most common PMQR determinant, detected in 88% of all PMQR positive isolates (n=103). This is in concordance with other studies which reported qnrB determinant as the most prevailing PMQR determinant among Enterobacter spp. isolates. A survey from France showed that qnr, especially qnrB determinants, was predominant in carbapenem resistant E. cloacae isolates.27 The same finding was reported from authors in Tunisia and Mexico.22,23In a recent survey from Iran, qnr genes were the predominant PMQR determinants in E. cloacae (60.3%). The authors found qnrB1, qnrS1, and qnrB4 alone or in combinations, to be the most common genes.28
In our study, three variants of qnrB аlleles were detected: predominantly qnrB1 (61%; 63/103), followed by qnrB9 (27%; 28/103), and qnrB4 in a single isolate only. In addition, one qnrB9 allele was found in a combination with qnrS1. QnrB1 genes were commonly detected in Enterobacter spp. isolates. They prevailed in isolates from Iran,28 Tunisia,22 Italy,29 and Algeria30 In our study, qnrB4 was found in one isolate only. This is in contrast with reports from some European countries, where qnrB4 is the main detected allele.31,32 QnrB4 gene was recently detected in two Bulgarian NDM-1 producing isolates of Klebsiella pneumoniae.33 Other qnrB alleles (qnrB10, qnrB13, and qnrB18) were found in Citrobacter freundii isolates from Bulgaria.34
Until now, 94 qnrB alleles (https://www.lahey.org/qnrstudies/) have been reported with qnrB1, qnrB2, qnrB4, qnrB5, qnrB6, and qnrB19 being the most common.5–7 In addition to this allele group, the present study has added qnrB9 as the second most commonly detected gene (27%). This allele has been rarely reported and found mostly in E. coli35 and in environmental and fecal carriage isolates of C. freundii.36–38 Very recently, qnrB9 was detected in Enterobacter spp. clinical isolates.39
The attempts to transfer the quinolone resistance determinants by conjugation experiments were successful. QnrB1 gene was transferred rarely than QnrB9 and the qnrB1 transconjugants showed presence of replicon type HI2. In all cases the transconjugants were blaCTX-M-15 positive and demonstrated resistance to aminoglycosides. QnrB alleles have been found often on plasmids carrying TEM, CTX-M (especially CTX-M-15), SHV, VEB, IMP, DHA, OXA-48, and KPC-3.5,6 The results from the epidemiological typing revealed that the qnrB positive isolates belonged to different clones. We did not find any association between a particular qnrB allele and a specific clone. However, almost half of the isolates belonged to the predominant clone A (53%). We may conclude that the qnrB distribution in this study was mediated by both plasmid and clonal dissemination.
In concordance with other reports, qnrA1 and qnrS1 genes were rarely found in our study.6,22,28 Similar to qnrB, all isolates with qnrA1 determinants belonged to the main clone A. This fact showed the possibility for this clone to acquire different PMQR determinants. QnrS1 was predominantly detected in clone C isolates.
Interestingly, we revealed a very low number of aac(6ʹ)-Ib-cr positive isolates. The acetyltransferase aac(6ʹ)-Ib-cr was produced only in combination with qnrA1 (n=4) or qnrA1 and qnrS1 (n=1). It has been reported commonly in E. coli isolates, less frequently in K. pneumoniae, and rarely in E. cloacae.5,6 However, some studies among Enterobacter spp. isolates showed higher rates for this enzyme.6,27,31 Aac(6ʹ)-Ib-cr enzyme has been previously reported in Bulgaria, mainly in E. coli, but also in E. aerogenes, K. pneumoniae, and C. freundii.34
To the best of our knowledge, the present study is the first report of qnrB1, qnrB9, qnrA, and qnrS in Bulgaria. Similar to other surveys, we did not find oqxAB, qepA, qnrD, and qnrC.6,22
The present study investigated 175 third-generation cephalosporin resistant clinical isolates of Enterobacter spp. ESBL production was identified in 87%. Only 23 isolates were possible AmpC overproducers. CTX-M-15 was the predominant ESBL type in E. cloacae complex isolates. E. aerogenes mostly produced CTX-M-3 enzymes. These results confirm the findings of our previous report in 201440 and demonstrate a significant increase in the rate of CTX-M-15 producing E. cloacae isolates (45% in 2011/2012 vs 76% in the present study) (p<0.0001).
In the present study, a significant association between CTX-M-15 production and the presence of PMQR determinants was found: 88% of all PMQR positive isolates were CTX-M-15 producers vs 56% of all PMQR negative isolates (p<0.0001). CTX-M-15 was associated with qnrB1 or qnrB9. The single qnrB4 positive isolate detected in this study was a co-producer of DHA-1 and CTX-M-3 enzyme. The association between qnrB4 and blaDHA-1 has been reported for K. pneumoniae.41 In addition, Potron et al31 found an association between qnrB4 and SHV-12 ESBL in Enterobacter spp. isolates. In our study, qnrS1 was associated with CTX-M-3 production. SHV-12 ESBL producers predominantly showed the presence of qnrA1 allele (five Enterobacter spp. from all six had qnrA1), although this association is not significant due to the small number of isolates. Similar data were reported from Korea and Japan.41,42
In addition to the PMQR determinants, substitutions in QRDR were detected in 53% of all ciprofloxacin resistant isolates (n=103). The most frequent substitutions in gyrA were Ser83Phe (49%), followed by Ser83Ile (22%), Ser83Tyr (22%), Asp87Ala (22%), and Asp87Gly (7%) (Table 4). For ParC, the Ser80Ile substitution was the single type mutation identified in 44% of all isolates (Table 4). Ser83Phe and Ser83Ile have been reported as common mutations in gyrA among E. cloacae isolates from France21,27 and Tunizia22
In our study, Ser83Phe mutation (Group D, Table 4) was associated with isolates from clone A (31 of 34 members). The isolates positive for this mutation could be divided into two groups: with and without PMQR determinants (Table 4). For the qnr positive group, slightly increased MICs to ciprofloxacin were observed, but both groups demonstrated levofloxacin susceptibility.
Asp87Gly mutation was found in five qnrA1 positive isolates from clone A. These isolates showed slightly elevated MICs to both ciprofloxacin and levofloxacin.
The substitutions Ser83Ile (gyrA) and Ser80Ile (parC) were identified in 15 isolates (Group B, Table 4) with higher MICs for nalidic acid (>256 mg/L) and ciprofloxacin (>32 mg/L). The MICs to levofloxacin were slighly increased (3–8 mg/L) and were higher in PMQR positive isolates. Interestingly, this group was presented predominantly by CTX-M-3 producers and the single DHA-1 positive isolate.
Three substitutions – two in GyrA (Ser83Tyr, Asp87Ala) and one in ParC (Ser80Ile) were identified in 15 isolates with very high MICs to the three tested quinolones (Group A, Table 4). Isolates with three substitutions and respectively higher resistance rates were associated with only one clone – V. In this group, only seven isolates were qnr positive.
In addition, ParC mutations were found in combination with GyrA substitutions. This is in agreement with other studies which demonstrated that the bacterial gyrase is the primary affected enzyme, while topoisomarase IV is the secondary target.5,7
The present results showed that a single mutation in QRDR resulted in the increase in the MICs for nalidixic acid only, while the higher level of quinolone resistance was associated with two or more mutations. This finding is in concordance with those of other authors.5–7 The relatively small number of isolates with two and more mutations is in concordance with not very high resistance to levofloxacin, which gave more therapeutic options.
In conclusion, a high proportion of combined third-generation cephalosporin and quinolone resistant Enterobacter spp. was detected. The rate of ESBL producers in the isolates was high (87%), CTX-M-15 being the major enzyme identified. The predominant PMQR determinant was qnrB (qnrB1 and qnrB9). They were co-transferred with blaCTX-M-15 gene, showing possibility of both beta-lactams and quinolones to select resistant strains. The alterations in 83 and 87 positions of gyrB in QRDR, and in 80 positions of parC gene were associated with a high level of quinolone resistance. The results showed the need for more prudent use of quinolones and third-generation cephalosporins because of the risk of promoting dissemination and selection of multiple resistance determinants (ESBL, PMQR) among Enterobacter spp. isolates.
Acknowledgments
This study was funded by grant number 8383/07.12.2016 and contract number D-59/2.05. 2017 from the Medical University of Sofia, Sofia, Bulgaria.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Davin-Regli A, Pages JM. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front Microbiol. 2015;6:392. doi:10.3389/fmicb.2015.00392
2. Mezzatesta ML, Gona F, Stefani S. Enterobacter cloacae complex: clinical impact and emerging antibiotic resistance. Future Microbiol. 2012;7:887–902. doi:10.2217/fmb.12.61
3. Tacconelli E, Carrara E, Savoldi A, et al. WHO pathogens priority list working group. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
4. Potter RF, D‘Souza AW, Dantas G. The rapid spread of carbapenem-resistant Enterobacteriaceae. Drug Resist Updat. 2016;29:30–46. doi:10.1016/j.drup.2016.09.002
5. Rodríguez-Martínez JM, Machuca J, Cano ME, Calvo J, Martínez-Martínez L, Pascual A. Plasmid-mediated quinolone resistance: two decades on. Drug Resist Updates. 2016;29:13–29. doi:10.1016/j.drup.2016.09.001
6. Yanat B, Rodríguez-Martínez JM, Touati A. Plasmid-mediated quinolone resistance in Enterobacteriaceae: a systematic review with a focus on Mediterranean countries. Eur J Clin Microbiol Infect Dis. 2017;36(3):421–435. doi:10.1007/s10096-016-2847-x
7. Hooper D, Jacoby GA. G Jacoby mechanisms of drug resistance: quinolone resistance. Ann N Y Acad Sci. 2015;1354(1):12–31. doi:10.1111/nyas.2015.1354.issue-1
8. Dimitrova D, Markovska R, Stoeva T, et al. First report of DHA-1 producing Enterobacter cloacae complex isolate in Bulgaria. Folia Med (Plovdiv). Epub 2019 Mar 15.
9. Carattoli A, Bertini A, Villa L, Falbo V, Hopkins KL, Threlfall EJ. Identification of plasmids by PCR-based replicon typing. J Microbiol Methods. 2005;63:219–228. doi:10.1016/j.mimet.2005.03.018
10. Garcia-Fernandes A, Fortini D, Veldman K, Mevius D, Carattoli A. Characterization of plasmids harbouring qnrS1, qnrB2 and qnrB19 genes in Salmonella. J Antimicrob Chemother. 2009;63:274–281. doi:10.1093/jac/dkn470
11. Markovska R, Schneider I, Keuleyan E, et al. Extended-spectrum beta-lactamase-producing Enterobacteriaceae in Bulgarian hospitals. Microb Drug Resist. 2008;14(2):119–128. doi:10.1089/mdr.2008.0814
12. Pérez-Pérez FJ, Hanson ND. Detection of plasmid-mediated AmpC beta-lactamase genes in clinical isolates by using multiplex PCR. J Clin Microbiol. 2002;40(6):2153–2162.
13. Yan JJ, Ko WC, Jung YC, Chuang CL, Wu JJ. Emergence of Klebsiella pneumoniae isolates producing inducible DHA-1 beta-lactamase in a university hospital in Taiwan. J Clin Microbiol. 2002;40(9):3121–3126.
14. Li B, Yi Y, Wang Q, et al. Analysis of drug resistance determinants in Klebsiella pneumoniae isolates from a tertiary-care hospital in Beijing, China. PLoS One. 2012;7(7):e42280. doi:10.1371/journal.pone.0042280
15. Kim HB, Park CH, Kim CJ, Kim EC, Jacoby GA, Hooper DC. Prevalence of plasmid-mediated quinolone resistance determinants over a 9-year period. Antimicrob Agents Chemother. 2009;53(2):639–645. doi:10.1128/AAC.01051-08
16. Deguchi T, Yasuda M, Nakano M, et al. Detection of mutations in the gyrA and parC genes in quinolone-resistant clinical isolates of Enterobacter cloacae. J Antimicrob Chemother. 1997;40(4):543–549.
17. Jacoby G, Walsh K, Mills D, et al. qnrB, another plasmid-mediated gene for quinolone resistance. Antimicrob Agents Chemother. 2006;50:1178–1182. doi:10.1128/AAC.50.4.1178-1182.2006
18. Robicsek A, Strahilevitz J, Sahm D, Jacoby G, Hooper D. qnr prevalence in ceftazidime-resistant Enterobacteriaceae isolates from the United States. Antimicrob Agents Chemother. 2006;50:2872–2874. doi:10.1128/AAC.01647-05
19. Tran J, Jacoby G. Mechanism of plasmid-mediated quinolone resistance. Proc Natl Acad Sci U S A. 2002;99:5638–5642. doi:10.1073/pnas.082092899
20. Hata M, Suzuki M, Matsumoto M, et al. Cloning of a novel gene for quinolone resistance from a transferable plasmid in Shigella flexneri 2b. Antimicrob Agents Chemother. 2005;49:801–803. doi:10.1128/AAC.49.2.801-803.2005
21. Lascols C, Robert J, Cattoir V, et al. Type II topoisomerase mutations in clinical isolates of Enterobacter cloacae and other enterobacterial species harbouring the qnrA gene. Int J Antimicrob Agents. 2007;29(4):402–409. doi:10.1016/j.ijantimicag.2006.11.008
22. Ferjani S, Saidani M, Amine FS, Boutiba-Ben Boubaker I. Prevalence and characterization of plasmid-mediated quinolone resistance genes in extended-spectrum β-lactamase-producing Enterobacteriaceae in a Tunisian hospital. Microb Drug Resist. 2015;21(2):158–166. doi:10.1089/mdr.2014.0053
23. Silva-Sanchez J, Barrios H, Reyna-Flores F, et al., ; . Prevalence and characterization of plasmid-mediated quinolone resistance genes in extended-spectrum β-lactamase-producing Enterobacteriaceae isolates in Mexico. Microb Drug Resist. 2011;17(4):497–505. doi:10.1089/mdr.2011.0086
24. Guessennd N, Bremont S, Gbonon V, et al. [Qnr-type quinolone resistance in extended-spectrum beta-lactamase producing enterobacteria in Abidjan, Ivory Coast]. Pathol Biol (Paris). 2008;56(7–8):439–446. doi:10.1016/j.patbio.2008.07.025
25. Allou N, Cambau E, Massias L, Chau F, Fantin B. Impact of low-level resistance to fluoroquinolones due to qnrA1 and qnrS1 genes or a gyrA mutation on ciprofloxacin bactericidal activity in a murine model of Escherichia coli urinary tract infection. Antimicrob Agents Chemother. 2009;53(10):4292–4297. doi:10.1128/AAC.01664-08
26. Domínguez-Herrera J, Velasco C, Docobo-Pérez F, et al. Impact of qnrA1, qnrB1 and qnrS1 on the efficacy of ciprofloxacin and levofloxacin in an experimental pneumonia model caused by Escherichia coli with or without the GyrA mutation Ser83Leu. J Antimicrob Chemother. 2013;68(7):1609–1615. doi:10.1093/jac/dkt063
27. Guillard T, Cholley P, Limelette A, et al. Fluoroquinolone Resistance Mechanisms and population structure of Enterobacter cloacae non-susceptible to Ertapenem in North-Eastern France. Front Microbiol. 2015;6:1186; print. doi:10.3389/fmicb.2015.01186
28. Peymani A, Farivar TN, Najafipour R, Mansouri S. High prevalence of plasmid-mediated quinolone resistance determinants in Enterobacter cloacae isolated from hospitals of the Qazvin, Alborz, and Tehran provinces, Iran. Rev Soc Bras Med Trop. 2016;49(3):286–291. Qnr B1 common Iran свалена. doi:10.1590/0037-8682-0454-2015
29. Perilli M, Forcella C, Celenza G, et al. Evidence for qnrB1 and aac(6‘)-Ib-cr in CTX-M-15-producing uropathogenic Enterobacteriaceae in an Italian teaching hospital. Diagn Microbiol Infect Dis. 2009;64(1):90–93. doi:10.1016/j.diagmicrobio.2009.01.009
30. Iabadene H, Messai Y, Ammari H, et al. Dissemination of ESBL and Qnr determinants in Enterobacter cloacae in Algeria. J Antimicrob Chemother. 2008;62:133–136. doi:10.1093/jac/dkn145
31. Potron A, Poirel L, Bernabeu S, Monnet X, Richard C, Nordmann P. Nosocomial spread of ESBL-positive Enterobacter cloacae co-expressing plasmid-mediated quinolone resistance Qnr determinants in one hospital in France. J Antimicrob Chemother. 2009;64(3):653–654. doi:10.1093/jac/dkp222
32. Müller S, Oesterlein A, Frosch M, Abele-Horn M, Valenza G. Characterization of extended-spectrum beta-lactamases and qnr plasmid-mediated quinolone resistance in German isolates of Enterobacter species. Microb Drug Resist. 2011;17(1):99–103. doi:10.1089/mdr.2010.0114
33. Todorova B, Sabtcheva S, Ivanov IN, et al. First clinical cases of NDM-1-producing Klebsiella pneumoniae from two hospitals in Bulgaria. J Infect Chemother. 2016;22:837–840. doi:10.1016/j.jiac.2016.03.014
34. Sabtcheva S, Kaku M, Saga T, Ishii Y, Kantardjiev T. High prevalence of the aac(6‘)-Ib-cr gene and its dissemination among Enterobacteriaceae isolates by CTX-M-15 plasmids in Bulgaria. Antimicrob Agents Chemother. 2009;53(1):335–336. doi:10.1128/AAC.00584-0 8
35. Coban AY, Nohut OK, Tanrıverdi Çaycı Y, et al. [Investigation of plasmid-mediated quinolone resistance determinants in enterobacteriaceae: a multicenter study]. Mikrobiyol Bul. 2012;46(3):366–374.
36. Zhang R, Ichijo T, Huang YL, et al. High prevalence of qnr and aac(6‘)-Ib-cr genes in both water-borne environmental bacteria and clinical isolates of Citrobacter freundii in China. Microbes Environ. 2012;27(2):158–163.
37. Anssour L, Messai Y, Derkaoui M, et al. ESBL, plasmidic AmpC, and associated quinolone resistance determinants in coliforms isolated from hospital effluent: first report of qnrB2, qnrB9, qnrB19, and blaCMY-4 in Algeria. J Chemother. 2014;26(2):74–79. doi:10.1179/1973947813Y.0000000115
38. Liu L, Chen D, Liu L, et al. Genetic diversity, multidrug resistance, and virulence of Citrobacter freundii from diarrheal patients and healthy individuals. Front Cell Infect Microbiol. 2018;10(8):233. doi:10.3389/fcimb.2018.00233
39. Osei Sekyere J, Amoako DG, Galdiero M. Genomic and phenotypic characterisation of fluoroquinolone resistance mechanisms in Enterobacteriaceae in Durban, South Africa. PLoS One. 2017;12(6):e0178888. doi:10.1371/journal.pone.0178888
40. Markovska RD, Stoeva TJ, Bojkova KD, Mitov IG. Epidemiology and molecular characterization of extended-spectrum beta-lactamase-producing Enterobacter spp., Pantoea agglomerans, and Serratia marcescens isolates from a Bulgarian hospital. Microb Drug Resist. 2014;20(2):131–137. doi:10.1089/mdr.2013.0102
41. Jeong HS, Bae IK, Shin JH, et al. Prevalence of plasmid-mediated quinolone resistance and its association with extended-spectrum beta-lactamase and AmpC beta-lactamase in Enterobacteriaceae. Korean J Lab Med. 2011;31(4):257–264. doi:10.3343/kjlm.2011.31.4.257
42. Kanamori H, Yano H, Hirakata Y, et al. Molecular characteristics of extended-spectrum beta-lactamases and qnr determinants in Enterobacter species from Japan. PLoS One. 2012;7(6):e37967. doi:10.1371/journal.pone.0037967
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.