Back to Journals » Cancer Management and Research » Volume 12
Quercetin Inhibits Epithelial-to-Mesenchymal Transition (EMT) Process and Promotes Apoptosis in Prostate Cancer via Downregulating lncRNA MALAT1
Authors Lu X, Chen D, Yang F, Xing N
Received 10 December 2019
Accepted for publication 21 February 2020
Published 9 March 2020 Volume 2020:12 Pages 1741—1750
DOI https://doi.org/10.2147/CMAR.S241093
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Rudolph Navari
Xinxing Lu,1 Dong Chen,2 Feiya Yang,2 Nianzeng Xing1,2
1Department of Urology, Beijing Chaoyang Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Department of Urology, Chinese Academy of Medical Sciences Cancer Institute and Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, People’s Republic of China
Correspondence: Nianzeng Xing
Department of Urology, Chinese Academy of Medical Sciences Cancer Institute and Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing, People’s Republic of China
Email [email protected]
Background: Prostate cancer (PC) is one of the most common carcinomas in men worldwide. The lack of effective therapies urges the development of novel therapeutic options against PC. Quercetin (Quer) is a flavonoid compound that has been shown to effectively inhibit PC in vitro and in vivo. However, the underlying mechanisms await elucidation. Long non-coding RNA MALAT1 has been reported as an oncogenic target in multiple types of cancers, including PC. Previous data showed that quercetin promoted the apoptosis of fibroblast-like synoviocytes by upregulating MALAT1 in rheumatoid arthritis. However, we speculate that mechanisms are different in PC.
Materials and Methods: Human PC cell line PC-3 and its xenograft tumor were chosen as in vitro and in vivo models for PC. A series of in vitro and in vivo functional experiments were carried out to elucidate the role of MALAT1 in quercetin treatment against PC. Western blot was performed to measure the expression of related proteins to explore underlying molecular mechanisms.
Results: We showed for the first time that MALAT1 expression was significantly downregulated in quercetin-treated PC cells in a dose- and time-dependent manner. Also, quercetin inhibited the proliferation of PC cells and the growth of xenograft tumors. Moreover, quercetin suppressed EMT process, promoted apoptosis and deactivated PI3K/Akt signaling pathway during the progression of PC. MALAT1 overexpression in PC cells resulted in the resistance against quercetin treatment.
Conclusion: Our study illustrated, for the first time, that MALAT1 played an important role in quercetin treatment against PC by inhibiting EMT process and promoting apoptosis, providing a new molecular basis for the application of quercetin in PC treatment.
Keywords: prostate cancer, quercetin, lncRNA, MALAT1, PI3K/Akt, epithelial-to-mesenchymal transition, apoptosis
Introduction
Prostate cancer (PC) is the most common malignancy and the second leading cause of cancer death in men in the US with an estimated 174,650 new cases and 31,620 deaths in 2019.1 The incidence of PC has also been increasing rapidly in developing countries, such as China.2 PC could be treated by radical prostatectomy or radiation at an early stage, but most PC patients ultimately develop local recurrence and metastasis. Although substantial progress has been made over the past decades, there is little improvement in the survival rates of PC.3
Quercetin (3,3ʹ,4ʹ,5,7-pentahydroxyflavone) is a flavonoid compound that has attracted enormous attention in cancer treatment.4,5 Our previous data and other study showed that quercetin suppressed the proliferation of human PC cells via multiple signaling pathway.6 However, detailed mechanisms of how quercetin inhibits cancer development remain unrevealed.
Long noncoding RNAs (lncRNAs) are a class of endogenous noncoding RNAs with >200 nucleotides.7,8 They are involved in cellular proliferation, apoptosis, and migration in a variety of cancers.9–11 Metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) was first shown to be highly expressed in non-small cell lung cancer with a potential role in cell invasion and metastasis.12 It has also been found to be overexpressed and served as a potential therapeutic target in PC.13–15 Pan et al reported that quercetin promoted the apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis by upregulating lncRNA MALAT1.16 However, the relationship between lncRNA MALAT1 and quercetin treatment in PC is still unknown.
In this study, we demonstrated that quercetin inhibited EMT process and promoted apoptosis in PC via downregulating lncRNA MALAT1.
Materials and Methods
Cell Culture
Human prostate cancer PC-3 (androgen independent) cell lines were obtained from Peking Union Medical College and maintained in RPMI-1640 medium (Hyclone, Logan, UT) supplemented with 10% fetal bovine serum (Hyclone) in a 5% CO2 humidified incubator at 37°C. Quercetin (Sigma, St. Louis, MO, USA) was dissolved in DMSO to a concentration of 10 mM as a working solution. The working solution was diluted in culture medium to reach indicated concentrations. The same volume of DMSO without quercetin was used as vehicle control.
Cell Transfection
PC-3 cells in the logarithmic growth phase were seeded in six-well plates until 75% confluency. The overexpressing plasmids (pcDNA3.1-MALAT1) were synthesized by Fenhui Biotechnologies Inc (Hunan, China). The pcDNA3.1-MALAT1 or vector was transfected into cells using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. After 6-h incubation, the medium was replaced. Cells transfected with recombinant pcDNA3.1 plasmid were cultured with 1 lg/mL puromycin (Beyotime, Shanghai, China) and placed in an incubator containing 95% air and 5% CO2 at 37°C.
RNA Isolation and qPCR
To determine the expression levels of MALAT1, total RNAs were extracted from tissues and cells using Trizol agent (Invitrogen, USA) according to the manufacturer’s protocol and reversely transcribed to complementary DNA using M-MLV Reverse transcriptase (Invitrogen, USA). Then, real-time PCR was performed using Power SYBR Green PCR Master Mix (Applied Biosystems, USA) with designated primers. The relative RNA levels were normalized to GAPDH. The relative gene expression was analyzed by the 2−ΔΔCt method. Each experiment was repeated three times. The primers for MALAT1 or GAPDH were as follows: MALAT1, forward: 5ʹ-CTT AAGCGCAGCGCCATTTT-3ʹ and reverse: 5ʹ-CCTCCAAACCCCAAGACCAA-3ʹ; GAPDH, forward: 5ʹ-CAACTCCCTCAAGATTGTCAGCAA-3ʹ and reverse: 5ʹ-GGCATGGACTGTGGTCATGA-3ʹ.
Cell Proliferation Assay (CCK-8 Assay)
CCK-8 kit (KeyGEN BioTECH, China) was used to measure cell proliferation ability. Cells in the logarithmic growth phase were seeded into 96-well plates at the density of 3×103 cells per 100 µL. Twelve hours later, cells were treated with indicated concentrations of quercetin (Quer/µM). Two hours before the termination of each time point, 10 µL CCK-8 was added to each well and co-cultured with cells for another 2 h. The optical density (OD) of each well was measured with a microplate reader (Bio-Rad Laboratories, Inc., Berkeley, CA, USA) at the wavelength of 450 nm.
Colony Formation Assay
After the treatment with indicated concentrations of quercetin (Quer/µM) for 48 h, cells were trypsinized and dispensed into individual wells of 6-well culture dishes at a density of 200 cells per well, followed by incubation for 10 days without changing the medium. Cell colonies were fixed by 10% formaldehyde for 10 min and stained with 0.5% crystal violet for 5 mins. The number of colonies containing >50 cells was counted under a dissecting microscope. The percentage of cell survival was calculated. The experiment was performed in triplicate.
Cell Migration Assay (Wound Healing Assay)
Cell migration capacity was measured by wound-healing assay. Cells were grown in six-well plates until reaching 90% confluency. A sterile 200 μL pipette tip was used to scratch a vertical line on the plate. Cells were washed with PBS twice. Then, cells were treated with serum-free 1640 containing indicated ingredients for 48 h. Images were obtained in triplicate for each condition using an inverted microscope at 0 h (baseline) and 48 h (end-point). The wound-healing area was calculated by Image J software.
Cell Invasion Assay (Transwell Assay)
Transwell assay was performed to assess cell invasion capability. The 24-well BioCoat cell culture inserts (BD Biosciences, Bedford, MA, USA) with a polyethylene terephthalate membrane (8-μm porosity) were used, in which the membranes of the upper chamber were coated with Matrigel (1mg/mL, BD) and incubated at 37°C for 6 h. Cells were treated with indicated ingredients for 24 h. Then about 1×105 cells were added into the upper chamber in serum-free media and 0.6 mL complete culture medium was added to the lower chamber. After 24-h incubation, cells were washed twice with PBS. The remaining cells were wiped away using cotton swabs. Cells were fixed by paraformaldehyde and stained with 0.1% crystal violet. The number of invasion cells on the bottom surface of the membrane was counted under a light microscope.
Apoptosis Analysis by TUNEL Assay
Cell apoptosis was detected by TUNEL assay. Cells were cultured in 12-well plates for 24 h followed by the exposure of designated stimuli for 48 h. Then cells were harvested and apoptosis was measured using TUNEL cell apoptosis in situ detection kit (KeyGEN, Nanjing, China) according to the manufacturer’s instructions. Positively stained cells were counted using a microscope.
Western Blot Analysis
Cells were treated with indicated reagents for 48 h. Protein lysates were harvested using ice-cold RIPA buffer (Beyotime Institute of Biotechnology, Beijing, China) containing protease inhibitor cocktail (Roche, Switzerland). The protein concentrations were quantified by Bradford Protein Assay (Bio-Rad Laboratories, USA). Then protein samples were separated by SDS-PAGE and transferred onto a PVDF membrane. Nonspecific binding sites were blocked with 5% (w/v) fetal bovine serum at room temperature for 2 h. Then, the membrane was incubated with a mouse monoclonal antibody at 4°C overnight. The primary antibodies (1:1000; Abcam, Cambridge, MA, USA) were as follows: anti-β-actin, anti-p-Akt, anti-Akt, anti-Bax, anti-bcl-2, anti-E-cadherin and anti-N-cadherin. After three washes with TBST buffer, the membranes were incubated with HRP-labelled goat anti-mouse IgGs (1:2000; Abcam) at room temperature for 1 h. Protein expression was detected by ECL and photographed by BioSpectrum Gel Imaging System (UVP, Upland, CA). Beta-actin was used as a loading control.
Animal Study
Twenty male BALB/c nude mice (6-week old) were obtained from the ministry of experimental animals of Capital Medical University. Mice were randomly assigned into four groups: (1) Vector group (PC-3 cells with Vector + intraperitoneal injection of vehicle every other day), (2) MALAT1 group (MALAT1 overexpression PC-3 cells + intraperitoneal injection of vehicle every other day), (3) Quercetin group (PC-3 cells with vector + intraperitoneal injection of 75 mg/kg quercetin every other day), (4) Quercetin + MALAT1 group (MALAT1 overexpression PC-3 cells + intraperitoneal injection of 75 mg/kg quercetin every other day). The therapeutic schedule was based on previous data from other researchers.17 Mice were housed in a pathogen-free environmental condition. Quercetin was dissolved in 25% hydroxypropyl-β-cyclodextrin (HPβCD, w/v in ddH2O). The same volume of HPβCD without quercetin was used as the vehicle control. Wild-type PC-3 cells or MALAT1-overexpressed PC-3 cells (1×105) were suspended in 100 μL PBS were injected subcutaneously into the right axillary fossa of mice. When the volume of xenograft tumor reached about 100 mm3, intervention treatment started. Tumor size was measured every week using a caliper, and tumor volume was calculated according to the formula: L×S2×0.5 (L = the longest diameter; S = the shortest diameter of tumor).
Immunohistochemistry (IHC) Stain
At the end of each animal study, xenograft tumors were fixed in 10% phosphate-buffered formalin and paraffin-embedded for immunohistochemical detection. Sections (5 µm) were deparaffinized with xylene, rehydrated in an alcohol gradient, immersed in 3% H2O2, and then incubated with primary antibody Ki67 (1:1000, Abcam). Subsequently, the sections were incubated with a secondary antibody using the ABC kit (Vector Laboratories. Burlingame, CA, USA) at room temperature for 1 h. The number of Ki67 positive cells was analyzed from three random high-power fields of each slide.
Statistical Analysis
All statistical analysis was performed using GraphPad Prism 6.0 software (GraphPad Software, CA, USA). Student’s t-test was used to analyze the differences between two groups and one-way ANOVA was used for multiple comparisons. P<0.05 was considered statistically significant.
Results
Effect of Quercetin on PC Cell Proliferation and MALAT1 Expression
The proliferation of PC-3 cells was inhibited by quercetin in a dose- and time-dependent manner (Figure 1A). Likewise, the expression of MALAT1 was dose- and time-dependently down-regulated by quercetin treatment (Figure 1B). These results suggested that MALAT1 might play a pivotal role in quercetin treatment in PC cells.
 |
Figure 1 Quercetin inhibited proliferation and colony formation of prostate cancer cells via down-regulating MALAT1. (A) CCK-8: PC-3 cells were treated with indicated concentrations of quercetin to examine the cell viability. (B) qPCR: MALAT1 expression was inhibited by quercetin in a dose- and time-dependent manner. (C, D) qPCR: MALAT1 overexpression validation and MALAT1 expression of each group. (E) CCK-8: PC-3 with vector cells or MALAT1 overexpressed PC-3 cells (MALAT1) were treated with blank or 50μM quercetin (50Q) for 48 h to measure the expression of MALAT1. Cell viability was measured. (F, G) colony formation assay: MALAT1 overexpression promoted colony formation ability and quercetin treatment reduced the number of colony, whereas MALAT1 overexpression significantly rescued colony formation capacity under the treatment of quercetin. *P<0.05, **P<0.01, ***P<0.001, compared with NC (negative control) or Vector group. #P<0.05, ##P<0.01, ###P<0.001, compared with the NC or 50Q+Vector group. |
Quercetin Inhibited PC Cell Proliferation via Downregulating MALAT1
We overexpressed MALAT1 in PC-3 cells (Figure 1C) and treated cells with quercetin. Our results showed that MALAT1 was significantly down-regulated in quercetin-treated PC-3 cells as compared to the non-quercetin-treated group (Figure 1D, P< 0.001). However, no significant difference was found in the expression of MALAT1 between 50Q + MALAT1 group and Vector group (Figure 1D, P> 0.05). To further ascertain that quercetin inhibited cell proliferation by targeting MALAT1, CCK-8 and colony formation assays were performed. The cell proliferation in Vector group was lower than that in MALAT1 group, but was much higher than that in Quercetin group (Figure 1E, P< 0.001). The overexpression of MALAT1 recovered the proliferation in PC-3 cells, which showed no significant difference as compared to Vector group (Figure 2A, P> 0.05). The colony formation assay also showed consistent results (Figure 1F and G). These data suggested that quercetin might inhibit the proliferation of PC cells via targeting MALAT1.
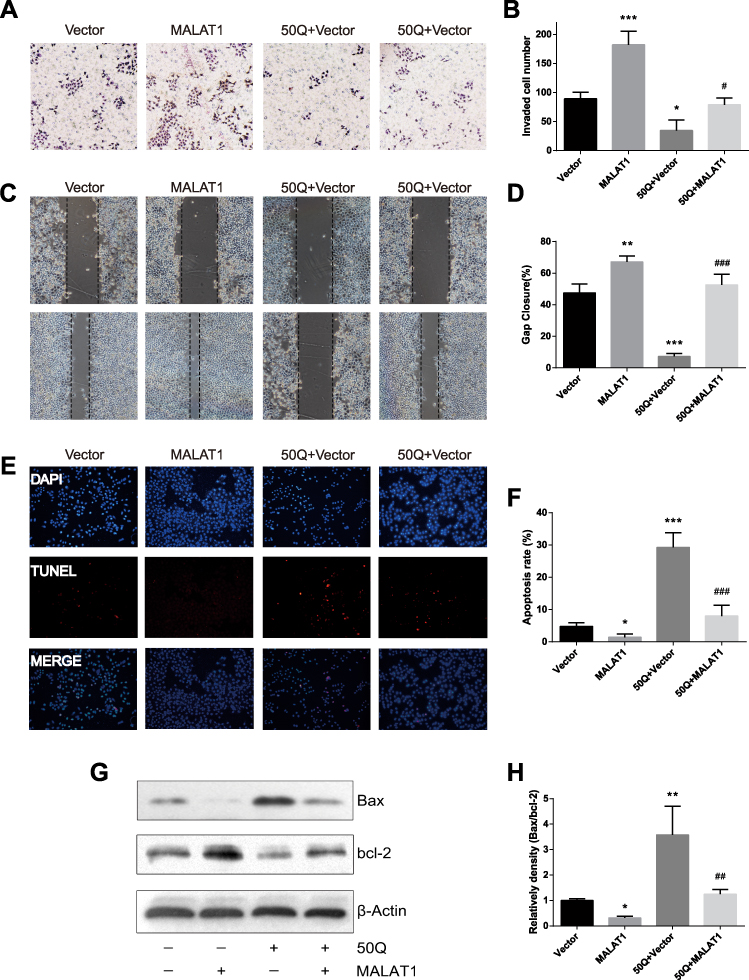 |
Figure 2 Quercetin suppressed invasion, migration and promoted apoptosis of prostate cancer cells via down-regulating MALAT1. (A, B) Transwell assay: MALAT1 overexpression promoted invasion and quercetin treatment impaired the invasive capacity of PC-3, whereas MALAT1 overexpression significantly rescued cell-invasive capacity in PC-3 cells under the treatment of quercetin. (C, D) Wound-healing assay: likewise, migration was measured. (E, F) TUNEL stain was employed to analyze cell apoptosis. (G, H) Apoptosis protein (Bax and Bcl-2) expression was detected by Western blotting. β-actin was used as the loading control. *P<0.05, **P<0.01, ***P<0.001, compared with Vector group. #P<0.05, ##P<0.01, ###P<0.001, compared with 50Q+Vector group. |
Quercetin Suppressed the Invasion and Migration of PC Cells via Downregulating MALAT1
The invasive ability of PC-3 cells was assessed by transwell invasion assay (Figure 2A and B). Compared to Vector group, the invasive capacity was significantly enhanced in MALAT1 group. The invasion potential of PC-3 cells was significantly inhibited by quercetin treatment, but restored with the overexpression of MALAT1. To elucidate the migration capacity of PC-3 cells, wound-healing assay was performed. The wound was nearly closed in MALAT1 group, but was wide open in quercetin-treated cells. When MALAT1 was overexpressed in PC-3 cells followed by the administration of quercetin, the wound healing was not significantly different from that of Vector group (P> 0.05, Figure 2C and D). Taken together, quercetin might target MALAT1 to suppress the invasion and migration of PC cells.
Quercetin Promoted the Apoptosis of PC Cells by Targeting MALAT1
Cell apoptosis was examined by TUNEL staining (Figure 2E and F) and Western blot (Figure 2G and H). The apoptotic rate in Vector group was slightly higher than that in MALAT1 group (P< 0.05, Figure 2E and F). Additionally, the percentage of apoptotic cells in quercetin group was much higher than that of Vector group (P< 0.001). In PC-3 cells overexpressing MALAT1, the number of apoptotic cells significantly decreased, and the apoptotic rate was comparable to that of Vector group (P> 0.05). The expressions of apoptotic proteins (Bax and Bcl-2) were detected by Western blotting (Figure 2G and H). The overexpression of MALAT1 protected PC-3 cells from apoptosis, while quercetin significantly induced apoptosis by regulating the expressions of pro-apoptotic protein Bax and anti-apoptotic protein Bcl-2. When MALAT1-overexpressed PC-3 cells were treated by quercetin, the ratio of Bax/Bcl-2 was comparable to that in Vector group (P> 0.05, Figure 2H). Taken together, these results indicated that MALAT1 might be an important target via which quercetin promoted the apoptosis of PC cells.
Quercetin Inhibited Tumor Growth by Targeting MALAT1 in vivo
To validate that quercetin inhibited tumor growth by targeting MALAT1 in vivo, wild-type PC-3 cells or MALAT1-overexpressed PC-3 cells were injected into male BALB/c nude mice to establish xenograft models. Mice in Quercetin group or Quercetin + MALAT1 group were intraperitoneally administered with quercetin (75 mg/kg) every other day, and the tumor size was recorded. Compared to Vector group, the tumor size and weight in MALAT1 group increased rapidly, while quercetin treatment significantly inhibited the tumor growth. The tumor size and weight in Quercetin + MALAT1 group showed no difference as compared to Vector group (P >0.05, Figure 3A–C). Likewise, the expression of MALAT1 was consistent with tumor size and weight (Figure 3D). In addition, immunohistochemistry analysis of Ki67 expression demonstrated that quercetin suppressed tumor cell proliferation via targeting MALAT1 in vivo (Figure 3E and F). In MALAT1 group, the overexpression of MALAT1 significantly elevated Ki67 expression as compared to Vector group (P >0.05, Figure 3E), while the production of Ki67 was significantly reduced by quercetin (Vector group vs Quercetin group: P<0.01, Figure 3E). In Quercetin + MALAT1 group, however, the expression of Ki67 was much higher than that in Quercetin group. Taken together, these results implied that quercetin inhibited tumor growth by targeting MALAT1 in vivo.
 |
Figure 3 Quercetin inhibits tumor growth, EMT process and PI3K/Akt signaling pathway via downregulating lncRNA MALAT1. (A) Photographs of dissected tumors in nude mice. (B) The tumor weight of the mice was measured in the end. (C) The tumor volume fluctuation of the mice was measured every week. (D) MALAT1 expression of tumor tissue. (E, F) Immunohistochemistry detected KI67-positive cells in PC-3 xenograft tumor tissue. (G–J) The expression of E-cadherin, N-cadherin, p-Akt and Akt was determined by Western blotting analysis. β-actin was used as the loading control. *P<0.05, **P<0.01, ***P<0.001, compared with Vector group. #P<0.05, ##P<0.01, ###P<0.001, compared with Quer+Vector group. |
Quercetin Inhibited EMT Process and PI3K/Akt Signaling Pathway via Downregulating MALAT1
To further explore the mechanism of quercetin treatment on prostate cancer, the expressions of p-AKT, AKT, E-cadherin and N-cadherin were determined by Western blot. PI3K/Akt signaling pathway plays a pivotal role in tumorigenesis.18–20 No significant difference was observed in AKT expression among the four groups. The level of phosphorylated Akt was significantly increased in MALAT1 group (Vector group vs MALAT1 group: P<0.01, Figure 3G and J). Quercetin significantly inhibited the phosphorylation of Akt (Vector group vs Quercetin group: P<0.05). However, Quercetin + MALAT1 group showed a much higher level of phosphorylated Akt than that of Quercetin group, suggesting that PC-3 cells gained resistance against quercetin treatment after the overexpression of MALAT1. Next, two important EMT process markers, E-cadherin and N-cadherin, were measured. The hallmark of EMT is the upregulation of N-cadherin followed by the downregulation of E-cadherin.21,22 The expression of E-cadherin was reduced followed by the upregulation of N-cadherin in MALAT1 group (Vector group vs MALAT1 group: P <0.05, Figure 3G–I). Quercetin increased E-cadherin expression and decreased the level of N-cadherin (Vector group vs quercetin group: P <0.05, Figure 3G–I). However, the effect of quercetin on the expression of E-Cadherin/N-Cadherin was inhibited by MALAT1 overexpression. The above findings indicated that quercetin might inhibit EMT process and PI3K/Akt signaling pathway via downregulating MALAT1.
Discussion
The incidence and mortality of PC are increasing rapidly in recent years, but current therapeutic approaches are still unsatisfactory for the management of PC.1,23 As a result, it is of great importance to develop novel treatment options for PC patients. Quercetin has presented its promising anticancer property in PC by inhibiting cell growth, invasion, and migration via targeting multiple signal pathways in vitro and in vivo.6,24 Moreover, unlike conventional chemotherapy, it is harmless to healthy cells.6
LncRNAs play pivotal roles in regulating chromatin dynamics, gene expression, cell growth, differentiation, and development. MALAT1 is a lncRNA associated with the progression of various human cancers.12,14,15 The upregulation of MALAT1 has been observed in PC cells relative to normal prostate epithelial cells or adjacent normal prostate tissues.25,26 Moreover, the expression level of MALAT-1 is correlated with higher Gleason score, PSA and tumor stage.25 MALAT-1 knockdown resulted in the inhibition of proliferation, invasion and migration, and an elevated apoptosis rate in PC cells.27 Previous studies showed that MALAT-1 promoted cancer progression mainly through the regulation of apoptosis-related genes, such as BAX, BCL2, CASP3, and CASP8.7 However, the relationship between quercetin treatment and MALAT1 is still unclear. Pan et al reported that quercetin promoted the apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis by upregulating MALAT1.16 The knockdown of MALAT1 reduced the activities of caspase-3 and caspase-9 and increased the Bcl-2/Bax ratio. This is inconsistent with the data obtained in PC cells. Thus, we hypothesized that quercetin might exert dual functions in the regulation of MALAT1. It might inhibit proliferation but promote apoptosis in PC via the downregulation of MALAT1.
In the present study, the results demonstrated that quercetin inhibited prostate cancer via the mechanisms as follows (Figure 4). We first showed that quercetin inhibited the proliferation of androgen-independent human PC-3 cells and the expression of MALAT1 in a dose- and time-dependent manner. Then, we treated wild-type and MALAT1-overexpressed PC-3 cells with or without quercetin. The overexpression of MALAT1 significantly promoted cell proliferation, colony formation, cell invasion and migration, but inhibited apoptosis in PC-3 cells. In quercetin-treated cells, however, the results were contrary to those observed in MALAT1 group. In Quercetin + MALAT1 group, cancer cells gained resistance against quercetin treatment, indicating that they were “rescued” by the overexpression of MALAT1. Similar results were found in nude mouse models. Quercetin treatment significantly decreased tumor volume, weight, as well as the expression of MALAT1 in vivo. MALAT1 overexpression, however, diminished the effect of quercetin treatment in xenograft mice. It was speculated that quercetin inhibited PC cell growth through the downregulation of MALAT1.
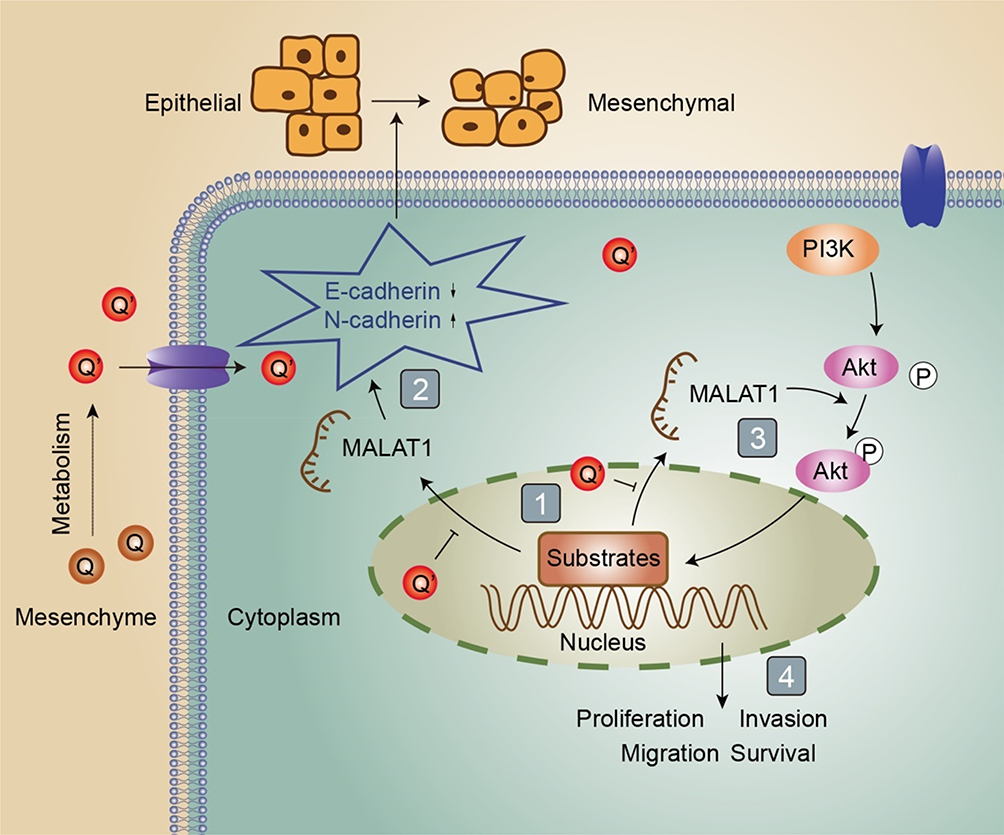 |
Figure 4 Schematic diagram. As depicted: (1) quercetin inhibited MALAT1 expression; (2) MALAT1 promoted EMT (epithelial-mesenchymal transition) process; (3, 4) MALAT1 promoted proliferation, invasion, migration and inhibited apoptosis via activating PI3K/Akt signal pathway. Q is short for quercetin; Q’ is short for quercetin metabolites; P is short for phosphorylation; The → indicates activation or induction, and → indicates inhibition. |
PI3K/Akt signal pathway is involved in the regulation of multiple biological processes, including apoptosis, metabolism and cell proliferation. The phosphorylation level of Akt plays an essential role in the activation of PI3K/Akt signal pathway.17,20 The hallmark of EMT is the upregulation of N-cadherin followed by the downregulation of E-cadherin.21,22 Here, we showed that the EMT process and the activity of PI3K/Akt signaling pathway were suppressed by quercetin administration. In Quercetin + MALAT1 group, however, the expressions of Bcl-2, N-cadherin, and p-Akt were upregulated and the levels of Bax and E-cadherin were downregulated. These results further proved our hypothesis that MALAT1 was an important target of quercetin treatment in the inhibition of EMT and PI3K/Akt signaling pathway.
In this study, we showed for the first time that MALAT1 expression was significantly downregulated in quercetin-treated PC cells at a dose- and time-dependent manner. Moreover, quercetin suppressed EMT process, promote apoptosis and deactivated PI3K/Akt signaling pathway in PC-3 cells through downregulation of MALAT1.
However, there were also some limitations in our study. For example, normal cells and standard chemotherapeutic agents were not included in the study. All experiments were carried out in PC-3 cell line. Another cell line, such as LNCaP, DU145 or 22Rv1, could be used for the validation of our conclusions. This is a preliminary study. A new article made by us is now in press, in which we showed that quercetin had anti-cancer effect against prostate cancer as good as first-line chemotherapeutic drug docetaxel in LNCaP and PC-3 cell lines and quercetin could also sensitize docetaxel and even reverse docetaxel resistance.28 Despite these inspiring results of in vitro and in vivo studies, it is still a question of how to use quercetin in humans. What is the ideal dose of quercetin with least toxicity and best therapeutic effect? Some previous reports revealed that there were not any side-effects even at high dose of 200 mg/kg given to rats or nude mice.29,30 And a prospective controlled trial showed that a consumption of 1000mg quercetin every day could be well tolerated and cure category III chronic prostatitis.31 Furthermore, bioavailability of quercetin is very low, which greatly discounted its anti-cancer effect.32 Recently, some novel nano-delivery systems were developed for quercetin treatment with better bioavailability.33 Therefore, to find new ways to improve the bioavailability of quercetin is also a good research direction.
In summary, our results indicated that MALAT1 could be an important target in quercetin treatment of PC, which might provide further support for the clinical use of quercetin for PC patients.
Ethic Statement
All procedures carried out in mice were approved by the Committee of Animal Experimentation and the Ethic Committee of Capital Medical University. The experiment was performed according to the NIH guidelines for animal care and use.34
Acknowledgment
We are very grateful to all individuals and groups involved in this study.
Funding
This research was financially supported by National Natural Science Foundation of China (Project number: 81772700) and Capital Science and Technology Leading Talent Project (Project number: Z181100006318007).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
2. Qi J, Wang L, Zhou M, et al. [Disease burden of prostate cancer among men in China, from 1990 to 2013]. Zhonghua Liu Xing Bing Xue Za Zhi. 2016;37(6):778–782. doi:10.3760/cma.j.issn.0254-6450.2016.06.007. Chinese.
3. Fleshner N. Defining high-risk prostate cancer: current status. Can J Urol. 2005;12 Suppl 1:
4. Botelho MF, Tralhao JG, Teixo RJ, et al. Quercetin in cancer treatment, alone or in combination with conventional therapeutics? Curr Med Chem. 2015;22(26):3025–3039.
5. Hollman PC, de Vries JH, van Leeuwen SD, Mengelers MJ, Katan MB. Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. Am J Clin Nutr. 1995;62(6):1276–1282. doi:10.1093/ajcn/62.6.1276
6. Yang F, Song L, Wang H, Wang J, Xu Z, Xing N. Quercetin in prostate cancer: chemotherapeutic and chemopreventive effects, mechanisms and clinical application potential. Oncol Rep. 2015;33(6):2659–2668. doi:10.3892/or.2015.3886
7. Gutschner T, Hämmerle M, Diederichs S. MALAT1—a paradigm for long noncoding RNA function in cancer. J Mol Med. 2013;91(7):791–801. doi:10.1007/s00109-013-1028-y
8. Michalik KM, You X, Manavski Y, et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ Res. 2014;114(9):1389–1397. doi:10.1161/CIRCRESAHA.114.303265
9. Jin Y, Feng S, Qiu S, Shao N, Zheng J. LncRNA MALAT1 promotes proliferation and metastasis in epithelial ovarian cancer via the PI3K-AKT pathway. Eur Rev Med Pharmacol Sci. 2017;21(14):3176–3184.
10. Tripathi V, Ellis JD, Shen Z, et al. The nuclear-retained noncoding RNA MALAT1 regulates alternative splicing by modulating SR splicing factor phosphorylation. Mol Cell. 2010;39(6):925–938. doi:10.1016/j.molcel.2010.08.011
11. Dong Y, Liang G, Yuan B, Yang C, Gao R, Zhou X. MALAT1 promotes the proliferation and metastasis of osteosarcoma cells by activating the PI3K/Akt pathway. Tumor Biol. 2015;36(3):1477–1486. doi:10.1007/s13277-014-2631-4
12. Ji P, Diederichs S, Wang W, et al. MALAT-1, a novel noncoding RNA, and thymosin β 4 predict metastasis and survival in early-stage non-small cell lung cancer. Oncogene. 2003;22(39):8031–8041. doi:10.1038/sj.onc.1206928
13. Dai X, Liang Z, Liu L, Guo K, Xu S, Wang H. Silencing of MALAT1 inhibits migration and invasion by sponging miR‑1‑3p in prostate cancer cells. Mol Med Rep. 2019;20(4):3499–3508. doi:10.3892/mmr.2019.10602
14. Dai X, Liu L, Liang Z, Guo K, Xu S, Wang H. Silencing of lncRNA MALAT1 inhibits cell cycle progression via androgen receptor signaling in prostate cancer cells. Pathol Res Pract. 2019;215(4):712–721. doi:10.1016/j.prp.2019.01.011
15. Zheng T, Ma G, Tang M, Li Z, Xu R. IL-8 Secreted from M2 macrophages promoted prostate tumorigenesis via STAT3/MALAT1 pathway. Int J Mol Sci. 2019;20(1):98. doi:10.3390/ijms20010098
16. Pan F, Zhu L, Lv H, Pei C. Quercetin promotes the apoptosis of fibroblast-like synoviocytes in rheumatoid arthritis by upregulating lncRNA MALAT1. Int J Mol Med. 2016;38(5):1507–1514. doi:10.3892/ijmm.2016.2755
17. Wang P, Henning SM, Magyar CE, Elshimali Y, Heber D, Vadgama JV. Green tea and quercetin sensitize PC-3 xenograft prostate tumors to docetaxel chemotherapy. J Exp Clin Cancer Res. 2016;35(1):73. doi:10.1186/s13046-016-0351-x
18. Schaefer T, Lengerke C. SOX2 protein biochemistry in stemness, reprogramming, and cancer: the PI3K/AKT/SOX2 axis and beyond. Oncogene. 2019:1–5.
19. Presti D, Quaquarini E. The PI3K/AKT/mTOR and CDK4/6 pathways in endocrine resistant HR+/HER2− metastatic breast cancer: biological mechanisms and new treatments. Cancers. 2019;11:1242. doi:10.3390/cancers11091242
20. Chen H, Zhou L, Wu X, et al. The PI3K/AKT pathway in the pathogenesis of prostate cancer. Front Biosci (Landmark Ed). 2016;21:1084–1091. doi:10.2741/4443
21. Knudsen KA, Wheelock MJ. Plakoglobin, or an 83-kD homologue distinct from beta-catenin, interacts with E-cadherin and N-cadherin. J Cell Biol. 1992;118(3):671–679. doi:10.1083/jcb.118.3.671
22. Gravdal K, Halvorsen OJ, Haukaas SA, Akslen LA. A switch from E-cadherin to N-cadherin expression indicates epithelial to mesenchymal transition and is of strong and independent importance for the progress of prostate cancer. Clin Cancer Res. 2007;13(23):7003–7011. doi:10.1158/1078-0432.CCR-07-1263
23. Miller KD, Nogueira L, Mariotto AB, et al. Cancer treatment and survivorship statistics, 2019. CA Cancer J Clin. 2019;69(5):363–385. doi:10.3322/caac.21565
24. Xing N, Chen Y, Mitchell SH, Young CY. Quercetin inhibits the expression and function of the androgen receptor in LNCaP prostate cancer cells. Carcinogenesis. 2001;22(3):409–414. doi:10.1093/carcin/22.3.409
25. Ren S, Liu Y, Xu W, et al. Long noncoding RNA MALAT-1 is a new potential therapeutic target for castration resistant prostate cancer. J Urol. 2013;190(6):2278–2287. doi:10.1016/j.juro.2013.07.001
26. Ren S, Wang F, Shen J, et al. Long non-coding RNA metastasis associated in lung adenocarcinoma transcript 1 derived miniRNA as a novel plasma-based biomarker for diagnosing prostate cancer. Eur J Cancer. 2013;49(13):2949–2959. doi:10.1016/j.ejca.2013.04.026
27. Wang D, Ding L, Wang L, et al. LncRNA MALAT1 enhances oncogenic activities of EZH2 in castration-resistant prostate cancer. Oncotarget. 2015;6(38):41045. doi:10.18632/oncotarget.5728
28. Lu X, Yang F, Chen D, et al. Quercetin reverses docetaxel resistance in prostate cancer via androgen receptor and PI3K/Akt signaling pathways. Int J Biol Sci. 2020;16(7):1121–1134. doi:10.7150/ijbs.41686
29. Sharmila G, Bhat F, Arunkumar R, et al. Chemopreventive effect of quercetin, a natural dietary flavonoid on prostate cancer in in vivo model. Clin Nutr. 2014;33(4):718–726. doi:10.1016/j.clnu.2013.08.011
30. Ma Z-S, Huynh TH, Ng CP, Do PT, Nguyen TH, Huynh H. Reduction of CWR22 prostate tumor xenograft growth by combined tamoxifen-quercetin treatment is associated with inhibition of angiogenesis and cellular proliferation. Int J Oncol. 2004;24(5):1297–1304.
31. Shoskes DA, Zeitlin SI, Shahed A, Rajfer J. Quercetin in men with category III chronic prostatitis: a preliminary prospective, double-blind, placebo-controlled trial. Urology. 1999;54(6):960–963. doi:10.1016/S0090-4295(99)00358-1
32. Hollman PC, Van Trijp JM, Mengelers MJ, De Vries JH, Katan MB. Bioavailability of the dietary antioxidant flavonol quercetin in man. Cancer Lett. 1997;114(1–2):139–140. doi:10.1016/S0304-3835(97)04644-2
33. Vaiserman A, Koliada A, Zayachkivska A, Lushchak O. Nanodelivery of natural antioxidants: an anti-aging perspective. Front Bioeng Biotechnol. 2019;7.
34. Council NR. Guide for the Care and Use of Laboratory Animals. National Academies Press; 2010.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.