Back to Journals » Journal of Inflammation Research » Volume 19
Qingyi Decoction Alleviates Alcoholic Pancreatitis by Improving Glycerolipid Homeostasis via the AMPK/SREBP-1c/PPARα Pathway
Authors Zhang J, Qi Y, Jiang Y, Wei T, Ma S, Fu K, Shen J ![]() , Chen C, Li X, Wang G, Leng A, Shang D, Qu J
, Chen C, Li X, Wang G, Leng A, Shang D, Qu J ![]()
Received 7 June 2025
Accepted for publication 27 November 2025
Published 17 March 2026 Volume 2026:19 537259
DOI https://doi.org/10.2147/JIR.S537259
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Jiahui Zhang,1,2,* Yueyi Qi,1,3,* Yuankuan Jiang,1,2,* Tianfu Wei,1,2 Shurong Ma,1,2 Kangzhe Fu,2 Jieyu Shen,1,2 Cai Chen,1,2 Xiaojie Li,1,3 Guanyu Wang,1,2 Aijing Leng,1,4 Dong Shang,1,2 Jialin Qu1
1Clinical Laboratory of Integrative Medicine, The First Affiliated Hospital of Dalian Medical University, Dalian, People’s Republic of China; 2Institute of Integrative Medicine, Dalian Medical University, Dalian, People’s Republic of China; 3Institute (College) of Pharmacy, Dalian Medical University, Dalian, People’s Republic of China; 4Department of Traditional Chinese Medicine, The First Affiliated Hospital of Dalian Medical University, Dalian, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Jialin Qu, Email [email protected] Dong Shang, Email [email protected]
Background: Alcoholic pancreatitis (AP) is one of the most common types of acute pancreatitis, which is commonly accompanied by lipid metabolism disorders. Qingyi Decoction (QYD) is a traditional Chinese medicine formula for the treatment of acute and chronic pancreatitis in clinical. However, previous studies have mostly focused on its efficacy in biliary pancreatitis, leaving its efficacy and mechanistic underpinnings in AP largely unexplored and unreported.
Methods: The mice with the AP model were established by intraperitoneal injections of a mixture of OA (150 mg/kg) and EtOH (1.35 g/kg) to evaluate the protective efficacy of QYD. The serum amylase, lipase, and pancreatic tissue myeloperoxidase (MPO) levels combined with histopathological analysis were used to evaluate the efficacy. Then, the differential lipids species associated with QYD treatment in improving AP were explored by a comprehensive lipidomics platform. Finally, the underlying mechanisms involving AMPK-mediated SREBP-1c/FASN and PPARα/CPT1A signaling pathways were systematically examined using Western blot and RT-qPCR techniques to determine both protein and mRNA expression levels.
Results: QYD administration significant reduced the serum levels of amylase and lipase as well as MPO activity in pancreatic tissue, and alleviated pancreatic tissue hispathological damage in AP mice. Additionally, QYD regulated the metabolism of lipids such as TAG, DAG and FFA, whose mechanism is closely associated with activation of AMPKα, leading to the concurrent suppression of downstream mediators in lipid synthesis (SREBP-1c, ACC1, and FASN) and enhancement of factors involved fatty acids β-oxidation (PPARα and CPT1A) at both proteins and mRNAs levels, respectively.
Conclusion: The study demonstrates that QYD could protect AP through improving lipid metabolism disorder via AMPK-mediated SREBP-1c/FASN and PPARα/CPT1A axis, which not only expands the clinical indications of QYD but also provides a novel therapeutic strategy and theoretical basis for the clinical treatment and drug development of AP.
Plain Language Summary: Qingyi Decoction (QYD) significantly reduced serum amylase and lipase levels, decreased pancreatic MPO activity, and alleviated pancreatic tissue damage in mice with alcoholic pancreatitis (AP).Improving glycerolipid homeostasis participated in the protective regulation of QYD against AP.The protection of QYD was achieved via inhibiting lipid synthesis and promoting fatty acid β-oxidation through the AMPK-mediated SREBP-1c/FASN and PPARα/CPT1A pathways.
Keywords: Qingyi Decoction, alcoholic pancreatitis, lipid metabolism, AMPK/SREBP-1c/PPARα pathway
Introduction
Acute pancreatitis is one of the common digestive system diseases caused by complex and diverse etiologies, with cholelithiasis, excessive alcohol consumption, and hyperlipidemia as the major nosogenesis,1,2 leading to approximately 220,000 people hospitalized each year.3 Meanwhile, excessive alcohol consumption has increased the risk of acute pancreatitis by 40% in the past 20 years.4 The pancreas, acting as another involved organ besides the liver, is also affected by alcohol. Excessive drinking often leads to recurrent episodes of acute pancreatitis and eventually develops into chronic pancreatitis due to the addiction of alcohol. Therefore, alcoholic pancreatitis (AP) has seriously impacted the quality of life of patients and socio-economic development.
In recent years, researchers have revealed that the imbalance of lipid metabolism is the main pathogenesis of AP. Clinical trials and animal experiments have consistently demonstrated that abnormal serum lipid metabolism in AP is characterized by elevated levels of polyunsaturated fatty acids and triglycerides (TAG). This dysregulation promotes free fatty acid (FFA)-induced toxicity and facilitates TAG accumulation, thereby exacerbating pancreatic injury.5–7 Lipids serve as essential components that maintain normal physiological functions and activities in the human body, and abnormalities in their metabolism participate in the occurrence and development of various diseases through inflammation, endoplasmic reticulum (ER) stress, oxidative stress, and autophagy inhibition, etc.8 As for the AP, the injury caused by ethanol is mainly attributed to the toxic fatty acid ethyl esters (FAEEs), which was produced by the esterification of fatty acids via non-oxidative pathways. FAEE could stimulate cholesterol ester synthesis and the release of FFAs, thereby regulating lipid metabolism. Moreover, existing studies have shown that the FAEE synthase is higher in patients with alcohol-related pancreatitis, which also induces the accumulation of FAEE and further exacerbates the severity of AP.9–11
AMP-activated protein kinase (AMPK), a “guardian” of lipid homeostasis discovered in recent years, has been considered a key enzyme that regulates cellular energy balance.12 Once activated via phosphorylation at Thr172, AMPK could promote energy metabolism by promoting fatty acid oxidation while concurrently reducing synthetic metabolism through suppressing the lipid synthesis pathways, which deems as the two critical processes in lipid metabolism.13 As its downstream regulatory factors with the same function, there has been sufficient evidence to suggest that downregulation of sterol regulatory element binding protein 1c (SREBP-1c), a master transcription factor for lipogenesis, leading to the downregulation of its downstream genes such as Acetyl-CoA carboxylase 1 (ACC1) and fatty acid synthase (FASN), which could inhibit lipid synthesis.14 Comparatively, the upregulation of peroxisome proliferatoractivated receptor alpha (PPARα) and carnitine o-palmitoyltransferase 1A (CPT1A) could induce fatty acids β-oxidation. PPARα, a nuclear receptor that transcriptionally upregulates genes involved in fatty acid oxidation (eg, CPT1A). As the rate-limiting enzyme responsible for mitochondrial fatty acid uptake, CPT1A activity is further potentiated through AMPK-mediated suppression of its inhibitor malonyl-CoA, thereby facilitating fatty acid β-oxidation.15 Their change could lead to a decrease in lipid accumulation and lipolysis both in vitro and in vivo, which makes lipid metabolism disorder tend to normal gradually.16 Accordingly, seeking effective drugs that could prevent and delay the progression of lipid disorder via regulating the above targets has exploitable potential for the treatment of AP.
Currently, the commonly adopted Western-style medicine treatment for acute pancreatitis primarily involves strategies such as inhibition of pancreatic secretion, anti-inflammatory, and antispasmodic treatment, etc.17 However, these approaches are largely supportive care in nature and are associated with a relatively high incidence of adverse reactions, as well as a substantial risk of persistent organ failure. Comparatively, traditional Chinese medicine (TCM) formula treatment has unique efficacy and advantages in the treatment of pancreatitis through clearing away heat and toxic material, invigorating the spleen, and clearing dampness, thereby decreasing the mortality rates caused by organ failure, infections, and local complications of the pancreas, improving prognosis as well as shortening the hospital stay.18 Besides, TCM has been included in Chinese guidelines for the diagnosis and treatment of acute pancreatitis (2013, Shanghai),18 which could be adjusted based on the chief complaint and accompanied symptoms of the patient by the personalized treatment principle of TCM basic theory.19 However, while these approaches are essential for managing the acute phase, they often lack the specificity required to precisely target the underlying etiology of AP—namely, the dysregulation of lipid metabolism induced by FAEEs and its subsequent pathological consequences.
Qingyi Decoction (QYD), a TCM formula used for over 60 years in our hospital, has been widely applied clinically with significant efficacy.20 It is derived from the modification of the classic prescriptions Da-Cheng-Qi Decoction and Da-Chai-Hu Decoction in Shang Han Lun, mainly composed of Rhei Radix et Rhizoma, Scutellariae Radix, Bupleuri Radix, Paeoniae Radix Alba, Aucklandiae Radix, Corydalis Rhizoma, Gardeniae Fructus and Natrii Sulfas.21 At present, QYD has demonstrated promising clinical benefits according to several observational studies and meta-analyses. Furthermore, its efficacy and mechanisms of action have been progressively validated in animal models, revealing associations with antiinflammatory effects, modulation of gut microbiota, regulation of cell apoptosis, and inhibition of ferroptosis, etc.20–24 However, current research on QYD has mainly focused on its efficacy in biliary pancreatitis, with no reported studies evaluating its therapeutic potential in alcohol-induced pancreatitis. Meanwhile, although clinical investigations have established its empirical benefits, the precise pharmacological mechanisms—particularly its effects on dysregulated lipid metabolism, a hallmark feature of pancreatitis—remain inadequately elucidated. Moreover, it is not clear whether lipid metabolism disorders participated in the protective effect of QYD against AP. Our team has characterized the chemical profile of QYD systematically and identified its chemical composition.21 Some identified components (baicalin, baicalein, emodin and quercetin) could promote lipid decomposition and inhibit lipid synthesis via regulating targets such as AMPK, SREBP-1c, CPT1A, etc.25–28 Combining the published efficacy of composed herbs (Scutellariae Radix, Bupleuri Radix, Gardeniae Fructus) on relieving alcohol toxicity,29 we hypothesize that QYD may have a wider range of clinical indications for the pancreatic diseases caused by other triggers.
Therefore, this study aims to verify whether QYD could protect against AP by evaluating the biochemical parameters in serum (amylase and lipase) and pancreatic tissue (myeloperoxidase, MPO) as well as histopathological analysis based on the established mice model with AP. Furthermore, the serum samples of mice were analyzed by a lipidomics platform based on an ultra-performance liquid chromatography-mass spectrometer (UPLC-MS/MS) to investigate the regulatory effects of QYD on lipid metabolism. Finally, Western blot and RT-qPCR were used to detect the expression of proteins and mRNAs in key metabolic pathways to verify the prediction results.
Methods and Materials
Preparation of QYD Extract and UHPLC-QTOF-MS Analysis
QYD is composed of eight herbs as shown in Table 1. They were obtained from the First Affiliated Hospital of Dalian Medical University (Dalian, Liaoning, China). The refluxing extraction was used for the preparation of QYD. Firstly, Chaihu, Huangqin, Muxiang, Yanhusuo, Zhizi, and Baishao in the prescription were soaked in a 10-fold mass of water (m/v =1:10, 870 mL) for 1 h, subjected to hot reflux extraction for 1 h, and filtered (through six layers of absorbent gauze) sequentially, which yields the solution I. The residue was further extracted with an 8-fold mass of water for 0.5 h using the same method, and Dahuang was then added to the extract and co-extracted for 0.5 h. After filtration, the solution II was obtained. The two extracts were combined, and Mangxiao was added while hot. The combined solution was concentrated to 120 mL (equal to 1 g of crude drug/mL) under a rotary evaporator and lyophilized using a vacuum freeze drier. The extract was obtained with a yield of 32.5% and kept in the desiccator before use.
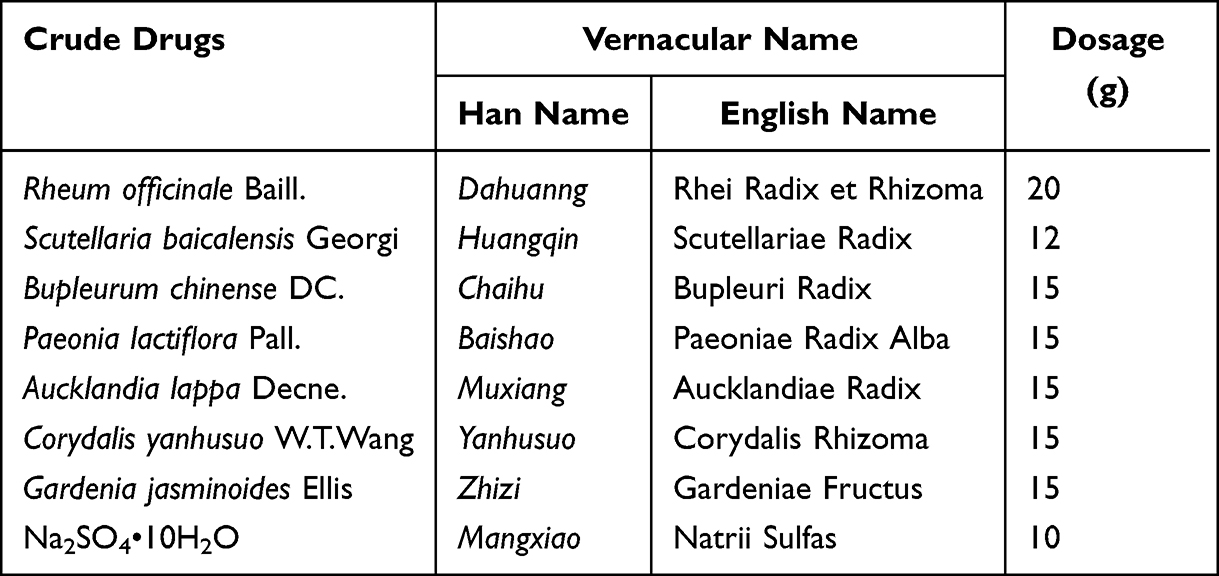 |
Table 1 The Name of Crude Drugs and Their Dosage |
The chemical profile of prepared freeze-dried powder was determined using UHPLC/QTOF-MS after ultrasound extraction with 80% methanol. The condition of chromatographic separation and mass spectrometry detection was the same as the method mentioned previously.20 The chemical profile and composition information of QYD is shown in Figure S1 and Table S1.
Animal Model and Drug Treatments
All animal experiment procedures were approved by the Animal Ethics Committee of Dalian Medical University (License number: AEE22119), and conducted in accordance with International Council for Laboratory Animal Science Basic Principles for Animal Welfare. Male C57BL/6 mice (23–25 g) were purchased from Liaoning Changsheng Bioscience Co., Ltd. (Liaoning, China). Mice were adaptively fed in a breeding room where controlled temperature (22 ± 2°C), humidity (55 ± 5%), lighting (12 h light-dark cycles), and libitum access to water and standard laboratory chow for one week.
The mice were randomly divided into five experimental groups (15 mice per group) employing a computer-generated random number table to ensure unbiased allocation, including control (CON), model (OA/EtOH), QYD low-dose (QYD-L, 15.21 g/kg), QYD high-dose (QYD-H, 30.42 g/kg), and dexamethasone (DEX). The animal model of AP was established following the protocol established by Huang et al9 which involved sequential intraperitoneal injections of a mixture of OA (150 mg/kg) and EtOH (1.35 g/kg) at hourly intervals. All mice were fasted for 12 hours before modeling. The CON, OA/EtOH, and DEX groups were given double-distilled water at the dose of 0.015 mL/g body weight by intragastrical administration one hour before modeling, and QYD-L and QYD-H groups received QYD by the same method. Then, 200 μL of normal saline was injected immediately prior at the injection site to avoid potential local damage by ethanol to peritoneal organs, and intraperitoneal injection of OA/EtOH injections was followed to induce the model. The dexamethasone group was given intraperitoneal injection of dexamethasone (5 mg/kg, concentration 5 mg/mL) immediately after the first injection of modeling drugs.
After modeling 12 h, serum and pancreatic tissues of mice from each group were collected (Figure 1).
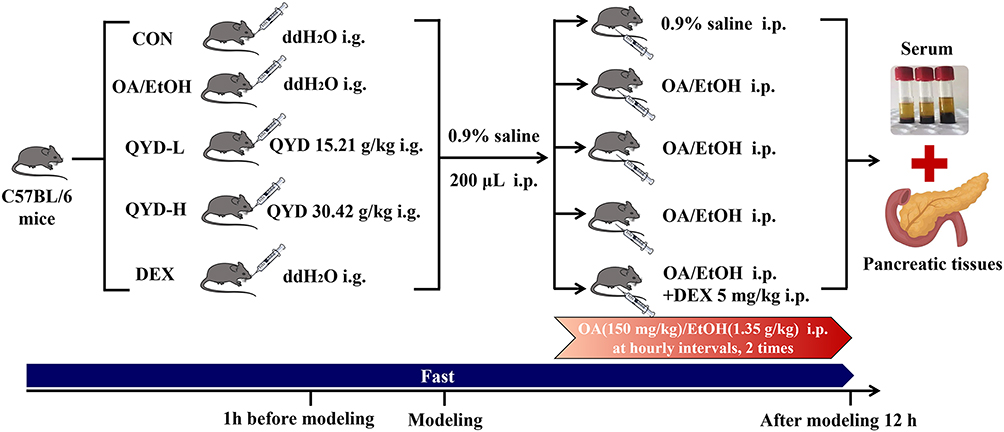 |
Figure 1 Schematic overview of the experimental design and procedures used in the animal experiment. |
Serum Amylase, Lipase, and Pancreatic Tissue Myeloperoxidase (MPO) Test
The levels of serum amylase and lipase in each group were measured by α-amylase and lipase assay kit (Nanjing, China). The MPO of pancreatic tissue was measured by an MPO assay kit according to the product instructions.
Histopathology
After euthanizing the mice, the pancreatic tissues were fixed by perfusion with 4% paraformaldehyde. Subsequently, ethanol gradient treatment was performed followed by embedding in paraffin. Paraffin sections with a thickness of 4 μm were prepared using a paraffin microtome, followed by hematoxylin and eosin (H&E) staining. To observe the changes in the pancreatic tissue structure, the sections were analyzed under a light microscope.
Serum Samples Preparation and Detection for Lipidomics Analysis
The methyl tert-butyl ether (MTBE) extraction method was used to extract the lipids from serum as described in Figure 2. Besides, the preparation of quality control (QC) samples was carried out by combining equal aliquots of 5 μL from each sample and pretreated in the same way, and the QC sample was injected before, during and after the analyzed sequence.
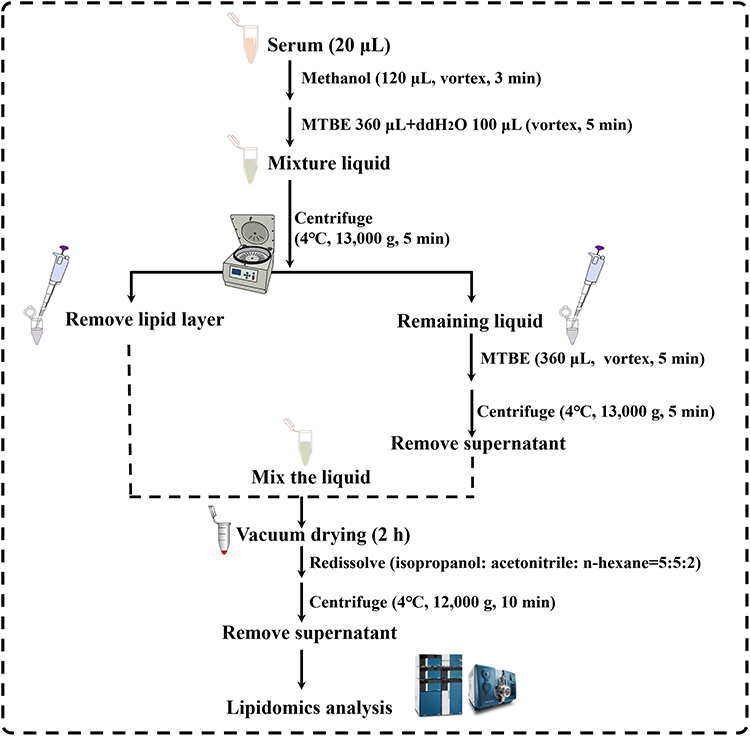 |
Figure 2 Workflow diagram illustrating the preparation of serum samples for lipidomic analysis. |
The SCIEX JasperTM HPLC coupled with a triple quadrupole mass spectrometer (AB SCIEX, Framingham, MA, USA) was used to analyze the serum samples, and data were collected using the selected reaction monitoring (SRM) mode. Waters ACQUITY UPLC® HSS C18 column (2.1 × 100 mm, 1.8 μm) was used for the chromatographic separation. The elution adopts a binary gradient system, where phase A (acetonitrile: water = 60:40, v: v) and phase B (isopropanol: acetonitrile = 90:10, v: v) both containing 5 mM ammonium acetate severed as the mobile phase. The gradient elution program was as follows: 0–2 min, 20% B; 2–5 min, 20–70%B; 5–18 min, 70–95%B; 18–18.6 min, 95%B; 18.6–19 min, 95–20%B; 19–22 min, 20%B.
MS/MS data in both positive and negative ion modes were acquired by Analyst MD Software 3.0.3 (AB SCIEX, USA), and the total ion chromatogram was processed for noise filtering, baseline correction, chromatographic peak identification, retention time correction, peak alignment as well as peak integration using MultiQuant MD 3.0.3 (AB SCIEX, USA). The detected ions were extracted into an Excel file in a two-dimensional matrix format, including mass-to-charge ratio (m/z), retention time, peak area, etc.
Data Processing
The data and method were evaluated using the IPOS website (http://82.157.20.231:3838/ipos/). QC samples demonstrated <30% coefficient of variation (CV) after rigorous filtering.
Multivariate analysis was conducted through MetaboAnalyst 6.0 (https://www.metaboanalyst.ca/) software was used for data normalization. Subsequently, samples from different groups were analyzed by principal component analysis (PCA), partial least squares-discriminant analysis (PLS-DA), and orthogonal partial least squares-discriminant analysis (OPLS-DA) to summarize their differences and filter out the potential biomarkers preliminarily. R2Y and Q2 are two major parameters that were used to assess the quality of the statistical model. Then, lipid molecules with variable importance in the project (VIP) >1, P value <0.05, and fold change (FC) >2 or FC <0.5 were screened out as significant lipid differential metabolites. Finally, clustering analysis was performed based on the screened results to demonstrate the changes in differential metabolites and clustering patterns more comprehensively and intuitively.
RT-qPCR
The total RNA was extracted from pancreatic tissue using the TRIzol method. Following quantification based on purity and concentration measurements, they were reversely transcribed into cDNA in a 20 μL reaction system according to kit instructions. The relative mRNA expression levels of target genes were quantified using the comparative 2–ΔΔCt method, with β-actin serving as the reference gene. The primers were purchased from Sangon Biotech Biological Co., Ltd. (Shanghai, China), and their sequences were listed in Table 2.
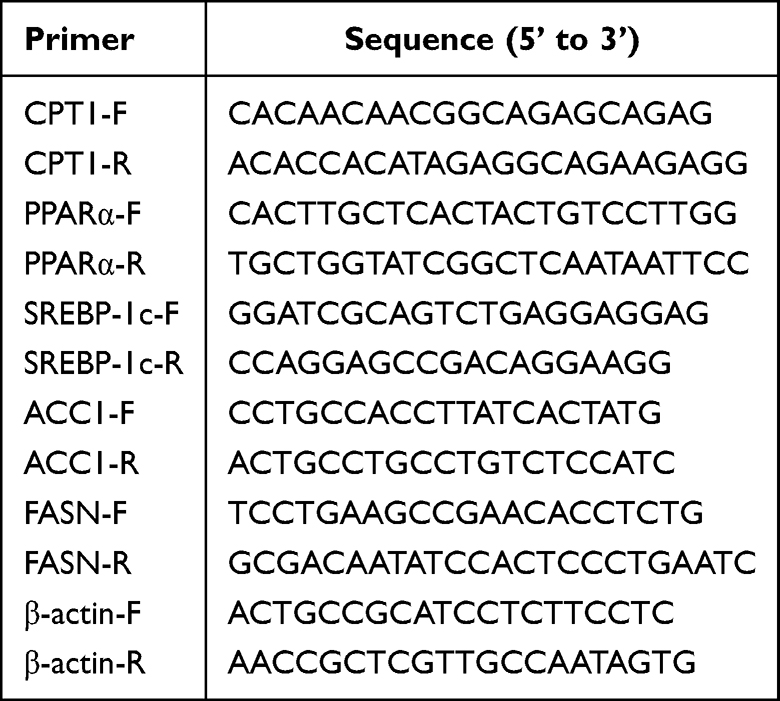 |
Table 2 Primer Sequences |
Western Blot
Protein extraction from pancreatic tissue derived from C57BL/6 mice in each group was carried out using a whole protein extraction kit (Solarbio, Nanjing). After centrifugation (4°C, 12,000 rpm, 10 min), the supernatant of the protein sample was collected and concentration was quantitatively determined using the BCA method. After protein denaturation, SDS-PAGE was performed and the protein on the PAGE glue was transferred onto the polyvinylidene fluoride membrane. Subsequently, membranes were blocked with a 5% skim milk solution for 1.5 h. Antibodies AMPKα (Rabbit, Proteintech, 1:1000), p-AMPKα (Rabbit, CST, 1:1000), SREBP-1c (Rabbit, Affinity, 1:1000), ACC1 (Rabbit, ABclonal, 1:1000), FASN (Rabbit, Proteintech, 1:5000), PPARα (Rabbit, Proteintech, 1:1000) and CPT1A (Mouse, Affinity, 1:5000) were added overnight at 4°C, and horseradish peroxidase (HRP) conjugated secondary antibodies (ABclonal, 1:10000) was incubated for 1 h at room temperature. The PVDF membranes were subjected to ultrasensitive ECL luminescent solution, exposure, and photography under the gel imaging system. All Western blot data were normalized to a corresponding housekeeping protein (β-actin). The band intensities of target proteins were quantified using ImageJ software and are expressed as the ratio of target protein intensity to β-actin intensity.
Statistical Analysis
Statistical analyses were carried out using GraphPad Prism software 8.0.2 (GraphPad Software, SanDiego, CA), and the data were presented as mean ± standard deviation (SD). Statistical significance was calculated by 2-tailed Student’s t-test or one-way analysis of variance (ANOVA) with Dunnett’s multiple-comparison test. A threshold of P < 0.05 was considered statistically significant. Significance levels in the figures were denoted as follows: P < 0.05 (*), P < 0.01 (**), P < 0.001 (***). To mitigate potential bias, all personnel involved in experimental interventions and outcome assessments were blinded to group assignments throughout the study. Additionally, data analysts were kept blinded to the experimental conditions during statistical evaluation.
Results
QYD Decreased the Levels of Amylase, Lipase, and MPO in OA/EtOH-Induced AP Mice
Serum amylase, lipase, and pancreatic tissue MPO were measured to evaluate the effect of QYD on the pancreatic injury degree of mice. As shown in Figure 3(A–C), the levels of amylase, lipase, and MPO in mice with the OA/EtOH group were significantly increased compared to the CON (P<0.001), indicating that the model has been successfully established.
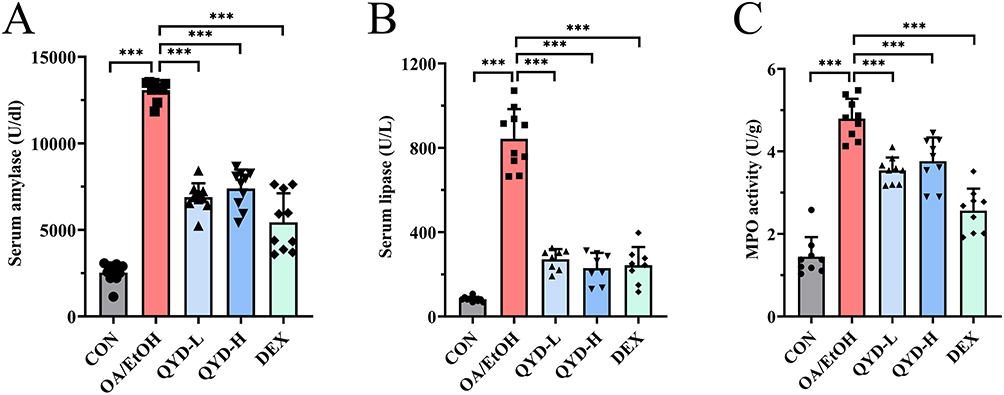 |
Figure 3 The protective effects of QYD against OA/EtOH-induced AP. (A) serum amylase activity. (B) serum lipase activity. (C) pancreatic tissue myeloperoxidase activity. (n=8~10, ***P<0.001). |
Compared to the OA/EtOH, all these biochemical parameters were significantly decreased by the QYD-L, QYD-H, and DEX (P<0.001). Moreover, the effects of QYD-L and QYD-H are equivalent, while they were slightly weaker than DEX for the amylase and MPO levels.
QYD Alleviated Pancreatic Tissue Damage in OA/EtOH-Induced AP Mice
Histopathological changes in the pancreas were visualized by H&E staining. The pancreatic tissues of mice in the control group showed normal morphology, whereas edema (red arrow), inflammatory infiltration (blue arrow), and significant necrosis (green arrow) were observed in the OA/EtOH group. However, the necrosis of pancreatic tissue in the QYD-L, QYD-H, and DEX groups was significantly alleviated compared to the OA/EtOH, and the severity of inflammatory infiltration, edema, and vacuolization was also improved, indicating that QYD and dexamethasone could improve pancreatic tissue damage to a certain extent (Figure 4).
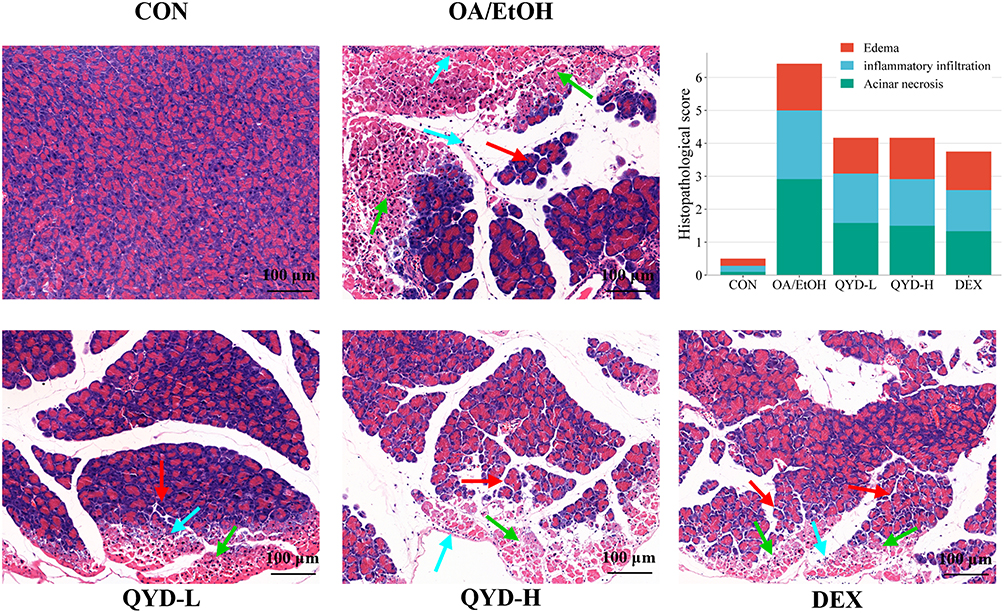 |
Figure 4 H&E staining and pathological scoring of pancreas tissue in each group. Red arrow points to edema, blue arrow points to inflammatory infiltration, green arrow points to acinar necrosis. (n=6, Scale bar: 100 μm). |
Combining the results of the previous chemical index detection, we found that the efficacy between the QYD-L and QYD-H groups is equivalent. Therefore, the samples from the QYD-L group (ie, the normal dose equivalent to clinical efficacy) were employed for further serum lipidomics analysis and named “QYD”.
Stability and Repeatability of the Serum Lipidomics Analytical Method
To investigate the stability and reliability of the established method, QC samples were inserted across the analytical batch. Briefly, each group samples (n=14) were interspersed with a QC sample. The Spearman correlation coefficient analysis of QC samples conducted through the IPOS was used for evaluating the stability of samples and apparatus. As shown in Figure S2, the correlation coefficient between the first and last QC samples is 0.9957, indicating a strong correlation between them and great reproducibility. Besides, the relative bias (Figure S3) and proportion of relative standard deviations lower than 30% (Figure S4) for QC samples were, respectively, smaller than 3.0% and more than 90%.
Identified Lipid Metabolites Based on the LC-MS/MS Method
Based on LC-MS/MS, a total of 897 lipid metabolites were detected from four groups of samples (a total of 56) in both positive and negative ion modes. The classification and quantity of each lipid metabolite are shown in Figure 5, among which TAG (48.05%), PE (11.15%), PC, and SM (both 6.13%) rank among the top 3 subclasses.
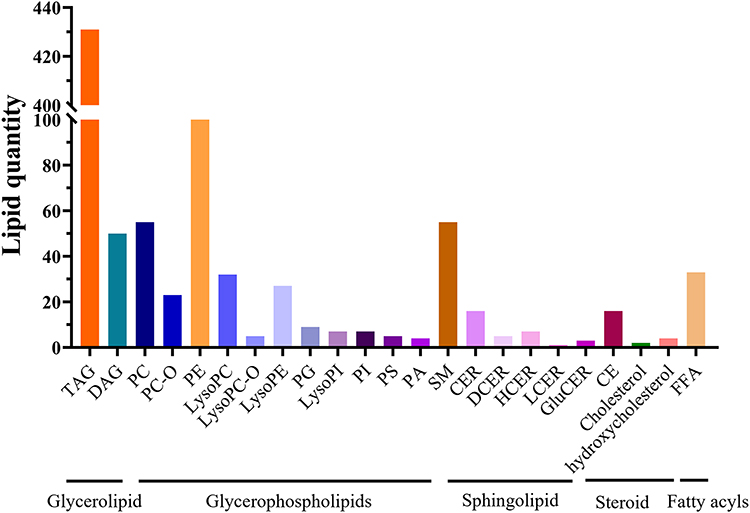 |
Figure 5 The identified lipid species. |
QYD Modulates OA/EtOH-Disrupted Glycerolipid Homeostasis
The differential expression of lipid molecules detected in the serum among different groups was analyzed. As presented in volcano plots (Figure 6A–C), 448 lipid molecules with significant differences were screened when comparing the OA/EtOH with the CON group, among which 431 were upregulated and 17 were downregulated. Besides, 116 lipid molecules with significant differences were screened between the QYD and OA/EtOH groups, among which 11 were upregulated and 105 were downregulated. Meanwhile, 334 lipid molecules with significant differences were screened between the DEX and OA/EtOH groups, among which 9 were upregulated and 325 were downregulated.
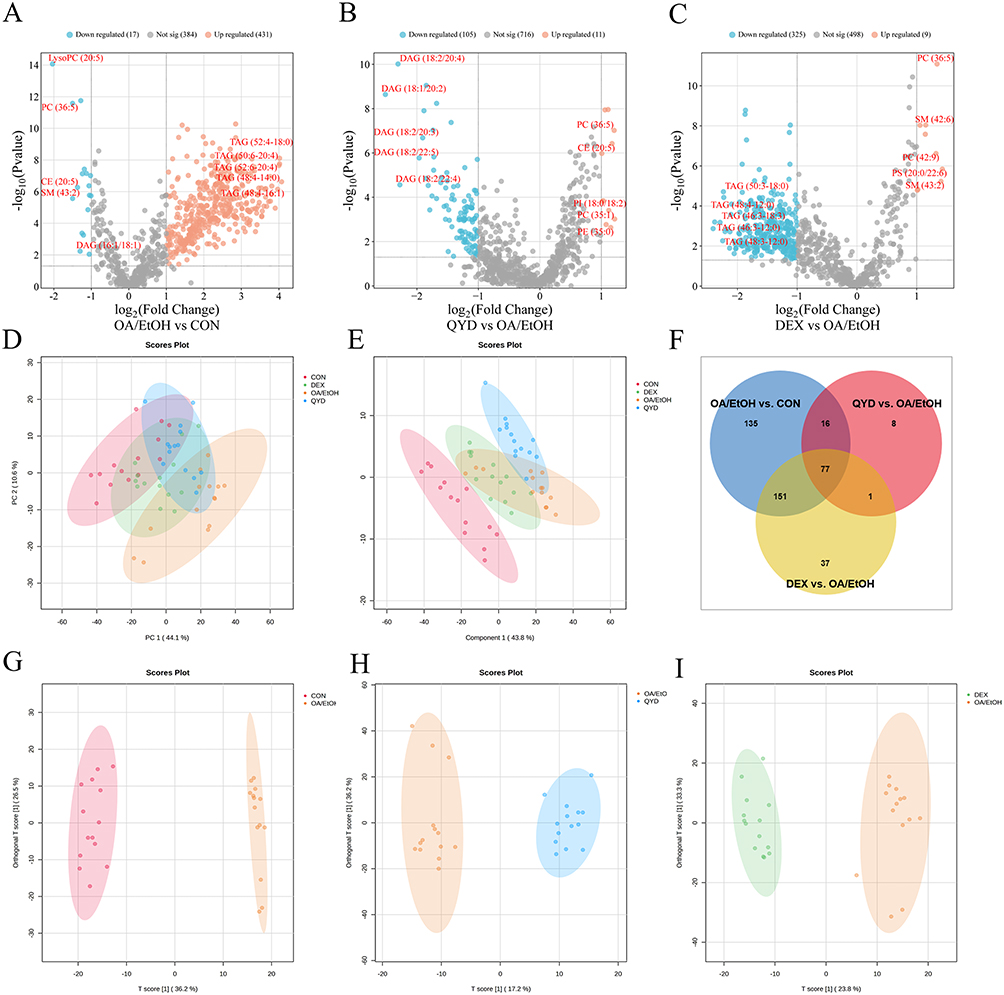 |
Figure 6 Comparative lipidomic profiling across experimental groups. (A-C) Volcano plots. (D) PCA score plots. (E) PLS-DA score plots. (F) Venn diagram of differential lipid metabolites. (G-I) OPLS-DA score plots. |
Furthermore, an overview of the lipidomics profiles in all groups was obtained by multidimensional statistical analysis. Unsupervised PCA (Figure 6D) and PLS-DA (Figure 6E) showed that the CON and OA/EtOH groups can be well separated. Meanwhile, the QYD and DEX groups tended to be closer to the CON compared to the OA/EtOH, indicating their intervention on lipid metabolism in AP mice to a certain extent. OPLS-DA (Figure 6G–6I) analysis showed an obvious diversity of lipid metabolites between the two groups. Moreover, no over-fitting was observed in the PLS-DA and OPLS-DA models according to the Cross-Validation and permutation test (Figure S5).
The lipid molecules with VIP>1, P value<0.05, FC>2.0 or FC<0.5 were screened for differential lipid metabolites. As shown in Figure 6F, 379, 102, and 266 differential metabolites between CON vs OA/EtOH, QYD vs OA/EtOH, and DEX vs OA/EtOH groups were screened. And 93 differential metabolites were shared between the CON, QYD, and OA/EtOH groups (Table 3). Among them, TAG and DAG are the main species, while other lipid molecules such as FFA (24:4), PC (36:5), LysoPC (20:5), LysoPE (20:5), SM (43:2), and CE (20:5) also show significant changes (Figure 7A–7H). Additionally, the top 20 metabolites with VIP values were displayed in the heatmap to more intuitively illustrate the regulatory effect of QYD on lipid metabolism (Figure 7I). Finally, enrichment analysis shows that QYD can improve the lipid synthesis and decomposition imbalance induced by OA/EtOH, more are reflected in glycerolipid metabolism (Figure 7J).
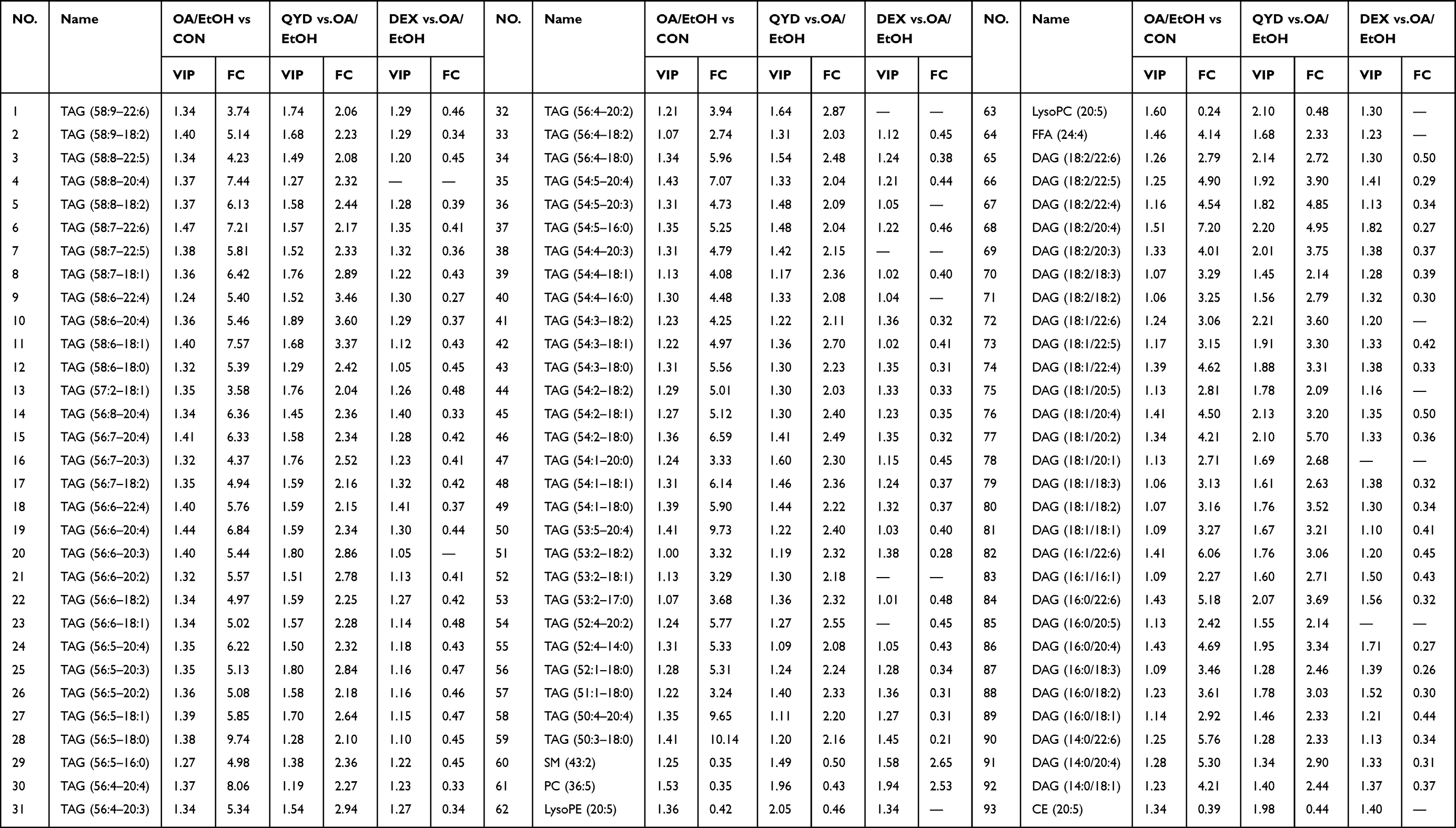 |
Table 3 Differential Lipid Metabolites of the Mice Serum in Each Group |
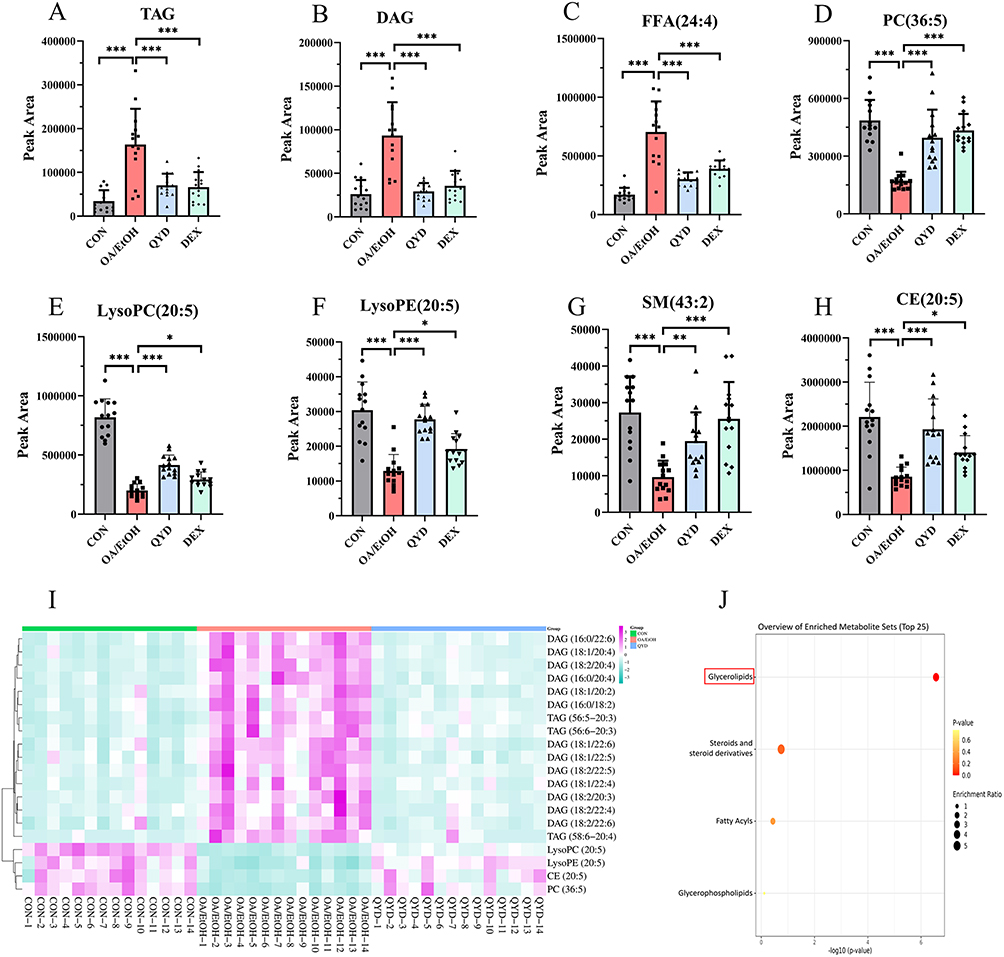 |
Figure 7 Visualization of differentially abundant lipid metabolites. (A–H) Peak areas of candidate differentially abundant lipid metabolites. (I) Heatmap visualization of the top 20 differentially abundant lipid metabolites across the CON, OA/EtOH, and QYD groups (n=14, *P<0.05, **P<0.01, ***P<0.001). (J) Enrichment analysis, the red colored rectangular box highlights the most significantly enriched metabolite. |
QYD Regulates the AMPK/SREBP-1c/PPARα Pathway in OA/EtOH-Induced AP
In order to identify the potential mechanisms by which QYD improves lipid metabolism disorder and to validate our hypotheses, the mRNA expression of key signaling molecules related to lipid metabolism including SREBP-1c, ACC1, FASN, PPARα, and CPT1A we conducted by RT-qPCR analysis experiments.
The expression of lipid synthesis-related mRNA SREBP-1c (P<0.001), ACC1 (P<0.01), and FASN (P<0.001) in the OA/EtOH were significantly higher than those in the CON, while they were significantly decreased by the QYD (P<0.05 or P<0.01 or P<0.001), but no significant decrease was observed in the DEX. Comparatively, the expression of fatty acid β-oxidation related mRNA PPARα and CPT1A in the OA/EtOH were significantly lower than those in the CON (P<0.001), while they were upregulated by both QYD and DEX intervention (P<0.01 or P<0.001). These data suggested that phosphorylation of AMPKα, inhibition of SREBP-1c/FASN and activation of PPARα/CPT-1A signaling pathway participated in the mechanism of QYD on AP (Figure 8).
 |
Figure 8 (A–E) The mRNA levels of SREBP-1c, ACC1, FASN, PPARα, and CPT1A in pancreatic tissues measured by RT-qPCR (n=3, *P<0.05, **P<0.01, ***P<0.001, ns indicates no statistical significance). |
To validate our hypothesis more comprehensively and the reliability of the Western blot results, the protein levels of AMPKα, p-AMPKα, and the above signaling molecules were conducted (Figure 9). As expected, the expression levels of p-AMPKα/AMPKα in the OA/EtOH group were significantly reduced compared to the CON group (P<0.01), while they were increased in the QYD-L, QYD-H, and DEX groups (P<0.01 or P<0.05). SREBP-1c, ACC1, and FASN in the OA/EtOH group were significantly increased in the CON (P<0.001 or P<0.01), while they were also reduced in the QYD (P<0.05 or P<0.01). Concerns were raised that the DEX group had no significant difference compared with the OA/EtOH group.
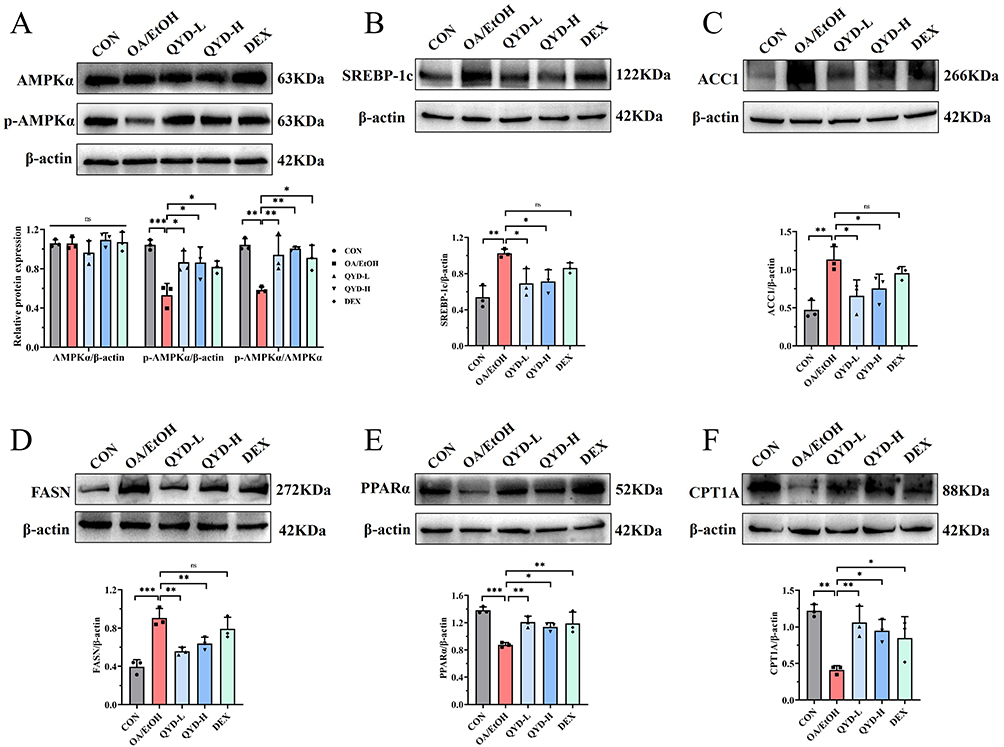 |
Figure 9 (A–F) The protein levels of AMPKα, p-AMPKα, SREBP-1c, ACC1, FASN, PPARα, and CPT1A in pancreatic tissues detected by Western blot (n=3, *P<0.05, **P<0.01, ***P<0.001, ns indicates no statistical significance). |
The levels of PPARα and CPT1A in the OA/EtOH were significantly downregulated compared to the CON (P<0.01 or P<0.001), while they were significantly downregulated in the QYD-L, QYD-H, and DEX groups (P<0.05 or P<0.01).
Discussion
Alcohol abuse stands as one of the most prevalent causes of pancreatitis.30 Contemporary epidemiological surveys indicate a steady rise in the incidence of acute pancreatitis over the years, rendering it one of the foremost considerations in adult acute or chronic pancreatitis.31 Furthermore, an increasing body of research suggests a close association between the onset and progression of AP and lipid metabolism disorders.5 However, due to a limited understanding of its mechanisms and pain origins, there are currently no effective drugs for preventing and treating AP, and existing medications for AP carry many adverse reactions.32 Hence, the imperative lies in developing novel methods for preventing and treating AP.
TCM formulas, recognized for their multi-target effects and favorable safety profiles, have demonstrated considerable potential in recent years for the prevention and treatment of pancreatitis.20 According to TCM theory, AP stems from the exposure to alcohol toxins that impair the intestines and stomach, resulting in a conflict with digestive qi and the subsequent accumulation of dampheat. The core pathogenesis is characterized by an interplay between damp-heat patterns and spleen deficiency.33 QYD, derived from the modifications of Da-Cheng-Qi-Decoction and Da-Chai-Hu-Decoction in Zhang Zhongjing’s Shang Han Lun, is believed to soothe the liver and regulate qi, clear away heat and toxic material, as well as purgation.34 Over the past decade, clinical and basic research on QYD has confirmed its effectiveness in the treatment of acute pancreatitis, including its anti-inflammatory effects (TNF-α, IL-6, NF-κB p65), regulation of cell apoptosis and intestinal flora, as well as inhibition of ferroptosis.20–24 Moreover, the monomeric components of QYD, such as baicalin, baicalein, emodin and quercetin, also regulate lipid metabolism through AMPK, SREBP-1c, CPT1A, etc.25–28 These findings suggest that the pharmacological mechanisms of QYD are not limited to what is described above, but perhaps also related to regulating lipid metabolism.
Considering that lipid metabolism is also a pioneer pathogenesis of AP,5 our focus is attracted to the QYD regulating lipid metabolism in AP. However, the efficacy of QYD in preventing and treating AP and its potential pharmacological mechanisms remain unclear. To investigate the effects of QYD on AP, this study utilized a lipidomics platform developed by UPLC-MS/MS and molecular biology techniques (Western blot and RT-qPCR) based on the OA/EtOH mice model to elucidate the pharmacological effects and mechanisms of QYD.
Amylase, lipase, and MPO are commonly used indicators in clinical practice to reflect pancreatitis,35 while pancreatic tissue histopathology examination serves as the gold standard for assessing the severity of pancreatic injury, widely applied in the evaluation of pancreatic injury animal models. In this study, the levels of amylase, lipase, and MPO in the model group of mice were significantly elevated, consistent with the experimental results of Huang Wei and others. H&E of pancreatic tissue confirmed that QYD intervention could reverse the damage, which is manifested by improved edema, necrosis, inflammation, and vacuolization. Moreover, the lack of obvious dose dependency between low and high doses of QYD also suggests the rationality of clinical dosages of QYD.
Lipidomics can be used to identify differential lipid metabolites.36 Given the association between the occurrence and development of AP and lipid metabolism disorders, lipidomics analysis was conducted. By comparing the changes in lipid subclasses between the QYD group and the OA/EtOH group, important lipid subclasses potentially involved in relevant biological processes were screened (Figure 5). We found that there were 93 differential metabolites shared by the CON group, QYD group, and OA/EtOH group, belonging to 8 lipid subclasses (TAG, DAG, FFA, CE, LPE, PC, LPC, SM). TAG, the predominant form of stored fat in adipocytes, constitute a major energy reservoir and are hydrolyzed into DAG and FFA to supply energy upon demand.37,38 Meanwhile, DAG serves multiple biological roles, such as acting as a structural component of cellular membranes, a precursor for glycerol phospholipid synthesis, and a lipid second messenger in cellular signaling pathways.39 Likewise, our research indicated that the reduction of TAG, DAG, and FFA contributes to alleviating pancreatic inflammation and tissue damage, as well as reducing the risk of pancreatic steatosis. Additionally, as important components of the cell membrane, CE, SM, PC, LPC, and LPE all play a crucial role in cell signal transduction, metabolic function regulation, and eventually maintaining the stability and physiological function of the cell membrane.40,41 Therefore, changes in the relative peak areas of the above lipid subclasses may reflect alterations in lipid functions. In this study, the significantly reduced TAG, DAG, and FFA levels suggested that QYD could improve the imbalance of lipid synthesis and decomposition caused by AP, especially embedded in the glycerolipid metabolism.
Meanwhile, dexamethasone—a widely used clinical agent for treating inflammatory, allergic, neoplastic, and autoimmune disorders—has been reported to modulate fatty acid metabolism.42 Our results revealed that dexamethasone also regulates TAG and DAG levels, with a more pronounced downregulatory effect on specific lipid species including TAG (51:1–18:0), TAG (53:1–18:2), and DAG (16:0/18:3) compared to the QYD group. Although dexamethasone is generally associated with elevated triglycerides and FFAs in clinical contexts, studies have reported conflicting effects under specific conditions. For instance, tumor-bearing mice treated with high-dose dexamethasone exhibited significantly reduced serum levels of triglycerides and FFA, along with increased hepatic PPARα expression.43 Furthermore, Djouadi et al demonstrated that dexamethasone activated PPARα gene expression in both the renal cortex and medulla in rats.44 These findings suggest that the dual-regulatory role of dexamethasone in lipid metabolism may be dose- and context-dependent. As for the present research, dexamethasone appears to promote lipid breakdown likely through enhancing the fatty acid β-oxidation pathway, thereby modulating TAG and other lipid levels. RT-qPCR and Western blot analyses support this mechanism that dexamethasone did not trigger significant changes in the expression levels of SREBP-1c, FASN, and ACC1, whereas PPARα and CPT1A were upregulated. This indicates that dexamethasone modulates lipid homeostasis primarily through stimulating fatty acid oxidation rather than suppressing synthesis. These results also imply that the mechanism of dexamethasone may differ between single high-dose and chronic administration regimens. However, this hypothesis requires further experimental validation.
AMPKα, a crucial regulator of energy metabolism homeostasis, plays a significant role in the pathogenesis of various diseases.45 Srinivasan et al explored the role of AMPKα in ER stress and its connection with the occurrence and progression of chronic AP in vitro.46 The results revealed that AMPKα inactivation participated in the acinar cell damage induced by ethanol, which leads to pancreatitis. Furthermore, AMPKα activation can weaken the formation of FAEEs, ER stress, oxidative stress, lipid synthesis, and promote lipid decomposition, thereby improving inflammatory responses and mitochondrial dysfunction.47,48 Consistent with this, up-regulated expression levels of AMPKα and its downstream targets (SREBP-1c, ACC1, and FASN) as well as down-regulated levels of PPARα and CPT1A were observed in the model group in the present study, and QYD intervention could reverse these changes. In summary, the above results suggest that QYD activates AMPKα to inhibit lipid synthesis, promote fatty acid decomposition and oxidation, and alleviate lipid toxicity and lipid metabolism disorders, ultimately delaying the occurrence and development of AP.
Despite the promising findings, this study has several limitations. First, mechanistic insights are primarily derived from an in vivo model, and future studies utilizing isolated pancreatic acinar cells or other in vitro systems are warranted to identify direct cellular targets and elucidate finer molecular events. Second, the lipidomics analysis conducted in this study was untargeted. Although this strategy offers a broad and unbiased overview of lipid profiles, further targeted validation employing authentic standards is necessary to definitively verify changes in individual lipid species. Third, owing to its multi-herb compatibility nature, QYD is characterized by a highly complex chemical composition.21,49 Although a standardized extraction procedure was followed, batch-to-batch variability remains a practical consideration. Moreover, the specific bioactive constituents responsible for QYD’s therapeutic effects against alcohol-induced pancreatitis not yet been fully elucidated. Future investigations should prioritize the identification and functional characterization of the key active compounds in QYD to enhance reproducibility and establish stringent quality control standards. Furthermore, it remains to be established whether QYD ameliorates AP through additional AMPKα-dependent mechanisms (eg, modulation of ER stress and autophagy) and whether long-term administration or combination strategies, especially with antiinflammatory agents, may lead to enhanced or synergistic therapeutic effects. From a translational standpoint, although human pharmacokinetic studies and early-phase clinical trials for QYD have not yet been initiated, such clinical assessments are essential to advance its potential therapeutic application. Our research team is actively seeking opportunities to launch these critical investigations.
Conclusions
This study demonstrates that QYD exerts a protective effect against AP through improving lipid metabolism dysfunction via AMPK-mediated SREBP-1c/FASN and PPARα/CPT1A pathway (Figure 10). Notably, our findings underscore the therapeutic potential of targeting lipid metabolism in AP. These results not only provide an experimental and theoretical groundwork for the expanded application of QYD in pancreatitis treatment but also offer novel perspectives for clinical intervention and drug development. Moreover, the multi-target and favorable safety profile of QYD represent a distinct advantage over single-target agents. We believe that modulation of lipid metabolism holds substantial translational promise, and further clinical trials evaluating QYD and other therapeutics agents targeting this pathway are warranted.
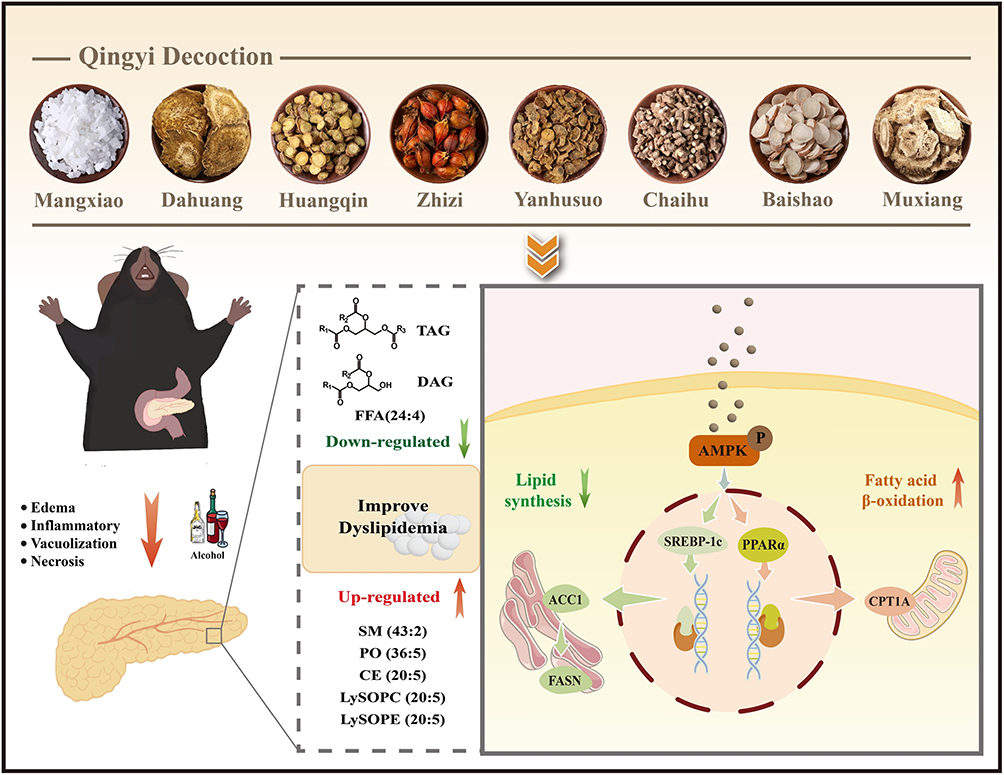 |
Figure 10 The molecular mechanism underlying the alleviation of AP by QYD. |
Abbreviations
ACC1, acetyl-CoA carboxylase 1; OA, Oleic Acid; AMPK, AMP-activated protein kinase; OPLS-DA, orthogonal partial least squares-discriminant analysis; AP, alcoholic pancreatitis; PCA, principal component analysis; CPT1A, carnitine palmitoyl transferase 1A; PLS-DA, partial least squares-discriminant analysis; CV, coefficient of variation; PPARα, peroxisome proliferator-activated receptor alpha; DEX, dexamethasone; QC, quality control; EtOH, ethyl Alcohol; QYD, Qingyi Decoction; FAEEs, fatty acid ethyl esters; RSD, relative standard deviation; FASN, fatty acid synthase; RT-qPCR; real-time fluorescence quantitative technology; FC, fold change; SREBP-1c, sterol regulatory element binding protein 1c.
H&E, hematoxylin and eosin; TCM, traditional Chinese medicine; MPO, myeloperoxidase; VIP, variable importance in the project.
Data Sharing Statement
Data supporting the findings of this study will be made available upon reasonable request on Jialin Qu ([email protected]). The complete raw and processed data from lipidomics analysis to the OMIX database (https://ngdc.cncb.ac.cn/omix/) under the accession number OMIX013713.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Natural Science Foundation of Liaoning Province (No. 2023-MSLH033), National Natural Science Foundation of China (No. 82374248), Dalian Medical University Interdisciplinary Research Cooperation Project Team Funding (No. JCHZ 2023005), and Life and Health Guidance Program Project of Dalian.
Disclosure
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
1. Colvin SD, Smith EN, Morgan DE, et al. Acute pancreatitis: an update on the revised Atlanta classification. Abdom Radiol. 2020;45(5):1222–19. doi:10.1007/s00261-019-02214-w
2. Valverde-López F, Martínez-Cara JG, Redondo-Cerezo E. Acute pancreatitis. Med Clin. 2022;158(11):556–563. doi:10.1016/j.medcli.2021.12.012
3. Mederos MA, Reber HA, Girgis MD. Acute Pancreatitis: a Review. JAMA. 2021;325(4):382–390. doi:10.1001/jama.2020.20317
4. Iannuzzi JP, King JA, Leong JH, et al. Global incidence of acute pancreatitis is increasing overtime: a systematic review and meta-analysis. Gastroenterology. 2022;162(1):122–134. doi:10.1053/j.gastro.2021.09.043
5. Khan J, Solakivi T, Seppänen H, et al. Serum lipid and fatty acid profiles are highly changed in patients with alcohol induced acute pancreatitis. Pancreatology. 2012;12(1):44–48. doi:10.1016/j.pan.2011.12.006
6. Marosvolgyi T, Horvath G, Dittrich A, et al. Fatty acid composition of plasma lipid classes in chronic alcoholic pancreatitis. Pancreatology. 2010;10(5):580–585. doi:10.1159/000289466
7. de Oliveira C, Khatua B, Noel P, et al. Pancreatic triglyceride lipase mediates lipotoxic systemic inflammation. J Clin Invest. 2020;130(4):1931–1947. doi:10.1172/JCI132767
8. Yoon H, Shaw JL, Haigis MC, et al. Lipid metabolism in sickness and in health: emerging regulators of lipotoxicity. Mol Cell. 2021;81(18):3708–3730. doi:10.1016/j.molcel.2021.08.027
9. Huang W, Booth DM, Cane MC, et al. Fatty acid ethyl ester synthase inhibition ameliorates ethanolinduced Ca2+-dependent mitochondrial dysfunction and acute pancreatitis. Gut. 2014;63(8):13131324. doi:10.1136/gutjnl-2012-304058
10. Javed MA, Wen L, Awais M, et al. TRO40303 ameliorates alcohol-induced pancreatitis through reduction of fatty acid ethyl ester-induced mitochondrial injury and necrotic cell death. Pancreas. 2018;47(1):18–24. doi:10.1097/mpa.0000000000000953
11. Vela S, Guerra A, Farrell G, et al. Pathophysiology and biomarker potential of fatty acid ethyl ester elevation during alcoholic pancreatitis. Gastroenterology. 2021;161(5):1513–1525. doi:10.1053/j.gastro.2021.07.029
12. Garcia D, Shaw RJ. AMPK: mechanisms of cellular energy sensing and restoration of metabolic balance. Mol Cell. 2017;66(6):789–800. doi:10.1016/j.molcel.2017.05.032
13. Wang Q, Liu S, Zhai A, et al. AMPK-mediated regulation of lipid metabolism by phosphorylation. Biol Pharm Bull. 2018;41(7):985–993. doi:10.1248/bpb.b17-00724
14. Meng Z, Liu Q, Sun F, et al. Hepatitis C virus nonstructural protein 5A perturbs lipid metabolism by modulating AMPK/SREBP-1c signaling. Lipids Health Dis. 2019;18(1):191. doi:10.1186/s12944019-1136-y
15. Dong J, Li M, Peng R, et al. ACACA reduces lipid accumulation through dual regulation of lipid metabolism and mitochondrial function via AMPK- PPARα- CPT1A axis. J Transl Med. 2024;22(1):196. doi:10.1186/s12967-024-04942-0
16. Steinberg GR, Hardie DG. New insights into activation and function of the AMPK. Nat Rev Mol Cell Biol. 2023;24(4):255–272. doi:10.1038/s41580-022-00547-x
17. Nesvaderani M, Eslick GD, Cox MR. Acute pancreatitis: update on management. Med J Aust. 2015;202(8):420–423. doi:10.5694/mja14.01333
18. Jin T, Li L, Zhang X, et al. Interpretation of integrated traditional Chinese and western medicine practice guidelines for diagnosis and treatment of acute pancreatitis. Chin. J. Bases Clin. Gen. Surg. 2024;31(2):205–211. doi:10.7507/1007-9424.202310043
19. Pancreas Study Group, Chinese Society of Gastroenterology, Chinese Medical Association. et al. Chinese guidelines for the management of acute pancreatitis (Shanghai, 2013). J Clin Hepatol. 2013;29(9):656–660. doi:10.3969/j.issn.1001-5256.2013.09.006
20. Wang G, Shang D, Zhang G, et al. Effects of QingYi decoction on inflammatory markers in patients with acute pancreatitis: a meta-analysis. Phytomedicine. 2022;95:153738. doi:10.1016/j.phymed.2021.153738
21. Wei TF, Zhao L, Huang P, et al. Qing-yi decoction in the treatment of acute pancreatitis: an integrated approach based on chemical profile, network pharmacology, molecular docking andexperimental evaluation. Front Pharmacol. 2021;12:590994. doi:10.3389/fphar.2021.590994
22. Ge P, Luo Y, Yang Q, et al. Ferroptosis in rat lung tissue during severe acute pancreatitis-associated acute lung injury: protection of qingyi decoction. Oxid Med Cell Longev. 2023;2023:5827613. doi:10.1155/2023/5827613
23. Liu G, Zhang J, Chen H, et al. Effects and mechanisms of alveolar type II epithelial cell apoptosis in severe pancreatitis-induced acute lung injury. Exp Ther Med. 2014;7(3):565–572. doi:10.3892/etm.2013.1453
24. Wang Z, Liu J, Li F, et al. Mechanisms of qingyi decoction in severe acute pancreatitis-associated acute lung injury via gut microbiota: targeting the short-chain fatty acids-mediated AMPK/NFκB/NLRP3 pathway. Microbiol Spectr. 2023;11(4):e0366422. doi:10.1128/spectrum.03664-22
25. Dai J, Liang K, Zhao S, et al. Chemoproteomics reveals baicalin activates hepatic CPT1 to ameliorate diet-induced obesity and hepatic steatosis. Proc Natl Acad Sci U S A. 2018;115(26):E5896–e5905. doi:10.1073/pnas.1801745115
26. Liu J, Shi Z, Yang Q, et al. Research progress on mechanism of emodin in treatment of metabolic associated fatty liver disease. Drugs Clinic. 2022;37(8):1898–1902. doi:10.7501/j.issn.16745515.2022.08.039
27. Liu M, Wang X, Yang Q, et al. Quercetin ameliorates BPA-induced hepatic lipid accumulation in rat offspring via AMPK-SREBP1-FAS/ACC pathway. Acta Nutrimenta Sinica. 2024;46(2):155–162. doi:10.13325/j.cnki.acta.nutr.sin.2024.02.004
28. Xing Y, Ren X, Li X, et al. Baicalein enhances the effect of acarbose on the improvement of nonalcoholic fatty liver disease associated with prediabetes via the inhibition of de novo lipogenesis. J Agric Food Chem. 2021;69(34):9822–9836. doi:10.1021/acs.jafc.1c04194
29. Liu RH, Wang XL. Experimental study of the alleviate ethanol intoxication effect of radix bupleuri and radix scutellariae in compatibility. Pharmacol Clin Chin Mater Med. 2004;20(4):3–4. doi:10.13412/j.cnki.zyyl.2004.04.002
30. Pandol SJ, Lugea A, Mareninova OA, et al. Investigating the pathobiology of alcoholic pancreatitis. Alcohol Clin Exp Res. 2011;35(5):830–837. doi:10.1111/j.1530-0277.2010.01408.x
31. Hanck C, Whitcomb DC. Alcoholic pancreatitis. Gastroenterol Clin North Am. 2004;33(4):751765. doi:10.1016/j.gtc.2004.07.002
32. Pfutzer RH, Schneider A. Treatment of alcoholic pancreatitis. Dig Dis. 2005;23(3–4):241–246. doi:10.1159/000090171
33. Li J, Chen J, Tang W. Consensus on diagnosis and treatment of acute pancreatitis with integrated traditional Chinese and western medicine (2017). Chin J Integr Trad West Med Dig. 2017;25(12):901909. doi:10.3969/j.issn.1671-038X.2017.12.03
34. Chen W, Yang X, Huang L, et al. Qing-Yi decoction in participants with severe acute pancreatitis: a randomized controlled trial. Chin Med. 2015;10:11. doi:10.1186/s13020-015-0039-8
35. Wang Y, Li T. Research progress on the development of myeloperoxidase inhibitors. J Nanjing Med Univ. 2023;43(12):1756–1763. doi:10.7655/NYDXBNS20231223
36. Avela HF, Sirén H. Advances in lipidomics. Clin Chim Acta. 2020;510:123–141. doi:10.1016/j.cca.2020.06.049
37. Frühbeck G, Méndez-Giménez L, Fernández-Formoso JA, et al. Regulation of adipocyte lipolysis. Nutr Res Rev. 2014;27(1):63–93. doi:10.1017/s095442241400002x
38. Yang A, Mottillo EP. Adipocyte lipolysis: from molecular mechanisms of regulation to disease and therapeutics. Biochem J. 2020;477(5):985–1008. doi:10.1042/bcj20190468
39. Eichmann TO, Lass A. DAG tales: the multiple faces of diacylglycerol--stereochemistry, metabolism, and signaling. Cell Mol Life Sci. 2015;72(20):3931–3952. doi:10.1007/s00018-015-19823
40. Escribá PV, Busquets X, Inokuchi J, et al. Membrane lipid therapy: modulation of the cell membrane composition and structure as a molecular base for drug discovery and new disease treatment. Prog Lipid Res. 2015;59:38–53. doi:10.1016/j.plipres.2015.04.003
41. Melero A, Jiménez-Rojo N. Cracking the membrane lipid code. Curr Opin Cell Biol. 2023;83:102203. doi:10.1016/j.ceb.2023.102203
42. Lettéron P, Brahimi-Bourouina N, Robin MA, et al. Glucocorticoids inhibit mitochondrial matrix acyl-CoA dehydrogenases and fatty acid beta-oxidation. Am J Physiol. 1997;272(5 Pt 1):1141–1150. doi:10.1152/ajpgi.1997.272.5.G1141
43. Xu L, Xia H, Ni D, et al. High-dose dexamethasone manipulates the tumor microenvironment and internal metabolic pathways in anti-tumor progression. Int J Mol Sci. 2020;21(5):1846. doi:10.3390/ijms21051846
44. Djouadi F, Bastin J. PPARalpha gene expression in the developing rat kidney: role of glucocorticoids. J Am Soc Nephrol. 2001;12(6):1197–1203. doi:10.1681/asn.V1261197
45. Hardie DG, Ross FA, Hawley SA. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol. 2012;13(4):251–262. doi:10.1038/nrm3311
46. Srinivasan MP, Bhopale KK, Caracheo AA, et al. Activation of AMP-activated protein kinase attenuates ethanol-induced ER/oxidative stress and lipid phenotype in human pancreatic acinar cells. Biochem Pharmacol. 2020;180:114174. doi:10.1016/j.bcp.2020.114174
47. Jeon SM. Regulation and function of AMPK in physiology and diseases. Exp Mol Med. 2016;48(7):e245. doi:10.1038/emm.2016.81
48. Wu S, Zou MHAMPK. Mitochondrial function, and cardiovascular disease. Int J Mol Sci. 2020;21(14). doi:10.3390/ijms21144987
49. Ma S, Wei T, Zhang B, et al. Integrated pharmacokinetic properties and tissue distribution of multiple active constituents in Qing-Yi Recipe: a comparison between granules and decoction. Phytomedicine. 2024;129:155645. doi:10.1016/j.phymed.2024.155645
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.