Back to Journals » Journal of Inflammation Research » Volume 18
Purinergic Receptors in Dendritic Cells
Authors Wang M, Zhao X, Hou S ![]() , Wu Z, Yin HY
, Wu Z, Yin HY ![]()
Received 23 June 2025
Accepted for publication 17 September 2025
Published 30 September 2025 Volume 2025:18 Pages 13423—13432
DOI https://doi.org/10.2147/JIR.S549102
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Miaomiao Wang,1 Xiaoxiao Zhao,1 Shuai Hou,1 Zhongbo Wu,2 Hai-yan Yin1
1School of Acupuncture and Tuina, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan, People’s Republic of China; 2Brain Disease Research Center, Tongren People’s Hospital, Tongren, Guizhou, People’s Republic of China
Correspondence: Hai-yan Yin, School of Acupuncture and Tuina, Chengdu University of Traditional Chinese Medicine, NO. 1166 Liutai Avenue, Wenjiang District, Chengdu, Sichuan, 611137, People’s Republic of China, Email [email protected]
Abstract: Dendritic cells (DCs) are regarded as highly effective antigen-presenting cells (APCs) and play a crucial role in immunomodulation. A growing body of research focuses on extracellular purines and their purinergic receptors in DCs. In this review, we provide an overview of the expression and function of purinergic receptors (P1 and P2) in DCs. To date, four P1 receptors (A1, A2A, A2B, A3), five P2X receptors (P2X1, P2X4, P2X5, P2X6, P2X7) and eight P2Y receptors (P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, P2Y14) have been reported to be expressed in DCs, with expression levels varying according to DC developmental stages (immature vs mature) and subsets. Functionally, P1 receptors are preferentially activated by adenosine(ADO) generated via ectonucleotidases, promoting the release of cytokines such as IL-10 and IL-23, and enhancing the activation, migration, and antigen presentation of DCs. In contrast, extracellular ATP-activated P2 receptors increase the secretion of pro-inflammatory cytokines including TNF-α, IL-1β, IL-18, and IL-12, while simultaneously inhibiting DC migration and antigen presentation efficiency. Therefore, only under the conditions that immune cells express the relevant receptors and ectonucleotidases dynamically regulate the ATP/ADO ratio can a mutually restrictive Yin-Yang relationship between P1 and P2 receptors be established, thereby safeguarding systemic immune homeostasis.
Keywords: dendritic cells, purinergic receptors, immunomodulation, yin and yang
Introduction
Dendritic cells (DCs) play a critical role in the immune system, serving as a bridge that connects innate and adaptive immunity.1 They can induce either effective immune response or promote immune tolerance under different pathological conditions. DC immunotherapy has emerged as a safe and promising therapeutic approach for cancer, as demonstrated in early clinical trials.2 Based on phenotypic and functional criteria, DCs in both humans and rodents can be divided into three major subsets: type 1 conventional dendritic cells (cDC1), type 2 conventional dendritic cells (cDC2), and plasmacytoid dendritic cells (pDCs).3 These subsets play distinct roles in immune responses. cDC1 is primarily responsible for antigen cross-presentation, whereby it presents intracellular antigens via MHC class I molecules to CD8+ T cells, thereby initiating anti-tumor immune responses, the subset exhibits strong pro-inflammatory properties and can enhance cellular immune reactions through the production of cytokines such as IL-12.4 In contrast, cDC2 primarily presents antigens to CD4+ T cells via MHC class II molecules, directing their differentiation into helper T cell subsets such as Th1, Th2, or Th17.5 Depending on microenvironmental signals, cDC2 may either promote inflammatory responses or contribute to the establishment of immune tolerance. On the other hand, pDCs are key producers of type I interferons (IFNs) and play crucial roles in antiviral immune responses as well as the regulation of antitumor immunity.6 Additionally, under inflammatory conditions in vivo, Ccr2+ monocytes originating from the bone marrow are recruited and differentiate into monocyte-derived dendritic cells (moDCs); this process can be triggered by exogenous stimuli such as the recall antigen (mBSA),7 LPS,8 or pathogen infections,9 thereby rapidly providing a population of DCs endowed with both phagocytic and antigen-presenting functions at the inflammatory site. To mediate innate and adaptive immunity, DCs undergo immature and mature stages.10 DCs in resting state are referred to as immature DCs (iDCs), while those in an activated state are called mature DCs (mDCs). Immature DCs are primarily found in peripheral tissues and feature short dendritic projections on their surface.10 They possess a high capacity for endocytosis but express low levels of co-stimulatory molecules and chemokine receptors.11 Under conditions of weak co-stimulatory signals or an adenosine-rich microenvironment, immature DCs continuously present self-antigens and secrete suppressive factors such as IL-10 and TGF-β, thereby exerting a tolerogenic function that drives naïve CD4+ T cells to differentiate into regulatory T cells and actively maintains peripheral immune tolerance.12,13 When an organism is attacked by pathogens, iDCs capture the antigens and subsequently begin to migrate and undergo a series of changes that lead to their transformation into mDCs.14 Mature DCs are primarily located in lymph nodes and exhibit long dendritic projections.15 They are characterized by the up regulation of major histocompatibility complex (MHC) class I and II, co-stimulatory molecules and inflammatory cytokines16–19, which contribute to initiating the adaptive immune response by binding to the T cell receptor (TCR) to active T cell.11 Notably, DCs detect and respond to pathogens through pattern recognition receptors (PRRs).20 The engagement of different pattern recognition receptors (PRRs) on dendritic cells (DCs) induces the secretion of distinct patterns of cytokines and inflammatory mediators. This not only promotes their own activation and maturation but also modulates the immune microenvironment through both cell-cell contacts and soluble factors. Consequently, PRR-stimulated DCs guide the differentiation of CD4+ T cells into various helper T cell subsets and regulate the activation and cytotoxic functions of CD8+ T cells, thereby orchestrating both the type and magnitude of the adaptive immune response.21–23 Overall, the diverse functionalities and adaptability of DCs underscore their essential role in orchestrating immune responses and maintaining homeostasis within the immune system.
An increasing number of studies are focusing on purinergic signaling and DCs. The key com ponents of the purinergic signaling include purine substances and their corresponding purinergic receptors.24 Purine substances, such as ATP, ADP and AMP, and ADO, serve as important cellular messengers within the purinergic pathway;25 Extracellular ATP (eATP) and ADO levels are precisely regulated by the ectonucleotidases CD39 (which hydrolyzes ATP/ADP to AMP) and CD73 (which hydrolyzes AMP to ADO), forming the key “CD39/CD73–ATP–ADO” metabolic axis that modulates immune responses through distinct purinergic receptors.26 Purinergic receptors can be categorized into two major classes, P1 (adenosine receptors) and P2 (adenine nucleotide receptors). Currently, four subtypes of P1 receptors are recognized: A1, A2A, A2B, and A3 receptors.27 P2 receptors are further divided into two classes: P2X and P2Y. P2X receptors are ligand-gated ion channels and consist of seven subtypes (P2X1-7), while P2Y receptors are G protein-coupled receptors with eight isoforms (P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, and P2Y14).28 In DCs, high concentrations of eATP engages P2 receptors, driving DC maturation and the secretion of pro-inflammatory cytokines.29 Conversely, the sequential hydrolysis of eATP by CD39 and CD73 generates adenosine, which activates P1 receptors and thereby suppresses immune responses, promotes a tolerogenic phenotype, and facilitates the differentiation of regulatory T cells.30 Purinergic signaling coordinates many aspects of cellular behavior,31 including proliferation, differentiation, migration, apoptosis, and various physiological processes. Furthermore, Purinergic signaling can mediate pathological dysregulation, leading to disease such as neurodegenerative disorders, immune diseases, inflammation, and cancer.31 Multiple lines of evidence suggest that purinergic receptors are potential targets for disease treatment, and specific purine agonists and antagonists show promising therapeutic potential.32
Accumulating research indicates that purinergic receptors may play a significant role in DCs concerning antigen presentation, cytokine release, and other functions. To gain a comprehensive understanding of this field, we made this review on purinergic receptors in DCs focusing on two main aspects: the expression of purinergic receptors on DCs, and the impact of purinergic receptors on various aspects of dendritic cell function.
Expression of Purinergic Receptors in DCs
It has been reported that DCs and purinergic signaling are widely present in the bodies of mammals. Notably both purine substances and their receptors are abundantly expressed in DCs.
Most data on purinergic receptor expression have been derived from studies on human DCs. Panther et al found that A1, A2A, and A3 receptors were expressed in iDCs derived from peripheral blood, whereas A2B receptors was not detected.33 After these iDCs were induced into mDCs by LPS, the mRNA expression of A1 and A3 decreased, while the expression of A2A obviously increased.33 Fossetta and his et.al also observed high expression of the A3 receptor in human iDCs, alongside high expression of the A2A receptor in LPS-activated mDCs.34 Schnurr35 showed that immature pDCs expressed A1 receptors not A2b or A3 receptors; Additionally, some donors exhibited low levels of A2A mRNA expression, while others had none detected. Neither A2b nor A3 receptors were found in mature pDCs.35 This pattern differed from immature moDCs, which expressed mRNA for A1, A2A and A3 receptors.33,34 In human moDCs, three P2X subtypes (P2X1, P2X4, and P2X7) and four P2Y subtypes (P2Y1, P2Y2, P2Y11), have been detected, with particularly high levels of the P2X7 receptor.36 Berchtold et al identified four P2X subtypes (P2X1, P2X4, P2X5, and P2X7) and six P2Y subtypes (P2Y1, P2Y2, P2Y4, P2Y6, P2Y10, P2Y11) in moDCs under both mature and immature states, as determined by RT-PCR.37 Shin38 found that immature pDCs express several P2Y receptor subtypes, namely P2Y4, P2Y6, P2Y12, P2Y13 and P2Y14, with the highest expression level observed for P2Y14. However, pDCs do not express receptors with high affinity for ATP, such as P2Y2 and P2Y11. Upon maturation, the mRNA expression of P2Y12, P2Y13, and P2Y14 decreases, while P2Y4 and P2Y6 remain unaffected.38
The expression of purinergic receptors in mouse DCs appears to differ from that in human DCs. In vitro cultured BMDCs of mice have detectable levels of A2A, A2B, and A3 receptors; however, A1 receptor have not consistently been found.39 Previous reports indicated that A1 receptors could be detected in resting cultured mouse BMDCs.40 In cultured mouse XS-106 (a Langerhans cell line), functional A2A and A3 receptors were inferred to be expressed in DCs through the use of selective agonists of A1, A2A, and A3.41 Subsequently, Professor Granstein42 and his team discovered that the XS-106 cell line also expresses P2 receptors. Through RT-PCR, they revealed the presence of mRNA for P2X1, P2X7, P2Y1, P2Y2, P2Y4 and P2Y11 in this cell line. Hayato et al found that P2rx4 and P2rx7 were expressed in mouse BMDCs, and co-expression of these two receptors was observed.43 Co-expression of P2rx4 and P2rx7 was also noted in rat liver DCs.44 Specifically, RT-PCR detection revealed that among the seven P2X receptor subtypes (P2X1 to P2X7), DCs in the liver of rat two days after birth express only the P2rx4, P2rx6 and P2rx7. The study also utilized immunofluorescence to observe changes in receptors during development, revealing a significant increase in the expression level of P2X6 receptor, while the expression level of P2X4 and P2X7 remained stable This indicates the specificity and dynamics of P2X receptor expression in DCs at different developmental stages.
To date, many purinergic receptors (both P1 and P2) have been reported to be widely expressed in DCs across different mammalian species, including humans, mice and rats (Table 1). However, there is limited research on the expression of purinergic receptors in rat DCs. Furthermore, the expression of purinergic receptors varies among DCs from different species and, notably, even within the same species. The expression of purinergic signaling differs in DCs from various sources (peripheral blood, bone marrow, skin, lymph, etc.) and in different state (mature vs immature). It also important to noted that most of the expression data for purinergic receptors have been obtained from in vitro cultured DCs. With continuous advancements in molecular detection technologies, the application of single-cell sequencing, two-photon microscopy, and other techniques may enable more accurate, real-time, and in vivo expression data of purinergic receptors in DCs.
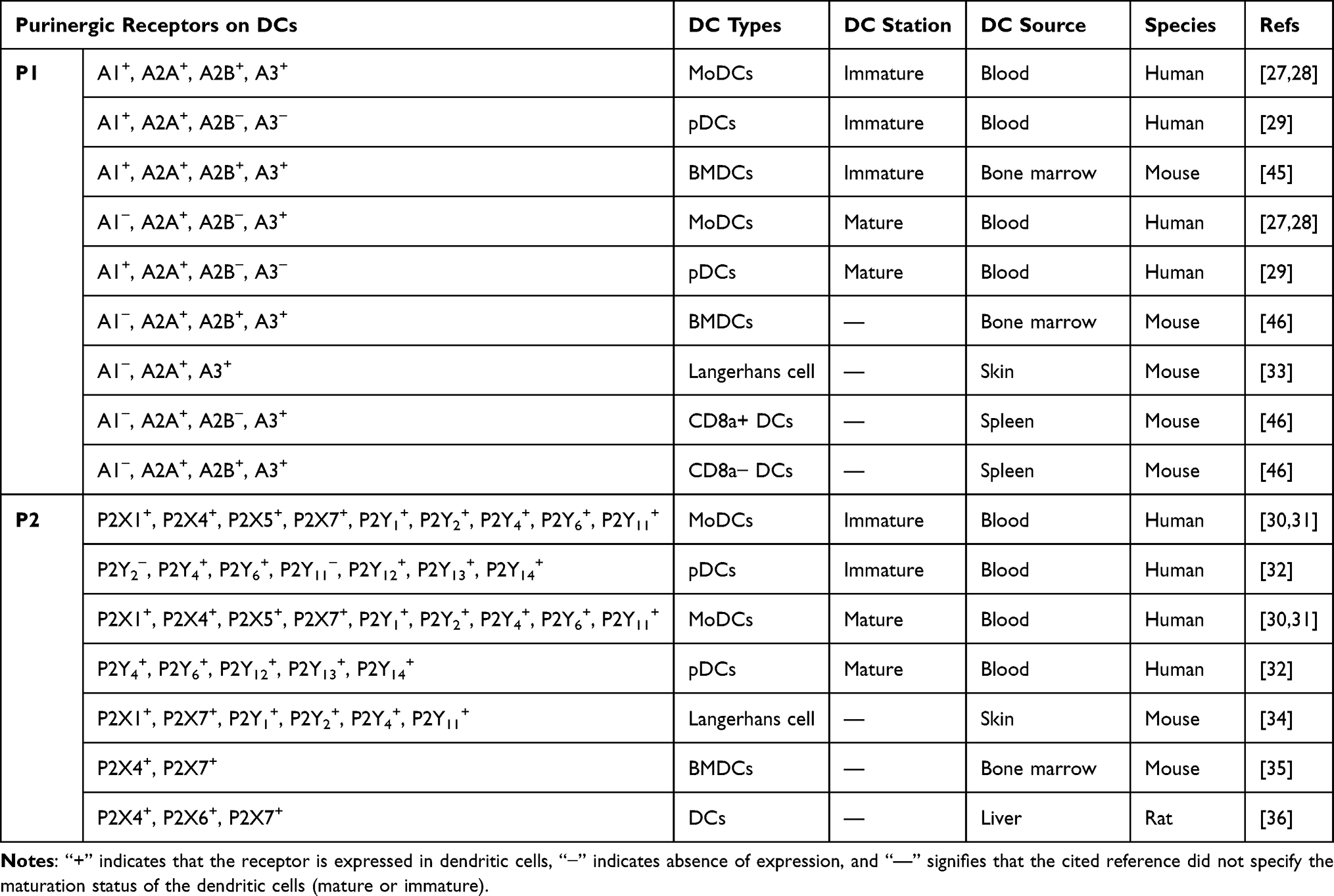 |
Table 1 Expression of Purinergic Receptors in Dendritic Cells |
Function of Purinergic Receptors in DCs
Function of P1 Receptors in DCs
P1 receptors, also known as adenosine receptors, can be selectively activated by adenosine and includes four isoforms, A1, A2A, A2B, and A3.47 It has been reported that P1 receptors play important roles in various type of DCs.
By analyzing human peripheral moDCs, researchers found that A1 and A3 receptors predom inantly induce chemotaxis, calcium transients, and actin polymerization in iDCs; In contrast, in mDCs, the A2A receptor enhances intracellular cAMP levels and inhibits IL-12 production.33 PDCs are specialized DCs primarily responsible for producing type I interferons (IFNs), such as IFN-α, which play a crucial role in antiviral immunity. In human pDCs, A1 receptors are involved in the migration of immature pDCs, while mature pDCs downregulate A1 receptors and upregulate A2A expression, leading to a reduction in the production of IL-6, IL-12, and IFN-α.35 MMP-9 plays a significant role in regulating the migration of DCs,48,49 and the A2B receptor indirectly affects the migratory capacity of DCs by modulating MMP-9 production. Specifically, under hypoxia conditions, A2B receptor expression on DCs is significantly increased, inhibiting MMP-9 production in moDCs through the cAMP/PKA signalling pathway, thereby slowing down DCs migration.50 In the hypoxic environment, ADO can inhibit IL-12 and TNF-α production by binding to the A2B receptor, thereby impairing Th1 cell differentiation. Moreover, the use of the A2B receptor antagonist MRS1754 can reverse this inhibitory effect.51 Additionally, hypoxic conditions can predispose DCs to exhibit a Th2-type stimulated phenotype.52 In studies on mouse DCs, ADO acts on the A1 receptor to inhibit vesicular cross-presentation of MHC-I molecules.40 Unlike human DCs, in mouse moDCs and pDCs, the A1 receptor mainly facilitates cell migration, whereas the migratory properties of mouse DCs are mediated by the A2A receptor.53 The A2A receptor also affects the antigen presenting capacity of DCs, which is enhanced when using a novel small-molecule A2AR antagonist AZD4635.54 The A2B receptor primarily influences the expression of co-stimulatory molecules and cytokine production on the surface of DCs. For example, it inhibits the expression of MHC-II, CD86 and CD40, thereby impairing DC activation and maturation, and reduces TNF-α, IL-12 production.55,56 In BMDCs, A2A and A2B receptors can target the PKA and Epac pathways to increase IL-10 secretion.57 Additionally, ADO impacts the ability of DCs to polarize T cells.58 Jeffrey59 and his et.al, through specific adenosine receptor knockout mice in their research, found that the A2B receptor promotes Th17 cell differentiation by stimulating DCs to secrete IL-6. Following this, researchers employed pharmacological approaches to explore the influence of the A2B receptor on the interaction between DCs and T cells. The study findings indicate that Th17 cells differentiation was significantly enhanced when BMDCs were treated with an A2B receptor agonist.60
Function of P2 Receptors in DCs
P2 receptors are expressed on the surface of various immune cells and are primarily activated by extracellular ATP and ADP. In physiological conditions, a large amount of ATP is stored intracellularly as an energetic source, while the eATP concentration is typically maintained in the the nanomolar range (1–100nM). However, when cells encounter pathogen-associated molecular patterns (PAMPs) released by pathogens, eATP levels may rise to the micromolar range, which is sufficient to activate immune responses and trigger various immune reactions, including cytokine release.61 In this context, eATP is regarded as a “danger signal”62 involved in the activation of the immune system. ATP exerts a series of effects on DCs by activating different P2 receptors, promoting antigen uptake, enhancing their chemotaxis, regulating cytokine release, and inducing cell death. In mouse BMDCs and spleen-derived DCs, researchers used FITC-dextran as a fluorescent antigen to explore the effects of P2 receptors on DCs. They found that activation of the P2ry12 receptor by ADP stimulated endocytosis and antigen presentation in iDCs.63 In another study,64 BMDCs from P2ry12-deficient mice showed a significant increase in IL-23 production compared to wild-type (WT) BMDCs. This increase in IL-23 promotes the differentiation of more native CD4+T cells into Th17 cells. Therefore, it can be concluded that genetic deletion of the P2ry12 affects the cytokine profile of BMDCs, disrupting the balance of T helper (Th) subsets and exacerbating experimental autoimmune encephalomyelitis (EAE). This imbalance ultimately exacerbates EAE. Conversely, DCs isolated from P2Y13-deficient mice did not exhibit significant functional differences compared to those from WT mice.63 Once DCs capture antigens, they increase the expression of cell surface co-stimulatory molecules and MHC molecules, allowing them to migrate to lymph nodes or sites of inflammation in preparation for an immune response. It was found that when immature moDCs were exposed to a low concentration (100 nM) of eATP, activation of the P2Y receptor triggered intracellular calcium mobilization, actin polymerization, and chemotaxis, collectively promoting the migration of DCs towards inflammatory areas.65 Similarly, extracellular UDP can also induce calcium transients, actin polymerization, and chemotaxis in iDCs by activating P2Y6 receptor, thereby facilitating their movement.66 Additionally, acute accumulation of ATP in the airways of mice after allergen provocation leads to DC recruitment and induces persistent airway inflammation.67 However, the specific P2 receptor mediating DC migration has not been clearly identified in some studies. Subsequent research demonstrated that ATP-induced DC migration requires the involvement of the P2Y2.68,69 Experiments using P2Y2 receptor-deficient mice exhibited a significant reduction in DC migration and a decrease in allergic airway inflammation.68 In addition to mediating the recruitment of DCs, P2Y2 can also induce the release of pro-inflammatory cytokines from DCs, exacerbating airway inflammation.70 Beyond directly regulating DCs migration, activation of P2 receptors on DCs can affect their mobility by modulating chemokine release. P2RY1 and P2RY11 inhibit the release of C-C motif chemokine ligand 2 (CCL2) and CCL3 from DCs,71 while P2Y11 and P2Y6 receptors promote the release of CCL20 from DCs.72 P2RY6 may also increase the release of C-X-C motif ligand 8 (CXCL8, also known as IL-8).66 These chemokines are capable of recruiting DCs and other immune cells to sites of inflammation.73 The P2X7 receptor is also involved in the regulation of DC migration. Specifically, ATP facilitates the rapid migration of DCs through Pannexin 1(Panx1) channels and P2rx7, aiding their homing to lymph nodes.74 In this process, functional Panx1 channels are essential for the in vivo migration of DCs, with P2X7 receptors acting as the main purinergic receptors involved in Panx1-dependent signaling. Furthermore, in DCs, the ATP/P2X7 axis may induce the expression and release of high mobility protein 1 (HMGB1), which promotes both DCs and macrophage migration and contributes to DCs maturation.75
During the migration of DCs, nucleotides finely regulate the immune response by mediating DC maturation and cytokine release. Specifically, ATP binding to the P2X receptors enhances the expression of surface markers CD83 and CD86 on DCs, thereby facilitating the maturation.76 In vitro experiments with human moDCs have shown that ATP increases intracellular cAMP levels through the activation of P2Y receptors, potentially including P2RY11,77 P2RX178 and P2RX3.78 This process promotes DC activation, as evidenced by the upregulation of CD83 and secretion of IL-12, further supporting DC maturation. Hypoxia decreases P2RY11 expression, leading to increased IL-12 secretion by DCs, and predisposing them to a pro-inflammatory phenotype, which may be involved in ischemia-reperfusion (I/R) injury following organ transplantation.79 However, it has also been demonstrated that ATP mediates immunosuppression by activating the P2RY11, promoting the upregulation of two targets (thrombospondin-1 [TSP-1] and indoleamine 2,3-dioxygenase [IDO]) in DCs.80 Schnurr’s study indicated that P2RY11 activation on DCs reduces their migration rate and prolongs their contact time with antigens, thereby enhancing the ability to recognize, bind and uptake antigens.81 In contrast, P2Y6 receptors inhibit antigen uptake by DCs as well as their differentiation and maturation, Specifically, it has been observed that BMDCs from P2Y6-deficient mice show significantly increased expression of CD40 and CD86 upon in vitro stimulation with LPS, along with higher levels of IL-12 and IL-23 detected in the supernatant fluid of P2ry6−/− BMDCs. This indicates a promotion ofTh1 and Th17 differentiation and exacerbation of EAE.82 Immature moDCs treated with UDP can activate P2RY14, leading to a significant increase in CD86 expression, which in turn promotes DC maturation and triggers an immune response.83 In another study, stimulation of the P2Y6 receptor was found to increase IL-8 secretion in DCs.84 Additionally, research has shown that P2RY4 and P2RY6, and P2RY14, inhibit the release of IFN-α in pDCs.38 Moreover, the P2X7 receptor is a potent inflammatory and immunostimulatory receptor.85 When expressed by DCs, P2X7 can induce DC maturation and pro-inflammatory cytokine release upon activation by ATP. Furthermore, previous studies have indicated that P2rx7 expression in murine is involved in antigen presentation processes.86 More recent research utilizing GM-CSF-induced BMDCs further demonstrates that the P2rx7 acts as a key molecule in dendritic cell-mediated cross-dressing, facilitating the transfer of preformed peptide–MHC complexes and thereby enhancing CD8+ T cell immune responses.87 Moreover, within the context of MVA viral vector vaccines, the P2rx7 indirectly yet enhances the cross-presentation capacity of BMDCs toward vaccine antigens.88 These findings collectively highlight the P2X7 receptor as a promising strategic target for optimizing T cell immunity in viral vector-based vaccines. The ATP/P2X7 axis typically functions to activate NLRP3 inflammatory vesicles in macrophages and monocytes. Upon activation of these inflammasomes, Caspase-1 is activated, which further processes the precursors interleukin (pro-IL)-1β and IL-18, ultimately promoting the release of these maturation cytokines.85 In vitro studies on BMDCs have found that treatment with P2X7R agonists results in increased secretion of IL-6 and IL-1β, as well as significantly elevated IL-17A production in CD4+ T cells, thereby inducing a more robust Th17 response.89 However, Yuan’s team later challenged the notion that the ATP-P2X7 axis induces TLR-mediated IL-1β production through the NLRP3 inflammasome in vivo. Using specific gene knockout mice, they discovered that intraperitoneal injection of LPS did not induce IL-1β production in the serum of NLRP3-deficient mice, while this was unaffected in P2rx7-deficient mice. Further in vitro experiments with BMDCs from these two types of mice yielded similar results. Ultimately, they concluded that IL-1β secretion by DCs is dependent on the NLRP3 inflammasome, but not on P2X7R.90 Additionally, the P2X7 receptor on DCs is also involved in their apoptotic process. Prolonged exposure of cells to a high concentration of ATP leads to the formation of a large pore by the P2X7 receptor in the plasma membrane, allowing the passage of molecules up to 900 Da. The long-term effect of this process is cell death.81
In summary, the expression and activation of P2 receptors in DCs are essential for regulating immune response, significantly affecting the maturation, migration, cytokine release, and apoptotic processes of these cells. P2X7R, in particular, plays a regulatory role in each of these processes. The multifaceted nature of P2 receptors in DCs also highlights their potential application in the treatment of immune-related diseases. To optimize therapies targeting purinergic receptors and enhance therapeutic efficacy, it is essential to understand the mechanisms of P2 receptors function in DCs.
Conclusions
In this discussion, we explore the multifaceted role of purinergic receptors in DCs, highlight ing the complexity and significance of this signaling system in immune regulation. To date, four P1 receptors (A1, A2A, A2B, A3), five P2X receptors (P2X1, P2X4, P2X5, P2X6, P2X7), and eight P2Y receptors (P2Y1, P2Y2, P2Y4, P2Y6, P2Y11, P2Y12, P2Y13, P2Y14) have been reported to be expressed in DCs. These receptors are not only involved in the maturation and migration of DCs but also influence cytokine secretion, as well as the proliferation and differentiation of T cells as shown in (Table 2). Purinergic receptors potentially regulate a wide range of DC functions through various signaling pathways (Figure 1). P1 receptors primarily promote the release of inflammatory cytokines (such as IL-10 and IL-23), as well as the activation, migration, and antigen presentation of DCs. In contrast, P2 receptors are mainly involved in increasing the secretion of pro-inflammatory cytokines (like TNF-α, IL-1β, IL-18, and IL-12) while inhibiting the activation, migration, antigen presentation of DCs.
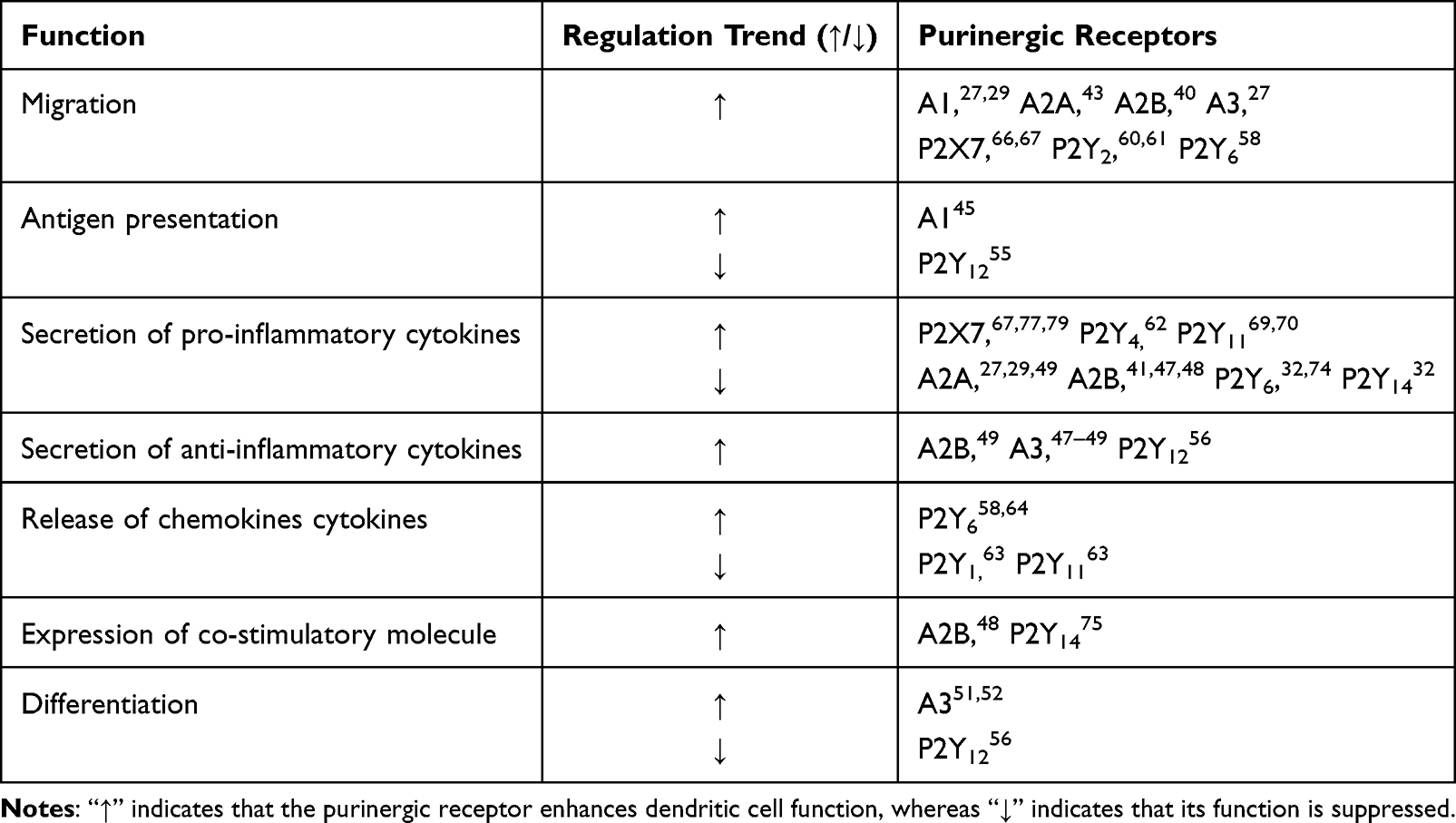 |
Table 2 Function of Purinergic Receptors in Dendritic Cells |
 |
Figure 1 Purinergic receptors in dendritic cell functions as opposing forces—Yin and Yang in immunomodulation. Abbreviations: P1, P1 purinoceptors; P2, P2 purinoceptors; eATP, Extracellular Adenosine Triphosphate; ADP, Adenosine Diphosphate; AMP, Adenosine Monophosphate; ADO, Adenosine. Notes: When tissues are damaged, levels of eATP increase significantly. The eATP is subsequently degraded into ADO by the exonucleases CD39 and CD73. The substantial increase of eATP and ADO activates their respective purinergic receptors (P1 and P2) In DCs. P1 receptors mainly promote the release of inflammatory factors (such as IL-10 and IL-23), as well as the activation, migration, and antigen presentation of DCs. In contrast, P2 receptors are mainly involved in enhancing the secretion of the pro-inflammatory factors (including TNF-α, IL-1β, IL-18, and IL-12) while inhibiting the activation, migration, antigen presentation of DCs. Therefore, P1 receptors and P2 receptors serve as complementary force in immunomodulation, playing an essential role in maintaining the body’s immune homeostasis. ↑ indicates enhanced function or increased release of inflammatory mediators, ↓ indicates reduced functionality. |
However, there are still some gaps in our understanding of the effects of purinergic receptors on DCs. Although it is known that these receptors influence DC function in various ways, the specific molecular mechanisms, particularly the detailed processes of downstream signaling pathways, require further elucidation. Additionally, DCs of different types and maturation stages may respond differently to purinergic signaling, and the mechanisms and biological significance of these differences need further exploration. Therefore, future studies should investigate the regulatory mechanisms of purinergic receptors and their functional changes in various disease states, aiming to identify new targets for the treatment of immune-related diseases.
Acknowledgments
We gratefully acknowledge the support provided by The Key Projects National Natural Science Foundation of China (NO. 81973969).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Lewis KL, Reizis B. Dendritic cells: arbiters of immunity and immunological tolerance. Cold Spring Harb Perspect Biol. 2012;4;8:a007401.
2. Chen J, Duan Y, Che J, Zhu J. Dysfunction of dendritic cells in tumor microenvironment and immunotherapy. Cancer Commun. 2024;44(9):1047–1070. doi:10.1002/cac2.12596
3. Guilliams M, Dutertre CA, Scott CL, et al. unsupervised high-dimensional analysis aligns dendritic cells across tissues and species. Immunity. 2016;45(3):669–684. doi:10.1016/j.immuni.2016.08.015
4. See P, Dutertre CA, Chen J, et al. Mapping the human DC lineage through the integration of high-dimensional techniques. Science. 2017;356(6342):eaag3009. doi:10.1126/science.aag3009
5. Plesca I, Müller L, Böttcher JP, Medyouf H, Wehner R, Schmitz M. Tumor-associated human dendritic cell subsets: phenotype, functional orientation, and clinical relevance. Eur J Immunol. 2022;52(11):1750–1758. doi:10.1002/eji.202149487
6. R B. Plasmacytoid dendritic cells: development, regulation, and function. Immunity. 2019. 50(1):37–50
7. Cook AD, Braine EL, Hamilton JA. Stimulus-dependent requirement for granulocyte-macrophage colony-stimulating factor in inflammation. J Immunol. 2004;173(7):4643–4651. doi:10.4049/jimmunol.173.7.4643
8. Cheong C, Matos I, Choi JH, et al. Microbial stimulation fully differentiates monocytes to DC-SIGN/CD209(+) dendritic cells for immune T cell areas. Cell. 2010;143(3):416–429. doi:10.1016/j.cell.2010.09.039
9. Greter M, Helft J, Chow A, et al. GM-CSF controls nonlymphoid tissue dendritic cell homeostasis but is dispensable for the differentiation of inflammatory dendritic cells. Immunity. 2012;36(6):1031–1046. doi:10.1016/j.immuni.2012.03.027
10. Bosteels V, Janssens S. Striking a balance: new perspectives on homeostatic dendritic cell maturation. Nat Rev Immunol. 2025;25:125–140. doi:10.1038/s41577-024-01079-5
11. ten Broeke T, Wubbolts R, Stoorvogel W. MHC class II antigen presentation by dendritic cells regulated through endosomal sorting. Cold Spring Harb Perspect Biol. 2013;5(12):a016873. doi:10.1101/cshperspect.a016873
12. Hawiger D, Inaba K, Dorsett Y, et al. Dendritic cells induce peripheral T cell unresponsiveness under steady state conditions in vivo. J Exp Med. 2001;194(6):769–779. doi:10.1084/jem.194.6.769
13. Audiger C, Rahman MJ, Yun TJ, Tarbell KV, Lesage S. The importance of dendritic cells in maintaining immune tolerance. J Immunol. 2017;198(6):2223–2231. doi:10.4049/jimmunol.1601629
14. Alvarez D, Vollmann EH, von Andrian UH. Mechanisms and consequences of dendritic cell migration. Immunity. 2008;29(3):325–342. doi:10.1016/j.immuni.2008.08.006
15. Randolph GJ, Angeli V, Swartz MA. Dendritic-cell trafficking to lymph nodes through lymphatic vessels. Nat Rev Immunol. 2005;5(8):617–628. doi:10.1038/nri1670
16. Steinman RM. Decisions about dendritic cells: past, present, and future. Annu Rev Immunol. 2012;30:1–22. doi:10.1146/annurev-immunol-100311-102839
17. Caux C, Massacrier C, Vanbervliet B, et al. Activation of human dendritic cells through CD40 cross-linking. J Exp Med. 1994;180(4):1263–1272. doi:10.1084/jem.180.4.1263
18. Tan JKH, O’Neill HC. Maturation requirements for dendritic cells in T cell stimulation leading to tolerance versus immunity. J Leukoc Biol. 2005;78(2):319–324. doi:10.1189/jlb.1104664
19. Jensen SS, Gad M. Differential induction of inflammatory cytokines by dendritic cells treated with novel TLR-agonist and cytokine based cocktails: targeting dendritic cells in autoimmunity. J Inflamm. 2010;7:37. doi:10.1186/1476-9255-7-37
20. Kawasaki T, Kawai T. Toll-like receptor signaling pathways. Front Immunol. 2014;5. doi:10.3389/fimmu.2014.00461
21. Yu M, Levine SJ. Toll-like receptor, RIG-I-like receptors and the NLRP3 inflammasome: key modulators of innate immune responses to double-stranded RNA viruses. Cytokine Growth Factor Rev. 2011;22(2):63–72. doi:10.1016/j.cytogfr.2011.02.001
22. Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563–604. doi:10.1146/annurev-immunol-020711-074950
23. Collin M, Bigley V. Human dendritic cell subsets: an update. Immunology. 2018;154(1):3–20. doi:10.1111/imm.12888
24. Bao X, Xie L. Targeting purinergic pathway to enhance radiotherapy-induced immunogenic cancer cell death. J Exp Clin Cancer Res. 2022;41(1):222. doi:10.1186/s13046-022-02430-1
25. Wang P, Jia J, Zhang D. Purinergic signalling in liver diseases: pathological functions and therapeutic opportunities. JHEP Rep. 2020;2(6):100165. doi:10.1016/j.jhepr.2020.100165
26. Antonioli L, Pacher P, Vizi ES, Haskó G. CD39 and CD73 in immunity and inflammation. Trends Mol Med. 2013;19(6):355–367.
27. Fredholm BB, IJzerman AP, Jacobson KA, Linden J, Müller CE. International union of basic and clinical pharmacology. LXXXI. nomenclature and classification of adenosine receptors--an update. Pharmacol Rev. 2011;63(1):1–34.
28. Ralevic V, Burnstock G. Receptors for purines and pyrimidines. Pharmacol Rev. 1998;50(3):413–492.
29. Silva-Vilches C, Ring S, Mahnke K. ATP and Its metabolite adenosine as regulators of dendritic cell activity. Front Immunol. 2018;9:417046
30. Silva-Vilches C, Ring S, Schrader J, et al. Production of extracellular adenosine by CD73+ dendritic cells is crucial for induction of tolerance in contact hypersensitivity reactions. J Invest Dermatol. 2019;139(3):541–551. doi:10.1016/j.jid.2018.10.016
31. Huang Z, Xie N, Illes P, et al. From purines to purinergic signalling: molecular functions and human diseases. Signal Transduct Target Ther. 2021;6(1):162. doi:10.1038/s41392-021-00553-z
32. Mahmood A, Iqbal J. Purinergic receptors modulators: an emerging pharmacological tool for disease management. Med Res Rev. 2022;42(4):1661–1703. doi:10.1002/med.21888
33. Panther E, Idzko M, Herouy Y, et al. Expression and function of adenosine receptors in human dendritic cells. FASEB J. 2001;15(11):1963–1970. doi:10.1096/fj.01-0169com
34. Fossetta J, Jackson J, Deno G, et al. Pharmacological analysis of calcium responses mediated by the human A3 adenosine receptor in monocyte-derived dendritic cells and recombinant cells. Mol Pharmacol. 2003;63(2):342–350. doi:10.1124/mol.63.2.342
35. Schnurr M, Toy T, Shin A, et al. Role of adenosine receptors in regulating chemotaxis and cytokine production of plasmacytoid dendritic cells. Blood. 2004;103(4):1391–1397. doi:10.1182/blood-2003-06-1959
36. Ferrari D, La Sala A, Chiozzi P, et al. The P2 purinergic receptors of human dendritic cells: identification and coupling to cytokine release. FASEB J. 2000;14(15):2466–2476. doi:10.1096/fj.00-0031com
37. Berchtold S, Ogilvie AL, Bogdan C, et al. Human monocyte derived dendritic cells express functional P2X and P2Y receptors as well as ecto-nucleotidases. FEBS Lett. 1999;458(3):424–428. doi:10.1016/S0014-5793(99)01197-7
38. Shin A, Toy T, Rothenfusser S, et al. P2Y receptor signaling regulates phenotype and IFN-alpha secretion of human plasmacytoid dendritic cells. Blood. 2008;111(6):3062–3069. doi:10.1182/blood-2007-02-071910
39. Addi AB, Lefort A, Hua X, et al. Modulation of murine dendritic cell function by adenine nucleotides and adenosine: involvement of the A 2B receptor. Eur J Immunol. 2008;38(6):1610–1620. doi:10.1002/eji.200737781
40. Chen L, Fredholm BB, Jondal M. Adenosine, through the A1 receptor, inhibits vesicular MHC class I cross-presentation by resting DC. Mol Immunol. 2008;45(8):2247–2254. doi:10.1016/j.molimm.2007.11.016
41. Dickenson JM, Reeder S, Rees B, Alexander S, Kendall D. Functional expression of adenosine A2A and A3 receptors in the mouse dendritic cell line XS-106. Eur. J. Pharmacol. 2003;474(1):43–51. doi:10.1016/S0014-2999(03)02041-7
42. Granstein RD, Ding W, Huang J, et al. Augmentation of cutaneous immune responses by ATPγS: purinergic agonists define a novel class of immunologic adjuvants. J Immunol. 2005;174(12):7725–7731. doi:10.4049/jimmunol.174.12.7725
43. Sakaki H, Fujiwaki T, Tsukimoto M, Kawano A, Harada H, Kojima S. P2X4 receptor regulates P2X7 receptor-dependent IL-1β and IL-18 release in mouse bone marrow-derived dendritic cells. Biochem. Biophys. Res. Commun. 2013;432(3):406–411. doi:10.1016/j.bbrc.2013.01.135
44. Xiang Z, Lv J, Jiang P, Chen C, Jiang B, Burnstock G. Expression of P2X receptors on immune cells in the rat liver during postnatal development. Histochem Cell Biol. 2006;126(4):453–463. doi:10.1007/s00418-006-0180-7
45. Di Virgilio F, Sarti AC, Grassi F. Modulation of innate and adaptive immunity by P2X ion channels. Curr Opinion Immunol. 2018;52:51–59. doi:10.1016/j.coi.2018.03.026
46. Zyma M, Pawliczak R. Characteristics and the role of purinergic receptors in pathophysiology with focus on immune response: purinergic receptors are widely expressed in immune cells and regulate high variety of their functions, taking part in pathogenesis of many diseases. Int Rev Immunol. 2020;39(3):97–117. doi:10.1080/08830185.2020.1723582
47. Fredholm BB, IJzerman AP, Jacobson KA, Klotz KN, Linden J. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev. 2001;53(4):527–552.
48. Ratzinger G, Stoitzner P, Ebner S, et al. Matrix metalloproteinases 9 and 2 are necessary for the migration of Langerhans cells and dermal dendritic cells from human and murine skin. J Immunol. 2002;168(9):4361–4371. doi:10.4049/jimmunol.168.9.4361
49. Osman M, Tortorella M, Londei M, Quaratino S. Expression of matrix metalloproteinases and tissue inhibitors of metalloproteinases define the migratory characteristics of human monocyte-derived dendritic cells. Immunology. 2002;105(1):73–82. doi:10.1046/j.0019-2805.2001.01349.x
50. Zhao P, Gang LX, Yang M, et al. Hypoxia suppresses the production of MMP-9 by human monocyte-derived dendritic cells and requires activation of adenosine receptor A2b via cAMP/PKA signaling pathway. Mol Immunol. 2008;45(8):2187–2195. doi:10.1016/j.molimm.2007.12.002
51. Yang M, Ma C, Liu S, et al. HIF-dependent induction of adenosine receptor A2b skews human dendritic cells to a Th2-stimulating phenotype under hypoxia. Immunol Cell Biol. 2010;88(2):165–171. doi:10.1038/icb.2009.77
52. Yang M, Ma C, Liu S, et al. Hypoxia skews dendritic cells to a T helper type 2-stimulating phenotype and promotes tumour cell migration by dendritic cell-derived osteopontin. Immunology. 2009;128(1 Suppl):e237–249. doi:10.1111/j.1365-2567.2008.02954.x
53. Ring S, Pushkarevskaya A, Schild H, et al. Regulatory T cell–derived adenosine induces dendritic cell migration through the epac-rap1 pathway. J Immunol. 2015;194(8):3735–3744. doi:10.4049/jimmunol.1401434
54. Borodovsky A, Barbon CM, Wang Y, et al. Small molecule AZD4635 inhibitor of A2AR signaling rescues immune cell function including CD103+ dendritic cells enhancing anti-tumor immunity. J Immunother Cancer. 2020;8(2):e000417. doi:10.1136/jitc-2019-000417
55. Figueiredo AB, Souza-Testasicca MC, Mineo TWP, Afonso LCC. Leishmania amazonensis-Induced cAMP triggered by adenosine A2B receptor is important to inhibit dendritic cell activation and evade immune response in infected mice. Front Immunol. 2017;8:849. doi:10.3389/fimmu.2017.00849
56. Wilson JM, Ross WG, Agbai ON, et al. The A2B adenosine receptor impairs the maturation and immunogenicity of dendritic cells. J Immunol. 2009;182(8):4616–4623. doi:10.4049/jimmunol.0801279
57. Kayhan M, Koyas A, Akdemir I, Savas AC, Cekic C. Adenosine receptor signaling targets both PKA and epac pathways to polarize dendritic cells to a suppressive phenotype. J Immunol. 2019;203(12):3247–3255. doi:10.4049/jimmunol.1900765
58. Panther E, Corinti S, Idzko M, et al. Adenosine affects expression of membrane molecules, cytokine and chemokine release, and the T-cell stimulatory capacity of human dendritic cells. Blood. 2003;101(10):3985–3990. doi:10.1182/blood-2002-07-2113
59. Wilson JM, Kurtz CC, Black SG, et al. The A2B adenosine receptor promotes th17 differentiation via stimulation of dendritic cell IL-6. J Immunol. 2011;186(12):6746–6752. doi:10.4049/jimmunol.1100117
60. Chen M, Liang D, Zuo A, Shao H, Kaplan HJ, Sun D. An A2B adenosine receptor agonist promotes th17 autoimmune responses in Experimental Autoimmune Uveitis (EAU) via dendritic cell activation. PLoS One. 2015;10(7):e0132348. doi:10.1371/journal.pone.0132348
61. Hamidzadeh K, Mosser DM. Purinergic signaling to terminate TLR responses in macrophages. Front Immunol. 2016;7:74. doi:10.3389/fimmu.2016.00074
62. Di Virgilio F. Purinergic mechanism in the immune system: a signal of danger for dendritic cells. Purinergic Signalling. 2005;1(3):205. doi:10.1007/s11302-005-6312-z
63. Ben Addi A, Cammarata D, Conley PB, Boeynaems JM, Robaye B. Role of the P2Y12 receptor in the modulation of murine dendritic cell function by ADP. J Immunol. 2010;185(10):5900–5906. doi:10.4049/jimmunol.0901799
64. Zhang J, Li Z, Hu X, et al. Knockout of P2Y12 aggravates experimental autoimmune encephalomyelitis in mice via increasing of IL-23 production and Th17 cell differentiation by dendritic cells. Brain Behav Immun. 2017;62:245–255. doi:10.1016/j.bbi.2016.12.001
65. Idzko M, Dichmann S, Ferrari D, et al. Nucleotides induce chemotaxis and actin polymerization in immature but not mature human dendritic cells via activation of pertussis toxin–sensitive P2y receptors. Blood. 2002;100(3):925–932. doi:10.1182/blood.V100.3.925
66. Idzko M, Panther E, Sorichter S, et al. Characterization of the biological activities of uridine diphosphate in human dendritic cells: influence on chemotaxis and CXCL8 release. J Cell Physiol. 2004;201(2):286–293. doi:10.1002/jcp.20070
67. Idzko M, Hammad H, Van Nimwegen M, et al. Extracellular ATP triggers and maintains asthmatic airway inflammation by activating dendritic cells. Nat Med. 2007;13(8):913–919. doi:10.1038/nm1617
68. Müller T, Robaye B, Vieira RP, et al. The purinergic receptor P2Y 2 receptor mediates chemotaxis of dendritic cells and eosinophils in allergic lung inflammation. Allergy. 2010;65(12):1545–1553. doi:10.1111/j.1398-9995.2010.02426.x
69. Klämbt V, Wohlfeil SA, Schwab L, et al. A novel function for P2Y2 in myeloid recipient-derived cells during graft-versus-host disease. J Immunol. 2015;195(12):5795–5804. doi:10.4049/jimmunol.1501357
70. Schneble D, El-Gazzar A, Kargarpour Z, et al. Cell-type-specific role of P2Y2 receptor in HDM-driven model of allergic airway inflammation. Front Immunol. 2023;14:1209097. doi:10.3389/fimmu.2023.1209097
71. Horckmans M, Marcet B, Marteau F, et al. Extracellular adenine nucleotides inhibit the release of major monocyte recruiters by human monocyte-derived dendritic cells. FEBS Lett. 2006;580(3):747–754. doi:10.1016/j.febslet.2005.12.091
72. Marcet B, Horckmans M, Libert F, Hassid S, Boeynaems JM, Communi D. Extracellular nucleotides regulate CCL20 release from human primary airway epithelial cells, monocytes and monocyte-derived dendritic cells. J Cell Physiol. 2007;211(3):716–727. doi:10.1002/jcp.20979
73. Sallusto F, Mackay CR, Lanzavecchia A. The role of chemokine receptors in primary, effector, and memory immune responses. Annu Rev Immunol. 2000;18:593–620. doi:10.1146/annurev.immunol.18.1.593
74. Sáez PJ, Vargas P, Shoji KF, Harcha PA, Lennon-Duménil AM, Sáez JC. ATP promotes the fast migration of dendritic cells through the activity of pannexin 1 channels and P2X 7 receptors. Sci Signal. 2017;10(506):eaah7107. doi:10.1126/scisignal.aah7107
75. Li R, Wang J, Li R, et al. ATP/P2X7-NLRP3 axis of dendritic cells participates in the regulation of airway inflammation and hyper-responsiveness in asthma by mediating HMGB1 expression and secretion. Exp. Cell. Res. 2018;366(1):1–15. doi:10.1016/j.yexcr.2018.03.002
76. La Sala A, Ferrari D, Corinti S, Cavani A, Di Virgilio F, Girolomoni G. Extracellular ATP induces a distorted maturation of dendritic cells and inhibits their capacity to initiate Th1 responses. J Immunol. 2001;166(3):1611–1617. doi:10.4049/jimmunol.166.3.1611
77. Wilkin F, Duhant X, Bruyns C, Suarez-Huerta N, Boeynaems JM, Robaye B. The P2Y11 Receptor Mediates the ATP-Induced Maturation of Human Monocyte-Derived Dendritic Cells. J Immunol. 2001;166(12):7172–7177. doi:10.4049/jimmunol.166.12.7172
78. Schnurr M, Then F, Galambos P, et al. Extracellular ATP and TNF-alpha synergize in the activation and maturation of human dendritic cells. J Immunol. 2000;165(8):4704–4709. doi:10.4049/jimmunol.165.8.4704
79. Chadet S, Ivanes F, Benoist L, et al. Hypoxia/Reoxygenation Inhibits P2Y11 receptor expression and its Immunosuppressive activity in human dendritic cells. J Immunol. 2015;195(2):651–660. doi:10.4049/jimmunol.1500197
80. Marteau F, Gonzalez NS, Communi D, Goldman M, Boeynaems JM, Communi D. Thrombospondin-1 and indoleamine 2,3-dioxygenase are major targets of extracellular ATP in human dendritic cells. Blood. 2005;106(12):3860–3866. doi:10.1182/blood-2005-05-1843
81. Schnurr M, Toy T, Stoitzner P, et al. ATP gradients inhibit the migratory capacity of specific human dendritic cell types: implications for P2Y11 receptor signaling. Blood. 2003;102(2):613–620. doi:10.1182/blood-2002-12-3745
82. Li Z, He C, Zhang J, et al. P2Y6 deficiency enhances dendritic cell-mediated Th1/Th17 differentiation and aggravates experimental autoimmune encephalomyelitis. J Immunol. 2020;205(2):387–397. doi:10.4049/jimmunol.1900916
83. Skelton L, Cooper M, Murphy M, Platt A. Human immature monocyte-derived dendritic cells express the G protein-coupled receptor GPR105 (KIAA0001, P2Y14) and increase intracellular calcium in response to its agonist, uridine diphosphoglucose. J Immunol. 2003;171(4):1941–1949. doi:10.4049/jimmunol.171.4.1941
84. Warny M, Aboudola S, Robson SC, et al. P2Y(6) nucleotide receptor mediates monocyte interleukin-8 production in response to UDP or lipopolysaccharide. J Biol Chem. 2001;276(28):26051–26056. doi:10.1074/jbc.M102568200
85. Di Virgilio F, Dal Ben D, Sarti AC, Giuliani AL, Falzoni S. The P2X7 Receptor in Infection and Inflammation. Immunity. 2017;47(1):15–31. doi:10.1016/j.immuni.2017.06.020
86. Mutini C, Falzoni S, Ferrari D, et al. Mouse dendritic cells express the P2X7 purinergic receptor: characterization and possible participation in antigen presentation. J Immunol. 1999;163(4):1958–1965. doi:10.4049/jimmunol.163.4.1958
87. Barrera-Avalos C, Briceño P, Valdés D, et al. P2X7 receptor is essential for cross-dressing of bone marrow-derived dendritic cells. iScience. 2021;24(12):103520. doi:10.1016/j.isci.2021.103520
88. Longo Y, Mascaraque SM, Andreacchio G, et al. The purinergic receptor P2X7 as a modulator of viral vector-mediated antigen cross-presentation. Front Immunol.
89. Yang Y, Story ME, Hao X, Sumpter TL, Mathers AR. P2X7 receptor expression and signaling on dendritic cells and CD4+ T cells is not required but can enhance Th17 Differentiation. Front Cell Dev Biol. 2022;10:687659. doi:10.3389/fcell.2022.687659
90. He Y, Franchi L, Núñez G. Toll-like Receptor Agonists Stimulate Nlrp3-dependent IL-1β Production Independently of the purinergic P2X7 Receptor in Dendritic Cells and in vivo. J Immunol. 2014;190(1):334–9
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.