Back to Journals » Journal of Pain Research » Volume 19
Pulsed Radiofrequency and Epigenetic Modulation of Pain Pathways: A Systematic Review Based on Preclinical Evidence
Authors Occhigrossi F, Mosca J, Micheli F, Gazzeri R, Robinson CL ![]() , Lo Bianco G
, Lo Bianco G ![]()
Received 17 November 2025
Accepted for publication 9 February 2026
Published 17 February 2026 Volume 2026:19 582117
DOI https://doi.org/10.2147/JPR.S582117
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor King Hei Stanley Lam
Felice Occhigrossi,1 Jacopo Mosca,2 Fabrizio Micheli,3 Roberto Gazzeri,1 Christopher L Robinson,4 Giuliano Lo Bianco5
1Pain Therapy Unit, San Giovanni-Addolorata Hospital, Rome, Italy; 2Pain Therapy Unit, Policlinico Umberto I Hospital, Rome, Italy; 3Unit of Interventional and Surgical Pain Management, Guglielmo da Saliceto Hospital, Piacenza, Italy; 4Division of Pain Medicine, Department of Anesthesiology and Critical Care Medicine, The Johns Hopkins University School of Medicine, Baltimore, Maryland, USA; 5Anesthesiology and Pain Department, Foundation G. Giglio Cefalù, Palermo, Italy
Correspondence: Felice Occhigrossi, Pain Therapy Unit, San Giovanni-Addolorata Hospital, Rome, Italy, Email [email protected]
Background: Pulsed radiofrequency (PRF) is used for refractory neuropathic pain, yet its mechanisms remain incompletely defined. The dorsal root ganglion (DRG) and peripheral nerves (eg, sciatic nerve) are common targets and key nodes in nociceptive transmission and neuro-immune crosstalk. Compared with implantable electrical neuromodulation (SCS/PNS), PRF provides non-invasive, percutaneous neuromodulation. Epigenetic regulation after PRF is plausible because chronic pain causes transcriptional modifications and electrical neuromodulation may influence chromatin-based gene control.
Objective: To summarize preclinical evidence on epigenetic and molecular changes after PRF applied to the DRG or peripheral nerves.
Methods: PubMed, Embase, and Scopus were searched (PRISMA-S); synthesis followed SWiM. Studies assessed direct epigenetic modifications and/or gene/protein expression changes after PRF at DRG or peripheral nerve targets. Risk of bias was evaluated using the JBI checklist for animal studies.
Results: Seventeen rodent studies were included. One study reported a direct epigenetic modification, showing that PRF reversed complete Freund’s adjuvant (CFA)-induced histone H3/H4 hypoacetylation at the Kcc2 locus, restoring KCC2 expression. Other studies linked PRF to attenuation of microglial/MAPK signaling, modulation of neurotrophic mediators, downregulation of pro-inflammatory cytokines, and normalization of sensory neuron excitability markers across DRG, sciatic nerve, and spinal dorsal horn tissue. Target-specific trends emerged: DRG PRF more consistently attenuated spinal microglial/MAPK signaling, whereas sciatic PRF emphasized normalization of DRG channels/neuropeptides. No human studies and no circulating epigenetic biomarkers were identified.
Conclusion: Preclinical data support a non-destructive PRF mechanism involving dampening of neuro-immune signaling and restoration of inhibitory tone. Direct epigenetic evidence remains limited (single study) and absent in humans. Priorities include standardized PRF parameter reporting, time-course and multi-omic profiling beyond Kcc2, and prospective clinical studies incorporating molecular endpoints and circulating epigenetic readouts.
Keywords: pulsed radiofrequency, dorsal root ganglion, neuropathic pain, epigenetics, histone acetylation, microglia, MAPK, KCC2, TRPV1, GDNF
Introduction
The clinical use of radiofrequency (RF) procedures in pain medicine has expanded, particularly for refractory neuropathic and inflammatory conditions.1–3 Clinical applications of pulsed and conventional RF have also been reported in human pain conditions, including knee osteoarthritis and lumbar facet joint pain.4–6 PRF, delivered at sub-neurodestructive temperatures, is applied to the dorsal root ganglion (DRG) and peripheral nerves to modulate nociceptive signaling without thermal lesioning. Despite widespread adoption, the mechanisms of action remain only partially understood.
Crucially, the current body of mechanistic evidence is exclusively preclinical in murine models; to our knowledge, no human studies have directly evaluated molecular or epigenetic effects of PRF at DRG or peripheral nerve targets. Across rodent experiments, PRF has been associated with changes in gene/protein expression within pain and neuroimmune pathways (eg, IRF8, BDNF, P2X3, TRPV1/CGRP, p‑ERK/p38/JNK, substance P, GABAB/KCC2) in DRG, nerve, and spinal cord tissues.7–13
Only one study has reported directly measured epigenetic modifications following PRF (histone acetylation at the Kcc2 locus),14 while the remaining literature infers mechanistic plausibility from downstream transcriptional or protein changes. Notably, no studies have measured circulating epigenetic biomarkers (eg, plasma/serum miRNAs) before and after PRF.
This systematic review synthesizes the available evidence on PRF at the DRG and peripheral nerves, focusing on directly measured epigenetic changes and molecular alterations (mRNA/proteins) within pain and neuroinflammation pathways. Our aim is to clarify the biological plausibility of PRF while highlighting key gaps—the absence of human data, the scarcity of direct epigenetic measurements, and the complete lack of circulating epigenetic readouts—to guide future translational research.
Methods
Study Design and Research Question
We conducted a systematic review in accordance with the PRISMA 2020 statement. The review protocol was not prospectively registered (eg, PROSPERO). The search strategy was documented following PRISMA-S,15 and the qualitative synthesis was reported according to SWiM16 (Figure 1). Our objective was to assess how RF—with a focus on PRF—applied to the DRG/nerve roots or to peripheral nerves (eg, sciatic or medial branches) is associated with epigenetic alterations and/or changes in gene/protein expression relevant to antinociceptive mechanisms. Risk of bias was assessed using the Joanna Briggs Institute (JBI) Critical Appraisal Checklist for experimental animal studies, supplemented by domains from the SYRCLE and ARRIVE guidelines.
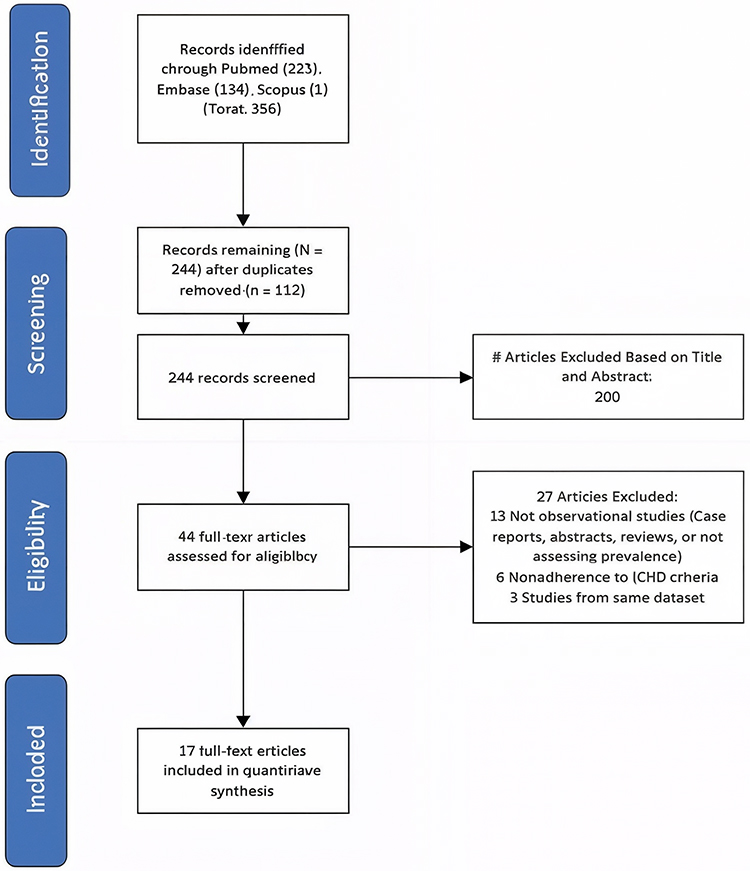 |
Figure 1 PRISMA-S 2020 flow diagram summarizing the study selection process. |
Information Sources and Search Strategy
We searched PubMed, Embase, and Scopus. Search strings combined RF terms (“pulsed radiofrequency”/“radiofrequency”) with epigenetic terms (epigenetic*histone*methyl*miRNA/lncRNA) and molecular terms (gene/protein expression), paired with pain/neuropath*/inflamm* keywords. The full PubMed search strategy is reported in Table 1, together with the complete search strategies for Embase and Scopus.
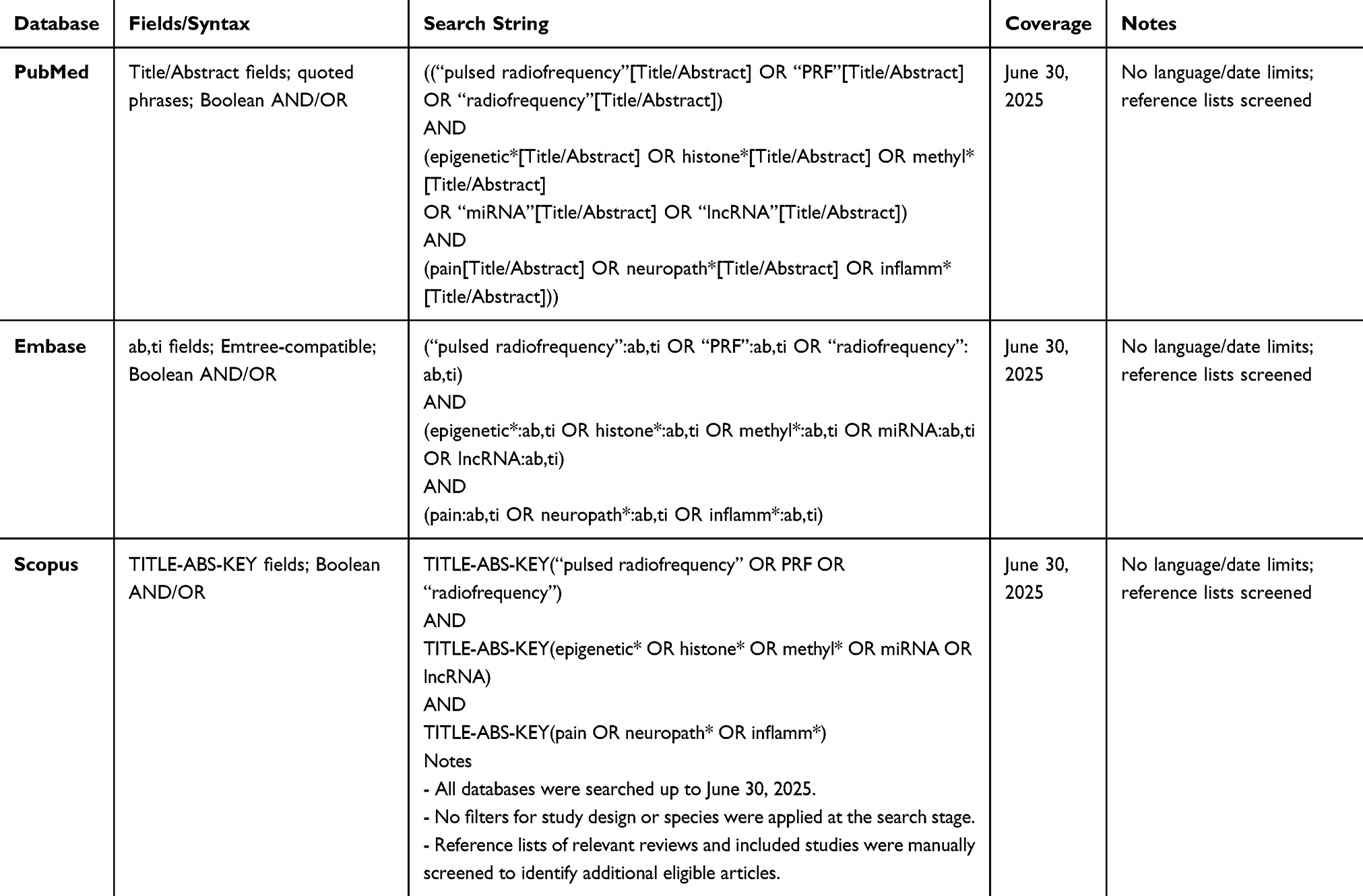 |
Table 1 The Full PubMed, Embase and Scopus Search Strategies |
Eligibility Criteria
We considered eligible preclinical and clinical studies investigating the effects of PRF delivered to the DRG/nerve roots or peripheral nerves on biological outcomes relevant to nociception. The primary outcome was the presence of directly measured epigenetic modifications, including DNA methylation, histone modifications (eg, via ChIP), and non-coding RNAs (miRNA/lncRNA), as well as potential circulating epigenetic biomarkers. In the absence of direct epigenetic measures, we included studies reporting changes in gene or protein expression within key pain- and neuroinflammation-related pathways (eg, IRF8, BDNF, KCC2, P2X3, TRPV1/CGRP, p-ERK/p38/JNK, substance P, GABAB). Reasons for exclusion were: absence of PRF procedure; absence of epigenetic/molecular measures; insufficient data.
Study Selection
Screening proceeded in two stages: first title/abstract review for preliminary eligibility; and then full-text assessment to confirm inclusion and allocate each study to the target (DRG/roots vs peripheral nerve) and outcome type (direct epigenetics vs expression).
Quality Assessment and Risk of Bias
Given the heterogeneity of study designs, we performed a qualitative appraisal across key domains: randomization and blinding, completeness of PRF parameter reporting, sample size, appropriateness of molecular methods (eg, antibody/primer specificity and normalization), use of controls, and reporting transparency. In addition, we applied the JBI Critical Appraisal Checklist for experimental animal studies, with each study evaluated across 10 domains (yes/no/unclear) and results summarized in Table 2. Most studies showed unclear risk in randomization and blinding, frequent incomplete reporting of PRF parameters, but consistent strengths in outcome validity and statistical analysis. As conceptual references, we also considered the SYRCLE and ARRIVE guidelines17–19 for animal studies and general risk-of-bias principles for clinical research.
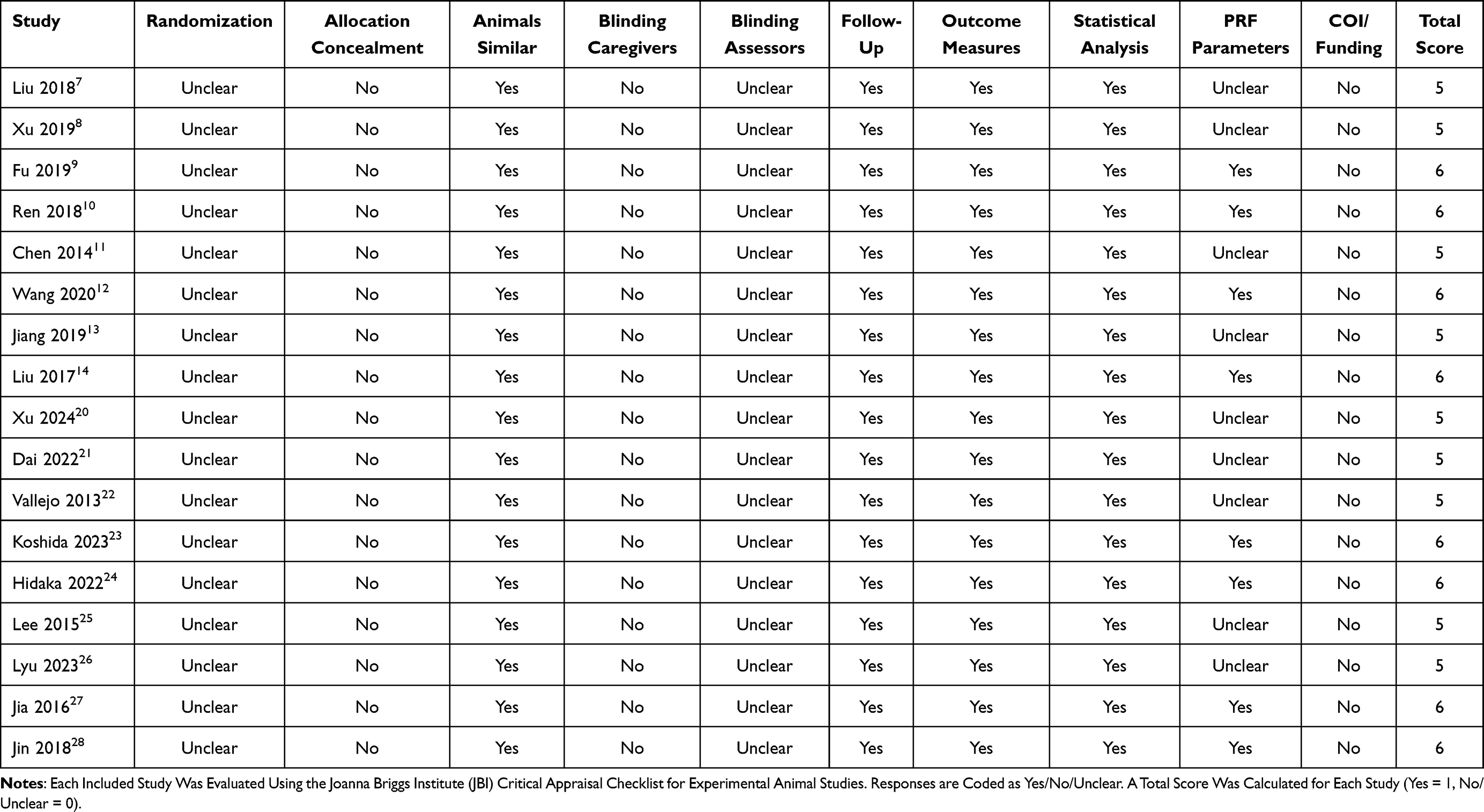 |
Table 2 JBI Critical Appraisal Checklist for Animal Studies |
The study selection process is illustrated in Figure 1, following the PRISMA 2020 guidelines. A total of 17 preclinical rodent studies met the eligibility criteria and were included in the final synthesis. The diagram shows the number of records identified through database searching (PubMed, Embase, and Scopus), the number of duplicates removed, records screened, full-text articles assessed for eligibility, and the final 17 preclinical studies included in the systematic review. Reasons for exclusion at the full-text stage are detailed in the diagram.
Study Selection and Characteristics
A total of 17 preclinical rodent studies met the eligibility criteria and were included in the final synthesis. The study selection process is illustrated in Figure 1 (PRISMA 2020 flow diagram). Screening proceeded in two stages: first title/abstract review for preliminary eligibility, followed by full-text assessment to confirm inclusion and allocation of each study to the target (dorsal root ganglion/nerve roots vs peripheral nerves) and outcome type (direct epigenetic measures vs molecular expression changes). Reasons for exclusion were recorded and are detailed in the PRISMA diagram.
The included studies involved PRF delivered to either the DRG/nerve roots or the sciatic/peripheral nerves. Molecular readouts comprised gene and protein expression in DRG, sciatic nerve, and spinal dorsal horn; only one study reported direct epigenetic modifications at the Kcc2 locus. Behavioral outcomes (mechanical and thermal thresholds) accompanied most experiments, while reporting of PRF parameters (eg, voltage, pulse width/interval, exposure time) was variably complete. Risk-of-bias assessment highlighted recurrent limitations, including incomplete reporting of PRF settings, lack of blinding and randomization details, and small sample sizes. Full JBI checklist results for each study are reported in Table 2.
Results
DRG/Nerve-Root Targets
Across DRG-targeted studies, convergent changes appeared in neuro-immune and synaptic pathways. In the spinal cord, PRF was associated with reductions in IRF8, microglial markers (Iba1), and phosphorylated MAPKs (p-p38 and p-ERK), changes that paralleled analgesic effects.7,8,20 BDNF modulation was also observed in line with pain relief.8,29 At the DRG level, Nav1.7 was consistently down-regulated, with high-voltage PRF showing stronger effects than standard settings.21 High-voltage PRF also increased GRK2 and reduced p-p38, consistent with dampened neuroinflammation.20 In mixed-target models, PRF reduced circulating IL-1β and TNF-α as well as spinal β-catenin, alongside behavioral improvement.13 Early transcriptomic profiling further demonstrated PRF-induced gene expression changes spanning nerve–DRG–spinal cord.22
Sciatic Nerve Target
When PRF was applied to the sciatic nerve, molecular alterations were observed across DRG, nerve, and spinal tissue. At the DRG, PRF normalized TRPV1/CGRP expression and modulated BDNF in resiniferatoxin-induced neuropathy.23 Early post-injury PRF produced greater suppression of p-ERK and Nav1.7 than delayed treatment.24 Expression of the purinoceptor P2X3 decreased in both DRG and dorsal horn.9 Neuropeptide and cytokine changes included reduced CGRP in DRG,10 decreased substance P in the dorsal horn,12 and down-regulation of TNF-α in sciatic nerve and spinal cord.25 Sciatic PRF also influenced the pain–depression interplay, reducing spinal IRF8 while increasing prefrontal BDNF, paralleling improvements in pain-induced depressive-like behavior.26 In inflammatory pain models, PRF attenuated CFA-induced hyperalgesia via spinal JNK modulation.11 Finally, studies targeting the ligation site demonstrated increased GDNF expression (both mRNA and protein), with associated behavioral improvements and ultrastructural recovery.27,28
Direct Epigenetic Readouts
Only one study to date has reported direct epigenetic measurements after PRF exposure. Liu and coworkers14 demonstrated that PRF attenuated CFA-induced histone H3/H4 hypoacetylation at the Kcc2 locus, thereby restoring KCC2 expression and inhibitory tone within spinal circuits. This finding provides the first mechanistic link between PRF delivery and epigenetic regulation of neuronal inhibition. Beyond this study, no experimental work has investigated DNA methylation, histone modifications at other loci, or non-coding RNAs in a hypothesis-driven, target-specific manner following PRF. Furthermore, no studies have evaluated circulating epigenetic biomarkers such as plasma or serum miRNAs, DNA methylation signatures, or histone marks, underscoring a critical gap for future translational research.
A schematic synthesis of these molecular and epigenetic changes induced by PRF across DRG, sciatic nerve, and spinal cord is presented in Figure 2.
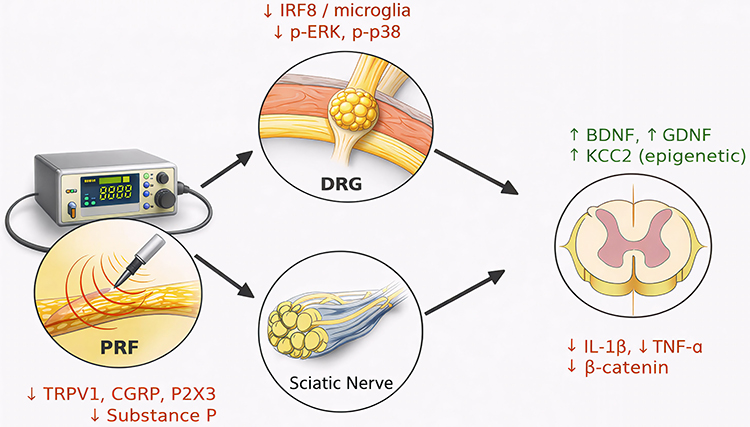 |
Figure 2 Mechanistic effects of pulsed radiofrequency (PRF) across DRG, sciatic nerve, and spinal cord. The red arrows indicate a decrease in concentration. The green arrows indicate an increase in concentration. |
The diagram illustrates convergent preclinical findings: suppression of microglial/IRF8 and MAPK signaling at the DRG; normalization of TRPV1, CGRP, P2X3, and Substance P at the sciatic nerve; and spinal changes including increased BDNF, GDNF, and KCC2 (epigenetic regulation), together with reductions in pro-inflammatory cytokines (IL-1β, TNF-α) and β-catenin. Arrows indicate the direction of proposed effects.
Discussion
This systematic review indicates that PRF applied to the DRG or peripheral nerves in rodent models is associated with gene expression changes across neuro-immune and synaptic pathways in the DRG, sciatic nerve, and spinal dorsal horn. Taken together, the data support a biologically plausible, non-neurodestructive mechanism of action in which PRF dampens microglial signaling and excitatory transmission while favoring inhibitory tone. At the same time, the evidence base remains preclinical, heterogeneous in reporting of PRF parameters, and—with one exception—limited to downstream transcriptional/protein readouts rather than direct epigenetic measures.
Multiple DRG-targeted studies converge on attenuation of microglia/IRF8 pathways and MAPK activation in the dorsal horn, paralleling behavioral analgesia. Reduced IRF8, Iba1, and p-p38/p-ERK suggest that PRF counters glial priming and inflammatory amplification within spinal circuits. In a mixed-target model, decreases in circulating IL-1β/TNF-α accompanied reductions in spinal β-catenin, linking peripheral immune tone to central signaling. These findings are mechanistically coherent with earlier tract-wide transcriptomic shifts after PRF across nerve → DRG → spinal cord (Table 3).
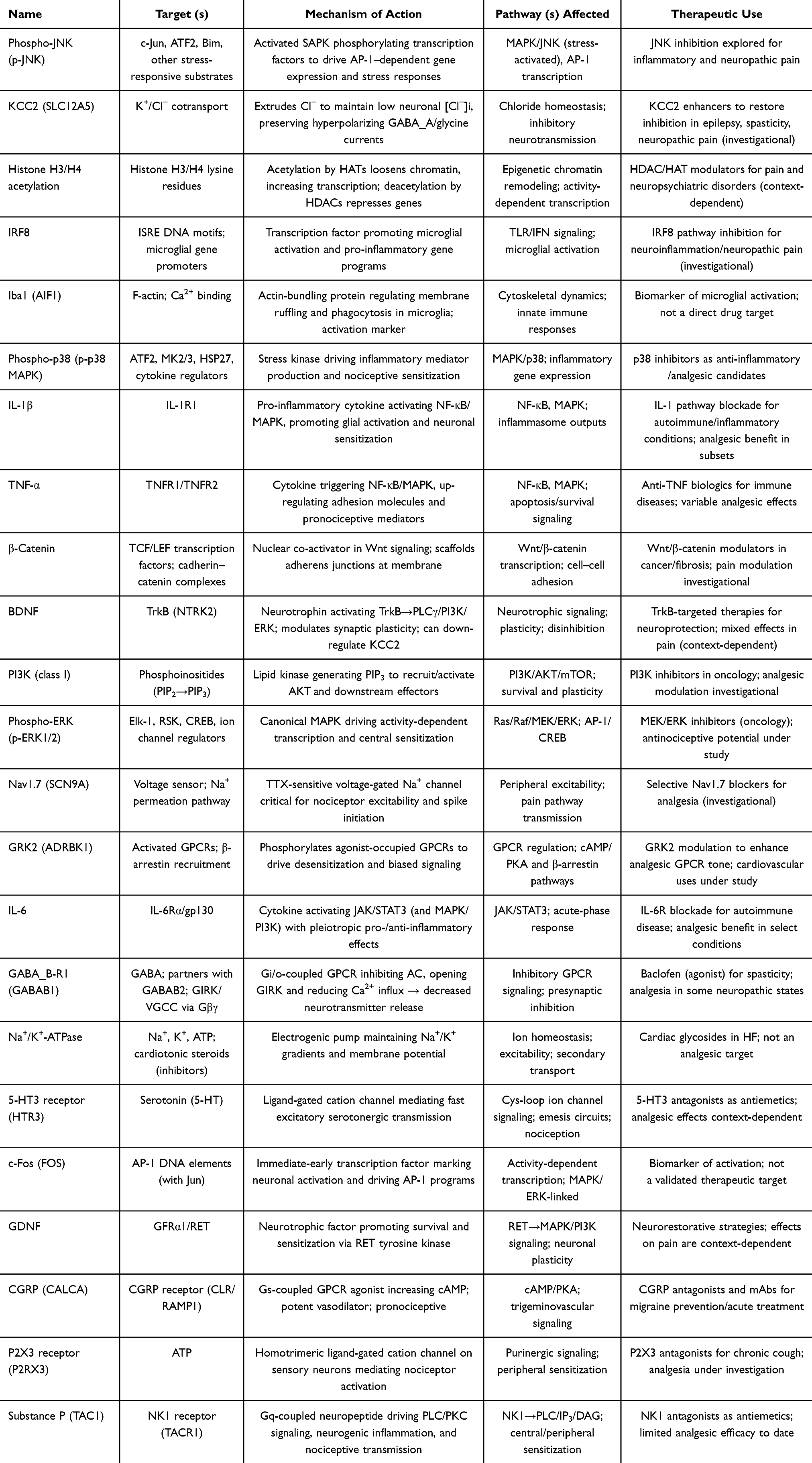 |
Table 3 Molecular/Signaling Factor Names, Targets, Mechanism of Action, Pathways Affected, and Therapeutic Uses |
At the DRG, high-voltage PRF (HV-PRF) down-regulated Nav1.7 and improved ultrastructure, whereas early-after-injury PRF more effectively suppressed p-ERK and Nav1.7 than delayed application, underscoring a timing/parameter dependence of molecular effects. Along the peptidergic axis, PRF lowered CGRP in the DRG and Substance P in the dorsal horn, while P2X3 purinoceptors were reduced in DRG and dorsal horn. In resiniferatoxin-induced neuropathy, PRF normalized TRPV1/CGRP and modulated BDNF. Two sciatic-nerve studies showed GDNF up-regulation with behavioral improvement and ultrastructural recovery. Together with DRG-targeted work reporting BDNF modulation30 and the pain–depression link (↓ spinal IRF8, ↑ prefrontal BDNF) after sciatic PRF, these results support a broader neuro-immune–neurotrophic model for PRF.
The most compelling mechanistic clue is restoration of spinal inhibitory tone. The only study with direct epigenetic measurement demonstrated that PRF countered CFA-induced histone (H3/H4) hypoacetylation at the Kcc2 locus, restoring KCC2 expression. This provides a concrete molecular bridge between PRF delivery and disinhibition reversal. Spinal met-enkephalin increased after PRF, supporting enhanced inhibitory transmission.31 Other studies align indirectly: reduced MAPK activation and decreased excitatory mediators (TRPV1/CGRP, Substance P, P2X3) are coherent with a shift toward inhibition. However, beyond Liu 2017, evidence remains indirect; no work has profiled DNA methylation, histone marks beyond H3/H4 at Kcc2, or noncoding RNAs in a hypothesis-driven, target-specific fashion after PRF.
DRG-directed PRF repeatedly impacts spinal microglial/MAPK pathways, supporting a principal site of action at primary afferent somata with downstream spinal effects. Sciatic-nerve PRF more often reports DRG peptide/channel normalization (TRPV1/CGRP, P2X3, Nav1.7) and neurotrophin shifts (GDNF, BDNF), yet spinal consequences (eg, ↓ Substance P, ↓ IRF8) still emerge. This pattern supports axon–soma–spinal coupling and suggests that procedural choice (DRG vs peripheral nerve) may be tailored to dominant pain mechanisms, though translational validation is lacking.
The superiority of HV-PRF for Nav1.7 and structural DRG effects, and the enhanced efficacy of earlier PRF after injury,4 together imply a window of plasticity and a dose–response relationship. Yet reporting of PRF settings (voltage, pulse width/interval, exposure time, tip temperature) is frequently incomplete, hampering cross-study comparison and precluding a formal exposure–response analysis.
Strengths of this work include convergent molecular findings across multiple labs and models (CCI, SNI, resiniferatoxin), consistent behavioral–molecular coupling, and one direct epigenetic demonstration. Limitations are the presence of exclusively rodent data, the heterogeneity of targets, time-points, and PRF parameters, the single time-point sampling in most studies, which obscures temporal trajectories. Sex, strain, and analgesic co-treatments are also variably reported, and blinding/randomization details are inconsistent, increasing risk of bias.
Our JBI risk-of-bias assessment confirmed moderate methodological quality across the included studies (Table 2). Most experiments lacked randomization, allocation concealment, and blinding, while reporting of PRF parameters was incomplete and conflicts of interest were seldom declared. These limitations reduce reproducibility and highlight the need for more rigorous experimental standards in future preclinical work.
The rodent data justify clinical plausibility that PRF may reduce pain by tuning neuro-immune signaling and restoring inhibitory balance without thermal lesioning. However, translation requires human evidence. Indeed, early clinical studies have already demonstrated analgesic benefits of PRF and innovative RF techniques in knee osteoarthritis and lumbar facet joint pain, further reinforcing the translational relevance of these preclinical findings.5,6,32
Conclusion
Across murine models, PRF at the DRG or peripheral nerves is consistently associated with molecular shifts spanning microglial/MAPK signaling, ion channels and neuropeptides (eg, P2X3, TRPV1/CGRP, Substance P), and restoration of inhibitory tone (KCC2), in parallel with behavioral analgesia. These findings support a biologically plausible, non-neurodestructive
Future research is needed in order to standardize PRF parameter, to replicate and extend locus-specific epigenetic findings (beyond Kcc2) with time-course designs, to incorporate unbiased multi-omic and single-cell approaches across DRG–spinal circuits and to embed mechanistic endpoints in prospective human studies.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Cohen SP, Bhaskar A, Bhatia A, et al. Consensus practice guidelines on interventions for lumbar facet joint pain from a multispecialty, international working group. Reg Anesth Pain Med. 2020;45(6):424–11. doi:10.1136/rapm-2019-101243
2. Vanneste T, Belba A, Kallewaard JW, et al. Comparison of cooled versus conventional radiofrequency treatment of the genicular nerves for chronic knee pain: a multicenter non-inferiority randomized pilot trial (COCOGEN trial). Reg Anesth Pain Med. 2023;48(5):197–204. doi:10.1136/rapm-2022-104054
3. Waicus S, Bhambra N. Pulsed radiofrequency treatment of the dorsal root ganglion in patients with chronic neuropathic pain: a narrative review. Indian J Pain. 2022;36(2):75–83. doi:10.4103/ijpn.ijpn_79_21
4. Yeh CC, Wu ZF, Chen JC, et al. Association between extracellular signal-regulated kinase expression and the anti-allodynic effect in rats with spared nerve injury by applying immediate pulsed radiofrequency. BMC Anesthesiol. 2015;15:92. doi:10.1186/s12871-015-0071-3
5. Leoni MLG, Schatman ME, Demartini L, Lo Bianco G, Terranova G. Genicular nerve pulsed dose radiofrequency (PDRF) compared to intra-articular and genicular nerve PDRF in knee osteoarthritis pain: a propensity score-matched analysis. J Pain Res. 2020;13:1315–1321. PMID: 32581573; PMCID: PMC7280065. doi:10.2147/JPR.S240138
6. Papa A, Di Dato MT, Lo Bianco G, et al. Intraarticular STP radiofrequency for painful osteoarthritis in the knee: a retrospective single center analysis. J Pain Res. 2021;14:2441–2447. PMID: 34413678; PMCID: PMC8370597. doi:10.2147/JPR.S317569
7. Liu R, Xu X, Xu Y, Fang X, Lin X. Pulsed radiofrequency on dorsal root ganglion relieved neuropathic pain associated with downregulation of the spinal interferon regulatory factor 8, microglia, p38MAPK expression in a CCI rat model. Pain Physician. 2018;21(4):E307–E322.
8. Xu X, Fu S, Shi X, Liu R. Microglial, BDNF, PI3K, and p-ERK in the spinal cord are suppressed by pulsed radiofrequency on dorsal root ganglion to ease SNI-induced neuropathic pain in rats. Pain Res Manag. 2019;2019:5948686. doi:10.1155/2019/5948686
9. Fu M, Meng L, Ren H, Luo F. Pulsed radiofrequency inhibits expression of P2X3 receptors and alleviates neuropathic pain induced by chronic constriction injury in rats. Chin Med J. 2019;132(14):1706–1712. doi:10.1097/CM9.0000000000000302
10. Ren H, Jin H, Jia Z, Ji N, Luo F. Pulsed radiofrequency applied to the sciatic nerve improves neuropathic pain by down-regulating the expression of calcitonin gene-related peptide in the dorsal root ganglion. Int J Med Sci. 2018;15(2):153–160. doi:10.7150/ijms.20501
11. Chen KH, Yang CH, Juang SE, et al. Pulsed radiofrequency reduced complete Freund’s adjuvant-induced mechanical hyperalgesia via the spinal c-Jun N-terminal kinase pathway. Cell Mol Neurobiol. 2014;34(2):195–203. doi:10.1007/s10571-013-0003-z
12. Wang JA, Niu SN, Luo F. Pulsed radiofrequency alleviated neuropathic pain by down-regulating the expression of substance P in chronic constriction injury rat model. Chin Med J. 2020;133(2):190–197. doi:10.1097/CM9.0000000000000619
13. Jiang R, Li P, Yao YX, et al. Pulsed radiofrequency to the dorsal root ganglion or the sciatic nerve reduces neuropathic pain behavior, decreases peripheral pro-inflammatory cytokines and spinal β-catenin in chronic constriction injury rats. Reg Anesth Pain Med. 2019;44(7):742–746. doi:10.1136/rapm-2018-100032
14. Liu CK, Liao WT, Chu YC, et al. Pulsed radiofrequency attenuates complete Freund’s adjuvant-induced epigenetic suppression of potassium chloride cotransporter 2 expression. Pain Med. 2017;18(4):807–813. doi:10.1093/pm/pnw243
15. Rethlefsen ML, Kirtley S, Waffenschmidt S, et al. PRISMA-S: an extension to the PRISMA statement for reporting literature searches in systematic reviews. Syst Rev. 2021;10(1):39. doi:10.1186/s13643-020-01542-z
16. Campbell M, McKenzie JE, Sowden A, et al. Synthesis without meta-analysis (SWiM) in systematic reviews: reporting guideline. BMJ. 2020;368:l6890. doi:10.1136/bmj.l6890
17. Hooijmans CR, Rovers MM, de Vries RBM, Leenaars M, Ritskes-Hoitinga M, Langendam MW. SYRCLE’s risk of bias tool for animal studies. BMC Med Res Methodol. 2014;14:43. doi:10.1186/1471-2288-14-43
18. Percie du Sert N, Hurst V, Ahluwalia A, et al. The ARRIVE guidelines 2.0: updated guidelines for reporting animal research. PLoS Biol. 2020;18(7):e3000410. doi:10.1371/journal.pbio.3000410
19. du Sert N P, Ahluwalia A, Alam S, et al. Reporting animal research: explanation and elaboration for the ARRIVE guidelines 2.0. PLoS Biol. 2020;18(7):e3000411. doi:10.1371/journal.pbio.3000411
20. Xu X, Chen R, Yu Y, Yang J, Lin C, Liu R. Pulsed radiofrequency on DRG inhibits hippocampal neuroinflammation by regulating spinal GRK2/p38 expression and enhances spinal autophagy to reduce pain and depression in male rats with spared nerve injury. Int Immunopharmacol. 2024;127:111419. doi:10.1016/j.intimp.2023.111419
21. Dai Z, Xu X, Chen Y, Lin C, Lin F, Liu R. Effects of high-voltage pulsed radiofrequency on the ultrastructure and Nav1.7 level of the dorsal root ganglion in rats with spared nerve injury. Neuromodulation. 2022;25(7):980–988. doi:10.1111/ner.13527
22. Vallejo R, Tilley DM, Williams J, Labak S, Aliaga L, Benyamin RM. Pulsed radiofrequency modulates pain regulatory gene expression along the nociceptive pathway. Pain Physician. 2013;16(5):E601–E613.
23. Koshida T, Maruta T, Tanaka N, et al. Changes in TRPV1 receptor, CGRP, and BDNF expression in rat dorsal root ganglion with resiniferatoxin-induced neuropathic pain: modulation by pulsed radiofrequency applied to the sciatic nerve. Acta Med Okayama. 2023;77(4):359–364. doi:10.18926/AMO/65741
24. Hidaka K, Maruta T, Koshida T, et al. Extracellular signal-regulated kinase phosphorylation enhancement and NaV1.7 sodium channel upregulation in rat dorsal root ganglia neurons contribute to resiniferatoxin-induced neuropathic pain: the efficacy and mechanism of pulsed radiofrequency therapy. Mol Pain. 2022;18:17448069221089784. doi:10.1177/17448069221089784
25. Lee JB, Byun JH, Choi IS, Kim Y, Lee JS. The effect of pulsed radiofrequency applied to the peripheral nerve in chronic constriction injury rat model. Ann Rehabil Med. 2015;39(5):667–675. doi:10.5535/arm.2015.39.5.667
26. Lyu C, Zhou J, Wu Q, Fang X. Spinal Interferon Regulatory Factor 8 and Brain-derived Neurotrophic Factor in the Prefrontal Cortex are Involved in Pain-induced Depression Relief via Ultrasound-guided Pulsed Radiofrequency in a Rat Spared Nerve Injury Model. Pain Phys. 2023;26(2):187–196.
27. Jia Z, Ren H, Li Q, Ji N, Luo F. Pulsed radiofrequency reduced neuropathic pain behavior in rats associated with upregulation of GDNF expression. Pain Physician. 2016;19(2):49–58.
28. Jin H, Ren H, Jia Z, Ji N, Luo F. Pulsed radiofrequency improves neuropathic pain in chronic constriction injury rats through the upregulation of the transcription and translation levels of glial cell line-derived neurotrophic factor. Pain Physician. 2018;21(1):33–40.
29. Yi HY, Hou XM, Guo YX, et al. Brain-derived neurotrophic factor in the ventrolateral orbital cortex regulates spared nerve injury-induced neuropathic pain and related emotional disorders in mice. Neuroscience. 2025;584:250–263. doi:10.1016/j.neuroscience.2025.08.032
30. Fang X, Xu X, Lin X, Liu R. Downregulated spinal IRF8 and BDNF in NAC are involved in neuropathic pain-induced depression relief via pulsed radiofrequency on dorsal root ganglion in rat SNI model. Brain Res Bull. 2019;146:192–200. doi:10.1016/j.brainresbull.2019.01.008
31. Wu B, Ni J, Zhang C, Fu P, Yue J, Yang L. Changes in spinal cord met-enkephalin levels and mechanical threshold values of pain after pulsed radio frequency in a spared nerve injury rat model. Neurol Res. 2012;34(4):408–414. doi:10.1179/1743132812Y.0000000026
32. Lo Bianco G, Misseri G, Stogicza AR, et al. Radiofrequency Ablation for Chronic Lumbar Zygapophyseal Joint Pain Using a V-Shaped Active Tip Needle: an Observational Retrospective Study. J Pain Res. 2023;16:1243–1255. PMID: 37069943; PMCID: PMC10105587. doi:10.2147/JPR.S406714
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.