Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 16
Proteomic Identification and Quantification of Secretory Proteins in Human Dermal Fibroblast-Conditioned Medium for Wound Repair and Hair Regeneration
Authors Suh SB, Ahn KJ ![]() , Kim EJ
, Kim EJ ![]() , Suh JY
, Suh JY ![]() , Cho SB
, Cho SB ![]()
Received 18 February 2023
Accepted for publication 27 April 2023
Published 1 May 2023 Volume 2023:16 Pages 1145—1157
DOI https://doi.org/10.2147/CCID.S407078
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Jeffrey Weinberg
Sang Bum Suh,1,* Keun Jae Ahn,2,* Eun Jeong Kim,1 Ji Youn Suh,1 Sung Bin Cho3
1BNV Biolab, Seoul, Korea; 2Department of Science Education, Jeju National University, Jeju, Korea; 3Yonsei Seran Dermatology and Laser Clinic, Seoul, Korea
*These authors contributed equally to this work
Correspondence: Sung Bin Cho, Yonsei Seran Dermatology and Laser Clinic, 224 Siheung-daero, Seoul, 08628, Korea, Tel +82 2-2135-1375, Fax +82 70-8250-1375, Email [email protected]
Background: Human dermal fibroblasts secrete numerous growth factors and proteins that have been suggested to promote wound repair and hair regeneration.
Methods: Human dermal fibroblast-conditioned medium (DFCM) was prepared, and proteomic analysis was performed. Secretory proteins in DFCM were identified using 1-dimensional sodium dodecyl sulphate-polyacrylamide gel electrophoresis, in-gel trypsin protein digestion, and quantitative liquid chromatography tandem mass spectrometry (LC-MS/MS). Identified proteins were analyzed using bioinformatic methods for the classification and evaluation of protein–protein interactions.
Results: Using LC-MS/MS, 337 proteins were identified in DFCM. Among them, 160 proteins were associated with wound repair, and 57 proteins were associated with hair regeneration. Protein–protein interaction network analysis of 160 DFCM proteins for wound repair at the highest confidence score (0.9) revealed that 110 proteins were grouped into seven distinctive interaction networks. Additionally, protein–protein interaction network analysis of 57 proteins for hair regeneration at the highest confidence score revealed that 29 proteins were grouped into five distinctive interaction networks. The identified DFCM proteins were associated with several pathways for wound repair and hair regeneration, including epidermal growth factor receptor, fibroblast growth factor, integrin, Wnt, cadherin, and transforming growth factor-β signaling pathways.
Conclusion: DFCM contains numerous secretory proteins that comprise groups of protein–protein interaction networks that regulate wound repair and hair regeneration.
Keywords: human dermal fibroblast, dermal fibroblast-conditioned media, proteomic analysis, secretory protein, wound repair, hair regeneration
Introduction
Fibroblasts secrete various growth factors, cytokines, chemokines, and other peptide or protein factors that play essential roles in regulating wound repair.1–5 Secretomes from fibroblasts include vascular endothelial growth factor (VEGF), fibroblast growth factor (FGF), hepatocyte growth factor (HGF), platelet-derived growth factor (PDGF), transforming growth factor (TGF)-β1, keratinocyte growth factor (KGF), interleukin (IL)-6, and IL-8.1–5 The secretion patterns of fibroblast-derived factors significantly varies among species, anatomic sites, sub-types of fibroblast, and experimental culture conditions for obtaining secretomes.4–7
Researchers have discovered that dermal fibroblast-conditioned medium (DFCM) contains numerous protein factors that reportedly promote wound repair.5–7 A previous proteomic study demonstrated that DFCM contains various secretory factors including fibronectin, fibulin-1, collagen type I α-1 chain (COL1A1), COL4A1, COL12A1, serum albumin, α-fetoprotein, pigment epithelium-derived factor, decorin, lactotransferrin, nucleobindin-1, inter-α-trypsin inhibitor heavy chain H2, pentraxin 3, α-2-HS-glycoprotein, α-2-macroglobin, TGF-β-induced protein ig-h3, pregnancy zone protein, and thrombospondin-1.5 Another growth factor G1 array study also revealed that DFCM contains numerous growth factors, which are quantitatively increased by 20% compared with control medium and include epidermal growth factor (EGF), basic FGF, FGF-7, insulin-like growth factor binding protein (IGFBP)-4, and IGFBP-6.8 Moreover, the outer root sheath (ORS) cells of hair follicles that were cultured with DFCM-supplemented cultured medium secreted notably higher amounts of growth factors compared with control ORS cells.8
In this study, we aimed to identify secretory proteins from human dermal fibroblasts, which have crucial roles in wound repair and hair regeneration, using proteomic analysis. To do so, DFCM was prepared to obtain fibroblast-derived secretory proteins under the culture conditions of low serum and high hypoxanthine and thymidine, as described in a previous study.8 Then, proteomic analysis was performed using 1-dimensional sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE), in-gel trypsin protein digestion, and quantitative liquid chromatography tandem mass spectrometry (LC-MS/MS). Proteins identified in DFCM were further analyzed using bioinformatic methods for the classification and evaluation of theoretical protein–protein interactions.
Materials and Methods
Culture of Human Dermal Fibroblasts
Neonatal human dermal fibroblasts (NHDF-Neo; Lonza, Walkersville, MD, USA) were mixed with 40-mL fibroblast growth medium (FGMTM; Lonza) at a 1:1 ratio. Cells (4 × 105 cells) were seeded into a 75-cm2-T-flask and incubated overnight at 37°C under 5% CO2. The medium was replaced with Opti-modified Eagle’s medium (Opti-MEM; Gibco BRL, Rockville, MD, USA) to create low-serum culture conditions and the cells were cultured for another 24 h to reach approximately 90% confluence. Cells were then washed with Dulbecco’s phosphate buffered saline (PBS; Santa Cruz Biotechnology, Santa Cruz, CA, USA) and treated with 0.15% trypsin (Gibco BRL) for 5 min at 37°C to detach cells from the surface of the culture flask. Medium was added to arrest trypsinization, and the cells were centrifuged for 5 min. The collected cells were resuspended in Opti-MEM and split into 75-cm2-T flasks. Cells were expanded until enough cells were prepared for experiments. To prepare DFCM, human dermal fibroblasts (passage 5) were cultured in 175-cm2-T flasks with Opti-MEM supplemented with 2-mM GlutaMAX, 2-ng/mL recombinant human EGF, and 2-ng/mL recombinant human FGF (all purchased from Lonza) without the addition of fetal bovine serum. The cells were incubated at 37°C in a 5% CO2 incubator for 24 h, and the medium was collected as DFCM.
Protein Filtration and Concentration for Proteomic Analysis
The prepared DFCM was filtered through a 3-kDa Amicon Ultra-15 centrifugal filter (Merck Millipore, Burlington, MA, USA) to concentrate the proteins. DFCM-containing tubes were centrifuged at 4000 × g for 40 min at 25°C. Proteins with molecular weights > 3 kDa retained on the membrane surface were collected for dialysis. A PlusOne Mini Dialysis Kit (GE Healthcare, Buckinghamshire, UK) with a 1-kDa cut-off was used to dialyze the DFCM, which allowed for the desalting and removal of small molecules. The dialyzed samples were stored at −80°C for proteomic analysis.
One-Dimensional Gel Electrophoresis and in-Gel Digestion
The protein components of DFCM (30 x concentrate), Opti-MEM, Opti-MEM supplemented with GlutaMAX, recombinant human EGF (2 ng/mL) and FGF (2 ng/mL) (30 x concentrate), EGF (0.1 mg/mL), FGF (0.1 mg/mL), and GlutaMAX were separated using 2% SDS-PAGE (Figure 1A). Then, the protein components (10 μg) of DFCM were additionally separated using 10% SDS-PAGE for in-gel digestion using a previously described method.9 Briefly, the gels were divided into four fractions according to molecular size after Coomassie blue staining (Figure 1B). Sliced gels were destained in a solution of 50% acetonitrile and 10 mM ammonium bicarbonate. The gels were then rinsed with distilled water, followed by 100% acetonitrile, to remove the destaining solution. Protein samples were sequentially treated with a reducing solution (10 mM dithiothreitol and 100 mM ammonium bicarbonate) and an alkylation solution (55 mM iodoacetamide). After the gels were washed with distilled water, tryptic digestion at a final concentration of 10 ng/mL was performed in 50 mM ammonium bicarbonate at 37°C for 12−16 h. Extraction of the tryptic peptides was performed in an extraction solution (50 mM ammonium bicarbonate and 50% acetonitrile containing 5% trifluoroacetyl (TFA)). The resulting peptide extracts were pooled and lyophilized. Tryptic peptides were dissolved in 0.5% TFA prior to further fractionation using LC-MS/MS.
 |
Figure 1 One-dimensional gel electrophoresis using human dermal fibroblast-conditioned medium (DFCM) preparation. (A) The protein components of (1) DFCM, (2) Opti-MEM, (3) Opti-MEM supplemented with GlutaMAX, recombinant human epidermal growth factor (EGF) and fibroblast growth factor (FGF), (4) EGF, (5) FGF, and (6) GlutaMAX were separated using 2% sodium dodecyl sulphate (SDS)-polyacrylamide gel electrophoresis (PAGE). (B) The protein components of DFCM were separated using 10% SDS-PAGE. Protein bands of DFCM were categorized into four groups according to the ranges of molecular weights. |
LC−MS/MS Analysis and Bioinformatic Analysis
Quantitative liquid chromatography tandem mass spectrometry analysis was performed according to a modified version of a previously published method.10 Tryptic peptide samples were loaded onto a trap column (Thermo Fisher Scientific, Bremen, Germany) for the enrichment of peptides and removal of chemical contaminants. Concentrated tryptic peptides were eluted from the column and directed onto a 150 mm × 75 µm i.d. C18 reverse phase column (Thermo Fisher Scientific) at a flow rate of 0.3 µL/min. Peptides were eluted using a gradient of 0–65% acetonitrile for 80 min. All MS and MS/MS spectra were acquired in data-dependent mode using a Q Exactive plus ESI Ion Trap mass spectrometer (Thermo Fisher Scientific). Each full MS scan (m/z range 150–2000) scan was followed by three MS/MS scans of the most abundant precursor ions in the MS spectra. The data from LC−MS/MS were searched using the Mascot database (http://www.matrixscience.com/) against the SwissProt Human Database. We estimated the false discovery rate of peptide spectrum matches using decoy databases. Proteins that were identified via LC−MS/MS were analyzed and classified using the Protein Analysis Through Evolutionary Relationships (PANTHER™) Classification System v.17.0 (http://pantherdb.org/) and Search Tool for Retrieval of Interacting Genes/Proteins (STRING®) v.11.5 protein interaction network (http://www.string-db.org/) to classify and predict protein–protein interactions.11,12
Results
LC-MS/MS Identification of Human Fibroblast-Derived Secretory Proteins in DFCM
Quantitative liquid chromatography tandem mass spectrometry using matrix-assisted laser desorption/ionization tandem time-of-flight (MALDI-TOF/TOF) was performed to identify the overall proteins in DFCM that contained human fibroblast-derived secretory proteins. To separate the protein mixtures, 1-dimensional SDS-PAGE was performed using the DFCM preparation. Protein bands were then categorized into four groups according to their molecular weights. Each group of bands was subjected to LC-MS/MS using MALDI-TOF/TOF, followed by SwissProt database searches. First, we identified 337 proteins, the known functions of which were reviewed from the UniProt Database to identify proteins potentially involved in wound repair and hair regeneration. A total of 160 unique proteins that were proposed to be involved in wound repair and 57 unique proteins that were deemed to be associated with hair regeneration were selected.
Protein–Protein Interaction Network Analysis of DFCM Proteins for Wound Repair
The 160 proteins of the wound repair group were further evaluated according to the results of protein–protein interaction analysis using the protein interaction network. At a medium confidence level (0.4) of interaction scores, a single protein–protein interaction network, which consisted of 110 proteins with notable connections among proteins, was identified, and the remaining 50 proteins without significant interactions were excluded (Figure 2A). With a high confidence score (0.7), 110 proteins were categorized into two distinct interaction networks without any disconnected proteins (Figure 2B). At the highest confidence score (0.9), 110 proteins were grouped into seven interaction networks (Figure 2C), and a list of proteins in each group is summarized in Table 1 and Table 2.
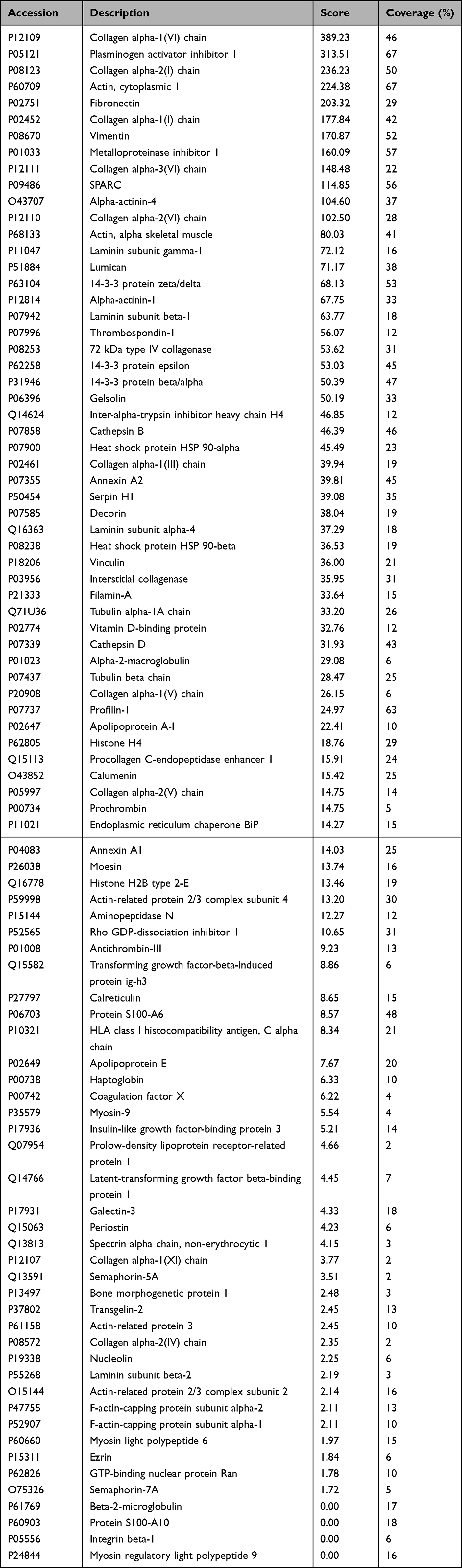 |
Table 1 The Major Distinctive Protein-Protein Interaction Network Group of 89 Human Dermal Fibroblast-Conditioned Medium (DFCM) Proteins for Wound Repair at a Confidence Score of 0.9 |
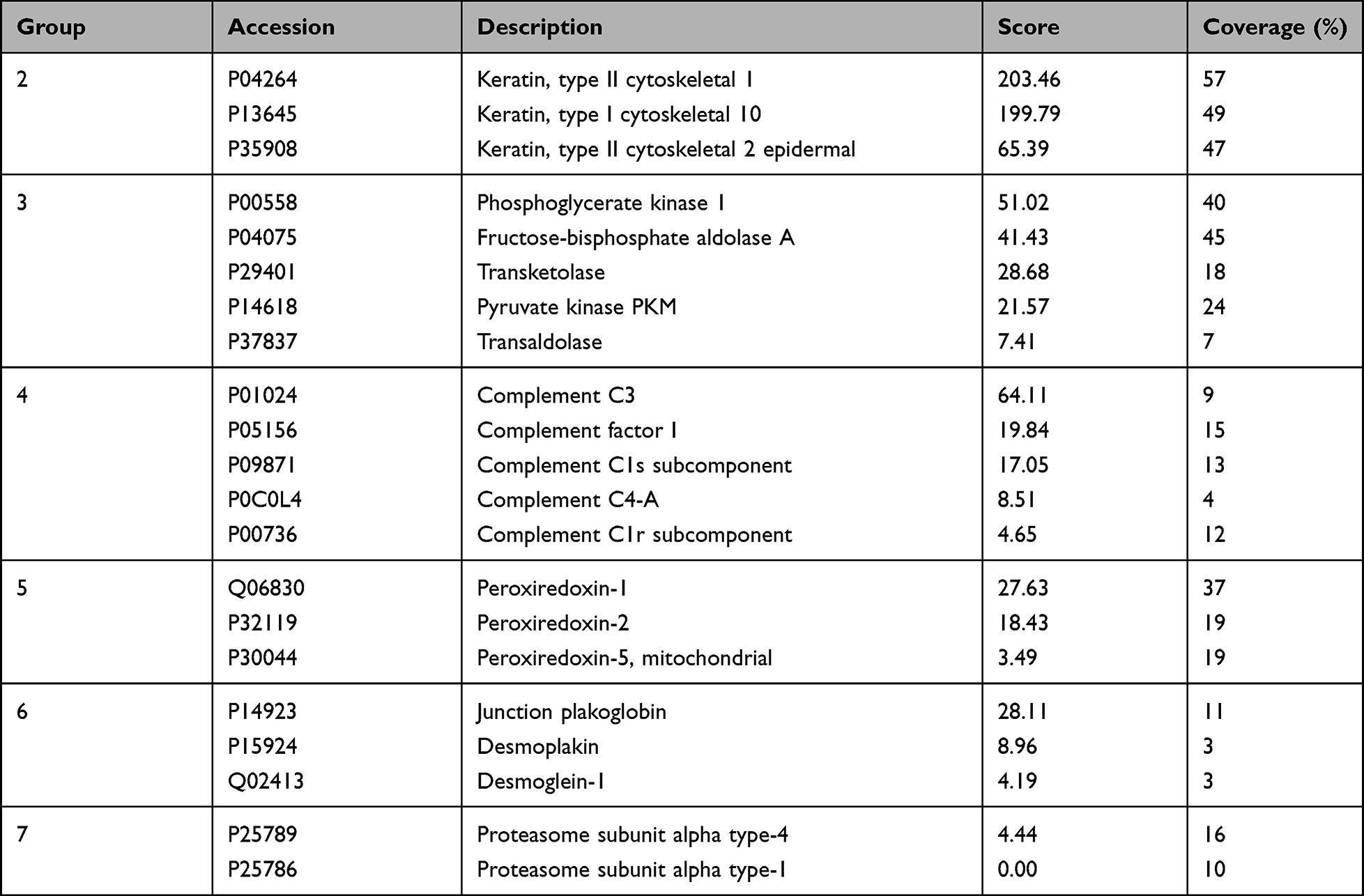 |
Table 2 The Other 6 Distinctive Protein-Protein Interaction Network Groups of 21 DFCM Proteins for Wound Repair at a Confidence Score of 0.9 |
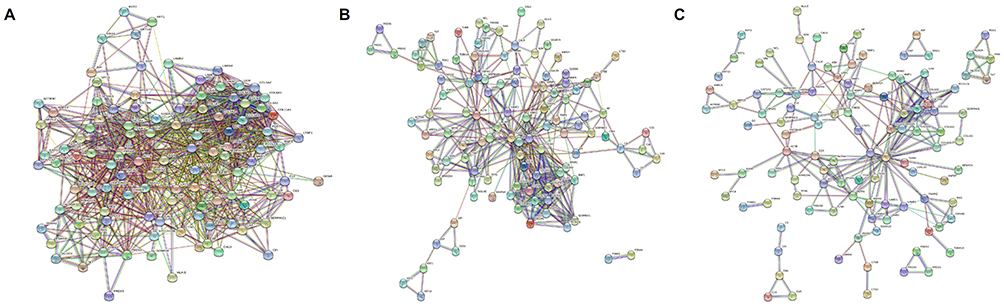 |
Figure 2 Protein–protein interaction network analysis of DFCM proteins for wound repair. (A) At a medium confidence level (0.4) of interaction scores, a single protein–protein interaction network, which consisted of 110 proteins with notable connections among them, was observed. (B) At a high confidence score (0.7), 110 proteins were categorized into two distinct interaction networks without any disconnected protein members. (C) At the highest confidence score (0.9), 110 proteins were grouped into seven interaction networks. |
Gene Ontology (GO) Protein Classification of DFCM Proteins for Wound Repair
Using the protein classification database, GO classified the identified proteins and classified 110 proteins in the DFCM into three categories: molecular function, cellular component, and biological process. The major molecular functions of these proteins included binding, catalytic activity, molecular function regulation, and molecular transducer activity (Figure 3A). The identified cellular components of these proteins included cellular anatomical entities and protein-containing complexes (Figure 3B). The major biological processes included cellular processes, biological regulation, responses to stimuli, and metabolic processes (Figure 3C). The protein classes identified in DFCMs that are important for wound healing were cytoskeletal proteins, extracellular matrix proteins, protein-modifying enzymes, and protein-binding activity modulators (Figure 4A). Interestingly, DFCM proteins were associated with several remarkable pathways during wound repair, which included EGF receptor signaling, FGF signaling, Wnt signaling, apoptosis signaling, and TGF-β signaling pathways (Figure 4B).
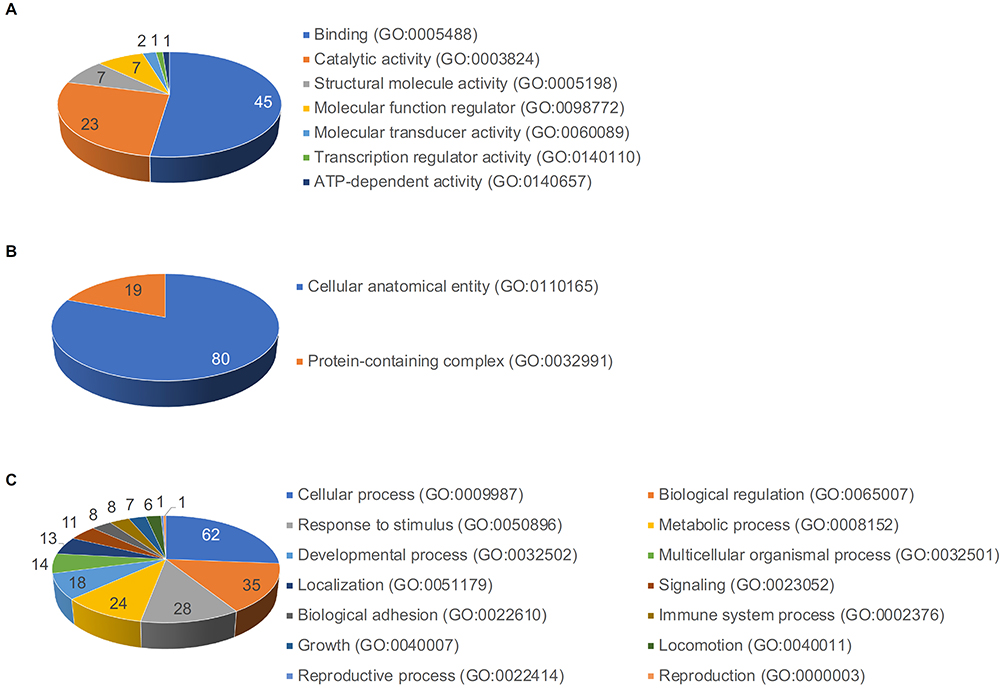 |
Figure 3 Gene Ontology (GO) protein classification of DFCM proteins for wound repair. GO classified the identified and selected 110 DFCM proteins in three categories: (A) molecular function, (B) cellular component, and (C) biological process. |
 |
Figure 4 GO protein classification of DFCM proteins for wound repair. (A) The identified protein classes in the DFCM that are important for wound healing were cytoskeletal protein, extracellular matrix protein, protein modifying enzyme, and protein-binding activity modulator. (B) DFCM proteins were associated with several pathways during wound repair. |
Protein–Protein Interaction Network Analysis of DFCM Proteins for Hair Regeneration
The 57 proteins of hair regeneration group were further evaluated according to the results of protein–protein interactions using the protein interaction network. At the medium confidence level of the interaction score, a single protein–protein interaction network, which consisted of 54 proteins with notable connections among proteins, was observed, and the other three proteins without significant interactions were excluded (Figure 5A). At a high confidence score, 40 of the 54 proteins were categorized into four distinctive interaction networks, and 14 proteins without notable interactions were excluded (Figure 5B). At the highest confidence score, 29 of the 40 proteins were grouped into five interaction networks (Figure 5C and Table 3), and the other 11 proteins without notable interactions were excluded.
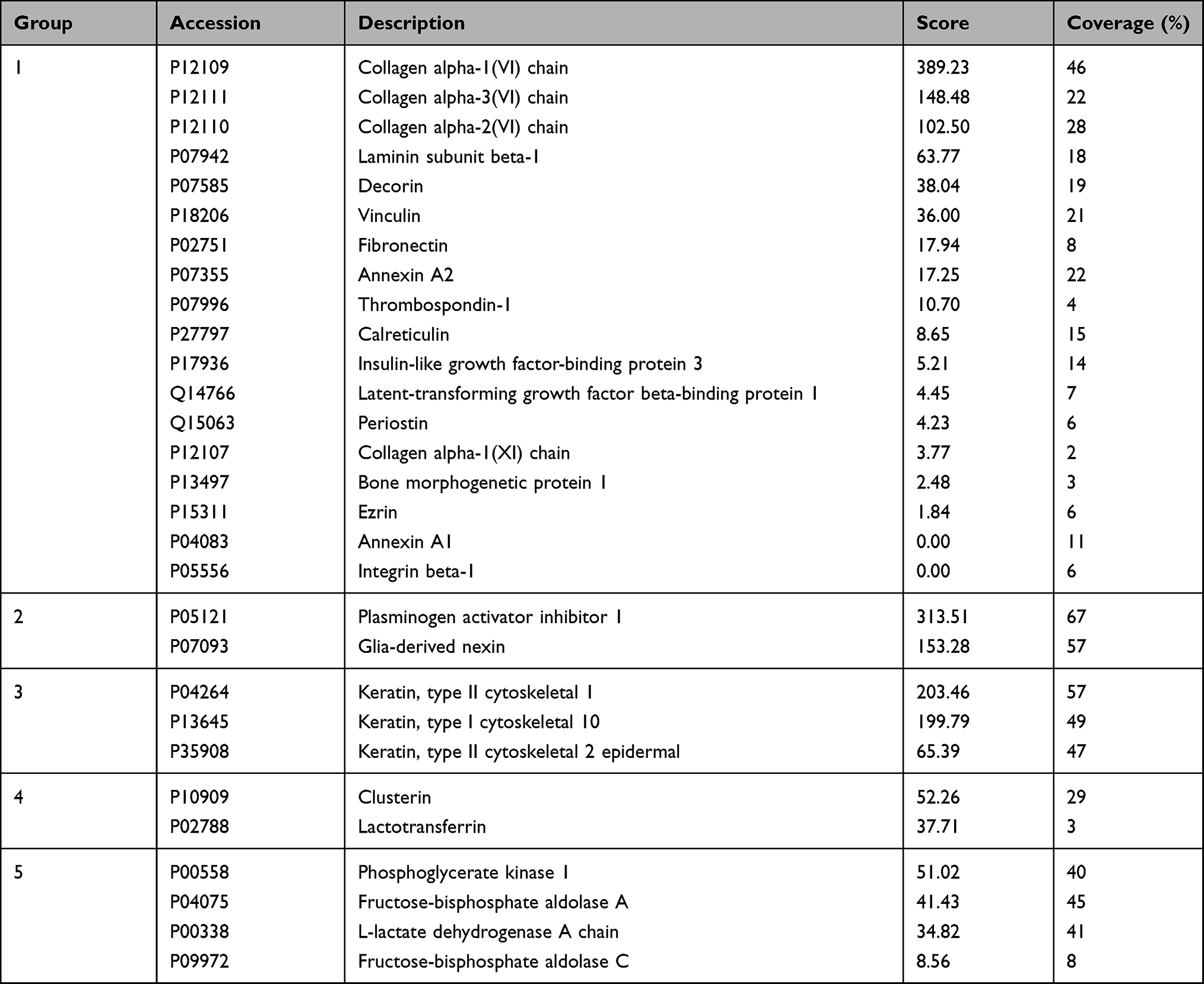 |
Table 3 The 5 Distinctive Protein-Protein Interaction Network Groups of 29 DFCM Proteins for Hair Regeneration at a Confidence Score of 0.9 |
 |
Figure 5 Protein–protein interaction network analysis of DFCM proteins for hair regeneration. (A) At a medium confidence of interaction score, a single protein–protein interaction network which consisted of 54 proteins with notable connections among proteins was observed. (B) At a high confidence score, 40 of 54 proteins were categorized into four distinct interaction networks. (C) At the highest confidence score, 29 of 40 proteins were grouped into five interaction networks. |
GO Protein Classification of DFCM Proteins for Hair Regeneration
Using the protein classification database, GO classified the identified and selected 29 proteins for hair regeneration in the DFCM on three categories: molecular function, cellular component, and biological process. The major molecular functions of these proteins included binding, catalytic activity, molecular function regulation, and structural molecule activity (Figure 6A). The identified cellular components of these proteins included cellular anatomical entities and protein-containing complexes (Figure 6B), whereas the major biological processes included cellular, biological regulation, developmental, and metabolic processes (Figure 6C). The protein classes identified in DFCMs that were important for hair regeneration were extracellular matrix proteins, metabolite interconversion enzymes, cell adhesion molecules, and protein-binding activity modulators (Figure 7A). Additionally, DFCM proteins were associated with several pathways during hair regeneration, which included the integrin signaling pathway, p53 pathway, fructose galactose metabolism, and TGF-β signaling pathways (Figure 7B).
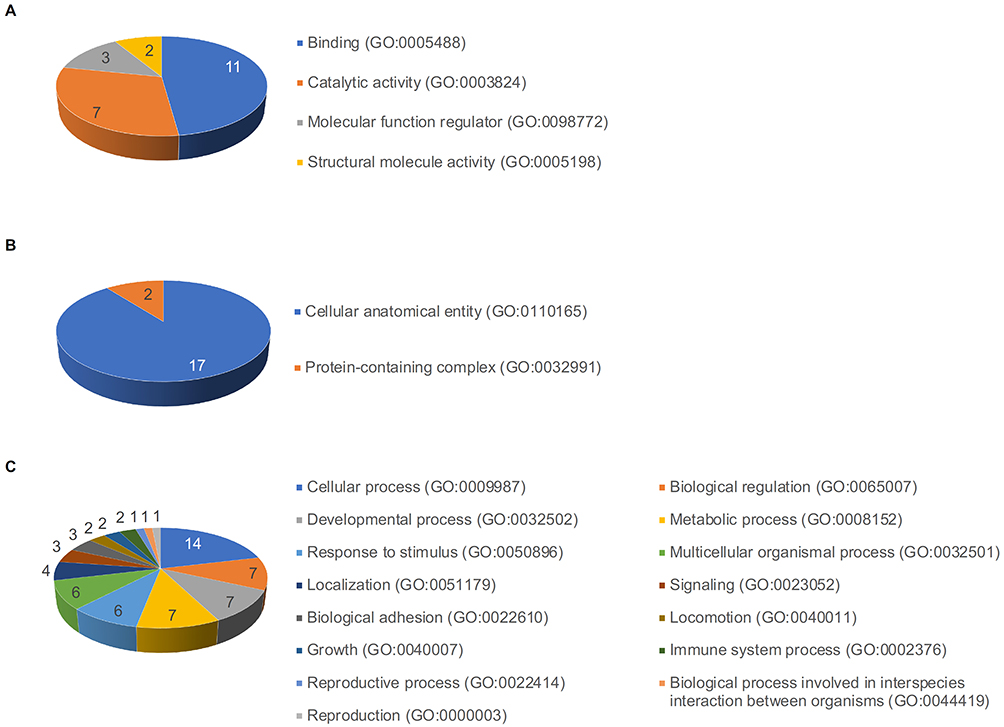 |
Figure 6 GO protein classification of DFCM proteins for hair regeneration. GO classified the 29 identified and selected proteins present in DFCM into three categories: (A) molecular function, (B) cellular component, and (C) biological process. |
 |
Figure 7 GO protein classification of DFCM proteins for hair regeneration. (A) The identified protein classes in the DFCM that are important for hair regeneration included extracellular matrix protein, metabolite interconversion enzyme, cell adhesion molecule, and protein-binding activity modulator. (B) DFCM proteins were associated with several pathways during hair regeneration. |
Discussion
The DFCM preparation in our study was obtained under culture conditions using Opti-MEM without the addition of fetal bovine serum to obtain fibroblast-secreted proteins containing no or minimal serum proteins.8,13 Although the culture medium was supplemented with recombinant human EGF and FGF, we believe that majority of the supplementary growth factors were used to induce protein secretion from fibroblasts. A previous study demonstrated that wound healing properties are improved in low- and high-energy nitrogen plasma-treated human dermal fibroblasts by culturing with the DFCM preparation.13 In our cell viability study and wound healing assays, the DFCM culture media was similarly effective compared with that of Opti-MEM, which was supplemented with synthetic growth factors.13 Moreover, the migration rates were higher in plasma-treated human dermal fibroblasts, which were further cultured with DFCM, compared with other culture conditions.13 Another study revealed that DFCM contains quantitatively elevated levels of growth factors, and DFCM remarkably induces growth factor secretion by the cells, which were cultured with DFCM.8 However, whether multiple factors in DFCM protect high-energy plasma-treated fibroblasts from apoptosis or stimulate cell proliferation remains to be elucidated. Furthermore, the mechanisms of action of factors other than growth factors in wound repair remain unclear.
In this study, we identified 337 secretory proteins from human dermal fibroblasts via proteomic analysis using DFCM preparations. Those proteins were further grouped according to the results of protein–protein interaction network analysis at the highest confidence score using the bioinformatic method. Then, 110 and 29 proteins that were associated with wound repair and hair regeneration, respectively, were selected for bioinformatic classification. Our study revealed that DFCM proteins were associated with various pathways, including EGF receptor, FGF, Wnt, apoptosis, and TGF-β signaling pathways, for promoting wound repair. Additionally, DFCM proteins are associated with the integrin signaling pathway, which activate signal transduction mediators to enhance wound closure.14,15 Moreover, the DFCM proteins in this study regulated cytoskeletal responses via Rho guanosine triphosphatases (GTPases). The Rho family of GTPases is necessary for the regulation of actin and myosin cytoskeletal dynamics during wound healing.16,17
In our bioinformatics study, DFCM proteins were also associated with the Wnt and cadherin signaling pathways. The stimulation of human dermal fibroblasts enhances migration, proliferation, and collagen synthesis by the cells, and gene expression of collagen I, III, cyclin-1, PCNA, and N-cadherin is also increased.18 Wnts are key regulators of cell proliferation and differentiation.19 Hence, the activation of the Wnt signaling pathway plays an essential role during the proliferative phase of wound repair.20 Wnt signaling and cadherin-mediated cell adhesion are significantly involved in the organization and maintenance of hematopoietic and germline stem cells.19 Moreover, dysregulation of Wnt signaling is associated with cancer induction and tumor progression. Additionally, the downregulation of cadherin expression can promote tumorigenesis.19 Therefore, we suggest that DFCM proteins can not only promote tissue regeneration during the wound healing process but can also inhibit the progression and migration of dysregulated cells.
Laser-assisted, power-, and energy-regulated wounding in the epidermis effectively induces Wnt-dependent hair follicle neogenesis.21,22 Additionally, the dermis harbors a CD49fhigh cell population, which bears an epidermal stem cell marker of mesenchymal stem cells.23 CD49fhigh cells act on the formation of hair follicle-like budding along with hair follicular epithelial cells.23 However, CD49fhigh cell-associated follicular neogenesis requires the activation of signaling pathways for regulating hair follicle development.23 Our previous study demonstrated that DFCM increases mRNA expression of KGF, EGF, and VEGF in ORS cells and KGF in dermal papilla cells.8 Moreover, DFCM activates β-catenin expression and inhibits Dickkopf-related protein 1 expression in dermal papilla cells.8 As the activation of Wnt/β-catenin signaling is important in wound repair, hair growth, and hair follicle morphogenesis, DFCM induces hair regeneration by upregulating growth factor expression and activating signaling pathways.8,24
The identified proteins in our study were functionally categorized into wound repair and hair regeneration groups by bioinformatically reviewing the UniProt Database. Our data demonstrated that several peptide factors and signaling pathways are associated with hair regeneration. However, identified proteins in this study, which were proposed to be involved in wound repair, can also induce hair regeneration by accelerating mesenchymal cell proliferation and neovascularization.25–27 Therefore, although bioinformatical analyses could categorize DFCM proteins into different groups, we suggested that the roles of those proteins could overlap or interact between wound repair and hair regeneration.
Conclusion
In this study, we identified secretory proteins from human dermal fibroblasts by analyzing DFCM preparations using proteomic analysis. Our study revealed that hundreds of proteins were secreted by human dermal fibroblasts and these proteins presented significant protein–protein interaction networks. The identified proteins were further categorized into distinct groups based on their molecular functions, biological processes, cellular components, and protein classes. Bioinformatic analyses revealed that DFCM proteins represent components of various signaling pathways involved in wound repair and hair regeneration. However, further studies are required to investigate the precise role of each DFCM protein in wound repair and hair regeneration.
Data Sharing Statement
Data supporting the findings of this study are available upon request from the corresponding author. The data are not publicly available because of privacy and ethical restrictions.
Acknowledgments
We would like to thank Dr. Yeol-Gyun Lee, PhD (PROTEINWORKS Co., Ltd., Daejeon, Korea) for assistance in performing proteomic analysis. Keun Jae Ahn was supported by the 2022 Scientific Promotion Program funded by Jeju National University.
Author Contributions
All authors have made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the manuscript; gave final approval of the version to be published; agreed on the journal to which the manuscript would be submitted; and agreed to be accountable for all aspects of the work.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Lawlor KT, Kaur P. Dermal contributions to human interfollicular epidermal architecture and self-renewal. Int J Mol Sci. 2015;16:28098–28107. doi:10.3390/ijms161226078
2. Leary T, Jones PL, Appleby M, Blight A, Parkinson K, Stanley M. Epidermal keratinocyte self-renewal is dependent upon dermal integrity. J Invest Dermatol. 1992;99:422–430. doi:10.1111/1523-1747.ep12616134
3. Tuan TL, Keller LC, Sun D, Nimni ME, Cheung D. Dermal fibroblasts activate keratinocyte outgrowth on collagen gels. J Cell Sci. 1994;107:2285–2289. doi:10.1242/jcs.107.8.2285
4. Sorrell JM, Baber MA, Caplan AI. Site-matched papillary and reticular human dermal fibroblasts differ in their release of specific growth factors/cytokines and in their interaction with keratinocytes. J Cell Physiol. 2004;200:134–145. doi:10.1002/jcp.10474
5. Maarof M, Lokanathan Y, Ruszymah HI, Saim A, Chowdhury SR. Proteomic analysis of human dermal fibroblast conditioned medium (DFCM). Protein J. 2018;37:589–607. doi:10.1007/s10930-018-9800-z
6. Kubo K, Kuroyanagi Y. A study of cytokines released from fibroblasts in cultured dermal substitute. Artif Organs. 2005;29:845–849. doi:10.1111/j.1525-1594.2005.00138.x
7. Chowdhury SR, Aminuddin BS, Ruszymah BH. Effect of supplementation of dermal fibroblasts conditioned medium on expansion of keratinocytes through enhancing attachment. Indian J Exp Biol. 2012;50:332–339.
8. Shin JM, Lee YY, Kim KM, et al. The potential role of fibroblast-derived multi-peptide factors in activation of growth factors and β-catenin in hair follicle cells. J Cosmet Dermatol. 2022;21:6184–6190. doi:10.1111/jocd.15188
9. Yun SH, Park GW, Kim JY, et al. Proteomic characterization of the Pseudomonas putida KT2440 global response to a monocyclic aromatic compound by iTRAQ analysis and 1DE-MudPIT. J Proteomics. 2011;74:620–628.
10. Choi CW, Lee YG, Kwon SO, et al. Analysis of Streptococcus pneumoniae secreted antigens by immuno-proteomic approach. Diagn Microbiol Infect Dis. 2012;72:318–327. doi:10.1016/j.diagmicrobio.2011.12.013
11. Thio CL, Yusof R, Ashrafzadeh A, Bahari S, Abdul-Rahman PS, Karsani SA. Differential analysis of the secretome of WRL68 cells infected with the Chikungunya virus. PLoS One. 2015;10:e0129033. doi:10.1371/journal.pone.0129033
12. Creaney J, Dick IM, Leon JS, Robinson BW. A proteomic analysis of the malignant mesothelioma secretome using iTRAQ. Cancer Genomics Proteomics. 2017;14:103–117.
13. Lee SY, Kim DY, Suh SB, Suh JY, Cho SB. Effects of human fibroblast-derived multi-peptide factors on the proliferation and migration of nitrogen plasma-treated human dermal fibroblasts. Clin Cosmet Investig Dermatol. 2022;15:2465–2475. doi:10.2147/CCID.S383483
14. DiPersio CM, Zheng R, Kenney J, Van De Water L. Integrin-mediated regulation of epidermal wound functions. Cell Tissue Res. 2016;365:467–482. doi:10.1007/s00441-016-2446-2
15. Couture C, Desjardins P, Zaniolo K, Germain L, Guérin SL. Enhanced wound healing of tissue-engineered human corneas through altered phosphorylation of the CREB and AKT signal transduction pathways. Acta Biomater. 2018;73:312–325. doi:10.1016/j.actbio.2018.04.021
16. Abreu-Blanco MT, Verboon JM, Parkhurst SM. Coordination of Rho family GTPase activities to orchestrate cytoskeleton responses during cell wound repair. Curr Biol. 2014;24:144–155. doi:10.1016/j.cub.2013.11.048
17. Nakamura M, Verboon JM, Parkhurst SM. Prepatterning by RhoGEFs governs Rho GTPase spatiotemporal dynamics during wound repair. J Cell Biol. 2017;216:3959–3969. doi:10.1083/jcb.201704145
18. Hu L, Wang J, Zhou X, et al. Exosomes derived from human adipose mensenchymal stem cells accelerates cutaneous wound healing via optimizing the characteristics of fibroblasts. Sci Rep. 2016;6:32993. doi:10.1038/srep32993
19. Nelson WJ, Nusse R. Convergence of Wnt, beta-catenin, and cadherin pathways. Science. 2004;303:1483–1487. doi:10.1126/science.1094291
20. Yang HL, Tsai YC, Korivi M, Chang CT, Hseu YC. Lucidone promotes the cutaneous wound healing process via activation of the PI3K/AKT, Wnt/β-catenin and NF-κB signaling pathways. Biochim Biophys Acta Mol Cell Res. 2017;1864:151–168. doi:10.1016/j.bbamcr.2016.10.021
21. Ito M, Yang Z, Andl T, et al. Wnt-dependent de novo hair follicle regeneration in adult mouse skin after wounding. Nature. 2007;447:316–320. doi:10.1038/nature05766
22. Cho SB, Goo BL, Zheng Z, Yoo KH, Kang JS, Kim H. Therapeutic efficacy and safety of a 1927-nm fractionated thulium laser on pattern hair loss: an evaluator-blinded, split-scalp study. Lasers Med Sci. 2018;33:851–859. doi:10.1007/s10103-018-2437-5
23. Yang Z, Ma S, Cao R, et al. CD49fhigh defines a distinct skin mesenchymal stem cell population capable of hair follicle epithelial cell maintenance. J Invest Dermatol. 2020;140:544–555.e9. doi:10.1016/j.jid.2019.08.442
24. Maarof M, Chowdhury SR, Saim A, Bt Hj Idrus R, Lokanathan Y. Concentration dependent effect of human dermal fibroblast conditioned medium (DFCM) from three various origins on keratinocytes wound healing. Int J Mol Sci. 2020;21:2929. doi:10.3390/ijms21082929
25. Lago JC, Puzzi MB. The effect of aging in primary human dermal fibroblasts. PLoS One. 2019;14:e0219165. doi:10.1371/journal.pone.0219165
26. Temiz SA, Dursun R. Dermatolojide plateletten zengin plazma tedavisi. Selcuk Med J. 2020;36:274–281.
27. Li ZJ, Choi HI, Choi DK, et al. Autologous platelet-rich plasma: a potential therapeutic tool for promoting hair growth. Dermatol Surg. 2012;38:1040–1046. doi:10.1111/j.1524-4725.2012.02394.x
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Effects of Human Fibroblast-Derived Multi-Peptide Factors on the Proliferation and Migration of Nitrogen Plasma-Treated Human Dermal Fibroblasts
Lee SY, Kim DY, Suh SB, Suh JY, Cho SB
Clinical, Cosmetic and Investigational Dermatology 2022, 15:2465-2475
Published Date: 15 November 2022