Back to Journals » International Journal of Nanomedicine » Volume 18
Proteinase K/Retinoic Acid-Loaded Cationic Liposomes as Multifunctional Anti-Acne Therapy to Disorganize Biofilm and Regulate Keratinocyte Proliferation
Authors Tsai MJ ![]() , Lin CY, Trousil J, Sung CT, Lee MH, Fang JY
, Lin CY, Trousil J, Sung CT, Lee MH, Fang JY ![]() , Yang SC
, Yang SC
Received 12 April 2023
Accepted for publication 1 July 2023
Published 17 July 2023 Volume 2023:18 Pages 3879—3896
DOI https://doi.org/10.2147/IJN.S416966
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Yan Shen
Ming-Jun Tsai,1,2 Cheng-Yu Lin,3 Jiří Trousil,4 Calvin T Sung,5 Mei-Hua Lee,3 Jia-You Fang,3,6,7 Shih-Chun Yang8
1School of Pharmacy, Kaohsiung Medical University, Kaohsiung, Taiwan; 2Jhong Siao Urological Hospital, Kaohsiung, Taiwan; 3Pharmaceutics Laboratory, Graduate Institute of Natural Products, Chang Gung University, Kweishan, Taoyuan, Taiwan; 4Institute of Macromolecular Chemistry, Czech Academy of Sciences, Prague, Czech Republic; 5Department of Dermatology, University of California, Irvine, CA, USA; 6Research Center for Food and Cosmetic Safety and Research Center for Chinese Herbal Medicine, Chang Gung University of Science and Technology, Kweishan, Taoyuan, Taiwan; 7Department of Anesthesiology, Chang Gung Memorial Hospital, Taoyuan, Taiwan; 8Department of Microbiology, Soochow University, Taipei, Taiwan
Correspondence: Jia-You Fang, Pharmaceutics Laboratory, Graduate Institute of Natural Products, Chang Gung University, 259 Wen-Hwa 1st Road, Kweishan, Taoyuan, 333, Taiwan, Email [email protected] Shih-Chun Yang, Department of Microbiology, Soochow University, 70 Linhsi Road, Shihlin, Taipei, 111, Taiwan, Email [email protected]
Background: Simultaneous anti-Cutibacterium acnes and anti-inflammatory actions are highly beneficial in treating acne vulgaris. In this study, we present novel anti-acne nanovesicles based on liposomes loaded with proteinase K (PK), retinoic acid (RA), and soyaethyl morpholinium ethosulfate (SME) to achieve an effective and safe treatment.
Materials and Methods: This study examined in vitro planktonic and biofilm C. acnes elimination, as well as the keratinocyte proliferation suppression by liposomes. The multifunctional liposomes for treating C. acnes in mice were also evaluated.
Results: We acquired multifunctional liposomes with a size of 71 nm and zeta potential of 31 mV. The antimicrobial activity of SME was enhanced after liposomal encapsulation according to the reduction of minimum bactericidal concentration (MBC) by 6-fold. The multifunctional liposomes exhibited a synergistically inhibitory effect on biofilm C. acnes colonization compared with the liposomes containing PK or those containing SME individually. The adhesive bacterial colony in the microplate was lessened by 62% after multifunctional liposome intervention. All liposomal formulations tested here demonstrated no cytotoxicity against the normal keratinocytes but inhibited C. acnes-stimulated cell hyperproliferation. The in vitro scratch assay indicated that the liposomal RA—but not free RA—restrained keratinocyte migration. The animal study showed that free RA combined with SME and multifunctional nanovesicles had a similar effect on diminishing C. acnes colonies in the skin. On the other hand, liposomes exhibited superior performance in recovering the impaired skin barrier function than the free control. We also found that RA-loaded nanovesicles had greater skin tolerability than free RA.
Conclusion: The cationic liposomes containing dual PK and RA represented a potential treatment to arrest bacterial infection and associated inflammation in acne.
Keywords: liposome, acne, Cutibacterium acnes, proteinase K, retinoic acid, skin delivery
Introduction
Acne vulgaris is a chronic inflammatory and recurrent skin disorder involved in appendages. The major symptoms of acne include comedones, pustules, papules, and erythema. The sebaceous gland hyperactivity leads to increased sebum secretion, Cutibacterium acnes proliferation, and hyperkeratosis.1 Acne affects about 10% of the global population and >80% of young adolescents, ranking as the 8th most prevalent disease.2 Although not life-threatening, acne produces significant psychological effects of depression, anxiety, and insomnia.3 The pathogenesis of acne demonstrates that C. acnes colonization activates keratinocytes and sebocytes to induce inflammation, resulting in follicular occlusion and hyperkeratinization.4 C. acnes, a Gram-positive anaerobic bacterium, is reported to present >30% of the skin microbiota in acne patients.5 Acne vulgaris therapy focuses on the intervention of antibiotics and anti-inflammatory agents. However, the prolonged use of antibiotics causes skin toxicity and drug-resistant C. acnes development. The resistance of C. acnes to antibiotics such as clindamycin, erythromycin, and tetracycline has gradually emerged.6 The American Academy of Dermatology thus recommends the shortest possible times for the use of antibiotics on acne treatment.
C. acnes generates biofilm in the sebaceous glands to upgrade antibiotic resistance, comedones, and inflammation. This C. acnes biofilm incidence is greater in acne patients compared to healthy subjects.7 Biofilm is a community of bacteria encased in an extracellular polymeric substance (EPS) attached to a surface. Sessile C. acnes in the EPS matrix can withstand the immune response and become resistant to antibiotics that fail to deliver into the biomass.8 Thus, the novel acne treatment targeting C. acnes biofilm and the prevention of antibiotic resistance is urgent. The nanoparticles provide insight into improving the outcome of antibiofilm efficacy. Among these, liposomes have received significant attention in eradicating biofilm bacteria due to their capability to penetrate EPS and their versatility as antimicrobial agents.9 Liposomes are made of phospholipid bilayers surrounding an aqueous core. They are widely employed in industrial and clinical applications because of their possible scale-up, biocompatibility, high drug encapsulation, and controlled drug delivery. These lipid-based nanovesicles can facilely fuse with bacterial membranes and biofilm and are attributed to the affinity of phospholipid bilayers with biofilm.10 The extracellular proteins are vital for the early adhesion of C. acnes biofilm to the surface.11 Proteinase K (PK) is reported to be effective for digesting proteinaceous adhesins during the attachment of EPS.12 Therefore, we aimed to design antibiofilm liposomes by encapsulating PK into the nanovesicles. The cationic liposomes loaded with quaternary ammonium cationic surfactants developed as broad-spectrum antibacterial agents have been studied previously.13,14 Recently, soyaethyl morpholinium ethosulfate (SME) as a cationic surface active compound has broadened the perspective in nanomedicine due to its antibacterial activity and good biocompatibility.15,16 Hence, this study also utilized SME as the anti-C. acnes molecule in liposomes.
In addition to antimicrobial drugs, topical retinoic acid (RA) is the first-line treatment option for acne therapy. During acne therapy, RA inhibits keratinocyte proliferation, inflammation, and microcomedone production.17 The topical RA delivery poses challenges, including instability, poor absorption, and skin irritation. Most of the RA-treated acne patients meet cutaneous irritation, such as edema and peeling.18 The delivery systems based on nanocarriers can overcome the limitation of RA intervention.19 The available management methods for acne are still limited and remain unchanged for decades. Nevertheless, efficient and safe approaches with fewer complications can remarkably improve patient compliance.
This work aimed to formulate multifunctional liposomes containing PK, RA, and cationic surfactant as a combined acne therapy. We began this investigation by evaluating the effects of anti-C. acnes on the liposomes in both planktonic and biofilm forms. Then, we explored the proliferation and migration inhibition of versatile nanovesicles against keratinocytes. Liposomes have been used to deliver drugs into the skin by facilitating skin permeation and protecting drugs from degradation.20 Finally, we conducted the topical treatment of acne by liposomes in a mouse model to appraise the possible application.
Materials and Methods
Materials and Reagents
Soybean phosphatidylcholine (SPC, Phospholipon 80H) was purchased from American Lecithin Company. RA, cholesterol, and crystal violet were provided by Sigma-Aldrich. PK and cell counting kit-8 (CCK-8) were obtained from BioTools. SME was from Croda. Reinforced clostridial medium (RCM) was obtained from Merck. The PrestoBlue cell viability kit was provided by Invitrogen. The transwell plate for keratinocyte migration assay was purchased from Millipore. The IVISense Bacterial 250 fluorescent probe was bought from Perkin Elmer.
Preparation of Multifunctional Liposomes
The lipid-based nanovesicles were fabricated using the thin-film technique. SPC (230 mg), cholesterol (70 mg), and RA (5 mg) were dissolved in chloroform/methanol (2:1, 10 mL). This dispersion was deposited as a thin film in a round bottom flask using a rotary evaporator at 50°C under a vacuum. After 24 h, the thin film was hydrated by double-distilled water (ddH2O) under a probe-type sonication (VCX 600, Sonics and Materials) at 37 W for 30 min to produce the final nanovesicles (NV). Subsequently, PK (100 mg) and SME (30 mg) were added into ddH2O and chloroform/methanol, respectively, if necessary. The code name for the nanovesicles containing PK or SME alone was NV/PK and NV/SME, respectively. Meanwhile, the combined PK and SME in the nanovesicles had been given a code name as NV/PK/SME.
Physicochemical Features of the Liposomes
The average size, polydispersity index (PDI), and zeta potential of the prepared nanovesicles were determined using a laser scattering technique (Nano ZS90, Malvern). The nanodispersion was diluted 100-fold by ddH2O before the detection. The encapsulation percentage of RA in the liposomes was estimated by the ultracentrifugation method to separate the entrapped RA from the free RA. The nanovesicles were centrifuged at 48,000 xg and 4°C for 40 min. Further, the encapsulated drug in the precipitate and free drug in the supernatant were analyzed by high-performance liquid chromatography (HPLC, Hitachi Series-2). Then, the precipitate was dissolved by acetonitrile before analysis. The HPLC condition was described in a previous study.21
Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
C. acnes (ATCC6919) was cultivated in RCM at 37°C under anaerobic status for 3 days. The MIC of liposomes was described as the lowest SME concentration necessary to inhibit C. acnes growth by serial dilution. The bacteria were diluted in RCM to obtain the optical density at 600 nm (OD600) of 0.01. The C. acnes solution was exposed to several dilutions of liposomal dispersions and then incubated at 37°C for 48 h. We used a microplate reader (Epoch2, BioTek) to determine the MIC at 600 nm. The colony-forming unit (CFU) of C. acnes was counted after liposome treatment for 48 h. The highest dilution of liposomes, resulting in a 99.9% eradication of C. acnes number, was acknowledged as MBC value.
C. acnes Growth Curve
The growth inhibition of C. acnes by the lipid nanovesicles over 108 h was detected in 96-well microplates. The liposomes containing SPC at 0.72 μg/mL were inoculated with bacteria (OD600=0.01) at 37°C. The liposomes were diluted with RCM if necessary. The microplate reader was used to analyze each well’s absorbance (600 nm) and reveal the bacterial growth in real-time.
Disk Diffusion Assay
We placed C. acnes (OD600=0.7) on the agar plate. The prepared liposomes with different dilutions (1/32, 1/16, 1/8, 1/4, 1/2, and 1) and ddH2O were loaded onto the agar. After incubating for 48 h, the clear zone diameter without bacteria was measured.
C. acnes Viability Assay by SYTO 9
The viability of C. acnes after treatment of nanovesicles for 24 h was visualized using SYTO 9 green fluorescent nucleic acid staining. C. acnes was grown to OD600=0.01 and then treated with liposomes at 37°C for 24 h. The microbes were incubated with SYTO 9. After a 15-min reaction, the strain was observed under confocal microscopy (TCS SP2, Leica).
C. acnes Survival Outside and Inside the Biofilm
The bacteria were cultivated anaerobically using RCM at 37°C for 3 days. The culture was diluted to OD600=0.1 by RCM supplemented with 1% glucose. The bacterial suspension (1 mL) was pipetted into the 24-well microplate and incubated at 37°C for 3 days. The biofilm was developed on the well bottom after supernatant removal. The nanovesicles with different dilutions (1/32, 1/16, 1/8, and 1/4) were treated to the biofilm for 24 h. Then, the recovered C. acnes outside and inside the biofilm were loaded on the agar plate to measure CFU.
Biofilm Structure Observation by Scanning Electron Microscopy (SEM)
The SEM analysis was performed to visualize the effect of liposomes on biofilm formation. The C. acnes biofilm was treated with liposomes in the 24-well plate at 37°C for 24 h under an anaerobic condition. The well was washed twice with PBS and fixed in 3% glutaraldehyde and 2% paraformaldehyde for 2 h. After washing, the sample was dehydrated in a graded ethanol series (30%‒95%), then dried and coated with gold. The sample was visualized under SEM (SU-8220, Hitachi).
Biofilm Metabolic Activity
The C. acnes biofilm metabolism was determined using the PrestoBlue cell viability kit. PrestoBlue is a resazurin-based metabolism assay that detects metabolically active bacteria or cells. The C. acnes biofilm in the microtiter plate was treated with liposomes at 37°C for 10 h. After adding the PrestoBlue reagent (10%) to the biofilm, the plate was incubated for 1 h, and the fluorescence intensity was read at 570 nm (excitation) and 600 nm (emission) using a microplate reader.
Biofilm Adhesion Assay
C. acnes biofilm adhesion was assessed in a static condition in the microplate. The C. acnes (OD600=0.1) allowed adhering at 37°C for 1 h. Subsequently, the nanovesicles were treated to the biofilm for 10 h. After rinsing with PBS and staining with 0.1% crystal violet, the CFU of the adherent microbes was measured.22
Microfluidic Chamber Analysis
The C. acnes biofilm was developed in a flow cell chamber (Ibidi μ-Slide I0.2 Luer) for 2 days under an anaerobic condition at 37°C. The nanovesicles and SYTO 9 were injected into the biofilm-loaded flow chamber by a syringe pump with a flow rate of 5 μL/min. The liposome-treated biofilm attached to the chamber surface was visualized by confocal microscopy (TCS SP8X, Leica) from 45 to 225 min.
Keratinocyte Viability Assay
The human-derived keratinocytes (HaCaT, AddexBio) were maintained in DMEM with 10% heat-inactivated FBS and 100 U/mL antibiotic/antimycotic. The keratinocyte viability after the nanovesicle intervention was explored by employing a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. The HaCaT (1 x 105 cells/mL) was exposed with liposomes containing RA (0‒2 μg/mL) at 37°C for 24 h. Meanwhile, the MTT reagent (0.5 mg/mL) in the culture medium (100 μL) was incorporated into the cells and incubated for 3 h. The keratinocyte viability was detected using a microplate reader at 595 nm.
Keratinocyte Proliferation Assay
HaCaT cells (5 x 104 cells/mL) were plated in a 96-well plate for 24 h. Then, the heat-killed C. acnes (h-C. acnes, the multiplicity of infection (MOI)=75) and liposomes were added into the wells for 24 and 48 h, respectively. The h-C. acnes was prepared by incubation at 100°C for 30 min.23 The dead bacteria were stored at −20°C before use. The h-C. acnes-induced keratinocyte proliferation inhibited by RA-loaded nanovesicles was measured with CCK-8.
Keratinocyte Migration Assay
The keratinocyte migration inhibition by RA-loaded liposomes was studied by in vitro scratch assay. HaCaT cells (4 x 105 cells/mL) were cultured in culture-insert with 2 wells (Ibidi) at 37°C for 24 h. The insert was then transferred by sterilized tweezers, followed by cell washing with DMEM to remove the detached cells.24 Then, the h-C. acnes (MOI=75) and liposomes were added to the cell suspension. After a 20-h treatment, the image was taken with optical microscopy (Axiovert 200, Zeiss). The cell-free area was quantified using the ImageJ software.
Keratinocyte Transwell Invasion Assay
The transwell invasion is another platform that rates the keratinocyte migration inhibition by the RA-loaded nanovesicles. This experiment was conducted in a transwell plate with a pore size of 8 μm. In addition, the HaCaT suspension (5 x 105 cells/mL) was treated with h-C. acnes (MOI=75) or tumor necrosis factor-α (TNF-α, 20 ng/mL) for 24 h. The liposomes were also added into the cell suspension simultaneously. Then, the keratinocytes were diluted to 1×105 cells/mL and placed on the upper chamber of the Transwell. The culture medium was subsequently filled into the lower chamber. After being incubated for 24 h, the crystal violet (0.1%) was used to stain the migrated HaCaT in the lower well. The image was taken by optical microscopy. The cell number in the lower well was also quantified using a microplate reader at 595 nm.
Animals
The Balb/c mice and nude mice (6 weeks old) were purchased from the National Laboratory Animal Center (Taipei, Taiwan). The Institutional Animal Care and Use Committee of Chang Gung University approved all animal experiments. The experimental processes complied with the European Community legislation (Directive 86/609/EEC).
In vivo C. acnes Infection Treated by Topical Liposomes
The mouse was anesthetized to shave the dorsal skin. C. acnes suspension (1 x 107 CFU, 150 μL) was subcutaneously injected into the dorsal area. The liposomes (200 μL) were topically administered on the infected skin region every 24 h for 3 days, and the transepidermal water loss (TEWL) was recorded daily using Tewameter TM300 (Courage and Khazaka). After a 3-day treatment, the appearance of the skin was observed using a digital magnifier (Dino-Lite). The IVISense Bacterial 250 fluorescent probe was subcutaneously injected into the infection site to observe the C. acnes residence by the in vivo imaging system (IVIS, Lumina LT, PerkinElmer).
In vivo Skin Tolerance of Topical Liposomes
The liposomal or free RA (0.2 mL) was added on a gauze pad (1 x 1 cm2)—which was applied to the nude mice’s back—and then fixed with Tegaderm dressing. The pad was removed daily; new ones were applied for 5 consecutive days. The TEWL was monitored every day. At the end of the experiment (Day 5), the skin appearance was recorded using a digital magnifier. After excising the treated skin area, the skin was sectioned for hematoxylin and eosin (H&E) staining to examine the histological change after formulation administration.
Statistical Analysis
The Kruskal–Wallis test was employed to analyze the statistical difference in the data from different treatment groups. Meanwhile, a posthoc test to check the individual difference was carried out using Dunn’s test. A 0.001, 0.01, or 0.05 level of probability was considered statistically significant.
Results
Physicochemical Features of the Liposomes
All liposomes were prepared using the thin-film method. The size of blank liposomes (NV) was 82 nm, as determined by dynamic light scattering (Table 1). On the other hand, PK (NV/PK) incorporation increased the hydrodynamic diameter to 112 nm, whereas SME incorporation (NV/SME) reduced the vesicle size to 66±0.4 nm. The combined PK and SME in liposomes (NV/PK/SME) exhibited an average size of 71±0.6 nm. PDI was below 0.32 for all liposomes tested, verifying a narrow distribution and homogeneity in size. NV and NV/PK were negatively charged at −35±0.8 and −47±0.9 mV, respectively, because of SPC’s anionic feature. The nanovesicles became positively charged after adding cationic surfactant SME (NV/SME and NV/PK/SME). The RA loading did not significantly change the liposomal diameter and surface charge because of the very low percentage (0.05%) of RA in the liposomes. NV and NV/PK showed more than 80% encapsulation percentage for RA loading. SME incorporation (NV/SME and NV/PK/SME) reduced RA encapsulation to about 63%.
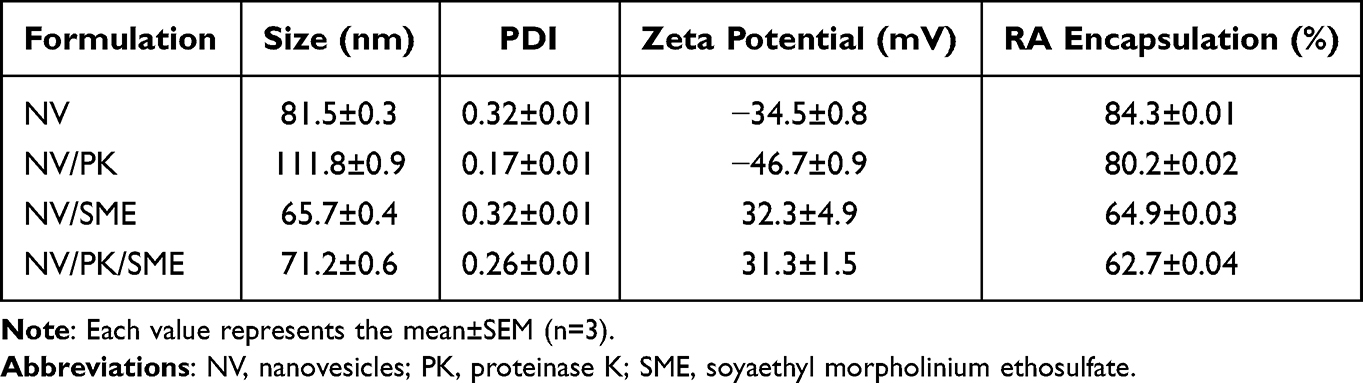 |
Table 1 The Characterization of Liposomes by Vesicle Size, Polydispersity Index (PDI), Zeta Potential, and RA Encapsulation Percentage |
Anti-C. acnes Activity of the Liposomes
We first measured MIC and MBC values of free and liposomal PK or SME. MIC is recognized as the lowest concentration to inhibit bacterial growth by serial dilution. Noteworthily, the formulations without SME (free PK, NV, and NV/PK) showed no effect on C. acnes growth suppression. The MIC levels of free SME were determined as 0.73‒1.47 μg/mL for planktonic C. acnes (Table 2). The SME inclusion in liposomes (NV/SME and NV/PK/SME) decreased MIC to 0.49 μg/mL. This 2‒4-fold reduction indicated that the nanovesicles offered dose attenuation in terms of SME concentration. MBC is the lowest bactericidal concentration that kills 99.9% of microbes. The MBC against C. acnes was 46.88‒93.75 μg/mL for free SME (Table 2). The SME loading in liposomes (NV/SME) led to greater bactericidal activity with MBC of 15.63‒31.25 μg/mL. Meanwhile, the PK inclusion in SME-loaded nanovesicles (NV/PK/SME) further enhanced the bactericidal effect (MBC=7.81‒15.63 μg/mL). The effect of the nanovesicles on the growth curve of C. acnes was assessed at an SME concentration of 15.63 μg/mL (MBC of the NV/SME). The time-kill curves for liposomal SME (NV/SME and NV/PK/SME) indicated that MBC was sufficient to inhibit almost all C. acnes (Figure 1A). The bacterial population incubated with the nanovesicles without SME (NV and NV/PK) demonstrated no C. acnes growth inhibition.
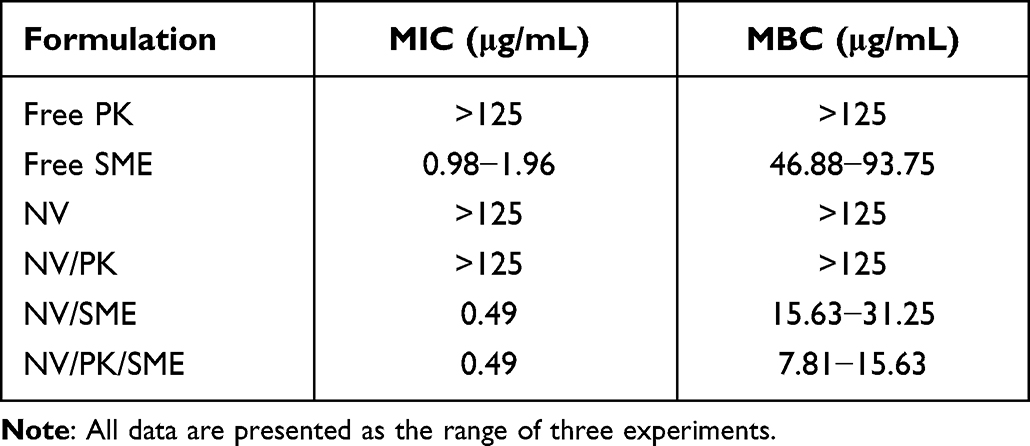 |
Table 2 The Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) of Free PK, Free SME, and Liposomes Based on SME Concentration (μg/Ml) |
 |
Figure 1 Determination of anti-C. acnes activity in planktonic or biofilm forms by liposomes: (A) the time-killing curves of liposomes during 108 h; (B) the diameter of inhibition zone (mm) after liposome treatment in the agar plate; (C) confocal image of live C. acnes stained with SYTO 9 after liposome treatment; (D) C. acnes CFU outside and inside the biofilm after liposome treatment; and (E) the biofilm structure observed by SEM after liposome treatment. All data are presented as the mean of three experiments±S.E.M. ***p < 0.001, **p < 0.01, or *p < 0.05 as compared to the untreated control group. |
The prepared liposomes were diluted to different folds by ddH2O to measure the C. acnes inhibition zone in the disk diffusion test. There is no clear inhibition zone in the agar for the liposomes without SME (NV and NV/PK) (Figure 1B). NV/SME and NV/PK/SME exhibited a large inhibition zone on the agar media in a concentration-dependent manner. The inhibition zone against C. acnes ranged between 8.3 and 14.5 mm for NV/SME. Adding PK in the liposomes (NV/PK/SME) further enlarged the inhibition zone to 8.8‒17.3 mm. The C. acnes viability was visualized using SYTO 9 as a fluorescent dye. In the confocal image, the blank liposomes (NV) could not diminish live bacteria compared to the untreated control (Figure 1C). An inferior green intensity due to SYTO 9 was detected after NV/PK and NV/SME treatments, suggesting the decrease of live C. acnes. Inserting the combined PK and SME in liposomes (NV/PK/SME) further decreased the green fluorescence, confirming the synergistic effect on C. acnes killing.
Furthermore, we examined the capability of liposomes to eradicate the C. acnes biofilm. The C. acnes CFU outside and inside the biofilm was counted with a log transformation. NV and NV/PK were devoid of the activity to lessen the viable bacteria outside the biofilm (the left panel of Figure 1D). The bacteria outside the biofilm showed a significant decrease in CFU upon treatment with SME-loaded liposomes (NV/SME and NV/PK/SME). The combination of PK and SME demonstrated a 2-log reduction of the C. acnes population outside the biofilm. The blank nanovesicles (NV) lacked the capacity to eradicate C. acnes inside the biofilm (the right panel of Figure 1D). A dose-dependent inhibition of biofilm C. acnes was found for NV/PK and NV/SME. The combined treatment using PK and SME appeared to be synergistic when C. acnes CFU was decreased inside the biofilm. SEM observation allows the understanding of biofilm structure change after liposome interaction. In the untreated control, C. acnes formed a dense biofilm composed of microbe aggregate with a pleomorphic rod shape (Figure 1E). The three-dimensional and well-structured network was still observed after the treatment of blank liposomes (NV). Treatment with NV/PK resulted in disaggregated and elongated bacteria with wrinkled and punctured membranes. NV/SME-treated biofilm was associated with a disintegrated cluster, along with the destabilization of the bacterial membrane. NV/PK/SME abolished the biofilm matrix together with the elongated and ruptured C. acnes appearance.
Biofilm Metabolic Activity and Adhesion
An initial surface contact followed by irreversible adhesion is required for biofilm formation. Here we examined whether the liposomes could prevent the adherence of biofilm in the microtiter plate. The biofilm adhesion percentage estimated by PrestoBlue determined the adhesion level by the metabolic activity of C. acnes. The biofilm treated with NV revealed an adhesion percentage similar to the untreated control (Figure 2A). We found that when biofilm was incubated in the presence of NV/PK, the PK-loaded liposomes suppressed the adhesion to 50±7.2%. Compared to the control, the adhesion treated with NV/SME and NV/PK/SME displayed a metabolic activity of 60±3.2% and 52±5.1%, respectively. We also determined the C. acnes CFU adhered in the microplate after a 10-h liposome treatment, while the blank liposomes (NV) were devoid of C. acnes adherence (Figure 2B). PK incorporation (NV/PK) significantly decreased the adherence to 48±8.1%. The bacterial adhesion was inhibited by SME-loaded liposomes (NV/SME and NV/PK/SME) to a similar level to NV/PK. The flow cell chamber assay was established to monitor the effect of liposomes on surface attachment and biofilm formation. The biofilm patch was formed by the preferential adhesion of C. acnes to the microfluidic channel. The continuous flow of ddH2O (control) did not eliminate the biofilm attached to the channel surface (Figure 2C). A similar phenomenon was observed in the confocal image of NV-treated biofilm. A partial dispersal of the adhered biofilm occurred after the continuous injection of NV/PK and NV/SME into the flow cell. Biofilm observation by confocal microscopy displayed a significant matrix degradation and structural integrity loss after combined treatment with PK and SME (NV/PK/SME). This phenomenon suggested a synergistic effect on biofilm destruction.
 |
Figure 2 Determination of anti-adhesion activity of biofilm C. acnes by liposomes: (A) the adhesion percentage of the biofilm detected by PrestoBlue after liposome treatment; (B) the adhesive C. acnes CFU percentage of the biofilm detected after liposome treatment; and (C) the biofilm mass attached on the surface of microfluidic chamber observed by confocal microscopy. All data are presented as the mean of three experiments±S.E.M. ***p < 0.001 or **p < 0.01 as compared to the untreated control group. |
Keratinocyte Proliferation Regulation by the Liposomes
The multifunctional liposomes were loaded with RA to treat acne-associated keratinocyte proliferation and migration. The biocompatibility of the nanovesicles on the human keratinocyte cell line was tested using an MTT assay. Five different concentrations (0.1‒2 μg/mL) of RA loaded in liposomes were incubated with keratinocytes for 24 h. The result showed that the liposomes were not cytotoxic at concentrations up to 2 μg/mL, as the cell viability was kept close to the untreated control (Figure 3A). The keratinocyte viability was found to be >80% for all RA liposomes tested. This finding suggests that the liposomes can be used for inhibiting keratinocyte activation at ≤2 μg/mL without causing damage to mammalian cells. The C. acnes-induced proliferation and migration of keratinocytes are crucial factors in acne development. We conducted a CCK-8 assay to investigate the effect of RA-loaded nanovesicles on keratinocyte cell proliferation regulation. HaCaT cell proliferation was increased by 1.3-fold after a 24-h stimulation with h-C. acnes (the left panel of Figure 3B). Meanwhile, free RA at 2 μg/mL did not reduce the h-C. acnes-induced proliferation. Treatment with different types of RA-loaded liposomes significantly suppressed proliferation. The liposomes containing SME (NV/SME and NV/PK/SME) showed greater proliferation repression than those without SME (NV and NV/PK). The free RA exhibited the ability to suppress proliferation as the incubation time was prolonged to 48 h (the right panel of Figure 3B). The inhibition level of the liposomes without SME (NV and NV/PK) was comparable to that of free RA. Similar to the 24-h treatment, the inhibition was pronounced with 48 h of incubation by SME-containing nanovesicles (NV/SME and NV/PK/SME).
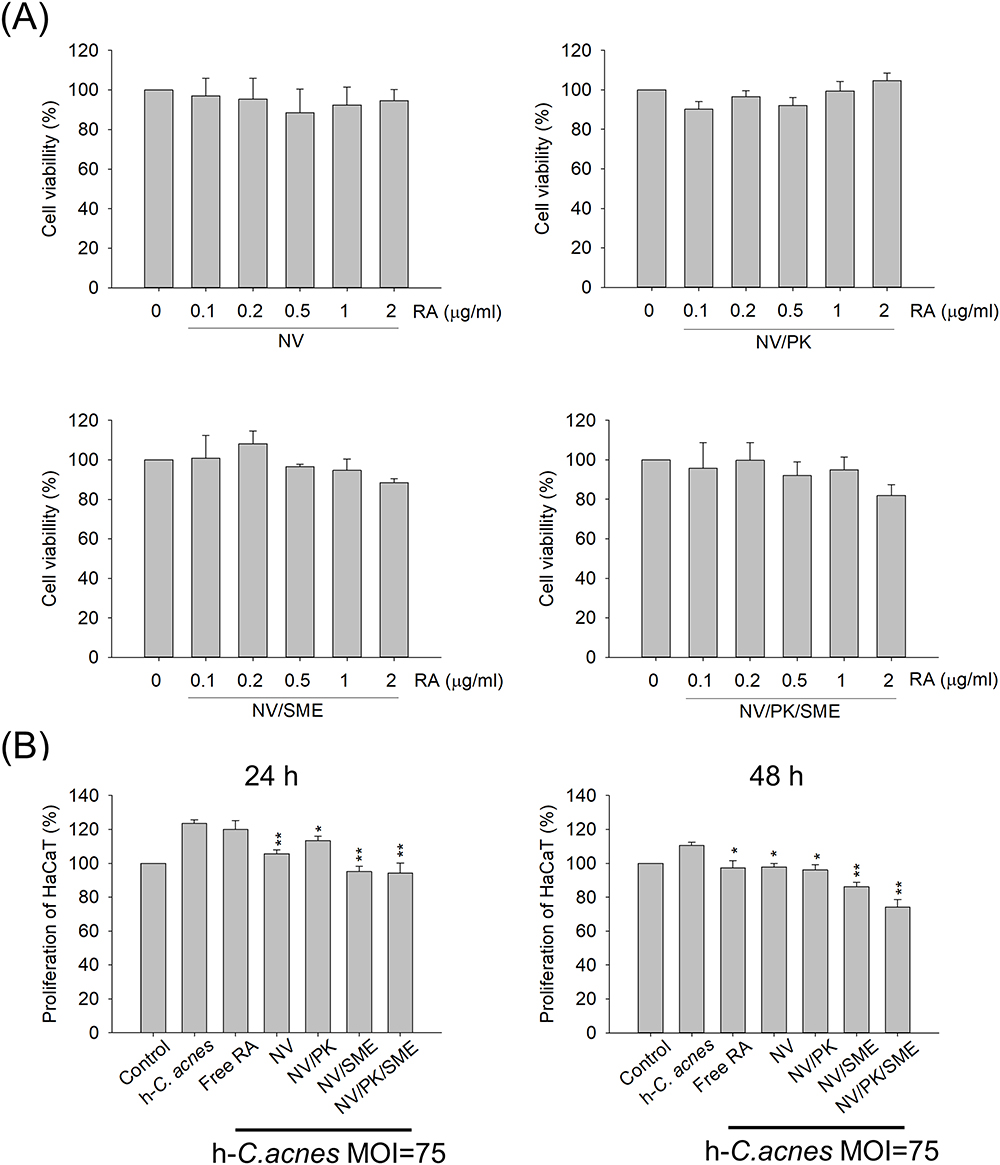 |
Figure 3 The effect of liposomes on the cytotoxicity and proliferation regulation of human keratinocytes: (A) the cell viability of keratinocytes determined by MTT assay; and (B) the proliferation of h-C. acnes-stimulated keratinocytes after liposome treatment. All data are presented as the mean of three experiments±S.E.M. **p < 0.01 or *p < 0.05 as compared to the h-C. acnes treatment group without liposome intervention. |
The cell-based scratch analysis was utilized to examine cell proliferation and migration. This evaluation can unveil whether a specific compound or liposomal formulation slows down the rate of proliferation or migration. The HaCaT cells that underwent no treatment (control) displayed wound closure after a 20-h incubation (Figure 4A). A further wound closure due to the proliferation and migration of keratinocytes was visualized 20 h later using h-C. acnes. No significant suppression in scratch closure was detected in the free RA (2 μg/mL) treatment. Migratory keratinocytes were inhibited by all nanoformulations tested. We calculated the remaining cell-free area in the culture insert (Figure 4B). The areas for the four liposomal formulations were approximately 90%‒98%. The transwell invasion test is another evaluation platform that detects cell migration. Keratinocytes were subjected to the upper layer of the transwell; the number of keratinocytes migrating to the bottom layer was appraised. The crystal violet colorimetric analysis indicated a significant cell migration after a 24-h incubation (Figure 4C). This migration was exceptionally elevated with h-C. acnes stimulation by 1.5-fold compared with the basal control. A substantial suppression was noticed after free RA and equivalent NV/PK/SME liposomes were treated. The violet crystal quantification result showed that free and liposomal RA reduced the migration by 49% and 63%, respectively (Figure 4D). The migration of HaCaT cells was increased by activating another stimulator TNF-α (Figure 4E). Free RA and NV/PK/SME provided considerably significant inhibition on the migration. The migration was decreased by 57% and 60% with the treatment of free and liposomal RA, respectively (Figure 4F).
 |
Figure 4 The effect of liposomes on the proliferation and migration regulation of human keratinocytes: (A) the migration of h-C. acnes-stimulated keratinocytes after liposome treatment as determined by in vitro scratch assay; (B) the measurement of remaining cell free area based on the in vitro scratch assay; and (C) the migration of h-C. acnes-stimulated keratinocytes after liposome treatment as determined by in vitro transwell assay; (D) the measurement of crystal violet-stained h-C. acnes-stimulated keratinocytes in the bottom layer based on the in vitro transwell assay; (E) the migration of TNF-α-stimulated keratinocytes after liposome treatment as determined by in vitro transwell assay; and (F) the measurement of crystal violet-stained TNF-α-stimulated keratinocytes in the bottom layer based on the in vitro transwell assay. All data are presented as the mean of three experiments±S.E.M. ***p < 0.001 or **p < 0.01 as compared to the stimulator treatment group without liposome intervention. |
In vivo C. acnes Infection Treated by Topical Liposomes
The in vivo anti-C. acnes effect of the multifunctional nanovesicles (NV/PK/SME) loaded with RA was evaluated in mice. Free RA combined with SME was also tested at a dose equivalent to that in liposomes. The animals were subcutaneously infected with C. acnes, followed by topical application of free drugs or liposomes. In the microscopic appearance of the mouse skin, the subcutaneous invasion of C. acnes resulted in the skin abscess with erythema signs (the arrow in Figure 5A). Both free SME/RA and liposomes were protected against the cutaneous damage caused by infection. The TEWL was significantly increased by C. acnes injection (Figure 5B), suggesting a skin barrier dysfunction after the infection. The drug formulations partially recovered the increased TEWL, with liposomes showing superior recovery than free SME/RA. The C. acnes colonization in the infected skin area was 5.5×105 CFU/cm2 after bacterial invasion for 3 days (Figure 5C). A 2-log reduction of CFU was found after topical delivery of the free drug group. The number of CFU in the skin treated with the multifunctional liposomes was significantly lower than that of the control with a 1.5-log reduction. The XenoLight Bacterial kit was injected into the C. acnes-infected region to visualize the bacterial aggregate by IVIS. There were some bacteria-associated signals in the healthy animals (Figure 5D and E), which could be derived from the resident bacterial flora other than C. acnes. The bacterial administration increased the fluorescence signal in the skin, suggesting a successful C. acnes colonization. The topical delivery of free SME/RA and nanovesicles decreased C. acnes accumulation with a comparable level. Both the fluorescence zone and intensity were lowered by drug treatment.
 |
Figure 5 In vivo anti-acne effect of liposomes in the C. acnes-infected mouse model: (A) the appearance of mouse skin surface; (B) the TEWL level of the mouse skin; (C) C. acnes CFU in skin; (D) the average radiance intensity of C. acnes burden (labeled with XenoLight Bacterial Detection Probe) in skin of the mouse quantified by in vivo imaging system; and (E) the C. acnes burden in skin of the mouse observed by in vivo imaging system. All data are presented as the mean of six experiments±S.E.M. ***p < 0.001 or **p < 0.01 as compared to the C. acnes infection group without RA treatment. |
In vivo Skin Tolerance of Topical Liposomes
The possible skin irritation elicited by topical RA was analyzed in vivo. The photograph of the mouse dorsal skin treated with ddH2O for 5 days (control) exhibited an unharmed appearance with a smooth surface (Figure 6A). Some skin lesions were detectable in the free RA-treated group that revealed erythema, crust, and edema in the superficial layer. We observed fewer skin lesions in the liposome-treated group than in the free RA. The ddH2O treatment (control) slightly increased TEWL on Day 1 and Day 2; then, the value recovered to baseline control after 3 days (Figure 6B). Free RA greatly disrupted the skin barrier function as evidenced by a large increase of TEWL. This increase could be partially relieved after encapsulation of RA into nanovesicles. The ddH2O application exhibited no histopathological change based on H&E staining (Figure 6C). Free RA induced scaling, epidermal hyperplasia, and immune cell infiltration after a consecutive 5-day challenge. In the case of topical liposome application, the skin section manifested persistence against epidermal thickening and inflammatory cell diffusion. The signs of these damages showed less severity than those of free RA treatments.
 |
Figure 6 The in vivo skin tolerance of the liposomes in the healthy mouse: (A) the appearance of mouse skin surface; (B) the TEWL level of the mouse skin; and (C) the H&E-stained histology of mouse skin treated by liposomes. All data are presented as the mean of five experiments±S.E.M. |
Discussion
The dual therapy of antibacterial and anti-inflammatory agents is commonly used for acne treatment. However, the currently available treatments are often unsatisfactory for complete response because of antibiotic resistance and skin irritation.25 The failure of antibiotic acne treatment can be associated with biofilm-mediated resistance. While most antimicrobial agents are designed to eradicate planktonic microbes, they are usually devoid of treating biofilm infection.10 One of the attempts to overcome the resistance threat is designing antibacterial nanoparticles. In this study, we developed cationic liposomes incorporated with PK as antibiofilm formulations to efficiently eliminate C. acnes. The combined PK and SME revealed a synergistic effect in diminishing the C. acnes burden in the biofilm. This combination also synergistically disintegrated the biofilm architecture. The liposomes were also loaded with RA to prevent keratinocyte proliferation and migration induced by C. acnes. Topical RA treatment is ideal for providing increased local drug bioavailability. Skin irritation during topical RA therapy can lower compliance with long-term treatment.26 An efficient topical anti-acne therapy should deliver the drugs to the target skin site with minimal irritation. Notably, our liposomes effectively lessened C. acnes accumulation and skin toxicity in the mouse model.
SME was the main active in the liposomes for eradicating C. acnes. The cationic surfactants can be utilized as capping agents to change the physicochemical characteristics of the nanoparticles. The nanovesicle size was decreased by SME incorporation because of the emulsification effect of the surfactant to reduce surface-free energy. All liposomal formulations developed in this study exhibited a size of <120 nm, confirming the nano-sized range of the prepared products. The surface charge is an index used to probe the stability of nanoparticles. It is generally regarded that the nanovesicles with zeta potential above +30 mV or below −30 mV are highly stable due to the electrostatic repulsion.27 Meanwhile, the zeta potential data implied high stability of the as-prepared liposomes. Due to the extremely high lipophilicity, RA was expected to be accommodated in the phospholipid bilayers of liposomes. The RA encapsulation could reach >80% in liposomes; however, SME incorporation reduced the encapsulation percentage to 60%. Because free RA could be removed by ultracentrifugation, the complete encapsulated form of RA in liposomes was used throughout all experiments. The liposomes were diluted by ddH2O for drug dose calibration to assure the equivalent RA concentration in different formulations for testing.
Our data demonstrated that the cationic liposomes facilely inhibited C. acnes growth in the planktonic form. The bacterial membrane behaves as a barrier to retain cellular homeostasis. Thus, the damage in the bacterial membrane can disrupt the physiological function, leading to cell death. It has been reported that liposomes are fusogenic to bacterial membranes due to the resemblance between the phospholipid bilayers and bacterial plasma membrane.28 This fusion effect promotes the bactericidal activity of antibiotics against drug-resistant pathogens.9,29 A similar result was found in our study, where the liposomal SME was superior to free SME in eradicating planktonic C. acnes. The anti-C. acnes activity of SME was attributed to the electrostatic interaction between the protonated amine moieties of SME and anionic teichoic acid in the bacterial membrane,16 resulting in membrane disturbance and intracellular protein leakage. Teichoic acid in the Gram-positive bacteria membrane is a common binding site of the nanovesicles.30 Gram-positive bacteria tend to possess looser cell walls than Gram-negative bacteria. The nanovesicles possibly penetrated across the loose membrane, permitting the entrance into the cytoplasm and consequently disorganizing the bacterial structure. The nanoparticle size affects the interaction with the microbial membrane.31 The liposomes with SME showed a very small size of about 70 nm that could invade the internal components of bacteria.
The transition of planktonic bacteria to biofilm involves adhesion to the surface. The biofilm network of C. acnes is composed of proteins, extracellular DNA, and carbohydrates.32 These components not only develop the mature biofilm but also play a vital role in early surface attachment and colonization. C. acnes biofilm acts as an adhesive in the sebum, generating microcomedone.11 We found that the biofilm was disorganized and detached from the microplate well after the treatment of PK- and/or SME-loaded nanovesicles based on the PrestoBlue and CFU assays. Once delivered into the biofilm, the effect of antimicrobial agents depends on bacterial membrane damage or metabolic process interference.33 The PrestoBlue assay indicated the reduction of metabolic activity in the adhered C. acnes biofilm treated by PK/SME-loaded liposomes. This phenomenon may have resulted from the death of biofilm C. acnes or merely the metabolism disturbance in live bacteria without death. The multifunctional liposomes significantly reduced microbial CFU in the adhered biofilm, suggesting that C. acnes death—but not metabolism disturbance—was the predominant mechanism of liposome-mediated detachment. Biofilm-associated protein (Bap) is a multi-domain and cell surface-anchored protein that plays a crucial capacity in the early adhesion of biofilm development. PK is reported to be effective for Bap degradation.34 It also enzymatically degrades proteinaceous adhesin to hamper biofilm development.35 In addition to PK, we found that SME in liposomes retarded the biofilm adhesion, possibly due to the C. acnes killing by the cationic surfactant, lessening the bacterial colonization to form biofilm.
Although some antibiotics can eradicate C. acnes, the clinical efficacy is usually limited because of the biofilm-associated recolonization within a few weeks.8 Once the biofilm is established, the thick EPS prevents host defense and antibiotic penetration, resulting in antibiotic resistance. It appeared that not only the adhesion inhibition but the multifunctional liposomes were also successful in abolishing pre-existing biofilm. The multifunctional liposomes disrupted the biofilm structure and killed the biofilm C. acnes. There is a complex channel network inside the C. acnes biofilm network.9,16,36 The nanoparticles with positive charge can interact with negatively charged EPS for facile biofilm entrance.37 PK was expected to impair the biofilm structure after liposome delivery into the biofilm. PK is proven to cleave the C-terminal peptide bonds of amino acids for protein digestion.38 Biofilm is produced by viable microbes. Thus, enzymatic treatment should be combined with antimicrobial agents for successful biofilm dispersal. SME in liposomes played a major role as anti-C. acne agents to eliminate biofilm. The biofilm CFU result verified the synergistic antibiofilm activity of the combined PK and SME.
Besides the enzymatic degradation of biofilm, the role of PK in the liposomes should be probed. We observed that NV/PK could decrease live C. acnes stained by SYTO 9. However, NV/PK demonstrated no bactericidal effect in the MIC/MBC and growth curve. SYTO 9 is a green dye binding to bacterial nucleic acids to determine cell viability. It is reported that serine proteases such as PK degrade proteins of the bacterial membrane to cause the disturbance of cell wall maintenance and membrane construction.39 Our SEM image illustrated a damaged bacterial envelope in the C. acnes biofilm after a challenge by NV/PK. Proteinases may also degrade the synthase of DNA and RNA polymers.40 This effect contributed to the less SYTO 9 fluorescence in C. acnes after NV/PK intervention. It was speculated that this degradation could mildly decrease nucleic acids but maintain bacterial viability, as detected in MIC/MBC and the time-killing curve. Nevertheless, we found a stronger bactericidal effect of cationic liposomes (NV/SME) after PK incorporation (NV/PK/SME) based on MBC. This finding indicated that PK assisted in the anti-C. acnes activity of SME.
Keratinocytes account for 95% of all cells in the cutaneous epidermis. They play a central role in the pathogenesis of acne. C. acnes activates Toll-like receptor (TLR)-2 and −4 in keratinocytes to evoke an inflammatory response and oxidative stress, consequently promoting keratinocyte proliferation and migration.41 C. acnes can directly activate keratinocytes to prompt local inflammation by releasing cytokines interleukin (IL)-1β, IL-6, and TNF-α.42 This inflammation precedes microcomedone generation to serve as a trigger for follicular hyperkeratinization. RA inhibits keratinocyte inflammation through the downregulation of TLR-2 expression, controlling the signs of acne vulgaris.43 The regulation of keratinocyte proliferation by RA is mediated by the binding to specific nuclear receptors, eliciting the transcription of target genes. Our findings showed that free RA at 2 μg/mL was defective in inhibiting keratinocyte proliferation in the CCK-8 assay. RA incorporation into nanovesicles was favorable for restraining the proliferation and migration of HaCaT cells, with the SME-loaded liposomes (NV/SME and NV/PK/SME) proving the highest inhibition. This incorporation into liposomes protected RA from degradation. The antiproliferative effect of the liposomes could possibly be related to the cellular uptake, enabling prompt internalization in the cytosol and RA release to interact with more targets. The positive charge of the nanoparticles facilitates cellular ingestion due to the enhanced affinity with negatively charged cytomembrane.44 The PK presence in nanovesicles generally signified no effect on keratinocyte proliferation and migration.
RA incorporation into nanovesicles such as liposomes is proven to ameliorate the absorption into the skin.45 Our in vivo study found that the topical application of multifunctional liposomes (NV/PK/SME) led to the keratinization normalization and disintegrated barrier recovery in the C. acnes-infected mice. The bacterial invasion in the skin was also reduced after topical liposome application. The liposomes can augment the drug contact time on the skin surface, after which the nanovesicles fuse with the intercellular lipids of superficial skin. This effect causes a looser and permeable skin barrier for improved drug delivery.46 The intercellular space between the keratinocytes in the stratum corneum is reported to be 50‒70 nm.47 Thus, multifunctional nanovesicles of approximately the same size could pass through this space and into the deeper skin strata for treating acne. The positive charge of the multifunctional liposomes was also preferable in penetrating across the negatively charged lipid bilayers of stratum corneum.48 On the other hand, the positive charge is beneficial for liposome transport into the appendages since the hair is also negatively charged. Another advantage of topical liposome delivery is its occlusive feature to form a dense film on the skin surface. This film retards TEWL and enriches skin hydration to enhance drug permeation through the skin.49 Our data confirmed the reduced TEWL after liposome administration. C. acnes is largely found in the lipid-rich area of the skin, reaching a density of about 106 CFU/cm2.50 Notably, the C. acnes colonization after subcutaneous bacterial injection in the present study approached this value. C. acnes impairs epidermal barrier property by stimulating keratinocyte proliferation.51 It was inferred that topical liposomes could facilely penetrate the skin to inhibit C. acnes infection, arresting inflammation and keratinocyte hyperproliferation to effectively treat the infected wound.
The first-line therapy for acne is topical RA medication, which usually makes retinoid dermatitis of irritation, erythema, and eschar.19 Its combination with antibiotics or benzoyl peroxide causes more adverse effects, inflicting a higher incidence of withdrawal than monotherapy.52 An effective topical acne therapy should deliver RA to the target site and prevent skin irritation with good tolerability. Topical RA exfoliates the superficial skin layer by loosening intercellular connections in the stratum corneum, leading to increased TEWL.43 Although RA effectively inhibits keratinocyte hyperproliferation in acne skin, this drug enhances skin proliferation in healthy skin.53 RA can regulate keratinocyte proliferation and differentiation based on different conditions. The in vitro MTT assay showed that the liposomes were not cytotoxic to the HaCaT cell line. The in vivo tolerance displayed that the liposomes prevented RA-related skin irritation by reducing epidermal hyperplasia and immune cell infiltration. This irritation prevention could be ascribed to the RA encapsulation in the nanovesicles to avoid direct contact of the acidic moiety with the skin. A previous study26 has also proven that the cream form with RA molecules encapsulated inside the oil droplets is better tolerated than the gel form.
The antibacterial effect of SME achieved the anti-acne potential of the multifunctional nanovesicles collaborated with the inflammation inhibition of RA. Moreover, the PK inclusion in the liposomes reduced the adhesion and development of C. acnes biofilm, ameliorating the susceptibility to antibacterial agents and the host immune system. C. acnes is also responsible for many infectious diseases, including folliculitis, periodontitis, and rhinosinusitis.25 The developed liposomes are expected to have a wide application against different types of C. acnes invasion. However, further investigations are still needed to explore the possible application. Currently, available management methods for acne, such as RA, benzoyl peroxide, and azelaic acid, have led to skin irritation. If patients’ skin demonstrates intolerance, it is recommended to terminate the topical therapy. The liposomes developed in the study were promising in improving tolerability and patient compliance because of their therapeutic efficacy and safety.
Conclusion
This study designed multifunctional nanovesicles to suppress C. acnes infection and the associated keratinocyte proliferation. An important finding of this work is that the liposomes had a strong anti-C. acnes potential against planktonic and biofilm forms. The fusion and electrostatic interaction between the cationic liposomes and negatively charged bacterial membrane and biofilm were the major mechanisms revealing greater antimicrobial activity than the free control. In addition, PK incorporation in the nanovesicles increased the susceptibility of bacterial effect and biofilm eradication. Further, the liposomes efficiently arrested the proliferation and migration of the activated keratinocytes. Liposomal RA improved skin tolerability compared to the free form. Moreover, the experimental data of the liposomes suggests that the versatile liposomes can potentially be used as an anti-acne agent against C. acnes.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The authors are grateful for the financial support from National Science and Technology Council, Taiwan (MOST-110-2320-B-182-011-MY3) and Chang Gung Memorial Hospital (CMRPD1M0541).
Disclosure
The authors have declared no competing interests.
References
1. Goodarzi A, Mozafarpoor S, Bodaghabadi M, Mohamadi M. The potential of probiotics for treating acne vulgaris: a review of literature on acne and microbiota. Dermatol Ther. 2020;33(3):e13279. doi:10.1111/dth.13279
2. McLaughlin J, Watterson S, Layton AM, Bjourson AJ, Barnard E, McDowell A. Propionibacterium acnes and acne vulgaris: new insights from the integration of population genetic, multi-omic, biochemical and host-microbe studies. Microorganisms. 2019;7:128.
3. Layton AM, Thiboutot D, Tan J. Reviewing the global burden of acne: how could we improve care to reduce the burden? Br J Dermatol. 2021;184:
4. Isard O, Knol AC, Ariès MF, et al. Propionibacterium acnes activates the IGF-1/IGF-1R system in the epidermis and induces keratinocyte proliferation. J Invest Dermatol. 2011;131:
5. Park SY, Kim HS, Lee SH, Kim S. Characterization and analysis of the skin microbiota in acne: impact of systemic antibiotics. J Clin Med. 2020;9:168.
6. Dréno B, Pécastaings S, Corvec S, Veraldi S, Khammari A, Roques C. Cutibacterium acnes (Propionibacterium acnes) and acne vulgaris: a brief look at the latest updates. J Eur Acad Dermatol Venereol. 2018;32(Suppl. 2):
7. Jahns AC, Lundskog B, Ganceviciene R, et al. An increased incidence of Propionibacterium acnes biofilms in acne vulgaris: a case-control study. Br J Dermatol. 2012;167:
8. Cavallo I, Sivori F, Truglio M, et al. Skin dysbiosis and Cutibacterium acnes biofilm in inflammatory acne lesions of adolescents. Sci Rep. 2022;12:21104.
9. Wang Y. Liposome as a delivery system for the treatment of biofilm-mediated infections. J Appl Microbiol. 2021;131:
10. Lin Y-K, Yang S-C, Hsu C-Y, Sung J-T, Fang J-Y. The antibiofilm nanosystems for improved infection inhibition of microbes in skin. Molecules. 2021;26(21):6392. doi:10.3390/molecules26216392
11. Kuehnast T, Cakar F, Weinhäupl T, et al. Comparative analyses of biofilm formation among different Cutibacterium acnes isolates. Int J Med Microbiol. 2018;308(8):
12. Eladawy M, El-Mowafy M, El-Sokkary MMA, Barwa R. Effects of lysozyme, proteinase K, and cephalosporins on biofilm formation by clinical isolates of Pseudomonas aeruginosa. Interdiscip Perspect Infect Dis. 2020;2020:6156720.
13. Campanhã MT, Mamizuka EM, Carmona-Ribeiro AM. Interactions between cationic liposomes and bacteria: the physical-chemistry of the bactericidal action. J Lipid Res. 1999;40:
14. Montefusco-Pereira CV, Formicola B, Goes A, et al. Coupling quaternary ammonium surfactants to the surface of liposomes improves both antibacterial efficacy and host cell biocompatibility. Eur J Pharm Biopharm. 2020;149:
15. Alalaiwe A, Wang PW, Lu PL, Chen YP, Fang JY, Yang SC. Synergistic anti-MRSA activity of cationic nanostructured lipid carriers in combination with oxacillin for cutaneous application. Front Microbiol. 2018;9:1493.
16. Fang JY, Chou WL, Lin CF, Sung CT, Alalaiwe A, Yang SC. Facile biofilm penetration of cationic liposomes loaded with DNase I/proteinase K to eradicate Cutibacterium acnes biofilm for treating cutaneous and catheter infections. Int J Nanomed. 2021;16:8121–8138.
17. Leyden J, Stein-Gold L, Weiss J. Why topical retinoids are mainstay of therapy for acne. Dermatol Ther. 2017;7:293–304.
18. Raminelli ACP, Romero V, Semreen MH, Leonardi GR. Nanotechnological advances for cutaneous release of tretinoin: an approach to minimize side effects and improve therapeutic efficacy. Curr Med Chem. 2018;25:3703–3718.
19. Milosheska D, Roškar R. Use of retinoids in topical antiaging treatments: a focused review of clinical evidence for conventional and nanoformulations. Adv Ther. 2022;39:5351–5375.
20. Filipczak N, Pan J, Yalamarty SSK, Torchilin VP. Recent advancements in liposome technology. Adv Drug Deliv Rev. 2020;156:4–22.
21. Lee WR, Huang TH, Hu S, et al. Laser-assisted nanoparticle delivery to promote skin absorption and penetration depth of retinoic acid with the AIM for treating photoaging. Int J Pharm. 2022;627:122162.
22. Polyudova TV, Eroshenko DV, Korobov VP. Plasma, serum, albumin, and divalent metal ions inhibit the adhesion and the biofilm formation of Cutibacterium (Propionibacterium) acnes. AIMS Microbiol. 2018;4:165–172.
23. Tsai PJ, Huang WC, Hsieh MC, Sung PJ, Kuo YH, Wu WH. Flavones isolated from Scutellariae radix suppress Propionibacterium acnes-induced cytokine production in vitro and in vivo. Molecules. 2016;21:15–26.
24. Cappiello F, Casciaro B, Mangoni ML. A novel in vitro wound healing assay to evaluate cell migration. J Vis Exp. 2018;133:e56825.
25. Achermann Y, Goldstein EJC, Coenye T, Shirtliff ME. Propionibacterium acnes: from commensal to opportunistic biofilm-associated implant pathogen. Clin Microbiol Rev. 2014;27:419–440.
26. Otlewska A, Baran W, Batycka-Baran A. Adverse events related to topical drug treatments for acne vulgaris. Expert Opin Drug Saf. 2020;19:513–521.
27. Salem HF, Kharshoum RM, Abou-Taleb HA, Farouk HO, Zaki RM. Fabrication and appraisal of simvastatin via tailored niosomal nanovesicles for transdermal delivery enhancement: in vitro and in vivo assessment. Pharmaceutics. 2021;13:138.
28. Eroǧlu I, Aslan M, Yaman Ü. Liposome-based combination therapy for acne treatment. J Liposome Res. 2020;30:263–273.
29. Veloso DFMC, Benedetti NIGM, Ávila RI, et al. Intravenous delivery of a liposomal formulation of voriconazole improves drug pharmacokinetics, tissue distribution, and enhances antifungal activity. Drug Deliv. 2018;25:1585–1594.
30. Kannan S, Solomon A, Krishnamoorthy G, Marudhamuthu M. Liposome encapsulated surfactant abetted copper nanoparticles alleviates biofilm mediated virulence in pathogenic Pseudomonas aeruginosa and MRSA. Sci Rep. 2021;11:1102.
31. Botequim D, Maia J, Lino MMF, et al. Nanoparticles and surfaces presenting antifungal, antibacterial and antiviral properties. Langmuir. 2012;28:7646–7656.
32. Jahns AC, Eilers H, Alexeyev OA. Transcriptomic analysis of Propionibacterium acnes biofilms in vitro. Anaerobe. 2016;42:111–118.
33. Wolfmeier H, Pletzer D, Mansour SC, Hancock RE. New perspectives in biofilm eradication. ACS Infect Dis. 2018;4:93–106.
34. Shukla SK, Rao TS. Dispersal of Bap-mediated Staphylococcus aureus biofilm by proteinase K. J Antibiot. 2013;66:55–60.
35. Nguyen UT, Burrows LL. DNase I and proteinase K impair Listeria monocytogenes biofilm formation and induce dispersal of pre-existing biofilms. Int J Food Microbiol. 2014;187:26–32.
36. Zerillo L, Polvere I, Varricchio R, et al. Antibiofilm and repair activity of ozonated oil in liposome. Microb Biotechnol. 2022;15:1422–1433.
37. Barros CHN, Hiebner DW, Fulaz S, Vitale S, Quinn L, Casey E. Synthesis and self-assembly of curcumin-modified amphiphilic polymeric micelles with antibacterial activity. J Nanobiotechnol. 2021;19:104.
38. Blackman LD, Qu Y, Cass P, Locock KES. Approaches for the inhibition and elimination of microbial biofilms using macromolecular agents. Chem Soc Rev. 2021;50:1587–1616.
39. Cho H, Choi Y, Min K, et al. Over-activation of a nonessential bacterial protease DegP as an antibiotic strategy. Commun Biol. 2020;3:547.
40. Culp E, Wright GD. Bacterial proteases, untapped antimicrobial drug targets. J Antibiot. 2017;70:366–377.
41. Zhu T, Fang F, Sun D, et al. Piceatannol inhibits P. acnes-induced keratinocyte proliferation and migration by downregulating oxidative stress and the inflammatory response. Inflammation. 2020;43:347–357.
42. Dell’Annunziata F, Cometa S, Marca RD, et al. In vitro antibacterial and anti-inflammatory activity of Arctostaphylos uva-ursi leaf extract against Cutibacterium acnes. Pharmaceutics. 2022;14:1952.
43. Zasada M, Budzisz E. Retinoids: active molecules influencing skin structure formation in cosmetic and dermatological treatments. Adv Dermatol Allergol. 2019;4:392–397.
44. Forroozandeh P, Aziz AA. Insight into cellular uptake and intracellular trafficking of nanoparticles. Nanoscale Res Lett. 2018;13:339.
45. Liu J, Zheng A, Peng B, Xu Y, Zhang N. Size-dependent absorption through stratum corneum by drug-loaded liposomes. Pharm Res. 2021;38:1429–1437.
46. Phatale V, Vaiphei KK, Jha S, Patil D, Agrawal M, Alexander A. Overcoming skin barriers through advanced transdermal drug delivery approaches. J Control Release. 2022;351:361–380.
47. Yoshioka C, Ito Y, Nagai N. Enhanced percutaneous absorption of cilostazol nanocrystals using aqueous gel patch systems and clarification of the absorption mechanism. Exp Ther Med. 2018;15:3501–3508.
48. Zhang ZJ, Michniak-Kohn M. Flavosomes, novel deformable liposomes for the co-delivery of anti-inflammatory compounds to skin. Int J Pharm. 2020;585:119500.
49. Yu YQ, Yang X, Wu XF, Fan YB. Enhancing permeation of drug molecules across the skin via delivery in nanocarriers: novel strategies for effective transdermal applications. Front Bioeng Biotechnol. 2021;9:646554.
50. Coenye T, Spittaels KJ, Achermann Y. The role of biofilm formation in the pathogenesis and antimicrobial susceptibility of Cutibacterium acnes. Biofilm. 2022;4:100063.
51. Kim H, Park SH, Oh SW, et al. Olfactory receptor OR7A17 expression correlates with all-trans retinoic acid (ATRA)-induced suppression of proliferation in human keratinocyte cells. Int J Mol Sci. 2021;22:12304.
52. Eichenfield DZ, Sprague J, Eichenfield LF. Management of acne vulgaris. A review. JAMA. 2021;326:2055–2067.
53. Kong R, Cui Y, Fisher GJ, et al. A comparative study of the effects of retinol and retinoic acid on histological, molecular, and clinical properties of human skin. J Cosmet Dermatol. 2015;15:49–57.
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.