Back to Journals » Journal of Inflammation Research » Volume 18
Programmed Cell Death in Diabetic Kidney Disease: Mechanisms and Therapeutic Targeting
Authors Tang S, Sun Y, Sun W, Kang X ![]() , Zhao X, Jiang L, Gao Q, An X, Ji H, Lian F
, Zhao X, Jiang L, Gao Q, An X, Ji H, Lian F
Received 10 June 2025
Accepted for publication 5 September 2025
Published 19 September 2025 Volume 2025:18 Pages 13001—13037
DOI https://doi.org/10.2147/JIR.S545938
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Wenjian Li
Shanshan Tang,1,* Yuting Sun,2,* Wenjie Sun,3,4,* Xiaomin Kang,2 Xuefei Zhao,2 Linlin Jiang,2 Qing Gao,2 Xuedong An,2 Hangyu Ji,2 Fengmei Lian2
1College of Traditional Chinese Medicine, Changchun University of Chinese Medicine, Changchun, People’s Republic of China; 2Guang’anmen Hospital, China Academy of Chinese Medical Sciences, Beijing, People’s Republic of China; 3The Affiliated Hospital of Shandong University of Traditional Chinese Medicine, Jinan, People’s Republic of China; 4Shandong University of Traditional Chinese Medicine, Jinan, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hangyu Ji, Email [email protected] Fengmei Lian, Email [email protected]
Abstract: The escalating incidence and mortality of diabetic kidney disease (DKD) underscore the critical need to elucidate its pathogenesis. Programmed cell death (PCD) plays a dual role in maintaining physiological homeostasis and driving pathological processes in DKD. Accumulating evidence demonstrates that apoptosis, autophagy, pyroptosis, and ferroptosis contribute directly or indirectly to DKD progression via distinct gene-regulated signaling pathways. Recently identified PCD modes (eg, necroptosis, parthanatos) remain poorly characterized in DKD, with emerging evidence suggesting crosstalk between different PCD pathways. This review synthesizes current knowledge on PCD-mediated DKD pathogenesis and PCD-targeted therapies, while highlighting research limitations (eg, unclear PCD interactions, translational gaps). We propose that dissecting the multifaceted roles of PCD in DKD will deepen mechanistic understanding and accelerate the development of novel therapeutics, offering significant scientific and clinical benefits.
Keywords: diabetic kidney disease, programmed cell death, apoptosis, autophagy, pyroptosis, ferroptosis
Introduction
Diabetic kidney disease (DKD) is the most common cause of end-stage renal disease (ESRD) and a major microvascular complication of diabetes.1 According to IDF data, the global diabetic population has reached 643 million and is projected to rise to 853 million by 2050, equivalent to one-eighth of the world’s population.2 Between 20% and 40% of diabetic patients develop DKD, associated with significantly increased morbidity and mortalityreflected in a 10-fold higher incidence of kidney failure compared to non-diabetics.3 By exploring mechanisms to identify novel therapeutic approaches for DKD, targeting programmed cell death (PCD) pathways offers a promising strategy to prevent, treat, or accelerate recovery by limiting renal parenchymal cell loss and inflammation.
Cell death, a fundamental process in physiological homeostasis and pathological states,4,5 is classified as accidental (ACD) or regulated (RCD).6 RCD, a genetically controlled suicide mechanism initiated upon failed adaptation to stress,7 is crucial for development and tissue renewal.8 Under pathological conditions like DKD, dysregulated RCD (synonymous with PCD) contributes to organ dysfunction.6,9–12
More than a dozen distinct PCD modes exist, each defined by specific molecular pathways and functional consequences.13 While evolved to protect organisms,14 aberrant PCD drives pathology. This review synthesizes current knowledge on both established PCD pathways (apoptosis, autophagy, pyroptosis, ferroptosis) and emerging forms (eg, necroptosis, parthanatos, alkaliptosis, oxeiptosis)13 within DKD pathogenesis. Critically, therapeutic targeting of PCD must balance efficacy with safety, considering impacts on physiological homeostasis.15 Understanding these mechanisms holds significant scientific and clinical potential for developing novel DKD therapies.
Apoptosis in Diabetic Kidney Disease
Apoptosis is one of the main causes of DKD cell death.16 Apoptosis comprises two main stages: initiation and execution. Based on the initiation mechanism, apoptotic pathways include receptor-mediated (exogenous), perforin/granzyme, and mitochondrial (intrinsic) pathways. The process involves receiving apoptotic signals, regulating molecular interactions, activating caspases, and culminating in a cascade reaction.17 High glucose stimulates podocytes to change albumin-related mitochondrial dynamics, leading to apoptosis.18 STZ-induced α-2u globulin nephropathy during DKD is associated with dysmolar deterioration, tubular adaptive injury, and mitochondria-driven apoptosis.19 CD36 promotes DKD progression by mediating apoptosis through the Wnt/β-catenin pathway;20 conversely, its inhibition ameliorates DKD by suppressing NLRP3 inflammasome activation, thereby reducing inflammation and tubular epithelial cell apoptosis.21 In DKD, extracellular vesicle (EV) transfer between small tubular epithelial cells and macrophages forms a negative feedback loop, promoting renal inflammation, apoptosis, and disease progression.22 The dysfunction of glomerular endothelial cells induced by the diabetic microenvironment secretes factors that cause podocyte apoptosis, and the increase of mitochondrial superoxide levels in glomerular endothelial cells leads to podocyte dysfunction and pathology, contributing to the loss of DKD glomerular filtration barrier.23 Different genes are closely related to DKD through the regulation of apoptosis. Figure 1 shows that different factors regulate DKD through apoptosis (Figure 1). In DKD mice, microtubule associated protein 4 (MAP4) phosphorylation (p-MAP4) was elevated, crotubules and F-actin filaments were reordered, and cell permeability was enhanced. It is also associated with dedifferentiation and apoptosis of podocytes.24 Under high glucose conditions, Smad3-dependent ezrin activation upregulates Nox4 expression and ROS production by suppressing PKA activity; MPC2 mediates mitochondrial dysfunction; and histone deacetylase 4 (HDAC4) promotes calcineurin upregulation, collectively contributing to podocyte apoptosis.25,26 In addition, pro-apoptotic factor Bim promotes the activation of nuclear factor of activated T cells 1 (NFAT2) and induces the downregulation of lncRNA NONHSAT179542.1, leading to cytoskeletal injury, and promoting high glucose-induced podocyte injury.27 Klotho activates Nuclear factor-erythroid 2-related factor 2 (Nrf2) signaling or regulates endoplasmic reticulum stress (ERS) and reactive oxygen species (ROS) and improves renal function and glomerulosclerosis in podocyte injury and DKD mice by regulating apoptosis.27 Firstly, the study of different genes promoting DKD through apoptosis was discussed. Notch signaling pathway regulates oxidative damage and apoptosis by regulating mitochondrial dynetin and biogenetic genes,28 Nrf2 regulates mitochondrial ribosomal protein L12 (MRPL12) transcription by changing mitochondrial function and apoptosis level,29 ATP binding cassette subfamily A member 1 (ABCA1) deficiency leads to apoptosis and inflammation, destroys endocalyx barrier, and induces ERS.30 A‐kinase anchoring protein (AKAP) 1 mediates mitochondrial fission by regulating the phosphorylation of dynamin‐related protein 1 (Drp1), which enhances podocyte apoptosis.31 BASP1 (brain acid-soluble protein) activates p53 and induces podocyte apoptosis by regulating Wilms’ tumor 1 transcription factor (WT1),32 gremlin 2, DAN family BMP antagonist (Grem2) and mannose receptor C-type 2 (MRC2) can induce apoptosis,33,34 ectodysplasin A2 receptor (EDA2R) can induce apoptosis and dedifferentiation of podocyte cells by enhancing ROS production,35 Decoy receptor 2 interacts with glucose regulated protein 78 (GRP78) to regulate the apoptosis phenotype mediated by caspase-3,36 renal (pro)renin receptor (pRR) induces mitochondrial dysfunction and upregulates the mitochondrial NADPH oxidase 4 (NOX4)/superoxide dismutase 2 (SOD2)/uncoupling protein 2 (UCP2) signaling pathway,37 up-regulation of cell division cycle 42 (Cdc42) can increase BCL2 (B-cell lymphoma-2) associated X (Bax) and cleaved caspase-3 levels and decrease the expression of Bcl-2, inducing podocyte apoptosis,38 Alk1 haploinsufficiency leads to glomerular apoptosis, glomerular basement membrane thickening, and podocyte loss,39 and X-C motif chemokine ligand 1 (XCL1) induces inflammation and cell apoptosis by regulating p53/Nuclear Factor -Kappa B (NF-κB) pathway,40 the PH domain and leucine rich repeat protein phosphatase 1 (PHLPP1)/ Nrf2-Mdm2 axis induces apoptosis of renal cells by regulating the forkhead box O1 (FoxO1) cytoplasmic shuttle,41 Rho associated coiled-coil containing protein kinase 2 (ROCK2) can inhibit peroxisome proliferator-activated receptors α (PPARα), rewiring cell programs, and negatively regulate transcription of fatty acid oxidation-related genes induce apoptosis,42 Sestrin-2 regulates mitochondrial dysfunction and apoptosis of podocytes through protein kinase AMP-activated catalytic subunit alpha 1 (AMPK),43 Myo-inositol Oxygenase (MIOX) overexpression can enhance apoptosis by regulating oxidative and ERS,44 methyltransferase 14 (METTL14) activates MAPK/mitogen-activated protein kinase 1 (ERK) pathway through m6A modification of taurine up-regulated 1 (TUG1) to promote apoptosis of renal tubular epithelial cells and ERS,45 promote DKD renal injury, and aggravate renal dysfunction. Inhibition of activating transcription factor 5 (ATF5) promotes mitochondrial ROS levels and apoptosis by regulating mitochondrial unfolded protein response,46 inducing DKD kidney injury. The progression of DKD in mice with phosphofurin acidic cluster sorting protein 2 (PACS-2) gene knockout was accelerated, with mitochondria-associated ER membranes (MAM) dysfunction apoptosis, and fibrosis.47 At the same time, regulating some genes can also treat DKD by controlling apoptosis. A carbohydrate responsive element binding protein (ChREBP) deficiency can inhibit oxidative stress (OS) and ERS, down-regulate the expression of Bax, upregulate the expression of Bcl-2, and down-regulate cleaved caspase-3 levels,48 METTL14 silence reduces ROS production and inflammatory cytokine levels and inhibits apoptosis.49 The complement component 1, q subcomponent (C1q)/Tumor Necrosis Factor-Related Protein-9 inhibits apoptosis of cells under OS,50 D(P)H: quinone oxidoreductase 1 (NQO1) enhances intracellular NAD+/NADH level regulation Sirtuin 1 (Sirt1), decreased Bax/Bcl-2 ratio and cleaved caspase-3 expression,51 GTS 21, a selective alpha 7 nicotinic acetylcholine receptor agonist, reduced diabetes-induced kidney injury by decreasing DKD mesangial matrix expansion and apoptosis, and Bax and cleaved caspase-3 expression,52 cancer susceptibility candidate 2 (CASC2) increased Bcl-2 levels by regulating the miR-144/suppressor of cytokine signaling 2 (SOCS2) axis. Decreased cleaved caspase-3 expression reduces apoptosis, inflammation, and fibrosis,53 and dopamine 1 receptor (D1R) activation protects DKD podocytes from apoptosis and oxidative damage through the PKA/NADPH oxidase 5 (NOX-5)/p38 MAPK axis.54 The absence of thioredoxin-interacting protein (TXNIP) mitigated podocellular apoptosis by inhibiting p38 AMPK or mechanistic target of rapamycin kinase (mTOR) signaling pathways,55 and silted cyclin G1 (CCNG1) improved the expression of Bcl-2, Bax, and p53 by modulating the MDM2/p53 signaling pathway.56 Down-regulation of SET domain containing 6 (SETD6) can improve mitochondrial dysfunction and inhibit apoptosis by activating the Nrf2-Keap1 signaling pathway.57 After RING-finger protein 166 (RNF166) knockout interacts with CYLD to reduce cleaved Caspase-9 expression and inhibit mitochondrial damage and apoptosis.58 The zinc transporter 7 in SLC30 family (SLC30A7) regulates apoptosis through NFE2 like bZIP transcription factor 2 (NFE2L2)/heme oxygenase 1 (HMOX1) signaling pathway,59 GLIS family zinc finger 2 (Glis2) alleviates apoptosis through β-catenin signaling pathway.60 HIF-1α (hypoxia inducible factor 1 subunit alpha) alleviates ROS accumulation and apoptosis through mitochondrial kinetic control mediated by HO-1 (heme oxygenase 1),61 Forkhead box A2 mediated upregulation of lncRNA SOX2OT inhibits OS and apoptosis by promoting SIRT1,62 while lack of complement factor B (CFB) improves apoptosis, ERS, and OS by inhibiting ceramide synthesis.63 INSL3 (insulin like 3) can inhibit the rate of apoptosis.64 Polysulfide-mediated sulfhydration of SIRT1 can inhibit OS, apoptosis, and inflammation by inhibiting the phosphorylation and acetylation of p65 NF-κB and signal transducer and activator of transcription 3 (STAT3).65 FoxO1 inhibits STAT1 level and attenuates apoptosis,66 FGF1ΔHBS inhibits apoptosis and inflammatory response by activating PPARα.67 Tumor Necrosis Factor Receptor-Associated Protein 1 alleviates OS and apoptosis by preventing abnormal opening of mPTP and maintaining mitochondrial structure and function.68 SIK2 (salt inducible kinase 2) overexpression activates HSF1 (heat shock transcription factor 1)/Hsp70 (heat shock protein 70) by inhibiting histone acetyltransferase activity of p300, thereby reducing ERS-mediated apoptosis of tubule epithelial cells,69 and SAR131675 inhibits EGF-C and VEGFR-3, reducing OS, apoptosis, and related inflammatory responses,70 Disulphide-bond A oxidoreductase-like protein (DsbA-L) by maintaining the mitochondrial associated ER membrane and the integrity of MAM plays an anti-apoptotic role,71 and the lack of TDAG51 (pleckstrin homology like domain family A member 1) activates Nrf2 through the Akt-GSK (inosine/guanosine kinase) 3β pathway, reducing apoptosis, OS and inflammation.72 Gasdermin D knockdown down-regulates the expression of Bax and caspase-3 through the C-Jun N-Terminal Kinase (JNK) pathway and reduces REDD1 (DNA damage inducible transcript 4)-regulated AKT/GSK-3β/Nrf2 pathway by regulating apoptosis and inflammation.73 Both decreasedDNA damage response 1 (REDD1) expression74 and Tripartite motif-containing protein 32 (TRIM32) inhibition75 confer protection against high glucose-induced podocyte injury through convergent potentiation of Nrf2 signaling via modulation of the AKT/GSK-3β pathway. Inhibition of high mobility group box 1 (HMGB1) can reduce podiocyte apoptosis by regulating autophagy flux,76 and ZnT8 can inhibit the expression of apoptotic protein through the (TNFAIP3) TNF alpha induced protein 3NF-κB pathway and play an anti-apoptotic role.77 IL-8 (interleukin-8)-CXCR1/2 axis blockade could alleviate podocyte apoptosis and DNA damage,78 and overexpression of PCK1 (phosphoenolpyruvate carboxykinase 1) could reduce apoptosis and collagen IV deposition around the tube, protecting mitochondrial defects.79 ANRIL knockdown can improve metabolic pathway, apoptosis, extracellular matrix synthesis and degradation, NF-κB-related pathway, AGE (advanced glycation end products)-RAGE (receptor for advanced glycation end products) interaction, etc.;80 TFPI2 (tissue factor pathway inhibitor 2) interacts with TGF-β2 (transforming growth factor beta 2) pathway to promote EMT of DKD and knockdown TFPI2 can reduce apoptosis.81 (+)-trans-Cannabidiol-2-hydroxy pentyl is a dual CB1R antagonist/CB2R agonist to reduce CD3+T cell infiltration by inhibiting apoptosis and inflammatory response.82
 |
Figure 1 Shows that different factors regulate DKD through apoptosis. By Figdraw. |
miRNA Regulates Cell Apoptosis and DKD
miRNA regulates apoptosis and DKD. Sequencing of patients with type 2 DKD showed that mirnas of urinary extracellular vesicles such as hsa-miR-375, hsa-miR-503, and hsa-miR-451a were involved in apoptosis and inflammation and promoted the occurrence of DKD83 and MicroRNA-494-3p promotes apoptosis by targeting SOCS6 (suppressor of cytokine signaling 6).84 Mir-27a-3p regulation prohibition and TMBIM6 (transmembrane BAX inhibitor motif containing 6) inhibition of extracellular matrix accumulation, mitochondrial dysfunction, ERS, and apoptosis.85 Noncoding RNA XIST can regulate apoptosis through the miR-423-5p/HMGA2 (high mobility group AT-hook 2) axis.86 MicroRNA‐182-5p and CD2AP (CD2 associated protein) dysregulation induce podocyte apoptosis,87 and miR-770-5p promotes podocyte apoptosis and inflammatory response by targeting TIMP3 (TIMP metallopeptidase inhibitor 3).88 CircHIPK3/FUS complex leads to the upregulation of ectodysplasin A2 receptor (EDA2R) and promotes the activation of apoptosis.89 Long Noncoding RNA (lncRNA) ENSG00000254693 interacts with HuR to induce inflammation and apoptosis,90 LncRNA MALAT1 induces apoptosis, ROS production, and inflammation through activation of LIN28A (lin-28 homolog A) and the Nox4/AMPK/mTOR signaling axis91 and increases apoptosis and inflammation through miR-15b-5p/TLR4 (toll like receptor 4) signaling axis.92 LncRNA NEAT1 regulates inflammation, OS, and apoptosis through the miR-423-5p/GLIPR2 (GLI pathogenesis related 2) axis,93 miR-20a over-expression enhanced cell proliferation, inhibited cell apoptosis, and suppressed the inflammatory response of HK-2 cells,94 and high glucose induces mesangial cell apoptosis through miR-15b-5p.95 High glucose-induced KCNQ1OT1 (KCNQ1 opposite strand/antisense transcript 1) increases Sema3A (semaphorin 3A) expression through sponging miR-23b-3p, promoting inflammatory response and apoptosis;96 Circ_0003928 acted as a sponge for miR-506-3p, HDAC4 (histone deacetylase 4) regulates OS and apoptosis,97 DACH1 (dachshund family transcription factor 1), a novel target of miR-218, promote apoptosis and inflammatory response,98 encourages and is implicated in preventing DKD kidney injury and renal dysfunction. Circ_LARP4 is decreased in the DKD model, after sponging miR-424 treatment, Circ_LARP4 overexpression leads to increased apoptosis.99 LncRNA PVT1 silencing enhances Bcl-2 expression by upregating FOXA1. Down-regulation of Bax and cleaved caspase-3 expression inhibited podocyte apoptosis,100 and Circ_0000491 inhibited apoptosis, inflammation, OS, and fibrosis by regulating miR-455-3p/Hmgb1 (high mobility group box 1) signaling axis.101 The miR‐181b promotes cell survival and inhibits apoptosis,102 LncRNA Airn alleviates the decline of cell viability and inhibits apoptosis by binding Igf2bp2 (insulin like growth factor 2 mRNA binding protein 2) and promoting the translation of Igf2 and Lamb2 (laminin subunit beta 2);103 LncRNA TUG1 reduces ERS and apoptosis by inhibiting PU.1/RTN1 (reticulon 1) signaling pathway.104 Circ_0008529 knockdown can reduce inflammation and apoptosis through the Circ_0008529-mediated miR-485 5p/WNT2B (Wnt family member 2B) signaling pathway,105 and mirNA-483-5p targets HDCA4 (histone deacetylase 4) to inhibit inflammation, ROS production, and apoptosis.106 miRNA 342 targets SOX6 (SRY-box transcription factor 6) to inhibit apoptosis,107 upregulates miR-20a to inhibit CXCL8 (C-X-C motif chemokine ligand 8) expression and inhibit apoptosis and inflammation,94 and miR-34b improves inflammation and apoptosis through IL-6R (interleukin 6 receptor) /JAK2 (Janus kinase 2) /STAT3 (signal transducer and activator of transcription 3) signaling pathway.108 GDF11 (growth differentiation factor 11), a target of miR-32-5p, inhibits mitochondrial dysfunction and apoptosis through the phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway,109 and inhibition of miR-17-92 Cluster inhibits cell apoptosis, inflammation and fibrosis.110 Hsa_circ_0003928 inhibits apoptosis and OS through the axis of miR 136 5p/PAQR3 (progestin and adipoQ receptor family member 3),111 and microRNA-29b-3p inhibits apoptosis and inflammatory response through modification of EZH2 (enhancer of zeste 2 polycomb repressive complex 2 subunit).112 LncRNA UCA1 inhibits apoptosis and inflammation of renal tubular epithelial cells by targeting microRNA-206.113 CircRNA circ_0000712 inhibits apoptosis, OS, and inflammation by targeting miR-879-5p /SOX6 axis,114 and miR 1423p inhibits apoptosis and OS by targeting cell division 1 (BOD1).115 Mir-1297 inhibits inflammation and apoptosis by targeting COL1A2 (collagen type I alpha 2 chain),116 and miR-590-3p inhibits apoptosis, inflammation, and OS by targeting C-X3-C motif chemokine ligand 1 (CX3CL1).117 The microRNA-126 inhibits inflammation and apoptosis through the VEGF-mediated PI3K/AKT signaling pathway118 and inhibits DKD both in vivo and in vitro.
Drugs and Compounds Ameliorate DKD by Modulating Cell Apoptosis
Drugs and compounds improve DKD by regulating apoptosis. Sodium-glucose cotransporter type 2 inhibitors (SGLT2), dapagliflozin inhibits inflammation-related expressions by improving the expression of apoptotic markers Bcl-2 and Bax119 and inhibits of ERS.120 Empagliflozin inhibits apoptosis and improves mitochondrial function by regulating mitochondrial fission and fusion;121 ursolic acid and empagliflozin inhibit abnormal apoptosis of glomerular cells induced by high glucose. Reducing inflammation and OS can improve the renal histopathological changes of DKD.122 Liraglutide promotes the browning of white fat and inhibits apoptosis of podocytes in DKD mice through GLP-1R.123 Telmisartan can inhibit dimerization of angiotensin type-1 receptor and adiponectin receptor-1 and alleviate cell apoptosis.124 Fenofibrate inhibited apoptosis through AMPK/FOXA2 (forkhead box A2) /MCAD (medium-chain acyl-CoA dehydrogenase) pathway,125 and Tacrolimus reduced Bax/Bcl-2 and cleaved caspase-3 levels by down-regulating TRPC6 (transient receptor potential cation channel subfamily C member 6), reducing the percentage of apoptosis.126 Tacrolimus can reduce inflammatory markers and apoptosis by inhibiting nuclear factor of activated T cells 1 (NFATc1) /TRPC6 pathway.127 Verapamil can inhibit apoptosis,128 hydralazine can inhibit XO/NADPH glycosylase and activate Nrf-2/HO-1 to reduce ROS production. Down-regulation of the poly (ADP-ribose) polymerase (PARP) /caspase-3 signaling pathway can reduce apoptosis,129 and Linagliptin can inhibit podocyte apoptosis by regulating the insulin receptor substrate 1 (IRS1) /Akt signaling pathway.130 Galantamine can reduce inflammation and apoptosis by regulating the activity of p38 MAPK and caspase-1 pathway131 and play a role in treating DKD.
In addition, the combination of drugs has also shown therapeutic effects. Cyproheptadine, a SET7/9 inhibitor, reduced apoptosis and inflammation and alleviated DKD tubular epithelial cell fibrosis by decreasing H3K4Me1 expression and E.R. stress.132 LCZ696 (valsartan/sacubitril) inhibits the expression of Bax and caspase-3 protein, enhances the expression of Bcl-2 protein, improves apoptosis and inflammation, and protects DKD kidney injury.133 The alpha-lipoic acid (ALA) supplementation inhibits renal fibers through anti-inflammatory, antioxidant, and anti-apoptotic effects and delays the progression of DKD.134 The sinapic acid and ellagic acid can synergistically inhibit caspase 3-mediated apoptosis, improve DNA damage and structural changes, and improve renal function in DKD patients.135 Paricalcitol and omega-3 fatty acids can reduce TGF-β1/iNOS/NGAL (neutrophil gelatinase-associated lipocalin) /KIM-1 (kidney injury molecule 1) /caspase-3 and apoptosis index and protect DKD through anti-inflammatory, antioxidant, and anti-apoptosis effects.136 Intermedia (IMD) intervention in DKD rats has been shown to block endoplasmic reticulum stress, alleviate podocyte apoptosis and F-actin rearrangement, reduce diaphragm protein synthesis, and protect DKD.137 Renalase inhibits apoptosis and improves DKD tubulointerstitial fibrosis through the p38 MAPK signaling pathway.138 Praliciguat inhibits inflammation and apoptosis and delays the progression of DKD.139 Pyruvate can inhibit ERS and apoptosis and alleviate the damage of renal tubular epithelial cells induced by high glucose.140 Hyperoside targets miR-499e5p/APC axis to reverse the increase of Bax and the decrease of Bcl-2, enhance the activity caspase-3, inhibit cell apoptosis, and alleviate kidney damage and fibrosis in DKD mice.141 Phosphocreatine protects DKD by reducing the expression of Bax/Bcl-2 ratio, caspase-9, and caspase-3 through the ERK/Nrf2/HO-1 signaling pathway and reducing the production of ROS.142 The prostaglandin E1 (PGE1) reduced Bax, caspase-3, and cleaved caspase-3 levels by inhibiting the JNK/Bim pathway and inhibited apoptosis to protect DKD rats from proximal renal tubule injury.143 CY-09 inhibits NLRP3, reduces caspase-1 and apoptosis in a dose-dependent manner, and alleviates DKD renal injury.144 β-Amyrin regulates the miR-181b-5p/HMGB2 axis to inhibit the inflammatory response and apoptosis and alleviate the renal histopathological changes of DKD.145 Inhibition of caspase-3 by Z-DEVD-FMK can improve proteinuria, renal function, and tubulointerstitial fibrosis of DKD.146 SC preparations mitigate renal cell apoptosis and mitochondrial dysfunction through Nrf2-dependent mechanisms.147 Finerenone ameliorates apoptosis in diabetic nephropathy by suppressing macrophage mineralocorticoid receptor (MR) and its downstream G protein subunit alpha i2 (Gnαi2) signaling, thereby reducing inflammation.148 Isoferulic acid (IFA) ameliorates apoptosis in diabetic nephropathy by inhibiting the CXCL12/CXCR4 signaling axis and its downstream PI3K/Akt/mTOR/p53/CASK pathways, while activating autophagy.149 β-Sitosterol ameliorates apoptosis in diabetic nephropathy by inhibiting the TLR4/NF-κB signaling pathway, thereby reducing oxidative stress and inflammation in podocytes and renal tubular cells exposed to high glucose.150 Micheliolide ameliorates apoptosis in diabetic kidney disease by activating the Nrf2/Keap1 antioxidant pathway and inhibiting the TLR4/MyD88/NF-κB inflammatory axis.151
Traditional Chinese medicine (TCM) compounds, single TCM, and TCM extracts can improve DKD by regulating apoptosis. Loganin and catalpol inhibit podocyte apoptosis and improve kidney injury in DKD mice by targeting AGE-RAGE and its downstream pathways p38 MAPK and Nox4.152 Huperzine A (Hup A) ameliorates apoptosis in diabetic nephropathy by modulating the Apoe/Apoc2 pathway and restoring lipid metabolism, while concurrently enhancing microbial homeostasis.153 Grifola frondosa (PGF) inhibits the TLR4/NF-κB pathway, reduces inflammatory response and apoptosis, and improves early DKD.154 Paclitaxel can inhibit the expression of inflammatory cytokines, down-regulate the expression of apoptosis markers Bax and caspase-3 and improve the damage and fibrosis of DKD podocytes.155 Resveratrol relies on the activation of AMPK to inhibit OS-mediated podocellular apoptosis and alleviate DKD kidney injury.156 Sinomenine can inhibit OS, reduce renal cell apoptosis, and improve DKD renal fibrosis by regulating the JAK2/STAT3/SOCS1 (suppressor of cytokine signaling 1) pathway.157 Puerarin alleviates podocyte apoptosis and DKD kidney injury by interacting with Guanidine nucleotide-binding protein Gi subunit alpha-1 (Gnai1) subunit.158 Diosgenin inhibits ROS production through the regulation of NOX4 and mitochondrial respiratory chain, down-regulates the expression of caspase 3 and caspase 9, upregulates the expression of Bcl-2, inhibits mitochondrial apoptosis, and improves DKD kidney injury through ERS.159 Salidroside inhibits the expression of inflammatory factors and OS markers through the Akt/GSK-3β signaling pathway, attenuates apoptosis characteristics, and improves DKD renal dysfunction.160 Hedysarum polypores polysaccharides can reduce apoptosis and inflammatory infiltration by inhibiting the HMGB1/RAGE/TLR4 pathway and improving DKD renal fibrosis.161 Grape seed Proanthidin extract targets p66Shc to improve mitochondrial dynamic homeostasis, apoptosis, and ROS production and treat DKD.162 Cordyceps sinensis can down-regulate the expression of Bax and caspase-3, inhibit apoptosis and cell proliferation, and inhibit DKD.163 Through anti-apoptosis and antioxidant effects, pectin-lyase-modified ginseng extract and Ginsenoside Rd protect against DKD renal dysfunction.164 Akebia Saponin D inhibits renal tubule cell apoptosis and OS by activating the Nrf2/HO-1 pathway, inhibiting the NF-κB pathway, and improving renal injury in DKD mice.165 Ginsenoside Rh1 improves cytochrome c, Bax, Bcl-2, Bcl-XL, cleaved-caspase 9, and cleaved-caspase 3 in the NF-κB signaling pathway through AMPK/PI3K/Akt. DKD is improved by inhibiting inflammation and apoptosis.166 Panax japonicus C.A. Meyer (P.J.) regulates the Bcl-2/caspase-3 signaling pathway, inhibits apoptosis, and alleviates kidney damage in DKD mice.167 20(S)-Ginsenoside Rg3 reduces apoptosis, inhibits inflammatory response, and alleviates renal histological changes in DKD rats.168 Jujuboside A (JuA) down-regulates the expression of apoptotic proteins Bax, CytC, Apaf-1, and caspase 9 by inhibiting mitochondrial and ERS, inhibits apoptosis, and improves renal pathological damage in DKD rats.169 Phillygenin (PHI) inhibits inflammation and apoptosis in vitro and alleviates diabetic kidney injury in db/db mice by interfering TLR4/MyD88/NF-κB and PI3K/AKT/GSK3β signaling pathways.170
In addition, modern novel therapies have also been shown to treat DKD by modulating apoptosis. The human umbilical cord-derived mesenchymal stem cells (hucMSCs) inhibited apoptosis by regulating Bax level through Nrf2 activation and alleviated renal oxidative damage of DKD.171 Umbilical Cord-Derived Mesenchymal Stem Cells activate apoptosis signal-regulating kinase 1 (ASK1), which inhibits apoptosis and improves renal cell injury and proteinuria in DKD rats.172 Mitochondria transfer from mesenchymal stem cells can enhance the expression of mitochondrial superoxide dismutase 2 and Bcl-2 and inhibit ROS production. Structural and functional repair of proximal renal tubule epithelial cell damage in DKD rats through antioxidant and anti-apoptotic effects.173 Extracellular vesicles of podocytes treated with high glucose-induced apoptosis of proximal tubule epithelial cells and induced DKD.174 Table 1 summarizes different interventions that delay the progression of DKD by regulating apoptosis (Table 1).
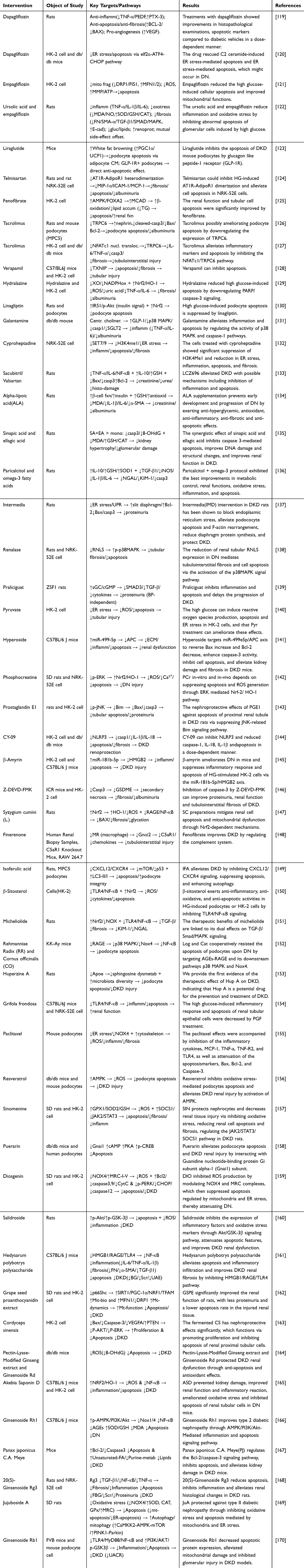 |
Table 1 Different Intervention Methods Delayed the Progression of DKD by Regulating Apoptosis |
Autophagy in Diabetic Kidney Disease
Autophagy is vital in recycling excess or damaged cellular components and maintaining homeostasis and survival. During the process of autophagy, misfolded proteins and damaged organelles are brought into the double-membrane vesicles of the autophagosome for degradation by lysosomes.175 Autophagy is divided into three main forms: Macroautophagy (hereafter referred to as autophagy), in which cellular cargo is isolated within a double-membrane vesicle called an autophagosome, and selection of the contents of the autophagosome can be carried out in a relatively non-selective manner. It may also involve the elimination of strictly regulated individual cellular components. In contrast, chaperone-mediated autophagy (CMA) is protein-specific autophagy; heat-shock cognate protein HSPA8/HSC70 first identified KFERQ-like motif-bearing proteins. The pathway is formed by oligomerizing the protein-bound LAMP2A (lysosomal-associated membrane protein 2A) and translocation into lysosomal degradation. Finally, microautophagy involves direct isolation of cellular material (including the KFERQ-flagged protein or many cytoplasmic contents) through membrane invasions formed on the surface of late endosomes or lysosomes.176 The role of autophagy in cell death is mainly divided into autophagy-dependent cell death (ADCD) (or autophagic cell death, ACD) and autophagy-mediated cell death (AMCD).177 Bioinformatics analysis of the data set of DKD patients showed that autophagy inhibition occurred in both glomeruli and renal tubules, of which podocyte inhibition was the most obvious, and the level of p62 protein in glomeruli was one of the predictive indicators for DKD patients to enter the stage of massive proteinuria.178 Clock-dependent regulation of autophagy is essential for podocyte survival, and loss of circadian control of autophagy plays an important role in DKD podocyte injury and proteinuria.179 A retrospective study included 120 patients with diabetes without albuminuria, diabetes with microalbuminuria, diabetes with macroalbuminuria, and a healthy control group. Studies have found that RUBCN (rubicon autophagy regulator), mTOR, and SESN2 (sestrin 2) are overexpressed in DKD patients, and RUBCN/ SESN2-mediated autophagy inhibition can be used as a biomarker for DKD.180 The prolyl4-hydroxylase subunit beta (P4HB) was identified as a new autophagy-associated biomarker, and P4HB in renal tubules was associated with DKD renal function.181 Figure 2 shows that different factors regulate DKD through autophagy (Figure 2). Some factors promote the development of DKD by inhibiting autophagy. The increased expression of NBR1 and the decreased expression of autophagy related 4B cysteine peptidase (ATG4B) and VPS37 in kidney tissue are closely related to the inhibition of the autophagy pathway,182 and the lack of IL-17 (interleukin 17) may be related to the occurrence or progression of DKD by inhibiting the formation of Autophagy in DKD mice.183 When mice were injected with macrophage-derived exosomes stimulated by high glucose, renal dysfunction and mesangial matrix expansion were observed in mice, which may be related to the activation of NLRP3 inflammasome and the defects of autophagy.184 In cadmium exposure, abnormal autophagy associated proteins and autophagosome count inhibit autophagy,185 and PACS-2 deficiency inhibits autophagy of the ER by TFEB /FAM134B pathway.186 GPR43 (free fatty acid receptor 2) activation-mediated lipotoxicity inhibits autophagy by regulating the ERK/EGR1 (early growth response 1) pathway,187 and high glucose inhibits autophagy by activating the JAK/STAT pathway in mice and podocytes.188 Allograft inflammatory factor-1 (AIF-1) regulates autophagy, OS, and inflammatory response through the miR-34a/ATG4B pathway.189 Activating transcription factor 4 (ATF4) directly inhibit autophagy,190 and ceramide synthase 6 (CERS6) -derived ceramides are involved in the pathogenesis of DKD by inhibiting mitochondrial autophagy mediated by PTEN-induced kinase 1 (PINK1).191
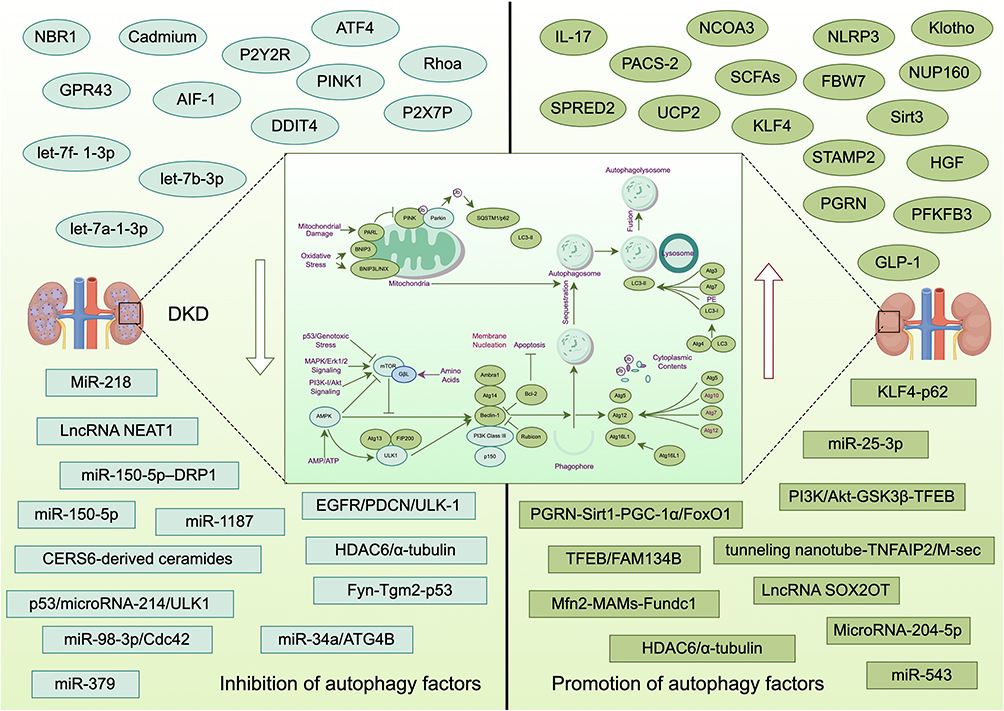 |
Figure 2 Shows that different factors regulate DKD through autophagy. By Figdraw. |
DKD Regulates miRNA-Modulated Autophagy
DKD is involved in regulating autophagy at the miRNA level.MiR-1187 can inhibit autophagy levels in podocytes and glomerulus of DKD mice with high glucose exposure,192 MiR-218 can inhibit Spred2-mediated autophagy and promote OS and inflammation.193 LncRNA NEAT1 inhibits mitochondrial autophagy through the miR-150-5p-DRP1 axis,194 and the p53/microRNA-214/ULK1 axis damages renal tubular autophagy.195 Arsenic alters autophagy through miRNA-mRNA axles of let-7a-1-3p, let-7b-3p, let-7f-1-3p, miR-98-3p/Cdc42, Mapk1, and Rhoa,196 which aggravate kidney damage in DKD. P2X7P is elevated in DKD condition, and podocyte autophagy is inhibited by Akt-mTOR pathway,15,197 and UCP2 deficiency inhibits podocyte autophagy.198 Lack of nuclear receptor coactivator 3 (NCOA3) inhibits podocyte autophagy,199 and the activation of EGFR signal in podocyte accelerates the progression of DKD by inhibiting autophagy to a certain extent by increasing the expression of Rubicon.200 Studies have also shown that mitochondrial biogenic markers are reduced in podocytes treated with high glucose, and PINK1/parkin-dependent mitochondrial autophagy is inhibited in cells, which is the mechanism of glomerular and podocyte injury induced by high glucose.201 DNA damage-inducing transcription factor 4 (DDIT4) participates in the vitamin D receptor (VDR) -mTOR pathway, and DDIT4 processing participates in DKD by regulating autophagy.202 AdipoRon is an autophagy-promoting adiponectin receptor activator, which can relieve the high glucose-induced decrease in the level of lipid deposition in HK-2 cells, suggesting that autophagy-mediated lipid deposition (ELD) plays a key role in DKD renal ELD and lipid-related kidney injury.203 In DKD mice, Fyn regulates autophagy-mediated p53 expression through Transglutaminase 2 (Tgm2), suggesting the role of the Fyn-TGm2-p53 axis in developing DKD.204
Many studies have shown that different factors delay the development of DKD by inducing autophagy. Studies have suggested that different genes regulate autophagy. Overexpression of STAMP2 can promote autophagy by inhibiting mTOR and activating AMPK/SIRT1 signaling pathway,205 and progranulin (PGRN) can maintain mitochondrial homeostasis through mitochondrial biogenesis mediated by PGRN-SIRT1-PGC-1α /FoxO1 signaling and mitochondrial autophagy.206 The KLF4-p62 axis regulates the mTOR/S6K pathway and autophagy,207 and hepatocyte growth factor (HGF) improved the podocyte autophagy lysosomal pathway by regulating the PI3K/Akt-GSK3β-TFEB axis.208 Klotho improves renal tubule autophagy through AMPK and ERK pathways,209 NUP160 regulates autophagy by activating the JAK2/STAT3 signaling pathway,210 and FBW7 (F-box and WD repeat domain containing 7) gene overexpression can enhance autophagy by inhibiting the mTOR signaling pathway and improve inflammation and fibrosis.211 SCFAs enhance podocyte autophagy through the histone deacetylase 2 (HDAC2)/unc-51 axis like ULK1,212 α-tubulin deacetylation mediated by HDAC6 regulates autophagy and enhances podocyte motility.213 In the absence of P2Y2R, the expression of SIRT-1 and FOXO3a is increased, which enhances the autophagy reaction.214 KLF4 activates podocyte autophagy through the mTOR signaling pathway,215 and Sirt3 promotes autophagy by inhibiting the Notch 1/ HIS1 signaling pathway.216 GLP-1 regulates LC3 (light chain 3) and p62 through the AMPK-mTOR signaling pathway to promote autophagy progression.217 6 phosphofructo 2 kinase/fructose 2,6 bios Phase 3 (PFKFB3) silences can induce autophagy,218 tunneling nanotube-TNF alpha induced protein 2 (TNFAIP2) /M-sec system mediates the exchange of autophagosome and lysosome,219 The activated mitofusin 2 (MFN2) -AMS-FundC1 (FUN14 domain containing 1) pathway restores the activity of mitochondrial ATP and complex V and the integrity of mitochondria-associated MAM, promoting the recovery of autophagy,220 thereby mediating the protective effect of DKD. LncRNA SOX2OT regulates Akt-mTOR-mediated autophagy,221 miR-543 increases autophagy by targeting TSPAN8,222 and miR-379 inhibition enhances adaptive mitochondrial autophagy through FIS1.223 MicroRNA-204-5p restores autophagy by regulating the Keap1/Nrf2 pathway,224 and silencing miR-150-5p exerts reno-protective effects in DKD by targeting SIRT1 to promote its interaction with p53, thereby suppressing p53 acetylation and eliciting AMPK-dependent autophagy in podocytes and renal tissue,225 M2 macrophage-derived exosomal miR-25-3p attenuates high glucose-induced podocyte injury by activating autophagy through suppression of DUSP1 expression,226 protection of DKD from the epigenetic level.
Various Drugs or Components Protect DKD by Activating Autophagy
Studies have shown that different drugs or components protect DKD by activating autophagy. Empagliflozin and DPP4 Inhibitor Linagliptin activated glomerulus autophagy and restored autophagosomes, autopilots, and autophagosomes, Engglitazin can reduce albumin exposure and prevent proximal tubular autophagy stagnation.227,228 Ferulic acid promotes the expression of LC3, Inhibit the levels of p62, NLRP3 and IL-1β, enhance autophagy and inhibit inflammation,229 aagliprazin can inhibit the expression of HMGB1 in kidney and restore autophagy,230 montelukast can inhibit HMGB1, TLR4, NF-κB, NLRP3, and IL-1β, and can stimulate autophagy.231 Celastrol inhibits the thickening of the glomerular basement membrane in DKD rats through the PI3K/AKT pathway and protects podocyte homeostasis.232 Soluble epoxide hydrolase (sEH) inhibitor t-AUCB can restore autophagy flux and improve mitochondrial function.233 Metformin-activated AMPK mitigates DKD progression by enhancing renal autophagy, suppressing partial EMT in renal tubular epithelial cells ((RTECs)), and attenuating tubulointerstitial fibrosis(TIF).234 Visceral adipose tissue-derived serine protease inhibitor (vaspin) acts by improving ERS, autophagy injury and lysosome dysfunction of DKD.235 Vitamin D enhanced autophagy activity and improved the number of autophagosomes by improving the expression of LC3, Beclin-1, and Vsp34.236–238 Isoorientin acted as an autophagy activator by activating autophagy and inhibiting the PI3K-AKT-TSC2-mTOR pathway.239 Melatonin enhances autophagy and improves mitochondrial dysfunction by upregulating the AMPK/SIRT1 axis,240 and Dihydromyricetin regulates the PI3K/AKT/mTOR signaling pathway and promotes autophagy by regulating miR-155-5p/PTEN signaling.241 Metformin can regulate the AMPK-mTOR-autophagy axis,242 and Cordyceps militaris polysaccharides can improve the autophagy defect in DKD mice and increase the renal autophagy rate.243 Cyclocarya paliurus triterpenic acids fraction inhibited kidney damage by autophagy regulated by AMPK-mTOR,244 while isorhamnetin improved the kidney lipid profile of DKD rats by regulating autophagy epigenetic regulatory factors, and autophagosomes increase in the kidney.245 Zinc oxide nanoparticles can enhance autophagy activity, reduce inflammation, and regulate the correlation between autophagy and Nrf2/TXNIP/NLRP3 signal transmission.246 Carbon monoxide improves autophagy-mediated partly by dissociated Beclin-1-Bcl-2 complex through senescence-related secretory phenotype (SASP).247 Epidermal growth factor (EGF) protects against high glucose-induced podocyte injury by promoting cell proliferation, suppressing cell apoptosis, and modulating autophagy via the PI3K/AKT/mTOR pathway.248 Calcium Dobesilate (CaD) can restore autophagy by blocking VEGF/VEGFR2 and inhibiting the PI3K/AKT/mTOR signaling pathway,249 and Exogenous spermine can promote autophagy by regulating the AMPK/mTOR signaling pathway.250 Sinensetin can significantly restore the autophagy activity of podocytes induced by high glucose.251 Paecilomyces cicadae-fermented Radix astragali can enhance autophagy by inhibiting PI3K/AKT/mTOR signaling pathway,252 Huang Gui Solid Dispersion (HGSD) activated autophagy by enhancing AMPK phosphorylation level and autophagy-related protein expression.253 Korean red ginseng (KRG) blocks the activation of TGF-β1 by inducing autophagy,254 and Nuciferine (NF) regulates autophagy through PI3K Akt mTOR pathway, inhibits the expression of TGF β and p-smad3, and improves stage IV renal fibrosis in DKD mice.255 Astragalus mongholicus Bunge and Panax notoginseng (Burkill) F.H. Chen Formula upregulated autophagy by inhibiting mTOR and activating the PINK1/Parkin signaling pathway;256 Icariin can restore autophagy by regulating the miR-192-5p/GLP-1R pathway,257 dasatinib and quercetin (DQ) can activate autophagy and Notch pathways to alleviate podocyte dedifferentiation.258 Geniposide can enhance ULK1-mediated autophagy,259 and the Dendrobium mixture can activate autophagy by inhibiting the PI3K/Akt/mTOR signaling pathway.260 Tripterygium glycoside (TG) upregulates autophagy through the mTOR/Twist1 pathway.261 TCM and their active chemical components protect DKD by activating autophagy. Novel therapies also have a similar effect. Intravenous injection of hucMSCs activates Autophagy through a para-secretory mode of action.262 The hUC-MSCs by reducing circulating TGF-β1 levels and restoring intracranial autophagy,263 the Placental Mesenchymal Stem Cells (MSCs) regulate mitochondrial autophagy by regulating the SIRT1-PGC-1alpha-TFAM pathway,264 and the Placenta-derived mesenchymal stem cells enhance podocyte autophagy by regulating the SIRT1/FOXO1 pathway.265 Human placental mesenchymal stem cells (hP-MSCs) derived Extracellular vesicles(EVs) can mitigate renal injury in DKD by modulating the miR-99b-5p/mTOR/autophagy pathway.266 Table 2 summarizes different interventions that delay the progression of DKD by regulating autophagy (Table 2).
 |
Table 2 Different Interventions Delayed the Progression of DKD by Regulating Autophagy |
Pyroptosis in Diabetic Kidney Disease
For the clinical transformation and application of pyroptosis, it is essential to deepen the theoretical knowledge of the pyroptosis pathway and develop targeted drugs.267 Start with It is triggered by a variety of inflammasomes When the inflammasome assembly is completed, caspases are activated, and gastrin proteins are cleaved to produce toxic fragments mediating cell membrane perforation.268 Pyroptosis is a major response to harmful damage such as pathogen ligands, abnormal levels of host metabolism, and environmental stimulation.269 As part of the innate immune system, the primary role of pyroptosis is to protect the host from pathogens.270 Under normal circumstances, moderate pyroptosis helps the host to defend against pathogen infection, but excessive pyroptosis can lead to uncontrolled inflammatory response, massive cell death, and serious tissue damage.271 For the clinical transformation and application of pyroptosis, deepening the theoretical knowledge of the pyroptosis pathway and developing targeted drugs267 is essential. Figure 3 shows that different factors regulate DKD through pyroptosis (Figure 3). Activation of caspase-1 induces maturation of IL-1β, and activation of caspase-3 mediates cleavage of GSDME (gasdermin E) and cytoplasmic release of mIL-1β, suggesting that GSDME has a pyrogenic effect and is upregulated in DKD model.272 GSDMD (gasdermin D) expression is positively correlated with tubular injury. In addition, DKD animal models have shown that TLR4 can aggravate tubular injury and fibrosis through the GSDMD-mediated pyrogen pathway.273 The non-classical pathway of caspase-11/GSDMD promotes scorification of glomerular endothelial cells (GECs) during DKD, thickening of glomerular basement membrane, extracellular matrix proliferation, and blurring of glomerular boundaries in DKD mice.274 Different factors induce DKD by participating in the pyroptosis pathway. miR-21-5p in macrophage-derived extracellular vesicles affects pyroptosis by regulating A20,275 circACTR2 is upregulated by participating in inflammation and pyroptosis pathways.276 Mir-6675p can also promote inflammatory response and cause pyroptosis.277 CircRNA COL1A2 mediates hyperglycemic-induced pyroptosis and OS by regulating the miR-424-5p/SGK1 (serum/glucocorticoid regulated kinase 1) axis.278 LncRNA NEAT1 mediates the miR-34c/NLRP3 axis to regulate caspase1 expression and promote cell pyroptosis.279 LncRNA-antisense noncoding RNA in the INK4 locus facilitated Caspase1-mediated oxidative destruction through the miR-497/thioredoxin-interacting protein axis.280 LncRNA MALAT1 promotes cell pyrogen via sponging miR-30c targeting NLRP3,281 CircLARP1B/miR-578/TLR4 axis inhibition blocks the cell cycle of G0-G1 phase and promotes cell pyrogen and the release of inflammatory factors.282 WTAP (WT1 associated protein) can promote the m6A methylation of NLRP3 mRNA and induce pyroptosis,283 TRIM29 (tripartite motif containing 29) can promote podocyte pyroptosis by activating the NF-κB /NLRP3 pathway.284 RBM15 (RNA binding motif protein 15), an m6A-related gene, can activate the AGE-RAGE pathway by promoting inflammation, OS, and pyroptosis.285 Alpha‐kinase 1 (ALPK1) /NF-κB pathway activates the caspase-1-GSDMD pyroptosis pathway,286 and ERS is involved in pyroptosis through the NF-κB/NLRP3 pathway.287 GECs indicate that charge-related pyroptosis is involved in neutrophil extracellular traps (NETs).288 The pRR promotes the pyroptosis of renal tubular epithelial cells through the dipeptidyl peptidase 4 (DPP4), which mediates signal transduction.289 Gsk-3β-hif-1α signaling pathway mediates pyroptosis.290 Lysophosphatidic Acid induces NLRP3 inflammatory activation by down-regulating EzH2/H3K27me3 and upregulating Egr1 expression, leading to endocytosis.291 Dihydroxyacetone phosphate (DHAP) leads to pyroptosis by regulating the mTORC1 pathway and promotes podiocyte injury.292 Discoid domain receptor 1 (DDR1) may promote pyroptosis through the NF-κB/NLRP3 pathway293 and induce the occurrence and development of DKD. Overexpression of miR-30e-5p or knockdown of ELAV-like RNA binding protein 1 (ELAVL1) can directly inhibit the pyro reaction,294 and down-regulation of KCNQ1OT1 can upregulate the expression of miR-506-3p. Inhibition of pyroptosis of HK-2 cells induced by high glucose295 and the down-regulation of ADAM metallopeptidase domain 10 can reduce inflammation, apoptosis, and pyroptosis by inhibiting AMPK signaling pathway,296 FOXM1 (forkhead box M1) activates SIRT4 through transcription and inhibits NF-κB/NLRP3. Alleviating podocyte pyroptosis,297 X inactive specific transcript (XIST) silences inhibit TLR4 by upregulation of miR-15b-5p and improve DKD kidney injury by inhibiting NLRP3/ caspase-1-mediated pyroptosis.298 Pyrroloquinoline quinone (PQQ) inhibited the pyroptosis pathway by alleviating mitochondrial dysfunction and reducing ROS production and NF-κB and caspase-1 expression.299 Hyperoside regulates extracellularly regulated protein kinases 1/2/mitogen-activated protein kinase signaling the pathway is used to inhibit OS, thereby alleviating the pyroptosis of renal tubular epithelial cells.300 Syringaresinol inhibits the NLRP3/caspase-1/GSDMD pyroptosis pathway through upregulating the Nrf2 signaling pathway.301 Hirudin regulates interferon regulatory factor 2 (Irf2) to inhibit GSDMD-mediated pyroptosis,302 Carnosine targeting Caspase-1-mediated pyroptosis,303 Sodium butyrate (NaB) regulates typical pyroptosis pathways of caspase-1/GSDMD,304 while Puerarin inhibits caspase-1-mediated pyroptosis.305 The GLP-1 analog liraglutide regulates NLRP3-induced inflammation and cell pyroptosis.306 VX-765 is a caspase-1 inhibitor and can interfere with DKD models in vivo and in vitro through cell pyroptosis.307 Dapagliflozin improved podocytosis through HO-1.308 Atorvastatin inhibit podocytosis through MALAT1/miR-200c/NRF2.309 Human Umbilical Cord Mesenchymal Stem Cells inhibited the pyroptosis of renal tubular epithelial cells under DKD conditions through miR-342-3p/ Caspase1 signaling pathway.310 TCM and active ingredients have also been shown to protect DKD by inhibiting the pathway of pyroptosis. Chuanxiong active components can improve the pyroptosis level under high glucose.311 Triptolide (TP) inhibits OS and pyroptosis through Nrf2/ROS/NLRP3 axis,312 and Astragaloside IV (AS) increased the expression of sirtuin 6 (SIRT6). The expression of hypoxia-inducible factor 1 subunit alpha (HIF-1a) was decreased, and ROS was inhibited from alleviating podocyte pyroptosis.313 Biochanin A regulates the pyroptosis cascade and NF-kB/NLRP3 axis of renal tubular epithelial cells314 and improve DKD renal dysfunction and renal injury. Table 3 summarizes different interventions that delay the progression of DKD by regulating pyroptosis (Table 3).
 |
Table 3 Different Intervention Methods Delayed the Progression of DKD by Regulating Pyroptosis |
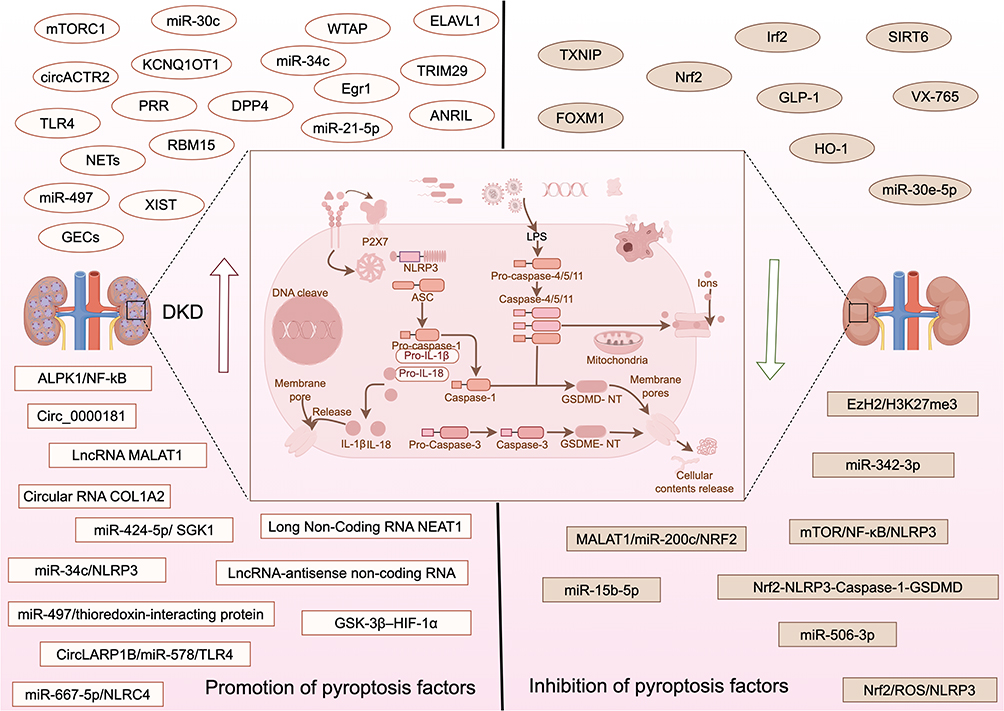 |
Figure 3 Shows that different factors regulate DKD through pyroptosis. By Figdraw. |
Ferroptosis in Diabetic Kidney Disease
The regulation of ferroptosis is closely related to iron metabolism disorder, lipid metabolism abnormality, oxidation, and antioxidant imbalance. Iron has two oxidation states: ferrous (Fe2+) and iron (Fe3+). The reductase of the kernel can reduce Fe3+ to Fe2+, and when Fe2+ is abnormally elevated, it will promote ferroptosis.315 The typical characteristic of ferroptosis was the inactivation of glutathione peroxidase 4 (GPX4) and the damage of cystine transport System XC- (xCT). These include the solute carrier family 7-member 11 (Slc7a11), increased cell membrane density, dysregulation of iron homeostasis, and accumulation of lipid peroxides, leading to mitochondrial atrophy.316–318 Proteinuria in DKD patients is related to the reabsorption of a large amount of transferrin into renal tubular epithelial cells and the degradation of lysozyme, resulting in excessive Fe2+ retention.319 Progressive iron overload can produce nephrotoxicity, which is manifested by the accumulation of Fe2+ in cells that catalyzes the production of many ROS through the Fenton reaction, and ROS can directly attack the phospholipids containing PUFAs (polyunsaturated fatty acids) in cell membranes, causing phospholipperoxidation and triggering ferroptosis in cells.320 Figure 4 shows that different factors regulate DKD through ferroptosis (Figure 4). Recent studies have found that iron overload exists in various DKD models, such as db/db, streptozotocin induction, and the ZSF1 model. In addition, the expression of ACSL4 (acyl-CoA synthetase long-chain family member 4), prostaglandin-endoperoxide synthase 2 (PTGS2), NCOA4 (nuclear receptor coactivator 4), and other ferroptosis markers were increased, while the expression of anti-lipid oxide GPX4 was decreased.321–324 In addition, GPX4 is significantly associated with proteinuria, ACR, serum creatinine (Scr), eGFR, and glomerular sclerosis rates, and patients with low GPX4 have a higher incidence of ESRD.325 A cross-sectional study showed that for T2DM patients, with the increase of urinary albumin excretion rate (UAER), the levels of GPX4, Fe, and Tf gradually decreased, while the levels of ACSL4 increased. At the same time, with the decrease in estimated glomerular filtration rate (eGFR), GPX4 and Tf levels gradually decreased, while ACSL4 levels increased, and UAER and ACSL4 were independently and positively correlated.326 Bioinformatic analysis of renal tubulointerstitial injury in DKD patients showed that D44, PTEN, ALOX5, and NCF2 were negatively correlated with eGFR, while VEGFA and DDIT3 were positively correlated with eGFR. These genes may be molecular mechanisms involved in the occurrence and development of DKD.327 A prospective observational study of 118 patients showed that the combined detection of GPX4, ACSL4, MDA, and ROS had good predictive value for DKD. In addition, ferritin level, serum iron, transferrin, and GPX4 were independently correlated with massive proteinuria.328 Compared with non-diabetic patients, the mRNA expressions of xCT and GPX4 in renal biopsy tissue of diabetic patients were decreased. Ferrostatin 1 (Fer-1) was found increased in the in vivo and in vitro DKD models with reduced lipid peroxidation, ferroptosis, and renal tissue injury.329
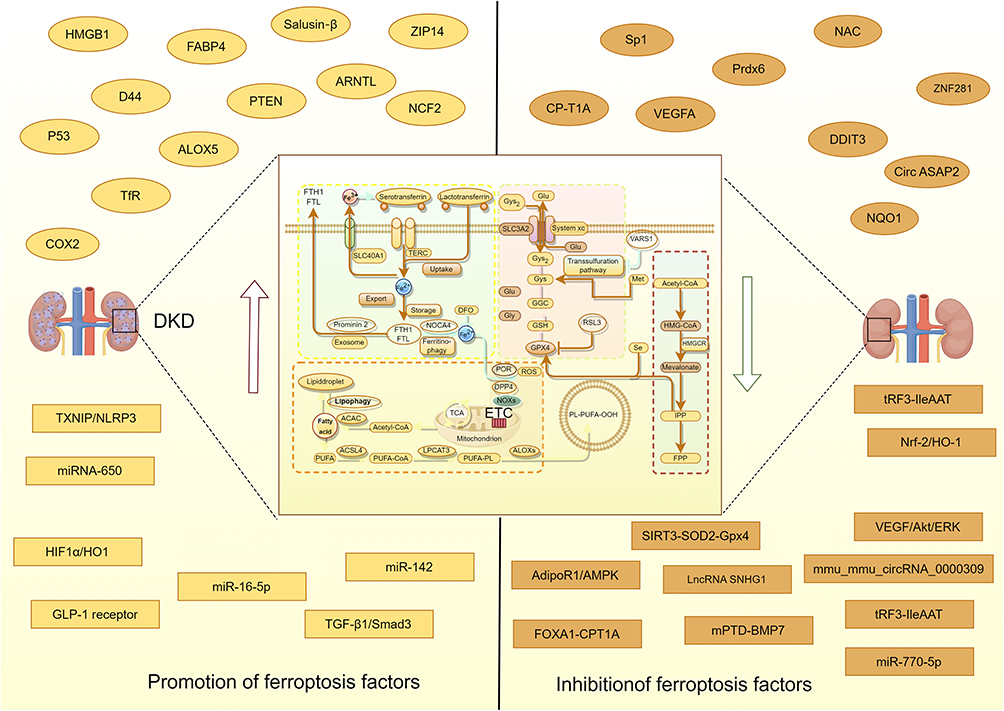 |
Figure 4 Shows that different factors regulate DKD through ferroptosis. By Figdraw. |
Diverse Factors Promote DKD Progression by Inducing Ferroptosis
Different factors promote the progression of DKD by inducing ferroptosis. HMGB1 regulates ferroptosis through the Nrf2 pathway,330 IME imbalance causes ferroptosis,331 and the expression of fatty acid binding protein 4 (FABP4) increases. Inhibit the expression of carnitine palmitoyltransferase-1A (CP-T1A), reduce superoxide dismutase, increase mitochondrial damage, and induce ferroptosis.332 ZRT/IRT-like protein 14 (ZIP14) is involved in iron accumulation and ferroptosis as a transporter.333 Salusin β is involved in ferroptosis in a Nrf 2 dependent manner,334 accelerating renal dysfunction and kidney injury in DKD.KAT2A promotes ferroptosis in DKD through H3K79succ-mediated upregulation of SAT2, thereby driving renal injury and inflammation.335 Different factors that inhibit ferroptosis have therapeutic effects on DKD. The mmu_mmu_circRNA_0000309, a circular RNA, regulates the ferroptosis pathway by regulating the miR-188-3p/GPX4 signaling pathway,336 LncRNA SNHG1 down-regulates the targeting of miR-16-5p/ACSL4 axis to inhibit ferroptosis.337 Circ ASAP2 inhibits inflammation and ferroptosis through SOX2/SLC7A11 mediated by miR-770-5p.338 Preupregulation of peroxiredoxin 6 (Prdx6) mediated by specificity protein 1 (Sp1) alleviated OS and ferroptosis pathway,339 while Vitexin decreased ROS, Fe2+ and MDA levels by activating GPX4.340 Down-regulation of Aryl hydrocarbon receptor nuclear translocator-like protein 1 (ARNTL) reduces OS and ferroptosis and alleviates mitochondrial morphological changes.341 N-acetylcysteine (NAC) activates the SIRT3-SOD2-Gpx4 signaling pathway to maintain mitochondrial REDOX homeostasis and reduce ferroptosis,342 GLP-1 drug somaluretide (SMG) further reduces renal inflammation and fibrosis by inhibiting iron death.343 The tRF3-IleAAT inhibits ferroptosis by targeting zinc finger protein 281 (ZNF281),344 and Umbelliferone inhibits ferroptosis by down-regulating ACSL4 and upregulating GPX4 by activating the Nrf-2/HO-1 pathway.345 Puerarin (PUR) inhibits excessive extracellular matrix (ECM) accumulation to prevent DKD.346 Protein transduction domain (PTD)-fused BMP7 in micelles (mPTD-BMP7) can accelerate the regeneration of the diabetic pancreas and inhibit ferroptosis,347 Aspirin supposes the activation of cyclooxygenase-2 (COX2), which is a potential target of ferroptosis and inhibits ferroptosis,348 Carnosine is a scavenger of iron ion and reactive oxygen species and acts as NrF2-mediated ferroptosis.349 Canagliflozin improves the fatty acid oxidation (FAO) and ferroptosis of renal tubular epithelial cells through the FOXA1-Carnitine palmitoyl transferase 1A (CPT1A) axis.350 Dapagliflozin improves DKD by inhibiting ferroptosis, promoting β-hydroxybutyrate (BHB) production, and regulating Ca2+/calmodulin-dependent protein kinase 2 (CaMKK2).351 Total flavones from Abelmoschus manihot (TFA) and daglizin can improve iron deposition, lipid peroxidation capacity, and expression of ferroptosis-related proteins, inhibit ferroptosis,352 and treat DKD.
TCM has been shown to have a regulatory effect on ferroptosis to treat DKD. Rosa laevigata Michx. inhibits ferroptosis.353 Natural botanical PDB (and its active constituents quercetin, kaempferol, β-sitosterol) inhibits ferroptosis in diabetic kidney disease by activating the Nrf2 signaling pathway, upregulating HO-1/GPX4/SLC7A11 and restoring glutathione homeostasis.354 Rhodiola decreased iron load and inhibited lipid peroxidation by inhibiting TFR1 expression.355,356 Leonurus reduces Fe2+ and ROS by upregulating GPX4 levels.357 Glycyrrhiza inhibits NCOA4 expression, upregulates GPX4 level, and mediates Slc7a11/SLC3A2 to down-regulate TFR1 protein expression.358 Quercetin inhibits ferroptosis in renal tubular epithelial cells by regulating the Nrf2/HO-1 signaling pathway to treat DKD.359 In addition, the sleeve gastrectomy (SG), a type of bariatric surgery, inhibits ferroptosis by inhibiting the TGF-β1/Smad3 signaling pathway, improves metabolic parameters and glucose homeostasis in DKD rats, reduces the impact of DKD on renal function indicators and tissue morphology, and reduces renal tubules injury.360 Table 4 summarizes different interventions that delay the progression of DKD by regulating ferroptosis (Table 4).
 |
Table 4 Different Interventions Delayed the Progression of DKD by Regulating Ferroptosis |
Other PCD in Diabetic Kidney Disease
Mitotic catastrophe (MC) causes the loss of podocytes in the urine of diabetic patients.361 The senescence of RTECs plays a crucial role in the progression of DKD, and insufficient mitophagy is closely related to RTEC senescence. Studies have shown that yeast mitochondrial escape 1-like 1 (YME1L) mediated senescence plays an anti-aging role in renal tubular cells in diabetic conditions and hinders the progression of DKD.362 Elevated circulating growth hormone levels (GH) are associated with diabetic podocyte damage and proteinuria. Studies have shown that GH induces mitotic mutations in glomerular podocytes, leading to proteinuria.363 Murine double minute 2 (MDM2) is a cell cycle regulator widely expressed in kidney cells, including podocytes.364,365 MDM2 is involved in high glucose-induced podocyte mitosis mutation through the Notch1 signaling pathway.366
In addition, several PCD modes have been discovered, including necroptosis, parthanatos, entotic cell death, mitotic cell death, lysosome-dependent cell death, alkalosis, and oxeiptosis. A microarray-based transcriptome analysis and single-core RNA sequencing analysis revealed that DKD is associated with the four core PCD pathways: entotic cell death, apoptosis, necroptosis, and pyroptosis. The WGCNA algorithm was further applied to screen four core death genes (CASP1, CYBB, PLA2G4A, and CTSS), and the CDS risk score was constructed based on these genes. The CDS risk score was more efficient in diagnosing DKD patients.367 There are still some unclear relationships between PCD and DKD, which may provide new ideas for further exploration of the pathogenesis of DKD.
Discussion
DKD progression is fundamentally driven by dysregulated PCD pathways, where apoptosis, autophagy, pyroptosis, and ferroptosis collectively exacerbate renal injury through distinct mechanisms. Apoptosis induces podocyte and tubular cell loss via mitochondrial dysfunction (eg, Bim/NFAT2) and caspase activation; impaired autophagy accelerates damage by accumulating toxic cellular debris, exhibiting dual roles (deficiency in PINK1/Parkin mitophagy vs detrimental overactivation); pyroptosis amplifies inflammation through NLRP3/GSDMD-driven gasdermin cleavage, though species-specific GSDME effects exist; while ferroptosis promotes iron-dependent lipid peroxidation (GPX4↓/ACSL4↑ correlating with proteinuria). Critically, these pathways exhibit extensive crosstalk (eg, NLRP3 pyroptosis → ferroptosis via ROS; autophagy defects → ERS-dependent apoptosis), forming an interconnected network that amplifies DKD pathogenesis.
Therapeutically, targeting PCD holds significant promise: Pharmacological agents (eg, SGLT2 inhibitors restoring autophagy flux, GLP-1 analogs suppressing pyroptosis), natural compounds (eg, berberine modulating ferroptosis via Nrf2), and RNA-based strategies (eg, miR-30e-5p inhibiting apoptosis/pyroptosis) demonstrate efficacy in preclinical models. However, key controversies impede translation: Compartmentalized PCD effects (GSDMD-tubular vs GSDME-glomerular), stage-dependent autophagy outcomes, and inconsistent ferroptosis biomarkers in human cohorts necessitate precision targeting. Clinically, interventions like combined enalapril/paricalcitol therapy synergistically attenuate multiple PCD pathways, improving proteinuria and glomerulosclerosis. Emerging approaches—including stem cell-derived extracellular vesicles and CRISPR-based gene editing—offer novel precision tools to reprogram PCD dynamics.
Nevertheless, key limitations persist. Mechanistic crosstalk between PCD modes remains incompletely mapped, necessitating multi-omics studies to decode signaling networks. Translational gaps are evident, as most evidence derives from animal models; validating urinary PCD biomarkers (eg, LC3B for autophagy, GPX4 for ferroptosis in human cohorts is imperative. Emerging PCD modes like necroptosis and parthanatos are underexplored in DKD, while clinical safety concerns—such as off-target effects of pan-caspase inhibitors (eg, Z-DEVD-FMK)—demand rigorous long-term assessment. Future research must prioritize combinatorial therapies, patient stratification based on PCD profiles, and human trials to translate these mechanistic insights into effective clinical strategies against DKD.
Conclusion
DKD progression is orchestrated by interconnected PCD pathways: apoptosis mediating cell loss, autophagy exhibiting dual protective/detrimental roles, pyroptosis driving inflammation, and ferroptosis accelerating lipid peroxidation. Their crosstalk creates an amplifying injury network. While therapeutic targeting shows promise, unresolved controversies—including stage-dependent effects, cell-type specificity, and biomarker gaps—demand patient-stratified combinatorial approaches for clinical translation.
Acknowledgments
Thank you Figdraw for your drawing assistance with the figures in this manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the National Natural Science Foundation of China (T2341018), 2015 Traditional Chinese Medicine Scientific Research (No.201507001-11); the National Natural Science Foundation of China under the Young Science Foundation Project (82305205), a multidisciplinary study on the treatment of obesity type 2 diabetes from the perspective of Tuyong (CI2023C049YLL); the Clinical Research Center Construction Project of Guang’anmen Hospital, CACMS (Grant No.2022LYJSZX01~2022LYJSZX29).
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Ruiz-Ortega M, Rodrigues-Diez RR, Lavoz C, Rayego-Mateos S. Special Issue “Diabetic nephropathy: diagnosis, prevention and treatment”. J Clin Med. 2020;9(3):813. doi:10.3390/jcm9030813
2. Schwarz P. IDF global clinical practice recommendations for managing type 2 diabetes - 2025. Diabet Res Clin Pract. 2025;222(Suppl 1):112158. doi:10.1016/j.diabres.2025.112158
3. IDF Virtual Congress 2023. Diabet Res Clin Pract. 2023;198:110632. doi:10.1016/j.diabres.2023.110632
4. Tummers B, Green DR. The evolution of regulated cell death pathways in animals and their evasion by pathogens. Physiol Rev. 2022;102(1):411–454. doi:10.1152/physrev.00002.2021
5. Wang H, Zhou X, Li C, et al. The emerging role of pyroptosis in pediatric cancers: from mechanism to therapy. J Hematol Oncol. 2022;15(1):140. doi:10.1186/s13045-022-01365-6
6. Galluzzi L, Vitale I, Aaronson SA, et al. Molecular mechanisms of cell death: recommendations of the nomenclature committee on cell death 2018. Cell Death Differ. 2018;25(3):486–541. doi:10.1038/s41418-017-0012-4
7. Zeng Q, Ma X, Song Y, Chen Q, Jiao Q, Zhou L. Targeting regulated cell death in tumor nanomedicines. Theranostics. 2022;12(2):817–841. doi:10.7150/thno.67932
8. Koren E, Fuchs Y. Modes of regulated cell death in cancer. Cancer Discov. 2021;11(2):245–265. doi:10.1158/2159-8290.CD-20-0789
9. Hotchkiss RS, Strasser A, McDunn JE, Swanson PE. Cell death. N Engl J Med. 2009;361(16):1570–1583. doi:10.1056/NEJMra0901217
10. Loftus LV, Amend SR, Pienta KJ. Interplay between cell death and cell proliferation reveals new strategies for cancer therapy. Int J Mol Sci. 2022;23(9):4723. doi:10.3390/ijms23094723
11. Garg JP, Vucic D. Targeting cell death pathways for therapeutic intervention in kidney diseases. Semin Nephrol. 2016;36(3):153–161. doi:10.1016/j.semnephrol.2016.03.003
12. Fuchs Y, Steller H. Programmed cell death in animal development and disease. Cell. 2011;147(4):742–758. doi:10.1016/j.cell.2011.10.033
13. Yuan J, Ofengeim D. A guide to cell death pathways. Nat Rev Mol Cell Biol. 2024;25(5):379–395. doi:10.1038/s41580-023-00689-6
14. Bedoui S, Herold MJ, Strasser A. Emerging connectivity of programmed cell death pathways and its physiological implications. Nat Rev Mol Cell Biol. 2020;21(11):678–695. doi:10.1038/s41580-020-0270-8
15. Sanz AB, Sanchez-Niño MD, Ramos AM, Ortiz A. Regulated cell death pathways in kidney disease. Nat Rev Nephrol. 2023;19(5):281–299. doi:10.1038/s41581-023-00694-0
16. Laustsen C, Nielsen PM, Qi H, Løbner MH, Palmfeldt J, Bertelsen LB. Hyperpolarized [1,4-13C]fumarate imaging detects microvascular complications and hypoxia mediated cell death in diabetic nephropathy. Sci Rep. 2020;10(1):9650. doi:10.1038/s41598-020-66265-6
17. Rodríguez-González J, Gutiérrez-Kobeh L. Apoptosis and its pathways as targets for intracellular pathogens to persist in cells. Parasitol Res. 2023;123(1):60. doi:10.1007/s00436-023-08031-x
18. Tagaya M, Kume S, Yasuda-Yamahara M, et al. Inhibition of mitochondrial fission protects podocytes from albumin-induced cell damage in diabetic kidney disease. Biochim Biophys Acta Mol Basis Dis. 2022;1868(5):166368. doi:10.1016/j.bbadis.2022.166368
19. Kengkoom K, Angkhasirisap W, Kanjanapruthipong T, et al. Streptozotocin induces alpha-2u globulin nephropathy in male rats during diabetic kidney disease. BMC Vet Res. 2021;17(1):105. doi:10.1186/s12917-021-02814-z
20. Zhang RD, Shi M. Occurrence and development of diabetic nephropathy caused by CD63 by inhibiting Wnt-β-catenin signaling pathway. Eur Rev Med Pharmacol Sci. 2020;24(1):284–294. doi:10.26355/eurrev_202001_19923
21. Hou Y, Wang Q, Han B, Chen Y, Qiao X, Wang L. CD36 promotes NLRP3 inflammasome activation via the mtROS pathway in renal tubular epithelial cells of diabetic kidneys. Cell Death Dis. 2021;12(6):523. doi:10.1038/s41419-021-03813-6
22. Jiang WJ, Xu CT, Du CL, et al. Tubular epithelial cell-to-macrophage communication forms a negative feedback loop via extracellular vesicle transfer to promote renal inflammation and apoptosis in diabetic nephropathy. Theranostics. 2022;12(1):324–339. doi:10.7150/thno.63735
23. Casalena GA, Yu L, Gil R, et al. The diabetic microenvironment causes mitochondrial oxidative stress in glomerular endothelial cells and pathological crosstalk with podocytes. Cell Commun Signal. 2020;18(1):105. doi:10.1186/s12964-020-00605-x
24. Li L, Feng Y, Zhang J, et al. Microtubule associated protein 4 phosphorylation-induced epithelial-to-mesenchymal transition of podocyte leads to proteinuria in diabetic nephropathy. Cell Commun Signal. 2022;20(1):115. doi:10.1186/s12964-022-00883-7
25. Guo W, Gao H, Pan W, Yu P, Che G. High glucose induces Nox4 expression and podocyte apoptosis through the Smad3/ezrin/PKA pathway. Biol Open. 2021;10(5):bio055012. doi:10.1242/bio.055012
26. Shi W, Huang Y, Zhao X, et al. Histone deacetylase 4 mediates high glucose-induced podocyte apoptosis via upregulation of calcineurin. Biochem Biophys Res Commun. 2020;533(4):1061–1068. doi:10.1016/j.bbrc.2020.09.121
27. Xu C, Zhou X, Xie T, et al. Renal tubular Bim mediates the tubule-podocyte crosstalk via NFAT2 to induce podocyte cytoskeletal dysfunction. Theranostics. 2020;10(15):6806–6824. doi:10.7150/thno.43145
28. Jing Z, Hu L, Su Y, Ying G, Ma C, Wei J. Potential signaling pathway through which Notch regulates oxidative damage and apoptosis in renal tubular epithelial cells induced by high glucose. J Recept Signal Transduction Res. 2021;41(4):357–362. doi:10.1080/10799893.2020.1810706
29. Gu X, Liu Y, Wang N, et al. Transcription of MRPL12 regulated by Nrf2 contributes to the mitochondrial dysfunction in diabetic kidney disease. Free Radic Biol Med. 2021;164:329–340. doi:10.1016/j.freeradbiomed.2021.01.004
30. Zhang J, Wu Y, Zhang J, Zhang R, Wang Y, Liu F. ABCA1 deficiency-mediated glomerular cholesterol accumulation exacerbates glomerular endothelial injury and dysfunction in diabetic kidney disease. Metabolism. 2023;139:155377. doi:10.1016/j.metabol.2022.155377
31. Chen Z, Ma Y, Yang Q, et al. AKAP1 mediates high glucose-induced mitochondrial fission through the phosphorylation of Drp1 in podocytes. J Cell Physiol. 2020;235(10):7433–7448. doi:10.1002/jcp.29646
32. Zhang Y, Xu C, Ye Q, et al. Podocyte apoptosis in diabetic nephropathy by BASP1 activation of the p53 pathway via WT1. Acta Physiol. 2021;232(1):e13634. doi:10.1111/apha.13634
33. Li L, Chen X, Zhang H, Wang M, Lu W. MRC2 Promotes Proliferation and Inhibits Apoptosis of Diabetic Nephropathy. Anal Cell Pathol. 2021;2021:6619870. doi:10.1155/2021/6619870
34. Wen H, Kumar V, Mishra A, et al. Grem2 mediates podocyte apoptosis in high glucose milieu. Biochimie. 2019;160:113–121. doi:10.1016/j.biochi.2019.02.015
35. Lan X, Kumar V, Jha A, et al. EDA2R mediates podocyte injury in high glucose milieu. Biochimie. 2020;174:74–83. doi:10.1016/j.biochi.2020.04.003
36. Chen J, Chen KH, Wang LM, Luo J, Zheng QY, He YN. Decoy receptor 2 mediates the apoptosis-resistant phenotype of senescent renal tubular cells and accelerates renal fibrosis in diabetic nephropathy. Cell Death Dis. 2022;13(6):522. doi:10.1038/s41419-022-04972-w
37. Li C, Matavelli LC, Akhtar S, Siragy HM. (Pro)renin receptor contributes to renal mitochondria dysfunction, apoptosis and fibrosis in diabetic mice. Sci Rep. 2019;9(1):11667. doi:10.1038/s41598-019-47055-1
38. Jiang S, Xu CM, Yao S, et al. Cdc42 upregulation under high glucose induces podocyte apoptosis and impairs β-cell insulin secretion. Front Endocrinol. 2022;13:905703. doi:10.3389/fendo.2022.905703
39. Lora Gil C, Henley N, Leblond FA, et al. Alk1 haploinsufficiency causes glomerular dysfunction and microalbuminuria in diabetic mice. Sci Rep. 2020;10(1):13136. doi:10.1038/s41598-020-68515-z
40. Zhang Y, Chen X, Fan Y, Liu J, Yuan L. XCL1 aggravates diabetic nephropathy-mediated renal glomerular endothelial cell apoptosis and inflammatory response via regulating p53/Nuclear Factor-Kappa B pathway. Nephron. 2022;146(1):84–98. doi:10.1159/000518172
41. Mathur A, Pandey VK, Khan MF, Kakkar P. PHLPP1/Nrf2-Mdm2 axis induces renal apoptosis via influencing nucleo-cytoplasmic shuttling of FoxO1 during diabetic nephropathy. Mol Cell Biochem. 2021;476(10):3681–3699. doi:10.1007/s11010-021-04177-3
42. Matoba K, Takeda Y, Nagai Y, et al. ROCK2-induced metabolic rewiring in diabetic podocytopathy. Commun Biol. 2022;5(1):341. doi:10.1038/s42003-022-03300-4
43. Lin Q, Ma Y, Chen Z, et al. Sestrin-2 regulates podocyte mitochondrial dysfunction and apoptosis under high-glucose conditions via AMPK. Int J Mol Med. 2020;45(5):1361–1372. doi:10.3892/ijmm.2020.4508
44. Sharma I, Deng F, Liao Y, Kanwar YS. Myo-inositol Oxygenase (MIOX) overexpression drives the progression of renal tubulointerstitial injury in diabetes. Diabetes. 2020;69(6):1248–1263. doi:10.2337/db19-0935
45. Zheng Y, Zhang Z, Zheng D, Yi P, Wang S. METTL14 promotes the development of diabetic kidney disease by regulating m6A modification of TUG1. Acta Diabetol. 2023;60(11):1567–1580. doi:10.1007/s00592-023-02145-5
46. Liu Y, Zhang L, Zhang S, et al. ATF5 regulates tubulointerstitial injury in diabetic kidney disease via mitochondrial unfolded protein response. Mol Med. 2023;29(1):57. doi:10.1186/s10020-023-00651-4
47. Xue M, Fang T, Sun H, et al. PACS-2 attenuates diabetic kidney disease via the enhancement of mitochondria-associated endoplasmic reticulum membrane formation. Cell Death Dis. 2021;12(12):1107. doi:10.1038/s41419-021-04408-x
48. Chen N, Song S, Yang Z, et al. ChREBP deficiency alleviates apoptosis by inhibiting TXNIP/oxidative stress in diabetic nephropathy. J Diabetes Complications. 2021;35(12):108050. doi:10.1016/j.jdiacomp.2021.108050
49. Li M, Deng L, Xu G. METTL14 promotes glomerular endothelial cell injury and diabetic nephropathy via m6A modification of α-klotho. Mol Med. 2021;27(1):106. doi:10.1186/s10020-021-00365-5
50. Hu H, Li W, Liu M, et al. C1q/tumor necrosis factor-related Protein-9 attenuates diabetic nephropathy and kidney fibrosis in db/db mice. DNA Cell Biol. 2020;39(6):938–948. doi:10.1089/dna.2019.5302
51. Qiu D, Song S, Wang Y, et al. NAD(P)H: quinone oxidoreductase 1 attenuates oxidative stress and apoptosis by regulating Sirt1 in diabetic nephropathy. J Transl Med. 2022;20(1):44. doi:10.1186/s12967-021-03197-3
52. Meng Q, Tian X, Li J, et al. GTS-21, a selective alpha7 nicotinic acetylcholine receptor agonist, ameliorates diabetic nephropathy in Leprdb/db mice. Sci Rep. 2022;12(1):22360. doi:10.1038/s41598-022-27015-y
53. Min XQ, Xie Y. LncRNA CASC2 alleviates the progression of diabetic nephropathy by regulating the miR-144/SOCS2 signalling axis. Kidney Blood Press Res. 2020;45(6):837–849. doi:10.1159/000508078
54. Shao X, Zhang X, Hu J, et al. Dopamine 1 receptor activation protects mouse diabetic podocytes injury via regulating the PKA/NOX-5/p38 MAPK axis. Exp Cell Res. 2020;388(2):111849. doi:10.1016/j.yexcr.2020.111849
55. Song S, Qiu D, Wang Y, et al. TXNIP deficiency mitigates podocyte apoptosis via restraining the activation of mTOR or p38 MAPK signaling in diabetic nephropathy. Exp Cell Res. 2020;388(2):111862. doi:10.1016/j.yexcr.2020.111862
56. Chen Y, Yan R, Li B, et al. Silencing CCNG1 protects MPC-5 cells from high glucose-induced proliferation-inhibition and apoptosis-promotion via MDM2/p53 signaling pathway. Int Urol Nephrol. 2020;52(3):581–593. doi:10.1007/s11255-020-02383-4
57. Wang X, Liu Q, Kong D, et al. Down-regulation of SETD6 protects podocyte against high glucose and palmitic acid-induced apoptosis, and mitochondrial dysfunction via activating Nrf2-Keap1 signaling pathway in diabetic nephropathy. J Mol Histol. 2020;51(5):549–558. doi:10.1007/s10735-020-09904-6
58. Hongbo M, Yanjiao D, Shuo W, Kun S, Yanjie L, Mengmeng L. Podocyte RNF166 deficiency alleviates diabetic nephropathy by mitigating mitochondria impairment and apoptosis via regulation of CYLD signal. Biochem Biophys Res Commun. 2021;545:46–53. doi:10.1016/j.bbrc.2020.12.014
59. Zhang X, Guan T, Yang B, Chi Z, Wan Q, Gu HF. SLC30A7 has anti-oxidant stress effects in high glucose-induced apoptosis via the NFE2L2/HMOX1 signal transductionpathway. Diabet Res Clin Pract. 2021;172:108445. doi:10.1016/j.diabres.2020.108445
60. He L, Li Q, Du C, Xue Y, Yu P. Glis2 inhibits the epithelial-mesenchymal transition and apoptosis of renal tubule cells by regulating the β-catenin signalling pathway in diabetic kidney disease. Biochem Biophys Res Commun. 2022;607:73–80. doi:10.1016/j.bbrc.2022.03.111
61. Jiang N, Zhao H, Han Y, et al. HIF-1α ameliorates tubular injury in diabetic nephropathy via HO-1-mediated control of mitochondrial dynamics. Cell Prolif. 2020;53(11):e12909. doi:10.1111/cpr.12909
62. Ye G, Hu ML, Xiao L. Forkhead box A2-mediated lncRNA SOX2OT up-regulation alleviates oxidative stress and apoptosis of renal tubular epithelial cells by promoting SIRT1 expression in diabetic nephropathy. Nephrology. 2023;28(3):196–207. doi:10.1111/nep.14139
63. Sun ZJ, Chang DY, Chen M, Zhao MH. Deficiency of CFB attenuates renal tubulointerstitial damage by inhibiting ceramide synthesis in diabetic kidney disease. JCI Insight. 2022;7(24):e156748. doi:10.1172/jci.insight.156748
64. Zhu J, Zheng X. Clinical value of INSL3 in the diagnosis and development of diabetic nephropathy. J Clin Lab Anal. 2021;35(9):e23898. doi:10.1002/jcla.23898
65. Sun HJ, Xiong SP, Cao X, et al. Polysulfide-mediated sulfhydration of SIRT1 prevents diabetic nephropathy by suppressing phosphorylation and acetylation of p65 NF-κB and STAT3. Redox Biol. 2021;38:101813. doi:10.1016/j.redox.2020.101813
66. Huang F, Wang Q, Guo F, et al. FoxO1-mediated inhibition of STAT1 alleviates tubulointerstitial fibrosis and tubule apoptosis in diabetic kidney disease. EBioMedicine. 2019;48:491–504. doi:10.1016/j.ebiom.2019.09.002
67. Lin Q, Chen O, Wise JP, et al. FGF1ΔHBS delays the progression of diabetic nephropathy in late-stage type 2 diabetes mouse model by alleviating renal inflammation, fibrosis, and apoptosis. Biochim Biophys Acta Mol Basis Dis. 2022;1868(8):166414. doi:10.1016/j.bbadis.2022.166414
68. Liu L, Zhang L, Zhao J, et al. Tumor necrosis factor receptor-associated protein 1 protects against mitochondrial injury by preventing high glucose-induced mPTP Opening in diabetes. Oxid Med Cell Longev. 2020;2020:6431517. doi:10.1155/2020/6431517
69. Liu B, Zhang L, Yang H, Chen X, Zheng H, Liao X. SIK2 protects against renal tubular injury and the progression of diabetic kidney disease. Transl Res. 2023;253:16–30. doi:10.1016/j.trsl.2022.08.012
70. Hwang SD, Song JH, Kim Y, et al. Inhibition of lymphatic proliferation by the selective VEGFR-3 inhibitor SAR131675 ameliorates diabetic nephropathy in db/db mice. Cell Death Dis. 2019;10(3):219. doi:10.1038/s41419-019-1436-1
71. Yang M, Zhao L, Gao P, et al. DsbA-L ameliorates high glucose induced tubular damage through maintaining MAM integrity. EBioMedicine. 2019;43:607–619. doi:10.1016/j.ebiom.2019.04.044
72. Liu C, Li Y, Wang X. TDAG51-Deficiency podocytes are protected from high-glucose-induced damage through Nrf2 activation via the AKT-GSK-3β Pathway. Inflammation. 2022;45(4):1520–1533. doi:10.1007/s10753-022-01638-9
73. Li H, Zhao K, Li Y. Gasdermin D protects mouse podocytes against high-glucose-induced inflammation and apoptosis via the C-Jun N-Terminal Kinase (JNK) pathway. Med Sci Monit. 2021;27:e928411. doi:10.12659/MSM.928411
74. Wang X, Yang J, Wang W, Li Y, Yang Y. Decreasing REDD1 expression protects against high glucose-induced apoptosis, oxidative stress and inflammatory injury in podocytes through regulation of the AKT/GSK-3β/Nrf2 pathway. Immunopharmacol Immunotoxicol. 2023;45(5):527–538. doi:10.1080/08923973.2023.2183351
75. Chen Z, Tian L, Wang L, et al. TRIM32 inhibition attenuates apoptosis, oxidative stress, and inflammatory injury in podocytes induced by high glucose by modulating the Akt/GSK-3β/Nrf2 pathway. Inflammation. 2022;45(3):992–1006. doi:10.1007/s10753-021-01597-7
76. Jin J, Gong J, Zhao L, Zhang H, He Q, Jiang X. Inhibition of high mobility group box 1 (HMGB1) attenuates podocyte apoptosis and epithelial-mesenchymal transition by regulating autophagy flux. J Diabetes. 2019;11(10):826–836. doi:10.1111/1753-0407.12914
77. Chi Y, Zhang X, Liang D, et al. ZnT8 exerts anti-apoptosis of kidney tubular epithelial cell in diabetic kidney disease through TNFAIP3-NF-κB signal pathways. Biol Trace Elem Res. 2023;201(5):2442–2457. doi:10.1007/s12011-022-03361-w
78. Loretelli C, Rocchio F, D’Addio F, et al. The IL-8-CXCR1/2 axis contributes to diabetic kidney disease. Metabolism. 2021;121:154804. doi:10.1016/j.metabol.2021.154804
79. Hasegawa K, Sakamaki Y, Tamaki M, Wakino S. PCK1 protects against mitoribosomal defects in diabetic nephropathy in mouse models. J Am Soc Nephrol. 2023;34(8):1343–1365. doi:10.1681/ASN.0000000000000156
80. Sooshtari P, Feng B, Biswas S, et al. ANRIL regulates multiple molecules of pathogenetic significance in diabetic nephropathy. PLoS One. 2022;17(8):e0270287. doi:10.1371/journal.pone.0270287
81. Guan G, Xie J, Dai Y, Han H. TFPI2 suppresses the interaction of TGF-β2 pathway regulators to promote endothelial-mesenchymal transition in diabetic nephropathy. J Biol Chem. 2022;298(3):101725. doi:10.1016/j.jbc.2022.101725
82. González-Mariscal I, Carmona-Hidalgo B, Winkler M, et al. (+)-trans-Cannabidiol-2-hydroxy pentyl is a dual CB1R antagonist/CB2R agonist that prevents diabetic nephropathy in mice. Pharmacol Res. 2021;169:105492.
83. Zapała B, Kamińska A, Piwowar M, Paziewska A, Gala-Błądzińska A, Stępień EŁ. miRNA signature of urine extracellular vesicles shows the involvement of inflammatory and apoptotic processes in diabetic chronic kidney disease. Pharm Res. 2023;40(4):817–832. doi:10.1007/s11095-023-03481-5
84. Xue X, Liu M, Wang Y, et al. MicroRNA-494-3p exacerbates renal epithelial cell dysfunction by targeting SOCS6 under high glucose treatment. Kidney Blood Press Res. 2022;47(4):247–255. doi:10.1159/000521647
85. Wu L, Wang Q, Guo F, et al. Involvement of miR-27a-3p in diabetic nephropathy via affecting renal fibrosis, mitochondrial dysfunction, and endoplasmic reticulum stress. J Cell Physiol. 2021;236(2):1454–1468.
86. Chen H, Guo Y, Cheng X. Long non-cording RNA XIST promoted cell proliferation and suppressed apoptosis by miR-423-5p/HMGA2 axis in diabetic nephropathy. Mol Cell Biochem. 2021;476(12):4517–4528. doi:10.1007/s11010-021-04250-x
87. Ming L, Ning J, Ge Y, Zhang Y, Ruan Z. Excessive apoptosis of podocytes caused by dysregulation of microRNA-182-5p and CD2AP confers to an increased risk of diabetic nephropathy. J Cell Biochem. 2019;120(10):16516–16523. doi:10.1002/jcb.28911
88. Wang L, Li H. MiR-770-5p facilitates podocyte apoptosis and inflammation in diabetic nephropathy by targeting TIMP3. Biosci Rep. 2020;40(4):BSR20193653. doi:10.1042/BSR20193653
89. Liu F, Huang J, Zhang C, et al. Regulation of podocyte injury by CircHIPK3/FUS complex in diabetic kidney disease. Int J Biol Sci. 2022;18(15):5624–5640. doi:10.7150/ijbs.75994
90. Yu Q, Lin J, Ma Q, et al. Long Noncoding RNA ENSG00000254693 promotes diabetic kidney disease via interacting with HuR. J Diabetes Res. 2022;2022:8679548. doi:10.1155/2022/8679548
91. Song P, Chen Y, Liu Z, et al. LncRNA MALAT1 aggravates renal tubular injury via activating LIN28A and the Nox4/AMPK/mTOR signaling axis in diabetic nephropathy. Front Endocrinol. 2022;13:895360. doi:10.3389/fendo.2022.895360
92. Yang Z, Song D, Wang Y, Tang L. lncRNA MALAT1 promotes diabetic nephropathy progression via miR-15b-5p/TLR4 signaling axis. J Immunol Res. 2022;2022:8098001. doi:10.1155/2022/8098001
93. Wu X, Fan D, Chen B. LncRNA NEAT1 accelerates the proliferation, oxidative stress, inflammation, and fibrosis and suppresses the apoptosis through the miR-423-5p/GLIPR2 axis in diabetic nephropathy. J Cardiovasc Pharmacol. 2022;79(3):342–354. doi:10.1097/FJC.0000000000001177
94. Bai Y, Li H, Dong J. Up-regulation of miR-20a weakens inflammation and apoptosis in high-glucose-induced renal tubular cell mediating diabetic kidney disease by repressing CXCL8 expression. Arch Physiol Biochem. 2022;128(6):1603–1610. doi:10.1080/13813455.2020.1785506
95. Tsai YC, Kuo MC, Hung WW, et al. High glucose induces mesangial cell apoptosis through miR-15b-5p and promotes diabetic nephropathy by extracellular vesicle delivery. Mol Ther. 2020;28(3):963–974. doi:10.1016/j.ymthe.2020.01.014
96. Fei B, Zhou H, He Z, Wang S. KCNQ1OT1 inhibition alleviates high glucose-induced podocyte injury by adsorbing miR-23b-3p and regulating Sema3A. Clin Exp Nephrol. 2022;26(5):385–397. doi:10.1007/s10157-021-02173-x
97. Liu Q, Cui Y, Ding N, Zhou C. Knockdown of circ_0003928 ameliorates high glucose-induced dysfunction of human tubular epithelial cells through the miR-506-3p/HDAC4 pathway in diabetic nephropathy. Eur J Med Res. 2022;27(1):55. doi:10.1186/s40001-022-00679-y
98. Zhang YL, Wang JM, Yin H, Wang SB, He CL, Liu J. DACH1, a novel target of miR-218, participates in the regulation of cell viability, apoptosis, inflammatory response, and epithelial-mesenchymal transition process in renal tubule cells treated by high-glucose. Ren Fail. 2020;42(1):463–473. doi:10.1080/0886022X.2020.1762647
99. Wang Y, Qi Y, Ji T, et al. Circ_LARP4 regulates high glucose-induced cell proliferation, apoptosis, and fibrosis in mouse mesangial cells. Gene. 2021;765:145114. doi:10.1016/j.gene.2020.145114
100. Liu DW, Zhang JH, Liu FX, et al. Silencing of long noncoding RNA PVT1 inhibits podocyte damage and apoptosis in diabetic nephropathy by upregulating FOXA1. Exp Mol Med. 2019;51(8):1–15. doi:10.1038/s12276-019-0259-6
101. Wang J, Yang S, Li W, Zhao M, Li K. Circ_0000491 promotes apoptosis, inflammation, oxidative stress, and fibrosis in high glucose-induced mesangial cells by regulating miR-455-3p/Hmgb1 axis. Nephron. 2022;146(1):72–83. doi:10.1159/000516870
102. Zhu FX, Wu HL, Chen JX, Han B, Guo YF. Dysregulation of microRNA-181b and TIMP3 is functionally involved in the pathogenesis of diabetic nephropathy. J Cell Physiol. 2019;234(10):18963–18969. doi:10.1002/jcp.28536
103. Jing F, Zhao J, Jing X, Lei G. Long noncoding RNA Airn protects podocytes from diabetic nephropathy lesions via binding to Igf2bp2 and facilitating translation of Igf2 and Lamb2. Cell Biol Int. 2020;44(9):1860–1869. doi:10.1002/cbin.11392
104. Meng D, Wu L, Li Z, et al. LncRNA TUG1 ameliorates diabetic nephropathy via inhibition of PU.1/RTN1 signaling pathway. J Leukoc Biol. 2022;111(3):553–562. doi:10.1002/JLB.6A1020-699RRR
105. Wang W, Lu H. High glucose-induced human kidney cell apoptosis and inflammatory injury are alleviated by Circ_0008529 knockdown via Circ_0008529-mediated miR-485-5p/WNT2B signaling. Appl Biochem Biotechnol. 2022;194(12):6287–6301. doi:10.1007/s12010-022-04088-z
106. Liu L, Chen H, Yun J, et al. miRNA-483-5p Targets HDCA4 to Regulate Renal Tubular Damage in Diabetic Nephropathy. Horm Metab Res. 2021;53(8):562–569. doi:10.1055/a-1480-7519
107. Jiang ZH, Tang YZ, Song HN, Yang M, Li B, Ni CL. miRNA-342 suppresses renal interstitial fibrosis in diabetic nephropathy by targeting SOX6. Int J Mol Med. 2020;45(1):45–52. doi:10.3892/ijmm.2019.4388
108. Lv N, Li C, Liu X, Qi C, Wang Z. miR-34b alleviates high glucose-induced inflammation and apoptosis in human HK-2 cells via IL-6R/JAK2/STAT3 signaling pathway. Med Sci Monit. 2019;25:8142–8151. doi:10.12659/MSM.917128
109. Wang H, Zhang Y, Liu H, Li S. GDF11, a target of miR-32-5p, suppresses high-glucose-induced mitochondrial dysfunction and apoptosis in HK-2 cells through PI3K/AKT signaling activation. Int Urol Nephrol. 2023;55(7):1767–1778. doi:10.1007/s11255-023-03495-3
110. Fan X, Hao Z, Li Z, Wang X, Wang J. Inhibition of miR-17~92 cluster ameliorates high glucose-induced podocyte damage. Mediators Inflamm. 2020;2020:6126490. doi:10.1155/2020/6126490
111. Zhang W, Zhang L, Dong Q, Wang X, Li Z, Wang Q. Hsa_circ_0003928 regulates the progression of diabetic nephropathy through miR-136-5p/PAQR3 axis. J Endocrinol Invest. 2023;46(10):2103–2114. doi:10.1007/s40618-023-02061-z
112. Zhao Y, Li D, Zhou P, Zhao Y, Kuang J. microRNA-29b-3p attenuates diabetic nephropathy in mice by modifying EZH2. Hormones. 2023;22(2):223–233. doi:10.1007/s42000-022-00426-2
113. Yu R, Zhang Y, Lu Z, Li J, Shi P, Li J. Long-chain non-coding RNA UCA1 inhibits renal tubular epithelial cell apoptosis by targeting microRNA-206 in diabetic nephropathy. Arch Physiol Biochem. 2022;128(1):231–239. doi:10.1080/13813455.2019.1673431
114. Zhao L, Chen H, Zeng Y, et al. Circular RNA circ_0000712 regulates high glucose-induced apoptosis, inflammation, oxidative stress, and fibrosis in (DN) by targeting the miR-879-5p/SOX6 axis. Endocr J. 2021;68(10):1155–1164. doi:10.1507/endocrj.EJ20-0739
115. Zhao N, Luo Q, Lin R, Li Q, Ma P. MiR-142-3p ameliorates high glucose-induced renal tubular epithelial cell injury by targeting BOD1. Clin Exp Nephrol. 2021;25(11):1182–1192. doi:10.1007/s10157-021-02102-y
116. Wang S, Sun K, Hu H, et al. MiR-1297 attenuates high glucose-induced injury in HK-2 cells via targeting COL1A2. Nephrology. 2021;26(7):623–631. doi:10.1111/nep.13881
117. Yun J, Ren J, Liu Y, et al. MicroRNA (miR)-590-3p alleviates high-glucose induced renal tubular epithelial cell damage by targeting C-X3-C motif chemokine ligand 1 (CX3CL1) in diabetic nephropathy. Bioengineered. 2022;13(1):634–644. doi:10.1080/21655979.2021.2012548
118. Lou Z, Li Q, Wang C, Li Y. The effects of microRNA-126 reduced inflammation and apoptosis of diabetic nephropathy through PI3K/AKT signalling pathway by VEGF. Arch Physiol Biochem. 2022;128(5):1265–1274. doi:10.1080/13813455.2020.1767146
119. Elkazzaz SK, Khodeer DM, El Fayoumi HM, Moustafa YM. Role of sodium glucose cotransporter type 2 inhibitors dapagliflozin on diabetic nephropathy in rats; Inflammation, angiogenesis and apoptosis. Life Sci. 2021;280:119018. doi:10.1016/j.lfs.2021.119018
120. Shibusawa R, Yamada E, Okada S, et al. Dapagliflozin rescues endoplasmic reticulum stress-mediated cell death. Sci Rep. 2019;9(1):9887. doi:10.1038/s41598-019-46402-6
121. Lee WC, Chau YY, Ng HY, et al. Empagliflozin protects HK-2 cells from high glucose-mediated injuries via a mitochondrial mechanism. Cells. 2019;8(9):1085. doi:10.3390/cells8091085
122. Wu X, Li H, Wan Z, et al. The combination of ursolic acid and empagliflozin relieves diabetic nephropathy by reducing inflammation, oxidative stress and renal fibrosis. Biomed Pharmacother. 2021;144:112267. doi:10.1016/j.biopha.2021.112267
123. Wang J, Zhou Y, Long D, Wu Y, Liu F. GLP-1 receptor agonist, liraglutide, protects podocytes from apoptosis in diabetic nephropathy by promoting white fat browning. Biochem Biophys Res Commun. 2023;664:142–151. doi:10.1016/j.bbrc.2023.04.012
124. Zha D, Yao T, Bao L, Gao P, Wu X. Telmisartan attenuates diabetic nephropathy progression by inhibiting the dimerization of angiotensin type-1 receptor and adiponectin receptor-1. Life Sci. 2019;221:109–120. doi:10.1016/j.lfs.2019.01.044
125. Tang C, Deng X, Qu J, et al. Fenofibrate attenuates renal tubular cell apoptosis by up-regulating MCAD in diabetic kidney disease. Drug Des Devel Ther. 2023;17:1503–1514. doi:10.2147/DDDT.S405266
126. Ma R, Wang Y, Xu Y, et al. Tacrolimus protects podocytes from apoptosis via downregulation of TRPC6 in diabetic nephropathy. J Diabetes Res. 2021;2021:8832114. doi:10.1155/2021/8832114
127. Zhang S, Wang H, Liu Y, et al. Tacrolimus ameliorates tubulointerstitial inflammation in diabetic nephropathy via inhibiting the NFATc1/TRPC6 pathway. J Cell Mol Med. 2020;24(17):9810–9824. doi:10.1111/jcmm.15562
128. Song Y, Guo F, Zhao Y, et al. Verapamil ameliorates proximal tubular epithelial cells apoptosis and fibrosis in diabetic kidney. Eur J Pharmacol. 2021;911:174552. doi:10.1016/j.ejphar.2021.174552
129. Chang TT, Chiang CH, Chen C, Lin SC, Lee HJ, Chen JW. Antioxidation and Nrf2-mediated heme oxygenase-1 activation contribute to renal protective effects of hydralazine in diabetic nephropathy. Biomed Pharmacother. 2022;151:113139. doi:10.1016/j.biopha.2022.113139
130. Mima A, Yasuzawa T, Nakamura T, Ueshima S. Linagliptin affects IRS1/Akt signaling and prevents high glucose-induced apoptosis in podocytes. Sci Rep. 2020;10(1):5775. doi:10.1038/s41598-020-62579-7
131. Meng Q, Ma J, Suo L, Pruekprasert N, Chakrapani P, Cooney RN. Galantamine improves glycemic control and diabetic nephropathy in Leprdb/db mice. Sci Rep. 2023;13(1):15544. doi:10.1038/s41598-023-42665-2
132. Sankrityayan H, Kale A, Shelke V, Gaikwad AB. Cyproheptadine, a SET7/9 inhibitor, reduces hyperglycaemia-induced ER stress alleviating inflammation and fibrosis in renal tubular epithelial cells. Arch Physiol Biochem. 2024;130(4):411–419. doi:10.1080/13813455.2022.2105365
133. Mohany M, Ahmed MM, Al-Rejaie SS. The Role of NF-κB and Bax/Bcl-2/Caspase-3 signaling pathways in the protective effects of Sacubitril/Valsartan (Entresto) against HFD/STZ-induced diabetic kidney disease. Biomedicines. 2022;10(11):2863. doi:10.3390/biomedicines10112863
134. Dugbartey GJ, Alornyo KK, N’guessan BB, Atule S, Mensah SD, Adjei S. Supplementation of conventional anti-diabetic therapy with alpha-lipoic acid prevents early development and progression of diabetic nephropathy. Biomed Pharmacother. 2022;149:112818. doi:10.1016/j.biopha.2022.112818
135. Altındağ F, Özdek U. Synergistic effects of sinapic acid and ellagic acid ameliorate streptozotocin-induced diabetic nephropathy by inhibiting apoptosis, DNA damage, and structural deterioration in rats. Hum Exp Toxicol. 2021;40(12_suppl):S290–S299. doi:10.1177/09603271211040825
136. El-Boshy M, Alsaegh A, Qasem AH, et al. Enhanced renoprotective actions of Paricalcitol and omega-3 fatty acids co-therapy against diabetic nephropathy in rat. J Adv Res. 2021;38:119–129. doi:10.1016/j.jare.2021.08.010
137. Wang Y, Tian J, Mi Y, et al. Experimental study on renoprotective effect of intermedin on diabetic nephropathy. Mol Cell Endocrinol. 2021;528:111224. doi:10.1016/j.mce.2021.111224
138. Zhang L, Zang CS, Chen B, Wang Y, Xue S, Wu MY. Renalase regulates renal tubular injury in diabetic nephropathy via the p38MAPK signaling pathway. FASEB J. 2023;37(10):e23188. Erratum in: FASEB J. 2024;38(1):e23336. doi: 10.1096/fj.202302343. doi:10.1096/fj.202300708R
139. Liu G, Shea CM, Jones JE, et al. Praliciguat inhibits progression of diabetic nephropathy in ZSF1 rats and suppresses inflammation and apoptosis in human renal proximal tubular cells. Am J Physiol Renal Physiol. 2020;319(4):F697–F711. doi:10.1152/ajprenal.00003.2020
140. Zhang XM, Wang YZ, Tong JD, et al. Pyruvate alleviates high glucose-induced endoplasmic reticulum stress and apoptosis in HK-2 cells. FEBS Open Bio. 2020;10(5):827–834. doi:10.1002/2211-5463.12834
141. Zhou J, Zhang S, Sun X, Lou Y, Bao J, Yu J. Hyperoside ameliorates diabetic nephropathy induced by STZ via targeting the miR-499-5p/APC axis. J Pharmacol Sci. 2021;146(1):10–20. doi:10.1016/j.jphs.2021.02.005
142. Shopit A, Niu M, Wang H, et al. Protection of diabetes-induced kidney injury by phosphocreatine via the regulation of ERK/Nrf2/HO-1 signaling pathway. Life Sci. 2020;242:117248. doi:10.1016/j.lfs.2019.117248
143. Zhang YH, Zhang YQ, Guo CC, et al. Prostaglandin E1 attenuates high glucose-induced apoptosis in proximal renal tubular cells by inhibiting the JNK/Bim pathway. Acta Pharmacol Sin. 2020;41(4):561–571. doi:10.1038/s41401-019-0314-9
144. Yang M, Zhao L. The selective NLRP3-inflammasome inhibitor CY-09 ameliorates kidney injury in diabetic nephropathy by inhibiting NLRP3- inflammasome activation. Curr Med Chem. 2023;30(28):3261–3270. doi:10.2174/0929867329666220922104654
145. Xu W, Zhang H, Zhang Q, Xu J. β-Amyrin ameliorates diabetic nephropathy in mice and regulates the miR-181b-5p/HMGB2 axis in high glucose-stimulated HK-2 cells. Environ Toxicol. 2022;37(3):637–649. doi:10.1002/tox.23431
146. Wen S, Wang ZH, Zhang CX, Yang Y, Fan QL. Caspase-3 promotes diabetic kidney disease through gasdermin E-mediated progression to secondary necrosis during apoptosis. Diabetes Metab Syndr Obes. 2020;13:313–323. doi:10.2147/DMSO.S242136
147. Bangar NS, Dixit A, Apte MM, Tupe RS. Syzygium cumini (L.) skeels mitigate diabetic nephropathy by regulating Nrf2 pathway and mitocyhondrial dysfunction: in vitro and in vivo studies. J Ethnopharmacol. 2025;336:118684. doi:10.1016/j.jep.2024.118684
148. Li ZH, Sun ZJ, Tang SCW, Zhao MH, Chen M, Chang DY. Finerenone alleviates over-activation of complement C5a-C5aR1 axis of macrophages by regulating G Protein Subunit Alpha i2 to improve diabetic nephropathy. Cells. 2025;14(5):337. doi:10.3390/cells14050337
149. Liu J, Chang A, Peng H, et al. Isoferulic acid regulates CXCL12/CXCR4-mediated apoptosis and autophagy in podocyte and mice with STZ-induced diabetic nephropathy. Int Immunopharmacol. 2025;144:113707. doi:10.1016/j.intimp.2024.113707
150. Yang S, Zhang Y, Zheng C. β-sitosterol mitigates apoptosis, oxidative stress and inflammatory response by inactivating TLR4/NF-кB pathway in cell models of diabetic nephropathy. Cell Biochem Biophys. 2025;83(1):1249–1262. doi:10.1007/s12013-024-01559-4
151. Zhang C, Sun B, Wang L, Korla PK, Liu C. Micheliolide mitigates diabetic nephropathy by modulating oxidative stress and inflammation in rats. Phytomedicine. 2025;145:157025. doi:10.1016/j.phymed.2025.157025
152. Chen Y, Chen J, Jiang M, et al. Loganin and catalpol exert cooperative ameliorating effects on podocyte apoptosis upon diabetic nephropathy by targeting AGEs-RAGE signaling. Life Sci. 2020;252:117653. doi:10.1016/j.lfs.2020.117653
153. Chen X, Zhang Y, Cao Z, et al. Huperzine A targets Apolipoprotein E: a potential therapeutic drug for diabetic nephropathy based on omics analysis. Pharmacol Res. 2024;208:107392. doi:10.1016/j.phrs.2024.107392
154. Jiang T, Shen S, Wang L, Zhao M, Li Y, Huang S. Grifola frondosa polysaccharide ameliorates early diabetic nephropathy by suppressing the TLR4/NF-κB pathway. Appl Biochem Biotechnol. 2022;194(9):4093–4104. doi:10.1007/s12010-022-03976-8
155. Son SS, Kang JS, Lee EY. Paclitaxel ameliorates palmitate-induced injury in mouse podocytes. Med Sci Monit Basic Res. 2020;26:e928265. doi:10.12659/MSMBR.928265
156. Wang F, Li R, Zhao L, Ma S, Qin G. Resveratrol ameliorates renal damage by inhibiting oxidative stress-mediated apoptosis of podocytes in diabetic nephropathy. Eur J Pharmacol. 2020;885:173387. doi:10.1016/j.ejphar.2020.173387
157. Zhu M, Wang H, Chen J, Zhu H. Sinomenine improve diabetic nephropathy by inhibiting fibrosis and regulating the JAK2/STAT3/SOCS1 pathway in streptozotocin-induced diabetic rats. Life Sci. 2021;265:118855. doi:10.1016/j.lfs.2020.118855
158. Zhu Q, Yang S, Wei C, et al. Puerarin attenuates diabetic kidney injury through interaction with Guanidine nucleotide-binding protein Gi subunit alpha-1 (Gnai1) subunit. J Cell Mol Med. 2022;26(14):3816–3827. doi:10.1111/jcmm.17414
159. Zhong Y, Wang L, Jin R, et al. Diosgenin inhibits ROS generation by modulating NOX4 and mitochondrial respiratory chain and suppresses apoptosis in diabetic nephropathy. Nutrients. 2023;15(9):2164. doi:10.3390/nu15092164
160. Pei D, Tian S, Bao Y, Zhang J, Xu D, Piao M. Protective effect of salidroside on streptozotocin-induced diabetic nephropathy by inhibiting oxidative stress and inflammation in rats via the Akt/GSK-3β signalling pathway. Pharm Biol. 2022;60(1):1732–1738. doi:10.1080/13880209.2022.2116055
161. Xu C, Chen Y, Liu Z, Fu X. Hedysarum polybotrys polysaccharide attenuates renal inflammatory infiltration and fibrosis in diabetic mice by inhibiting the HMGB1/RAGE/TLR4 pathway. Exp Ther Med. 2023;26(4):493. doi:10.3892/etm.2023.12192
162. Song Y, Yu H, Sun Q, et al. Grape seed proanthocyanidin extract targets p66Shc to regulate mitochondrial biogenesis and dynamics in diabetic kidney disease. Front Pharmacol. 2023;13:1035755. doi:10.3389/fphar.2022.1035755
163. Zhang Y, Xu L, Lu Y, et al. Protective effect of Cordyceps sinensis against diabetic kidney disease through promoting proliferation and inhibiting apoptosis of renal proximal tubular cells. BMC Complement Med Ther. 2023;23(1):109. doi:10.1186/s12906-023-03901-4
164. Jung E, Pyo MK, Kim J. Pectin-Lyase-modified ginseng extract and ginsenoside Rd inhibits high glucose-induced ROS production in mesangial cells and prevents renal dysfunction in db/db mice. Molecules. 2021;26(2):367. doi:10.3390/molecules26020367
165. Lu C, Fan G, Wang D. Akebia Saponin D ameliorated kidney injury and exerted anti-inflammatory and anti-apoptotic effects in diabetic nephropathy by activation of NRF2/HO-1 and inhibition of NF-KB pathway. Int Immunopharmacol. 2020;84:106467. doi:10.1016/j.intimp.2020.106467
166. Su WY, Li Y, Chen X, et al. Ginsenoside Rh1 improves type 2 diabetic nephropathy through AMPK/PI3K/Akt-mediated inflammation and apoptosis signaling pathway. Am J Chin Med. 2021;49(5):1215–1233. doi:10.1142/S0192415X21500580
167. Wang T, Huang X, Zhai K, et al. Integrating metabolomics and network pharmacology to investigate Panax japonicus prevents kidney injury in HFD/STZ-induced diabetic mice. J Ethnopharmacol. 2023;303:115893. doi:10.1016/j.jep.2022.115893
168. Zhou T, Sun L, Yang S, et al. 20(S)-Ginsenoside Rg3 protects kidney from diabetic kidney disease via renal inflammation depression in diabetic rats. J Diabetes Res. 2020;2020:7152176. doi:10.1155/2020/7152176
169. Zhong Y, Luo R, Liu Q, et al. Jujuboside A ameliorates high fat diet and streptozotocin induced diabetic nephropathy via suppressing oxidative stress, apoptosis, and enhancing autophagy. Food Chem Toxicol. 2022;159:112697. doi:10.1016/j.fct.2021.112697
170. Feng Q, Yu X, Xie J, et al. Phillygenin improves diabetic nephropathy by inhibiting inflammation and apoptosis via regulating TLR4/MyD88/NF-κB and PI3K/AKT/GSK3β signaling pathways. Phytomedicine. 2025;136:156314. doi:10.1016/j.phymed.2024.156314
171. Nie P, Bai X, Lou Y, et al. Human umbilical cord mesenchymal stem cells reduce oxidative damage and apoptosis in diabetic nephropathy by activating Nrf2. Stem Cell Res Ther. 2021;12(1):450. doi:10.1186/s13287-021-02447-x
172. Chen L, Xiang E, Li C, et al. Umbilical cord-derived mesenchymal stem cells ameliorate nephrocyte injury and proteinuria in a diabetic nephropathy rat model. J Diabetes Res. 2020;2020:8035853. doi:10.1155/2020/8035853
173. Konari N, Nagaishi K, Kikuchi S, Fujimiya M. Mitochondria transfer from mesenchymal stem cells structurally and functionally repairs renal proximal tubular epithelial cells in diabetic nephropathy in vivo. Sci Rep. 2019;9(1):5184. doi:10.1038/s41598-019-40163-y
174. Huang Y, Li R, Zhang L, et al. Extracellular vesicles from high glucose-treated podocytes induce apoptosis of proximal tubular epithelial cells. Front Physiol. 2020;11:579296. doi:10.3389/fphys.2020.579296
175. Mohammadi-Motlagh HR, Sadeghalvad M, Yavari N, et al. β cell and autophagy: what do we know? Biomolecules. 2023;13(4):649. doi:10.3390/biom13040649
176. Klionsky DJ, Petroni G, Amaravadi RK, et al. Autophagy in major human diseases. EMBO J. 2021;40(19):e108863. doi:10.15252/embj.2021108863
177. Liu S, Yao S, Yang H, Liu S, Wang Y. Autophagy: regulator of cell death. Cell Death Dis. 2023;14(10):648. doi:10.1038/s41419-023-06154-8
178. Wang X, Zeng HX, Jiang L, Liu XQ, Huang YB, Wu YG. Clinical significance of glomerular autophagy in evaluation of diabetic kidney disease progression. Diabetes Metab Syndr Obes. 2022;15:1945–1959. doi:10.2147/DMSO.S366907
179. Wang L, Tian H, Wang H, et al. Disrupting circadian control of autophagy induces podocyte injury and proteinuria. Kidney Int. 2024;105(5):1020–1034. doi:10.1016/j.kint.2024.01.035
180. Watany MM, El-Horany HE, Elhosary MM, Elhadidy AA. Clinical application of RUBCN/SESN2 mediated inhibition of autophagy as biomarkers of diabetic kidney disease. Mol Med. 2022;28(1):147. doi:10.1186/s10020-022-00580-8
181. Bai F, Yu K, Yang Y, et al. Identification and validation of P4HB as a novel autophagy-related biomarker in diabetic nephropathy. Front Genet. 2022;13:965816. doi:10.3389/fgene.2022.965816
182. Huang Q, Fei X, Zhong Z, et al. Stratification of diabetic kidney diseases via data-independent acquisition proteomics-based analysis of human kidney tissue specimens. Front Endocrinol. 2022;13:995362. doi:10.3389/fendo.2022.995362
183. Kim KH, Hong GL, Jung DY, Karunasagara S, Jeong WI, Jung JY. IL-17 deficiency aggravates the streptozotocin-induced diabetic nephropathy through the reduction of autophagosome formation in mice. Mol Med. 2021;27(1):25. doi:10.1186/s10020-021-00285-4
184. Liu Y, Li X, Zhao M, et al. Macrophage-derived exosomes promote activation of NLRP3 inflammasome and autophagy deficiency of mesangial cells in diabetic nephropathy. Life Sci. 2023;330:121991. doi:10.1016/j.lfs.2023.121991
185. Ma Y, Yue C, Sun Q, et al. Cadmium exposure exacerbates kidney damage by inhibiting autophagy in diabetic rats. Ecotoxicol Environ Saf. 2023;267:115674. doi:10.1016/j.ecoenv.2023.115674
186. Yang J, Li L, Li C, et al. PACS-2 deficiency aggravates tubular injury in diabetic kidney disease by inhibiting ER-phagy. Cell Death Dis. 2023;14(10):649. doi:10.1038/s41419-023-06175-3
187. Lu J, Chen PP, Zhang JX, et al. GPR43 activation-mediated lipotoxicity contributes to podocyte injury in diabetic nephropathy by modulating the ERK/EGR1 pathway. Int J Biol Sci. 2022;18(1):96–111. doi:10.7150/ijbs.64665
188. Chen D, Liu Y, Chen J, et al. JAK/STAT pathway promotes the progression of diabetic kidney disease via autophagy in podocytes. Eur J Pharmacol. 2021;902:174121. doi:10.1016/j.ejphar.2021.174121
189. Jianbing H, Xiaotian L, Jie T, et al. The effect of allograft inflammatory factor-1 on inflammation, oxidative stress, and autophagy via miR-34a/ATG4B pathway in diabetic kidney disease. Oxid Med Cell Longev. 2022;2022:1668000. doi:10.1155/2022/1668000
190. Liang Q, Liu T, Guo T, et al. ATF4 promotes renal tubulointerstitial fibrosis by suppressing autophagy in diabetic nephropathy. Life Sci. 2021;264:118686. doi:10.1016/j.lfs.2020.118686
191. Wang X, Song M, Li X, et al. CERS6-derived ceramides aggravate kidney fibrosis by inhibiting PINK1-mediated mitophagy in diabetic kidney disease. Am J Physiol Cell Physiol. 2023;325(2):C538–C549. doi:10.1152/ajpcell.00144.2023
192. Chen B, He Q. miR-1187 induces podocyte injury and diabetic nephropathy through autophagy. Diab Vasc Dis Res. 2023;20(3):14791641231172139. doi:10.1177/14791641231172139
193. Fu L, Huang X, Zhang J, Lin Z, Qin G. MiR-218 promotes oxidative stress and inflammatory response by inhibiting SPRED2-mediated autophagy in HG-induced HK-2 cells. Adv Clin Exp Med. 2022;31(9):1011–1022. doi:10.17219/acem/147891
194. Yang DY, Zhou X, Liu ZW, Xu XQ, Liu C. LncRNA NEAT1 accelerates renal tubular epithelial cell damage by modulating mitophagy via miR-150-5p-DRP1 axis in diabetic nephropathy. Exp Physiol. 2021;106(7):1631–1642. doi:10.1113/EP089547
195. Ma Z, Li L, Livingston MJ, et al. p53/microRNA-214/ULK1 axis impairs renal tubular autophagy in diabetic kidney disease. J Clin Invest. 2020;130(9):5011–5026. doi:10.1172/JCI135536
196. Zhang X, Jackson S, Liu J, et al. Arsenic aggravates the progression of diabetic nephropathy through miRNA-mRNA-autophagy axis. Food Chem Toxicol. 2024;187:114628. doi:10.1016/j.fct.2024.114628
197. Qian C, Lu J, Che X, et al. P2X7R/AKT/mTOR signaling mediates high glucose-induced decrease in podocyte autophagy. Free Radic Biol Med. 2023;204:337–346. doi:10.1016/j.freeradbiomed.2023.05.015
198. Yang Q, Yang S, Liang Y, et al. UCP2 deficiency impairs podocyte autophagy in diabetic nephropathy. Biochim Biophys Acta Mol Basis Dis. 2023;1869(5):166705. doi:10.1016/j.bbadis.2023.166705
199. Xie Y, Yuan Q, Cao X, et al. Deficiency of nuclear receptor coactivator 3 aggravates diabetic kidney disease by impairing podocyte autophagy. Adv Sci. 2024;11(19):e2308378. doi:10.1002/advs.202308378
200. Li Y, Pan Y, Cao S, et al. Podocyte EGFR inhibits autophagy through upregulation of rubicon in type 2 diabetic nephropathy. Diabetes. 2021;70(2):562–576. doi:10.2337/db20-0660
201. Audzeyenka I, Rachubik P, Typiak M, et al. Hyperglycemia alters mitochondrial respiration efficiency and mitophagy in human podocytes. Exp Cell Res. 2021;407(1):112758. doi:10.1016/j.yexcr.2021.112758
202. Lu HT, Jiao YY, Yu TY, et al. Unraveling DDIT4 in the VDR-mTOR pathway: a novel target for drug discovery in diabetic kidney disease. Front Pharmacol. 2024;15:1344113. doi:10.3389/fphar.2024.1344113
203. Han Y, Xiong S, Zhao H, et al. Lipophagy deficiency exacerbates ectopic lipid accumulation and tubular cells injury in diabetic nephropathy. Cell Death Dis. 2021;12(11):1031. doi:10.1038/s41419-021-04326-y
204. Uehara R, Yamada E, Okada S, et al. Fyn phosphorylates Transglutaminase 2 (Tgm2) and modulates autophagy and p53 expression in the development of diabetic kidney disease. Cells. 2023;12(8):1197. doi:10.3390/cells12081197
205. Song FQ, Song M, Ma WX, et al. Overexpressing STAMP2 attenuates diabetic renal injuries via upregulating autophagy in diabetic rats. Biochem Biophys Res Commun. 2021;579:47–53. doi:10.1016/j.bbrc.2021.09.026
206. Zhou D, Zhou M, Wang Z, et al. PGRN acts as a novel regulator of mitochondrial homeostasis by facilitating mitophagy and mitochondrial biogenesis to prevent podocyte injury in diabetic nephropathy. Cell Death Dis. 2019;10(7):524. doi:10.1038/s41419-019-1754-3
207. Wang X, Su W, Ma M, et al. The KLF4-p62 axis prevents vascular endothelial cell injury via the mTOR/S6K pathway and autophagy in diabetic kidney disease. Endokrynol Pol. 2022;73(5):837–845. doi:10.5603/EP.a2022.0072
208. Hou B, Li Y, Li X, et al. HGF protected against diabetic nephropathy via autophagy-lysosome pathway in podocyte by modulating PI3K/Akt-GSK3β-TFEB axis. Cell Signal. 2020;75:109744. doi:10.1016/j.cellsig.2020.109744
209. Xue M, Yang F, Le Y, et al. Klotho protects against diabetic kidney disease via AMPK- and ERK-mediated autophagy. Acta Diabetol. 2021;58(10):1413–1423. doi:10.1007/s00592-021-01736-4
210. Xie J, Yuan Y, Yao G, Yu W, Zhu Q. Role and mechanism of NUP160-regulated autophagy in pathogenesis of diabetic nephropathy. Iran J Kidney Dis. 2023;17(6):327–334. doi:10.52547/ijkd.7884
211. Gao C, Fan F, Chen J, et al. FBW7 regulates the autophagy signal in mesangial cells induced by high glucose. Biomed Res Int. 2019;2019:6061594. doi:10.1155/2019/6061594
212. Ma X, Wang Q. Short-chain fatty acids attenuate renal fibrosis and enhance autophagy of renal tubular cells in diabetic mice through the HDAC2/ULK1 axis. Endocrinol Metab. 2022;37(3):432–443. doi:10.3803/EnM.2021.1336
213. Liang T, Qi C, Lai Y, et al. HDAC6-mediated α-tubulin deacetylation suppresses autophagy and enhances motility of podocytes in diabetic nephropathy. J Cell Mol Med. 2020;24(19):11558–11572. doi:10.1111/jcmm.15772
214. Dusabimana T, Kim SR, Park EJ, et al. P2Y2R contributes to the development of diabetic nephropathy by inhibiting autophagy response. Mol Metab. 2020;42:101089. doi:10.1016/j.molmet.2020.101089
215. Gong J, Zhan H, Li Y, Zhang W, Jin J, He Q. Krüppel-like factor 4 ameliorates diabetic kidney disease by activating autophagy via the mTOR pathway. Mol Med Rep. 2019;20(4):3240–3248. doi:10.3892/mmr.2019.10585
216. Wang Y, Chang J, Wang ZQ, Li Y. Sirt3 promotes the autophagy of HK-2 human proximal tubular epithelial cells via the inhibition of Notch-1/Hes-1 signaling. Mol Med Rep. 2021;24(3):634. doi:10.3892/mmr.2021.12273
217. Yang S, Lin C, Zhuo X, et al. Glucagon-like peptide-1 alleviates diabetic kidney disease through activation of autophagy by regulating AMP-activated protein kinase-mammalian target of rapamycin pathway. Am J Physiol Endocrinol Metab. 2020;319(6):E1019–E1030. doi:10.1152/ajpendo.00195.2019
218. Zhu Z, Liu Q, Sun J, Bao Z, Wang W. Silencing of PFKFB3 protects podocytes against high glucose-induced injury by inducing autophagy. Mol Med Rep. 2021;24(5):765. doi:10.3892/mmr.2021.12405
219. Barutta F, Bellini S, Kimura S, et al. Protective effect of the tunneling nanotube-TNFAIP2/M-sec system on podocyte autophagy in diabetic nephropathy. Autophagy. 2023;19(2):505–524. doi:10.1080/15548627.2022.2080382
220. Chen H, Zhang H, Li AM, et al. VDR regulates mitochondrial function as a protective mechanism against renal tubular cell injury in diabetic rats. Redox Biol. 2024;70:103062. doi:10.1016/j.redox.2024.103062
221. Chen K, Yu B, Liao J. LncRNA SOX2OT alleviates mesangial cell proliferation and fibrosis in diabetic nephropathy via Akt/mTOR-mediated autophagy. Mol Med. 2021;27(1):71. doi:10.1186/s10020-021-00310-6
222. Zhuang L, Ge X, Hu X, Yang Q, Pei X, Jin G. miR-543 regulates high glucose-induced fibrosis and autophagy in diabetic nephropathy by targeting TSPAN8. BMC Nephrol. 2022;23(1):89. doi:10.1186/s12882-022-02716-8
223. Kato M, Abdollahi M, Tunduguru R, et al. miR-379 deletion ameliorates features of diabetic kidney disease by enhancing adaptive mitophagy via FIS1. Commun Biol. 2021;4(1):30. Erratum in: Commun Biol. 2021;4(1):175. doi: 10.1038/s42003-021-01691-4. doi:10.1038/s42003-020-01516-w
224. Dong J, Liu M, Bian Y, et al. MicroRNA-204-5p ameliorates renal injury via regulating Keap1/Nrf2 pathway in diabetic kidney disease. Diabetes Metab Syndr Obes. 2024;17:75–92. doi:10.2147/DMSO.S441082
225. Dong W, Zhang H, Zhao C, Luo Y, Chen Y. Silencing of miR-150-5p ameliorates diabetic nephropathy by targeting SIRT1/p53/AMPK pathway. Front Physiol. 2021;12:624989. doi:10.3389/fphys.2021.624989
226. Huang H, Liu H, Tang J, et al. M2 macrophage-derived exosomal miR-25-3p improves high glucose-induced podocytes injury through activation autophagy via inhibiting DUSP1 expression. IUBMB Life. 2020;72(12):2651–2662. doi:10.1002/iub.2393
227. Korbut AI, Taskaeva IS, Bgatova NP, et al. SGLT2 inhibitor empagliflozin and DPP4 inhibitor linagliptin reactivate glomerular autophagy in db/db mice, a model of type 2 diabetes. Int J Mol Sci. 2020;21(8):2987. doi:10.3390/ijms21082987
228. Matsui S, Yamamoto T, Takabatake Y, et al. Empagliflozin protects the kidney by reducing toxic ALB (albumin) exposure and preventing autophagic stagnation in proximal tubules. Autophagy. 2025;21(3):583–597. doi:10.1080/15548627.2024.2410621
229. Ma R, He Y, Fang Q, Xie G, Qi M. Ferulic acid ameliorates renal injury via improving autophagy to inhibit inflammation in diabetic nephropathy mice. Biomed Pharmacother. 2022;153:113424. doi:10.1016/j.biopha.2022.113424
230. Feng B, Yang F, Liu J, Sun Q, Meng R, Zhu D. Amelioration of diabetic kidney injury with dapagliflozin is associated with suppressing renal HMGB1 expression and restoring autophagy in obese mice. J Diabetes Complications. 2023;37(3):108409. doi:10.1016/j.jdiacomp.2023.108409
231. Awad AM, Elshaer SL, Gangaraju R, Abdelaziz RR, Nader MA. Ameliorative effect of montelukast against STZ induced diabetic nephropathy: targeting HMGB1, TLR4, NF-κB, NLRP3 inflammasome, and autophagy pathways. Inflammopharmacology. 2024;32(1):495–508. doi:10.1007/s10787-023-01301-1
232. Nie Y, Fu C, Zhang H, et al. Celastrol slows the progression of early diabetic nephropathy in rats via the PI3K/AKT pathway. BMC Complement Med Ther. 2020;20(1):321. doi:10.1186/s12906-020-03050-y
233. Jiang XS, Xiang XY, Chen XM, et al. Inhibition of soluble epoxide hydrolase attenuates renal tubular mitochondrial dysfunction and ER stress by restoring autophagic flux in diabetic nephropathy. Cell Death Dis. 2020;11(5):385. doi:10.1038/s41419-020-2594-x
234. Wang F, Sun H, Zuo B, et al. Metformin attenuates renal tubulointerstitial fibrosis via upgrading autophagy in the early stage of diabetic nephropathy. Sci Rep. 2021;11(1):16362. doi:10.1038/s41598-021-95827-5
235. Nakatsuka A, Yamaguchi S, Eguchi J, et al. A Vaspin-HSPA1L complex protects proximal tubular cells from organelle stress in diabetic kidney disease. Commun Biol. 2021;4(1):373. doi:10.1038/s42003-021-01902-y
236. Li A, Yi B, Han H, et al. Vitamin D-VDR (vitamin D receptor) regulates defective autophagy in renal tubular epithelial cell in streptozotocin-induced diabetic mice via the AMPK pathway. Autophagy. 2022;18(4):877–890. doi:10.1080/15548627.2021.1962681
237. Shi L, Xiao C, Zhang Y, et al. Vitamin D/vitamin D receptor/Atg16L1 axis maintains podocyte autophagy and survival in diabetic kidney disease. Ren Fail. 2022;44(1):694–705. doi:10.1080/0886022X.2022.2063744
238. Song Z, Xiao C, Jia X, et al. Vitamin D/VDR protects against diabetic kidney disease by restoring podocytes autophagy. Diabetes Metab Syndr Obes. 2021;14:1681–1693. doi:10.2147/DMSO.S303018
239. Kong Z, Xiao M, Wang B, et al. Renoprotective effect of isoorientin in diabetic nephropathy via activating autophagy and inhibiting the PI3K-AKT-TSC2-mTOR pathway. Am J Chin Med. 2023;51(5):1269–1291. doi:10.1142/S0192415X23500581
240. Siddhi J, Sherkhane B, Kalavala AK, Arruri V, Velayutham R, Kumar A. Melatonin prevents diabetes-induced nephropathy by modulating the AMPK/SIRT1 axis: focus on autophagy and mitochondrial dysfunction. Cell Biol Int. 2022;46(12):2142–2157. doi:10.1002/cbin.11899
241. Guo L, Tan K, Luo Q, Bai X. Dihydromyricetin promotes autophagy and attenuates renal interstitial fibrosis by regulating miR-155-5p/PTEN signaling in diabetic nephropathy. Bosn J Basic Med Sci. 2020;20(3):372–380. doi:10.17305/bjbms.2019.4410
242. Sun T, Liu J, Xie C, Yang J, Zhao L, Yang J. Metformin attenuates diabetic renal injury via the AMPK-autophagy axis. Exp Ther Med. 2021;21(6):578. doi:10.3892/etm.2021.10010
243. Chen DD, Xu R, Zhou JY, et al. Cordyceps militaris polysaccharides exerted protective effects on diabetic nephropathy in mice via regulation of autophagy. Food Funct. 2019;10(8):5102–5114. doi:10.1039/c9fo00957d
244. Zhang XX, Jiang CH, Liu Y, et al. Cyclocarya paliurus triterpenic acids fraction attenuates kidney injury via AMPK-mTOR-regulated autophagy pathway in diabetic rats. Phytomedicine. 2019;64:153060. doi:10.1016/j.phymed.2019.153060
245. Matboli M, Ibrahim D, Hasanin AH, et al. Epigenetic modulation of autophagy genes linked to diabetic nephropathy by administration of isorhamnetin in type 2 diabetes mellitus rats. Epigenomics. 2021;13(3):187–202. doi:10.2217/epi-2020-0353
246. Abd El-Khalik SR, Nasif E, Arakeep HM, Rabah H. The prospective ameliorative role of zinc oxide nanoparticles in STZ-induced diabetic nephropathy in rats: mechanistic targeting of autophagy and regulating Nrf2/TXNIP/NLRP3 inflammasome signaling. Biol Trace Elem Res. 2022;200(4):1677–1687. doi:10.1007/s12011-021-02773-4
247. Chen L, Mei G, Jiang C, et al. Carbon monoxide alleviates senescence in diabetic nephropathy by improving autophagy. Cell Prolif. 2021;54(6):e13052. doi:10.1111/cpr.13052
248. Sun Y, Deng M, Ke X, et al. Epidermal growth factor protects against high glucose-induced podocyte injury possibly via modulation of autophagy and PI3K/AKT/mTOR signaling pathway through DNA methylation. Diabetes Metab Syndr Obes. 2021;14:2255–2268. doi:10.2147/DMSO.S299562
249. Wang Y, Lu YH, Tang C, et al. Calcium dobesilate restores autophagy by inhibiting the VEGF/PI3K/AKT/mTOR signaling pathway. Front Pharmacol. 2019;10:886. doi:10.3389/fphar.2019.00886
250. Zhang X, Zhang L, Chen Z, et al. Exogenous spermine attenuates diabetic kidney injury in rats by inhibiting AMPK/mTOR signaling pathway. Int J Mol Med. 2021;47(3):27. doi:10.3892/ijmm.2021.4860
251. Kong Z, Lv W, Wang Y, et al. Sinensetin ameliorates high glucose-induced diabetic nephropathy via enhancing autophagy in vitro and in vivo. J Biochem Mol Toxicol. 2023;37(10):e23445. doi:10.1002/jbt.23445
252. Yang F, Qu Q, Zhao C, et al. Paecilomyces cicadae-fermented Radix astragali activates podocyte autophagy by attenuating PI3K/AKT/mTOR pathways to protect against diabetic nephropathy in mice. Biomed Pharmacother. 2020;129:110479. doi:10.1016/j.biopha.2020.110479
253. Zhang M, Zhang Y, Xiao D, et al. Highly bioavailable berberine formulation ameliorates diabetic nephropathy through the inhibition of glomerular mesangial matrix expansion and the activation of autophagy. Eur J Pharmacol. 2020;873:172955. doi:10.1016/j.ejphar.2020.172955
254. Karunasagara S, Hong GL, Park SR, et al. Korean red ginseng attenuates hyperglycemia-induced renal inflammation and fibrosis via accelerated autophagy and protects against diabetic kidney disease. J Ethnopharmacol. 2020;254:112693. doi:10.1016/j.jep.2020.112693
255. Li T, Chen H, Guo Y, et al. Nuciferine restores autophagy via the PI3K-AKT-mTOR pathway to alleviate renal fibrosis in diabetic kidney disease. J Agric Food Chem. 2025;73(9):5223–5235. doi:10.1021/acs.jafc.4c08844
256. Wen D, Tan RZ, Zhao CY, et al. Astragalus mongholicus Bunge and Panax notoginseng (Burkill) F.H. Chen formula for renal injury in diabetic nephropathy-in vivo and in vitro evidence for autophagy regulation. Front Pharmacol. 2020;11:732. doi:10.3389/fphar.2020.00732
257. Jia Z, Wang K, Zhang Y, et al. Icariin ameliorates diabetic renal tubulointerstitial fibrosis by restoring autophagy via regulation of the miR-192-5p/GLP-1R pathway. Front Pharmacol. 2021;12:720387. doi:10.3389/fphar.2021.720387
258. Zhu X, Zhang C, Liu L, Xu L, Yao L. Senolytic combination of dasatinib and quercetin protects against diabetic kidney disease by activating autophagy to alleviate podocyte dedifferentiation via the Notch pathway. Int J Mol Med. 2024;53(3):26. doi:10.3892/ijmm.2024.5350
259. Dusabimana T, Park EJ, Je J, et al. Geniposide improves diabetic nephropathy by enhancing ULK1-mediated autophagy and reducing oxidative stress through AMPK activation. Int J Mol Sci. 2021;22(4):1651. doi:10.3390/ijms22041651
260. Chen Y, Zheng YF, Lin XH, Zhang JP, Lin F, Shi H. Dendrobium mixture attenuates renal damage in rats with diabetic nephropathy by inhibiting the PI3K/Akt/mTOR pathway. Mol Med Rep. 2021;24(2):590. doi:10.3892/mmr.2021.12229
261. Tao M, Zheng D, Liang X, et al. Tripterygium glycoside suppresses epithelial-to-mesenchymal transition of diabetic kidney disease podocytes by targeting autophagy through the mTOR/Twist1 pathway. Mol Med Rep. 2021;24(2):592. doi:10.3892/mmr.2021.12231
262. Li X, Guo L, Chen J, et al. Intravenous injection of human umbilical cord-derived mesenchymal stem cells ameliorates not only blood glucose but also nephrotic complication of diabetic rats through autophagy-mediated anti-senescent mechanism. Stem Cell Res Ther. 2023;14(1):146. doi:10.1186/s13287-023-03354-z
263. He J, Liu B, Du X, et al. Amelioration of diabetic nephropathy in mice by a single intravenous injection of human mesenchymal stromal cells at early and later disease stages is associated with restoration of autophagy. Stem Cell Res Ther. 2024;15(1):66. doi:10.1186/s13287-024-03647-x
264. Han X, Wang J, Li R, et al. Placental mesenchymal stem cells alleviate podocyte injury in diabetic kidney disease by modulating mitophagy via the SIRT1-PGC-1alpha-TFAM pathway. Int J Mol Sci. 2023;24(5):4696. doi:10.3390/ijms24054696
265. Liu H, Wang J, Yue G, Xu J. Placenta-derived mesenchymal stem cells protect against diabetic kidney disease by upregulating autophagy-mediated SIRT1/FOXO1 pathway. Ren Fail. 2024;46(1):2303396. doi:10.1080/0886022X.2024.2303396
266. Li R, Tao H, Pan K, et al. Extracellular vesicles derived from mesenchymal stem cells alleviate renal fibrosis via the miR-99b-5p/mTOR/autophagy axis in diabetic kidney disease. Stem Cell Res Ther. 2025;16(1):142. doi:10.1186/s13287-025-04265-x
267. Liang X, Qin Y, Wu D, Wang Q, Wu H. Pyroptosis: a double-edged sword in lung cancer and other respiratory diseases. Cell Commun Signal. 2024;22(1):40. doi:10.1186/s12964-023-01458-w
268. Frank D, Vince JE. Pyroptosis versus necroptosis: similarities, differences, and crosstalk. Cell Death Differ. 2019;26(1):99–114. doi:10.1038/s41418-018-0212-6
269. Zhou J, Qiu J, Song Y, et al. Pyroptosis and degenerative diseases of the elderly. Cell Death Dis. 2023;14(2):94. doi:10.1038/s41419-023-05634-1
270. Yang F, Bettadapura SN, Smeltzer MS, Zhu H, Wang S. Pyroptosis and pyroptosis-inducing cancer drugs. Acta Pharmacol Sin. 2022;43(10):2462–2473. doi:10.1038/s41401-022-00887-6
271. Rao Z, Zhu Y, Yang P, et al. Pyroptosis in inflammatory diseases and cancer. Theranostics. 2022;12(9):4310–4329. doi:10.7150/thno.71086
272. Li S, Feng L, Li G, et al. GSDME-dependent pyroptosis signaling pathway in diabetic nephropathy. Cell Death Discov. 2023;9(1):156. doi:10.1038/s41420-023-01452-8
273. Yuan S, Wang Y, Li Z, et al. Gasdermin D is involved in switching from apoptosis to pyroptosis in TLR4-mediated renal tubular epithelial cells injury in diabetic kidney disease. Arch Biochem Biophys. 2022;727:109347. doi:10.1016/j.abb.2022.109347
274. Shao Y, Deng S, Tang W, et al. Molecular mechanism of GSDMD mediated glomerular endothelial cells pyroptosis: an implying in the progression of diabetic nephropathy. Int Immunopharmacol. 2023;122:110632. doi:10.1016/j.intimp.2023.110632
275. Ding X, Jing N, Shen A, et al. MiR-21-5p in macrophage-derived extracellular vesicles affects podocyte pyroptosis in diabetic nephropathy by regulating A20. J Endocrinol Invest. 2021;44(6):1175–1184. doi:10.1007/s40618-020-01401-7
276. Wen S, Li S, Li L, Fan Q. circACTR2: a novel mechanism regulating high glucose-induced fibrosis in renal tubular cells via pyroptosis. Biol Pharm Bull. 2020;43(3):558–564. doi:10.1248/bpb.b19-00901
277. Li Y, Yu W, Xiong H, Yuan F. Circ_0000181 regulates miR-667-5p/NLRC4 axis to promote pyroptosis progression in diabetic nephropathy. Sci Rep. 2022;12(1):11994. doi:10.1038/s41598-022-15607-7
278. Zhuang L, Jin G, Qiong W, Ge X, Pei X. Circular RNA COL1A2 Mediates high glucose-induced oxidative stress and pyroptosis by regulating MiR-424-5p/SGK1 in diabetic nephropathy. Appl Biochem Biotechnol. 2023;195(12):7652–7667. doi:10.1007/s12010-023-04501-1
279. Zhan JF, Huang HW, Huang C, Hu LL, Xu WW. Long non-coding RNA NEAT1 regulates pyroptosis in diabetic nephropathy via mediating the miR-34c/NLRP3 axis. Kidney Blood Press Res. 2020;45(4):589–602. doi:10.1159/000508372
280. Wang J, Zhao SM. LncRNA-antisense non-coding RNA in the INK4 locus promotes pyroptosis via miR-497/thioredoxin-interacting protein axis in diabetic nephropathy. Life Sci. 2021;264:118728. doi:10.1016/j.lfs.2020.118728
281. Liu C, Zhuo H, Ye MY, Huang GX, Fan M, Huang XZ. LncRNA MALAT1 promoted high glucose-induced pyroptosis of renal tubular epithelial cell by sponging miR-30c targeting for NLRP3. Kaohsiung J Med Sci. 2020;36(9):682–691. doi:10.1002/kjm2.12226
282. Du Y, Feng Y, Cai Y, Tian C. CircLARP1B promotes pyroptosis of high glucose-induced renal mesangial cells by regulating the miR-578/TLR4 axis. Int Urol Nephrol. 2024;56(1):283–293. doi:10.1007/s11255-023-03672-4
283. Lan J, Xu B, Shi X, Pan Q, Tao Q. WTAP-mediated N6-methyladenosine modification of NLRP3 mRNA in kidney injury of diabetic nephropathy. Cell Mol Biol Lett. 2022;27(1):51. doi:10.1186/s11658-022-00350-8
284. Xu X, Qin Z, Zhang C, et al. TRIM29 promotes podocyte pyroptosis in diabetic nephropathy through the NF-kB/NLRP3 inflammasome pathway. Cell Biol Int. 2023;47(6):1126–1135. doi:10.1002/cbin.12006
285. Qin Y, Wu S, Zhang F, Zhou X, You C, Tan F. N6-methyladenosine methylation regulator RBM15 promotes the progression of diabetic nephropathy by regulating cell proliferation, inflammation, oxidative stress, and pyroptosis through activating the AGE-RAGE pathway. Environ Toxicol. 2023;38(11):2772–2782. doi:10.1002/tox.23917
286. Cui X, Li Y, Yuan S, et al. Alpha-kinase1 promotes tubular injury and interstitial inflammation in diabetic nephropathy by canonical pyroptosis pathway. Biol Res. 2023;56(1):5. doi:10.1186/s40659-023-00416-7
287. Li Q, Zhang K, Hou L, et al. Endoplasmic reticulum stress contributes to pyroptosis through NF-κB/NLRP3 pathway in diabetic nephropathy. Life Sci. 2023;322:121656. doi:10.1016/j.lfs.2023.121656
288. Zheng F, Ma L, Li X, et al. Neutrophil extracellular traps induce glomerular endothelial cell dysfunction and pyroptosis in diabetic kidney disease. Diabetes. 2022;71(12):2739–2750. doi:10.2337/db22-0153
289. Xie S, Song S, Liu S, et al. (Pro)renin receptor mediates tubular epithelial cell pyroptosis in diabetic kidney disease via DPP4-JNK pathway. J Transl Med. 2024;22(1):26. doi:10.1186/s12967-023-04846-5
290. Wan J, Jiang Z, Liu D, Pan S, Zhou S, Liu Z. Inhibition of the glycogen synthase kinase 3β-hypoxia-inducible factor 1α pathway alleviates NLRP3-mediated pyroptosis induced by high glucose in renal tubular epithelial cells. Exp Physiol. 2022;107(12):1493–1506. doi:10.1113/EP090685
291. Kim D, Ban KY, Lee GH, Jun HS. Lysophosphatidic acid induces podocyte pyroptosis in diabetic nephropathy by an increase of Egr1 expression via downregulation of EzH2. Int J Mol Sci. 2023;24(12):9968. doi:10.3390/ijms24129968
292. Zhang Z, Hu H, Luo Q, et al. Dihydroxyacetone phosphate accumulation leads to podocyte pyroptosis in diabetic kidney disease. J Cell Mol Med. 2024;28(3):e18073. doi:10.1111/jcmm.18073
293. Zhao W, He C, Jiang J, et al. The role of discoid domain receptor 1 on renal tubular epithelial pyroptosis in diabetic nephropathy. Korean J Physiol Pharmacol. 2022;26(6):427–438. doi:10.4196/kjpp.2022.26.6.427
294. Lv J, Hao YN, Wang XP, Lu WH, Xie LY, Niu D. Bone marrow mesenchymal stem cell-derived exosomal miR-30e-5p ameliorates high-glucose induced renal proximal tubular cell pyroptosis by inhibiting ELAVL1. Ren Fail. 2023;45(1):2177082. doi:10.1080/0886022X.2023.2177082
295. Zhu B, Cheng X, Jiang Y, et al. Silencing of KCNQ1OT1 decreases oxidative stress and pyroptosis of renal tubular epithelial cells. Diabetes Metab Syndr Obes. 2020;13:365–375. doi:10.2147/DMSO.S225791
296. Sui C, Zhou D. ADAM metallopeptidase domain 10 knockdown enables podocytes to resist high glucose stimulation by inhibiting pyroptosis via MAPK pathway. Exp Ther Med. 2023;25(6):260. doi:10.3892/etm.2023.11959
297. Xu X, Zhang L, Hua F, et al. FOXM1-activated SIRT4 inhibits NF-κB signaling and NLRP3 inflammasome to alleviate kidney injury and podocyte pyroptosis in diabetic nephropathy. Exp Cell Res. 2021;408(2):112863. doi:10.1016/j.yexcr.2021.112863
298. Xu J, Wang Q, Song YF, et al. Long noncoding RNA X-inactive specific transcript regulates NLR family pyrin domain containing 3/caspase-1-mediated pyroptosis in diabetic nephropathy. World J Diabetes. 2022;13(4):358–375. doi:10.4239/wjd.v13.i4.358
299. Qu X, Zhai B, Liu Y, et al. Pyrroloquinoline quinone ameliorates renal fibrosis in diabetic nephropathy by inhibiting the pyroptosis pathway in C57BL/6 mice and human kidney 2 cells. Biomed Pharmacother. 2022;150:112998. doi:10.1016/j.biopha.2022.112998
300. Zhang K, Li M, Yin K, et al. Hyperoside mediates protection from diabetes kidney disease by regulating ROS-ERK signaling pathway and pyroptosis. Phytother Res. 2023;37(12):5871–5882. doi:10.1002/ptr.7993
301. Li G, Liu C, Yang L, et al. Syringaresinol protects against diabetic nephropathy by inhibiting pyroptosis via NRF2-mediated antioxidant pathway. Cell Biol Toxicol. 2023;39(3):621–639. doi:10.1007/s10565-023-09790-0
302. Han J, Zuo Z, Shi X, et al. Hirudin ameliorates diabetic nephropathy by inhibiting Gsdmd-mediated pyroptosis. Cell Biol Toxicol. 2023;39(3):573–589. doi:10.1007/s10565-021-09622-z
303. Zhu W, Li YY, Zeng HX, et al. Carnosine alleviates podocyte injury in diabetic nephropathy by targeting caspase-1-mediated pyroptosis. Int Immunopharmacol. 2021;101(Pt B):108236. doi:10.1016/j.intimp.2021.108236
304. Gu J, Huang W, Zhang W, et al. Sodium butyrate alleviates high-glucose-induced renal glomerular endothelial cells damage via inhibiting pyroptosis. Int Immunopharmacol. 2019;75:105832. doi:10.1016/j.intimp.2019.105832
305. Chen Q, Wang L, Wei X, et al. Puerarin alleviates diabetic nephropathy by inhibiting Caspase-1-mediated pyroptosis. J Pharm Pharmacol. 2024;76(3):213–223. doi:10.1093/jpp/rgad113
306. Shi S, Chen X, Yu W, Ke X, Ma T. Protective effect of GLP-1 analog liraglutide on podocytes in mice with diabetic nephropathy. Endocr Connect. 2023;12(10):e230284. doi:10.1530/EC-23-0284
307. Wen S, Deng F, Li L, Xu L, Li X, Fan Q. VX-765 ameliorates renal injury and fibrosis in diabetes by regulating caspase-1-mediated pyroptosis and inflammation. J Diabetes Investig. 2022;13(1):22–33. PMID: 34494385; PMCID: PMC8756311. doi:10.1111/jdi.13660
308. Zhang Z, Ni P, Tang M, Song Y, Liu C, Zhao B. Dapagliflozin alleviates renal podocyte pyroptosis via regulation of the HO-1/NLRP3 axis. Mol Med Rep. 2023;28(5):200. doi:10.3892/mmr.2023.13087
309. Zuo Y, Chen L, He X, et al. Atorvastatin regulates MALAT1/miR-200c/NRF2 activity to protect against podocyte pyroptosis induced by high glucose. Diabetes Metab Syndr Obes. 2021;14:1631–1645. doi:10.2147/DMSO.S298950
310. Zheng S, Zhang K, Zhang Y, et al. Human umbilical cord mesenchymal stem cells inhibit pyroptosis of renal tubular epithelial cells through miR-342-3p/Caspase1 signaling pathway in diabetic nephropathy. Stem Cells Int. 2023;2023:5584894. doi:10.1155/2023/5584894
311. Li C, Ma J, Mali N, et al. Relevance of the pyroptosis-related inflammasome drug targets in the Chuanxiong to improve diabetic nephropathy. Mol Med. 2022;28(1):136. doi:10.1186/s10020-022-00567-5
312. Lv C, Cheng T, Zhang B, Sun K, Lu K. Triptolide protects against podocyte injury in diabetic nephropathy by activating the Nrf2/HO-1 pathway and inhibiting the NLRP3 inflammasome pathway. Ren Fail. 2023;45(1):2165103. doi:10.1080/0886022X.2023.2165103
313. Zhang M, Liu W, Liu Y, et al. Astragaloside IV inhibited podocyte pyroptosis in diabetic kidney disease by regulating SIRT6/HIF-1α axis. DNA Cell Biol. 2023;42(10):594–607. doi:10.1089/dna.2023.0102
314. Ram C, Gairola S, Verma S, et al. Biochanin A ameliorates nephropathy in high-fat diet/streptozotocin-induced diabetic rats: effects on NF-kB/NLRP3 axis, pyroptosis, and fibrosis. Antioxidants. 2023;12(5):1052. doi:10.3390/antiox12051052
315. Tang D, Chen X, Kang R, Kroemer G. Ferroptosis: molecular mechanisms and health implications. Cell Res. 2021;31(2):107–125. doi:10.1038/s41422-020-00441-1
316. Dixon SJ, Lemberg KM, Lamprecht MR, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–1072. doi:10.1016/j.cell.2012.03.042
317. Yang WS, SriRamaratnam R, Welsch ME, et al. Regulation of ferroptotic cancer cell death by GPX4. Cell. 2014;156(1–2):317–331. doi:10.1016/j.cell.2013.12.010
318. Ke B, Tian M, Li J, Liu B, He G. Targeting programmed cell death using small-molecule compounds to improve potential cancer therapy. Med Res Rev. 2016;36(6):983–1035. doi:10.1002/med.21398
319. Siddiqui K, Joy SS, George TP, Mujammami M, Alfadda AA. Potential role and excretion level of urinary transferrin, KIM-1, RBP, MCP-1 and NGAL markers in diabetic nephropathy. Diabetes Metab Syndr Obes. 2020;13:5103–5111. doi:10.2147/DMSO.S282166
320. van Swelm RPL, Wetzels JFM, Swinkels DW. The multifaceted role of iron in renal health and disease. Nat Rev Nephrol. 2020;16(2):77–98. doi:10.1038/s41581-019-0197-5
321. Feng X, Wang S, Sun Z, et al. Ferroptosis enhanced diabetic renal tubular injury via HIF-1α/HO-1 pathway in db/db mice. Front Endocrinol. 2021;12:626390. doi:10.3389/fendo.2021.626390
322. Wang Y, Bi R, Quan F, et al. Ferroptosis involves in renal tubular cell death in diabetic nephropathy. Eur J Pharmacol. 2020;888:173574. doi:10.1016/j.ejphar.2020.173574
323. Chaudhary K, Chilakala A, Ananth S, et al. Renal iron accelerates the progression of diabetic nephropathy in the HFE gene knockout mouse model of iron overload. Am J Physiol Renal Physiol. 2019;317(2):F512–F517. doi:10.1152/ajprenal.00184.2019
324. Dominguez JH, Liu Y, Kelly KJ. Renal iron overload in rats with diabetic nephropathy. Physiol Rep. 2015;3(12):e12654. doi:10.14814/phy2.12654
325. Wang YH, Chang DY, Zhao MH, Chen M. Glutathione peroxidase 4 is a predictor of diabetic kidney disease progression in type 2 diabetes mellitus. Oxid Med Cell Longev. 2022;2022:2948248. doi:10.1155/2022/2948248
326. Zhao P, Lv X, Zhou Z, Yang X, Huang Y, Liu J. Indexes of ferroptosis and iron metabolism were associated with the severity of diabetic nephropathy in patients with type 2 diabetes mellitus: a cross-sectional study. Front Endocrinol. 2023;14:1297166. doi:10.3389/fendo.2023.1297166
327. Ma LL, Bai Y, Liu WH, Diao ZL. Bioinformatics analysis of potential key ferroptosis-related genes involved in tubulointerstitial injury in patients with diabetic nephropathy. Ren Fail. 2023;45(1):2199095. doi:10.1080/0886022X.2023.2199095
328. Wu Y, Sun Y, Wu Y, Zhang K, Chen Y. Predictive value of ferroptosis-related biomarkers for diabetic kidney disease: a prospective observational study. Acta Diabetol. 2023;60(4):507–516. doi:10.1007/s00592-022-02028-1
329. Kim S, Kang SW, Joo J, et al. Characterization of ferroptosis in kidney tubular cell death under diabetic conditions. Cell Death Dis. 2021;12(2):160. Erratum in: Cell Death Dis. 2021;12(4):382. doi: 10.1038/s41419-021-03667-y. doi:10.1038/s41419-021-03452-x
330. Wu Y, Zhao Y, Yang HZ, Wang YJ, Chen Y. HMGB1 regulates ferroptosis through Nrf2 pathway in mesangial cells in response to high glucose. Biosci Rep. 2021;41(2):BSR20202924. doi:10.1042/BSR20202924
331. Ni L, Cao J, Yuan C, Zhou LT, Wu X. Expression of ferroptosis-related genes is correlated with immune microenvironment in diabetic kidney disease. Diabetes Metab Syndr Obes. 2022;15:4049–4064. doi:10.2147/DMSO.S388724
332. Chen J, Wu K, Lei Y, et al. Inhibition of fatty acid β-oxidation by fatty acid binding protein 4 induces ferroptosis in HK2 cells under high glucose conditions. Endocrinol Metab. 2023;38(2):226–244. doi:10.3803/EnM.2022.1604
333. Wu K, Fei L, Wang X, et al. ZIP14 is involved in iron deposition and triggers ferroptosis in diabetic nephropathy. Metallomics. 2022;14(7):mfac034. doi:10.1093/mtomcs/mfac034
334. Wang WJ, Jiang X, Gao CC, Chen ZW. Salusin-β participates in high glucose-induced HK-2 cell ferroptosis in a Nrf-2-dependent manner. Mol Med Rep. 2021;24(3):674. doi:10.3892/mmr.2021.12313
335. Peng Q, Zhang H, Li Z. KAT2A-mediated H3K79 succinylation promotes ferroptosis in diabetic nephropathy by regulating SAT2. Life Sci. 2025;376:123746. PMID: 40409584. doi:10.1016/j.lfs.2025.123746
336. Jin J, Wang Y, Zheng D, Liang M, He Q. A novel identified circular RNA, mmu_mmu_circRNA_0000309, involves in germacrone-mediated improvement of diabetic nephropathy through regulating ferroptosis by targeting miR-188-3p/GPX4 signaling axis. Antioxid Redox Signal. 2022;36(10–12):740–759. doi:10.1089/ars.2021.0063
337. Fang X, Song J, Chen Y, et al. LncRNA SNHG1 knockdown inhibits hyperglycemia induced ferroptosis via miR-16-5p/ACSL4 axis to alleviate diabetic nephropathy. J Diabetes Investig. 2023;14(9):1056–1069. doi:10.1111/jdi.14036
338. Li Q, Meng X, Hua Q. Circ ASAP2 decreased inflammation and ferroptosis in diabetic nephropathy through SOX2/SLC7A11 by miR-770-5p. Acta Diabetol. 2023;60(1):29–42. doi:10.1007/s00592-022-01961-5
339. Zhang Q, Hu Y, Hu JE, et al. Sp1-mediated upregulation of Prdx6 expression prevents podocyte injury in diabetic nephropathy via mitigation of oxidative stress and ferroptosis. Life Sci. 2021;278:119529. doi:10.1016/j.lfs.2021.119529
340. Zhang S, Zhang S, Wang H, Chen Y. Vitexin ameliorated diabetic nephropathy via suppressing GPX4-mediated ferroptosis. Eur J Pharmacol. 2023;951:175787. doi:10.1016/j.ejphar.2023.175787
341. Peng Z, Xiao H, Liu H, et al. Downregulation of ARNTL in renal tubules of diabetic db/db mice reduces kidney injury by inhibiting ferroptosis. Cell Signal. 2023;111:110883. doi:10.1016/j.cellsig.2023.110883
342. Li Q, Liao J, Chen W, et al. NAC alleviative ferroptosis in diabetic nephropathy via maintaining mitochondrial redox homeostasis through activating SIRT3-SOD2/Gpx4 pathway. Free Radic Biol Med. 2022;187:158–170. doi:10.1016/j.freeradbiomed.2022.05.024
343. Tian S, Zhou S, Wu W, et al. GLP-1 receptor agonists alleviate diabetic kidney injury via β-Klotho-mediated ferroptosis inhibition. Adv Sci. 2025;12(4):e2409781. doi:10.1002/advs.202409781
344. Qiao YY, Ji JL, Hou WL, et al. tRF3-IleAAT reduced extracellular matrix synthesis in diabetic kidney disease mice by targeting ZNF281 and inhibiting ferroptosis. Acta Pharmacol Sin. 2024;45(5):1032–1043. doi:10.1038/s41401-024-01228-5
345. Jin T, Chen C. Umbelliferone delays the progression of diabetic nephropathy by inhibiting ferroptosis through activation of the Nrf-2/HO-1 pathway. Food Chem Toxicol. 2022;163:112892. doi:10.1016/j.fct.2022.112892
346. Hou B, Ma P, Yang X, et al. In silico prediction and experimental validation to reveal the protective mechanism of Puerarin against excessive extracellular matrix accumulation through inhibiting ferroptosis in diabetic nephropathy. J Ethnopharmacol. 2024;319(Pt 2):117281. doi:10.1016/j.jep.2023.117281
347. Song SH, Han D, Park K, et al. Bone morphogenetic protein-7 attenuates pancreatic damage under diabetic conditions and prevents progression to diabetic nephropathy via inhibition of ferroptosis. Front Endocrinol. 2023;14:1172199. doi:10.3389/fendo.2023.1172199
348. Wu Z, Li D, Tian D, Liu X, Wu Z. Aspirin mediates protection from diabetic kidney disease by inducing ferroptosis inhibition. PLoS One. 2022;17(12):e0279010. doi:10.1371/journal.pone.0279010
349. Zhang S, Li Y, Liu X, et al. Carnosine alleviates kidney tubular epithelial injury by targeting NRF2 mediated ferroptosis in diabetic nephropathy. Amino Acids. 2023;55(9):1141–1155. doi:10.1007/s00726-023-03301-5
350. Gan T, Wang Q, Song Y, et al. Canagliflozin improves fatty acid oxidation and ferroptosis of renal tubular epithelial cells via FOXA1-CPT1A axis in diabetic kidney disease. Mol Cell Endocrinol. 2024;582:112139. doi:10.1016/j.mce.2023.112139
351. Tian Y, Zhou C, Yan Q, et al. Dapagliflozin improves diabetic kidney disease by inhibiting ferroptosis through β-hydroxybutyrate production. Ren Fail. 2025;47(1):2438857. doi:10.1080/0886022X.2024.2438857
352. Wang MZ, Cai YF, Fang QJ, et al. Inhibition of ferroptosis of renal tubular cells with total flavones of Abelmoschus manihot alleviates diabetic tubulopathy. Anat Rec. 2023;306(12):3199–3213. Erratum in: Anat Rec. 2025;308(8):2275-2277. doi: 10.1002/ar.25617. doi:10.1002/ar.25123
353. Zhang T, Sun W, Wang L, et al. Rosa laevigata Michx. Polysaccharide Ameliorates diabetic nephropathy in mice through inhibiting ferroptosis and PI3K/AKT pathway-mediated apoptosis and modulating tryptophan metabolism. J Diabetes Res. 2023;2023:9164883. doi:10.1155/2023/9164883
354. Liu Y, Cai Y, Wei X, et al. Molecular mechanisms of potentilla discolor bunge in regulating ferroptosis to alleviate DKD via the Nrf2 signaling pathway. J Ethnopharmacol. 2025;350:120035. PMID: 40414576. doi:10.1016/j.jep.2025.120035
355. Wang Y, Chen Z, Luo J, et al. Salidroside postconditioning attenuates ferroptosis-mediated lung ischemia-reperfusion injury by activating the Nrf2/SLC7A11 signaling axis. Int Immunopharmacol. 2023;115:109731. Erratum in: Int Immunopharmacol. 2023;117:110002. doi: 10.1016/j.intimp.2023.110002. doi:10.1016/j.intimp.2023.109731
356. Yang S, Pei T, Wang L, et al. Salidroside alleviates renal fibrosis in SAMP8 mice by inhibiting ferroptosis. Molecules. 2022;27(22):8039. doi:10.3390/molecules27228039
357. Hu J, Gu W, Ma N, Fan X, Ci X. Leonurine alleviates ferroptosis in cisplatin-induced acute kidney injury by activating the Nrf2 signalling pathway. Br J Pharmacol. 2022;179(15):3991–4009. doi:10.1111/bph.15834
358. Tan H, Chen J, Li Y, et al. Glabridin, a bioactive component of licorice, ameliorates diabetic nephropathy by regulating ferroptosis and the VEGF/Akt/ERK pathways. Mol Med. 2022;28(1):58. doi:10.1186/s10020-022-00481-w
359. Feng Q, Yang Y, Qiao Y, et al. Quercetin ameliorates diabetic kidney injury by inhibiting ferroptosis via activating Nrf2/HO-1 signaling pathway. Am J Chin Med. 2023;51(4):997–1018. doi:10.1142/S0192415X23500465
360. Liu C, Zhong M, Jin X, et al. Sleeve gastrectomy links the attenuation of diabetic kidney disease to the inhibition of renal tubular ferroptosis through down-regulating TGF-β1/Smad3 signaling pathway. J Endocrinol Invest. 2024;47(7):1763–1776. doi:10.1007/s40618-023-02267-1
361. Hara M, Oohara K, Dai DF, Liapis H. Mitotic catastrophe causes podocyte loss in the urine of human diabetics. Am J Pathol. 2019;189(2):248–257. doi:10.1016/j.ajpath.2018.10.016
362. Luo Y, Zhang L, Su N, Liu L, Zhao T. YME1L-mediated mitophagy protects renal tubular cells against cellular senescence under diabetic conditions. Biol Res. 2024;57(1):10. doi:10.1186/s40659-024-00487-0
363. Nishad R, Mukhi D, Singh AK, et al. Growth hormone induces mitotic catastrophe of glomerular podocytes and contributes to proteinuria. Cell Death Dis. 2021;12(4):342. PMID: 33795655; PMCID: PMC8016968. doi:10.1038/s41419-021-03643-6
364. Mulay SR, Thomasova D, Ryu M, et al. Podocyte loss involves MDM2-driven mitotic catastrophe. J Pathol. 2013;230(3):322–335. doi:10.1002/path.4193
365. Haupt Y, Maya R, Kazaz A, Oren M. Mdm2 promotes the rapid degradation of p53. Nature. 1997;387(6630):296–299. doi:10.1038/387296a0
366. Tang H, Lei CT, Ye C, et al. MDM2 is implicated in high-glucose-induced podocyte mitotic catastrophe via Notch1 signalling. J Cell Mol Med. 2017;21(12):3435–3444. doi:10.1111/jcmm.13253
367. Luo Y, Liu L, Zhang C. Identification and analysis of diverse cell death patterns in diabetic kidney disease using microarray-based transcriptome profiling and single-nucleus RNA sequencing. Comput Biol Med. 2024;169:107780. doi:10.1016/j.compbiomed.2023.107780
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.