Back to Journals » Drug Design, Development and Therapy » Volume 14
Pristimerin Suppresses Trophoblast Cell Epithelial–Mesenchymal Transition via miR-542-5p/EGFR Axis
Authors Shu C, Yu X, Cheng S ![]() , Jing J, Hu C
, Jing J, Hu C ![]() , Pang B
, Pang B ![]()
Received 4 August 2020
Accepted for publication 18 October 2020
Published 2 November 2020 Volume 2020:14 Pages 4659—4670
DOI https://doi.org/10.2147/DDDT.S274595
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Jianbo Sun
Chang Shu,1,* Xiaowei Yu,2,* Shihuan Cheng,3 Jili Jing,2 Cong Hu,2,3 Bo Pang4,5
1Department of Obstetrics and Gynecology, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China; 2Centre for Reproductive Medicine, Centre for Prenatal Diagnosis, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China; 3Department of Rehabilitation, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China; 4Central Laboratory, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China; 5Department of Cardiology, The First Hospital of Jilin University, Changchun, Jilin 130021, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Cong Hu; Bo Pang Email [email protected] [email protected]
Background: Ectopic pregnancy (EP) is an ectopic embryo implantation occurred outside the uterine cavity. Nowadays, more attention have garnered in fast and effective treatment with less side effects. Pristimerin is known as the clinical application for anti-cancer, and the effect on EP therapy is still unclear.
Materials and Methods: Trophoblast cell line HTR-8/SVneo was used; then, we performed cell counting kit-8 assay, wound healing assay, flow cytometry and real-time polymerase chain reaction analysis (RT-PCR) to detect the cell viability, migration ability, apoptosis and epithelial–mesenchymal transition (EMT) under pristimerin treatment. In addition, public bioinformatic database was used to discover the connection between molecular and genes. Finally, we used miRNA transfection and RT-PCR techniques to determine the underlying molecular mechanism.
Results: We revealed that pristimerin inhibited trophoblast cells proliferation, migration and EMT, while induced trophoblast cell apoptosis. Furthermore, expression of miR-542-5p, AGO2 and EGFR was suppressed in HTR-8/SVneo cells post pristimerin treatment, and miR-542-5p silence showed the same effect. Combing pristimerin treatment and miR-542-5p silence showed a synergistic action.
Conclusion: Pristimerin could be an effective treatment to block embryo implantation by miR-542-5p and EGFR down-regulation.
Keywords: pristimerin, trophoblast cell, unruptured ectopic pregnancy, miR-542-5p, EGFR
Introduction
Ectopic pregnancy (EP) is an ectopic embryo implantation occurred outside the uterine cavity.1,2 Although the incidence rate of EP is no more than 2% of all pregnancies, still, it is one of the dominant pregnancy-related diseases that associates with reproductive female mortality.2–4 EP pathology is caused by extravillous trophoblast cells proliferation and invasion, eventually resulted in local hemorrhage and rupture. Laparoscopic resection is the most conventional and effective treatment for EP, however, complications may occur subsequently, such as infections, secondary pelvic adhesions, and infertility. In recent years, methotrexate (MTX) is considered to be another effective treatment to EP. Although the anti-tumor function may cause trophoblast cells necrosis and shedding, then embryo tissue absorption. The adverse effects of MTX, such as elevated liver enzymes, nausea, and stomatitis are more intractable.5,6 Therefore, to explore new drugs to block trophoblast growth and induce apoptosis for the treatment of EP is an urgent need.
Pristimerin (20α-3-hydroxy-2-oxo-24-nor-friedela-1-10,3,5,7-tetraen-carboxylic acid-29-methyl ester) belongs to the Celastraceae and Hippocrateaceae families, and naturally occurring quinone methide triterpenoid. Various pharmacological effects of pristimerin have been discovered, such as anti-protozoal, anti-angiogenic, insecticidal, anti-inflammatory, and anti-cancer roles.7 Recently, the promoting apoptosis, inhibiting cell growth, migration and implantation activity in cancer cell lines has been confirmed, more and more researches are focused on the clinical application of pristimerin in cancer treatment.8–13 However, the role of pristimerin in trophoblast cells is yet to be discovered.
Trophoblast invasion is regulated by the interactions from trophoblastic and uterine factors at the maternal-fetal interface.14,15 A complex network of cell types, mediators, and signaling pathways regulating trophoblast invasiveness has been described, and a link has been established between regulation of non-coding RNAs (ncRNAs) and embryo development.16 MicroRNAs (miRNAs) belong to ncRNAs.17 The primarily function of miRNAs is to destabilization or translational repression mRNA transcripts by targeting the 3ʹ untranslated region (3ʹUTR) in the cytoplasm,18,19 then the downstream mRNA expression is decreased at the post-transcriptional level,20 result in both mRNA degradation and protein translation inhibition.21 Recent studies also suggest that miRNAs have also been confirmed to regulate gene transcription by positively targeting promoter elements, as RNA activation (RNAa).22–25 AGO2 interacted with miRNAs in the nucleus.26,27 Abundant evidence has shown that miRNAs play crucial roles in an amount of key biological processes leading to multiple diseases. Interestingly, miRNAs participate in different kinds of cellular processes, even associate with disease progression and severity, probably as suppressor genes or oncogenes.28 It means that miRNAs can not only be a crucial component related to diseases pathophysiological development but also be a therapy target. As a novel anti-cancer pharmaceutical, pristimerin treatment may play a central regulatory role with differentially expressed miRNAs, eventually alleviate, or slow disease progression.29,30
Epidermal growth factor receptor (EGFR) gene is a cell surface protein that binds to epidermal growth factor, thus inducing receptor dimerization and tyrosine autophosphorylation leading to cell proliferation.31 EGFR was associated with several signaling pathways, including HIF-1 pathway, PI3K-Akt pathway, ErbB pathway, FoxO pathway, MAPK pathway, estrogen pathway and focal adhesion pathway.32 Previous literatures revealed that the intentional use of EGFR inhibitors could effectively result in placental growth regressed both in vivo and in vitro.33
In the current study, we test and verify miR-542-5p as a silence signal of EGFR. Furthermore, the miR-542-5p/EGFR signaling axis regulated by pristimerin treatment in trophoblast cells resulted in HTR-8/SVneo cells EMT suppressed. Thus, pristimerin could be an effective treatment for EP.
Materials and Methods
Materials
Pristimerin (purity ≥ 99%) was reconstructed to 20 mM stock solution with dimethyl sulfoxide (DMSO), purchased from Paypaytech Inc. (Shenzhen, China). Stored at −20°C in small aliquots.
Cells and Cell Culture
Trophoblast cell line HTR-8/SVneo was purchased from the Shanghai Institute of Cell Biology, Chinese Academy of Sciences (Shanghai, China). Cells were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum, at 37 °C, 5% CO2 cell culture conditions.
Animals
Six- to eight-week-old C57BL/6 mice were obtained from Beijing Vital River Laboratory Animal Technology. All mice were maintained under specific pathogen-free (SPF) conditions with a 22°C 12-h light/dark cycle at the animal facility of Institute of Translational Medicine, Jilin University. Each female mouse was mated with one male mice. The morning of vaginal plug detection was considered as day 0.5. Then, the pregnant mice were separated into individual cages. The mice were divided into control and experimental groups which intraperitoneally injected once by DMSO or 5μg/30g pristimerin on day 4, separately.34,35 All pregnant mice were sacrificed on day 13.5 via 5% isoflurane inhalation. Then, the abdomen was dissected, the gravid uterus was removed. The number of total implantation sites and viable implantation sites per pregnant mouse was examined. The resorption embryos were identified by their smaller size and darker appearance. The frequency of fetal resorption was calculated on the day13.5 (number of resorptions/total number of formed fetuses and resorptions) × 100%.36 All animal experiments were approved by the Ethics Committee of the First Hospital of Jilin University on Laboratory Animal Care (No. 2018–018) and were performed according to the guidelines of the Institute for Laboratory Animal Research of the First Hospital of Jilin University Laboratory Animal Center (Changchun, China).
Cell Viability Detection
1−2×104 cells/hole were plated into 96-hole microplate, cells were treated with gradient concentration (0−4 μM) of pristimerin 24 h later, triplicates per concentration. 24 h after that, 10 μL Cell Counting Kit-8 (CCK-8; Beyotimes, Wuhan, China) was added to each well. After incubated for 45 minutes, optical density was examined by spectrophotometer (Bio-Rad, Hercules, CA, USA) at 450 nm. Medium alone without HTR-8/SVneo cells was set as blank, whereas 0 μM pristimerin group was set as control. The following formula was calculated for cell viability: Cell viability (% of control) = (A450 of pristimerin pritimerin treated cells- A450 of blank)/(A450 of control cells- A450 of blank) ×100%.
Wound Healing Assay
1−2×105 cells/hole were plated into 6-hole plates. After the cells were adhered for 18 h, a straight scratch was made on the cell monolayer using a sterile 1000-µL pipette tip, then change the culture to serum-free 1640 with 0 or 0.5 μM pristimerin. 24 h later, ImageJ 1.52a software (National institutes of Health, USA) was used to measure wound closure area.
Cell Apoptosis Detection by Flow Cytometry
1−2×104 cells/hole were plated into 96-hole microplate, HTR-8/SVneo cells were treated with 0 or 0.5 μM pristimerin for 24 h. Then, cell apoptosis was evaluated by Annexin V-FITC/PI apoptosis detection kit (BD Biosciences) following the manufacturer’s instructions.37 Finally, analyzed by using a FACS LSRFortessa flow cytometer and FlowJo 10.0 software package (Treestar Inc., Ashland, OR, USA).38
Real-Time Polymerase Chain Reaction Analysis (RT-PCR)
AxyPrep Multisource Total RNA Miniprep Kit (Axygen, Union City, CA, USA) was used for purifying total RNA of HTR-8/SVneo cells. PrimeScript RT Reagent Kit with gDNA Eraser (Takara Bio, Kusatsu, Japan) was used for converting RNA to cDNA. Next, polymerase chain reaction was performed using SYBR Green based on the instruction manual (Takara Bio, Kusatsu, Japan). Primer sequences were explored in Primerbank (https://pga.mgh.harvard.edu/primerbank/), purchased fromComate Bioscience (Changchun, China). Relative target mRNA expression was quantified using the ∆∆Ct method. β-actin and U6 were used as housekeeping genes for mRNA and miRNA, respectively. The primer sequences are shown in Table 1.
 |
Table 1 Primer Sequence |
Bioinformatics Analysis
Gene expression in tissues was explored by using National Center for Biotechnology Information Gene database (https://www.ncbi.nlm.nih.gov/gene/). Bio-project PRJEB4337 was found, which showed RNA-seq of tissue samples from 95 human individuals representing 27 different tissues in order to determine tissue-specificity of EGFR.
RNA Interactome Database (RNAInter, http://www.rna-society.org/raid/) is a comprehensive resource for RNA interactome data obtained from the literature and other databases, containing over 41 million RNA-associated interactions of RNA–DNA interactions, RNA–protein interactions and RNA–RNA interactions.39 With detailed interactome information, such as miR-542-5p and EGFR, visualized interaction network and RNA dynamic expression, enhanced search functions, and embedded RNA interactome tools, RNAInter depicts a system-level RNA interactome landscape with guides and help researchers to perform further studies.
Transfection with Hsa-miR-542-5p Inhibitor
5×104 cells/hole were plated into 24-hole microplate with 1640 medium. 24 h later, 1μL HiPerFect reagent (Qiagen, Shanghai, China) was mixed with 2μL 20 nM negative control or hsa-miR-542-5p inhibitor (GenePharma, Shanghai, China) for transfection, then, added 60μL 1640 medium for 10 minutes, finally added the mixture above to 150μL medium with HTR-8/SVneo cells. The efficiency of transfection was confirmed by RT-PCR, 48 h after. siRNA primer sequences are shown in Table 2.
 |
Table 2 siRNA Primer Sequences |
Statistical Analysis
Statistical analysis of the data was performed using GraphPad Prism 8.0 (San Diego California, USA). Student’s t-test was used for statistical comparisons. Results were shown as mean ± SD. p-value <0.05 were considered to denote statistical significance. All experiments were performed at least three times.
Results
Pristimerin Blocked the Growth and Migration of Trophoblast Cells
CCK-8 assay was used to investigate the potential role of pristimerin in cell viability (all p<0.05 with 0.25−4 μM concentrations of pristimerin Figure 1A) in HTR-8/SVneo cells. The result showed that pristimerin significantly inhibited trophoblast cells proliferation, even in a very low dose.
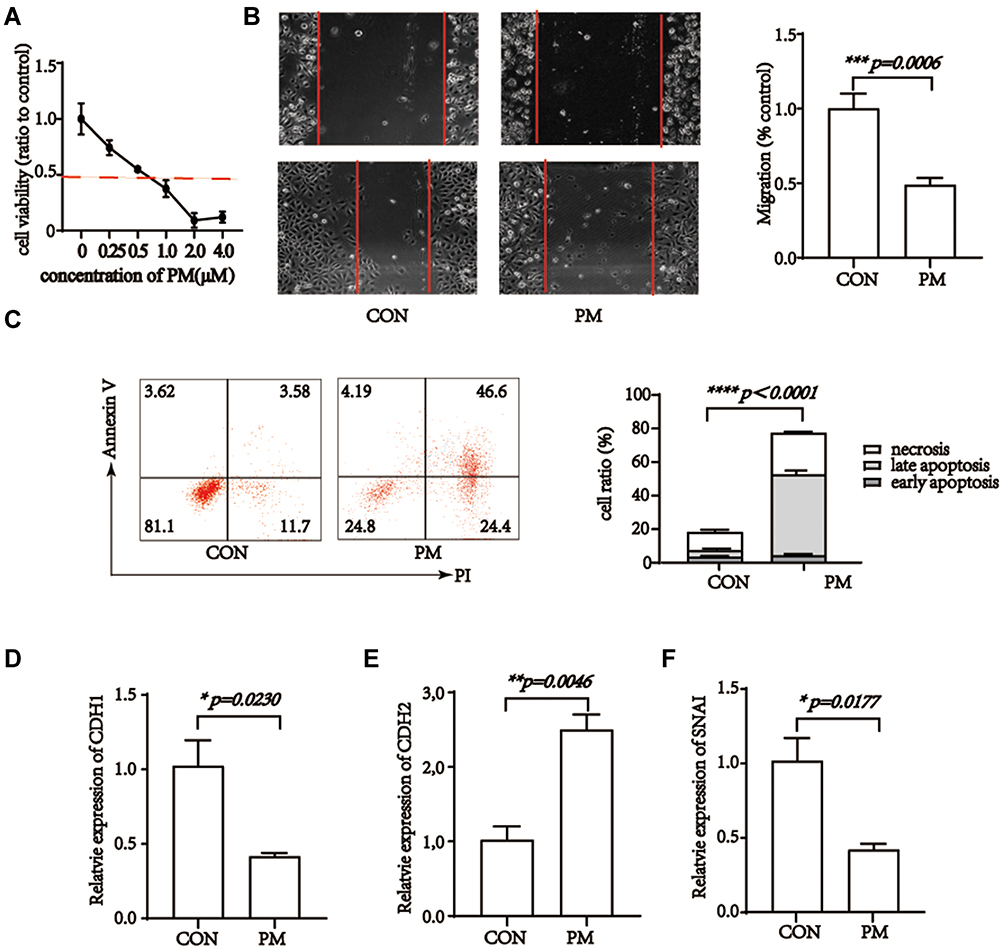 |
Figure 1 Pristimerin blocks cellular viability, migration, epithelial-mesenchymal transition and induces apoptosis of trophoblast cells. (A) Pristimerin treatment blocks cell viability of HTR-8/SVneo cells in a dose-dependent manner. (B) Pristimerin inhibits migration ability of HTR-8/SVneo cells. p=0.0006. (C) Pristimerin induces apoptosis of HTR-8/SVneo cells. p<0.0001. (D–F) Pristimerin treatment elevates expression of CDH1 and SNAI while decreases expression of CDH2 in HTR-8/SVneo cells. CON=control; PM=pristimerin. p=0.023, p=0.0177, and p=0.0046, respectively. |
Migration and invasion into endomembrane is the dominant character of trophoblast cells.40 We evaluated the migration ability of HTR-8/SVneo cells by wound healing assay, under IC50 of pristimerin treatment, HTR-8/SVneo cells migration ability was obviously declined (Figure 1B). It indicated that pristimerin had the ability to impede trophocyte cells migration.
Pristimerin Induced Apoptosis, and EMT of Trophoblast Cells
Annexin V-FITC/PI double-staining was used to further evaluate function of pristimerin on HTR-8/SVneo cells. Pristimerin treatment remarkably induced cell apoptosis and necrosis, especially late apoptosis (Figure 1C).
Our experiments showed the anti-growth effect of pristimerin on trophoblast cells. Next, we determined the effect of pristimerin on EMT process of trophoblast cells by RT-PCR. We extracted mRNA from pristimerin and control groups, qualified CDH1, CDH2 and SNAI. Statistical analysis showed up-regulation of mesenchymal associated gene CDH1 and SNAI, while epithelial-associated gene CDH2 down-regulated (Figure 1D−F).
Pristimerin Induced Fetal Resorption in Mice
To confirm the impacts of pristimerin on reproductive outcomes, pregnant mice were intraperitoneally injected by pristimerin (15 μg/30 g) or DMSO on day 4, and sacrificed on day13.5. Next, the percentage of fetal resorption per mouse, viable implantation sites, and the number of total implantation sites were measured. Compared with the control group, pristimerin-treated group experienced a significant decrease in viable implantation sites, placental hemorrhage and an obviously embryo development retardation (Figure 2).
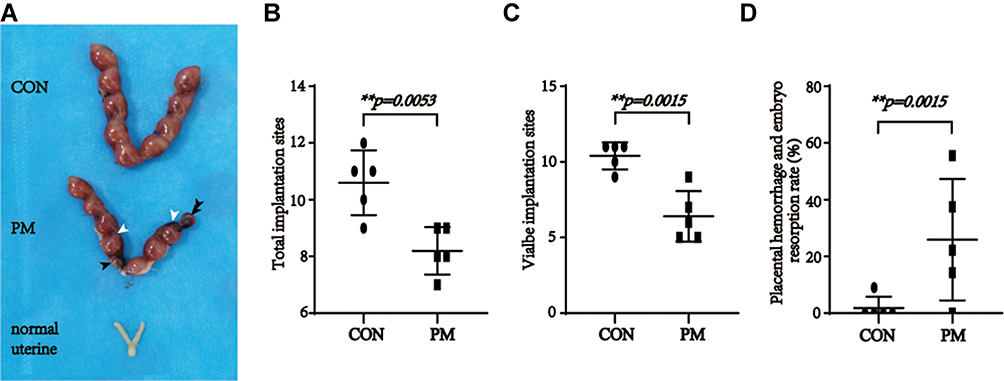 |
Figure 2 Maternal pristimerin administration results in fetal and placental resorption. (A) The uterine of control pregnancy mice, pristimerin treated pregnancy mice and normal mice. n = 5, respectively. Black arrows indicate the absorbed embryos. White arrows indicate the placental hemorrhage. (B) Statistic analysis of the number of total implantation sites. p= 0.0053. (C) Statistic analysis of the number of viable implantation sites. p=0.0015. (D) Statistic analysis of placental hemorrhage and embryo resorption rate. p=0.0384. |
Pristimerin Down-Regulated miR-542-5p and EGFR in Trophoblast Cells
In our previous study, bioinformatic analysis showed that 13 genes are targeted by pristimerin, including CYP3A4, PTPN1, TNF, NOS2, ZEB1, EGFR, AGO2, NUP62, MPO, MAPK3, CYCS, IL-10 and FASN.29 Among these genes, EGFR, known as epidermal growth factor receptor, is a transmembrane glycoprotein and the founding member of the ErbB tyrosine kinase receptors.41 RNA-seq revealed that EGFR expression in placenta was the highest compared to other human tissues42,43 (Figure 3A). EGFR plays critical roles in placental development and survival. Mount of evidence illustrated that AGO2 can also exist in the nucleus and interacted with miRNAs.26,27 Bioinformatic analysis by RNAInter showed that the predominant 100 genes binding to miR-542-5p contained AGO2 and EFGR, whereas EFGR was also predicted to bind miR-542-5p (Figure 3B−C). To inquire whether AGO2 is involved in our miRNA function and investigate the potential role of pristimerin, the expression level of miR-542-5p, AGO2 and EGFR with or without pristimerin treatment in HTR-8/SVneo cells were analyzed by RT-PCR. The results indicated that similar with anti-cancer effect, pristimerin treatment significantly decreased miR-542-5p, AGO2 and EGFR levels in trophoblast cells (Figure 3D−F). Our findings suggested that under pristimerin treatment, miR-542-5p, AGO2 and EGFR both declined, which probably associated with EP occurrence.
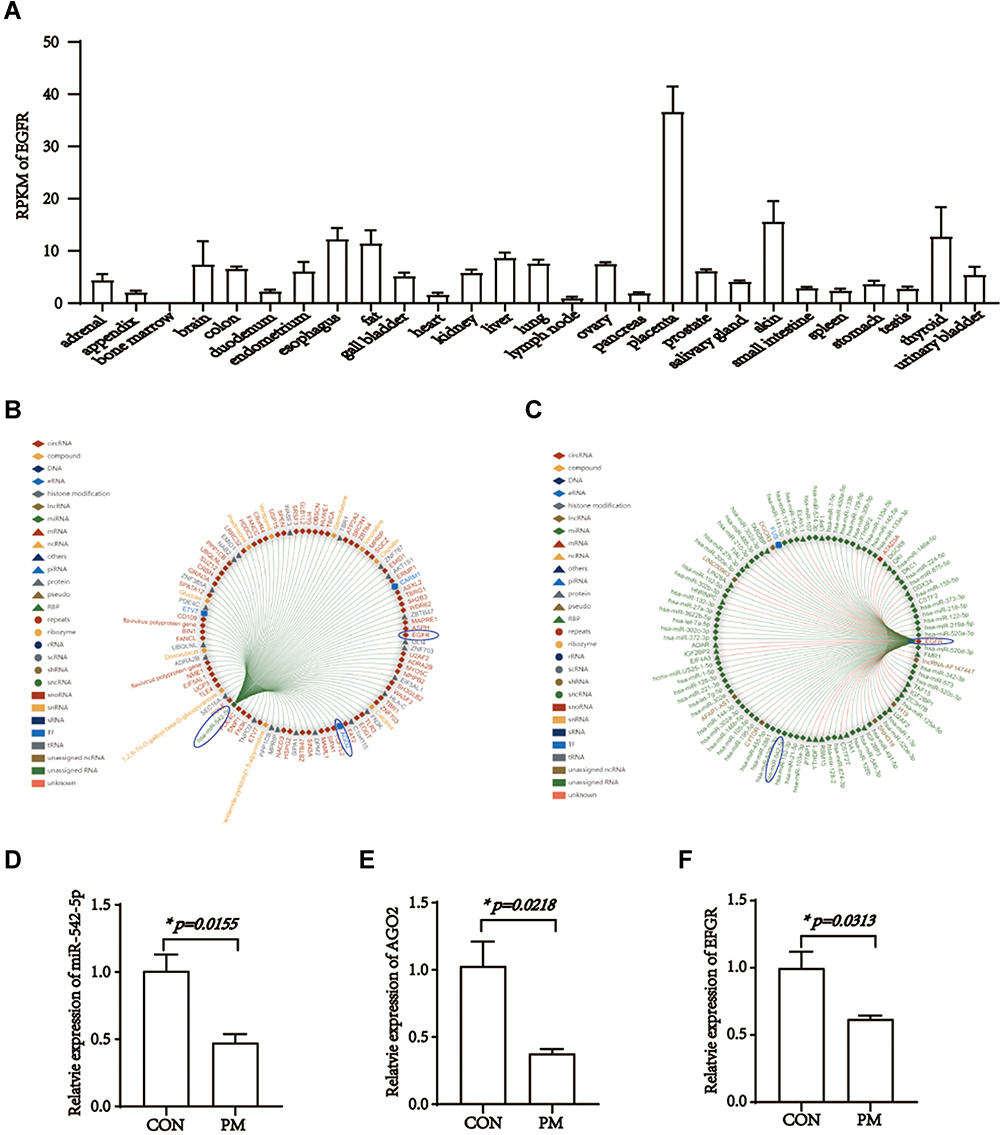 |
Figure 3 EGFR differential expression in tissues, interacts with mi4-542-5p and is down-regulated under pristimerin treatment. (A) Expression of EGFR in different tissues. (B) The top 100 genes interaction with miR-542-5p. (C) The top 100 genes interaction with EGFR. (D) Expression of miR-542-5p is decreased in HTR-8/SVneo 24h post-treated with pristimerin, p=0.0155. (E) Expression of AGO2 is decreased in HTR-8/SVneo 24h post-treated with pristimerin, p=0.0218. (F) Expression of EGFR is decreased in HTR-8/SVneo 24h post-treated with pristimerin, p=0.0313.*p<0.05. RPKM, reads per kilobase per million reads placed. |
miR-542-5p Targeted AGO2 and EGFR in Trophoblast Cells
Literatures associated with miR-542 were focused on survival, proliferation, metastasis, migration, and angiogenesis of tumor cells.44–46 Hence, we hypothesized that pristimerin might directly modulate miR-542-5p to target AGO2 and EGFR, resulted in the proliferation, migration and cell cycle were all inhibited, meanwhile EMT was induced. Next, to confirm miR-542-5p is the target miRNA of pristimerin, we silenced miR-542-5p and observed the phenomenon of trophoblast cells (transfect efficiency is shown in Figure 4A). Our results revealed that miR-542-5p silence induced apoptosis, and inhibited migration in HTR-8/SVneo cells (Figure 4B and C). EMT evaluation by RT-PCR, also showed mesenchymal-associated gene CDH1 and SNAI increased, whereas epithelial associated gene CDH2 reduced (Figure 4D−F). Next, we found AGO2 and EGFR mRNA levels were both positive related with miR-542-5p. It indicated that miR-542-5p might be the upstream of AGO2 and EGFR (Figure 5A and B). Finally, the synergistic effect of pristimerin treatment and miR-542-5p silence in trophoblast cells was evaluated. miR-542-5p negative control (NC) or inhibitor was transfected to HTR-8/SVneo cells for 24h, then treated with or without pristimerin (0.5 μM) for another 48h, RT-PCR was performed to determine EMT associated genes and AGO2/EGFR expression. hsa-miR-542-5p NC transfection with 0μM pristimerin treated group was set as control. HTR-8/SVneo human trophoblast cells with hsa-miR-542-5p NC transfection plus pristimerin treated showed lower expression of CDH1, SNAI, AGO2 and EGFR compared to control, meanwhile, hsa-miR-542-5p silence with pristimerin treated showed even lower expression of CDH1, SNAI, AGO2 and EGFR compared with hsa-miR-542-5p NC transfection with pristimerin treated group (Figure 5C−G). Our results suggested that pristimerin treated and miR-542-5p silence had profoundly the same effect to decease AGO2/EGFR expression, and inhibit trophoblast cells EMT.
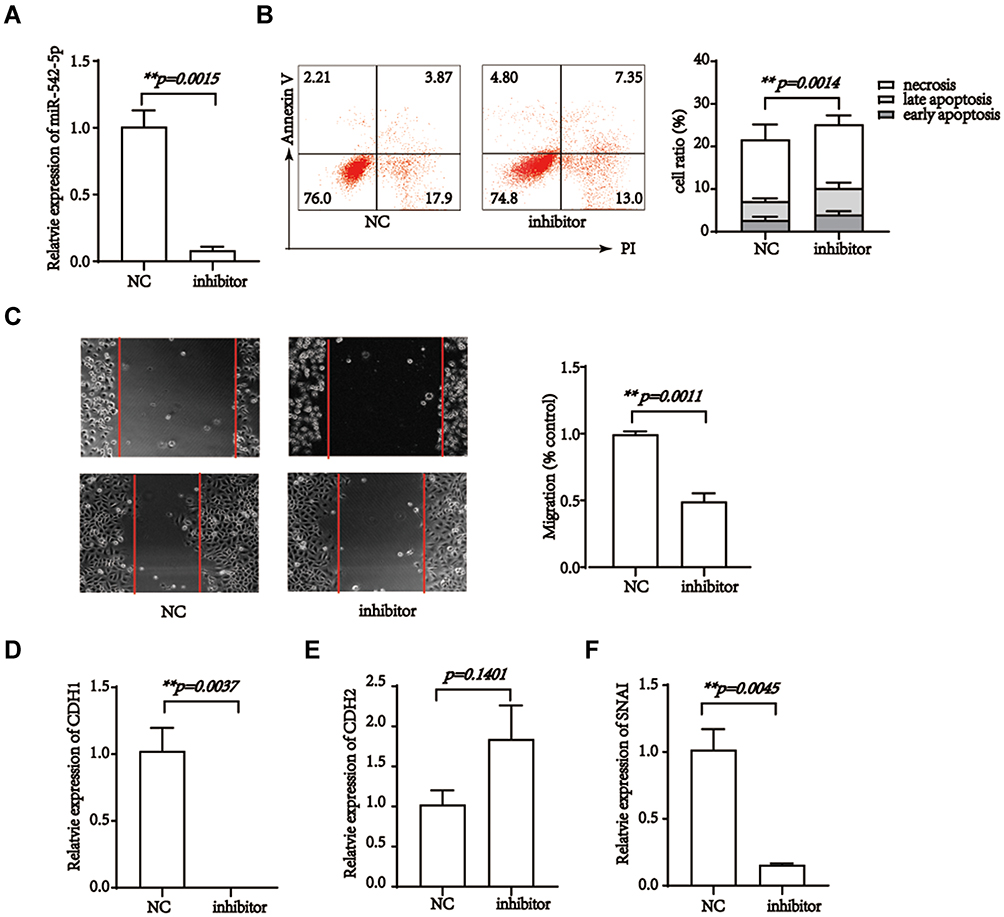 |
Figure 4 miR-542-5p silence inhibits the migration, epithelial-mesenchymal transition and induce apoptosis of trophoblast cells. (A) Relative expression of miR-542-5p in HTR-8/SVneo cells, after transfected with miR-542-5p NC or inhibitor for 24h. (B) miR-542-5p silence inhibits the migratory potential of HTR-8/SVneo cells. p=0.0011. (C) miR-542-5p silence induces HTR-8/SVneo cells apoptosis. p =0.0014. (D–F) miR-542-5p silence increased the expression of CDH1 and SNAI while decreases expression of CDH2 in HTR-8/SVneo cells. p=0.0037, p=0.0045, and p=0.1401, respectively. **p<0.01. |
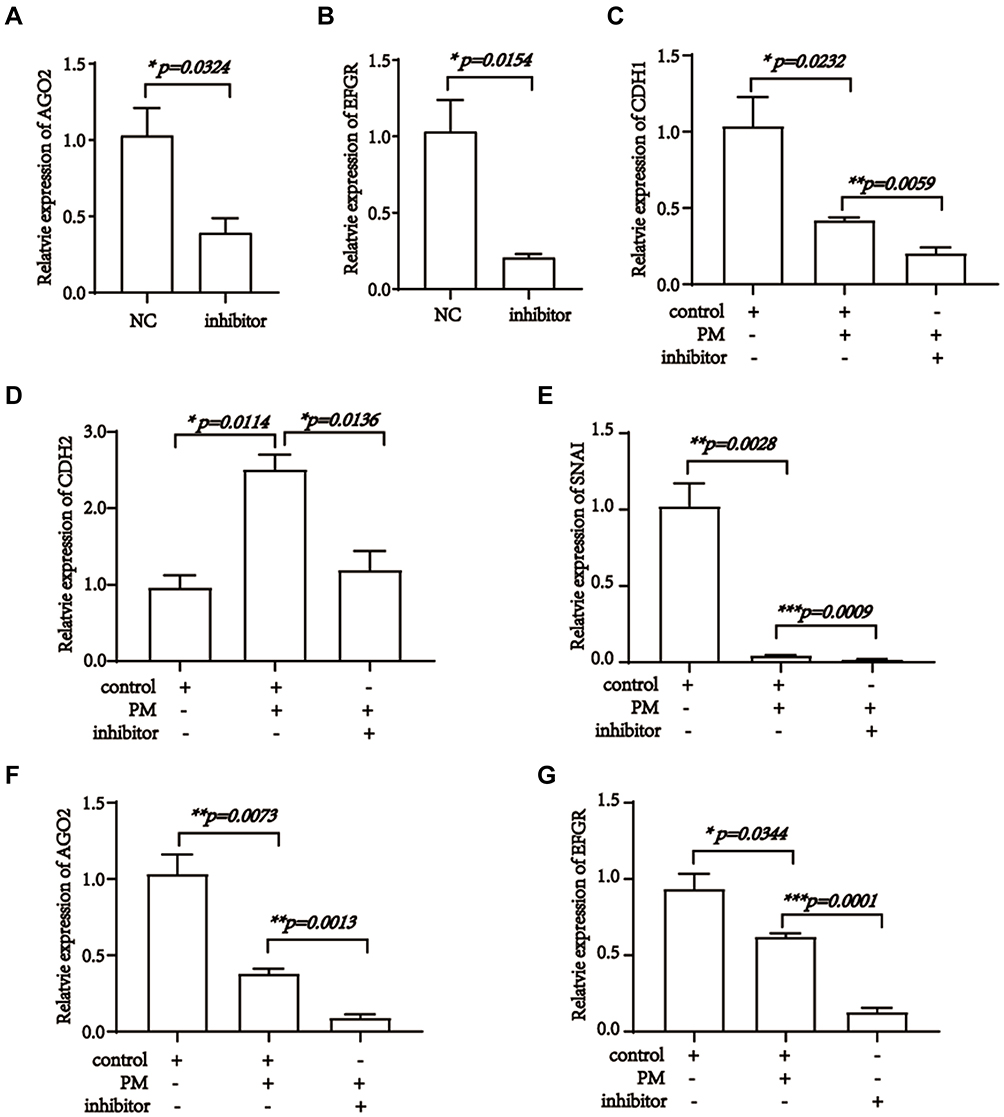 |
Figure 5 Pristimerin targeted miR-542-5p, AGO2 and EGFR in trophoblast cells. (A) Expression of AGO2 decreases post miR-542-5p inhibitor transfection, p=0.0324. (B) Expression of EGFR decreases post miR-542-5p inhibitor transfection, p=0.0154. (C) Expression of CDH1 48 h after miR-542-5p NC or inhibitor transfection, and 24 h post-treated with or without pristimerin (0.5 μM) in HTR-8/SVneo. p=0.0205, p=0.0232, p=0.0059, respectively. (D) Expression of CDH2 48 h after miR-542-5p NC or inhibitor transfection, and 24 h post-treated with or without pristimerin (0.5 μM) in HTR-8/SVneo. p=0.0114, p=0.0136, respectively. (E) Expression of SNAI 48 h after miR-542-5p NC or inhibitor transfection, and 24 h post-treated with or without pristimerin (0.5 μM) in HTR-8/SVneo. p=0.0028, p=0.0009, respectively. (F) Expression of AGO2 48 h after miR-542-5p NC or inhibitor transfection, and 24 h post-treated with or without pristimerin (0.5 μM) in HTR-8/SVneo. p=0.0073, p=0.0013, respectively. (G) Expression of EGFR 48 h after miR-542-5p NC or inhibitor transfection, and 24 h post-treated with or without pristimerin (0.5 μM) in HTR-8/SVneo. p=0.02344, p=0.0001, respectively. *p<0.05; **p<0.01; ***p<0.001. |
Discussion
Despite EP can be early and precisely diagnosis, fast and effective treatment with less side effects is still under exploration. Pristimerin purified from traditional medicinal plants has the activity to inhibit tumor angiogenesis and prevent chemotherapy resistance.12,47 In the current study, we find pristimerin inhibit trophoblast cells proliferation and migration, induce apoptosis and EMT, while miR-542-5p/AGO2/EGFR expression down-regulated. Nowadays, placenta-enriched molecules, such as EGFR is confirmed to be a target gene for EP therapy.41 In addition, we point out that miR-542-5p is the crucial molecular under pristimerin treatment to induce trophoblast cells EMT through EGFR signaling.
Embryo implantation includes the process of embryo location, adhesion and successful invasion into decidua. It has similar proliferation, adhesion and invasion characteristics with tumorigenesis.48 In tumor, angiogenesis, degeneration of extracellular matrix, migration and proliferation of vascular endothelial cells, are the basis of epithelial–mesenchymal transition (EMT).49–51 EMT refers to the basal polarity of epithelial cells down-regulating, resulting in the transformation into stromal cells, which endows cells with the ability to transfer and invade.52 Epithelial cells mainly expressed E-cadherin protein (encoded by CDH1 gene), while stromal cells mainly expressed N-cadherin protein (encoded by CDH2 gene).53 Snail, as the main transcription factor of E-cadherin, is as high as E-cadherin in EMT.54 The key to successful pregnancy is the development of blastocyst to the right state and the coordinated embryo EMT with implantation and embryo morphogenesis. EMT is a biologic process that polarized epithelial cells undergo multiple biochemical changes to gain a mesenchymal cell phenotype. The epithelial-like trophectoderm cells transform into invasive trophoblast giant cells, known as the yolk sac placenta, to provide a network of anastomotic channels surrounding the embryo and anchor the placenta. EMT is the most commonly used and significant indicator of endometrial receptivity.55,56 At the maternal fetal interface, after the blastocyst adheres to decidual tissue, extravillous trophoblasts (EVT) also need to undergo EMT to invade the endometrial matrix, complete embryo implantation, and transform the maternal spiral artery into a low resistance, high flow blood vessel, so that the maternal blood flows through the trophoblast layer and establishes the utero placental circulation.16,57 The moderate invasion of EVT and the establishment of blood supply between mother and fetus play an important role in embryo implantation.
Poor EVT migration and invasion often result in failure to establish the maternal–fetal connection and are associated with pre-eclampsia, fetal growth restriction and recurrent miscarriage,58 meanwhile, it could be used as an effective treatment for EP. ncRNAs and transcription factors are reported to play important roles in EVT invasion during early pregnancy and may be involved in the pathogenesis of embryo loss.16,59 In the present study, we test and verified expression of hsa-miR-542-5p was significantly decreased post pristimerin treatment, meanwhile trophoblast cells viability reduced. It indicated that hsa-miR-542-5p might unfacilitated trophoblast cells proliferation. Additionally, the effect of hsa-miR-542-5p on EP associated gene EGFR was detected. Our results suggested that hsa-miR-542-5p silence and pristimerin treatment shared synergistic effect, decreasing HTR-8/SVneo cells EMT potentially achieved by its down-regulation on AGO2 and EGFR gene expression. The function of inhibiting trophoblast cells migration and EMT indicated that it may act as a novel therapeutic target in EP. Here, we provided a mechanism basis for pristimerin application as a medical therapy for EP, however, further researches are still needed.
Conclusions
In conclusion, the current study revealed pristimerin may acted as a treatment to block embryo implantation through miR-542-5p/EGFR axis, which could also be a potential novel therapy for EP.
Acknowledgment
We acknowledge the technical support of Central Laboratory, The First Hospital of Jilin University.
Funding
The research is supported by grant from NSFC (81671592), Finance Department of Jilin Province (JLSWSRCZX2020-089), Education Department of Jilin Province (JJKH20201064KJ), and Science and Technology Department of Jilin Province (20180201028YY).
Disclosure
The authors report no conflicts of interest for this work.
References
1. Chouinard M, Mayrand M-H, Ayoub A, Healy-Profitós J, Auger N. Ectopic pregnancy and outcomes of future intrauterine pregnancy. Fertility Sterility. 2019;112(1):112–119. doi:10.1016/j.fertnstert.2019.03.019
2. American College of Obstetricians and Gynecologists’ Committee on Practice Bulletins—Gynecology. ACOG practice bulletin no. 210: fecal incontinence. Obstet Gynecol. 2019;133(4):e260–e273. doi:10.1097/AOG.0000000000003187
3. Brady PC. New evidence to guide ectopic pregnancy diagnosis and management. Obstetrical Gynecological Survey. 2017;72(10):618–625. doi:10.1097/OGX.0000000000000492
4. Creanga AA, Syverson C, Seed K, Callaghan WM. Pregnancy-related mortality in the United States, 2011–2013. Obstet Gynecol. 2017;130(2):366–373. doi:10.1097/AOG.0000000000002114
5. Mol F, Mol BW, Ankum WM, van der Veen F, Hajenius PJ. Current evidence on surgery, systemic methotrexate and expectant management in the treatment of tubal ectopic pregnancy: a systematic review and meta-analysis. Human Reproduction Update. 2008;14(4):309–319. doi:10.1093/humupd/dmn012
6. Uludag SZ, Kutuk MS. Comparison of systemic and local methotrexate treatments in cesarean scar pregnancies: time to change conventional treatment and follow-up protocols. European J Obstetrics, Gynecology, Reproductive Biology. 2016;206:131–135. doi:10.1016/j.ejogrb.2016.09.010
7. Brinker AM, Ma J, Lipsky PE, Raskin I. Medicinal chemistry and pharmacology of genus tripterygium (celastraceae). Phytochemistry. 2007;68(6):732–766. doi:10.1016/j.phytochem.2006.11.029
8. Yousef BA, Hassan HM, Zhang LY, Jiang ZZ. Pristimerin exhibits in vitro and in vivo anticancer activities through inhibition of nuclear factor-small ka, CyrillicB signaling pathway in colorectal cancer cells. Phytomedicine. 2018;40:140–147. doi:10.1016/j.phymed.2018.01.008
9. Liu YB, Gao X, Deeb D, Arbab AS, Gautam SC. Pristimerin induces apoptosis in prostate cancer cells by down-regulating Bcl-2 through ROS-dependent ubiquitin-proteasomal degradation pathway. J Carcinogenesis Mutagenesis. 2013;3(Suppl 6):005.
10. Wu CC, Chan ML, Chen WY, Tsai CY, Chang FR, Wu YC. Pristimerin induces caspase-dependent apoptosis in MDA-MB-231 cells via direct effects on mitochondria. Molecular Cancer Therapeutics. 2005;4(8):1277. doi:10.1158/1535-7163.MCT-05-0027
11. Tiedemann RE, Schmidt J, Keats JJ, et al. Identification of a potent natural triterpenoid inhibitor of proteosome chymotrypsin-like activity and NF-kappaB with antimyeloma activity in vitro and in vivo. Blood. 2009;113(17):4027–4037.
12. Xie G, Yu X, Liang H, et al. Pristimerin overcomes adriamycin resistance in breast cancer cells through suppressing Akt signaling. Oncol Lett. 2016;11(5):3111–3116. doi:10.3892/ol.2016.4335
13. Yousef BA, Hassan HM, Zhang LY, Jiang ZZ. Anticancer potential and molecular targets of pristimerin: a mini- review. Current Cancer Drug Targets. 2017;17(2). doi:10.2174/1568009616666160112105824
14. Ma XL, Li X, Tian FJ, et al. Upregulation of RND3 affects trophoblast proliferation, apoptosis, and migration at the maternal-fetal interface. Front Cell Dev Biol. 2020;8:153.
15. Zhang Y, He XY, Qin S, et al. Upregulation of PUM1 expression in preeclampsia impairs trophoblast invasion by negatively regulating the expression of the lncRNA HOTAIR. Mol Ther. 2020;28(2):631–641. doi:10.1016/j.ymthe.2019.11.025
16. Tian FJ, He XY, Wang J, et al. Elevated tristetraprolin impairs trophoblast invasion in women with recurrent miscarriage by destabilization of HOTAIR. Mol Ther Nucleic Acids. 2018;12:600–609. doi:10.1016/j.omtn.2018.07.001
17. Paulmurugan R, Oronsky B, Brouse CF, Reid T, Knox S, Scicinski J. Real time dynamic imaging and current targeted therapies in the war on cancer: a new paradigm. Theranostics. 2013;3(6):437–447. doi:10.7150/thno.5658
18. Fabian M, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annual Review Biochemistry. 2010;79:351–379. doi:10.1146/annurev-biochem-060308-103103
19. Pasquinelli A. MicroRNAs and their targets: recognition, regulation and an emerging reciprocal relationship, Nature reviews. Genetics. 2012;13(4):271–282.
20. RD T, SB Y, Tang Y, O’Gorman WE, Chen C-Z. The potential functions of primary microRNAs in target recognition and repression. EMBO J. 2010;29(19):3272–3285. doi:10.1038/emboj.2010.208
21. Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–233. doi:10.1016/j.cell.2009.01.002
22. Matsui M, Chu Y, Zhang H, et al. Promoter RNA links transcriptional regulation of inflammatory pathway genes. Nucleic Acids Research. 2013;41(22):10086–10109. doi:10.1093/nar/gkt777
23. Huang V, Place R, Portnoy V, et al. Upregulation of Cyclin B1 by miRNA and its implications in cancer. Nucleic Acids Research. 2012;40(4):1695–1707. doi:10.1093/nar/gkr934
24. Xiao M, Li J, Li W, et al. MicroRNAs activate gene transcription epigenetically as an enhancer trigger. RNA Biology. 2017;14(10):1326–1334. doi:10.1080/15476286.2015.1112487
25. Turner M, Jiao A, Slack F. Autoregulation of lin-4 microRNA transcription by RNA activation (RNAa) in C. elegans. Cell Cycle. 2014;13(5):772–781. doi:10.4161/cc.27679
26. Nishi K, Nishi A, Nagasawa T, Ui-Tei K. Human TNRC6A is an Argonaute-navigator protein for microRNA-mediated gene silencing in the nucleus. RNA. 2013;19(1):17–35. doi:10.1261/rna.034769.112
27. Zisoulis D, Kai Z, Chang R, Pasquinelli A. Autoregulation of microRNA biogenesis by let-7 and Argonaute. Nature. 2012;486(7404):541–544. doi:10.1038/nature11134
28. Hayes J, Peruzzi PP, Lawler S. MicroRNAs in cancer: biomarkers, functions and therapy. Trends Molecular Medicine. 2014;20(8):460–469. doi:10.1016/j.molmed.2014.06.005
29. Li Z, Hu C, Zhen Y, Pang B, Yi H, Chen X. Pristimerin inhibits glioma progression by targeting AGO2 and PTPN1 expression via miR-542-5p. Biosci Rep. 2019;39:5.
30. Tu Y, Tan F, Zhou J, Pan J. Pristimerin targeting NF-kappaB pathway inhibits proliferation, migration, and invasion in esophageal squamous cell carcinoma cells. Cell Biochem Funct. 2018;36(4):228–240. doi:10.1002/cbf.3335
31. Jiang X, Xu Y, Yuan L, et al. TGFβ2-mediated epithelial-mesenchymal transition and NF-κB pathway activation contribute to osimertinib resistance. Acta pharmacologica Sinica. 2020. doi:10.1038/s41401-020-0457-8
32. Liu J, Zhao M. Prioritization of susceptibility genes for ectopic pregnancy by gene network analysis. International J Molecular Sciences. 2016;17:2.
33. Nilsson U, Johns T, Wilmann T, et al. Effects of gefitinib, an epidermal growth factor receptor inhibitor, on human placental cell growth. Obstet Gynecol. 2013;122(4):737–744. doi:10.1097/AOG.0b013e3182a1ba56
34. Deng Q, Bai S, Gao W, Tong L. Pristimerin inhibits angiogenesis in adjuvant-induced arthritic rats by suppressing VEGFR2 signaling pathways. Int Immunopharmacol. 2015;29(2):302–313. doi:10.1016/j.intimp.2015.11.001
35. El-Agamy DS, El-Harbi KM, Khoshhal S, et al. Pristimerin protects against doxorubicin-induced cardiotoxicity and fibrosis through modulation of Nrf2 and MAPK/NF-kB signaling pathways. Cancer Manag Res. 2019;11:47–61. doi:10.2147/CMAR.S186696
36. Hosseini M, Ali-Hassanzadeh M, Nadimi E, Karbalay-Doust S, Noorafshan A, Gharesi-Fard B. Stereological study of the placental structure in abortion-prone mice model (CBA/J×DBA/2J). Ann Anatomy. 2020;230:151508. doi:10.1016/j.aanat.2020.151508
37. KH L. Systematic dissection of the metabolic-apoptotic interface in aml reveals heme biosynthesis to be a regulator of drug sensitivity. Cell Metabolism Undefined. 2019. undefined.
38. JM S, HT X. YF, Induction of cell-cycle arrest and apoptosis in human cholangiocarcinoma cells by pristimerin. J Cellular Biochemistry Undefined. 2019. undefined.
39. Lin Y, Liu T, Cui T, et al. RNAInter in 2020: RNA interactome repository with increased coverage and annotation. Nucleic Acids Research. 2020;48:D189–D197. doi:10.1093/nar/gkz804
40. Wang D, Na Q, Song G, Wang L. Human umbilical cord mesenchymal stem cell-derived exosome-mediated transfer of microRNA-133b boosts trophoblast cell proliferation, migration and invasion in preeclampsia by restricting SGK1. Cell Cycle. 2020;1–15.
41. Whigham C, MacDonald T, Walker S, Hannan N, Tong S, Kaitu’u-Lino T. The untapped potential of placenta-enriched molecules for diagnostic and therapeutic development. Placenta. 2019;84:28–31. doi:10.1016/j.placenta.2019.02.002
42. Wu C, Orozco C, Boyer J, et al. BioGPS: an extensible and customizable portal for querying and organizing gene annotation resources. Genome Biology. 2009;10(11):R130. doi:10.1186/gb-2009-10-11-r130
43. Fagerberg L, Hallström B, Oksvold P, et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Molecular Cellular Proteomics. 2014;13(2):397–406. doi:10.1074/mcp.M113.035600
44. Rosa*MBA, Sorrentino†A, Sthandier*O, et al.The interplay between the master transcription factor PU.1 and miR-424 regulates human monocyte/macrophage differentiation.PNAS 2007;104(50):19849–19854.doi:10.1073/pnas.0706963104
45. Schmeier S, MacPherson CR, Essack M, et al. Deciphering the transcriptional circuitry of microRNA genes expressed during human monocytic differentiation. BMC Genomics. 2009;10:595. doi:10.1186/1471-2164-10-595
46. Singh R, Kaushik S, Wang Y. Autophagy regulates lipid metabolism. Nature. 2009;458(7242):1131–1135. doi:10.1038/nature07976
47. Mu X, Shi W, Sun L, Li H, Jiang Z, Zhang L. Pristimerin, a triterpenoid, inhibits tumor angiogenesis by targeting VEGFR2 activation. Molecules. 2012;17(6):6854. doi:10.3390/molecules17066854
48. Shi Z, She K, Li H, Yuan X, Han X, Wang Y. MicroRNA-454 contributes to sustaining the proliferation and invasion of trophoblast cells through inhibiting Nodal/ALK7 signaling in pre-eclampsia. Chemico-Biological Interactions. 2019;298:8–14. doi:10.1016/j.cbi.2018.10.012
49. CL D, Colson A, Hubinont C, Debieve F. Impaired vascular endothelial growth factor expression and secretion during in vitro differentiation of human primary term cytotrophoblasts. Angiogenesis. 2020;23(2):221–230. doi:10.1007/s10456-019-09702-z
50. Zhang Q, Agoston AT, Pham TH. Acidic bile salts induce epithelial to mesenchymal transition via vegf signaling in non-neoplastic barrett’s cells. Gastroenterology. 2019;156(1):130–144.e10. doi:10.1053/j.gastro.2018.09.046
51. CJ G, Xie F, Zhang B. High glucose promotes epithelial-mesenchymal transition of uterus endometrial cancer cells by increasing ER/GLUT4-mediated vegf secretion. Cellular Physiology Biochemistry. 2018;50(2):706–720. doi:10.1159/000494237
52. Horikawa N, Abiko K, Matsumura N, et al. Expression of vascular endothelial growth factor in ovarian cancer inhibits tumor immunity through the accumulation of myeloid-derived suppressor cells. Clinical Cancer Research. 2017;23(2):587–599.
53. HPT N. N-cadherin identifies human endometrial epithelial progenitor cells by in vitro stem cell assays. Human Reproduction. 2017;32(11):2254–2268.
54. Taki M, Abiko K, Baba T. Snail promotes ovarian cancer progression by recruiting myeloid-derived suppressor cells via CXCR2 ligand upregulation. Nature Communications. 2018;9(1):1685. doi:10.1038/s41467-018-03966-7
55. Bu Z, Wang K, Dai W, Sun Y. Endometrial thickness significantly affects clinical pregnancy and live birth rates in frozen-thawed embryo transfer cycles. Gynecological Endocrinology. 2016;32(7):524–528. doi:10.3109/09513590.2015.1136616
56. Simeonov M, Sapir O, Lande Y, et al. The entire range of trigger-day endometrial thickness in fresh IVF cycles is independently correlated with live birth rate. Reprod Biomed Online. 2020;41:239–247. doi:10.1016/j.rbmo.2020.04.008
57. Bai R, Kusama K, Nakamura K, et al. Down-regulation of transcription factor OVOL2 contributes to epithelial-mesenchymal transition in a noninvasive type of trophoblast implantation to the maternal endometrium. FASEB J. 2018;32(6):3371–3384.
58. Zhang J, Mo HQ, Tian FJ, et al. EIF5A1 promotes trophoblast migration and invasion via ARAF-mediated activation of the integrin/ERK signaling pathway. Cell Death Dis. 2018;9(9):926. doi:10.1038/s41419-018-0971-5
59. Tian FJ, Cheng YX, Li XC, et al. The YY1/MMP2 axis promotes trophoblast invasion at the maternal-fetal interface. J Pathol. 2016;239(1):36–47. doi:10.1002/path.4694
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.