Back to Journals » Infection and Drug Resistance » Volume 11
Prevalence of resistance-associated substitutions to direct-acting antiviral agents in hemodialysis and renal transplant patients infected with hepatitis C virus
Authors Tavares RCF, Feldner AC, Pinho JR ![]() , Malta FM
, Malta FM ![]() , Carvalho-Filho RJ
, Carvalho-Filho RJ ![]() , Santana RA
, Santana RA ![]() , Fusco Duarte de Castro V, Dastoli GT, Custódio Lima J
, Fusco Duarte de Castro V, Dastoli GT, Custódio Lima J ![]() , Ferraz ML
, Ferraz ML ![]()
Received 7 May 2018
Accepted for publication 30 July 2018
Published 25 October 2018 Volume 2018:11 Pages 1993—2000
DOI https://doi.org/10.2147/IDR.S169512
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Rita Chelly Felix Tavares,1 Ana Cristina de Castro Amaral Feldner,1 João Renato Rebello Pinho,2,3 Fernanda de Mello Malta,3 Roberto José Carvalho-Filho,1 Rúbia Anita Ferraz Santana,2 Vanessa Fusco Duarte de Castro,2 Gregório Tadeu Fernando Dastoli,2 Juliana Custódio Lima,1 Maria Lucia Cardoso Gomes Ferraz1
1Gastroenterology Division, Federal University of Sao Paulo, São Paulo, SP, Brazil; 2Albert Einstein Diagnostic Medicine, Albert Einstein Hospital São Paulo, SP, Brazil; 3Laboratory of Tropical Gastroenterology and Hepatology “João Alves de Queiroz and Castorina Bittencourt Alves,” Institute of Tropical Medicine, Department of Gastroenterology, University of São Paulo, São Paulo, SP, Brazil
Background: Direct-acting antiviral agents (DAAs) permit the use of interferon (IFN)-free regimens to treat hepatitis C (HCV) in patients with chronic kidney disease (CKD) on hemodialysis (HD) or renal transplant (RTx) recipients, with excellent response rates and safety. However, the occurrence of basal or therapy-induced resistance-associated substitutions (RASs) to DAAs can result in treatment failure. The aim of this study was to estimate the prevalence of RASs to NS3A, NS5A and NS5B inhibitors, and particularly the Q80K polymorphism, in CKD patients on HD and RTx recipients infected with HCV.
Patients and methods: HD and RTx patients infected with HCV-genotype 1 (GT1) were subjected to sequencing of the NS3, NS5A and NS5B regions.
Results: Direct sequencing of NS3 protease, NS5A and NS5B was performed in 76 patients (HD, n=37; RTx, n=39). The overall prevalence of RASs was 38.2%, but only 5.3% of the patients had mutations in more than one region. Substitutions were detected in NS3A (17.8%), NS5A (21.9%) and NS5B (8.4%). Q80K was detected in 1.5 % of the patients. Highly inhibitory RASs were uncommon (L31M, 2.6%; L159F+C316N, 2.6%). RASs were more prevalent in HCV-GT1a (42.9%) than in HCV-GT1b (32.4%), P=0.35. RASs were detected in 52.4% of treatment-naive patients and 27.8% of peg-IFN/ribavirin-experienced patients (P=0.12). The presence of RASs was associated with time of RTx (P=0.01).
Conclusion: The Q80K polymorphism was uncommon in our sample of HD and RTx patients. Despite the high prevalence of naturally occurring RASs, most of the substitutions detected were associated with a low level of resistance to DAAs.
Keywords: HCV, treatment, DAA resistance, hepatitis
Erratum for this paper has been published
Introduction
The prevalence of chronic infection with hepatitis C virus (HCV) in patients with chronic kidney disease (CKD) on hemodialysis (HD) continues to be higher than in the general population, ranging from 3.8% to 27.3% depending on the geographic region.1 The main reasons for the higher prevalence of HCV infection in this population are previous exposure to blood and blood products, nosocomial transmission of HCV in HD units and longer time on dialysis.2 Genotype 1 (GT1) predominates in dialysis patients and renal transplant (RTx) recipients, regardless of geographic distribution, and subtype 1a is the most frequent.3,4
Chronic infection with HCV is an independent risk factor for increased mortality in dialysis patients,5 and is associated with cirrhosis complications, development of hepatocellular carcinoma and death due to cardiovascular diseases.6 In RTx recipients, the presence of HCV is an independent risk factor for graft loss7 and increased morbidity and mortality.8
The treatment of HCV has progressed considerably in the last two decades, particularly in the last 3 years. In this respect, the better understanding of the replication mechanisms of HCV resulted in the development of direct-acting antiviral agents (DAAs), which specifically block viral proteins.9 The first DAAs introduced were first-wave NS3 protease inhibitors (eg, telaprevir and boceprevir), which are used in triple combination therapy with pegylated-interferon (IFN) and ribavirin (RBV). However, their use was limited, especially in the CKD population because of significant worsening of anemia.10 The second generation of DAAs permitted the use of IFN-free regimens resulting from the combination of two or three drugs acting at different sites, such as protease inhibitors, NS5A inhibitors, and nucleotide and non-nucleotide inhibitors of NS5B. These treatments became promising for both HD patients and RTx recipients, showing high sustained virological response (SVR) rates without considerably increasing adverse effects or renal graft rejection.11–14
However, concern related to the use of DAAs is the emergence of resistance-associated substitutions (RASs), which have resulted in the failure of many treatment strategies. These substitutions can be naturally occurring or induced and might be selected during treatment.15 Inherent characteristics of HCV, such as a high viral replication rate and the lack of RNA polymerase-dependent proofreading mechanisms,16,17 associated with the genetic barrier characteristics of each inhibitor, contribute to the short-term development of these RASs whose frequency, duration and inhibitory potential are variables.18–22
Many studies have reported the prevalence of naturally occurring RASs in different populations, but there are no sequencing data of RASs to DAAs for the specific dialysis and RTx populations. Therefore, the aim of this study was to estimate the prevalence of naturally occurring RASs to DAAs in CKD patients on HD and RTx recipients infected with HCV, and to determine the association between the presence of RASs and demographic and clinical characteristics.
Patients and methods
Patients
From August 2014 to January 2015, patients of both sexes (age: 18–75 years) with CKD undergoing HD and RTx recipients infected with HCV-GT1 (anti-HCV and positive HCV PCR), seen at the Hepatitis Sector of Hospital São Paulo (UNIFESP, São Paulo, Brazil), were consecutively included in the study. Patients coinfected with HIV and HBV and those treated previously with DAAs were excluded. The study was approved by the local Ethics Committee (Comitê de Ética em Pesquisa Unifesp/HSP-HU) with the number 292365/2014. All patients signed the free informed consent form before the collection of data and serum.
NS3, NS5A and NS5B amplification
Viral RNA was extracted from 200 µL of each plasma sample using the QIAamp MinElute Virus Spin Kit (Qiagen, Hilden, Germany) according to manufacturer recommendations. cDNA synthesis and the first round of PCR of the NS3, NS5A and NS5B genes were performed using the SuperScript III One-Step RT-PCR system with Platinum Taq High Fidelity DNA Polymerase (Thermo Fisher Scientific, Waltham, MA, USA). Next, a nested PCR was performed using Platinum Taq DNA Polymerase (Thermo Fisher Scientific). The reaction conditions were as follows: 50°C for 30 minutes; 94°C for 2 minutes; 40 cycles at 94°C for 15 seconds, primer annealing for 30 seconds and 68°C for 3 minutes, and 68°C for 5 minutes. The annealing temperatures of NS3, NS5A and NS5B primers are shown in Table 1.23–25 Positive and negative controls were always included to control for possible contaminations.
 | Table 1 NS5A and NS5B primers used to amplification Notes: Data from Ewing et al28 and Tamura et al.29 Abbreviation: NS, nonstructural protein. |
Sequencing by the Sanger method
The nested PCR products were purified with 4 µL of ExoSAP-IT reagent (Affymetrix, Santa Clara, CA, USA) according to manufacturer instructions. A cycle-sequencing reaction was performed using the BigDye® Terminator v3.1 Kit (Thermo Fisher Scientific) and the same primers as employed in the nested PCR. Direct sequencing by the Sanger method26 was performed in an ABI 3500 genetic analyzer (Thermo Fisher Scientific). The sequences were generated from the two DNA strands to ensure the best quality of the results obtained.
Sequence analysis
The sequences obtained were first analyzed using the Phred-Phrap-Consed programs.27,28 The Phred program measures the quality of the data by attributing a score of 0–50 to each base of the chromatogram. Next, the Phrap program assembles a consensus sequence from the alignment of various overlapping sequences validated by Phred. Bases with a score ≥20 are considered for assembly of the consensus sequence. HCV genotype/subtype was determined by NS5B sequence analysis using maximum likelihood phylogenetic tree 1000 replicates (bootstrap values ≥70% were considered significant; data not shown) with the MEGA 5 software;29 reference sequences were downloaded from NCBI Genbank. In the cases where the NS5B region was not amplified, 5′UTR and NS3 sequences were used to genotyping. The resistance substitutions were determined by aligning the sequences with reference genomes NC_004102 for GT-1a and AJ238799 for GT-1b using the MEGA 5.29 The nucleotides were translated into amino acids and analyzed regarding the presence of substitutions previously identified to confer resistance clinically relevant to NS3 protease inhibitors (positions: 36, 54, 55, 80, 155, 156 and 168)30,31 and NS5A (positions: 28, 30, 31 and 93) and NS5B inhibitors (positions: 159, 282 and 316).22,30–33
Statistics analysis
Patients with and without RASs to DAAs were compared regarding age, sex, GT1 subtype, type of population (HD x RTx), duration of infection, length of time on HD, time after RTx and previous treatment without DAAs. The SPSS 20 software (IBM Corporation, Armonk, NY, USA) was used for statistical analysis. Descriptive statistics included absolute frequencies, means and SD. The Mann–Whitney test was applied for variables with non-normal distribution. Categorical variables were compared by the chi-squared test or Fisher’s exact test, adopting a level of significance of P-value less than 0.05 for all tests.
Results
Seventy-six patients (60.5% males, mean age of 52.2±9.9 years) were included in the study. Thirty-seven (48.7%) had CKD and were on HD and 39 (51.3%) were RTx recipients. Genotype 1 was detected in all patients included, with GT1a in 55.3% and GT1b in 48.7%. There were 52.6% treatment-naive patients and 47.4% Peg-IFN/RBV-experienced patients (Table 2).
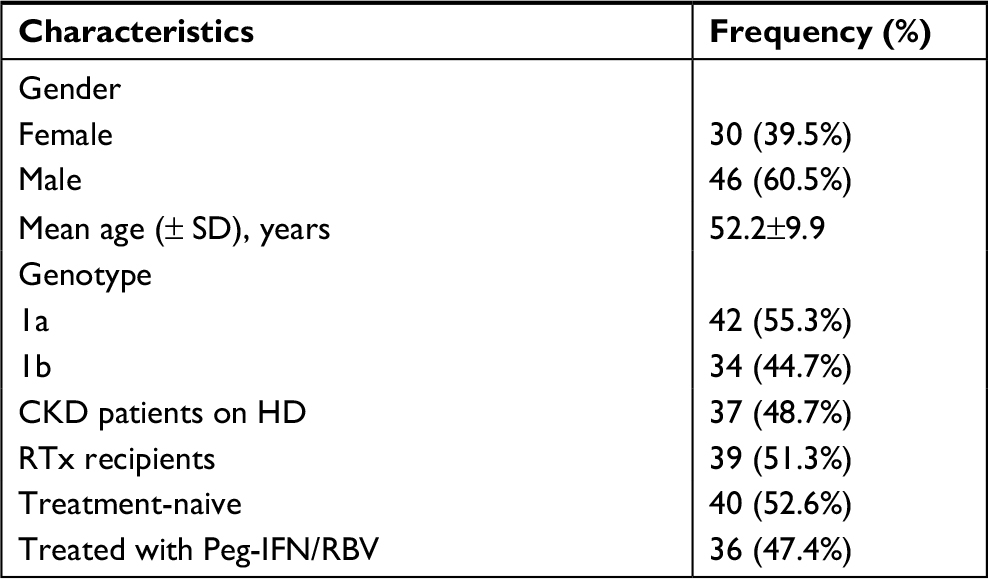 | Table 2 Demographic and epidemiologic characteristics of the population studied (n=76) Abbreviations: CKD, chronic kidney disease; HD, hemodialysis; Peg-IFN, pegylated interferon; RBV, ribavirin; RTx, renal transplant. |
Normal levels of alanine aminotransferase and aspartate aminotransferase were observed in CKD patients on HD, while these levels were elevated (1.2 times the upper limit of normal) in RTx recipients. Analysis of mean values and SD of the international normalized ratio (1±0.1 and 1.1±0.1), albumin (4±0.4 and 3.9±0.6 g/dL) and total bilirubin (0.5±0.3 and 0.7±0.4 mg/dL) in HD patients and RTx recipients, respectively, showed good hepatocellular function in both groups.
The immunosuppression scheme among RTx patients was based on calcineurin inhibitors in 30 patients (77%): with cyclosporin in 15 and with tacrolimus in 15. Other combinations (azathioprine and prednisone and mycophenolate and prednisone) were used in nine patients (23%).
Among the 76 patients evaluated, direct sequencing and amplification of NS3 protease were obtained for 64, of the NS5A region for 73 and of the NS5B region for 72. The overall prevalence of naturally occurring RASs was 38.2% (29/76). Only 5.3% of the patients had combined mutations in two regions (NS3A+NS5A) and none of them exhibited combined mutations in three regions. The following RASs in the NS5A region were the most frequent, observed in 21.9% of the patients: Q30 A/H/L; L31M; H58P/R; P58R/S; and P58S+Y93F (Table 3). RASs to NS3A protease inhibitors were detected in 17.2% of the participants (V55A; V36L; I170L; T54S; S122N; M175L; and Q80K) and substitutions in the NS5B region in only 8.4% (C316N; L159F+C316N) (Tables 4 and 5). In addition, the frequency of RASs conferring high-level resistance, including the Q80K polymorphism (1.6%), L31M (2.6%) and L159F and C316N (2.6%), was low.
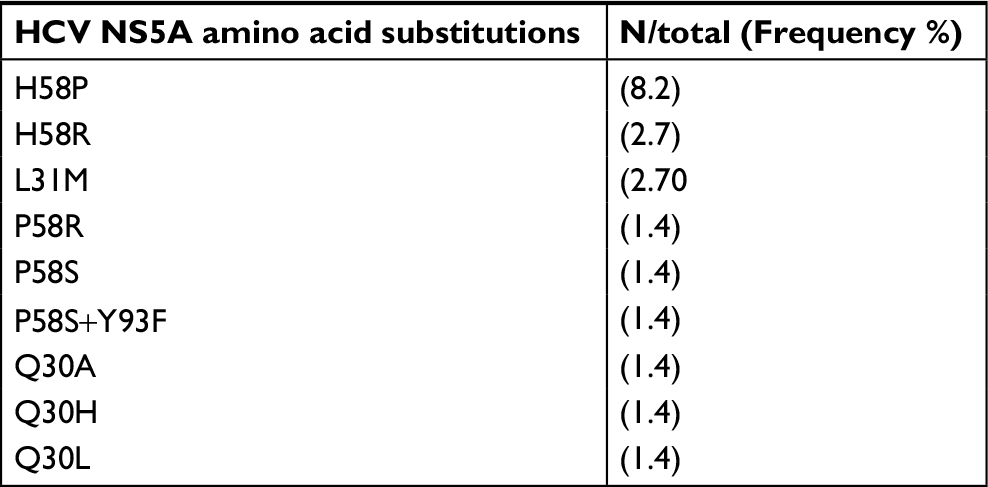 | Table 3 Frequency of resistance-associated substitutions in the NS5A region (n=73) Abbreviation: NS, nonstructural protein. |
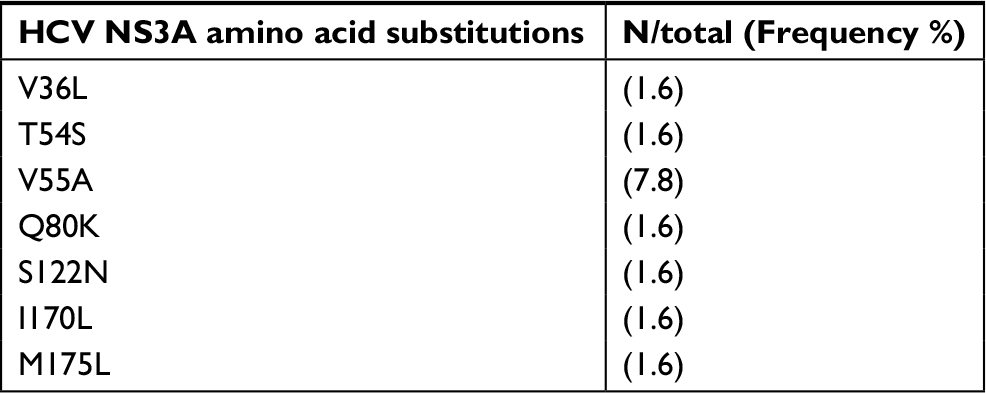 | Table 4 Frequency of resistance-associated substitutions to NS3A protease inhibitors (n=64) Abbreviation: NS, nonstructural protein. |
 | Table 5 Frequency of resistance-associated substitutions in the NS5B region (n=73) Abbreviation: NS, nonstructural protein. |
Table 6 shows the results of comparison of the different variables between patients with and without RASs. These substitutions were detected at the same frequency in males (37%) and females (40%), and the mean age was similar for patients with and without mutations (54.7 vs 55 years).
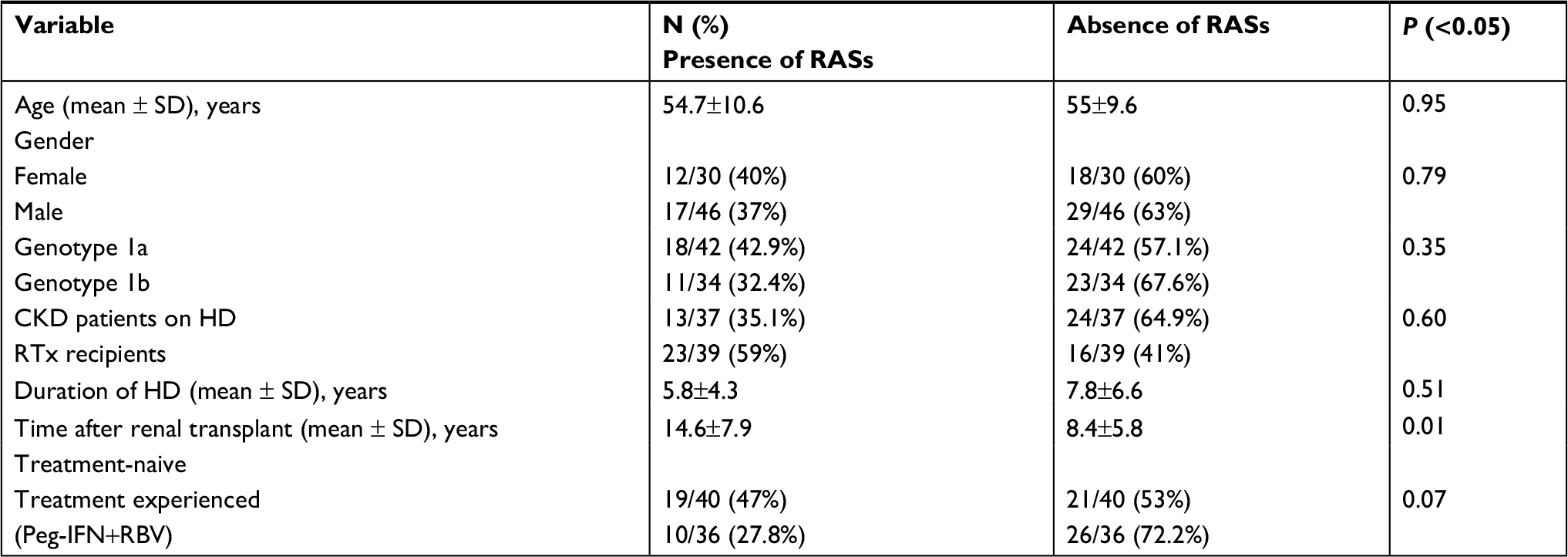 | Table 6 Comparative analysis of demographic and epidemiological variables related to hepatitis C virus according to the presence or absence of RASs to direct-acting antiviral agents (n=76) Abbreviations: RASs, resistance-associated substitutions; CKD, chronic kidney disease; HD, hemodialysis; RTx, renal transplant; Peg-IFN, pegylated interferon; RBV, ribavirin. |
RASs were found in 35.1% of CKD patients on HD and in 59% of RTx recipients, with no significant difference between groups (P=0.6). There was no association between the length of time on HD and presence of RASs, but a significant association was observed between the time after transplant and presence of RASs (P=0.01). The mean duration of HCV infection (in years), defined as the date of first transfusion or onset of HD, was similar in patients with and without mutations (P=0.7).
Comparison of the prevalence of RAS between patients immunosuppressed with calcineurin inhibitors (77%) vs patients immunosuppressed with non-calcineurin inhibitors (23%) showed a numerical difference in RAS frequency: 68% vs 31.2%, but without statistical significance (P=0.09)
With respect to treatment, RASs were detected in 47.5% (19/40) of patients naive to any treatment and in 27.8% (10/36) of Peg-IFN/RBV-experienced patients, with the difference being not significant (P=0.07)
The distribution of RASs differed according to GT1 subtype, but the difference was not significant (P=0.35). In this respect, 42.9% (18/42) of patients with GT1a carried the following RASs: NS3A: V55A, Q80K, I170L, T54S and S122N; NS5A: Q30A, Q30H, Q30L, L31M, H58P and H58R. By contrast, 32.4% (11/34) of patients with GT1b had the following mutations: NS3A: V55A, V36L and M175L; NS5A: L31M, P58R, P58S and P58S+Y93F; NS5B: C316N and L159F+C316N. V55A and L31M were observed in both subtypes and mutations in the NS5B region were only found in GT1b.
The only patient carrying the Q80K polymorphism (GT1a) was an RTx recipient and Peg-IFN/RBV-experienced who had not used protease inhibitors.
We did not detect other RASs conferring a high level of resistance, such as R155K and A156T, which are more implicated in resistance to protease inhibitors, Y93H (resistance to NS5A), S282T (resistance to sofosbuvir), or any mutation related to resistance to paritaprevir (Y56H and D168 A/V/Y).
Discussion
Until recently, the treatment of HCV in HD patients and RTx recipients was based on the use of IFN and RBV. In HD patients RBV is associated to poor tolerance, even with the use of low doses of the drug24 and in RTX recipients IFN is associated with a percentage of graft rejection.25 The use of IFN-free treatment regimens has opened up new perspectives for patients on HD and RTx recipients. Studies on DAAs involving these special populations are scarce, but promising results are available for HD and RTx patients. Even in the case of sofosbuvir regimens whose use is limited in patients with estimated glomerular filtration rate <30%,34 several studies report a considerable increase in the SVR (>90%) in the absence of significant worsening of renal refunction in most cases and tolerable adverse events.11–13,35,36
However, the emergence of RASs to DAAs, either naturally occurring or therapy induced, has resulted in the failure of some treatments.22,34,37–39
Treatment-induced viral resistance can be defined as the positive selection of viral variants with reduced susceptibility to DAAs.40 The presence of naturally occurring RASs to DAAs has been reported in patients from Europe, the US and Japan, while there are few studies demonstrating naturally occurring GT1 mutations in Latin America.
No study has so far evaluated the occurrence of these mutations in the exclusive population of CKD patients on HD and RTx recipients. The present study evaluated the frequency of these RASs in this group of patients in which infection with HCV is more prevalent and GT1a predominates.1,3,17
In the present study, direct sequencing of the NS3A (n=64), NS5A (n=73) and NS5B (n=72) regions in CKD patients on HD and RTx recipients demonstrated an overall frequency of naturally occurring RASs of 38.2%. Only 5.3% of the participants had combined mutations in two regions (NS3A+NS5A). The frequencies of RASs in the NS3A, NS5A and NS5B regions were 17.2%, 21.9% and 8.4%, respectively.
These rates are higher than those reported in the literature for patients without kidney disease. On the other hand, the prevalence of combined RASs in different regions was similar to previously reported data.15,20,39,41,42 This difference might be attributed to the deficient immune response observed in HD patients and RTx recipients,43 which could reduce the natural clearance of these emerging substitutions during viral replication. HCV has a high replication activity, and a large number of viral variants are continuously produced. These variants should be cleared by the immune system.44 Another paper from our group demonstrated that HCV genome variability and number of mutations were higher in HD patients when compared to non-HD patients.40
There was no significant difference between the prevalence of RAS when comparing HD to RTx, but numerically the number of RAS was higher among RTx patients. Furthermore, the prevalence was significantly associated to time of transplantation, suggesting the role of the duration of the immunosuppression, more than type or intensity, on the emergence of RAS. Regarding the type of immunosuppression, there was no difference on the emergence of RAS when comparing calcineurin inhibitors and non-calcineurin inhibitors schemes. There is no available information in the literature about the role of these specific drugs on RAS occurrence, and the effect of immunosuppression on the emergence of hepatitis C RASs is still under debate. In HIV/HCV patients with variable degrees of impairment of immune response, the prevalence of basal RAVs seems to be no different from that observed in immunocompetent monoinfected patients.45
The individual frequencies of each RAS (Tables 3–5) were similar to those reported in the literature for patients infected with GT1 without kidney disease.21,34,37,46,47 Furthermore, the types of natural RAS observed in NS3, NS5A and NS5B HCV proteins are in line with those described in previous studies in the same region.41–44
With respect to the impact of naturally occurring RASs on treatment, phenotypic analyses have shown that some substitutions confer a high level of resistance in vitro and have a greater impact in clinical practice.18–21,48 In the present study, among the RAAs found, the prevalence of those conferring high-level resistance, such as the Q80K polymorphism (1.6%) and substitutions L31M (2.6%) and L159F+C316N (2.6%), was low.
The global prevalence of the Q80K polymorphism ranges from 3% to 47% depending on the geographic region. In North America, this polymorphism is found in 48% of patients infected with HCV-GT1a. In South America and Europe, the polymorphism occurs in 9% and 19% of patients, respectively.20,47,49 The Q80K polymorphism is found almost exclusively in patients carrying GT1a and acts by reducing in vitro the susceptibility to simeprevir, with an 11-fold increase in the IC50% and a negative impact on the SVR.48,50 In the present study, the Q80K polymorphism was detected in only 1.6% of patients, corroborating the results of two previous studies conducted in the same geographic region that found this polymorphism to be uncommon.51,52
We detected no other RAAs that confer high-level resistance to NS5A inhibitors such as Y93H or S282T related to resistance to sofosbuvir, which are associated with high rates of treatment failure if present.18,21,39 Likewise, no amino acid substitution related to resistance to paritaprevir (D168A/V/Y and Y56H) was detected. This drug is part of IFN-free 3D regimens that are safe for use in patients with CKD.22
The frequency of RASs differed between GT1 subtypes, with a slightly higher frequency in patients carrying GT1a (42.9%) compared to GT1b (32.4%). Some RASs in the NS3A and NS5A regions occurred in both subtypes (V55A and L31M). Genotype GT1a has a lower genetic barrier than GT1b and consequently exhibits a higher frequency of RASs. Several studies have shown a higher SVR in patients with GT1b compared to those with GT1a.9,19,22,38 However, some RASs related to resistance to NS5B inhibitors are detected specifically in patients with GT1b, such as C316N.47,49,53 In the present study, all RASs in the NS5B region were only detected in patients carrying GT1b, with C316N being the most frequent substitution in the region studied.
The frequency of RASs was significantly higher in patients with a longer time after RTx, a finding supporting the hypothesis that a greater state of immunosuppression may reduce the natural clearance of these substitutions that emerge during viral replication.54 However, there was no association between the presence of RASs and the length of time on dialysis or duration of HCV infection. Furthermore, we observed that RASs were present in patients naive to any treatment (52.4%) and in those previously treated with Peg-IFN and RBV (27.8%). Thus, no significant association was found between the presence of mutations and previous exposure to Peg-IFN and RBV, as also demonstrated in other studies.15,32,55,56
This study has some limitations. The study included only GT1 patients, and the importance of RAS in GT3 patients was not evaluated. The number of studied patients was small, and we could not amplify the three regions of the genome in all patients. The differences between HD patients and Rtx patients have to be confirmed in other studies as well as the differences when comparing these special populations with non-immunocompromised patients.
Conclusion
In summary, this is the first study to analyze the presence of RASs to DAAs used for the treatment of hepatitis C in a population consisting exclusively of CKD patients on HD and RTx recipients infected with HCV-GT1. In this population, the presence of RASs was common and the prevalence of the Q80K polymorphism was low. Although naturally occurring RASs are more frequently detected in patients without kidney disease, most of the RASs found conferred low-level resistance to DAAs, indicating a high chance of response to treatment in this special population. RASs to protease and NS5A inhibitors predominated in patients with GT1a, while those in the NS5B region were exclusively found in patients with GT1b. A significant association was observed between the presence of substitutions and time after RTx, demonstrating the role of the immune system in the control of RAS emergence. Further studies are needed to increase our understanding of the impact of these resistance substitutions in the population of CKD patients on HD and RTx recipients.
Acknowledgment
The authors would like to thank Coordination for the Improvement of Higher Level- or Education- Personnel from Brazil government for the financial support.
Disclosure
The authors report no conflicts of interest in this work.
References
Kidney Disease: Improving Global Outcomes (KDIGO). KDIGO clinical practice guideline on the prevention, diagnosis, evaluation and treatment of hepatitis C in CKD. Public review draft; February 2017. Available from: https://kdigo.org/clinical_practice_guidelines/Hep%20C/KDIGO%202017%20Hep%20C%20GL%20Public%20Review%20Draft%20FINAL.pdf. Accessed October 5, 2018. | ||
Etik DO. Serkan Ocal, and Ahmet Sedat Boyacioglu. Hepatitis C infection in hemodialysis patients: A review. World J Hepatol. 2015;7(6):885–895. | ||
Perez RM, Ferraz ML, Figueiredo MS, et al. Unexpected distribution of hepatitis C virus genotypes in patients on hemodialysis and kidney transplant recipients. J Med Virol. 2003;69(4):489–494. | ||
Khedmat H, Amini M, Ghamar-Chehreh ME, Agah S. Hepatitis C virus infection in dialysis patients. Saudi J Kidney Dis Transpl. 2014;25(1):1–8. | ||
Fabrizi F, Takkouche B, Lunghi G, Dixit V, Messa P, Martin P. The impact of hepatitis C virus infection on survival in dialysis patients: meta-analysis of observational studies. J Viral Hepat. 2007;14(10):697–703. | ||
Marinaki S, Boletis JN, Sakellariou S, Delladetsima IK. Hepatitis C in hemodialysis patients. World J Hepatol. 2015;7(3):548–558. | ||
Fabrizi F, Martin P, Dixit V, Messa P. Meta-analysis of observational studies: hepatitis C and survival after renal transplant. J Viral Hepat. 2014;21(5):314–324. | ||
Morales JM, Fabrizi F. Hepatitis C and its impact on renal transplantation. Nat Rev Nephrol. 2015;11(3):172–182. | ||
Poveda E, Wyles DL, Mena Álvaro, Mena A, et al. Update on hepatitis C virus resistance to direct-acting antiviral agents. Antiviral Res. 2014;108:181–191. | ||
Wiegand J, Maasoumy B, Buggisch P, et al. Letter: Telaprevir triple therapy in chronic hepatitis C genotype 1 patients receiving haemodialysis. Aliment Pharmacol Ther. 2014;39(11):1342–1344. | ||
Bhamidimarri KR, Martin P. Finally, safe and effective treatment options for hepatitis C in hemodialysis patients. J Hepatol. 2016;65(1):7–10. | ||
Sawinski D, Kaur N, Ajeti A, et al. Successful treatment of hepatitis C in renal transplant recipients with direct-acting antiviral agents. Am J Transplant. 2016;16(5):1588–1595. | ||
Singh T, Guirguis J, Anthony S, et al. Sofosbuvir-based treatment is safe and effective in patients with chronic hepatitis C infection and end stage renal disease: a case series. Liver Int. 2016;36(6):802–806. | ||
Fabrizi F, Donato FM, Messa P. Direct-acting antivirals for hepatitis C virus in patients on maintenance dialysis. Int J Artif Organs. 2017;40(10):531–541. | ||
Bartels DJ, Zhou Y, Zhang EZ, et al. Natural prevalence of hepatitis C virus variants with decreased sensitivity to NS3.4A protease inhibitors in treatment-naive subjects. J Infect Dis. 2008;15:800–807. | ||
Simmonds P, Bukh J, Combet C, et al. Consensus proposals for a unified system of nomenclature of hepatitis C virus genotypes. Hepatology. 2005;42(4):962–973. | ||
Smith DB, Bukh J, Kuiken C, et al. Expanded classification of hepatitis C virus into 7 genotypes and 67 subtypes: updated criteria and genotype assignment web resource. Hepatology. 2014;59(1):318–327. | ||
Fridell RA, Wang C, Sun JH, et al. Genotypic and phenotypic analysis of variants resistant to hepatitis C virus nonstructural protein 5A replication complex inhibitor BMS-790052 in humans: in vitro and in vivo correlations. Hepatology. 2011;54(6):1924–1935. | ||
Halfon P, Locarnini S. Hepatitis C virus resistance to protease inhibitors. J Hepatol. 2011;55(1):192–206. | ||
Paolucci S, Fiorina L, Piralla P, et al. Naturally occurring mutations to HCV protease inhibitors in treatment naïve patients. Virol J. 2012;24:245. | ||
Pawlotsky JM. NS5A inhibitors in the treatment of hepatitis C. J Hepatol. 2013;59(2):375–382. | ||
Lontok E, Harrington P, Howe A, et al. Hepatitis C virus drug resistance-associated substitutions: State of the art summary. Hepatology. 2015;62(5):1623–1632. | ||
Peres-da-Silva A, de Almeida AJ, Lampe E. Mutations in hepatitis C virus NS3 protease domain associated with resistance to specific protease inhibitors in antiviral therapy naïve patients. Arch Virol. 2010;155(5):807–811. | ||
Brennan BJ, Wang K, Blotner S, et al. Safety, tolerability, and pharmacokinetics of ribavirin in hepatitis C virus-infected patients with various degrees of renal impairment. Antimicrob Agents Chemother. 2013;57(12):6097–6105. | ||
Kim E, Ko HH, Yoshida EM. Treatment issues surrounding hepatitis C in renal transplantation: a review. Ann Hepatol. 2011;10(1):5–14. | ||
Sanger WG, Eisen JD. Cryogenically preserved human semen: clinical applications. Nebr Med J. 1977;62(12):422-4. | ||
Ewing B, Green P. Base-calling of automated sequencer traces using phred. II. Error probabilities. Genome Res. 1998;8(3):186–194. | ||
Ewing B, Hillier L, Wendl MC, Green P. Base-calling of automated sequencer traces using phred. I. Accuracy assessment. Genome Res. 1998;8(3):175–185. | ||
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28(10):2731–2739. | ||
Nettles RE, Gao M, Bifano M, et al. Multiple ascending dose study of BMS-790052, a nonstructural protein 5A replication complex inhibitor, in patients infected with hepatitis C virus genotype 1. Hepatology. 2011;54(6):1956–1965. | ||
Paolucci S, Fiorina L, Mariani B, et al. Naturally occurring resistance mutations to inhibitors of HCV NS5A region and NS5B polymerase in DAA treatment-naïve patients. Virol J. 2013;10(1):355. | ||
Paolucci S, Fiorina L, Mariani B, et al. Naturally occurring resistance mutations to inhibitors of HCV NS5A region and NS5B polymerase in DAA treatment-naïve patients. Virol J. 2013;10:355. | ||
Sandres-Sauné K, Deny P, Pasquier C, et al. Determining hepatitis C genotype by analyzing the sequence of the NS5b region. J Virol Methods. 2003;109(2):187–193. | ||
Sarrazin C. The importance of resistance to direct antiviral drugs in HCV infection in clinical practice. J Hepatol. 2016;64(2):486–504. | ||
Roth D, Nelson DR, Bruchfeld A, et al. Grazoprevir plus elbasvir in treatment-naive and treatment-experienced patients with hepatitis C virus genotype 1 infection and stage 4-5 chronic kidney disease (the C-SURFER study): a combination phase 3 study. Lancet. 2015;386(10003):1537–1545. | ||
Kamar N, Marion O, Rostaing L, et al. Efficacy and Safety of Sofosbuvir-Based Antiviral Therapy to Treat Hepatitis C Virus Infection After Kidney Transplantation. Am J Transplant. 2016;16(5):1474–1479. | ||
Kieffer TL, George S. Resistance to hepatitis C virus protease inhibitors. Curr Opin Virol. 2014;8:16–21. | ||
Welsch C. Genetic barrier and variant fitness in hepatitis C as critical parameters for drug resistance development. Drug Discov Today Technol. 2014;11:19–25. | ||
Pawlotsky JM. Hepatitis C Virus Resistance to Direct-Acting Antiviral Drugs in Interferon-Free Regimens. Gastroenterology. 2016;151(1):70–86. | ||
Alves da Silva R. Molecular characterization of hepatitis C virus in end-stage renal disease patients under hemodialysis. Journal of Medical Virology 90. 2018;3:537–544. | ||
Noble CF, Malta F, Lisboa-Neto G, et al. Natural occurrence of NS5B inhibitor resistance-associated variants in Brazilian patients infected with HCV or HCV and HIV. Arch Virol. 2017;162(1):165–169. | ||
Peres-da-Silva A, de Almeida AJ, Lampe E. NS5A inhibitor resistance-associated polymorphisms in Brazilian treatment-naive patients infected with genotype 1 hepatitis C virus. J Antimicrob Chemother. 2015;70(3):726–730. | ||
Peres-da-Silva A, Brandão-Mello CE, Lampe E. Prevalence of sofosbuvir resistance-associated variants in Brazilian and worldwide NS5B sequences of genotype-1 HCV. Antivir Ther. 2017;22(5):447–451. Epub 2017 Jan 13. | ||
Moreira RC, de Torres Santos AP, Lisboa-Neto G, et al. Prevalence of naturally occurring amino acid substitutions associated with resistance to hepatitis C virus NS3/NS4A protease inhibitors in São Paulo state. Arch Virol. 2018. | ||
Deng H, Deng X, Liu Y, et al. Naturally occurring antiviral drug resistance in HIV patients who are mono-infected or co-infected with HBV or HCV in China. J Med Virol. 2018;90(7):1246–1256. | ||
Patiño-Galindo JÁ, Salvatierra K, González-Candelas F, López-Labrador FX. Comprehensive Screening for Naturally Occurring Hepatitis C Virus Resistance to Direct-Acting Antivirals in the NS3, NS5A, and NS5B Genes in Worldwide Isolates of Viral Genotypes 1 to 6. Antimicrob Agents Chemother. 2016;60(4):2402–2416. | ||
Paolucci S, Fiorina L, Piralla P, et al. Naturally occurring mutations to HCV protease inhibitors in treatment naïve patients. Virol J. 2012;24:245. | ||
Mccloskey RM, Liang RH, Joy JB, et al. Global origin and transmission of hepatitis C virus nonstructural protein 3 Q80K polymorphism. J Infect Dis. 2015;211(8):1288–1295. | ||
Poveda E, Wyles DL, Mena A, Pedreira JD, Castro-Iglesias A, Cachay E. Update on hepatitis C virus resistance to direct-acting antiviral agents. Antiviral Res. 2014;108:181–191. | ||
Sarrazin C, Lathouwers E, Peeters M, et al. Prevalence of the hepatitis C virus NS3 polymorphism Q80K in genotype 1 patients in the European region. Antiviral Res. 2015;116:10–16. | ||
Peres-da-Silva A, Almeida AJ, Lampe E. Genetic diversity of NS3 protease from Brazilian HCV isolates and possible implications for therapy with direct-acting antiviral drugs. Mem Inst Oswaldo Cruz. 2012;107(2):254–261. | ||
de Carvalho IM, Alves R, de Souza PA, et al. Protease inhibitor resistance mutations in untreated Brazilian patients infected with HCV: novel insights about targeted genotyping approaches. J Med Virol. 2014;86(10):1714–1721. | ||
Ahmed A, Felmlee DJ. Mechanisms of hepatitis C viral resistance to direct acting antivirals. Viruses. 2015;7(12):6716–6729. | ||
Buti M, Riveiro-Barciela M, Esteban R. Management of direct-acting antiviral agent failures. J Hepatol. 2015;63(6):1511–1522. | ||
Kuntzen T, Timm J, Berical A, et al. Naturally occurring dominant resistance mutations to hepatitis C virus protease and polymerase inhibitors in treatment-naïve patients. Hepatology. 2008;48(6):1769–1778. | ||
Sede MM, Laufer NL, Quarleri J. Previous failure of interferon-based therapy does not alter the frequency of HCV NS3 protease or NS5B polymerase inhibitor resistance-associated variants: longitudinal analysis in HCV/HIV co-infected patients. Int J Antimicrob Agents. 2015;46(2):219–224. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.