Back to Journals » Infection and Drug Resistance » Volume 19
Prevalence, Antimicrobial Susceptibility, and Molecular Characteristics of Ceftazidime-Avibactam-Resistant Klebsiella pneumoniae in a Chinese Hospital
Authors Li J, Wang H, Liu Z, Xia Y, Hu Y, Wang H, Xia F ![]() , Zou M
, Zou M
Received 5 December 2025
Accepted for publication 23 January 2026
Published 3 March 2026 Volume 2026:19 586812
DOI https://doi.org/10.2147/IDR.S586812
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Hazrat Bilal
Jun Li,1,2,* Haolan Wang,1,* Zhaojun Liu,1 Yubing Xia,1 Yongmei Hu,1,2 Haichen Wang,1,2 Fengjun Xia,1,2 Mingxiang Zou1,2
1Department of Clinical Laboratory, Xiangya Hospital, Central South University, Changsha, Hunan, 410008, People’s Republic of China; 2National Clinical Research Center for Geriatric Diseases (Xiangya Hospital), Changsha, Hunan, 410008, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Mingxiang Zou, Department of Clinical Laboratory, Xiangya Hospital, Central South University, Changsha, Hunan, 410008, People’s Republic of China, Tel +86 13907496278, Email [email protected]
Background: The mechanisms underlying ceftazidime-avibactam resistance in carbapenem-resistant Klebsiella pneumoniae (CRKPCZA-R) remain to be elucidated.
Methods: CRKPCZA-R isolates were screened from non-repetitive CRKP isolates at our hospital from January 1, 2018 to October 30, 2021. The antimicrobial susceptibility and molecular characteristics of CRKPCZA-R were analyzed by broth microdilution method and next-generation sequencing, respectively.
Results: In total, 67 of 623 CRKP isolates (10.8%) were identified as CRKPCZA-R. The susceptibility rates of CRKPCZA-R to polymyxin B, tigecycline, aztreonam-avibactam, and cefiderocol were 97.0% (65/67), 83.6% (56/67), 100.0% (67/67), and 94.0% (63/67), respectively. The most prevalent resistance gene was blaNDM-1 (44.8%, 30/67), followed by blaIMP-4 (9.0%, 6/67), blaNDM-5 (7.5%, 5/67), and blaNDM-4 (1.5%, 1/67). Furthermore, 37.3% (25/67) of the CRKPCZA-R isolates co-harbored more than two carbapenemase-encoding genes, mainly blaNDM-1 and blaKPC-2 (31.3%, 21/67). The enzyme inhibitor enhancement method detected carbapenemase activity with high sensitivity, except for isolates carrying two or more carbapenemases. Notably, 21 KL64-ST11 CRKPCZA-R isolates presented blaNDM-1, blaKPC-2, and ompk36 deletion, and 17 co-harbored two or more high virulence gene markers. Patients infected with these 21 isolates were older and experienced more serious illness compared to those infected with other drug-resistant isolates.
Conclusion: The detection rate of CRKPCZA-R was relatively high due to the metallo-β-lactamase-producing isolates. Although enzyme inhibitor enhancement method can detect carbapenemases with high sensitivity and specificity, and provide an important reference for drug selection, it is not as effective for isolates carrying two or more carbapenemases. Patients infected with CRKPCZA-R co-harboring both blaKPC-2 and blaNDM-1 should be closely monitored.
Plain Language Summary: The resistance mechanisms of CRKPCZA-R remain to be uncovered. In our study, the detection rate of CRKPCZA-R was relatively high due to the metallo-β-lactamase-producing isolates. The sensitivity of CRKPCZA-R to antibacterial agents was evaluated in vitro, and revealed high susceptibility to aztreonam-avibactam, cefiderocol, polymyxin B, and tigecycline. The enzyme inhibitor enhancement method plays an important role in guiding the selection of clinical antibiotics due to its high sensitivity and specificity. Specifically, when the inhibition zone diameter of K-B method CZA is 20-22 mm, CZA can be used alone if it produces serine enzyme, but not if it produces metalloenzyme. Patients infected with CRKPCZA-R harboring both blaKPC-2 and blaNDM-1 were older and experienced more serious illness compared to those infected with other drug-resistant isolates, and should therefore be monitored more closely.
Keywords: carbapenem-resistant Klebsiella pneumoniae, ceftazidime–avibactam, resistance mechanism, homology
Introduction
The global prevalence of carbapenem-resistant Klebsiella pneumoniae (CRKP) has increased in recent years, resulting in big challenges in the treatment of bacterial infections.1–3 On May 17, 2024, CRKP was listed as part of the critical group in the latest priority list of bacterial pathogens issued by World Health Organization (WHO).4
Ceftazidime-avibactam (CZA) is a novel β-lactamase inhibitor consisting of ceftazidime (CAZ) and avibactam (AVI). With the increasing prevalence of infections caused by bacterial isolates resistant to traditional antibiotics, there is an urgent need to design more effective therapeutic options.5 CZA is effective against a range of pathogens, including Enterobacterales and Pseudomonas aeruginosa,6,7 and has been recommended by the Infectious Diseases Society of America (IDSA) for treating infections caused by CRKP. However, the widespread use of CZA has led to the emergence of CZA-resistant CRKP (CRKPCZA-R) isolates in several countries, and the resistance rate in some regions is as high as 21%.8,9
The most common mechanism driving CZA resistance is the production of metallo-β-lactamases (MBLs), including New Delhi MBL, imipenemase, and Verona integron-encoded MBL. As AVI cannot bind to MBLs and block the hydrolysis of CAZ, MBL-producing isolates are naturally resistant to CZA.10 Nevertheless, AVI can protect CAZ from serinases. Although CZA has been effective in the treatment of infections caused by carbapenemase-producing CRKP, studies show that KPC-producing isolates can develop resistance to CZA. Selective mutations in one or more amino acids in the Ω loop of KPC-2 and KPC-3 can render them impervious to the inhibitory action of AVI, resulting in novel CZA-resistant CRKP variants.11 Novel KPC subtypes, including KPC-33, KPC-121, and KPC-125, have been continuously reported among CZA-resistant CRKP isolates.12–14 The CRKP isolates producing KPC-31 and KPC-33 variants, which are the result of D179Y and H274Y mutations, respectively, are currently the most common isolates isolated in clinics worldwide.15 Furthermore, increased expression of the blaKPC gene,16 mutations in LamB (outer membrane maltoprotein), and high expression of blaSHV-12 have also been associated with CZA resistance.17,18
In this study, we screened CRKPCZA-R isolates among non-repetitive clinical isolates of CRKP collected at our hospital from January 1, 2018 to October 30, 2021. The drug resistance genes, virulence-related genes, and sequence types (STs) of these isolates, and the clinical data of infected patients were analyzed to clarify the epidemiological characteristics of CRKPCZA-R, and provide an experimental basis for treating infections caused by CZA-resistant isolates.
Materials and Methods
Bacterial Isolates
In order to clarify the epidemiological characteristics of CRKPCZA-R, all non-repetitive clinical CRKP isolates were collected at our hospital between January 1, 2018 and October 30, 2021, and re-identified by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (Zhongyuan Huiji Biotechnology Co. Ltd, China) using Escherichia coli ATCC 25922 as the quality-control isolate. Patient data were also collected, including age, sex, ward, clinical diagnosis, underlying diseases, complications, prior invasive procedures, clinical outcomes, and antibiotic usage before and after isolation.
The quality control isolates (E. coli ATCC 25922, K. pneumoniae ATCC 700603, and P. aeruginosa ATCC 27853) were purchased from National Center for Clinical Laboratories (Beijing, China).
Antimicrobial Susceptibility Testing
The K-B method was used to screen CZA-resistant CRKP isolates (30/20-μg Disk, Oxoid). For isolates with inhibition zone diameters of 20–22 mm, the respective minimum inhibitory concentrations (MICs) of CZA were determined by the microdilution broth method as per the guidelines of the Clinical and Laboratory Standards Institute (CLSI).19 The K. pneumoniae ATCC 700603 isolate was also included in the drug sensitivity test for quality control.
The MICs of meropenem (MEM), imipenem (IPM), polymyxin B (POL), tigecycline (TGC), fosfomycin (FOS), aztreonam-avibactam (AZA), and cefiderocol against CRKPCZA-R were determined by the broth microdilution method. The antibiotics were purchased from Wenzhou Kont Biotechnology Co. Ltd. The MIC of fosfomycin (FOS) was detected by the agar dilution method using medium supplemented with glucose-6-phosphate. The MIC of cefiderocol was determined using iron-free medium. E. coli ATCC 25922 and P. aeruginosa ATCC 27853 were employed as the quality-control isolates. The drug sensitivity results of cefiderocol, MEM, IPM, and ATM were interpreted according to the CLSI M100-34 criteria.19 The results for TGC were evaluated according to the breakpoint set by the US Food and Drug Administration,20 and the latest breakpoints from the European Committee on Antimicrobial Susceptibility Testing were applied for AZA and POL.21 Currently, there are no established sensitivity breakpoints for FOS.
Phenotype Detection of Carbapenemases
Carbapenemase phenotypes were detected using the enzyme inhibitor enhancement test as previously described.22 Boric acid (APB) and ethylene diamine tetraacetic acid (EDTA) were purchased from Wenzhou Kont Biotechnology Co. Ltd. The isolates were cultured in the following conditions: A (IPM), B [IPM + APB (400 μg)], C [IPM + EDTA (292 μg)], and D [IPM + APB (400 μg) + EDTA (292 μg)], and the diameters of their respective inhibition zones were measured. The carbapenemase type produced by an isolate was determined according to the inhibition zone diameter as follows: class A carbapenemase - difference between the inhibition zone diameters of disk B and disk A ≥ 5 mm; class B metalloenzymes - difference between the inhibition zone diameters of disk C and disk A ≥ 5 mm; both class A carbapenemase and class B metalloenzyme - difference between the inhibition zone diameters of disk D and disk A ≥ 5 mm; no carbapenemase production - difference between the inhibition zone diameters of disks B, C, and D and that of disk A < 5 mm.
Next-Generation Sequencing (NGS)
Genomic DNA was extracted from the CRKPCZA-R isolates using the Ezup Column Bacterial Genomic DNA Extraction Kit (Shanghai Sangong Bioengineering Co. Ltd.) according to the manufacturer’s instructions, and sequenced by Wuhan Baiyi Huineng using the double-ended 150-bp read length dnbseq platform. The raw data were filtered using fastp and subjected to quality control using FastQC. The sequencing data were assembled and corrected using Unicycler (parameter: −1r1-2r2--keep3--modenomal) to obtain the final bacterial genome sequences. The single-nucleotide polymorphisms (SNPs) of the core genomes were identified using Snippy software (version 2, https://github.com/tseemann/snippy), and the phylogenetic trees were constructed according to the maximum likelihood method. The MLST types, drug resistance genes, and virulence-related genes of CRKPCZA-R isolates were analyzed by BacWGSTdb (BacWGSTdb 2.0, http://bacdb.cn/BacWGSTdb/). The capsule locus (KL) types were analyzed by BIGSdb-Pasteur (version 3, https://bigsdb.pasteur.fr/). The phylogenetic trees, MLST types, drug resistance genes, virulence-related genes, and serotypes were visualized using ChiPlot (version 2.6.1, https://www.chiplot.online/).
Results
Screening and Confirmation of CZA-Resistant CRKP Isolates
In total, 623 non-repetitive CRKP isolates were collected. According to the results of the K-B method, 59 CRKP isolates were identified as CRKPCZA-R (inhibition zone < 20 mm) and 539 were confirmed as CZA-sensitive CRKP (inhibition zone > 22 mm). Notably, there were 25 isolates with inhibition zone diameters of 20–22 mm, and the MICs of CZA were further determined by the microdilution broth method. Accordingly, 8 of these 25 unconfirmed isolates (32.0%) were identified as CRKPCZA-R, and 17 isolates (68.0%) were identified as CZA-sensitive CRKP. Overall, 67 CRKPCZA-R isolates (67/623, 10.8%) were detected.
Sensitivity of CZA-Resistant CRKP to Other Antimicrobial Agents
We also tested the sensitivity of CRKPCZA-R isolates to other antimicrobial agents. The CRKPCZA-R isolates were resistant to all carbapenems, and resistance rates to POL, TGC, AZA, cefiderocol were 97.0% (65/67), 83.6% (56/67), 100.0% (67/67), and 94.0% (63/67), respectively. Furthermore, the MIC50 and MIC90 for AZA were 0.125/4 μg/mL and 0.5/4 μg/mL, respectively, and that for FOS were 128 μg/mL and >512 μg/mL, respectively. The MIC50 and MIC90 for cefiderocol were 2 μg/mL and 4 μg/mL, respectively, although two isolates (C142 and C387) exhibited low-level resistance with MICs of 16 μg/mL.
Identification of Carbapenemases and Mechanisms of Drug Resistance in CRKPCZA-R Isolates
The carbapenemase phenotypes of the 67 CRKPCZA-R isolates were tested, and the results indicated that all but one isolate (R2) produced at least one carbapenemase. In addition, 51 isolates were positive only for metalloenzymes, none were positive only for serine enzymes, and 15 isolates were positive for metalloenzymes and serine enzymes.
The predominant carbapenemase-encoding gene in these CRKPCZA-R isolates was blaNDM-1 (44.8%, 30/67), followed by blaIMP-4 (9.0%, 6/67) and blaNDM-5 (7.5%, 5/67), and one isolate carried blaNDM-4. In addition, 37.3% (25/67) of the CRKPCZA-R isolates simultaneously harbored more than two types of carbapenemase-encoding genes, of which 21 isolates (31.3%) co-harbored blaNDM-1 and blaKPC-2, two isolates simultaneously carried blaKPC-2 and blaIMP-4, one isolate (R2) carried blaKPC-2 and blaIMP-26, and one isolate (R57) carried blaKPC-2, blaNDM-1, and blaIMP-4. The outer membrane protein-coding genes, including ompk35, ompk36, and ompk37, were also analyzed. No deletions were detected in any of these genes in 41 isolates (61.2%), while ompk36 and ompk35 were deleted in 25 isolates and one isolate, respectively. Furthermore, 21 CRKPCZA-R isolates with ompk36 deletion carried both blaKPC-2 and blaNDM-1, and 67 CRKPCZA-R isolates also carried other drug-resistance genes (Figure 1).
 |
Figure 1 Phylogenetic tree, STs, resistance genes, KL types, and virulence-related genes in the 67 CRKPCZA-R isolates. Abbreviations: KL, capsule locus; MLST, multilocus sequence typing; NA, not determined. |
The Mechanism of Drug Resistance in CRKP Isolates with Inhibition Zone Diameters of 20-22 mm
Twenty-five CRKP isolates with inhibition zone diameters of 20–22 mm to CZA were rechecked by the MIC method. Seven of these isolates, which exhibited an inhibition zone of 20 mm in the K-B test, were confirmed to be CZA-resistant and positive for carbapenemases, and also carried metalloenzyme-encoding genes. In addition, seven isolates had an inhibition zone of 21 mm, of which only one isolate exhibited resistance to CZA through metalloenzyme production and carried the blaIMP-4 gene. The other isolates were sensitive to CZA and produced serinase. Finally, 11 isolates with an inhibition zone of 22 mm were confirmed to be CZA-sensitive after rechecking, and were positive for serinase (Table 1), suggesting that the diameter of the inhibition zone of CZA is between 20–22 mm using the disk diffusion method. Therefore, in case there is no condition to detect the MIC of CZA in clinical laboratories, the detection of carbapenemase types may provide an important reference for drug selection, that is, if serinase carbapenemase is produced, CZA can be used for treatment, but if metalloenzyme is produced, it cannot be used alone.
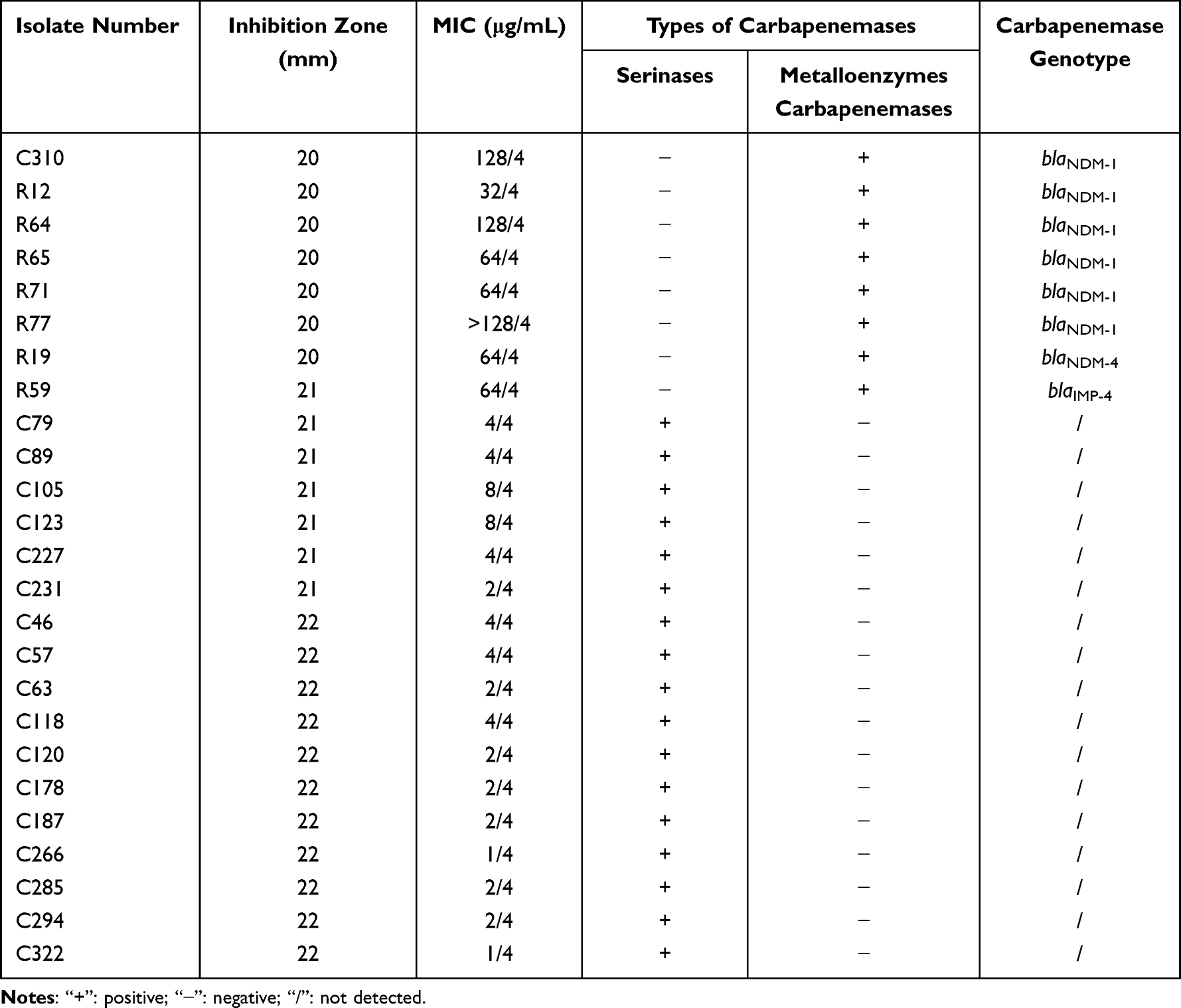 |
Table 1 The Drug Sensitivity, Carbapenemase Phenotypes, and Genotypes of 25 CRKP Isolates Should Be Rechecked by the MIC Method |
Homology Analysis
The results of MLST identified 27 STs among the 67 CRKPCZA-R isolates, whereas two isolates were not typed. ST11 was the most common type (35.8%, 24/67), followed by ST196 (7.5%, 5/67), ST20 (4.5%, 3/67), ST39 (4.5%, 3/67), and ST307 (4.5%, 3/67, Figure 1). In addition, 21 KL64-ST11 isolates belonged to the same clone cluster with ompk36 deletion, and co-harbored blaKPC-2 and blaNDM-1. Seventeen of these isolates co-harbored two or more hypervirulent marker genes, suggesting clonal transmission of hypervirulent KL64-ST11 CRKPCZA-R isolates within our hospital (Figure 1).
The Risk of Infection with KL64-ST11 CRKPCZA-R Isolates Harboring blaKPC-2 and blaNDM-1
Patients infected with CRKPCZA-R isolates co-harboring blaKPC-2 and blaNDM-1, and with ompk36 deletion, were generally older than those infected with other drug-resistant isolates. Notably, patients harboring these isolates often experienced more severe illness, resulting in higher ICU admission rates and a greater proportion of patients being discharged without recovery (Table 2).
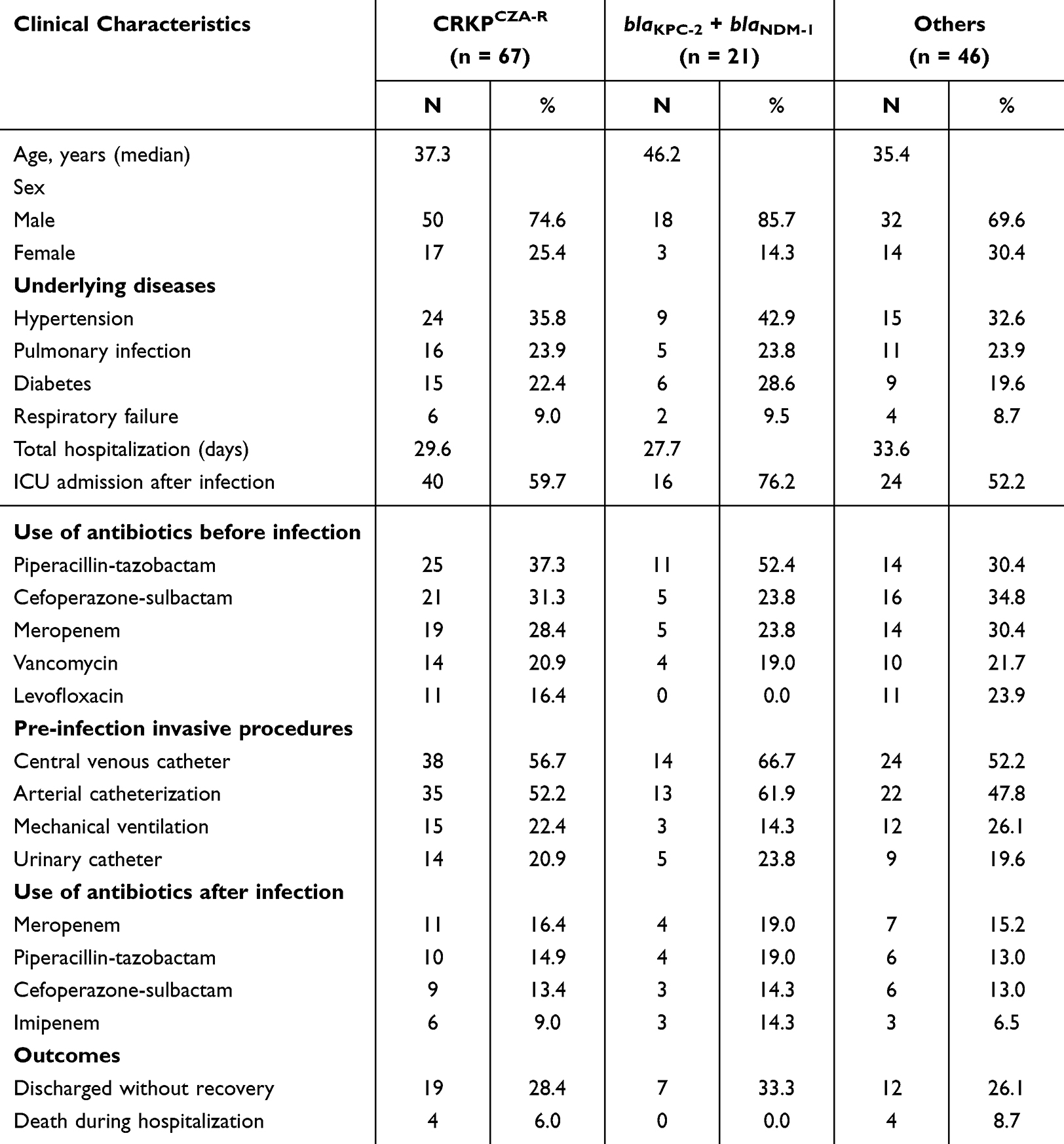 |
Table 2 Clinical Characteristics of Patients Infected with CRKPCZA-R with blaKPC-2 + blaNDM-1 and Others |
Interestingly, CRKPCZA-R isolates harboring both blaKPC-2 and blaNDM-1, and other resistance genes, exhibited similar MIC50 and MIC90 for IPM, MEM, TGC, POL, and AZA. However, the MIC50 for FOS ranged from 64 to 256 μg/mL, whereas no notable difference was observed in the MIC90. The MIC50 and MIC90 of cefiderocol for CRKPCZA-R carrying both blaKPC-2 and blaNDM-1 were lower compared to that for other isolates, and no intermediate or resistant isolates were identified, suggesting a potentially higher sensitivity to cefiderocol (Table 3).
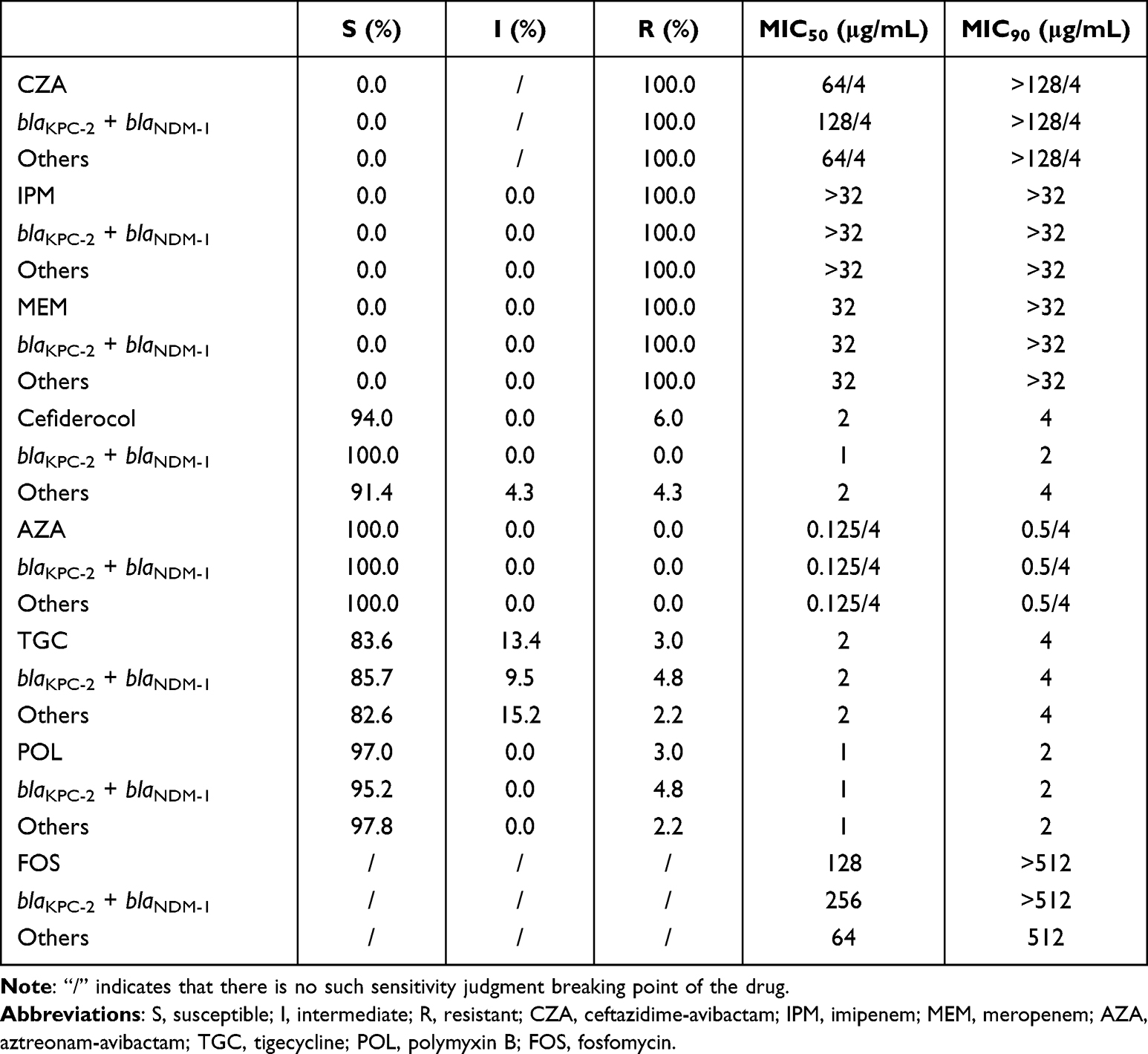 |
Table 3 Drug Sensitivity for CRKPCZA-R with blaKPC-2 + blaNDM-1 and Others |
We detected 27 serotypes, of which KL64 accounted for 37.3% of the isolates (25/67), followed by KL173 (7.5%, 5/67), KL23 (6.0%, 4/67), and KL46 (6.0%, 4/67). The marker genes of hypervirulent isolates, including rmpA, rmpA2, iroB, and iucA, were also analyzed. As shown in Figure 1, 32.8% (22/67) of the CRKPCZA-R isolates carried two or more hypervirulent marker genes, and four isolates carried iucA. None of these marker genes were detected in the remaining isolates. It is worth noting that 17/21 CRKPCZA-R isolates carrying blaKPC-2 and blaNDM-1 also harbored two or more hypervirulent marker genes and exhibited high virulence characteristics (Figure 1).
Discussion
The global emergence of CRKP in recent years has posed significant challenges to the treatment of bacterial infections.1–3,23 CZA, a novel β-lactamase inhibitor, has been recommended by the IDSA for the treatment of CRKP infections.6 However, its widespread use in clinics has led to the emergence of CZA-resistant isolates worldwide. In this study, we screened CRKPCZA-R isolates from the clinical isolates of CRKP collected at our hospital from January 1, 2018 to October 30, 2021. In addition, the clinical data of patients infected with CRKPCZA-R, and the drug resistance phenotypes and genotypes of these isolates were analyzed to clarify the clinical and molecular epidemiological characteristics of CRKPCZA-R infections.
CLSI recommends the K-B method and MIC test for detecting the sensitivity of bacterial isolates to CZA.19 Given its simplicity and affordability, the K-B method is more appropriate for screening CZA-resistant isolates, particularly in developing countries. Conversely, the MIC method is primarily used to confirm CZA resistance. In this study, 25 isolates with inhibition zone diameters of 20–22 mm in the K-B test were re-evaluated using the microdilution broth method; the isolates with an inhibition zone diameter of 20 mm were confirmed to be CZA-resistant, whereas those with zones measuring 22 mm were sensitive to CZA.23 However, one isolate with an inhibition zone of 21 mm was resistant to CZA, and it carried the blaIMP-4 gene and tested positive for carbapenemase. The sensitivity of this isolate could be attributed to PcW, an upstream promoter of blaIMP-4, which likely downregulated the carbapenemase gene.24 Therefore, in regions where CZA is not widely utilized, further verification using the MIC method is not required when the diameter of the inhibition zone of CZA is 20 or 22 mm. However, verification may be necessary when the diameter is 21 mm.
Since the treatment plans for carbapenem-resistant isolates depend on the types of carbapenemases, IDSA recommends clinical testing for carbapenem-resistant Enterobacterales (CRE).6 Enzyme inhibitor enhancement is a simple and cost-effective method for detecting carbapenemase phenotypes, and has significant clinical applicability.22 In our study, this method could detect carbapenemase activity with high sensitivity (82.1%), particularly for isolates harboring a single carbapenemase-encoding gene (100.0%). Nevertheless, its detection capability was limited when dealing with isolates carrying two or more carbapenemase-encoding genes. Furthermore, 10 blaKPC-2 + blaNDM-1 isolates and one blaKPC-2 + blaNDM-5 isolate were positive for metalloenzymes in the carbapenemase phenotype test. It is worth noting that the R2 isolate co-harbored blaKPC-2 and blaIMP-26, but tested negative for all carbapenemases. Therefore, in clinical practice, it is not feasible to rely solely on enzyme types to determine CZA sensitivity, particularly when the test results are negative. A comprehensive assessment should incorporate drug sensitivity results along with carbapenemase and genotype information.
MBL production is the primary mechanism of CZA resistance. In recent years, new mechanisms of drug resistance have been discovered, such as the presence of new subtypes of KPCs, and the absence of outer membrane proteins.11,14,16,17 In this study, all CRKPCZA-R isolates carried MBL-encoding genes, most commonly blaNDM-1 (44.8%, 30/67), followed by blaIMP-4 (9.0%, 6/67), blaNDM-5 (7.5%, 5/67), and blaNDM-4 (1.5%, 1/67). This suggests that MBL production is the primary mechanism underlying CZA resistance in the CRKP isolates of our hospital. The new subtypes of KPC were not detected in our hospital, which may be attributed to the fact that CZA was not widely used in our hospital at that time. It was worth noting that 35.8% (24/67) of the CRKPCZA-R isolates simultaneously carried more than two carbapenemase-encoding genes, of which 21 isolates co-harbored blaNDM-1 and blaKPC-2, two isolates carried blaKPC-2 and blaIMP-4, one isolate (R2) carried blaKPC-2 and blaIMP-26, and one isolate (R57) carried blaKPC-2, blaNDM-1, and blaIMP-4. Recent reports indicate a rise in multi-carbapenemase–producing isolates, which would pose a serious threat to public health.25,26
At present, there is a lack of effective and reliable antibacterial drugs for nosocomial CRKPCZA-R infections. The CRKPCZA-R isolates from our hospital were highly sensitive to POL and TGC. Although studies indicate that this drug combination might be effective against CRKPCZA-R, POL is associated with heterogeneous resistance and nephrotoxicity, and TGC has poor efficacy in treating severe infections, thereby precluding their use for CRKPCZA-R infections.6,27 In addition, some reports suggest that the combination of FOS and other antibiotics can be used to treat urinary tract infections.28 Currently, metalloenzyme-producing CRKP is clinically managed using the combination of ATM (which has good stability against MBLs29) and CZA. Nevertheless, this therapeutic regimen is not effective against all MBL-producing isolates, which might be associated with mutations arising at specific sites within PBP3, the target site of ATM.30 Recently, AZA, a novel composite antibacterial agent, has gained approval for market release in Europe. The MIC of AZA in this study was exceptionally low, thus demonstrating its immense potential in the treatment of CRKPCZA-R infections.31 In addition, cefiderocol monotherapy has been recommended by IDSA for the treatment of metalloenzyme-producing CRKP.6 Novel antibiotics such as cefepime-taniborbactam may also be sensitive to metalloenzyme-producing CRKP.32 In this study, the detection rate of blaNDM-1 and blaKPC-2 was higher. For these strains, the combination of ATM with innovative β-lactam/β-lactamase inhibitors could prove to be an effective treatment option.33
ST11 was the predominant ST, and accounted for 35.8% (24/67) of the CRKPCZA-R isolates. Notably, 17 ST11 CRKPCZA-R isolates concurrently harbored both blaKPC-2 and blaNDM-1 along with ompk36 deletion, in addition to two or more hypervirulent marker genes. These findings are indicative of the clonal transmission of KL64 ST11 hypervirulent CRKPCZA-R isolates within our hospital. Furthermore, patients infected with these isolates were older than those infected with other drug-resistant isolates, and exhibited more serious illness. The ICU occupancy rate of these patients was also higher, as was the proportion of patients who were discharged without recovery. In addition, the MIC50 of FOS was significantly higher for the KL64 ST11 CRKP isolates co-harboring blaKPC-2 and blaNDM-1 than for other drug-resistant isolates, suggesting that FOS should be used cautiously in the treatment of urinary tract infections caused by these isolates. Remarkably, cefiderocol exhibited lower MIC50 and MIC90 for isolates co-harboring blaKPC-2 and blaNDM-1 compared to other isolates, and no intermediate or resistant isolates were identified, suggesting a potentially higher sensitivity to cefiderocol. Previous studies revealed that the prevalent serotype associated with ST11 CRKP has shifted from KL47 to primarily KL64, which might be related to factors such as the superior antioxidant capabilities of the KL64 isolate, or recombination.1,34–36 Furthermore, KL64-ST11 CRKP exhibits remarkable genetic plasticity. Although IncFII/IncR plasmids carrying the blaKPC-2 gene cannot be transferred independently, they can form hybrid plasmids with IncC plasmids harboring blaNDM, thereby facilitating transfer.26 In this study, the isolates harboring these genes also featured ompk36 deletion. It remains to be confirmed whether ompk36 deletion facilitates the acquisition of drug resistance genes through horizontal transfer. Altogether, our findings suggest that the clonal transmission of KL64-ST11 CRKP in hospitals should be monitored closely.
Conclusion
CRKPCZA-R isolates had a high detection rate in our hospital, and the main drug resistance mechanism was MBLs production. Nevertheless, these isolates exhibit very high sensitivity to POL, TGC, AZA, and cefdizime. The enzyme inhibitor enhancement method can detect carbapenemases with high sensitivity and specificity and provide an important reference for drug selection, but it might have limited detection ability for isolates that simultaneously produce more than two carbapenemases. Patients infected with CRKPCZA-R co-harboring both blaKPC-2 and blaNDM-1 exhibited more serious disease; these isolates exhibit lower sensitivity to FOS, high sensitivity to cefiderocol, and clonal transmission, all of which warrant concern.
Abbreviations
CZA, Ceftazidime-avibactam; CRKP, K. pneumoniae; CRKPCZA-R, CZA-resistant CRKP; WHO, World Health Organization; AVI, avibactam; KPC, K. pneumoniae carbapenemase; IDSA, Infectious Diseases Society of America; MBL, metallo-β-lactamase; ST, sequence type; MIC, minimum inhibitory concentration; CLSI, Clinical and Laboratory Standards Institute; MEM, meropenem; IPM, imipenem; ATM, aztreonam; POL, polymyxin B; TGC, tigecycline; FOS, fosfomycin; AZA, aztreonam-avibactam; APB, boric acid; EDTA, Ethylene Diamine Tetraacetic Acid; NGS, Next-generation sequencing; CRE, Carbapenem resistant Enterobacterales.
Data Sharing Statement
The datasets generated and analyzed during the current study are available in the NCBI database with ID PRJNA1236832. https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA1236832.
Ethics Approval and Informed Consent
The study was approved by the Clinical Research Medical Ethics Subcommittee of the Medical Ethics Committee of Xiangya Hospital, Central South University, with ID 202209541. All methods were performed in accordance with the relevant guidelines and regulations. Written informed consent was obtained from all participants.
Acknowledgments
We would like to express our gratitude to the staff of the Department of Clinical Laboratory at Central South University’s Xiangya Hospital for their assistance with collecting and identifying the CRKP isolates. We thank Medjaden Inc. for scientific editing of this manuscript. We also thank Xuan, Li (GMA MSD, China) for scientific support.
Funding
The Natural Science Foundation of Hunan Province (Nos. 2022JJ70084 and 2023JJ30900) and Merck Sharp & Dohme (100759) supported this study. The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Shi Q, Ruan Z, Zhang P, et al. Epidemiology of carbapenem-resistant Klebsiella pneumoniae in China and the evolving trends of predominant clone ST11: a multicentre, genome-based study. J Antimicrob Chemother. 2024;79:2292–11. doi:10.1093/jac/dkae227
2. Budia-Silva M, Kostyanev T, Ayala-Montaño S, et al. International and regional spread of carbapenem-resistant Klebsiella pneumoniae in Europe. Nat Commun. 2024;15:5092. doi:10.1038/s41467-024-49349-z
3. Lee J, Sunny S, Nazarian E, et al. Carbapenem-resistant Klebsiella pneumoniae in large public acute-care healthcare system, New York, New York, USA, 2016-2022. Emerg Infect Dis. 2023;29:1973–1978. doi:10.3201/eid2910.230153
4. Available from: https://www.who.int/news/item/17-05-2024-who-updates-list-of-drug-resistant-bacteria-most-threatening-to-human-health.
5. Diaz Caballero J, Wheatley RM, Kapel N, et al. Mixed strain pathogen populations accelerate the evolution of antibiotic resistance in patients. Nat Commun. 2023;14:4083. doi:10.1038/s41467-023-39416-2
6. Tamma PD, Heil EL, Justo JA, et al. Infectious diseases society of America 2024 guidance on the treatment of antimicrobial-resistant gram-negative infections. Clin Infect Dis. 2024;2024:ciae403. doi:10.1093/cid/ciae403
7. Díaz Santos E, Mora Jiménez C, Río-Carbajo LD et al. Treatment of severe multi-drug resistant Pseudomonas aeruginosa infections. Med Intensiva. 2022;46:508–520. doi:10.1016/j.medin.2022.03.015
8. Wilson WR, Kline EG, Jones CE, et al. Effects of KPC variant and porin genotype on the in vitro activity of meropenem-vaborbactam against carbapenem-resistant Enterobacteriaceae. Antimicrob Agents Chemother. 2019;63. 10.1128/AAC.02048-18
9. Sader HS, Mendes RE, Duncan L, et al. Ceftazidime-avibactam, meropenem-vaborbactam, and imipenem-relebactam activities against multidrug-resistant Enterobacterales from United States Medical Centers (2018-2022). Diagn Microbiol Infect Dis. 2023;106:115945. doi:10.1016/j.diagmicrobio.2023.115945
10. Lin LY, Debabov D, Chang W, et al. Antimicrobial activity of ceftazidime-avibactam and comparators against pathogens harboring OXA-48 and AmpC alone or in combination with other β-Lactamases collected from phase 3 clinical trials and an international surveillance program. Antimicrob Agents Chemother. 2022;66:e0198521. doi:10.1128/aac.01985-21
11. Hemarajata P, Humphries RM. Ceftazidime/avibactam resistance associated with L169P mutation in the omega loop of KPC-2. J Antimicrob Chemother. 2019;74:1241–1243. doi:10.1093/jac/dkz026
12. Gaibani P, Amadesi S, Lazzarotto T, et al. Genome characterization of a Klebsiella pneumoniae co-producing OXA-181 and KPC-121 resistant to ceftazidime/avibactam, meropenem/vaborbactam, imipenem/relebactam and cefiderocol isolated from a critically ill patient. J Glob Antimicrob Resist. 2022;30:262–264. doi:10.1016/j.jgar.2022.06.021
13. Liao Q, Deng J, Feng Y, et al. Emergence of ceftazidime-avibactam resistance due to a novel blaKPC-2 mutation during treatment of carbapenem-resistant Klebsiella pneumoniae infections. J Infect Public Health. 2022;15:545–549. doi:10.1016/j.jiph.2022.04.002
14. Ding L, Shen S, Chen J, et al. Klebsiella pneumoniae carbapenemase variants: the new threat to global public health. Clin Microbiol Rev. 2023;36:e0000823. doi:10.1128/cmr.00008-23
15. Hobson CA, Pierrat G, Tenaillon O, et al. Klebsiella pneumoniae Carbapenemase variants resistant to ceftazidime-avibactam: an evolutionary overview. Antimicrob Agents Chemother. 2022;66:e0044722. doi:10.1128/aac.00447-22
16. Shen Z, Ding B, Ye M, et al. High ceftazidime hydrolysis activity and porin OmpK35 deficiency contribute to the decreased susceptibility to ceftazidime/avibactam in KPC-producing Klebsiella pneumoniae. J Antimicrob Chemother. 2017;72:1930–1936. doi:10.1093/jac/dkx066
17. Guo Y, Liu N, Lin Z, et al. Mutations in porin LamB contribute to ceftazidime-avibactam resistance in KPC-producing Klebsiella pneumoniae. Emerg Microbes Infect. 2021;10:2042–2051. doi:10.1080/22221751.2021.1984182
18. Cui Q, Wang C, Wang Q, et al. Ceftazidime/Avibactam Resistance in Carbapenemase-Producing Klebsiella pneumoniae. Emerg Infect Dis. 2023;29:2398–2400.
19. Clinical and laboratory standards institute (CLSI). Performance standards for antimicrobial susceptibility testing. 34th. Clin Lab Stand Inst. 2024.
20. The US Food and Drug Administration (USFDA). Tigecycline–Injection products [EB/OL]. 2023. Available from: https://www.fda.gov/drugs/development-resources/tigec-ycline-injection-products.
21. The European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clinical breakpoints- breakpoints and guidance [EB/OL]. 2023. Available from: https://w-ww.eucast.org/clinical_breakpoints.
22. Tsakris A, Poulou A, Pournaras S, et al. A simple phenotypic method for the differentiation of metallo-beta-lactamases and class a KPC carbapenemases in enterobacteriaceae clinical isolates. The J Antimicrob Chemother. 2010;65:1664–1671. doi:10.1093/jac/dkq210
23. Han R, Yang X, Yang Y, et al. Assessment of Ceftazidime-Avibactam 30/20-μg disk, etest versus broth microdilution results when tested against enterobacterales clinical isolates. Microbiol Spectr. 2022;10:e0109221. doi:10.1128/spectrum.01092-21
24. Kikuchi Y, Yoshida M, Kuwae A, Asami Y, Inahashi Y, Abe A. Correlation between the spread of IMP-producing bacteria and the promoter strength of bla(IMP) genes. J Antibiot. 2024;77:315–323.
25. Guo H, Wu Y, Li L, Wang J, Xu J, He F. Global emergence of carbapenem-resistant Klebsiella pneumoniae co-carrying multiple carbapenemases. Comput Struct Biotechnol J. 2023;21:3557–3563. doi:10.1016/j.csbj.2023.07.013
26. Wang Q, Liu Y, Chen R, et al. Genomic insights into the evolution and mechanisms of carbapenem-resistant hypervirulent klebsiella pneumoniae co-harboring bla(KPC) and bla(NDM): implications for public health threat mitigation. Ann Clin Microbiol Antimicrob. 2024;23:27. doi:10.1186/s12941-024-00686-3
27. Paul M, Carrara E, Retamar P, et al. European Society of Clinical Microbiology and Infectious Diseases (ESCMID) guidelines for the treatment of infections caused by multidrug-resistant Gram-negative bacilli (endorsed by European society of intensive care medicine). Clin Microbiol Infect. 2022;28:521–547.
28. Kaye KS, Rice LB, Dane AL, et al. Fosfomycin for Injection (ZTI-01) Versus Piperacillin-tazobactam for the treatment of complicated urinary tract infection including acute pyelonephritis: ZEUS, A Phase 2/3 randomized trial. Clin Infect Dis. 2019;69:2045–2056. doi:10.1093/cid/ciz181
29. Sempere A, Viñado B, Los-Arcos I, et al. Ceftazidime-avibactam plus aztreonam for the treatment of infections by VIM-type-producing gram-negative bacteria. Antimicrob Agents Chemother. 2022;66:e0075122.
30. Mushtaq S, Vickers A, Woodford N, Livermore DM. Activity of aztreonam/avibactam and ceftazidime/avibactam against Enterobacterales with carbapenemase-independent carbapenem resistance. Int J Antimicrob Agents. 2024;63:107081. doi:10.1016/j.ijantimicag.2023.107081
31. Yu W, Luo Q, Shen P, et al. New options for bloodstream infections caused by colistin- or ceftazidime/avibactam-resistant Klebsiella pneumoniae. Int J Antimicrob Agents. 2021;58:106458. doi:10.1016/j.ijantimicag.2021.106458
32. Castanheira M, Kimbrough JH, Morgan GM, et al. Activity of aztreonam-avibactam, cefiderocol, and cefepime-taniborbactam against a global collection of genetically characterized metallo-β-lactamase-producing Enterobacterales. Antimicrob Agents Chemother. 2026;70:e0084225. doi:10.1128/aac.00842-25
33. Fu Y, Zhu Y, Zhao F, et al. In vitro synergistic and bactericidal effects of aztreonam in combination with ceftazidime/ avibactam, meropenem/vaborbactam and imipenem/relebactam against dual-carbapenemase-producing enterobacterales. Infect Drug Resist. 2024;17:3851–3861. doi:10.2147/IDR.S474150
34. Wang R, Zhang A, Sun S, et al. Increase in antioxidant capacity associated with the successful subclone of hypervirulent carbapenem-resistant Klebsiella pneumoniae ST11-KL64. Nat Commun. 2024;15:67.
35. Chen T, Wang Y, Zhou Y, et al. Recombination drives evolution of carbapenem-resistant Klebsiella pneumoniae sequence type 11 KL47 to KL64 in China. Microbiol Spectr. 2023;11:e0110722.
36. Wang J, Feng Y, Zong Z. The origins of ST11 KL64 Klebsiella pneumoniae: a genome-based study. Microbiol Spectr. 2023;11:e0416522. doi:10.1128/spectrum.04165-22
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.