Back to Journals » Infection and Drug Resistance » Volume 13
Prevalence and Molecular Typing of Colistin-Resistant Pseudomonas aeruginosa (CRPA) Among β-Lactamase-Producing Isolates: A Study Based on High-Resolution Melting Curve Analysis Method
Authors Tahmasebi H ![]() , Dehbashi S
, Dehbashi S ![]() , Arabestani MR
, Arabestani MR
Received 28 May 2020
Accepted for publication 7 August 2020
Published 21 August 2020 Volume 2020:13 Pages 2943—2955
DOI https://doi.org/10.2147/IDR.S264796
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Suresh Antony
Hamed Tahmasebi,1 Sanaz Dehbashi,2 Mohammad Reza Arabestani2,3
1Microbiology Department, Faculty of Medicine, Zahedan University of Medical Sciences, Zahedan, Iran; 2Microbiology Department, Faculty of Medicine, Hamadan University of Medical Sciences, Hamadan, Iran; 3Nutrition Health Research Center, Hamadan University of Medical Sciences, Hamadan, Iran
Correspondence: Mohammad Reza Arabestani
Microbiology Department, Faculty of Medicine, Hamadan University of Medical Sciences, Pajoohesh Junction, Hamadan, Iran
Tel +98-918-866-2009
Email [email protected]
Background: The frequency and production of β-lactamase enzymes may be different in colistin-resistant Pseudomonas aeruginosa (CRPA) strains compared to susceptible strains. The purpose of this study was to investigate the relationship between colistin resistance and β-lactamase enzymes in different Sequence Types (ST) of P. aeruginosa.
Methods: A total of 101 P. aeruginosa isolates were collected from different samples. The antimicrobial susceptibilities of the bacterial isolates were examined by disk diffusion and MIC E-test methods. Also, real-time PCR and high-resolution melting curve analysis (HRMA) assay were performed to detect the resistance genes.
Results: Out of the 101 P. aeruginosa isolates, four isolates (3.96%) were resistant to colistin. Also, 39 isolates (38.61%) were considered as MDR, and eight isolates (7.92%) were considered as XDR. Further, 25 (24.75%) and 26 isolates (25.74%) were produced ESBL and carbapenemase enzymes, respectively. According to HRMA results, four isolates (3.96%) were positive for pmrA, three isolates (2.97%) were positive for mcr-1, 25 isolates (24.75%) were positive for blaTEM, 24 isolates (23.76%) were positive for blaSHV, 26 isolates (25.75%) were positive for blaKPC, and 23 isolates (22.77%) were positive for blaIMP genes. Furthermore, ST108 and ST250 showed the highest distribution in P. aeruginosa isolates. Also, ST217, ST1078, and ST3340 were reported as novel types in CRPA strains.
Conclusion: Concerns about the prevalence of CRPA strains should be taken seriously. Also, our results showed that the mcr-1 gene plays a vital role in the distribution of ESBL and KPC-producing P. aeruginosa strains.
Keywords: Pseudomonas aeruginosa, high-resolution melting curve analysis assay, HRMA assay, β-lactamases, colistin resistance, sequence types
Introduction
Pseudomonas aeruginosa has been shown to produce the following β-lactamases: Class A extended-spectrum β-lactamases (ESBL) and Class B Metallo-β-lactamases that degrade all β-lactams, including the carbapenems.1,2 Carbapenemases, mostly classified as Ambler class B Metallo-β-lactamases (MBLs), are zinc-dependent and have an extensive substrate profile, such as carbapenems and expanded-spectrum cephalosporins.3,4 However, resistance to colistin in β-lactamase producing strains is higher than strains lacking this enzyme.5,6
The emergence of multi-drug resistance (MDR) strain of P. aeruginosa has resulted in the use of older drugs such as colistin.7 Colistin is bactericidal and displays excellent activity against pathogens such as P. aeruginosa, which is highly resistant to currently available antibiotics such as carbapenems.8 These modifications occur as a result of the activation of the PmrA-PmrB system—this system regulated by the PhoP-PhoQ system.9 In addition to the known chromosomal-mediated resistance mechanisms, a plasmid-mediated colistin resistance gene named mcr-1 was discovered in China and subsequently elsewhere around the world.10 Carbapenemase producers, mainly nosocomial P. aeruginosa isolates, are usually MDR, and therapeutic options for treating infections with carbapenemase producers remain limited.11
However, the sensitivity and specificity of phenotypic methods are less than molecular methods.12 The speed and accuracy of these methods are also low compared to new molecular assay.13 HRMA is the characteristic of a targeted PCR product melting profile resulting from gradually increasing melting temperature (Tm) by tiny increments in the presence of high-saturating DNA binding dye.14 Moreover, analysis based on HRMA has been used in different clinical laboratory applications, such as the identification of many different bacterial isolates.15 Understandably, HRMA assay has endless applications and just as many advantages. The technology that runs and analyzes samples undergoing HRMA is a cost-effective one. HRMA also is a straightforward technology that can perform both the PCR and HRM Analysis in one instrument.12
CRPA, have all been reported in clinical isolates; therefore, rapid and accurate identification of these strains become particularly important.9 Hence, we used the HRMA technique to identify the different isolates of P. aeruginosa in the clinical specimens. Hence, the present study involving the epidemiological surveillance of colistin-resistant P. aeruginosa from Hamadan, Iran. We further describe the molecular epidemiology of ESBL and KPC-producing strains leading to increase colistin resistance in P. aeruginosa.
Materials and Methods
Study Design and P. Aeruginosa Isolates
Out of the 402 isolates collected from different clinical specimens which were part of the routine hospital laboratory procedure, a total of 101 P. aeruginosa isolates were collected from different samples from November 2017 to December 2018, from different wards of the Hamedan Hospitals and University Centers, Iran. Biochemical tests were used to differentiate between closely related bacterial species or genera. These tests were conducted according to the standard specifications described and applied in the Microbiology Laboratory at Hamedan University of medical sciences. Colonies morphologically resembled P. aeruginosa and had a positive oxidase test selected for further identification. Then colonies were examined by various biochemical tests such as oxidase, catalase, urease, arginine dehydrogenase, and oxidation-fermentation. Bacterial strains were stored at −20°C in Brain-Heart Infusion (BHI) (Merck, Germany) broth supplemented with 25% v/v glycerol. This study was approved by the ethics committee of Hamadan University of Medical Sciences (No: IR.UMSHA.REC. 1396.637).
Determination of Antibiotic Profiles
The disk diffusion (Kirby-Bauer) method was used for the detection of antimicrobial resistance patterns. Nine separate antimicrobial susceptibility discs, including imipenem, doripenem, amikacin, ceftazidime, aztreonam, gentamicin, ciprofloxacin, piperacillin, and cefepime (MAST, UK) were used. Plates were incubated for 18–24 hours in ambient air at 35ºC±2 and the diameters of zones of inhibition around the antimicrobial discs recorded to the nearest whole millimeter using a graduated ruler. Results were reported as “sensitive,” “intermediate,” or “resistant” based on the cut-off values recommended by CLSI 2018.16
MIC test was performed for all the P. aeruginosa isolates based on the standard CLSI guidelines 2018 using E.test (Liofilchem, Italy) and broth microdilution method for the following antimicrobials: amikacin, ciprofloxacin, ceftazidime, aztreonam, imipenem, meropenem, cefoxitin, and colistin. For all phenotypic tests, P. aeruginosa ATCC 15,442 and P. aeruginosa ATCC 27,853 were used as a positive control, and P. aeruginosa PAO-1 was used as a negative control.
Phenotypic Detection of ESBL and KPC-Producing
Combined disc method (CDT) (Mast, UK) and modified Hodge test (MHT) was used for the detection of ESBL and KPC-producing strains, respectively. Discs containing cefotaxime and ceftazidime with and without clavulanic acid were used. An ESBL phenotype was defined as an increase of ≥ 5 mm in the zone around the disc containing clavulanic acid compared to the zones of corresponding discs without clavulanic acid.17 In each batch of the experiment, K. pneumoniae ATCC BAA-1705 was included as the positive control, and a carbapenemase-negative K. pneumoniae strain ATCC BAA-1706 was included as a negative control.
Extraction of DNA
Genomic DNA was isolated using the Qiagen miniprep kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. Also, a plasmid was isolated from 5mL of overnight culture using Qiagen miniprep kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DNA and plasmid concentrations were determined using a spectrophotometer Nanodrop-200 (Hangzhou Allsheng Instruments Co., Ltd, China), and the samples were stored at −20ºC for further DNA analysis.
Evaluation of Sensitivity and Specificity and HRMA Assay
Real-time PCR was carried out in 20µL reactions on the ABI step one plus (USA) instrument (Table 1). For each reaction, 4µL of the reaction mix (HOT FIREPol EvaGreen HRMA Mix), 1µL of each primer (5pmol).9,18-21 Furthermore, PCR grade water to a final volume of 12µL was added to 2µL of the target DNA. PCR amplification was carried out according to the following parameters; a denaturation cycle at 95°C for 10min following by 40 amplification cycles which include, denaturation at 95°C for 10 sec, annealing at 58°C for 30 sec with single fluorescence measurement and extension at 72°C for 20 sec. This was followed by a melting curve (Tm) analysis program for one cycle (95 °C for 1 sec, 70°C continuous using a temperature transition rate of 0.3ºC/sec and a cooling period of 40°C for 30 sec. Then, the detection of pathogens was performed by melting curve analysis. P. aeruginosa ATCC 15,442 and P. aeruginosa ATCC 27,853 were used as a positive control, and P. aeruginosa PAO-1 was used as a negative control.
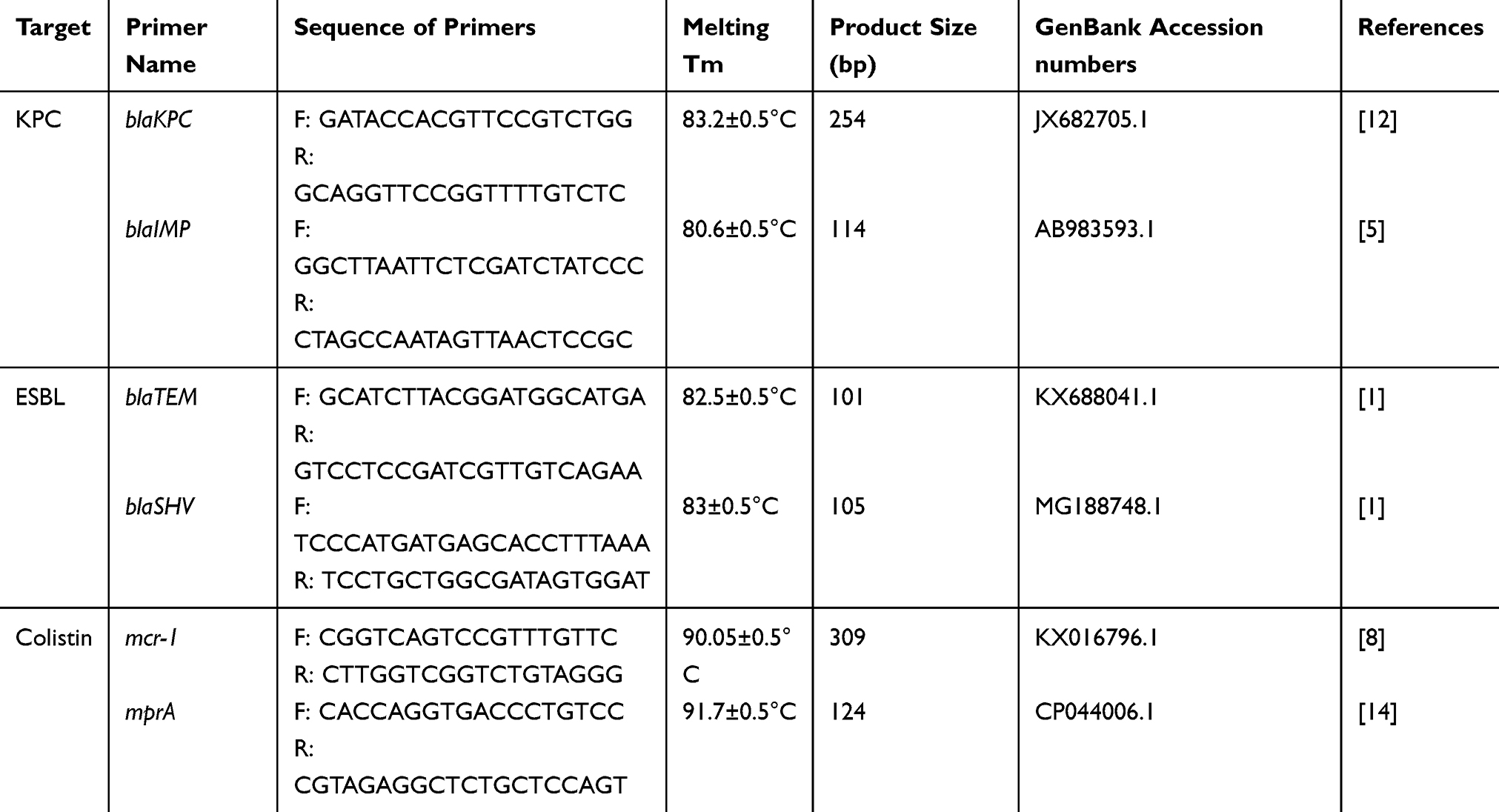 |
Table 1 Oligonucleotide Sequences Used in This Study |
MLST Sequences Typing
All strains of colistin and β-lactamase producers were selected for sequencing. The seven housekeeping genes: acsA, aroE, guaA, mutL, nuoD, ppsA, and trpE were selected for amplification and sequencing, according to the MLST scheme of P. aeruginosa website (http://pubmlst.org/paeruginosa/). PCR amplification reactions (volumes, nucleotides, and primer concentrations) were performed in an Eppendorf thermocycler as previously described. The amplified products were purified by Pishgam biotech co (Tehran, Iran).
Data Analysis
The WHO.net 2018 software was used for entry and preliminary analysis of microbiology data. SPSS software version 16 (SPSS, Chicago, IL) and chi-square and t-test were used to examine the relationship between different variables. The instrument measured the fluorescence, and ABI Thermo Fisher software plotted the fluorescence versus temperature in a graph. Reagent blanks were used to ensure there was no contamination in the reagents.
Results
Out of the 402 isolates collected from different clinical specimens, 101 P. aeruginosa isolates were identified as follows: 22 isolates (21.78%) from blood, 30 isolates (29.70%) from wounds, 18 isolates (17.82%) from the burn, 14 isolates (13.86%) from urine, seven isolates (6.93%) from the catheter, and ten isolates (9.9%) from discharge. Eighty-eight isolates (87.12%) were isolated from female patients, and 13 isolates were collected (12.87%) from male patients.
Prevalence of Antibiotic Resistance Profile
The resistance profile of P. aeruginosa isolates was detected as follows: 28 strains (27.72%) resistant to imipenem, 19 strains (18.81%) resistant to doripenem, 13 strains (12.87%) resistant to amikacin, 19 strains (18.81%) resistant to ceftazidime, 11 strains (10.89%) resistant to aztreonam, 96 strains (95.04%) resistant to gentamicin, 97 strains (96.03%) resistant to ciprofloxacin, 23 strains (22.77%) resistant to piperacillin, and 29 strains (28.71%) resistant to cefepime (Figure 1A). Besides, 39 isolates (38.61%) were resistant to six antibiotics and were considered as MDR strains, and eight isolates (7.92%) were resistant to all antibiotics and were considered as XDR strains (Figure 1).
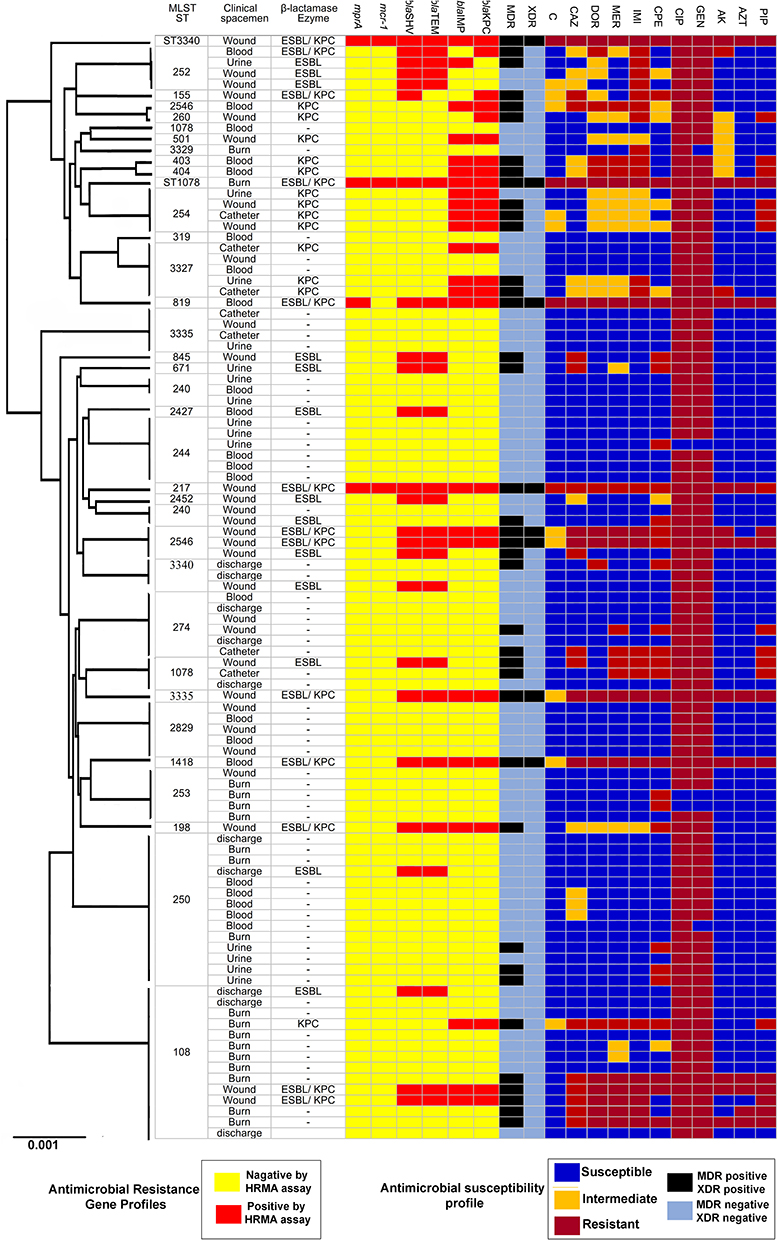 |
Figure 1 Analysis of 101 sequenced P. aeruginosa genomes. A maximum-likelihood tree and MLST analysis, presence of beta-lactamases, and enzymes. |
Out of the 101 isolates, 28 isolates (27.72%) showed MIC >8 µg/mL for imipenem, 20 isolates (19.18%) demonstrated MIC >32 µg/mL for ceftazidime, 13 isolates (12.87%) were detected with MIC >4 µg/mL for amikacin, 10 isolates (9.9%) indicated MIC>4 µg/mL for aztreonam, 98 isolates (96.03%) showed MIC >4 µg/mL for ciprofloxacin, and 4 isolates (3.96%) were observed with MIC >4 µg/mL for colistin (Figure 1).
Prevalence of ESBL and KPC-Producing Strains
Out of the 101 isolates of P. aeruginosa, 25 isolates (24.75%) were considered as ESBL producing, and 26 isolates (25.74%) were considered as KPC-producing (carbapenemase-producing), and 13 isolates were considered as ESBLs/carbapenemase-producing.
Analytical Sensitivity and Specificity of Primers
The melting profile was characterized by a peak of 83.2±0.5°C for blaKPC gene, 80.6±0.5°C for blaIMP gene, 83±0.5°C for blaSHV gene, 82.5±0.5°C for blaTEM gene, 91.7±0.5°C for pmrA gene, and 90.0± 0.5°C for mcr-1 gene. The results of the standard curves for the various dilutions of DNA are shown in Figures 2 and 3. Moreover, the primers used were not able to detect Staphylococcus aureus at specific Tm (Figure 4).
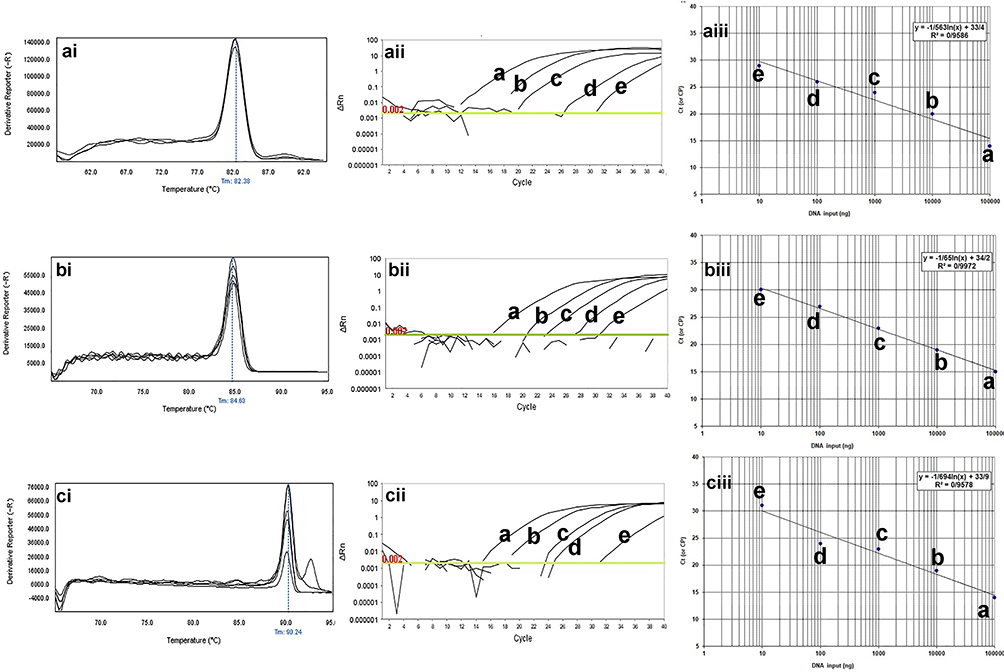 |
Figure 2 The qPCR dilution series for analytical sensitivity of real-time PCR for mcr-1, blaTEM, and blaSHV genes in clinical isolates of P. aeruginosa. The melting profiles of mcr-1, blaTEM, and blaSHV genes showed (i) Figures. The Ct value of different dilutions was shown in (ii) Figures. The primer efficiency slope and standard curves of mcr-1, blaTEM, and blaSHV genes are shown in (iii) Figures. The melting temperature profile was as follows: 83±0.5°C for blaSHV gene, 82.5±0.5°C for blaTEM gene, and 90.0± 0.5°C for mcr-1 gene. Also, the efficiency of the primer was 98.19% for the blaSHV gene, 99.97% for the blaTEM gene, and 97.01% for the mcr-1 gene. For evaluation of the sensitivity of the real-time PCR assays, a series of five-fold diluted DNA of the positive controls were initially tested on the ABI Step-One Plus system. The mean of a: 100,000 ng; b: 10,000 ng; c: 1000 ng; d: 100 ng; e: 10 ng of DNA dilutions. Horizontal lines represent the cycle threshold of real-time PCR. The melting and slope temperature profile were as follows: 83.2±0.5°C for blaKPC gene, 80.6±0.5°C for blaIMP gene, 83±0.5°C for blaSHV gene, 82.5±0.5°C for blaTEM gene, 91.7±0.5°C for pmrA gene, and 90.0± 0.5°C for mcr-1 gene. |
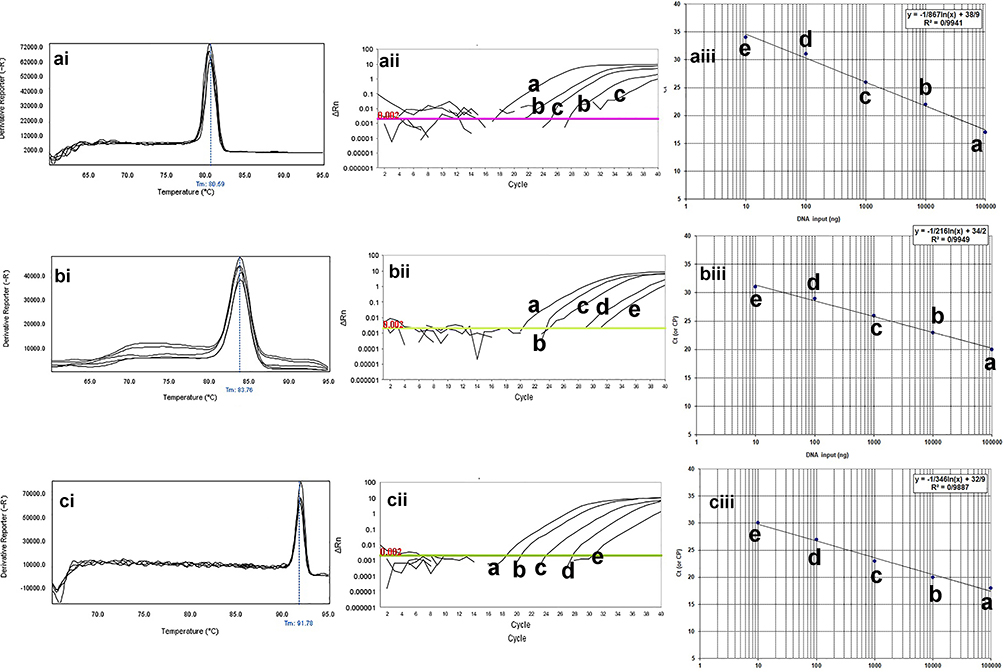 |
Figure 3 The qPCR dilution series for analytical sensitivity of real-time PCR for blaIMP, blaKPC, and mprA genes in clinical isolates of P. aeruginosa. The melting profiles of blaIMP, blaKPC, and mprA genes showed (i) Figures. The Ct value of different dilutions was shown in (ii) Figures. The primer efficiency slope and standard curves of blaIMP, blaKPC, and mprA genes are shown in (iii) Figures. The melting temperature profile was as follows: 83.2±0.5°C for blaKPC gene, 80.6±0.5°C for blaIMP gene, and 91.7±0.5°C for pmrA gene. Also, the efficiency of the primer was 99.31% for the blaKPC gene, 97.99% for the blaIMP gene, and 98.12% for the mprA gene. For evaluation of the sensitivity of the real-time PCR assays, a series of five-fold diluted DNA of the positive controls were initially tested on the ABI Step-One Plus system. The mean of a: 100,000 ng; b: 10,000 ng; c: 1000 ng; d: 100 ng; e: 10 ng of DNA dilutions. Horizontal lines represent the cycle threshold of real-time PCR. |
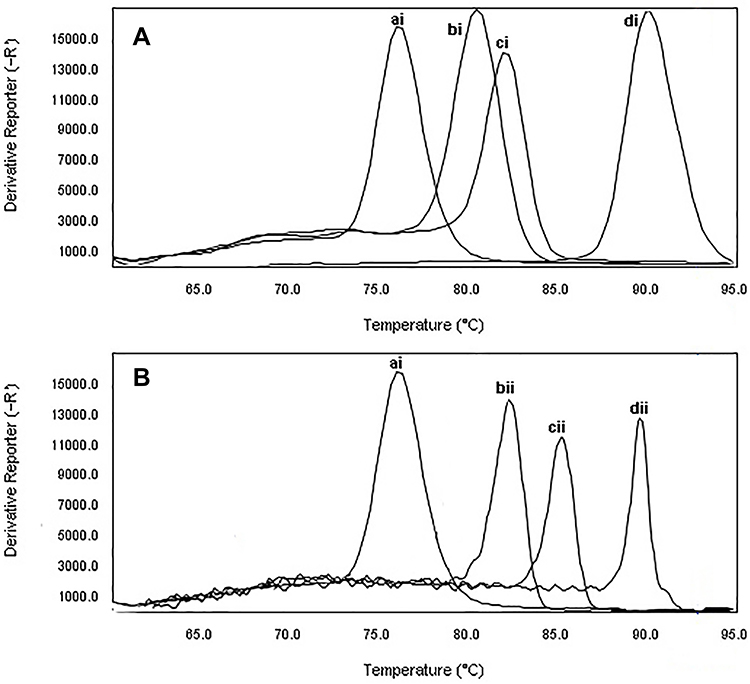 |
Figure 4 The specificity of real-time PCR for primers used to detect of CRPA and β-lactamase producing P. aeruginosa. Melting curve analysis showing the melting temperature peaks (Tm) of blaIMP (bi), blaKPC(ci), mprA(di), blaTEM(bii), blaSHV(cvi), and mcr-1(dii). ai: Staphylococcus aureus ATCC25923. |
Detection ESBL and KPC Producer, and Colistin-Resistant Strains by HRMA Assay
The frequency of beta-lactamase and colistin genes by the HRMA method was as follows: 25 isolates (24.75%) carrying blaTEM gene, 26 isolates (25.74%) carrying blaKPC gene, 24 isolates (23.76%) carrying blaSHV gene, 23 isolates (22.77%) carrying blaIMP gene, four isolates (3.96%) carrying pmrA gene, and three isolates (2.97%) carrying mcr-1 gene (Figures 4 and 5).
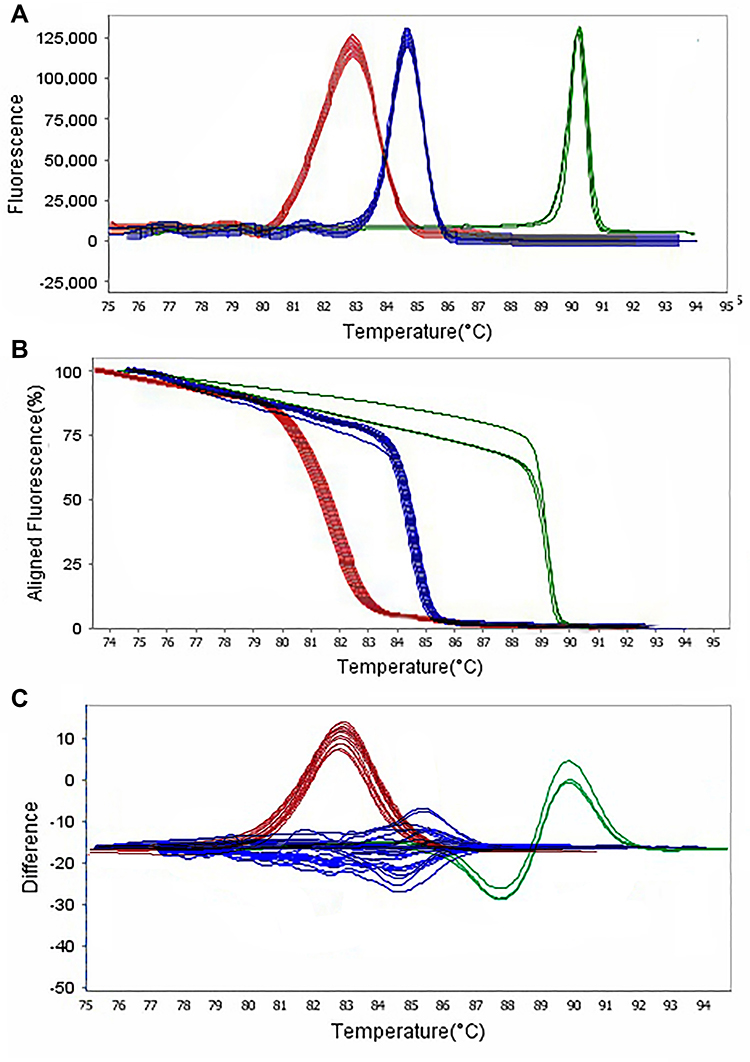 |
Figure 5 Representative high resolution melting graphs corresponding to one high resolution melting analysis of MDR and XDR strains of P. aeruginosa (n = 39). Curves of tested samples identified as blaIMP carrier are shown in red, blaKPC carrier are shown in blue, and mprA carrier are shown in green. β-lactamases genes were amplified successfully using the EvaGreen Dye in the ABI Step-OnePlus machine by one-sept protocol. (A) Melting curves; (B) Normalized plot; and (C) Difference plot. Positive control: P. aeruginosa NCTC 13,359. |
Phylogenetic Analysis of the MLST Sequences
Among the 101 isolates that were genotyped, 35 different STs were identified. ST108 and ST250 were found in many strains. Therefore, a lower level of genetic diversity was observed among the isolates were obtained from male patients comparing to the isolates were collected from female patients. In total, 40% of all colonized female patients harbored P. aeruginosa STs that were also found in other male patients, and 30% were colonized with isolates with unique STs. Moreover, ST217, ST1078, and ST3340 were characterized as novel types among the CRPA, ESBL, and carbapenemase-producing P. aeruginosa. Typing by MLST further illustrated the enormous diversity within the strains, as isolates with similar β-lactamase genes had different STs (Figure 1).
Allelic profiles were investigated to find a representative isolate circulating among the different samples, including wound, blood, and urine culture. The MLST revealed a new ST, namely ST3340, belonging to a clonal complex previously has not been described. Further, 20 ST types were identified, of which two were shared between multiple clinical samples. ST108 and ST250 were common among wound, urine, burn, and discharge specimens, and ST254 and ST274 were shared by the wound, catheter, burn, and discharge specimens (Figure 1).
Analytical Analysis
The most prevalent STs, ST1078 and ST3340, had a higher prevalence in the blood and wound infections. The type of the samples and antibiotics residence were correlated with the carriage of STs (P =0.001). In general, ST3340 strains were more sequence type in MDR and XDR strains. Further, Both ST1078 and ST3340 strains were more often resistant to colistin (P = 0.001) (Table 2).
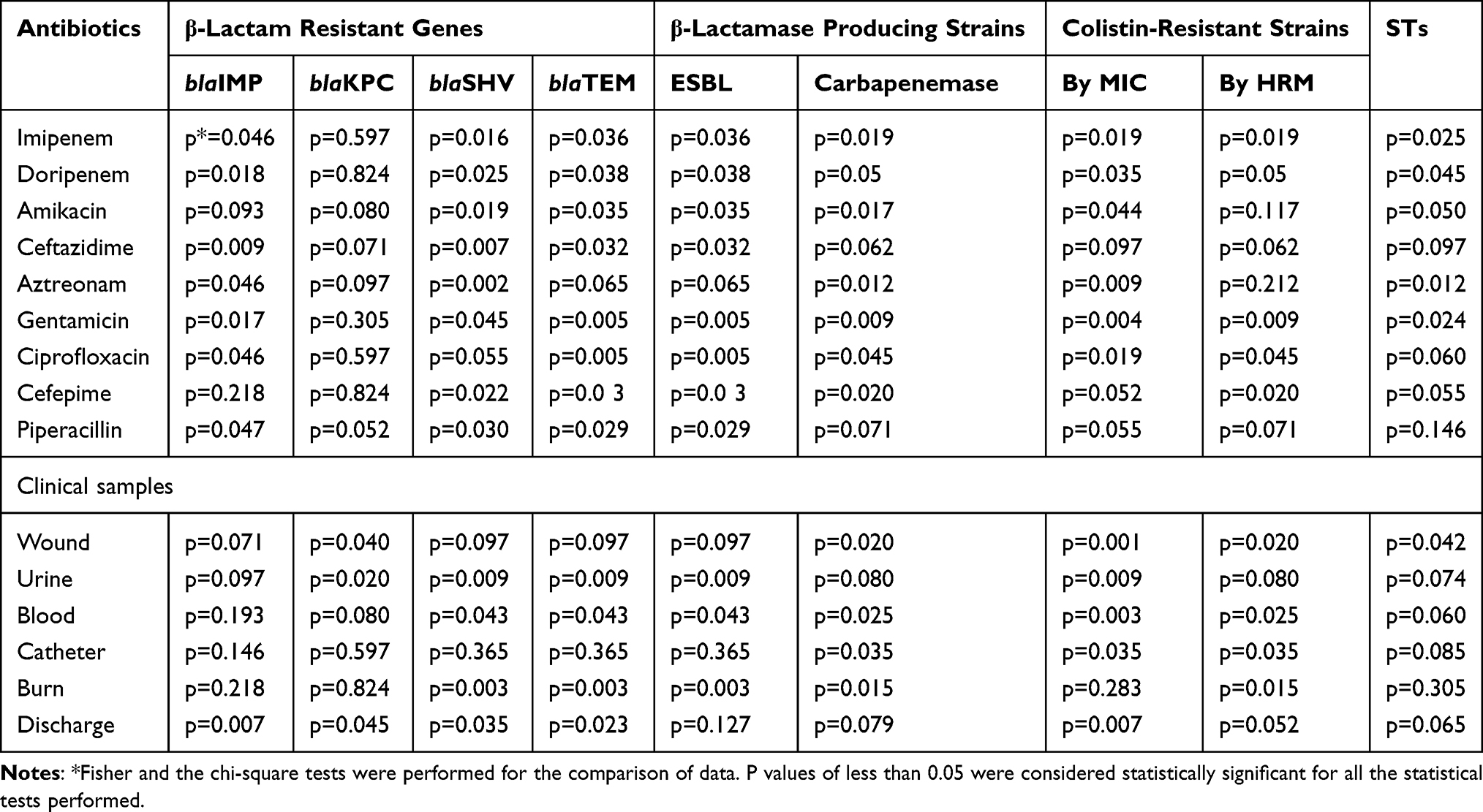 |
Table 2 Analysis of Variables Significantly Associated STs, Antimicrobial Resistance and Clinical Samples in Clinical Isolates of P. aeruginosa |
Discussion
P. aeruginosa is an important pathogen associated with severe infections in healthcare settings and a significant cause of morbidity and mortality. However, colistin is rapidly emerging as the last resort antibiotic for the treatment of MDR/XDR P. aeruginosa.15
In this study, according to phenotypic methods, gentamicin and ciprofloxacin-resistant strains had the highest frequency with 95.04% and 96.03%, respectively, and amikacin (6.66%) and colistin (3.96%) showed the lowest abundance. Moreover, MDR and XDR strains were counted as 38.61% and 7.92% of isolates, respectively. These results are in agreement with different studies of Venezuela,22 China,23 and Germany,24 that demonstrated high rates of antibacterial resistance of P. aeruginosa, where they showed similar resistance to the antibiotics examined in this study. In contrast to the current study, some reports showed a low rate of resistance in the last three years, except for colistin, in which the same findings were described.25,26 Also, the prevalence of KPC, ESBL-producing, and CRPA strains (as illustrated in Figure 1) was 25.74%, 24.75%, and 3.96%, respectively, which was in line with other studies.27,28 However, other studies described the different abundance of P. aeruginosa isolates of KPC, ESBL-producer, and resistant to colistin.1,2
In the current study, the distribution of ESBL and KPC-producing strains were reported 58.92%, 48.71% from MDR strains, and 100% from XDR strains of P. aeruginosa, respectively. Moreover, in MDR strains, 33.33% of isolates were KPC-ESBL producing strain, and in XDR, 100% of P. aeruginosa isolates were KPC-ESBL producers. Those findings correlate with other reports indicated that the prevalence of MDR and XDR strains in KPC-ESBL producing strains of P. aeruginosa higher than that of non- KPC-ESBL producing strains.1,29 Therefore, based on Table 2, there was a significant relationship between the prevalence of MDR-XDR strains and the production of ESBL and carbapenemases producing P. aeruginosa (p=0.001). Some researchers reported that the same result and stated that the frequency of β-lactamases in XDR strains is higher than MDR ones.1,3,30,31 However, to increase the predictive value of this association, precise and sensitive molecular methods should be used in epidemiological studies.32
In this study, we developed an HRMA assay for the detection of ESBL producing, KPC-producing, and CRPA strains by melting curve analysis technology. Based on Melting temperatures, as showed in Figures 2 and 3, melt curves of gene amplification were equal to 91.7±0.5°C for pmrA gene and 90.0± 0.5°C for mcr-1 gene. Besides, DNA melting temperatures for the blaTEM, blaSHV, blaKPC, and blaIMP genes were 82.5±0.5°C, 83±0.5°C, 83.2±0.5°C, and 80.6±0.5°C, respectively. According to our study in Figure 5, CRPA strains had a prevalence of 2.44% and detected by the HRMA method in agreement with a similar study,11,12 as well as Sekyere in a study showed that different methods for identifying colistin resistance strains are not of the same sensitivity and specificity.18 Also, the mechanism of colistin resistance in P. aeruginosa has not been fully understood, and some functions may be performed by genes not cover in this study.
According to Figures 5 and 6, HRMA results revealed that 88.23%, 82.35%, and 94.11% of P. aeruginosa were positive for blaTEM and blaSHV genes. Likewise, the frequency of MDR-ESBL producing, MDR-KPC producing, XDR-KPC producing, and XDR-KPC producing strains were reported in 83.52%, 76.47%, 87.5%, and 37.5% of P. aeruginosa, respectively. A strong and significant relationship was observed between the MDR-XDR strains and the production KPC and ESBL producer in P. aeruginosa strains, whereas the frequency of KPC-ESBL producing strains in MDR and XDR strains were 83.52% and 62.5%, respectively. Besides, our phenotypic and HRMA findings confirmed a significant association between colistin resistance and the prevalence of beta-lactamases. Also, the high prevalence of CRPA was reported in MDR and XDR strains. Thus, carbapenem resistance has led to the emergence of the colistin. On the other hand, the emergence of colistin resistance in XDR-KPC producing strains is higher than in MDR-KPC producing strains and is mediated by the mcr-1 gene. The mcr-1 gene can be transmitted through the plasmid to colistin-sensitive strains.33–36 Another concern is the emergence of colistin resistance during therapy among KPC-producing strains of P. aeruginosa in Switzerland,37 USA,38 and Australia.8
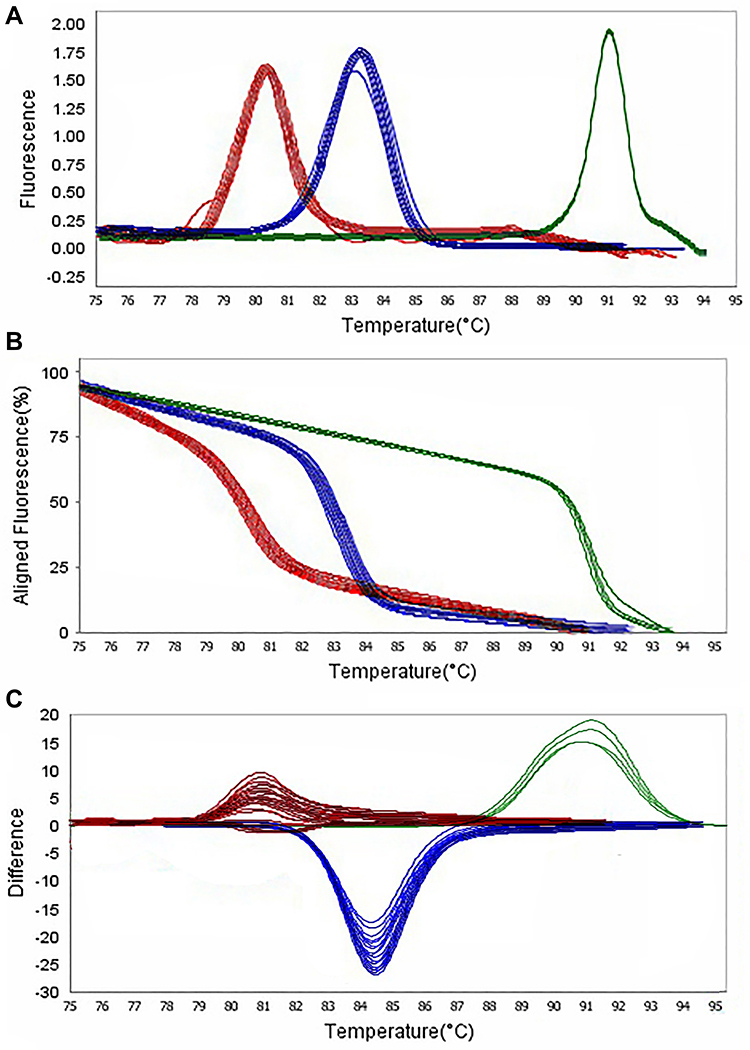 |
Figure 6 Representative high resolution melting graphs corresponding to one high resolution melting analysis of MDR and XDR strains of P. aeruginosa (n = 39). Curves of tested samples identified as blaTEM carrier are shown in red, blaSHV carrier are shown in blue, and mcr-1 carrier are shown in green. β-lactamases genes were amplified successfully using the EvaGreen Dye in the ABI Step-OnePlus machine by one-sept protocol. (A) Melting curves; (B) Normalized plot; and (C) Difference plot. Positive control: P. aeruginosa NCTC 13,359. |
As the main benefit of this study was to develop an HRMA assay to detect low levels KPC-producing, ESBL producing simultaneously, and CRPA strains, we next selected the KPC/IMP/mprA primer set1 and TEM/SHV/mcr-1 primers set2 combination for a multiplex assay assessment. Although the results of the HRMA assay and phenotypic methods for identifying β-lactamase producing strains in P. aeruginosa showed close results; however, the HRMA assay has several advantages. The significant advantage of the HRMA assay in this study was the identification of co-existence of CRPA and β-lactamase producing strains in a short time. Bentaleb et al39 and Otaguiri et al40 demonstrated that HRMA based methods have an improved level of sensitivity when compared to culture and, thus, can detect lower levels of bacteria.
In Korea, it has previously been argued that some STs are highly abundant in CRPA strains.6 In the present study, Figure 1 reported that the most prevalent STs, ST108, and ST250, had a higher prevalence in the blood and burn-wound infections. The gender of the patients was correlated with the carriage of STs (P = 0.001). Compared to strains of all other STs, strains of ST1078 and ST3340 could be characterized by a specific antibiotic resistance profile. In general, ST3340 strains were more resistant to other antibiotics and susceptible to amikacin. Both ST1078 and ST3340 strains were more often resistant to colistin. In contrast to this study, some researchers reported ST235 and ST108 as the essential MDR strains in colistin-sensitive P. aeruginosa.5,6 This difference may be due to resistance to colistin in P. aeruginosa.
In this study, we would like to point out these limitations; the role of colistin resistance in the prevalence of P. aeruginosa β-lactamase producing strains needs further studies. Besides, the HRMA method is limited due to the use of dedicated primers and sensitive fluorescent dyes. It should be noted that software analysis of the melting curve of HRMA has a significant impact on the quality of the results.
Conclusions
Colistin resistance in clinical isolates of Pseudomonas aeruginosa is one of the most critical recent threats. The results of the present study showed that colistin resistance is directly related to ESBL and KPC enzymes. Analysis of new sequence types (ST3340 and ST1078) from P. aeruginosa isolates revealed that both STs were resistant to colistin and producer of ESBL and KPC enzymes. Those observations suggest that colistin resistance may provide resistance to beta-lactam and pneumonia antibiotics. Moreover, colistin resistance was observed only in MDR-ESBL producing, MDR-KPC producing, XDR-KPC producing, XDR-KPC producing. These indicate that the optimization of an accurate and sensitive method for the identification of CRPA strains is essential. Compared with phenotypic methods, HRMA assay can identify colistin and β-lactam resistant strains simultaneously with high speed and accuracy. Moreover, we reported that the acquisition of a novel ST of colistin-resistant from Iran, further indicating that the emergence of CRPA strains among important human pathogens is becoming broadened.
Abbreviations
MDR, multiple drug resistance; XDR, extensively drug-resistant; KPC, Klebsiella pneumoniae carbapenemase; ESBL, extended-spectrum β-lactamases; SHV, sulfhydryl variable; TEM, temoneira; mcr-1, mobilized colistin resistance-1; MIC, minimum inhibitory concentration; HRMA, high-resolution melting analysis; STs, sequence type; MLST, multilocus sequence typing; CRPA, colistin-resistant P. aeruginosa; CDT, combined disk method; TSB, tryptic soy broth; WHO, World Health Organization.
Ethics Approval and Consent to Participate
This study was approved by the ethics committee of Hamadan University of Medical Sciences (No: IR.UMSHA.REC. 1396.637). Written informed consent was taken from all participants.
Acknowledgment
The authors would like to acknowledge the Vice-Chancellor of Hamadan University of Medical Sciences for the funding and support of the study.
Author Contributions
All authors made substantial contributions to conception and design, acquisition of data, or analysis and interpretation of data; took part in drafting the article or revising it critically for important intellectual content; agreed on the journal to which the article will be submitted; gave final approval of the version to be published; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest for this work.
References
1. Peymani A, Naserpour-Farivar T, Zare E, Azarhoosh KH. Distribution of bla(TEM), bla(SHV), and bla(CTX-M) genes among ESBL-producing P. aeruginosa isolated from Qazvin and Tehran hospitals, Iran. J Prev Med Hyg. 2017;58(2):E155–E160.
2. Kaur A, Singh S. Prevalence of Extended Spectrum Betalactamase (ESBL) and Metallobetalactamase (MBL) producing pseudomonas aeruginosa and acinetobacter baumannii isolated from various clinical samples. J Pathog. 2018;2018:7. doi:10.1155/2018/6845985
3. Pattnaik D, Panda SS, Singh N, Sahoo S, Mohapatra I, Jena J. Multidrug resistant, extensively drug resistant and pan drug resistant gram negative bacteria at a tertiary care centre in Bhubaneswar. ijcmph. 2019;6(2):6. doi:10.18203/2394-6040.ijcmph20190170
4. Kosykowska E, Dzieciątkowski T, Mlynarczyk G. Rapid detection of NDM, VIM, KPC and IMP carbapenemases by real-time PCR. J Bacteriol Parasitol. 2016;7(06):6. doi:10.4172/2155-9597.1000299
5. Guzvinec M, Izdebski R, Butic I, et al. Sequence types 235, 111, and 132 predominate among multidrug-resistant pseudomonas aeruginosa clinical isolates in Croatia. Antimicrob Agents Chemother. 2014;58(10):6277–6283. doi:10.1128/AAC.03116-14
6. Wi YM, Choi J-Y, Lee J-Y, et al. Emergence of colistin resistance in Pseudomonas aeruginosa ST235 clone in South Korea. Int J Antimicrob Agents. 2017;49(6):767–769. doi:10.1016/j.ijantimicag.2017.01.023
7. Roschanski N, Fischer J, Guerra B, Roesler U. Development of a multiplex real-time PCR for the rapid detection of the predominant beta-lactamase genes CTX-M, SHV, TEM and CIT-type AmpCs in Enterobacteriaceae. PLoS One. 2014;9(7):e100956. doi:10.1371/journal.pone.0100956
8. Nang SC, Li J, Velkov T. The rise and spread of mcr plasmid-mediated polymyxin resistance. Crit Rev Microbiol. 2019;45(2):131–161. doi:10.1080/1040841x.2018.1492902
9. Lee J-Y, Ko KS. Mutations and expression of PmrAB and PhoPQ related with colistin resistance in Pseudomonas aeruginosa clinical isolates. Diagn Microbiol Infect Dis. 2014;78(3):271–276. doi:10.1016/j.diagmicrobio.2013.11.027
10. Hameed F, Khan MA, Muhammad H, Sarwar T, Bilal H, Rehman TU. Plasmid-mediated mcr-1 gene in Acinetobacter baumannii and Pseudomonas aeruginosa: first report from Pakistan. Rev Soc Bras Med Trop. 2019;52. doi:10.1590/0037-8682-0237-2019
11. Chabou S, Leangapichart T, Okdah L, Le Page S, Hadjadj L, Rolain JM. Real-time quantitative PCR assay with Taqman((R)) probe for rapid detection of MCR-1 plasmid-mediated colistin resistance. New Microbes New Infect. 2016;13:71–74. doi:10.1016/j.nmni.2016.06.017
12. Tahmasebi H, Dehbashi S, Arabestani MR. New approach to identify colistin-resistant Pseudomonas aeruginosa by high-resolution melting curve analysis assay. Lett Appl Microbiol. 2020;70(4):290–299. doi:10.1111/lam.13270
13. Landolt P, Stephan R, Scherrer S. Development of a new High Resolution Melting (HRM) assay for identification and differentiation of Mycobacterium tuberculosis complex samples. Sci Rep. 2019;9(1):1850. doi:10.1038/s41598-018-38243-6
14. Tahmasebi H, Dehbashi S, Arabestani MR. High resolution melting curve analysis method for detecting of carbapenemases producing pseudomonas aeruginosa. JKIMSU. 2018;7(4):70–77.
15. Dehbashi S, Tahmasebi H, Arabestani MR. The clinical utility of analysis high resolution melting curve assay for simultaneous identification of methicillin and mupirocin resistant in coagulase-negative Staphylococci. Clin Lab. 2019;65(10/2019):10. doi:10.7754/Clin.Lab.2019.190314
16. Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—10th Ed. M07-A11. Wayne, PA: Clinical and Laboratory Standards Institute; 2018.
17. Teklu DS, Negeri AA, Legese MH, Bedada TL, Woldemariam HK, Tullu KD. Extended-spectrum beta-lactamase production and multi-drug resistance among Enterobacteriaceae isolated in Addis Ababa, Ethiopia. Antimicrob Resist Infect Control. 2019;8(1):39. doi:10.1186/s13756-019-0488-4
18. Osei Sekyere J. Mcr colistin resistance gene: a systematic review of current diagnostics and detection methods. MicrobiologyOpen. 2019;8(4):e00682–e. doi:10.1002/mbo3.682
19. Lund M, Petersen MB, Jorgensen AL, Paulmann D, Wang M. Rapid real-time PCR for the detection of IMP, NDM, VIM, KPC and OXA-48 carbapenemase genes in isolates and spiked stool samples. Diagn Microbiol Infect Dis. 2018;92(1):8–12. doi:10.1016/j.diagmicrobio.2018.04.002
20. Cherkaoui A, Emonet S, Renzi G, Schrenzel J. Characteristics of multidrug-resistant Acinetobacter baumannii strains isolated in Geneva during colonization or infection. Ann Clin Microbiol Antimicrob. 2015;14(1):42. doi:10.1186/s12941-015-0103-3
21. Roschanski N, Fischer J, Guerra B, Roesler U, Bereswill S. Development of a multiplex real-time PCR for the rapid detection of the predominant beta-lactamase genes CTX-M, SHV, TEM and CIT-type AmpCs in Enterobacteriaceae. PLoS One. 2014;9(7):e100956–e. doi:10.1371/journal.pone.0100956
22. Teixeira B, Rodulfo H, Carreño N, Guzmán M, Salazar E, De Donato M. Aminoglycoside resistance genes in pseudomonas aeruginosa isolates from cumana, venezuela. Rev Inst Med Trop Sao Paulo. 2016;58:13. doi:10.1590/S1678-9946201658013
23. Rahim K, Saleha S, Basit A, et al. Pseudomonas aeruginosa as a powerful biofilm producer and positive action of amikacin against isolates from chronic wounds. Jundishapur J Microbiol. 2017;10(10):e57564. doi:10.5812/jjm.57564
24. Yayan J, Ghebremedhin B, Rasche K, Webber MA. Antibiotic resistance of pseudomonas aeruginosa in pneumonia at a single University Hospital center in germany over a 10-year period. PLoS One. 2015;10(10):e0139836. doi:10.1371/journal.pone.0139836
25. Sala A, Di Ianni F, Pelizzone I, et al. The prevalence of Pseudomonas aeruginosa and multidrug resistant Pseudomonas aeruginosa in healthy captive ophidian. PeerJ. 2019;7:e6706–e. doi:10.7717/peerj.6706
26. Dou Y, Huan J, Guo F, Zhou Z, Shi Y. Pseudomonas aeruginosa prevalence, antibiotic resistance and antimicrobial use in Chinese burn wards from 2007 to 2014. J Int Med Res. 2017;45(3):1124–1137. doi:10.1177/0300060517703573
27. Ribeiro PCS, Monteiro AS, Marques SG, et al. Phenotypic and molecular detection of the blaKPC gene in clinical isolates from inpatients at hospitals in São Luis, MA, Brazil. BMC Infect Dis. 2016;16(1):737. doi:10.1186/s12879-016-2072-3
28. Farajzadeh Sheikh A, Shahin M, Shokoohizadeh L, Halaji M, Shahcheraghi F, Ghanbari F. Molecular epidemiology of colistin-resistant Pseudomonas aeruginosa producing NDM-1 from hospitalized patients in Iran. Iran J Basic Med Sci. 2019;22(1):38–42. doi:10.22038/ijbms.2018.29264.7096
29. Kotwal A, Biswas D, Kakati B, Singh M. ESBL and MBL in cefepime resistant pseudomonas aeruginosa: an update from a Rural Area in Northern India. J Clin Diagn Res. 2016;10(4):DC09–DC11. doi:10.7860/JCDR/2016/18016.7612
30. von Tippelskirch P, Gölz G, Projahn M, et al. Prevalence and quantitative analysis of ESBL and AmpC beta-lactamase producing Enterobacteriaceae in broiler chicken during slaughter in Germany. Int J Food Microbiol. 2018;281:82–89. doi:10.1016/j.ijfoodmicro.2018.05.022
31. Beyene D, Bitew A, Fantew S, Mihret A, Evans M, Ruiz-Rodriguez M. Multidrug-resistant profile and prevalence of extended spectrum β-lactamase and carbapenemase production in fermentative gram-negative bacilli recovered from patients and specimens referred to National Reference Laboratory, Addis Ababa, Ethiopia. PLoS One. 2019;14(9):e0222911–e. doi:10.1371/journal.pone.0222911
32. Bajpai T, Pandey M, Varma M, Bhatambare G. Prevalence of TEM, SHV, and CTX-M Beta-Lactamase genes in the urinary isolates of a tertiary care hospital. Avicenna J Med. 2017;7(1):12–16. doi:10.4103/2231-0770.197508
33. Büchler AC, Gehringer C, Widmer AF, Egli A, Tschudin-Sutter S. Risk factors for colistin-resistant Enterobacteriaceae in a low-endemicity setting for carbapenem resistance – a matched case–control study. Eurosurveillance. 2018;23(30):1700777. doi:10.2807/1560-7917.ES.2018.23.30.1700777
34. Cienfuegos AV, Ocampo AM, Higuita LF, Jiménez N. Risk factors for the acquisition of colistin and carbapenem-resistant Klebsiella pneumoniae in Medellín-Colombia, an endemic region for carbapenem resistance. Int J Infect Dis. 2016;53:39–40. doi:10.1016/j.ijid.2016.11.106
35. Liu YY, Wang Y, Walsh TR, et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: a microbiological and molecular biological study. Lancet Infect Dis. 2016;16(2):161–168. doi:10.1016/s1473-3099(15)00424-7
36. Tran DM, Larsson M, Olson L, et al. High prevalence of colonisation with carbapenem-resistant Enterobacteriaceae among patients admitted to Vietnamese hospitals: risk factors and burden of disease. J Infect. 2019;79(2):115–122. doi:10.1016/j.jinf.2019.05.013
37. Bontron S, Poirel L, Nordmann P. Real-time PCR for detection of plasmid-mediated polymyxin resistance (mcr-1) from cultured bacteria and stools. J Antimicrob Chemother. 2016;71(8):2318–2320. doi:10.1093/jac/dkw139
38. Daniels JB, Campbell D, Boyd S, et al. Development and validation of a clinical laboratory improvement amendments-compliant multiplex real-time PCR assay for detection of mcr genes. Microb Drug Resist. 2019;25(7):991–996. doi:10.1089/mdr.2018.0417
39. Bentaleb EM, El Messaoudi MD, Abid M, et al. Plasmid-based high-resolution melting analysis for accurate detection of rpoB mutations in Mycobacterium tuberculosis isolates from Moroccan patients. BMC Infect Dis. 2017;17(1):548. doi:10.1186/s12879-017-2666-4
40. Otaguiri ES, Morguette AEB, Morey AT, et al. Development of a melting-curve based multiplex real-time PCR assay for simultaneous detection of Streptococcus agalactiae and genes encoding resistance to macrolides and lincosamides. BMC Pregnancy Childbirth. 2018;18(1):126. doi:10.1186/s12884-018-1774-5
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.