Back to Journals » Infection and Drug Resistance » Volume 13
Prevalence and Distribution Characteristics of blaKPC-2 and blaNDM-1 Genes in Klebsiella pneumoniae
Authors Zhang X, Li F, Cui S, Mao L, Li X, Awan F, Lv W, Zeng Z
Received 3 April 2020
Accepted for publication 26 June 2020
Published 20 August 2020 Volume 2020:13 Pages 2901—2910
DOI https://doi.org/10.2147/IDR.S253631
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Suresh Antony
Xiufeng Zhang,1,2,* Fangping Li,3,* Shiyun Cui,1,2 Lisha Mao,4 Xiaohua Li,3 Furqan Awan,1,2 Weibiao Lv,5 Zhenling Zeng1,2
1College of Veterinary Medicine, Guangdong Provincial Key Laboratory of Veterinary Pharmaceutics Development and Safety Evaluation, National Risk Assessment Laboratory for Antimicrobial Resistance of Microorganisms in Animals, South China Agricultural University, Guangzhou 510642, People’s Republic of China; 2Guangdong Laboratory for Lingnan Modern Agriculture, Guangzhou 510642, People’s Republic of China; 3Guangdong Provincial Key Laboratory of Plant Molecular Breeding, State Key Laboratory for Conservation and Utilization of Subtropical Agro-Bioresources, South China Agricultural University, Guangzhou 510642, People’s Republic of China; 4Department of Clinical Laboratory, Cancer Hospital of Guangxi Medical University, Guangxi Medical University, Nanning 530021, People’s Republic of China; 5Department of Clinical Laboratory, Shunde Hospital, Southern Medical University (The First People’s Hospital of Shunde), Foshan 528000, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Weibiao Lv; Zhenling Zeng Email [email protected]; [email protected]
Background: Carbapenem-resistant Klebsiella pneumoniae infections have caused major concern and posed a global threat to public health. As blaKPC-2 and blaNDM-1 genes are the most widely reported carbapenem resistant genes in K. pneumonia, it is crucial to study the prevalence and geographical distribution of these two genes for further understanding of their transmission mode and mechanism.
Purpose: Here, we investigated the prevalence and distribution of blaKPC-2 and blaNDM-1 genes in carbapenem-resistant K. pneumoniae strains from a tertiary hospital and from 1579 genomes available in the NCBI database, and further analyzed the possible core structure of blaKPC-2 or blaNDM-1 genes among global genome data.
Materials and Methods: K. pneumoniae strains from a tertiary hospital in China during 2013– 2018 were collected and their antimicrobial susceptibility testing for 28 antibiotics was determined. Whole-genome sequencing of carbapenem-resistant K. pneumoniae strains was used to investigate the genetic characterization. The phylogenetic relationships of these strains were investigated through pan-genome analysis. The epidemiology and distribution of blaKPC-2 and blaNDM-1 genes in K. pneumoniae based on 1579 global genomes and carbapenem-resistant K. pneumoniae strains from hospital were analyzed using bioinformatics. The possible core structure carrying blaKPC-2 or blaNDM-1 genes was investigated among global data.
Results: A total of 19 carbapenem-resistant K. pneumoniae were isolated in a tertiary hospital. All isolates had a multi-resistant pattern and eight kinds of resistance genes. The phylogenetic analysis showed all isolates in the hospital were dominated by two lineages composed of ST11 and ST25, respectively. ST11 and ST25 were the major ST type carrying blaKPC-2 and blaNDM-1 genes, respectively. Among 1579 global genomes data, 147 known ST types (1195 genomes) have been identified, while ST258 (23.6%) and ST11 (22.1%) were the globally prevalent clones among the known ST types. Genetic environment analysis showed that the ISKpn7-dnaA/ISKpn27 -blaKPC-2-ISkpn6 and blaNDM-1-ble-trpf-nagA may be the core structure in the horizontal transfer of blaKPC-2 and blaNDM-1, respectively. In addition, DNA transferase (hin) may be involved in the horizontal transfer or the expression of blaNDM-1.
Conclusion: There was clonal transmission of carbapenem-resistant K. pneumoniae in the tertiary hospital in China. The prevalence and distribution of blaKPC-2 and blaNDM-1 varied by countries and were driven by different transposons carrying the core structure. This study shed light on the genetic environment of blaKPC-2 and blaNDM-1 and offered basic information about the mechanism of carbapenem-resistant K. pneumoniae dissemination.
Keywords: Klebsiella pneumoniae, blaKPC-2, *blaNDM-1, bioinformatics
Introduction
Klebsiella pneumonia, as a member of ESKAPE pathogens (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species), is the most common factor of nosocomial infections. Infections caused by K. pneumoniae are associated with a high risk of mortality and increased economic costs.1 In recent years, K. pneumoniae has been considered as a growing global threat due to its high level of resistance, particularly to those last resort antibiotics, such as carbapenems.2 In 2017, carbapenem-resistant K. pneumoniae (CRKP) has been listed in the critical priority tier of pathogens and the highest priority in new antibiotic development by WHO.3
Carbapenem resistance in K. pneumoniae involves multiple mechanisms, including the production of carbapenemases, alterations in outer membrane permeability, and the upregulation of efflux systems.4 The latter two mechanisms are often combined with other types of β-lactamases (eg, AmpC, ESBLs).5 Carbapenemases are carbapenem hydrolyzing β-lactamases and the most frequently reported in K. pneumoniae are ambler molecular class A (KPC), class B (VIM, IMP, NDM), and class D (OXA-48-like) types.6,8 The KPC-type β-lactamases were the most prevalent mechanism for carbapenem resistance in K. pneumoniae. Even though there are more than 20 different KPC variants reported, KPC-2 and -3 remain the most commonly identified variants.9 The class B β-lactamases identified in K. pneumoniae have also been found in other Enterobacteriaceae worldwide.10 The majority of NDM-1-producing K. pneumoniae strains also carried a diversity of other resistance genes but mostly remained susceptible to agents such as colistin, fosfomycin, and tigecycline.8,11 These different carbapenemase genes circulating within K. pneumoniae are often associated with mobile structures, including insertion sequences (IS), plasmids, and transposons, which caused serious challenges to clinic treatment.12
Nosocomial infection outbreaks caused by CRKP have been rapidly emerging in many countries and they are more often found in critically ill patients dwelling in intensive care units.2,13 Recently, genotypic characterization has shown that several high-risk clones or ST types were reported for the global distribution of CRKP.14 The pandemic of KPC-producing K. pneumoniae is primarily driven by clonal complex 258 (CC258).15 CC258 consists of the predominant clone ST258 and its single-locus variants (ST11, ST340, and ST512). A previous study indicated that ST11 is an emerging high-risk clone in KPC-producing K. pneumoniae, while NDM-1-producing K. pneumoniae reported widely in ST14, ST25, and ST340.4 Hence, it is crucial to carry out molecular mechanism research and epidemiological investigation on carbapenem resistance genes among K. pneumoniae in the world.
In this study, we conducted a 6-year-long longitudinal study of a tertiary care hospital in China, to study the genetic characterization of carbapenem-resistant K. pneumoniae using Whole-genome sequencing (WGS) and bioinformatic analyses. In addition, we combined the NCBI global K. pneumoniae genomes data to demonstrate the distribution and spread characteristics of blakPC-2 and blaNDM-1 around the world. The study aims to provide a theoretical basis for the rational use of clinical antibiotics and the reduction of the outbreak of nosocomial infection.
Materials and Methods
Strains Collection and Antimicrobial Susceptibility Testing
K. pneumoniae strains were collected from patients from Shunde hospital in China between 2013 and 2018. These clinical samples were routinely obtained in the clinical microbiology laboratory. Antimicrobial susceptibility testing was performed by a BD PhoenixTM 100 Automated Identification and Susceptibility Testing system (BD, USA) according to CLSI and EUCAST guidelines.16,17 The minimum inhibitory concentrations (MICs) were determined for 28 different antibiotics; amoxicillin, ampicillin, ampicillin/sulbactam, cefazolin, cefepime, cefotaxime, cefoperazone/sulbactam, ceftazidime, cefuroxime, aztreonam, piperacillin, piperacillin/tazobactam, imipenem, meropenem, chloramphenicol, florfenicol, ciprofloxacin, levofloxacin, fosfomycin, amikacin, gentamicin, neomycin, streptomycin, polymyxin B, colistin, tigecycline, tetracycline, and trimethoprim/sulfamethoxazole. For tigecycline, the EUCAST clinical MIC breakpoint for Enterobacteriaceae was used as the breakpoint for K. pneumoniae (a MIC > 2.0 mg/L is resistant).18 The reference strains Escherichia coli ATCC 25,922 was used as control strain.
Whole-Genome Sequencing (WGS)
Bacterial DNA of carbapenem-resistant K. pneumoniae (CRKP) strains were extracted using a HiPure Bacterial DNA Kit (MAGEN, China, http://www.magentec.com.cn/). The libraries were created using the VAHTSTM Universal DNA Library Prep kit for Illumina. The genomes were sequenced using the Illumina Hiseq 2500 system to obtain 2×150 bp reads. Processed reads after quality trimming were de novo assembled into contigs with a CLC Genomics Workbench 10.1 (CLC Bio, Aarhus, Denmark) and the genomes were annotated by the NCBI Prokaryotic Annotation Pipeline (PGAP).
Global Data from NCBI
As of November 2019, all K. pneumoniae genomes were downloaded from the NCBI database (https://www.ncbi.nlm.nih.gov/genome/?term=Klebsiella+pneumoniae). Then we used conventional python scripts to screen samples with complete information (including country, host, assembly level) for further analysis. A total of 1579 human samples from 44 countries, including 247 fully assembled (chromosome) level samples and 1332 scaffold level samples, were gathered to subsequent analysis (Supplementary Appendix 1).
Taxonomic Assignment
FastANI v1.1 (https://github.com/ParBLiSS/FastANI) with the core algorithm of BLAST-based ANI (ANIb) solver was used to identify the species of all isolates.19 Genome ASM24018v2 (https://www.ncbi.nlm.nih.gov/search/all/?term=%20ASM24018v2%20) was used as K. pneumoniae reference genome for species identification. Species were determined if the genome in question had >95% ANIb compared with reference genome.20 Software MLST v2.16.1 was used to perform multi-site sequence typing (MLST) of genomes.21 The ResFinder BLAST identification program (https://cge.cbs.dtu.dk/services/ResFinder/) and Isfinder (https://www-is.biotoul.fr/index.php) were used to identify acquired antibiotic resistance genes (ARGs) and mobile elements, respectively.22 The VFDB (http://www.mgc.ac.cn/VFs/) virulence database (setB) was conducted for bacterial virulence genes analysis.
Pan-Genome Analysis
Prokka v1.14 was used to produce gff file format for the contigs of K. pneumoniae genomes we collected.23 After that, core genome alignment was constructed with Roary v3.8.0 and PRANK v1.0.24 Core_genome_alignment.aln, the output file of Roary pipeline, was conveyed to fastGEAR to identify lineages by hierBAPS.25
blaKPC-2 and blaNDM-1 Loci Annotation and Comparison
Regular Python scripts and Easyfigv2.2.3 (http://easyfig.sourceforge.net/) were used to extract gene sequences. Seventy-six K. pneumoniae genomes carrying blaKPC-2 and 30 K. pneumoniae genomes carrying blaNDM-1 with complete chromosomal genomic data were selected for blaKPC-2 and blaNDM-1 gene environment analysis, respectively. Through python script, we extracted the upstream and downstream 2~6 kb sequences of the target gene for blast alignment and annotation.
Visualization of Data
ArcGIS 10.3, R package ggplot2 (http://had.co.nz/ggplot2), package pheatmap (https://stat.ethz.ch/pipermail/r-help/2012-November/330785.html) and conventional Python scripts were used for visualizing analysis. Statistical analyses were performed in R v3.6.2.
Nucleotide Accession Number
These assemblies sequence data of CRKP isolates from hospital were deposited in the GenBank database under BioProject accession PRJNA564463.
Results
CRKP Isolates of Hospital Had High Genotypic and Phenotypic Resistance
A total of 2846 K. pneumoniae strains were collected from hospital during 2013–2018 and 19 isolates showed resistance to carbapenems. The information of these strains is shown in Supplementary Appendix 2. ANI analysis confirmed these 19 strains (>98% ANIb) are K. pneumoniae. Antibiotic susceptibility testing results revealed that 94.7% (18/19) of CRKP strains showed multi-drug resistant phenotype (Figure 1B). All isolates showed 100% resistance against amoxicillin, ampicillin, ampicillin/sulbactam, cefazolin, cefepime, cefuroxime, cefoperazone/sulbactam, cefotaxime, ceftazidime, piperacillin, piperacillin/tazobactam, imipenem, and fosfomycin. None of them were resistant to polymyxins and tigecycline.
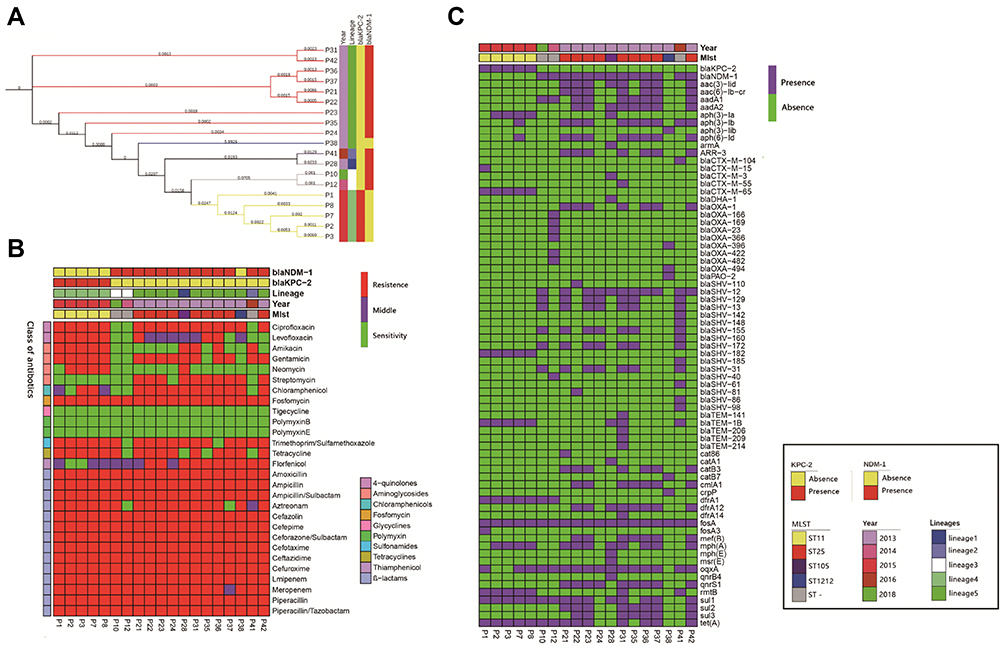 |
Figure 1 Results of phylogenetic trees; Bayesian classification, MIC test, and ARGs identification of 19 samples from ICU rooms. Sampling time, lineages of hierBAPS, presence/absence of blaKPC-2 and blaNDM-1 are annotated as colored bars next to the isolate number. (A) Phylogenetic trees of high abundance species from core genome alignments. Maximum likelihood phylogenetic trees from core genome alignments; Tree branches are colored by MLST types. (B) Results of MIC test; class of antibiotics and resistance strength are annotated as colored bars next to the main heatmap. (C) Results of ARGs identification; presence/absence of ARGs are annotated as colored bars next to the main heatmap. |
WGS analysis demonstrated that CRKP strains carried 29 unique ARGs for eight different classes of antibiotics. At least one kind of ß-lactamase genes, fosA gene, was detected in all strains. For carbapenem resistance, 26.3% (n=5) and 68.4%(n=13) of all strains carried blaKPC-2 and blaNDM-1 genes, respectively. In addition, NDM-producing strains contained more antibiotic resistance genes than KPC-producing isolates (P=0.002) (Figure 1C).
Two Lineages Dominated Hospital CRKP Populations
MLST typing showed that CRKP isolates were divided into four ST types and three unknown types. Most isolates belonged to ST25 (47.3%, n=9) and ST11 (26.3%, n=5) types, while two isolates were assigned to ST105 and ST1212, respectively. We further identified lineages with FastGEAR/BAPS results which revealed that 19 CRKP isolates contained five BAPS lineages with ST11 and ST25 relating to lineages 4 and 5, respectively. Lineages 4 and 5 represented 78.9% (n=15) of all isolates, and the differences between lineages were consistent with ST types and time collections (Figure 1A).
In addition, blaKPC-2 was only found in ST11 strains, whereas blaNDM-1 was found in ST25, ST105, and unknown ST samples (Figure 1A and C and Figure 2C). Hierarchical clustering of isolates based on ARG presence or phenotypic susceptibility indicated lineage was the major predictor of resistance-based clustering patterns (Figure 1B and C).
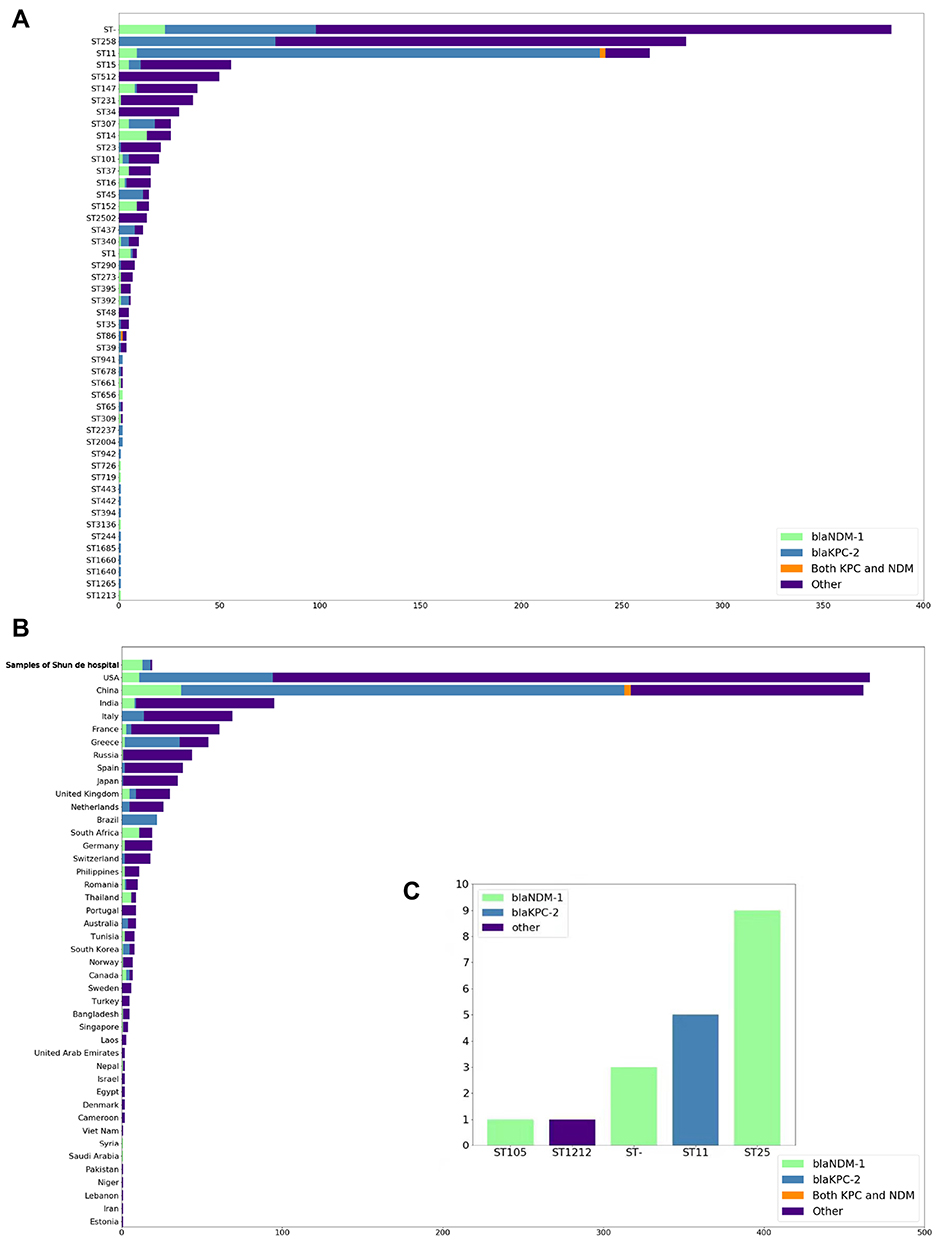 |
Figure 2 Distribution of K. pneumoniae and carbapenem resistance genes (CRGs) (blaKPC-2 and blaNDM-1); Types of CRGs are annotated by different colors. All STs are based on the Pubmlst MLST scheme. (A) Distribution of carbapenem resistance genes of CRGs in K. pneumoniae genomes from NCBI in the ST type with large abundance or with blaKPC-2 or blaNDM-1 for display. (B) Geographical distribution of CRGs in K. pneumoniae genomes from NCBI (only countries with ≥1 CRG-containing genome are shown). Countries are shown on the y-axis and the numbers on the x-axis indicate the number of CRGs. (C) Distribution of carbapenem resistance genes of CRGs in K. pneumoniae genomes from Shunde hospital. |
Geographical Genetic Distribution Characteristics of K. pneumoniae
Through MLST classification of 1579 K. pneumoniae genomes, a total of 147 known ST types (75.7%, n=1159) were identified and unknown ST types (24.3%) were found in 384 samples. Among the known ST type samples, the distribution of epidemic ST varied by counties (Figure 3). Of these, ST258 (23.6%, n=282) and ST11 (22.1%, n=264) were predominant, followed by ST15 (4.7%, n=56), ST512 (4.2%, n=50), ST147 (3.3%, n=39), and ST231 (3.1%, n=37). In addition, ST11 and ST258 were dominant in China (51.7%, n=231) and the United States (54.1%, n=252), respectively.
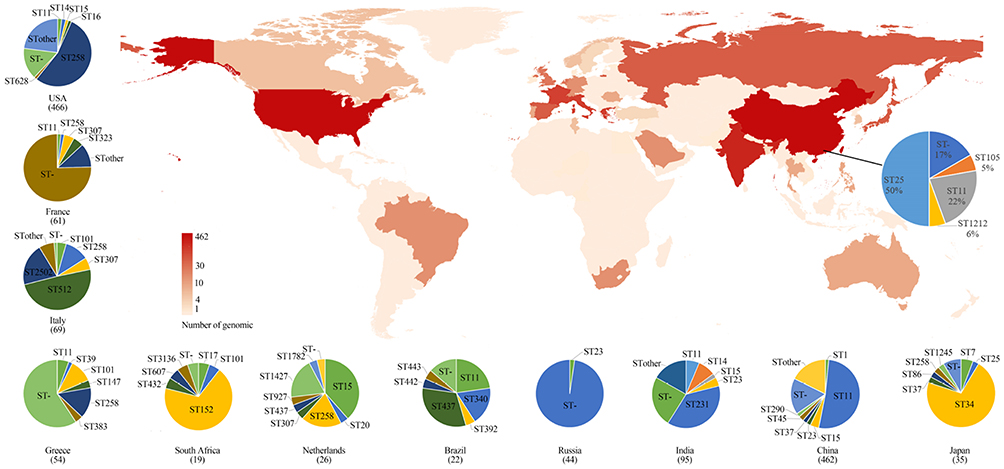 |
Figure 3 Geographic distribution and MLST typing results of K. pneumoniae samples in different countries and regions of genomic samples from NCBI and Shunde hospital; The color shades on the map represent the number of samples. The pie chart link with black line to China on the main map represents the MLST typing results of samples from Shunde hospital. The pie chart next to the main map represents MLST typing results of 13 countries. |
Then we detected the presence of blaKPC-2 and blaNDM-1 genes in 1579 genomes and found 462 genomes carried the blaKPC-2 gene and 106 had the blaNDM-1 gene, respectively. Gene blaKPC-2 was mostly detected in ST11 (88.2%), ST45 (80.0%), and ST437 (66.7%), while blaNDM-1 was mostly found in ST1 (66.7%) and ST14 (53.8%) (Figure 2A). Besides, only four genomes contained both genes and they were all isolated from China, with three ST11 isolates and one ST86 strain.
Among the dominant ST types, we found 88% of ST11 K. pneumoniae contained blaKPC-2 and 4% had blaNDM-1. However, in ST258 K. pneumoniae, only 28% of genomes carried the blaKPC-2 gene and none of them contained blaNDM-1. The presence of the blaKPC-2 and blaNDM-1 genes in China (9% and 62%) was significantly higher than those in the United States (2% and 18%) (Figure 2B).
Gene Environments of blaKPC-2 and blaNDM-1
For the blaKPC-2 gene, we found 97.4% (74/76) of the blaKPC-2 genes were located in the plasmid. Gene environment analysis found that the blaKPC-2 gene is more likely linked to mobile component ISKpn6 and ISKpn27 (46.1%, n=35)/ISKpn7-dnaA (51.3%, n=37), including those strains belonging to ST11 and ST258. Of these, ISKpn6 and ISKpn27/ISkpn7-dnaA are commonly located in blaKPC-2 gene 3ʹ and 5ʹ, respectively. Additionally, ISKpn7-dnaA, instead of ISKpn27, at blaKPC-2 gene 5ʹ was detected in the chromosome and plasmid of ST258 samples from the US and Australia. Moreover, in ST11 samples from China, the IS26 family transposon Tn3 (tnpR) was commonly found in the gene environment of blaKPC-2. In addition, the aminoglycoside resistance gene accA4 was also detected in the blaKPC-2 gene environment of ST258 samples from the US (Figure 4A).
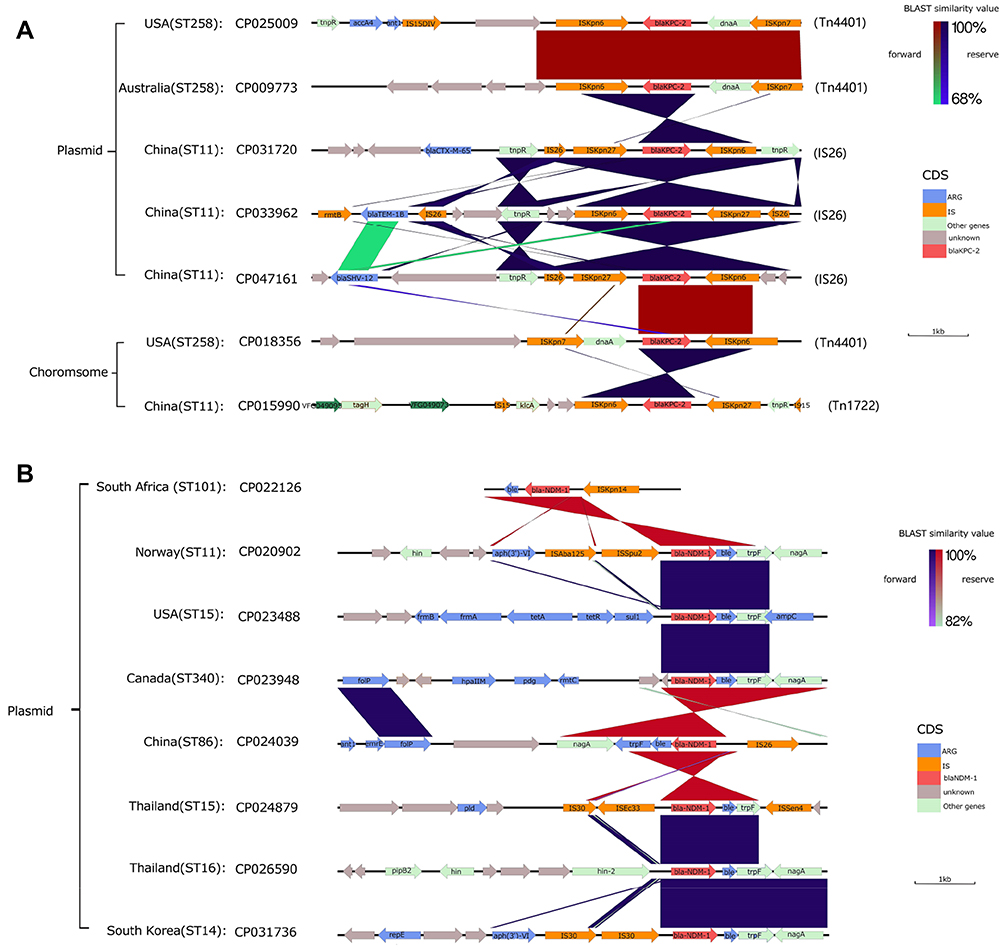 |
Figure 4 blaKPC-2 and blaNDM-1 gene environment and comparison of some representative samples; BLAST similarity values and the types of different CDS are annotated by different colors next to the main chart (A) Gene environment of blaKPC-2 (B) Gene environment of blaNDM-1. |
For the blaNDM-1 gene environment, the blaNDM-1 gene was all located in plasmids of all genomes (Figure 4B). In most samples, the blaNDM-1 was mostly link with IS30 family mobile elements such as ISAba125. However, the IS630 family mobile element ISSpu2 and DNA transferase (hin) encoding gene fragments were detected in 5ʹ of blaNDM-1 gene of ST11 genomes from Norway, two hins were found in 5ʹ of blaNDM-1 gene of ST16 genomes from Thailand. Antibiotic resistance genes were also detected in the blaNDM-1 gene environment, such as the tetracycline resistance gene (tet), aminoglycoside antibiotic resistance gene (aph), and bleomycin resistance (ble, trpF, and nagA).
Discussion
K. pneumoniae is responsible for human infections and the patient population is the most important reservoir in high-frequency nosocomial K. pneumoniae outbreaks.26 The emergence of K. pneumoniae carrying carbapenemases and their worldwide dissemination posed a significant public health crisis.27 As blaKPC-2 and blaNDM-1 are the most widely reported among CRKP worldwide, we used genomic and phylogenetic approaches to analyze CRKP isolated from hospital in China and uncover the distribution of these two genes in global genomes from NCBI.
Our findings indicated that the CRKP population from hospital is highly diverse, encompassing five lineages and different STs. All these CRKP isolates were resistant to multiple antibiotics and sensitive to polymyxins and tigecycline, suggesting resistance in CRKP isolates is serious. In particular, ST11 CRKP have been associated with the global dissemination of K. pneumoniae, all the ST types found in hospital were also widely reported in China.28 Interestingly, through core genome phylogenetic analysis, there was no significant genetic difference in the core genome except the sample P38, which indicated there is high homology between samples in different time of the hospitals. Interestingly, P38 did not contain blaKPC-2 or blaNDM-1, it only carried blaOXA-396 and blaOXA-494, which may relate to carbapenem resistance.
Hence, based on the actual collection time of the samples (at least in different months of the same year), we can infer that there are certain nosocomial infections and clonal transmission in the time segment of the hospital.
MLST typing results showed that blaKPC-2 was only detected in ST11-CRKP (5/19) strains from hospital. According to the global genome data, 88.25% (n=264) of ST11 strains contained blaKPC-2 and 4.5% (n=12) carried blaNDM-1. These results suggested that ST11 CRKP strains have obvious preference for carrying blaKPC-2, which is consistent with previous studies.29,30 In addition, ST11 also has a relatively high level of carrying blaNDM-1 in NDM-1 producing CRKP strains (n=106). In addition, the blaKPC-2 gene was also found in ST45, ST307, and ST437 K. pneumoniae isolates, among which ST437 was the dominant ST-type in Brazil. These three ST K. pneumoniae were widely distributed in the global scope and had no obvious locality, hence it was worth exploring the underlying spread mechanism.31,32
Notably, CRKP isolates in China carried a relatively high proportion of blaKPC-2 and blaNDM-1, especially these strains carrying both genes only found in China. Co-occurrence of blaNDM-1 and blaKPC-2 in a clinical K. pneumoniae isolate was considered to result in broad-spectrum antibiotic resistance profiles.33 This phenomenon has been reported in India, South Africa, and China.34,35 This may indicate that circulating isolates in China are more likely to develop pan drug resistance. Interestingly, all ST25 samples (9/19) from hospital carried blaNDM-1, while the NDM-1 producing. This preference has not been previously reported and is worth further exploration.
It was noted that blaKPC-2 together with its adjacent mobile component formed three core structures through genetic analysis, including the n3-ISKpn27-blaKPC-2-ISKpn6 chain, ISKpn6-blaKPC-2-ISKpn27-IS26, and ISKpn6-blaKPC-2-ISKpn7-dnaA. This phenomenon means that the core structures ISKpn27-blaKPC-2 -ISKpn6 and ISKpn6-blaKPC-2-ISKpn7-dnaA are likely to carry blaKPC-2 to transfer in the global scope. The first two structures were called Tn1722-based unit transposon and IS26-based composite transposon, respectively.36,38 The insertion sequences ISKpn6 and ISKpn7-dnaA were often reported in transposon Tn4401, which was the main genetic structure enhancing the spread of the blaKPC-type genes in different plasmid scaffolds.36 In addition, the presence of various resistance genes was detected in the blaKPC-2 environment of ST11 samples from China. The interaction between these resistance genes and blaKPC-2 is worthy to further investigate.
We found that there were ISAba125, trpF, ISSen4, and IS family elements detected in the environment of blaNDM-1. In addition, the upstream and downstream of the blaNDM-1 gene often contained transposons (Tn3) or inserted sequence fragments (IS30), which were often involved in horizontal transfer of drug resistance genes.39 In most NDM-1 producing samples, ble, trpF, and nagA are closely linked downstream, hence it can be considered that blaNDM-1-ble-trpf-nagA may be the core structure of horizontal transfer of blaNDM-1.40 It was noted that hin was also detected in 5ʹ of the blaNDM-1 gene of some ST11 and ST16 genomes. Hin is a special recombinant binding enzyme that can promote the inversion of DNA position, which has been reported to be involved in the inversion control of DNA segment H in the flagellum phase of Salmonella enterica.41 This hints that hin may be involved in the horizontal transfer or expression control of blaNDM-1.
Conclusion
In this study, we found all CRKP strains were dominated by two lineages composed of ST25 and ST11. Phylogenetic analysis showed that there was clonal transmission of CRKP in the hospital. Bioinformatic analysis of 1597 NCBI samples revealed that ST11 and ST258 were the most detected clones and both ST types had a preference for carrying the blaKPC-2 gene. The core structure of ISKpn27/ISKpn7-dnaA-blaKPC-2-ISKpn6 and blaNDM-1-ble-trpF-nagA is highly epidemic in KPC- and NDM-1-producing K. pneumoniae. Our study provides a complete genetic background and geographical distribution for further understanding the transmission mode and mechanism of blaKPC-2 and blaNDM-1.
Acknowledgment
This work was supported by the National Natural Science Foundation of China (Grant No. 31672608) and Guangdong Special Support Program Innovation Team 2019BT02N054.
Disclosure
All authors declare that they have no conflict of interest.
References
1. Martin RM, Bachman MA. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Front Cell Infect Microbiol. 2018;8:4. doi:10.3389/fcimb.2018.00004
2. Ripabelli G, Tamburro M, Guerrizio G, et al. Tracking multidrug-resistant Klebsiella pneumoniae from an Italian hospital: molecular epidemiology and surveillance by PFGE, RAPD and PCR-based resistance genes prevalence. Curr Microbiol. 2018;75(8):977–987. doi:10.1007/s00284-018-1475-3
3. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18(3):318–327. doi:10.1016/S1473-3099(17)30753-3
4. Pitout JDD, Nordmann P, Poirel L. Carbapenemase-producing Klebsiella pneumoniae, a key pathogen set for global nosocomial dominance. Antimicrob Agents Chemother. 2015;59(10):5873–5884. doi:10.1128/AAC.01019-15
5. Sawa T, Kooguchi K, Moriyama K. Molecular diversity of extended-spectrum beta-lactamases and carbapenemases, and antimicrobial resistance. J Intensive Care. 2020;8:13. doi:10.1186/s40560-020-0429-6
6. Candan ED, Aksöz N. Klebsiella pneumoniae: characteristics of carbapenem resistance and virulence factors. Acta Biochim Pol. 2015;62:867–874. doi:10.18388/abp.2015_1148
7. Di Tella D, Tamburro M, Guerrizio G, et al. Molecular epidemiological insights into colistin-resistant and carbapenemases-producing clinical Klebsiella pneumoniae Isolates. Infect Drug Resist. 2019;12:3783–3795. doi:10.2147/IDR.S226416
8. Lee CR, Lee JH, Park KS, et al. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: epidemiology, genetic context, treatment options, and detection methods. Front Microbiol. 2016;7:895. doi:10.3389/fmicb.2016.00895
9. Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13:785–796. doi:10.1016/S1473-3099(13)70190-7
10. Abbas HA, Kadry AA, Shaker GH, Goda RM. Impact of specific inhibitors on metallo-beta-carbapenemases detected in Escherichia coli and Klebsiella pneumoniae isolates. Microb Pathog. 2019;132:266–274. doi:10.1016/j.micpath.2019.05.022
11. Girmenia C, Serrao A, Canichella M. Epidemiology of carbapenem resistant Klebsiella pneumoniae infections in Mediterranean countries. Mediterr J Hematol Infect Dis. 2016;8(1):e2016032. doi:10.4084/MJHID.2016.032
12. Cerdeira LT, Lam MMC, Wyres KL, et al. Small IncQ1 and col-like plasmids harboring bla KPC-2 and non-Tn4401 elements (NTEKPC-IId) in high-risk lineages of Klebsiella pneumoniae CG258. Antimicrob Agents Chemother. 2019;63(3):e02140–18. doi:10.1128/AAC.02140-18
13. Abramowicz L, Gerard M, Martiny D, et al. Infections due to carbapenemase-producing bacteria, clinical burden, and impact of screening strategies on outcome. Med Mal Infect. 2020:
14. Ferrari C, Corbella M, Gaiarsa S, et al. Multiple Klebsiella pneumoniae KPC clones contribute to an extended hospital outbreak. Front Microbiol. 2019;10:2767. doi:10.3389/fmicb.2019.02767
15. Chen L, Mathema B, Chavda KD, et al. Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 2014;22(12):686–696. doi:10.1016/j.tim.2014.09.003
16. Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, M100. 28th Informational Supplement. Wayne, PA: Clinical and Laboratory Standards Institute; 2018.
17. EUCAST. European committee on antimicrobial susceptibility testing. Breakpoint tables for interpretation of MICs and zone diameters. Available from: http://www.eucast.org/fileadmin/src/media/PDF s/EUCAST_files/Breakpoint_tables/v_8.0_Breakpoint_Tables.pdf.
18. Juan C-H, Huang Y-W, Lin Y-T, et al. Risk factors, outcomes, and mechanisms of tigecycline-nonsusceptible Klebsiella pneumoniae bacteremia. Antimicrob Agents Chemother. 2016;60(12):7357–7363. doi:10.1128/AAC.01503-16
19. Goris J, Konstantinidis KT, Klappenbach JA, et al. DNA-DNA hybridization values and their relationship to whole-genome sequence similarities. Int J Syst Evol Microbiol. 2007;57(1):81–91. doi:10.1099/ijs.0.64483-0
20. D’Souza AW, Potter RF, Wallace M, et al. Spatiotemporal dynamics of multidrug resistant bacteria on intensive care unit surfaces. Nat Commun. 2019;10(1):4569. doi:10.1038/s41467-019-12563-1
21. Maiden MC, Van Rensburg MJJ, Bray JE, et al. MLST revisited: the gene-by-gene approach to bacterial genomics. Nat Rev Microbiol. 2013;11(10):728–736. doi:10.1038/nrmicro3093
22. Kichenaradja P, Siguier P, Perochon J, et al. ISbrowser: an extension of ISfinder for visualizing insertion sequences in prokaryotic genomes. Nucleic Acids Res. 2010;38(suppl_1):D62–D68. doi:10.1093/nar/gkp947
23. Seemann T. Prokka: rapid prokaryotic genome annotation. Bioinformatics. 2014;30(14):2068–2069. doi:10.1093/bioinformatics/btu153
24. Page AJ, Cummins CA, Hunt M, et al. Roary: rapid large-scale prokaryote pan genome analysis. Bioinformatics. 2015;31(22):3691–3693. doi:10.1093/bioinformatics/btv421
25. Mostowy R, Croucher NJ, Andam CP, et al. Efficient inference of recent and ancestral recombination within bacterial populations. Mol Bio Evol. 2017;34(5):1167–1182. doi:10.1093/molbev/msx066
26. Gorrie CL, Mirceta M, Wick RR, et al. Gastrointestinal carriage is a major reservoir of Klebsiella pneumoniae infection in intensive care patients. Clin Infect Dis. 2017;65(2):208–215. doi:10.1093/cid/cix270
27. Effah CY, Sun T, Liu S, et al. Klebsiella pneumoniae: an increasing threat to public health. Ann Clin Microbiol Antimicrob. 2020;19(1):1. doi:10.1186/s12941-019-0343-8
28. Gheitani L, Fazeli H, Moghim S, et al. Frequency determination of carbapenem-resistant Klebsiella pneumoniae (CRKP) isolated from hospitals in Isfahan of Iran and evaluation of synergistic effect of colistin and meropenem on them. Cell Mol Biol. 2018;64(1):70–74. doi:10.14715/cmb/2018.64.1.13
29. Zhan L, Wang S, Guo Y, et al. Outbreak by hypermucoviscous Klebsiella pneumoniae ST11 isolates with carbapenem resistance in a tertiary hospital in China. Front Cell Infect Microbiol. 2017;7:182. doi:10.3389/fcimb.2017.00182
30. Fu P, Tang Y, Li G, et al. Pandemic spread of blaKPC-2 among Klebsiella pneumoniae ST11 in China is associated with horizontal transfer mediated by IncFII-like plasmids. Int J Antimicrob Agents. 2019;54(2):117–124. doi:10.1016/j.ijantimicag.2019.03.014
31. Andrade LN, Curiao T, Ferreira JC, et al. Dissemination of blaKPC-2 by the spread of Klebsiella pneumoniae clonal complex 258 clones (ST258, ST11, ST437) and plasmids (IncFII, IncN, IncL/M) among Enterobacteriaceae species in Brazil. Antimicrob Agents Chemother. 2011;55(7):3579–3583. doi:10.1128/AAC.01783-10
32. Loconsole D, Accogli M, De Robertis AL, et al. Emerging high-Risk ST101 and ST307 carbapenem-resistant Klebsiella pneumoniae clones from bloodstream infections in southern Italy. Ann Clin Microbiol Antimicrob. 2020;19(1):24. doi:10.1186/s12941-020-00366-y
33. Kumarasamy K, Kalyanasundaram A. Emergence of Klebsiella pneumoniae isolate co-producing NDM-1 with KPC-2 from India. J Antimicrob Chemother. 2012;67(1):243–244. doi:10.1093/jac/dkr431
34. Brink AJ, Coetzee J, Clay CG, et al. Emergence of New Delhi metallo-beta-lactamase (NDM-1) and Klebsiella pneumoniae carbapenemase (KPC-2) in South Africa. J Clin Microbiol. 2012;50(2):525–527. doi:10.1128/JCM.05956-11
35. Gao H, Liu Y, Wang R, et al. The transferability and evolution of NDM-1 and KPC-2 co-producing Klebsiella pneumoniae from clinical settings. EBioMedicine. 2020;51:102599. doi:10.1016/j.ebiom.2019.102599
36. Wang L, Fang H, Feng J, et al. Complete sequences of KPC-2-encoding plasmid p628-KPC and CTX-M-55-encoding p628-CTXM coexisted in Klebsiella pneumoniae. Front Microbiol. 2015;6:838. doi:10.3389/fmicb.2015.00838
37. Li G, Zhang Y, Bi D, et al. First report of a clinical, multidrug-resistant Enterobacteriaceae isolate coharboring fosfomycin resistance gene fosA3 and carbapenemase gene blaKPC-2 on the same transposon, Tn1721. Antimicrob Agents Chemother. 2015;59(1):338–343. doi:10.1128/AAC.03061-14
38. Chen YT, Lin JC, Fung CP, et al. KPC-2-encoding plasmids from Escherichia coli and Klebsiella pneumoniae in Taiwan. J Antimicrob Chemother. 2014;69(3):628–631. doi:10.1093/jac/dkt409
39. Nordmann P, Poirel L, Walsh TR, et al. The emerging NDM carbapenemases. Trends Microbiol. 2011;19(12):588–595. doi:10.1016/j.tim.2011.09.005
40. Kutsukake K, Nakashima H, Tominaga A, et al. Two DNA invertases contribute to flagellar phase variation in Salmonella enterica serovar Typhimurium strain LT2. J bacteriol. 2006;188(3):950–957. doi:10.1128/JB.188.3.950-957.2006
41. Feng JA, Johnson RC, Dickerson RE. Hin recombinase bound to DNA: the origin of specificity in major and minor groove interactions. Science. 1994;263(5145):348–355. doi:10.1126/science.8278807
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.