Back to Journals » Journal of Inflammation Research » Volume 19
Postoperative Abdominal Infectious Complications Promote Colon Cancer Metastasis by Inducing MDSCs to Release Neutrophil Extracellular Traps
Authors Cheng T ![]() , Liu C, Zang J, Jin X, Wu Y, Huang W, Sui L, Luo X, Zhang X, Guo L, Tian W, Cui J, Ren J, Qi Z, Cao L, Yang L
, Liu C, Zang J, Jin X, Wu Y, Huang W, Sui L, Luo X, Zhang X, Guo L, Tian W, Cui J, Ren J, Qi Z, Cao L, Yang L
Received 16 December 2025
Accepted for publication 5 June 2026
Published 16 July 2026 Volume 2026:19 585132
DOI https://doi.org/10.2147/JIR.S585132
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Felix Marsh-Wakefield
Tianwei Cheng,1 Chang Liu,1 Juanhao Zang,1 Xiandu Jin,1 Yulin Wu,1 Wenjun Huang,1 Lele Sui,1 Xiyue Luo,1 Xiaoxiao Zhang,2 Lihong Guo,2 Wencong Tian,3 Jianlin Cui,1 Jiling Ren,4 Zhi Qi,1– 3 Lei Cao,1,3 Liang Yang1
1Department of Molecular Pharmacology, School of Medicine, Nankai University, Tianjin, 300071, People’s Republic of China; 2Shengli Oilfield Central Hospital, Shandong, 257034, People’s Republic of China; 3Department of General Surgery, Tianjin Union Medical Center, The First Affiliated Hospital of Nankai University, Nankai University, Tianjin, 300121, People’s Republic of China; 4Department of Pathogen Biology, Basic Medical College, Tianjin Medical University, Tianjin, 300070, People’s Republic of China
Correspondence: Liang Yang, Email [email protected] Lei Cao, Email [email protected]
Purpose: Surgical resection is the primary treatment for colon cancer. Postoperative abdominal infectious complications (AIC) are associated with poor prognosis, but their mechanistic role in promoting metastasis remains unclear. This study aimed to investigate the association between AIC and metastasis, focusing on the potential involvement of myeloid-derived suppressor cells (MDSCs) and neutrophil extracellular traps (NETs).
Patients and Methods: Peripheral blood from colon cancer patients with (AIC, n = 20) or without (Non-AIC, n = 20) postoperative abdominal infectious complications was analyzed by flow cytometry to assess MDSC frequency and NET markers. A mouse model of postoperative infection following tumor resection was established. Metastatic burden was evaluated using in vivo imaging and histopathological analysis. MDSC function and NET induction were further examined using in vitro co-culture assays.
Results: In patients, MDSC and NET markers were elevated on postoperative day 3. By day 7, levels remained significantly higher in AIC patients compared to Non-AIC patients. In mice, AIC triggered a systemic expansion of MDSCs in peripheral blood, bone marrow, and ascites, accompanied by increased NET formation. These AIC-induced MDSCs and NETs promoted tumor cell migration and accelerated liver and peritoneal metastasis in vivo. Critically, degrading NETs with DNase I markedly reduced metastatic burden without exacerbating sepsis.
Conclusion: Postoperative infection is associated with enhanced colon cancer metastasis, and experimental data indicate that the MDSC-NETs axis is involved in this process. Pharmacological disruption of NETs with DNase I represents a potential therapeutic strategy that warrants further investigation for improving postsurgical outcomes.
Keywords: Colon cancer, Postoperative abdominal infectious complication, Myeloid-derived suppressor cells, Neutrophil extracellular traps, Metastasis
Introduction
Colon cancer is the third most common malignancy and the second leading cause of cancer deaths worldwide, posing a major public health challenge.1,2 Surgical resection remains the primary treatment and can be performed via open, laparoscopic, or robotic approaches.3 Nevertheless, surgery alone is limited by a postoperative recurrence rate of 33.3% and a five-year survival rate of 10%–40%, highlighting the need to improve outcomes.4,5 Furthermore, the colon’s physiological and anatomical characteristics make colon procedures more prone to abdominal infection than gastric or small intestinal operations.6 Surgery induces a transient sterile inflammatory response mediated by damage-associated molecular patterns (DAMPs). By contrast, postoperative infectious complications, such as anastomotic leakage, introduce pathogen-associated molecular patterns (PAMPs), which amplify and sustain the inflammatory response.7,8 Clinical evidence indicates that postoperative abdominal infectious complications (AIC) can promote tumor recurrence and metastasis, thereby adversely affecting long-term survival.9,10 Nevertheless, the biological mechanisms by which postoperative infection facilitates metastatic progression remain incompletely understood.
Myeloid-derived suppressor cells (MDSCs) are immature myeloid cells with potent immunosuppressive activity that facilitate tumor progression.11,12 Studies using mouse models of peritonitis have shown that MDSCs in the spleen can remain elevated for several months after infection.13 Under infectious conditions, inflammatory factors such as IL-6 can significantly promote the proliferation and activation of MDSCs through signaling pathways such as STAT3.14–16 Clinical and preclinical studies show that MDSCs accumulate at metastatic sites and that their abundance correlates with poor prognosis and increased metastatic risk.17,18 Importantly, emerging evidence suggests that MDSCs can generate neutrophil extracellular traps (NETs) in response to specific microenvironmental cues.19,20 NETs are web-like structures composed of DNA backbones decorated with histones and granule proteins and are classically released by neutrophils during infection.21,22 NETs have been shown to facilitate metastasis by trapping circulating tumor cells, reactivating dormant tumor cells in the bone marrow, and promoting tumor cell migration through chemokine signaling.23,24 However, whether MDSC-mediated metastasis in the context of postoperative infection involves NET formation in colon cancer remains unclear.
In this study, we show that abdominal infectious complications after colon cancer surgery induce a systemic expansion of MDSCs and stimulate NET release. We further demonstrate that AIC-driven MDSCs and NETs promote metastatic progression in vitro and in vivo. Crucially, degrading NETs with DNase I reduces liver metastasis (LM) and peritoneal metastasis (PM) in AIC mice without worsening sepsis, suggesting a potential therapeutic strategy to improve outcomes in patients with colon cancer.
Materials and Methods
Clinical Samples
Plasma samples were collected from colon cancer patients who underwent curative surgical resection at the First Affiliated Hospital of Nankai University. Inclusion criteria were: (1) histologically confirmed colon adenocarcinoma; (2) no prior history of neoadjuvant chemotherapy or radiotherapy. Exclusion criteria included: (1) presence of other concurrent malignancies; (2) incomplete clinical or follow-up data.
Postoperative abdominal infectious complications (AIC) were defined as the occurrence of one or more of the following conditions: (1) Intra-abdominal infection or abscess: diagnosed by the presence of abdominal pain, fever (body temperature ≥38.0°C), purulent drainage from an intra-abdominal drain, or radiological evidence of intra-abdominal fluid collection or abscess on CT or ultrasound, with or without confirmation during relaparotomy; (2) Anastomotic leakage: defined by the presence of abdominal pain, fever, or gastrointestinal contents in the surgical drain, or extravasation of contrast on water-soluble contrast enema or CT; (3) Positive bacterial culture: confirmed by microbial culture of fluid obtained from postoperative abdominal puncture or drainage.
Patients meeting any of the above criteria were assigned to the AIC group. Patients who underwent surgery during the same period but developed no postoperative infectious complications, or developed non-infectious complications were assigned to the Non-AIC group. Based on these criteria, a total of 40 patients were included in the final analysis: 20 in the AIC group and 20 in the Non-AIC group. Baseline demographic, clinical, and pathological characteristics of the patient cohorts are summarized in Supplementary Table 1. The AIC and Non-AIC groups were well-balanced across most variables, with no significant differences observed (all P > 0.05). All participants provided written informed consent. The study protocol was approved by the Nankai University Institutional Review Board (approval number: NKUIRB2026125).
Animal Experiments
All mouse experiments were approved by the Institutional Animal Care and Use Committee of Nankai University (approval number: 2026-SYDWLL-000714). For the tumor resection mouse model, CT-26, 1×106 cells were subcutaneously implanted into the right flank of BALB/c mice (female, 6–8 weeks old, weighing 25 ± 1 g). 12 days later, 90% of the tumors were removed. The wound was then closed using standard suture techniques. For AIC model, we referred to cecal ligation puncture (CLP) model. After anesthetizing and laparotomy, the cecum without ligation was perforated (CP) with 25G needle. Sham mice were subjected to the same surgical procedure, including anesthesia and laparotomy, but without CP. After surgery, mice were euthanized by cervical dislocation to extract interested organs. To ensure uniform infection severity, peritoneal lavage was performed immediately after cecal puncture using 1 mL of sterile PBS. The lavage fluid was serially diluted, plated on solid culture medium, and incubated overnight at 37°C. Colony-forming units (CFUs) were counted the following day, and only mice with a confirmed bacterial load within the target range of 1 × 104 to 5 × 104 CFU/mL were included in the AIC group for subsequent experiments. For the LM and PM models, more than 1×106 CT-26 cells were injected into the spleen or abdomen under anesthesia three days after tumor resection surgery.25–27 Treatments including PBS or DNase I (2.5 mg/kg, MedChemExpress) were administrated intraperitoneally every 3 days. Mice were sacrificed after anesthesia at the end of the experiment, and blood and liver samples were collected for further analysis. The maximum allowable tumor diameter in any dimension must not exceed 20 mm. The anesthetic agent was isoflurane.
ELISA Assay
The levels of myeloperoxidase (MPO) in serum from patients and mice were tested by ELISA. The operation was strictly based on the instructions of the Human MPO ELISA Kit (ELK Biotechnology, 10187) and the mouse MPO ELISA Kit (ELK Biotechnology, 10310).
Flow Cytometry and MDSC Isolation
Venous peripheral blood samples were collected from Non-AIC and AIC patients and mice. Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Paque density gradient centrifugation. Cells were stained in FACS buffer at room temperature with fluorochrome-conjugated antibodies. For human MDSCs, we employed the internationally accepted consensus definition: Lin− HLA-DR− CD11b⁺ CD33⁺ cells (Supplementary Figure S1A). For mouse MDSCs, total MDSCs were identified as CD11b⁺ Gr-1⁺ cells, as Gr-1 recognizes both Ly6G and Ly6C epitopes (Figure S1C). For multicolor flow cytometry, compensation was performed automatically using single-stained compensation beads for each fluorochrome prior to data acquisition, and fluorescence-minus-one (FMO) controls were used to define positive gates. FACS sorting was performed on a BD FACSAria II, and data were analyzed using FlowJo software (v10.8.1).
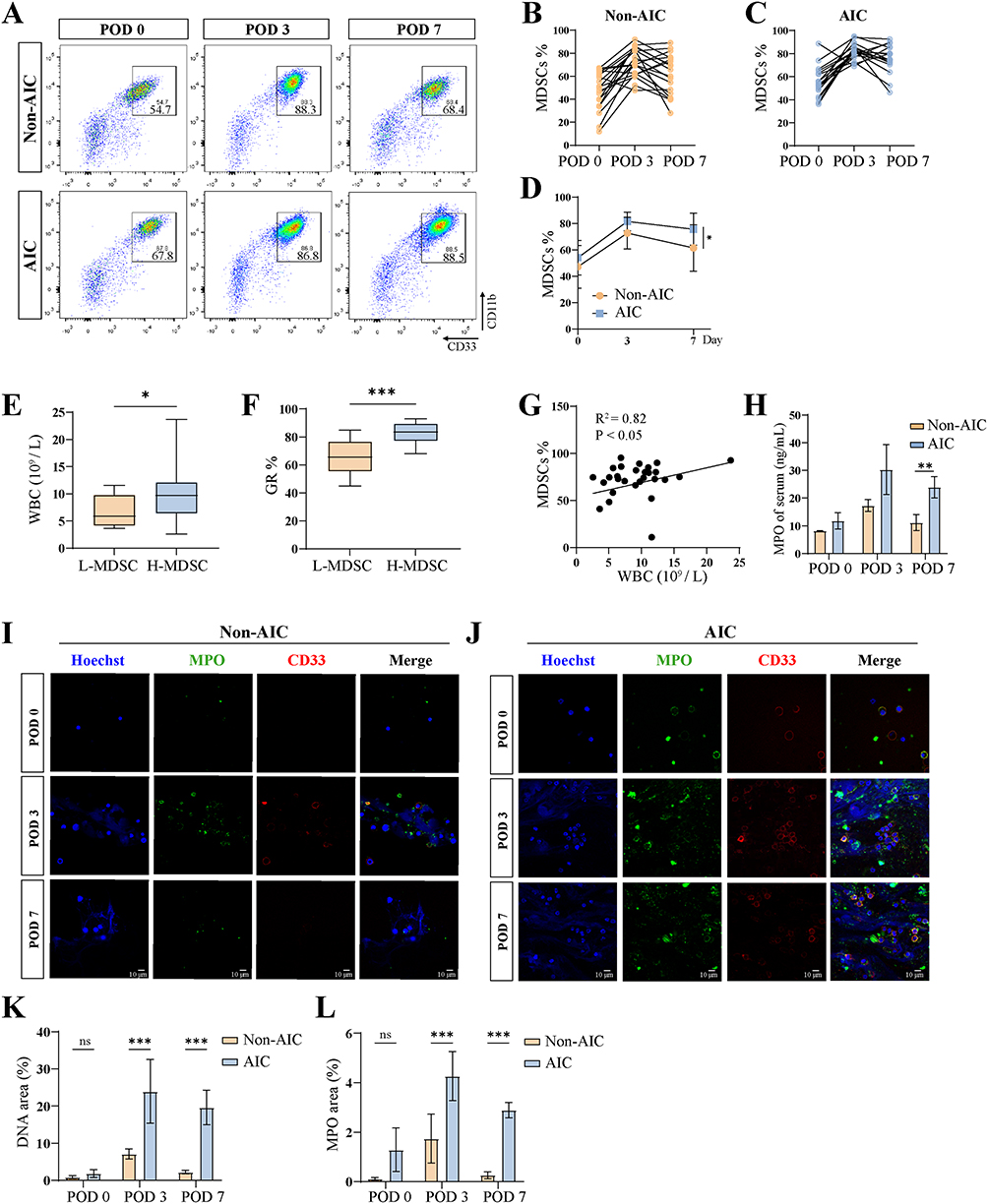 |
Figure 1 Postoperative AIC promotes sustained NET release from MDSCs. (A) Representative flow cytometric analyses of peripheral blood from patients with Non-AIC (n = 20) and AIC (n = 20). (B–D) The relative percentages of MDSCs are shown at each time point. (E) WBC counts in patients stratified into low- and high-MDSC (L-MDSC and H-MDSC) groups based on the median MDSCs level at POD 7. (F) Proportion of granulocyte in patients with L-MDSC and H-MDSC. (G) Correlation analysis between WBC counts and MDSCs %. (H) Serum levels of MPO in patients with Non-AIC and AIC (n = 20 per group). (I and J) Representative immunofluorescence co-staining of DNA (Hoechst) and MPO (myeloperoxidase) to assess NET formation in MDSCs isolated from peripheral blood of Non-AIC and AIC at each time point, scale bar = 10 μm. (K and L) Fluorescence quantification of DNA and MPO on POD 7 (n = 4 independent experiments). Data are presented as the mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant. |
Immunofluorescence Staining
Isolated MDSCs were fixed with 4% paraformaldehyde for 15 min at room temperature. After washing with PBS twice, samples were blocked with 5% goat serum for one hour at room temperature and were incubated with antibodies overnight at 4°C. After washing with PBS twice, samples were incubated with appropriate Alexa Fluor 488- or Alexa Fluor 594-conjugated secondary antibodies (Beijing Solarbio Science & Technology) for one hour at room temperature. Samples were stained with Hoechst 33342 (MedChemExpress, HY-15559) for 4 min at room temperature. Images were acquired with a FV1000 confocal microscope (Olympus, USA). For NET quantification, DNA (Hoechst 33342) area and MPO area were quantified using a semi-automated method applied to fluorescence microscopy images of MDSCs, as previously described.28,29 Quantification was performed using ImageJ software. The primary antibodies used in this investigation were: CD33 (ABclonal, A22704), MPO (Zen bioscience, R25062), CD11b (Solarbio, K01785RR). All outcome evaluations and preliminary statistical analyses were completed by researchers unaware of the grouping.
Wound Healing Assay and Transwell Assay
For transwell migration, MDSCs from patients and mice were placed in lower chambers. CT-26 cells were seeded in upper chambers for 24 hours incubation. Finally, four random fields were imaged for ImageJ analysis. For wound healing assay, Cells in the logarithmic growth phase were placed in six-well plates until they reached 95% confluence. A straight wound was created using a 200 μL sterile pipette tip across the cell layers after 24 hours of seeding. The cell debris was removed by washing with PBS, and the remaining cells were then incubated in a medium containing 1% FBS at 37 °C with 5% CO2. Photographs of the wounded cell layers were taken immediately after scratching and again 24 hours later.
Western Blotting
All cells were lysed with RIPA buffer supplemented with a protease inhibitor cocktail. The lysates were incubated on ice for 30 min and then centrifuged at 15,000 × g for 30 min at 4 °C. The protein concentration was determined using the bicinchoninic acid assay (BCA). Proteins were separated by 12% SDS-PAGE and subsequently transferred to PVDF membranes. The membranes were blocked with 5% skimmed milk (PSAITONG, S10191) for 2 h at room temperature, followed by incubation with primary antibodies at 4 °C overnight. The membranes were then incubated with horseradish peroxidase-conjugated secondary antibody for 1 h at room temperature. The target bands were detected by the ECL solution system (Shandong Sparkjade Biotechnology Co., Ltd.) and were analyzed using ImageJ. The primary antibodies used included Vimentin (Zen bioscience, R22775), GAPDH (Zen bioscience, R380646).
Statistical Analysis
All statistical analyses were conducted by GraphPad Prism 9.0. Data are shown as mean ± SEM. Student’s t-test was used to compare between two groups. For multiple-group comparisons, one-way analysis of variance was used, followed by Tukey’s post hoc multiple comparison test. Statistical significance was set at P < 0.05. A post-hoc power analysis was performed using an independent samples t-test based on the observed difference in MDSC frequency between the AIC and Non-AIC groups (Cohen’s d = 0.99). With a two-sided α of 0.05 and 20 patients per group, the achieved statistical power was 87%.
Results
Postoperative AIC Promotes Sustained NET Release from MDSCs
MDSCs were quantified by flow cytometry in the Non-AIC and AIC patients before surgery (POD 0) and on postoperative days 3 (POD 3) and 7 (POD 7). In both Non-AIC and AIC patients, MDSCs in peripheral blood increased significantly by the third day after surgery (Figure 1A–C). However, by POD 7, the MDSCs proportion was significantly higher in the AIC group than in the Non‑AIC group (Figure 1D). Expression of genes associated with the MDSC signature (ARG1 and NOS2) was obviously upregulated in PBMCs on POD 3 (Figure S1B). Based on the POD 7 MDSC levels, patients were classified into a low‑MDSC group (<72%, n = 20) and a high‑MDSC group (≥72%, n = 20). The high‑MDSC group exhibited significantly elevated peripheral white blood cell (WBC) and granulocyte (GR) counts (Figure 1E and F), and MDSC proportion correlated positively with WBC count (Figure 1G). Serum MPO, a marker of NETs, was also significantly higher in the AIC group than in the Non‑AIC group (Figure 1H). To investigate whether postoperative infection enhances NET release by MDSCs, we isolated MDSCs from patients through flow sorting, cultured for 2 hours, and performed immunofluorescence co‑staining for DNA and MPO. NET levels, quantified by DNA/MPO area, increased by POD 3 compared to baseline in both groups, with significantly higher levels in the AIC group than in the Non‑AIC group at this time point (Figure 1I and J). By POD 7, NET levels in the Non‑AIC group had returned to near‑baseline levels, whereas they remained elevated in the AIC group (Figure 1K and L). Together, these findings indicate that postoperative abdominal infection promotes sustained NET release from MDSCs.
Postoperative AIC Drives MDSC Expansion and NET Release in a Colon Cancer Mouse Model
To further investigate the impact of postoperative infection on MDSCs and NET formation, we established a mouse model of colon cancer by subcutaneous injection of CT-26 cells. Primary tumor resection was performed to simulate the clinical scenario of surgical tumor removal. A subset of mice subsequently underwent 25G needle cecal puncture without ligation (CP) to establish a post-colon-cancer-surgery infection model (Figure 2A). Serum ELISA revealed significantly higher IL-6 levels in the AIC group compared to the Non-AIC group, indicating an enhanced systemic inflammatory response (Figure 2B). Flow cytometric analysis on postoperative day 7 revealed that the proportion of MDSCs (CD11b⁺Gr-1⁺) in the spleen, bone marrow (BM), and PBMCs was significantly increased in the Non-AIC group compared with the control group. Furthermore, the proportion of MDSCs in the AIC group was markedly higher than that in the Non-AIC group (Figure 2C–F). Expression of genes associated with the MDSC signature (Arg1 and Nos2) was obviously upregulated in the AIC group (Figure S1D). MDSC proportions positively correlated with serum IL-6 levels (Figure 2G). Similarly, serum MPO levels were significantly higher in the AIC group than in the Non-AIC group, suggesting enhanced NET formation (Figure 2H). Immunofluorescence staining further demonstrated that NET release from MDSCs was more pronounced in the AIC group than in the Non-AIC group (Figure 2I–K). To determine whether IL-6, a key cytokine elevated in the serum of AIC mice (Figure 2B), directly drives NETs of MDSCs, we performed in vitro stimulation experiments. FACS-sorted MDSCs were cultured with recombinant IL-6, with or without Tocilizumab (IL-6 receptor inhibitor). As shown in Supplementary Figure S2A and B, IL-6 stimulation significantly increased extracellular DNA release and MPO from MDSCs compared to unstimulated controls, indicating enhanced NETs. Importantly, treatment with Tocilizumab markedly attenuated IL-6-induced NET release. These results demonstrate that IL-6 signaling is sufficient to induce NETs in MDSCs. Together, these findings validate our clinical observations, confirming that postoperative infection promotes MDSC expansion and enhances NET release.
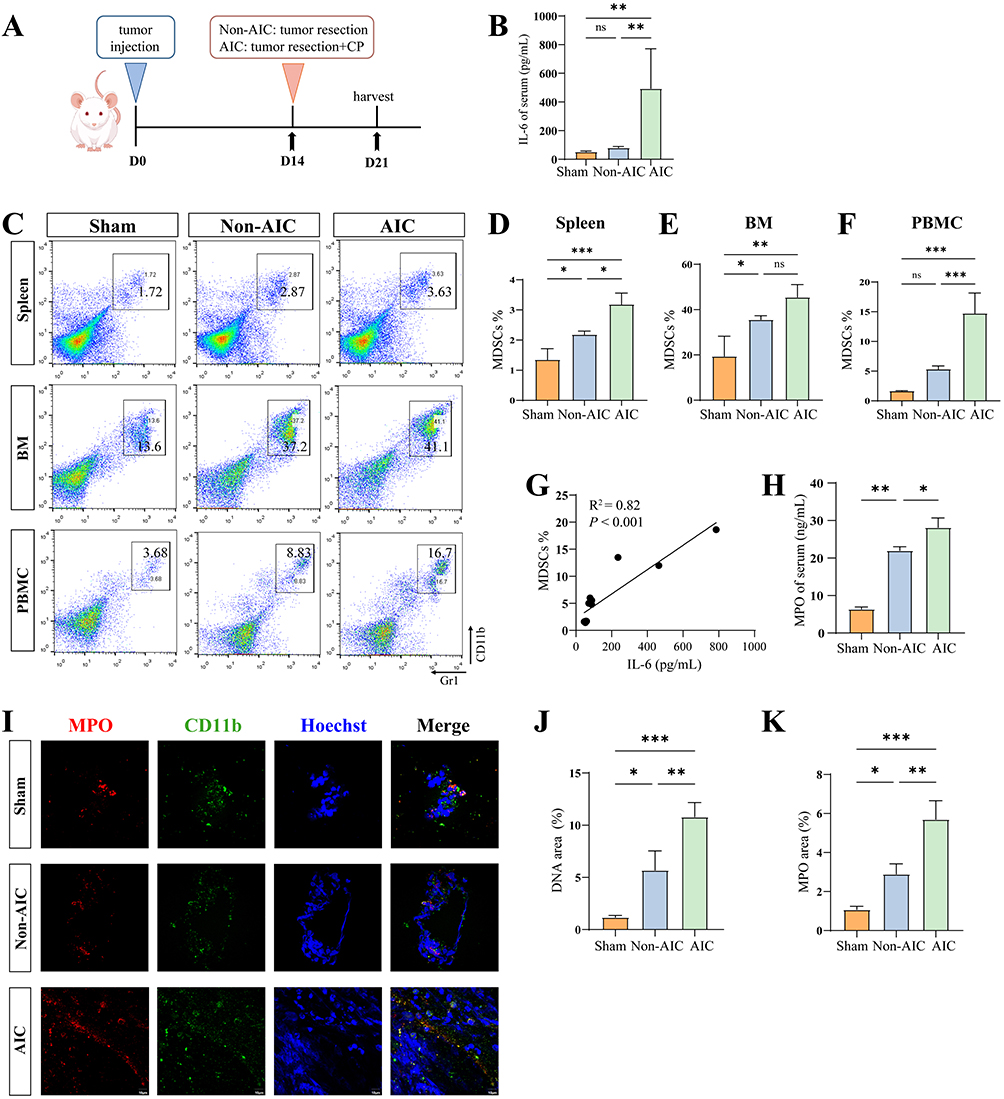 |
Figure 2 Postoperative AIC drives MDSC expansion and NET release in a colon cancer mouse model. (A) Schematic diagram of Non-AIC and AIC mouse model. (B) Serum levels of IL-6 in mice (n = 5 per group) with Non-AIC and AIC. (C–F) The relative percentages of MDSCs are shown in spleen, BM, and PBMCs from mice (n = 5 per group) at 7 days after surgery (MDSCs: CD11b+Gr1+). (G) Correlation analysis between levels of IL-6 and MDSCs %. (H) Serum levels of MPO in mice (n = 5 per group) with Non-AIC and AIC. (I–K) Representative immunofluorescence co-staining of DNA and MPO to assess NET formation in MDSCs isolated from peripheral blood of Non-AIC and AIC mice (n = 5 per group), scale bar = 10 μm. Data are presented as the mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant. |
Infection-Activated MDSCs Drive Tumor Migration Through Induction of Epithelial-Mesenchymal Transition in vitro
To test whether MDSCs from the postoperative infection microenvironment promote colon cancer cell migration, we co-cultured HCT116 cells with MDSCs isolated from AIC and quantified migration using a wound healing assay. AIC-derived MDSCs significantly increased HCT116 migration (Figure 3A and B). Western blot analysis showed that co-culture with MDSCsAIC elevated Vimentin, a principal mesenchymal marker of epithelial-mesenchymal transition (EMT), in HCT116 cells (Figure 3C and D). To explore the role of MDSC-derived NETs in facilitating tumor cell migration, MDSCs isolated from AIC mice were co-cultured with CT-26 colon cancer cells in the presence of DNase I (to degrade NETs) or the PAD4 inhibitor GSK484 (PAD4i, to inhibit NET formation). As illustrated in the wound healing assays (Figure 3E and F) and Transwell migration assays (Figure 3G and H), treatment with either DNase I or PAD4i significantly reduced the migration of CT-26 cells compared to the CT-26 + MDSCsAIC group. Notably, DNase I exhibited a stronger inhibitory effect on migration than PAD4i in the Transwell assay. Western blotting analysis (Figure 3I and J) showed that DNase I treatment markedly downregulated Vimentin expression, whereas no statistically significant change in Vimentin levels was observed following PAD4i treatment.
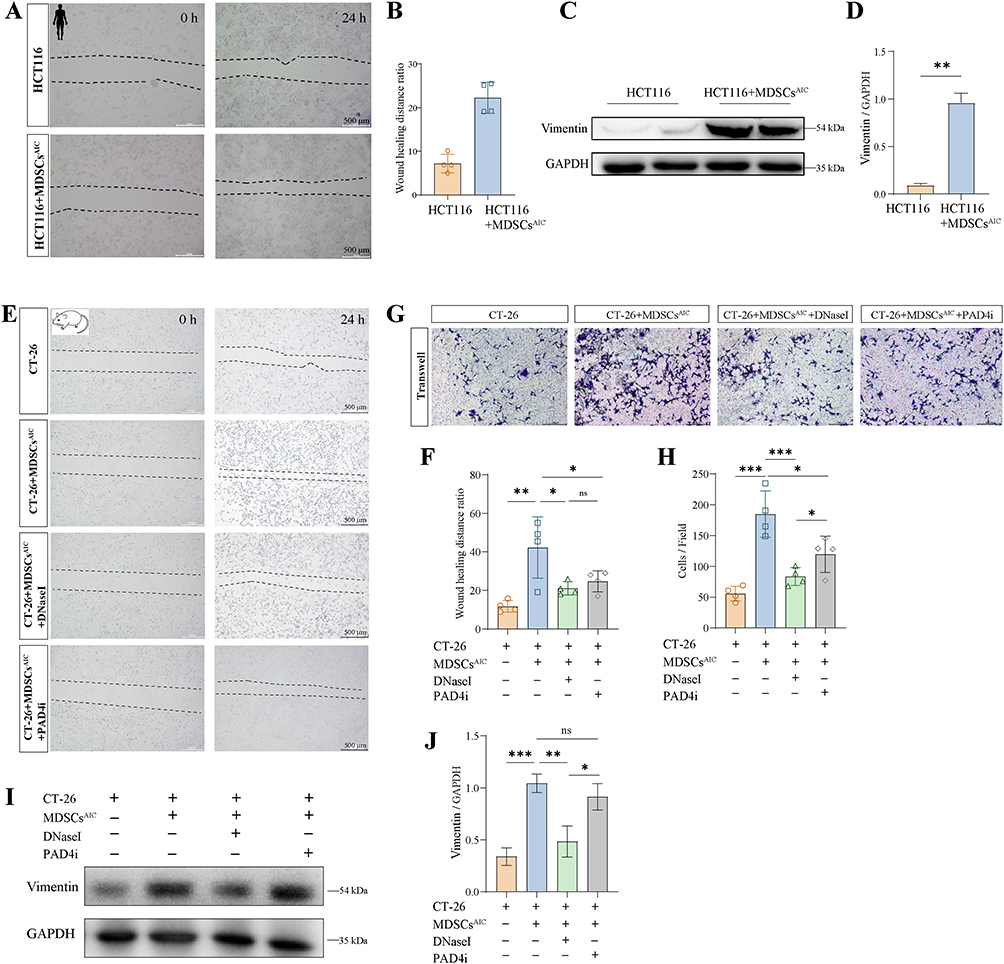 |
Figure 3 Infection-activated MDSCs drive tumor migration through induction of epithelial-mesenchymal transition in vitro. (A and B) HCT116 cells were co-cultured with MDSCs from AIC patients. Wound healing assays to assess the migration ability of HCT116. (C and D) Western blot analysis was conducted for Vimentin in HCT116. (E and F) Wound healing assay showing migration of CT‑26 cells co-cultured with MDSCs from AIC mice treated with DNase I (0.25 mg/mL) or PAD4i (10μM). (G and H) Transwell migration assay of CT‑26 cells co-cultured with MDSCs from Non-AIC or AIC mice under the same conditions. (I and J) Western blot analysis of Vimentin expression in CT-26 cells under the same conditions. n = 4 biologically independent experiments. Data are presented as mean ± SEM. *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant. |
Postoperative Infection Enhances Tumor Metastasis via Systemic Expansion of MDSCs in vivo
To assess how postoperative AIC influences tumor metastasis in vivo, we resected primary tumors in mice and then induced a postoperative infection using CP. We subsequently established LM and PM metastasis models (Figure 4A). In vivo imaging showed more extensive PM in the AIC group than in the Non-AIC group (Figure 4B and C). Macroscopic inspection and H&E staining of livers revealed a significantly greater area of hepatic metastatic nodules in the AIC group compared with the Non-AIC group (Figure 4D and E). PBMC, BM, and ascites (ASC) were collected from each group for flow cytometric analysis. This analysis showed a marked increase in the proportion of MDSCs across PBMC, BM, and ASC in the AIC group relative to the Non-AIC group (Figure 4F–I). These findings indicate that postoperative infection promotes tumor metastasis through the systemic expansion of MDSCs in vivo.
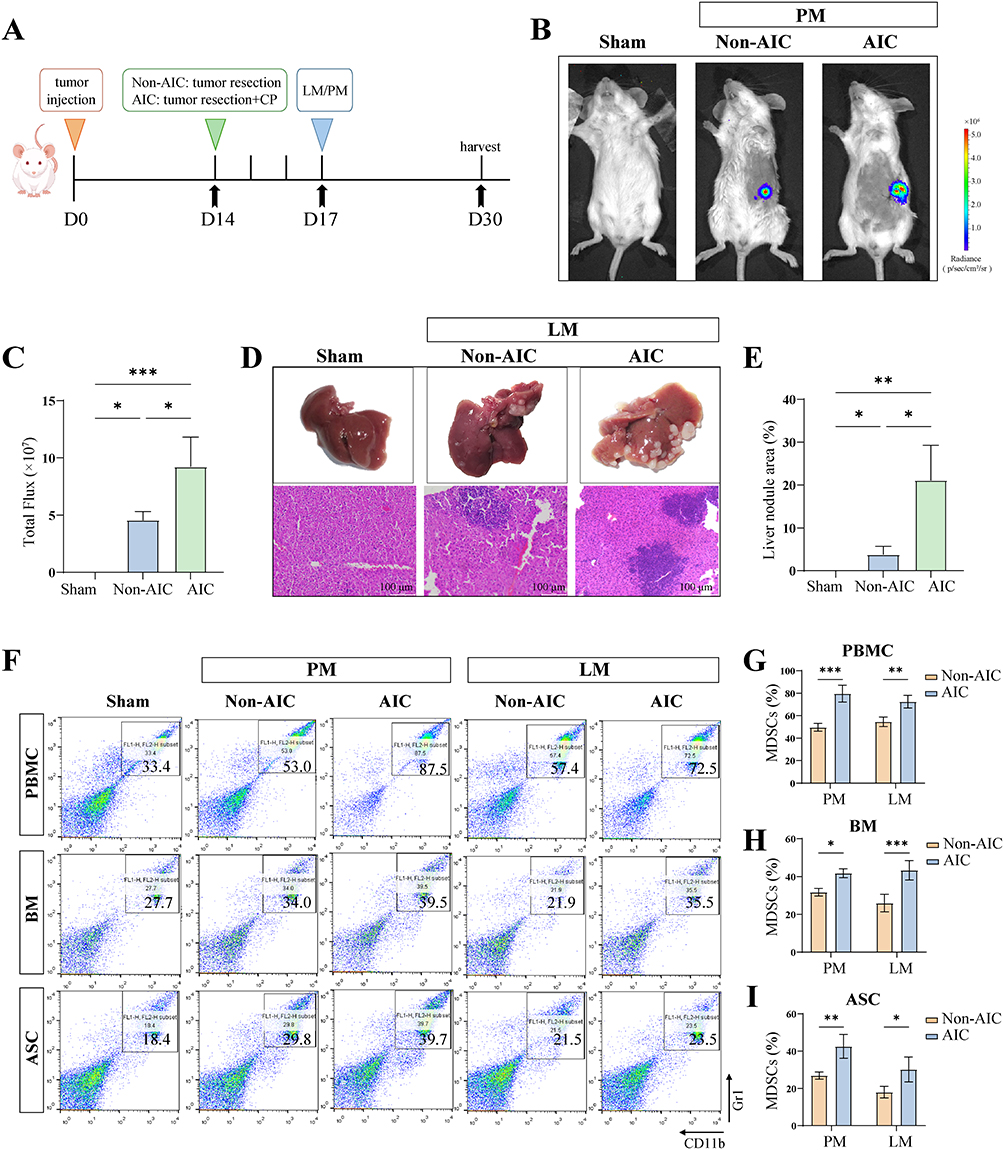 |
Figure 4 Postoperative infection enhances tumor metastasis via systemic expansion of MDSCs in vivo. (A) Experimental timeline. (B and C) Representative images and quantification of metastatic lesions in PM mice models (n = 4 per group). (D and E) Representative images and quantification of liver nodules and their HE stained images in LM mice (n = 4 per group), scale bar = 100 μm. (F) Representative flow cytometric analyses of MDSCs from mice with Non-AIC and AIC groups (n = 4 independent experiments). The relative percentages of MDSCs are shown in (G) PBMC, (H) BM, and (I) ASC. PBMCs and BM from LM mice, ASC from PM mice (n = 4 per group). Data are presented as the mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant. |
DNase I-Mediated NET Clearance Attenuates Postoperative AIC-Driven Metastasis
To assess the contribution of NETs to tumor metastasis, mice received postoperative treatment with the NETs-degrading agent DNase I. In vivo imaging showed that DNase I reduced peritoneal metastasis in the AIC group (Figure 5A and B). As shown in Figure 5C, DNase I treatment did not exacerbate weight loss in AIC mice. Consistently, analysis of hepatic metastases indicated that although the AIC group developed significantly larger liver nodules than the Non-AIC group, DNase I treatment substantially decreased the area of nodule in AIC mice (Figure 5D and E). The liver-to-body weight ratio was also significantly lower in the AIC+DNase I group than in the AIC group (Figure 5F). Flow cytometric analysis of peripheral blood from the LM model revealed a significant reduction in the proportion of MDSCs after DNase I treatment (Figure 5G and H). Correspondingly, serum MPO levels were reduced in the AIC+DNase I group, confirming decreased NET formation after treatment (Figure 5I). Together, these results indicate that targeting NETs with DNase I effectively mitigates postoperative AIC associated tumor metastasis.
 |
Figure 5 DNase I-mediated NET clearance attenuates postoperative AIC-driven metastasis. (A and B) Representative images and quantification of metastatic lesions in PM mice with Non-AIC, AIC and AIC+DNase I groups (n = 4 per group). (C) Body weight changes were monitored throughout the experimental period in the indicated groups (Sham, Non-AIC, AIC, and AIC+DNase I, n = 4 per group). (D and E) Representative images and quantification of liver nodules in LM mice with Non-AIC, AIC and AIC+DNase I groups (n = 4 per group). (F) Percentage of liver weight to body weight in LM mice with Non-AIC, AIC and AIC+DNase I groups (n = 4 per group). (G and H) Representative flow cytometric analyses of MDSCs are shown in PBMCs from LM mice with Non-AIC, AIC and AIC+DNase I groups (n = 4 per group). (I) Serum levels of MPO in LM mice with Non-AIC, AIC and AIC+DNase I groups (n = 4 per group). Data are presented as the mean ± SEM, *P < 0.05, **P < 0.01, ***P < 0.001, ns, not significant. |
Discussion
While surgery itself induces a sterile inflammatory response that is typically transient, the superimposition of postoperative infection (AIC) introduces a more intense and sustained inflammatory cascade. Based on standardized clinical criteria, patients were divided into AIC and Non-AIC groups. In Non-AIC patients, MDSCs showed a transient elevation on postoperative day 3, returning to near-baseline levels by day 7. In contrast, AIC patients exhibited persistently elevated MDSC levels at day 7. Notably, patients with high MDSC levels had significantly higher white blood cell and neutrophil counts than those with low MDSC levels. In our experimental models, AIC was associated with MDSC expansion and enhanced NETs release, and treatment with DNase I reduced liver and peritoneal metastasis in AIC-bearing mice.
MDSCs are immature myeloid cells that exhibit effective immunosuppressive properties and participate in the formation of immunosuppressive pathology under inflammatory and tumor-bearing conditions.30,31 There is growing evidence that MDSCs can also regulate the immune response to infection, acute and chronic inflammation, traumatic stress and sepsis.32–34 It is reported that patients with a continued increase in the percentage of MDSCs had extended ICU stays.35 In this study, we observed an increased proportion of MDSCs on POD 7 in colon cancer patients who developed postoperative infections, suggesting that AIC is associated with MDSC expansion. This finding was recapitulated in our mouse model, where MDSC percentages were similarly elevated in AIC mice.
Recent research has shown that tumor-associated MDSCs play an important role in tumor progression.36 In vitro, MDSCs from AIC patients or mice promoted tumor migration and EMT of tumor cells. In vivo results showed that the proportion of MDSCs in AIC mice increased, and tumor metastasis increased significantly. These results suggest that highly activated MDSCs generated due to infection are more likely to accumulate around residual tumors after surgery, leading to enhanced local tumor growth.
NETs are web-like structures composed of related proteins and DNA released by neutrophils and are involved in promoting tumor cell invasion and metastasis.37,38 In human solid tumors, the presence of NETs correlates positively with neutrophil abundance and is closely associated with unfavorable clinical outcomes.39,40 In this study, we provide evidence that circulating NETs levels are elevated in AIC mice, and that MDSCs isolated from these animals exhibit enhanced NET release capacity. Therapeutic degradation of NETs with DNase I was associated with reduced metastatic burden in AIC mice. Notably, the weaker effect of PAD4i compared to DNase I in the migration assays may be explained by the fact that DNase I directly degrades pre‑formed NETs, whereas PAD4i inhibits the formation of new NETs. Beyond trapping circulating tumor cells, NETs actively remodel the tumor microenvironment to promote metastasis through multiple mechanisms. They contribute to extracellular matrix degradation via proteases such as matrix metalloproteinase 9 (MMP9), facilitating tumor cell invasion.28 Furthermore, NET-associated proteins like cathelicidin and histones also promote angiogenesis, supporting the vascularization of micrometastases.39 Together, these functions suggest that NETs reprogram target organs into a pro-metastatic “soil”. In the context of AIC, sustained MDSC-derived NETs may thus promote metastasis not only by trapping tumor cells but also through matrix remodeling, enhanced angiogenesis, and local immune suppression.
Our data suggest that postoperative infection is associated with MDSC expansion and elevated NETs levels. IL-6 appears to be one of the drivers of NETs formation in this context, as in vitro stimulation with IL-6 induced NETs release from MDSCs. However, the AIC-induced cytokine storm is complex and comprises multiple factors beyond IL-6, and other inflammatory mediators likely contribute as well. NET formation can be induced by a variety of cytokines, including C5a.20,41,42 Therefore, identifying which specific cytokines drive NET formation in MDSCs will be an important direction for future research.
Metastasis is also driven by multiple interconnected mechanisms. Beyond the MDSC-NETs axis identified here, the tumor microenvironment (TME) comprises cancer-associated fibroblasts (CAFs), tumor-associated macrophages (TAMs), and diverse immune subsets, whose dynamic crosstalk critically shapes metastatic outcomes.43 Recent single-cell studies have shown that inflammatory CAFs interact metabolically and immunologically with myeloid cells, promoting immunosuppressive TAM subsets that collectively form a pro-metastatic niche.44 In parallel, several molecular pathways also contribute. The CXCL12-CXCR4 axis supports pre-metastatic niche formation and epithelial-mesenchymal transition (EMT), recruiting cancer cells to distant organs.45 Ferroptosis regulated by SLC7A11 has emerged as a key factor in colorectal cancer progression and therapy response.46 Exosome-mediated communication further conditions distant sites by transferring miRNAs and proteins that prime the pre-metastatic soil.47 In the context of AIC, this persistent inflammatory environment may simultaneously affect multiple TME components, and future studies employing single-cell approaches will be essential to dissect these complex cellular interactions. Our study now identifies the MDSC-NETs axis as an infection-triggered pathway that may intersect with these mechanisms, for example, through NETs modulating chemokine gradients, influencing exosome activity, or altering ferroptosis sensitivity. Understanding these intersections could guide combinatorial strategies to better prevent or treat postoperative metastasis in colon cancer.
This study has several limitations. First, while our in vitro functional data supports a role for MDSC-derived NETs in promoting tumor cell migration, definitive in vivo causal validation using conditional MDSC depletion models represents an important direction for future research. Second, the precise molecular signals that trigger MDSC activation and NETs release remain incompletely defined. Third, whether the composition and function of MDSC-derived NETs differ from those of neutrophil-derived NETs warrants further investigation. Finally, our clinical cohort, while sufficient for the primary analysis, is modest in size and derived from a single center; larger multicenter studies are needed to validate our findings.
Conclusion
This study identifies an association between postoperative infection in colon cancer and increased metastatic potential. Elevated MDSC levels and enhanced NET formation were observed in both AIC patients and mouse models. In vitro, MDSCs derived from AIC mice released NETs, accompanied by increased tumor cell migration and EMT; DNase I degraded NETs, and was associated with reduced metastatic burden in AIC mice. These findings suggest that the MDSC-NETs axis may play a role in metastasis associated with postoperative infection, warranting further investigation.
Data Sharing Statement
All relevant data that supports the findings of this study are available from the corresponding author upon reasonable request.
Ethics Statement
This study was approved by the Nankai University Institutional Review Board (approval number: NKUIRB2026125). All participants were informed and signed the written informed consent. This research was conducted in accordance with guidelines outlined in the Declaration of Helsinki.
All mouse experiments were approved by the Institutional Animal Care and Use Committee of Nankai University (approval number: 2026-SYDWLL-000714). The reporting of this study adheres to the ARRIVE guidelines.
Acknowledgments
We would like to thank the School of Medicine, Nankai University, for support in all animal experiments.
Author Contributions
Tianwei Cheng: Conceptualization, Methodology, Formal analysis, Writing – original draft, Writing – review & editing. Liang Yang, Zhi Qi, Jiling Ren, Lei Cao: Resources, Writing – review & editing. Chang Liu, Juanhao Zang, Xiandu Jin, Yulin Wu, Wenjun Huang, Lele Sui, Xiyue Luo: Data curation, Writing – review & editing. Xiaoxiao Zhang, Lihong Guo, Wencong Tian, Jianlin Cui: Formal analysis, Writing – review & editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (Nos. 32271327, 82572525, 82372195) Tianjin Natural Science Foundation (24JCYBJC01850, 24JCYBJC01010, 25JCZDJC00980), Tianjin Health Research Project (TJWJ2023XK014, TJWJ2025ZD013), Scientific Research Project of Tianjin Union Medical Center (2022PWXK03, 2022PWXK05) and Nankai University Optometry Institute Key Project (NKSGZ202413).
Disclosure
The authors report no conflicts of interest for this study.
References
1. Noronha MM, Costa Almeida LF, Cappellaro AP, et al. Neoadjuvant chemotherapy for colon cancer: a systematic review and meta-analysis of randomized controlled trials. Eur J Cancer. 2025;222:115476. doi:10.1016/j.ejca.2025.115476
2. Fillon M. Colon cancer blood test effective for average-risk population. CA Cancer J Clin. 2024;74(4):317–14. doi:10.3322/caac.21859
3. Ricciardi R, Seshadri-Kreaden U, Yankovsky A, et al. The COMPARE study: comparing perioperative outcomes of oncologic minimally invasive laparoscopic, da vinci robotic, and open procedures: a systematic review and meta-analysis of the evidence. Ann Surg. 2025;281(5):748–763. doi:10.1097/SLA.0000000000006572
4. Oppedijk V, van der Gaast A, van Lanschot JJ, et al. Patterns of recurrence after surgery alone versus preoperative chemoradiotherapy and surgery in the CROSS trials. J Clin Oncol. 2014;32(5):385–391. doi:10.1200/JCO.2013.51.2186
5. Duineveld LA, van Asselt KM, Bemelman WA, et al. Symptomatic and asymptomatic colon cancer recurrence: a multicenter cohort study. Ann Fam Med. 2016;14(3):215–220. doi:10.1370/afm.1919
6. Antonio N, Bønnelykke-Behrndtz ML, Ward LC, et al. The wound inflammatory response exacerbates growth of pre-neoplastic cells and progression to cancer. EMBO j. 2015;34(17):2219–2236. doi:10.15252/embj.201490147
7. Matsui R, Sagawa M, Sano A, et al. Impact of perioperative immunonutrition on postoperative outcomes for patients undergoing head and neck or gastrointestinal cancer surgeries: a systematic review and meta-analysis of randomized controlled trials. Ann Surg. 2024;279(3):419–428. doi:10.1097/SLA.0000000000006116
8. Xia X, Zhang Z, Zhu C, et al. Neutrophil extracellular traps promote metastasis in gastric cancer patients with postoperative abdominal infectious complications. Nat Commun. 2022;13(1):1017. doi:10.1038/s41467-022-28492-5
9. Koedam TWA, Bootsma BT, Deijen CL, et al. Oncological outcomes after anastomotic leakage after surgery for colon or rectal cancer: increased risk of local recurrence. Ann Surg. 2022;275(2):e420–e7. doi:10.1097/SLA.0000000000003889
10. Rosowski EE, Huttenlocher A. Neutrophils, wounds, and cancer progression. Dev Cell. 2015;34(2):134–136. doi:10.1016/j.devcel.2015.07.005
11. Hamilton-Reeves JM, Bechtel MD, Hand LK, et al. Effects of immunonutrition for cystectomy on immune response and infection rates: a pilot randomized controlled clinical trial. Eur Urol. 2016;69(3):389–392. doi:10.1016/j.eururo.2015.11.019
12. Tang F, Tie Y, Tu C, Wei X. Surgical trauma-induced immunosuppression in cancer: recent advances and the potential therapies. Clin Transl Med. 2020;10(1):199–223. doi:10.1002/ctm2.24
13. Ruan WS, Feng MX, Xu J, et al. Early activation of myeloid-derived suppressor cells participate in sepsis-induced immune suppression via PD-L1/PD-1 Axis. Front Immunol. 2020;11:1299. doi:10.3389/fimmu.2020.01299
14. Li H, Chen X, Zheng S, et al. The expansion of MDSCs induced by exosomal PD-L1 promotes the progression of gastric cancer. J Transl Med. 2024;22(1):821. doi:10.1186/s12967-024-05611-y
15. An J, Feng L, Ren J, et al. Chronic stress promotes breast carcinoma metastasis by accumulating myeloid-derived suppressor cells through activating β-adrenergic signaling. Oncoimmunology. 2021;10(1):2004659. doi:10.1080/2162402X.2021.2004659
16. Neo SY, Tong L, Chong J, et al. Tumor-associated NK cells drive MDSC-mediated tumor immune tolerance through the IL-6/STAT3 axis. Sci Transl Med. 2024;16(747):eadi2952. doi:10.1126/scitranslmed.adi2952
17. He S, Zheng L, Qi C. Myeloid-derived suppressor cells (MDSCs) in the tumor microenvironment and their targeting in cancer therapy. Mol Cancer. 2025;24(1):5. doi:10.1186/s12943-024-02208-3
18. Zheng L, Wu D, Xie H, Zhao H. Revisiting Tregs in cancer and beyond: immunological control and therapeutic potential. Front Immunol. 2025;16:1581093. doi:10.3389/fimmu.2025.1581093
19. Haykal T, Yang R, Tohme C, et al. Surgery-induced neutrophil extracellular traps promote tumor metastasis by reprogramming cancer cell lipid metabolism. Cancer Res. 2025;85:4995–5014. doi:10.1158/0008-5472.CAN-24-3393
20. Ortiz-Espinosa S, Morales X, Senent Y, et al. Complement C5a induces the formation of neutrophil extracellular traps by myeloid-derived suppressor cells to promote metastasis. Cancer Lett. 2022;529:70–84. doi:10.1016/j.canlet.2021.12.027
21. Herro R, Grimes HL. The diverse roles of neutrophils from protection to pathogenesis. Nat Immunol. 2024;25(12):2209–2219. doi:10.1038/s41590-024-02006-5
22. Froy O, Weintraub Y. The circadian clock, metabolism, and inflammation-the holy trinity of inflammatory bowel diseases. Clin Sci. 2025;139(13):777–790. doi:10.1042/CS20256383
23. Ma Y, Wei J, He W, Ren J. Neutrophil extracellular traps in cancer. MedComm. 2024;5(8):e647. doi:10.1002/mco2.647
24. Wang Y, Yang K, Li J, Wang C, Li P, Du L. Neutrophil extracellular traps in cancer: from mechanisms to treatments. Clin Transl Med. 2025;15(6):e70368. doi:10.1002/ctm2.70368
25. Xia X, Zhang Z, Zhu C, et al. Author Correction: neutrophil extracellular traps promote metastasis in gastric cancer patients with postoperative abdominal infectious complications. Nat Commun. 2025;16(1):2381. doi:10.1038/s41467-025-57805-7
26. Li Q, Zhu CC, Ni B, et al. Lysyl oxidase promotes liver metastasis of gastric cancer via facilitating the reciprocal interactions between tumor cells and cancer associated fibroblasts. EBioMedicine. 2019;49:157–171. doi:10.1016/j.ebiom.2019.10.037
27. Xia X, Wang S, Ni B, et al. Correction to: hypoxic gastric cancer-derived exosomes promote progression and metastasis via MiR-301a-3p/PHD3/HIF-1α positive feedback loop. Oncogene. 2021;40(41):6058. doi:10.1038/s41388-021-01989-x
28. Albrengues J, Shields MA, Ng D, et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science. 2018;361(6409)
29. Rebernick R, Fahmy L, Glover C, et al. DNA Area and NETosis Analysis (DANA): a high-throughput method to quantify neutrophil extracellular traps in fluorescent microscope images. Biol Proced Online. 2018;20:7. doi:10.1186/s12575-018-0072-y
30. Tong X, Xiao M, Yang J, et al. The TMBIM1-YBX1 axis orchestrates MDSC recruitment and immunosuppressive microenvironment in pancreatic cancer. Theranostics. 2025;15(7):2794–2813. doi:10.7150/thno.111180
31. Zhang X, Wang WB, Cai XY, et al. MNDA promotes immunosuppression in microsatellite instability-high colorectal cancer by facilitating PMN-MDSC infiltration via H3K18 lactylation. J Transl Med. 2025;23(1):1049. doi:10.1186/s12967-025-07097-8
32. Teng Y, Chen Y, Xia X, et al. PAD2-mediated citrullination of STAT3 enhances immunosuppressive function of PMN-MDSCs in tumor-bearing hosts. Int Immunopharmacol. 2025;164:115303. doi:10.1016/j.intimp.2025.115303
33. Zhang W, Fang X, Gao C, et al. MDSCs in sepsis-induced immunosuppression and its potential therapeutic targets. Cytokine Growth Factor Rev. 2023;69:90–103. doi:10.1016/j.cytogfr.2022.07.007
34. Zhou J, Yin H, Pan J, et al. Lnk deficiency attenuates the immunosuppressive capacity of MDSCs via ferroptosis to suppress tumor development. Cell Death Dis. 2025;16(1):610. doi:10.1038/s41419-025-07948-8
35. Mathias B, Delmas AL, Ozrazgat-Baslanti T, et al. Human myeloid-derived suppressor cells are associated with chronic immune suppression after severe sepsis/septic shock. Ann Surg. 2017;265(4):827–834. doi:10.1097/SLA.0000000000001783
36. Conche C, Finkelmeier F, Pešić M, et al. Combining ferroptosis induction with MDSC blockade renders primary tumours and metastases in liver sensitive to immune checkpoint blockade. Gut. 2023;72(9):1774–1782. doi:10.1136/gutjnl-2022-327909
37. Zhu T, Zou X, Yang C, et al. Neutrophil extracellular traps promote gastric cancer metastasis by inducing epithelial‑mesenchymal transition. Int J Mol Med. 2021;48(1)
38. Demkow U. Neutrophil extracellular traps (nets) in cancer invasion, evasion and metastasis. Cancers. 2021;13(17):4495. doi:10.3390/cancers13174495
39. Adrover JM, McDowell SAC, He XY, Quail DF, Egeblad M. NETworking with cancer: the bidirectional interplay between cancer and neutrophil extracellular traps. Cancer Cell. 2023;41(3):505–526. doi:10.1016/j.ccell.2023.02.001
40. Zhang Y, Hu Y, Ma C, et al. Diagnostic, therapeutic predictive, and prognostic value of neutrophil extracellular traps in patients with gastric adenocarcinoma. Front Oncol. 2020;10:1036. doi:10.3389/fonc.2020.01036
41. Tang J, Xia J, Gao H, et al. IL33-induced neutrophil extracellular traps (NETs) mediate a positive feedback loop for synovial inflammation and NET amplification in rheumatoid arthritis. Exp Mol Med. 2024;56(12):2602–2616. doi:10.1038/s12276-024-01351-7
42. Sun X, Gui Y, Yang T, et al. PD-L1(+) neutrophils induced NETs in malignant ascites is a potential biomarker in HCC. Cancer Immunol Immunother. 2024;73(12):254. doi:10.1007/s00262-024-03833-z
43. Mao X, Xu J, Wang W, et al. Crosstalk between cancer-associated fibroblasts and immune cells in the tumor microenvironment: new findings and future perspectives. Mol Cancer. 2021;20(1):131. doi:10.1186/s12943-021-01428-1
44. Peng Z, Fang C, Tong Z, Rao Q, Ren Z, Hu K. Crosstalk between cancer-associated fibroblasts and myeloid cells shapes the heterogeneous microenvironment of gastric cancer. Curr Genomics. 2024;25(5):390–411. doi:10.2174/0113892029300608240531111743
45. Yang P, Hu Y, Zhou Q. The CXCL12-CXCR4 signaling axis plays a key role in cancer metastasis and is a potential target for developing novel therapeutics against metastatic cancer. Curr Med Chem. 2020;27(33):5543–5561. doi:10.2174/0929867326666191113113110
46. Wang Y, Zhang Z, Sun W, et al. Ferroptosis in colorectal cancer: potential mechanisms and effective therapeutic targets. Biomed Pharmacother. 2022;153:113524. doi:10.1016/j.biopha.2022.113524
47. Zhao L, Gu C, Gan Y, Shao L, Chen H, Zhu H. Exosome-mediated siRNA delivery to suppress postoperative breast cancer metastasis. J Control Release. 2020;318:1–15. doi:10.1016/j.jconrel.2019.12.005
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.